 Ruimei Liu1
Ruimei Liu1 Wei Tang
Wei Tang Chen Zhang
Chen Zhang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 10 December 2021
Sec. Behavioral and Psychiatric Genetics
Volume 12 - 2021 | https://doi.org/10.3389/fgene.2021.781625
It has been proposed that immune abnormalities may be implicated with pathophysiology of schizophrenia. The nod-like receptor pyrin domain-contraining protein 3 (NLRP3) can trigger immune-inflammatory cascade reactions. In this study, we intended to identify the role of gene encoding NLRP3 (NLRP3) in susceptibility to schizophrenia and its clinical features. For the NLRP3 mRNA expression analysis, 53 drug-naïve patients with first-episode schizophrenia and 56 healthy controls were enrolled. For the genetic study, a total of 823 schizophrenia patients and 859 controls were recruited. Among them, 239 drug-naïve patients with first-episode schizophrenia were enrolled for clinical evaluation. There is no significant difference in NLRP3 mRNA levels between patients with schizophrenia and healthy controls (p = 0.07). We did not observe any significant differences in allele and genotype frequencies of rs10754558 polymorphism between the schizophrenia and control groups. We noticed significant differences in the scores of RBANS attention and total scores between the patients with different genotypes of rs10754558 polymorphism (p = 0.001 and p < 0.01, respectively). Further eQTL analysis presented a significant association between the rs10754558 polymorphism and NLRP3 in frontal cortex (p = 0.0028, p = 0.028 after Bonferroni correction). Although our findings did not support NLRP3 confer susceptibility to schizophrenia, NLRP3 may be a risk factor for cognitive impairment, especially attention deficit in this disorder.
Schizophrenia is a deliberating and severe neuropsychiatric disease with a cluster of psychotic symptoms and cognitive impairment. Early studies have documented that maternal infection during pregnancy is implicated with increased risk of schizophrenia in the offspring (Brown and Derkits, 2010). It is known that certain clinical and genetic features are sheared between some autoimmune diseases and schizophrenia (Benros et al., 2011). Therefore, it has been proposed that immunologic abnormalities is likely to be involved in the development of schizophrenia (Tomasik et al., 2016).
Evidence from genetic studies have indicated that the concordance rates of schizophrenia for monozygotic twins are around 40–50%, and its heritability is estimated approximately 80% (Sullivan et al., 2003). As such, genetic studies of schizophrenia may provide some clues to uncover the biological mechanism of this disorder (Dhindsa and Goldstein, 2016). Several genome-wide association studies (GWASs) have consistently reported that schizophrenia susceptibility genes are located in the region of extended human major histocompatibility complex (MHC) (Shi et al., 2009; Stefansson et al., 2009). A recent meta-analysis on postmortem brain studies showed a prominent increase in the expression of pro-inflammatory genes in schizophrenia patients (van Kesteren et al., 2017). Our previous studies have identified the important roles of complement factors and pro-inflammatory cytokines in the pathophysiology of schizophrenia (Ni et al., 2015; Wang et al., 2015; Zhang et al., 2017a; Zhang et al., 2017b; Zhang et al., 2018). These results strengthened the hypothesis of immune-inflammatory processes in the pathogenesis of schizophrenia.
Based on this hypothesis, maternal immune activation (MIA) animal model is used to mimick schizophrenia-like pathological, neurochemical changes and behaviors in rodents (Zuckerman and Weiner, 2005; Wolff et al., 2011). Recent studies demonstrated that nod-like receptor pyrin domain-contraining protein 3 (NLRP3) inflammasome is involved in the schizophrenia-like behaviors of MIA model (Ventura et al., 2020; Talukdar et al., 2021). The NLRP3 inflammasome plays a key role in the innate immune system that mediates pro-inflammatory cytokines (Ventura et al., 2020). The involvement of NLRP3 in major psychiatric disorders has been reported in patients with major depressive disorder (Alcocer-Gomez et al., 2014; Tian et al., 2021) and bipolar disorder (Kim et al., 2016; Scaini et al., 2019). However, the relationship of NLRP3 with schizophrenia is not well known.
The human gene encoding NLRP3 (NLRP3) is located on chromosome 1q44, where linkage findings were reported in schizophrenia (Saviouk et al., 2005). To the best of our knowledge, there is no genetic study to identify the role of NLRP3 in schizophrenia. In this study, we hypothesized that NLRP3 may confer susceptibility to schizophrenia. First, we examined the difference of NLRP3 mRNA expression between schizophrenia patients and healthy controls. Rs10754558 is a functional single-neucleotide polymorphism (SNP) and mapped in the 3′-untranslated region of NLRP3. Evidence showed that this SNP influences the stability of NLRP3 mRNA (Hitomi et al., 2009). Thus, we secondly sought to characterize the association of NLRP3 rs10754558 polymorphism with schizophrenia in Han Chinese. Finally, we planned to conduct an eQTL (expression quantitative trait loci) analysis to detect the role of the rs10754558 polymorphism in NLRP3 mRNA expression in brain.
