 Mojca Simčič
Mojca Simčič Barbara Luštrek1
Barbara Luštrek1 Betka Logar
Betka Logar- 1Department of Animal Science, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia
- 2Agricultural Institute of Slovenia, Ljubljana, Slovenia
The aim of this study was to estimate genetic parameters of 26 individual and four composite type traits in first parity Cika cows. An analysis of variance was performed with the generalized linear model procedure of the SAS/STAT statistical package, where the fixed effects of year of recording, cow’s age at recording and days after calving as a linear regression were included in the model. The variance components for the direct additive genetic effect and the herd effect in all type traits were estimated using the REML method in the VCE-6 software package. The estimated heritabilities ranged from 0.42 to 0.67 for the measured body frame traits, from 0.36 to 0.80 for the scored autochthonous traits, from 0.11 to 0.61 for the scored body frame traits, and from 0.20 to 0.47 for the scored udder traits. The estimated heritabilities for the composite traits called “autochthonous characteristics”, “muscularity”, “body frame” and “udder” were 0.55, 0.19, 0.19, and 0.26, respectively. The estimated genetic correlations among the measured body frame traits were positive and high, while the majority of them among the scored body frame traits were low to moderate. The estimated proportions of variance explained by the herd effect for the composite traits “autochthonous characteristics,” “muscularity,” “body frame” and “udder” were 0.09, 0.28, 0.14, and 0.10, respectively. The estimated heritabilities for the type traits of first parity Cika cows were similar to those reported for other breeds where breeding values have been routinely predicted for a long time. All estimated genetic parameters are already used for breeding value prediction in the Cika cattle population.
Introduction
Cika cattle is a Slovenian autochthonous dual-purpose breed. The breeding goal is to preserve the original type traits of the breed and to prevent an increase of inbreeding. Cika cattle are widespread all over Slovenia, especially in their regions of origin (Bohinj, Kamnik, Tolmin). The coat color pattern is red pied sided, which is typical for this breed and very different from other cattle breeds in Slovenia. Some Cika animals have a coat color pattern similar to Pinzgauer cattle and some to Tux-Zillertaler cattle from Austria (Sambraus, 1999). Based on microsatellite genotyping, Cika cattle form an Eastern Alpine breed cluster with Pinzgauer and Pustertaler cattle. Cika cattle are considered as an authentic and valuable genetic resource (Felius et al., 2011; Simčič et al., 2013). The possibility of excluding admixed animals (sires and sire dams) from the Cika cattle breeding program using SNP haplotypes has been reported in detail elsewhere (Simčič et al., 2015a,b).
Cika cattle breeding is based on the recording of type traits of all first parity cows (Žan Lotrič et al., 2010). The age of first parity cows at the recording day is restricted to a minimum of 560 days and there are no restrictions on the maximum age. Recording time is adjusted to the rearing technology, so the animals are not recorded during the grazing season. According to the breeding program, the recording of first parity cows is planned to take place 15–120 days after calving, which is often not possible due to the grazing season in the higher mountain pastures.
The current population size of Cika cattle is 5,531 animals (www.fao.org/dad-is), which is lower than in all other breeds in Slovenia. Therefore, all breeding animals are scored by a single expert. Compared to the small population size, a large number of sires (98 in the year 2020) is used for natural service and artificial insemination in the population. The effective population size is estimated to be 117 animals. Of the animals born in 2016, 96.6% had complete pedigree data in the second generation, while 65.7% of the animals had a complete pedigree in the sixth generation. The inbreeding coefficient in the population is estimated to be 2.1%.
Genetic parameters for type traits in dairy cattle breeds have been reported in first parity cows of Holstein-Friesian (Van Dorp et al., 1998; Rupp and Boichard, 1999; Larroque and Ducrocq, 2001; Neuenschwander et al., 2005; de Haas et al., 2007; Němcová et al., 2011), Jersey (Thomas et al., 1985; Norman et al., 1988; Rogers et al., 1991), Brown Swiss (Norman et al., 1988; Dal Zotto et al., 2007; de Haas et al., 2007; Samore et al., 2010), Red Holstein (de Haas et al., 2007), Ayrshire, Guernsey and Shorthorn (Norman et al., 1988), where correlations among type traits, milk yield, fertility and longevity were estimated.