We recruited schizophrenia patients from three mental institutions in Eastern China, including Jinhua Wenzhou Kangning Hospital, Jinhua Second Hospital and Shanghai Mental Health Center. The criteria of patients with schizophrenia have been documented in our previous studies (Zhang et al., 2016a; Wang et al., 2016; Lu et al., 2017; Wang et al., 2020). In brief, criteria are as followings: 1) met the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV) criteria for schizophrenia; 2) had a stable condition for more than 6 months prior to participate in this study; 3) had no physical illness or other psychiatric disorders. The healthy subjects were hospital staff and students in local medical schools, who have no physical or psychiatric diseases. All subjects were Han Chinese.
For the NLRP3 mRNA expression analysis, 53 drug-naïve patients with first-episode schizophrenia and 56 healthy subjects were included. For the genetic study, a total of 823 schizophrenia patients and 859 controls were recruited. Among them, 239 drug-naïve patients with first-episode schizophrenia were enrolled for clinical evaluation. Detailed information on the participants was previously described (Zhang et al., 2014).
The Positive and Negative Syndrome Scale (PANSS) was used to evaluate the severity of psychotic symptoms in schizophrenia patients. There is a correlation coefficient greater than 0.8 for the PANSS total scores in repeated assessments (Zhu et al., 2015). The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS) was used to evaluate cognitive performance in this study (Randolph et al., 1998).
RNA extraction and Quantitative real-time polymerase chain reaction (PCR)
The processes of fasting peripheral blood extraction, total RNA extraction, reverse transcription reaction and RT-PCR were documented in our previous study (Zhang et al., 2016b).
We detected the difference of NLRP3 expression in brain between schizophrenia patients and healthy subjects through SZDB database, a newly developed on-line platform for schizophrenia research (http://www.szdb.org/) (Wu et al., 2017).
SNP rs10754558 was genotyped using Improved Multiplex ligase Detection Rection (iMLDR) method, with technical support from Center for Human Genetics Resarch, Shanghai Genesky Biotech Co., Ltd.
We extracted the schizophrenia genetic data from the Psychiatric Genomics Consortium (PGC, http://www.broadinstitute.org/mpg/ricopili/) database and reanalyzed the data set as an independent sample to validate our genetic results.
It is known that schizophrenia is derived from abnormalities in brain. As such, we used the brain eQTL database (http://peana-od.inf.um.es:8080/UKBECv12/) for eQTL analysis of rs10754558 polymorphism in brain.
SHEsisplus (Shen et al., 2016) software was used to calculate the Hardy-Weinberg equilibrium (HWE) and compare differences of genotype and allele distribution between schizophrenia and control groups. ANCOVA was conducted to compare the difference of NLRP3 mRNA expression between schizophrenia patients and healthy controls, with covariates such as gender, smoking and age controlled in the model. Statistical significance of differences in clinical features between genotype groups was compared using ANCOVA with covariates (e.g., age, gender, course of disease, treatment duration and doses) to minimize their potential effects. Power analysis was carried out by using the Quanto software (http://hydra.usc.edu/GxE). SPSS 17.0 (SPSS, Inc. Chicago, IL, United States) was used to conduct all the statistical calculation. p values were two-tailed, and p < 0.05 means statistically significant. Bonferroni correction was performed during multiple comparison testing.
For the expression analysis, demographic characteristics of our sample were presented in Supplementary Table S1. Schizophrenia and control groups are well-matched with respect to age, sex and smoking. The RT-PCR results showed no significant difference in NLRP3 mRNA levels between schizophrenia and control groups (p = 0.07, Figure 1). We extracted brain NLRP3 mRNA expression data from SZDB database. We did not observe significant differences of NLRP3 mRNA expression in prefrontal cortex, hippocampus and striatum between schizophrenia patients and healthy controls (Supplementary Figure S1).
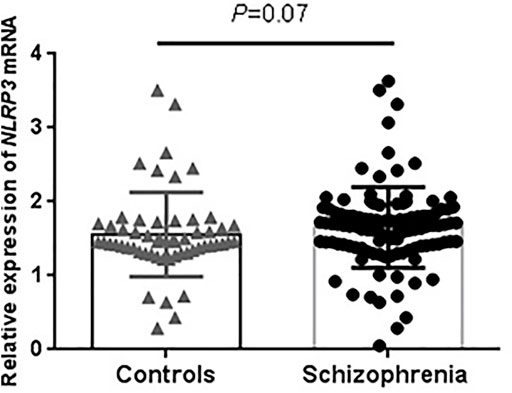
FIGURE 1. Peripheral expression levels of NLRP3 mRNA in drug-naïve patients with first-episode schizophrenia and healthy controls. NLRP3 mRNA level was normalized to that of GAPDH. Schizophrenia group (n = 53); Control group (n = 56).
For the genetic analysis, we enrolled 823 schizophrenia patients and 859 healthy controls in this study. Demographic characteristics are well matched in both groups. No deviation from HWE was observed in genotype distribution for rs10754558 polymorphism in control group. The results of rs10754558 polymorphism were presented in Table 1. No significant differences in allele and genotype frequencies were found between the schizophrenia and control groups. With false positive rate controlled as 0.05, the statistical power to detect the odds ratio (OR) value as 1.5 for risk allele was expected to be more than 85% in our samples under the log additive model. We further examined the genetic association between the rs10754558 polymorphism and schizophrenia in PGC database, and we also did not observe significant association of rs10754558 polymorphism with schizophrenia (Supplementary Figure S2).

TABLE 1. Distribution of rs10754558 genotypes and alleles in schizophrenia patients and healthy controls.
There were 239 drug-naïve patients with first-episode schizophrenia participated in the clinical evaluation. We examined the association of rs10754558 polymorphism with psychotic symptoms and cognitive performance in schizophrenia patients by comparing scores of the PANSS and RBANS with genotypes of SNP rs10754558. We observed significant differences in the scores of RBANS attention and total scores between the patients with different genotypes of rs10754558 polymorphism (p = 0.001 and p < 0.01, respectively), as shown in Table 2. Post-hoc comparisons showed that rs10754558 G/G or G/C carriers had lower RBANS attention and total scores than those with C/C carriers (Ps<0.01). Subsequently, we conducted an eQTL analysis to test the influence of rs10754558 polymorphism on NLRP3 mRNA expression in brain. Figure 2 showed that carriers with G allele have higher levels of NLRP3 expression than those without G allele in frontal cortex.
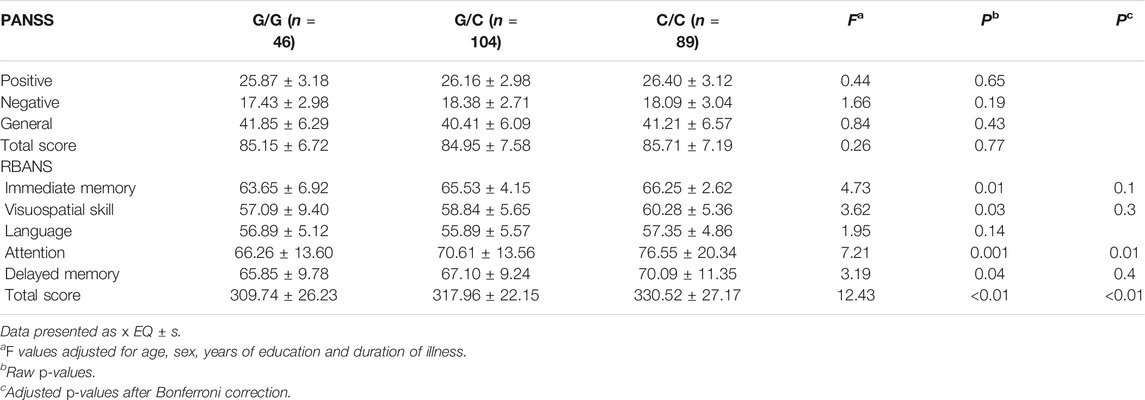
TABLE 2. PANSS and RBANS performance comparisons of rs10754558 genotypic groups in patients with schizophrenia.