In first parity cows of beef breeds, genetic parameters for type traits were studied in Belgian blue cattle with double muscling by Gengler et al. (1995), and in Spanish Asturiana de los Valles cattle by Guitiérrez and Goyache (2002), and Guitiérrez et al. (2002). Forabosco et al. (2004) indirectly estimated longevity using correlated type traits. Genetic parameters for type traits in first parity cows of some autochthonous cattle breeds were reported as well, e.g. in the Italian autochthonous breeds Chianina (Forabosco et al., 2004), Piemontese (Mantovani et al., 2010), Rendena (Mazza and Mantovani, 2012; Mazza et al., 2014), Cabanina (Comunod et al., 2013), Valdostana cattle (Mazza et al., 2015), Alpine Grey cattle (Mancin et al., 2021) and Italian Simmental (Frigo et al., 2013).
The aim of this pilot study was to estimate genetic parameters for the measured and scored type traits included in the routine breeding values estimation of Cika cows according to the breeding program.
Materials and Methods
Data
The data were obtained from the Central database for cattle kept by the Agricultural Institute in Slovenia, which included information from the type traits recording of Slovenian first parity Cika cows scored in the years from 2006 to 2019. We assumed that the type trait recording was intended for first parity cows, so we limited the age at recording to a maximum of 1,460 days or 4 years as recommended by de Haas et al. (2007) and a minimum of 607 days. On average, the animals were 1,030.7 ± 167.3 days old at recording. In addition, we excluded all cows younger than 4 years of age that were scored after the second parity. First parity cows that were not in the recommended stage of lactation on the day of recording according to the breeding program were not excluded from further analysis. Consequently, the recorded first parity cows were one to 446 days after calving, 227.5 ± 112.7 days on average. After applying these limitations, 1,815 first parity cows were included in the final analysis (Table 1).
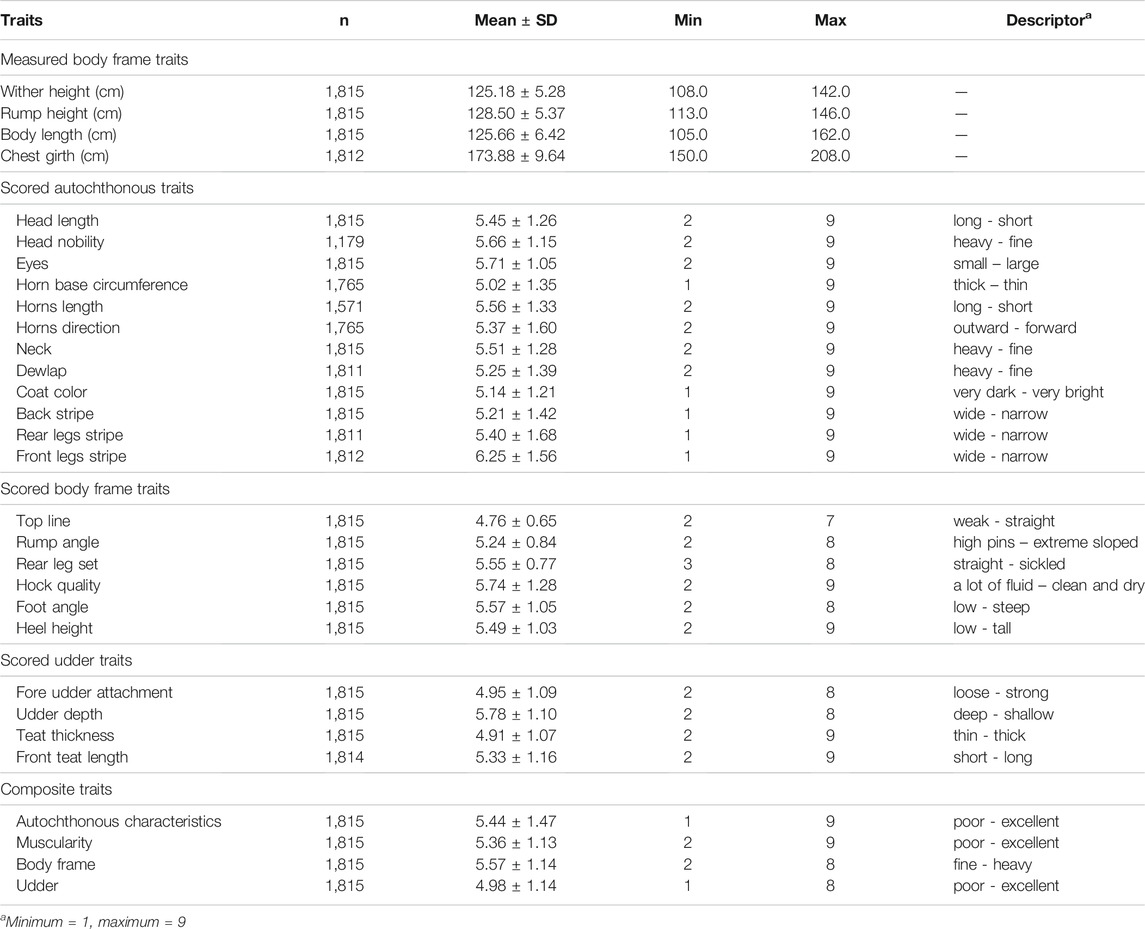
TABLE 1. Descriptive statistics of analyzed traits.