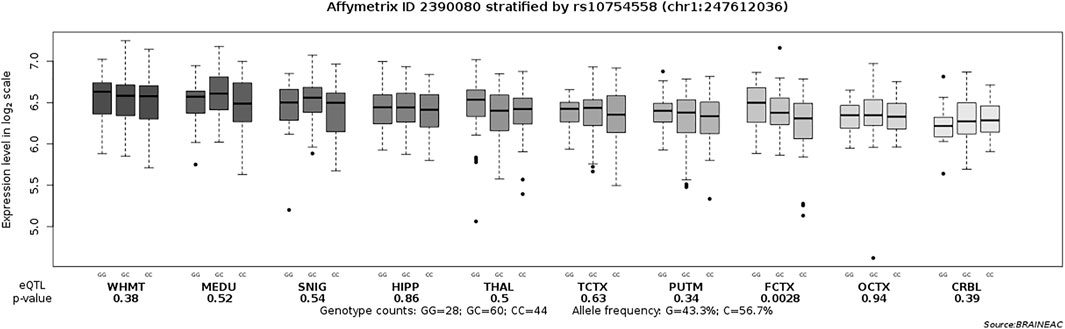
FIGURE 2. Association of rs10754558 polymorphism with the NLRP3 mRNA expression level in ten brain regions (Affymetrix ID 2390080). SNIG, substantia nigra; PUTM, putamen (at the level of the anterior commissure); MEDU, the inferior olivary nucleus (sub-dissected from the medulla); THAL, thalamus (at the level of the lateral geniculate nucleus); OCTX, occipital cortex; HIPP, hippocampus; FCTX, frontal cortex; TCTX, temporal cortex; WHMT, intralobular white matter; CRBL, cerebellar cortex. Data were extracted from the BRAINEAC database (http://peana-od.inf.um.es:8080/UKBECv12/).
There is a growing body of evidence implying that schizophrenia is attributed to a dysregulation of innate and adaptive immune system (Michel et al., 2012). Our previous studies have provided supportive evidence for the involvement of innate immune response in the pathogenesis of schizophrenia (Ni et al., 2015; Wang et al., 2015; Zhang et al., 2017b). NLRP3 is an intracellular protein involved in the initiation of the innate immune response (Bonar et al., 2012). It is known that NLRP3 mutation confers susceptibility to immune disorders (Pontillo et al., 2010). Accordingly, we employed this study to identify the role of NLRP3 in schizophrenia.
In this study, we found that peripheral NLRP3 mRNA level has no difference between patients with schizophrenia and healthy controls. We also did not observe the difference of NLRP3 mRNA level in brain between schizophrenia and controls. It has been well-established that MIA is associated with an increased risk of schizophrenia in offspring (Scola and Duong, 2017). Recent preclinical studies have found that activation of NLRP3 when LPS counts may be one of the potential mechanisms driving schizophrenia-like behavior in offspring in MIA model (Ventura et al., 2020; Talukdar et al., 2021). However, a postmortem study compared the expression of NLRP3-related protein in brain tissue from patients with schizophrenia and bipolar disorder in which increased level of NLRP3 was observed in patients with bipolar disorder, but not schizophrenia (Kim et al., 2016). Another comparative study also showed that peripheral level of NLRP3 is increased in patients with bipolar disorder, but not schizophrenia (Garcia-Alvarez et al., 2018). As such, our findings implied that NLRP3 mRNA level is not a risk factor for schizophrenia.
Another aim of this study is to detect the association of NLRP3 functional SNP rs10754558 with schizophrenia. SNP rs10754558 is located in the 3′-untranlated region (3′-UTR) of NLRP3 and affects expression of NLRP3 mRNA through altering its stability (Hitomi et al., 2009). This SNP has been reported to confer susceptibility to disorders of the central nervous system, such as Alzheimer’s disease (Tan et al., 2013), multiple sclerosis (Imani et al., 2018) and ischemic stroke (Lv et al., 2020). However, our results did not support the significance for rs10754558 polymorphism in the susceptibility to schizophrenia in either Han Chinese or European ancestry from PGC GWAS. This suggests that NLRP3 SNP rs10754558 is probably not a risk SNP for schizophrenia.
Schizophrenia is known to have variable clinical presentation. Therefore, we further sought to determine the role of rs10754558 in the expression of clinical features in a sample of drug-naïve patients with first-episode schizophrenia. We observed a significant association of rs10754558 polymorphism with RBANS attention and total scores in such patients, but not psychotic symptoms. This suggested that rs10754558 polymorphism may result in cognitive impairment, especially attention deficit in schizophrenia. We further carried out an eQTL analysis to detect the effect of rs10754558 polymorphism on NLRP3 mRNA expression in brain. Data showed that rs10754558 polymorphism is significantly associated with the NLRP3 mRNA expression in frontal cortex, and the risk allele G is associated with increased level of NLRP3 mRNA. There is accumulating evidence from magnetic resonance imaging (MRI) studies showing that frontal regions are critical for cognitive impairment in schizophrenia (Glascher et al., 2019). Our recent findings indicated that dysfunctional connectivity of frontal cortices is associated with cognitive impairment in patients with schizophrenia (Yu et al., 2021). North et al. (2021) found that elevated level of peripheral inflammation is a risk factor for cognitive deficits and brain structure in schizophrenia, especially reduced attention and cortical thickness. Meta-analysis also showed that attention deficit has a strongest relationship with peripheral inflammation in all cognitive domains tested in patients with schizophrenia (Bora, 2019). Our previous studies have demonstrated that elevated levels of pro-inflammatory cytokines are associated with cognitive impairment in schizophrenia (Zhang et al., 2017a; Zhang et al., 2018; Xu et al., 2019). NLRP3 is a critical regulator of pro-inflammatory cytokines production by activated caspase-1 to initiating an inflammatory form of cell death (Feng et al., 2021). Recent literature documented that enriched NLRP3 may lead to cognitive deficits, and inhibition of NLRP3 could ameliorate such impairment (Fang et al., 2021). As such, our results implied that increased level of NLRP3 mRNA may result in cognitive impairment, especially attention deficit in patients with schizophrenia, possibly through triggering a cascade of inflammatory responses in frontal cortex.