All first parity cows (1,815) with type traits and their known ancestors were included in the additive relationship matrix among all animals (2,953 animals in total). Almost 98% of the first parity cows had a known sire and 96% had a known dam. In total, 95% of the first parity cows had both parents known.
Estimation of Variance Components
To investigate the non-genetic effects to be included in the model, an analysis was performed using the GLM (generalized linear model) procedure in the SAS/STAT statistical package (version 9.4, SAS Institute Inc, 2001) with the statistical model
where yijk was the type trait, μ was the mean of the population, Yi was the fixed effect of the recording year (i = 2006, … , 2019), bI was the linear regression coefficient for the age at recording, xijk was the age at recording (days),
The matrix notation for the complete model including both non-genetic and genetic effects was expressed as:
Where y is an N x 1 vector of observations, β is the vector of systematic fixed effects of order p, q is the vector of herd effect considered as random effect, u is the vector of animal effects with order m, and e is the vector of residual effects. Likewise, X, W and Z are the corresponding incidence matrices with the appropriate dimensions.
The variance components and heritability for each individual and composite trait were estimated with the REML method in the VCE-6 software package (Groeneveld et al., 2010).
The analyzed type traits were divided into six groups according to specific body regions (Simčič et al., 2016). The measured body frame traits (wither height, rump height, body length, chest girth) were assigned to Group 1. The scored autochthonous traits were divided into three groups: head characteristics (head length, head nobility, eyes, horn base circumference, horns length, horns direction) were assigned to Group 2, neck characteristics (neck, dewlap) were assigned to Group 3, and coat color traits (coat color, back stripe, rear legs stripe, front legs stripe) were assigned to Group 4. The scored body frame traits (top line, rump angle, rear leg set, hock quality, foot angle, heel height, body frame as a composite trait) were assigned to Group 5. Udder traits (fore udder attachment, udder depth, teat thickness, front teat length, udder as a composite trait) were assigned to Group 6. The variance components and genetic parameters for these anatomically similar traits divided into six groups were estimated separately for each group using multivariate mixed models. The two remaining composite traits (autochthonous characteristics, muscularity) were analyzed with a univariate mixed model.
Results and Discussion
Variance Component Estimates
The heritabilities (h2) for all included type traits in the first parity Cika cows ranged between 0.11 and 0.80, while the proportions of variance explained by the herd effect ranged between 0.01 and 0.28. Considering the six different trait groups, the heritabilities for the measured body frame traits ranged from 0.42 to 0.67, for the scored autochthonous traits from 0.36 to 0.80, for the scored body frame traits from 0.11 to 0.61, and for the scored udder traits from 0.20 to 0.47. The estimated heritabilities for the composite traits of autochthonous characteristics, muscularity, body frame, and udder were 0.55, 0.19, 0.19, and 0.26, respectively (Table 2).
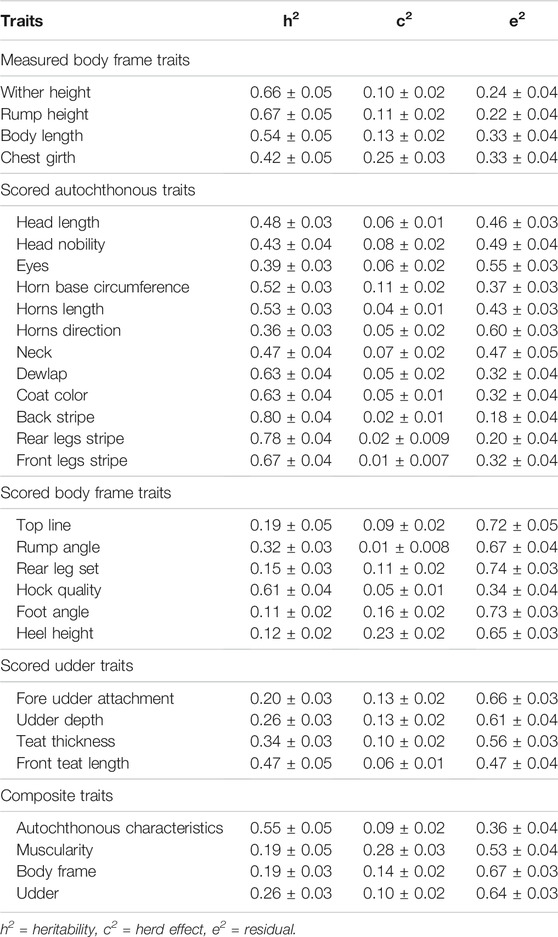
TABLE 2. Estimated proportions of variance components ± standard errors for type traits in the first parity Cika cows.
The estimated h2 for measured wither height and rump height in the first parity Cika cows were very similar, 0.66 ± 0.05 and 0.67 ± 0.05, respectively (Table 2). A lower h2 for wither height was estimated in first parity Piemontese cows (0.31 ± 0.02; Mantovani et al., 2010). A similar h2 for rump height was estimated in first parity Holstein cows (0.69 ± 0.03), American Brown Swiss cows (0.64 ± 0.02), and Red Holstein cows (0.74 ± 0.03) in Switzerland (de Haas et al., 2007), whereas the estimated h2 was lower in first parity Brown Swiss cows in Slovenia (0.46; Špehar et al., 2012). A lower h2 for rump height was estimated in first parity Rendena cows (0.52; Mazza et al., 2014), American Brown Swiss cows (0.32; Dal Zotto et al., 2007), and first parity Holstein-Friesian cows in the Czech Republic (0.45; Němcová et al., 2011). The estimated h2 for body length in the first parity Cika cows was 0.54 ± 0.05 (Table 2). A lower h2 for body length was estimated in Rendena cows (0.41; Mazza et al., 2014). The estimated h2 for chest girth in first parity Cika cows was 0.42 ± 0.05, which was only slightly higher than in Holstein (0.38 ± 0.02), American Brown Swiss (0.35 ± 0.02) and Red Holstein cows (0.36 ± 0.02) in Switzerland (de Haas et al., 2007).
The estimated h2 for head length in the first parity Cika cows was 0.48 ± 0.03 (Table 2). A lower h2 for head length was estimated in the Spanish Asturiana de los Valles (0.25 ± 0.02; Gutiérrez and Goyache, 2002), and in Piemontese cows (0.15 ± 0.02; Mantovani et al., 2010). The estimated h2 for the composite autochthonous trait in first parity Cika cows was 0.55 ± 0.05 (Table 2). In Asturiana de los Valles cows, the estimated h2 for a similar composite trait called “breed characteristics” was 0.33 ± 0.02 (Gutiérrez and Goyache, 2002). The estimated h2 for the coat color and coat pattern traits (coat color, back stripe, rear legs stripe, front legs stripe) in the first parity Cika cows were between 0.63 and 0.80.
The estimated h2 for the top line in the first parity Cika cows was 0.19 ± 0.05 (Table 2). A lower h2 for top line was estimated in first parity Asturiana de los Valles cows (0.11 ± 0.01; Gutiérrez and Goyache, 2002), Piemontese cows (0.07 ± 0.01; Mantovani et al., 2010), Brown Swiss cows in Slovenia (0.16; Špehar et al., 2012) and American Brown Swiss cows (0.10; Dal Zotto et al., 2007). Estimated h2 for rump angle in first parity Cika cows was 0.32 ± 0.03 (Table 2). A similar h2 for rump angle was estimated in first parity Rendena cows (0.36; Mazza et al., 2014) and Czech Holstein-Friesian cows (0.34; Němcová et al., 2011). The h2 for rump angle in first parity Brown Swiss cows in Slovenia was 0.22 (Špehar et al., 2012), and 0.24 in American Brown Swiss cows (Dal Zotto et al., 2007). The estimated h2 for rear leg set in the first parity Cika cows was 0.15 ± 0.03 (Table 2). Similar h2 for rear legs set was found in the Rendena cows (0.21; Mazza et al., 2014), Piemontese (0.12 ± 0.02; Mantovani et al., 2010), Slovenian Brown Swiss (0.13; Špehar et al., 2012), American Brown Swiss cattle (0.14; Dal Zotto et al., 2007) and Czech Holstein-Friesian cows (0.16; Němcová et al., 2011). The estimated h2 for hock quality in the first parity Cika cows was 0.61 ± 0.04 (Table 2). A lower h2 for hock quality was estimated in Slovenian Brown Swiss (0.11; Špehar et al., 2012) and American Brown Swiss cows (0.08; Dal Zotto et al., 2007). The estimated h2 for heel height in first parity Cika cows was 0.12 ± 0.02 (Table 2). An equal or very similar h2 for heel height was estimated in Rendena cows (0.12; Mazza et al., 2014), Piemontese (0.09 ± 0.01; Mantovani et al., 2010), Slovenian Brown Swiss (0.04; Špehar et al., 2012), American Brown Swiss (0.09; Dal Zotto et al., 2007) and in Czech Holstein-Friesian cows (0.10; Němcová et al., 2011). Likewise, the estimated h2 for the composite trait body frame in the first parity Cika cows was 0.19 ± 0.03, which was very similar to Rendena cows (0.18; Mazza et al., 2014), an autochthonous small frame cattle breed from north-eastern Italy.