Some limitation in this study should be mentioned. First, we evaluated the NLRP3 mRNA in peripheral level, not in brain. It is uncertain whether the peripheral level of NLRP3 mRNA is correlated with the level in the central nervous system. Second, the lack of a significant association of rs10754558 polymorphism and schizophrenia may be caused by the modest sample size, whereas the power reached statistical significance. Accordingly, our findings should be considered only preliminary and require further investigations for validation in independent samples. Third, this study employed samples from Eastern China and may not be representative of the Han Chinese population in general, nor other closely related populations in the area. Finally, we have analyzed only one functional SNP in NLRP3 to evaluate the genetic relationship with schizophrenia. This means further analyses need to be carried out with more variants in the NLRP3 for validation of our results.
In conclusion, we performed a comprehensive analysis to detect the association of NLRP3 with schizophrenia in Han Chinese. Although our findings did not support NLRP3 confer susceptibility to schizophrenia, NLRP3 may be a risk factor for cognitive impairment, especially attention deficit in this disorder. Further investigations are required for validation of our results in larger and independent sample across various ethnicities.
The original contributions presented in the study are publicly available. This data can be found here: https://datadryad.org/stash/share/vfrPQUqJNc28WuM76t41oWjYIBJY403RcXSMTr33pjM
The studies involving human participants were reviewed and approved by the Institutional Review Boards of the Shanghai Mental Health Center. The patients/participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
CZ and RL contributed to the overall design of the study. WT and WF wrote the protocol for the genotyping. RL, WT, WW, FX, and WF got involved sample collection. WW and CZ undertook the statistical analysis and interpretation of data. CZ and RL wrote the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the National Key Research and Development Program of China (2018YFC1314302), the National Natural Science Foundation of China (81471358 and 81771450), the Shanghai Science and Technology Commission Foundation (19411969300), and Medical and Public Health Program for Social Development in Wuxi (WX18IIAN032).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer LW declared a past co-authorship with the authors CZ, WT, WF, YZ to the handling editor.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We are deeply grateful to all participants.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2021.781625/full#supplementary-material
Alcocer-Gómez, E., de Miguel, M., Casas-Barquero, N., Núñez-Vasco, J., Sánchez-Alcazar, J. A., Fernández-Rodríguez, A., et al. (2014). NLRP3 Inflammasome Is Activated in Mononuclear Blood Cells from Patients with Major Depressive Disorder. Brain Behav. Immun. 36, 111–117. doi:10.1016/j.bbi.2013.10.017
Benros, M. E., Nielsen, P. R., Nordentoft, M., Eaton, W. W., Dalton, S. O., and Mortensen, P. B. (2011). Autoimmune Diseases and Severe Infections as Risk Factors for Schizophrenia: a 30-year Population-Based Register Study. Ajp 168, 1303–1310. doi:10.1176/appi.ajp.2011.11030516
Bonar, S. L., Brydges, S. D., Mueller, J. L., McGeough, M. D., Pena, C., Chen, D., et al. (2012). Constitutively Activated NLRP3 Inflammasome Causes Inflammation and Abnormal Skeletal Development in Mice. PLoS One 7, e35979. doi:10.1371/journal.pone.0035979
Bora, E. (2019). Peripheral Inflammatory and Neurotrophic Biomarkers of Cognitive Impairment in Schizophrenia: a Meta-Analysis. Psychol. Med. 49, 1971–1979. doi:10.1017/s0033291719001685
Brown, A. S., and Derkits, E. J. (2010). Prenatal Infection and Schizophrenia: a Review of Epidemiologic and Translational Studies. Ajp 167, 261–280. doi:10.1176/appi.ajp.2009.09030361
Dhindsa, R. S., and Goldstein, D. B. (2016). From Genetics to Physiology at Last. Nature 530, 162–163. doi:10.1038/nature16874
Fang, P., Chen, C., Zheng, F., Jia, J., Chen, T., Zhu, J., et al. (2021). NLRP3 Inflammasome Inhibition by Histone Acetylation Ameliorates Sevoflurane-Induced Cognitive Impairment in Aged Mice by Activating the Autophagy Pathway. Brain Res. Bull. 172, 79–88. doi:10.1016/j.brainresbull.2021.04.016
Feng, X., Luo, D., Wei, G., Zhan, F., Hua, F., and Xu, G. (2021). Non-coding RNAs: The Key Regulators in NLRP3 Inflammasome-Mediated Inflammatory Diseases. Int. Immunopharmacology 100, 108105. doi:10.1016/j.intimp.2021.108105
García-Álvarez, L., Caso, J. R., García-Portilla, M. P., de la Fuente-Tomás, L., González-Blanco, L., Sáiz Martínez, P., et al. (2018). Regulation of Inflammatory Pathways in Schizophrenia: A Comparative Study with Bipolar Disorder and Healthy Controls. Eur. Psychiatr. 47, 50–59. doi:10.1016/j.eurpsy.2017.09.007
Gläscher, J., Adolphs, R., and Tranel, D. (2019). Model-based Lesion Mapping of Cognitive Control Using the Wisconsin Card Sorting Test. Nat. Commun. 10, 20. doi:10.1038/s41467-018-07912-5
Hitomi, Y., Ebisawa, M., Tomikawa, M., Imai, T., Komata, T., Hirota, T., et al. (2009). Associations of Functional NLRP3 Polymorphisms with Susceptibility to Food-Induced Anaphylaxis and Aspirin-Induced Asthma. J. Allergy Clin. Immunol. 124, 779–785. doi:10.1016/j.jaci.2009.07.044
Imani, D., Azimi, A., Salehi, Z., Rezaei, N., Emamnejad, R., Sadr, M., et al. (2018). Association of Nod-like Receptor Protein-3 Single Nucleotide Gene Polymorphisms and Expression with the Susceptibility to Relapsing-Remitting Multiple Sclerosis. Int. J. Immunogenet. 45, 329–336. doi:10.1111/iji.12401
Kim, H. K., Andreazza, A. C., Elmi, N., Chen, W., and Young, L. T. (2016). Nod-like Receptor Pyrin Containing 3 (NLRP3) in the post-mortem Frontal Cortex from Patients with Bipolar Disorder: A Potential Mediator between Mitochondria and Immune-Activation. J. Psychiatr. Res. 72, 43–50. doi:10.1016/j.jpsychires.2015.10.015
Lu, W., Zhang, Y., Fang, X., Fan, W., Tang, W., Cai, J., et al. (2017). Genetic Association Analysis of microRNA137 and its Target Complex 1 with Schizophrenia in Han Chinese. Sci. Rep. 7, 15084. doi:10.1038/s41598-017-15315-7
Lv, J., Jiang, X., Zhang, J., Peng, X., and Lin, H. (2020). Combined Polymorphisms in Genes Encoding the Inflammasome Components NLRP3 and CARD8 Confer Risk of Ischemic Stroke in Men. J. Stroke Cerebrovasc. Dis. 29, 104874. doi:10.1016/j.jstrokecerebrovasdis.2020.104874
Michel, M., Schmidt, M. J., and Mirnics, K. (2012). Immune System Gene Dysregulation in Autism and Schizophrenia. Devel Neurobio 72, 1277–1287. doi:10.1002/dneu.22044
Ni, J., Hu, S., Zhang, J., Tang, W., Lu, W., and Zhang, C. (2015). A Preliminary Genetic Analysis of Complement 3 Gene and Schizophrenia. PLoS One 10, e0136372. doi:10.1371/journal.pone.0136372
North, H. F., Bruggemann, J., Cropley, V., Swaminathan, V., Sundram, S., Lenroot, R., et al. (2021). Increased Peripheral Inflammation in Schizophrenia Is Associated with Worse Cognitive Performance and Related Cortical Thickness Reductions. Eur. Arch. Psychiatry Clin. Neurosci. 271, 595–607. doi:10.1007/s00406-021-01237-z
Pontillo, A., Brandão, L. A., Guimarães, R. L., Segat, L., Athanasakis, E., and Crovella, S. (2010). A 3′UTR SNP in NLRP3 Gene Is Associated with Susceptibility to HIV-1 Infection. J. Acquir Immune Defic Syndr. 54, 236–240. doi:10.1097/qai.0b013e3181dd17d4
Randolph, C., Tierney, M. C., Mohr, E., and Chase, T. N. (1998). The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS): Preliminary Clinical Validity. J. Clin. Exp. Neuropsychol. 20, 310–319. doi:10.1076/jcen.20.3.310.823
Saviouk, V., Chow, E. W. C., Bassett, A. S., and Brzustowicz, L. M. (2005). Tumor Necrosis Factor Promoter Haplotype Associated with Schizophrenia Reveals a Linked Locus on 1q44. Mol. Psychiatry 10, 375–383. doi:10.1038/sj.mp.4001582
Scaini, G., Barichello, T., Fries, G. R., Kennon, E. A., Andrews, T., Nix, B. R., et al. (2019). TSPO Upregulation in Bipolar Disorder and Concomitant Downregulation of Mitophagic Proteins and NLRP3 Inflammasome Activation. Neuropsychopharmacol. 44, 1291–1299. doi:10.1038/s41386-018-0293-4
Scola, G., and Duong, A. (2017). Prenatal Maternal Immune Activation and Brain Development with Relevance to Psychiatric Disorders. Neuroscience 346, 403–408. doi:10.1016/j.neuroscience.2017.01.033
Shen, J., Li, Z., Chen, J., Song, Z., Zhou, Z., and Shi, Y. (2016). SHEsisPlus, a Toolset for Genetic Studies on Polyploid Species. Sci. Rep. 6, 24095. doi:10.1038/srep24095
Shi, J., Levinson, D. F., Duan, J., Sanders, A. R., Zheng, Y., Pe’er, I., et al. (2009). Common Variants on Chromosome 6p22.1 Are Associated with Schizophrenia. Nature 460, 753–757. doi:10.1038/nature08192
Stefansson, H., Ophoff, R. A., Ophoff, R. A., Steinberg, S., Andreassen, O. A., Cichon, S., et al. (2009). Common Variants Conferring Risk of Schizophrenia. Nature 460, 744–747. doi:10.1038/nature08186
Sullivan, P. F., Kendler, K. S., and Neale, M. C. (2003). Schizophrenia as a Complex Trait. Arch. Gen. Psychiatry 60, 1187–1192. doi:10.1001/archpsyc.60.12.1187
Talukdar, P. M., Abdul, F., Maes, M., Berk, M., Venkatasubramanian, G., Kutty, B. M., et al. (2021). A Proof-Of-Concept Study of Maternal Immune Activation Mediated Induction of Toll-like Receptor (TLR) and Inflammasome Pathways Leading to Neuroprogressive Changes and Schizophrenia-like Behaviours in Offspring. Eur. Neuropsychopharmacol. 52, 48–61. doi:10.1016/j.euroneuro.2021.06.009
Tan, M.-S., Yu, J.-T., Jiang, T., Zhu, X.-C., Wang, H.-F., Zhang, W., et al. (2013). NLRP3 Polymorphisms Are Associated with Late-Onset Alzheimer's Disease in Han Chinese. J. Neuroimmunology 265, 91–95. doi:10.1016/j.jneuroim.2013.10.002
Tian, H., Li, G., Xu, G., Liu, J., Wan, X., Zhang, J., et al. (2021). Inflammatory Cytokines Derived from Peripheral Blood Contribute to the Modified Electroconvulsive Therapy-Induced Cognitive Deficits in Major Depressive Disorder. Eur. Arch. Psychiatry Clin. Neurosci. 271, 475–485. doi:10.1007/s00406-020-01128-9
Tomasik, J., Rahmoune, H., Guest, P. C., and Bahn, S. (2016). Neuroimmune Biomarkers in Schizophrenia. Schizophrenia Res. 176, 3–13. doi:10.1016/j.schres.2014.07.025
van Kesteren, C. F. M. G., Gremmels, H., de Witte, L. D., Hol, E. M., Van Gool, A. R., Falkai, P. G., et al. (2017). Immune Involvement in the Pathogenesis of Schizophrenia: a Meta-Analysis on Postmortem Brain Studies. Transl Psychiatry 7, e1075. doi:10.1038/tp.2017.4
Ventura, L., Freiberger, V., Thiesen, V. B., Dias, P., Dutra, M. L., Silva, B. B., et al. (2020). Involvement of NLRP3 Inflammasome in Schizophrenia-like Behaviour in Young Animals after Maternal Immune Activation. Acta Neuropsychiatr. 32, 321–327. doi:10.1017/neu.2020.27
Wang, D., Tang, W., Zhao, J., Fan, W., Zhang, Y., and Zhang, C. (2020). A Comprehensive Analysis of the Effect of SIRT1 Variation on the Risk of Schizophrenia and Depressive Symptoms. Front. Genet. 11, 832. doi:10.3389/fgene.2020.00832
Wang, P., Cai, J., Ni, J., Zhang, J., Tang, W., and Zhang, C. (2016). The NCAN Gene: Schizophrenia Susceptibility and Cognitive Dysfunction. Ndt Volume 12, 2875–2883. doi:10.2147/ndt.s118160
Wang, S., Lu, H., Ni, J., Zhang, J., Tang, W., Lu, W., et al. (2015). An Evaluation of Association between Common Variants in C4BPB/C4BPA Genes and Schizophrenia. Neurosci. Lett. 590, 189–192. doi:10.1016/j.neulet.2015.02.005
Wolff, A. R., Cheyne, K. R., and Bilkey, D. K. (2011). Behavioural Deficits Associated with Maternal Immune Activation in the Rat Model of Schizophrenia. Behav. Brain Res. 225, 382–387. doi:10.1016/j.bbr.2011.07.033
Wu, Y., Yao, Y. G., and Luo, X. J. (2017). SZDB: A Database for Schizophrenia Genetic Research. Schizophr Bull. 43, 459–471. doi:10.1093/schbul/sbw102
Xu, F., Fan, W., Wang, W., Tang, W., Yang, F., Zhang, Y., et al. (2019). Effects of omega-3 Fatty Acids on Metabolic Syndrome in Patients with Schizophrenia: a 12-week Randomized Placebo-Controlled Trial. Psychopharmacology 236, 1273–1279. doi:10.1007/s00213-018-5136-9
Yu, L., Guo, L., Fang, X., Yang, F., Chen, Y., Wang, Y., et al. (2021). Altered Brain Activity in the Bilateral Frontal Cortices and Neural Correlation with Cognitive Impairment in Schizophrenia. Brain Imaging Behav. doi:10.1007/s11682-021-00516-6
Zhang, C., Cai, J., Zhang, J., Li, Z., Guo, Z., Zhang, X., et al. (2014). Genetic Modulation of Working Memory Deficits by Ankyrin 3 Gene in Schizophrenia. Prog. Neuro-Psychopharmacology Biol. Psychiatry 50, 110–115. doi:10.1016/j.pnpbp.2013.12.010
Zhang, C., Fang, X., Yao, P., Mao, Y., Cai, J., Zhang, Y., et al. (2017a). Metabolic Adverse Effects of Olanzapine on Cognitive Dysfunction: A Possible Relationship between BDNF and TNF-Alpha. Psychoneuroendocrinology 81, 138–143. doi:10.1016/j.psyneuen.2017.04.014
Zhang, C., Lu, W., Wang, Z., Ni, J., Zhang, J., Tang, W., et al. (2016a). A Comprehensive Analysis of NDST3 for Schizophrenia and Bipolar Disorder in Han Chinese. Transl Psychiatry 6, e701, e701.doi:10.1038/tp.2015.199
Zhang, C., Lv, Q., Fan, W., Tang, W., and Yi, Z. (2017b). Influence of CFH Gene on Symptom Severity of Schizophrenia. Ndt 13, 697–706. doi:10.2147/ndt.s132108
Zhang, C., Wu, Z., Hong, W., Peng, D., and Fang, Y. (2016b). Evaluating the Association between the SHANK3 Gene and Bipolar Disorder. Psychiatry Res. 244, 284–288. doi:10.1016/j.psychres.2016.07.058
Zhang, Y., Fang, X., Fan, W., Tang, W., Cai, J., Song, L., et al. (2018). Interaction between BDNF and TNF-α Genes in Schizophrenia. Psychoneuroendocrinology 89, 1–6. doi:10.1016/j.psyneuen.2017.12.024
Zhu, Y., Wang, Z., Ni, J., Zhang, Y., Chen, M., Cai, J., et al. (2015). Genetic Variant in NDUFS1 Gene Is Associated with Schizophrenia and Negative Symptoms in Han Chinese. J. Hum. Genet. 60, 11–16. doi:10.1038/jhg.2014.94
Keywords: NLRP3, cognitive impairment, schizophrenia, RBANS, polymorphism
Citation: Liu R, Tang W, Wang W, Xu F, Fan W, Zhang Y and Zhang C (2021) NLRP3 Influences Cognitive Function in Schizophrenia in Han Chinese. Front. Genet. 12:781625. doi: 10.3389/fgene.2021.781625
Received: 23 September 2021; Accepted: 19 November 2021;
Published: 10 December 2021.
Edited by:
Cunyou Zhao, Southern Medical University, ChinaReviewed by:
Qiang Wang, West China Hospital, Sichuan University, ChinaCopyright © 2021 Liu, Tang, Wang, Xu, Fan, Zhang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chen Zhang, emhhbmdjaGVuNjQ1QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.