The estimated h2 for udder attachment in the first parity Cika cows was 0.20 ± 0.03 (Table 2), which was lower than in Rendena cows (0.32; Mazza et al., 2014), and similar to Czech Holstein-Friesian (0.24; Němcová et al., 2011) as well as French Holstein cows (0.18; Rupp and Boichard, 1999). The estimated h2 for fore udder attachment was higher than in Slovenian Brown Swiss (0.14; Špehar et al., 2012) and American Brown Swiss cows (0.14; Dal Zotto et al., 2007). The estimated h2 for udder depth in the first parity Cika cows was 0.26 ± 0.03, similar to Rendena cows (0.27; Mazza et al., 2014), Slovenian Brown Swiss (0.22; Špehar et al., 2012), American Brown Swiss (0.23; Dal Zotto et al., 2007), Czech Holstein-Friesian (0.32; Němcová et al., 2011) and French Holstein cows (0.29; Rupp and Boichard, 1999). The estimated h2 for teat thickness in first parity Cika cows (0.34 ± 0.03; Table 2) was similar to Rendena (0.34; Mazza et al., 2014), Slovenian Brown Swiss (0.33; Špehar et al., 2012), American Brown Swiss (0.32; Dal Zotto et al., 2007), Czech Holstein-Friesian (0.28; Němcová et al., 2011) and French Holstein cows (0.30; Rupp and Boichard, 1999). Taking into account estimated composite trait “udder” in the first parity Cika cows (h2 = 0.26 ± 0.03; Table 2), the results were lower than in Rendena cows (0.37; Mazza et al., 2014), and higher than in Slovenian Brown Swiss cows (0.16; Špehar et al., 2012).
The estimated h2 for muscularity in the first parity Cika cows was 0.19 ± 0.05 (Table 3), which was similar to Asturiana de los Valles cows (0.22 ± 0.01; Gutiérrez and Goyache, 2002) and Brown Swiss cows (0.16; Špehar et al., 2012). On the other hand, h2 for muscularity in the first parity Cika cows was lower than in Rendena (0.31; Mazza et al., 2014), American Brown Swiss (0.42 ± 0.02) and Red Holstein cows (0.59 ± 0.03) (de Haas et al., 2007). The huge variability in the heritabilities for the trait muscularity could be due to the recording procedure, the degree of harmonization among the experts, the statistical models, and data quality.
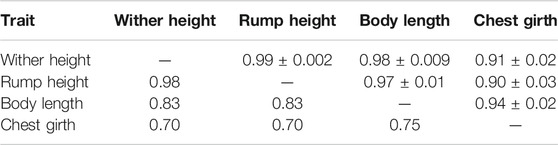
TABLE 3. Estimated genetic correlations ± standard errors (above diagonal) and phenotypic correlations (below diagonal) for measured body frame traits from Group 1.
The proportion of variance in the type traits of the first parity Cika cows explained by the effect of the herd (c2) ranged from 0.01 to 0.28 (Table 3). Interestingly, c2 was on average higher for measured body frame traits (0.10–0.25) compared to the scored body frame traits (0.01–0.23). Likewise, the scored autochthonous and udder traits had a low c2 (0.01–0.11 and 0.06 to 0.13, respectively). The estimated c2 for the composite traits (autochthonous characteristics, muscularity, body frame, udder) were 0.09, 0.28, 0.14, and 0.10, respectively. This might reflect the effect of the huge variability of production systems on the farms that cause variability in the body condition of the animals in the herd. The proportion of residual variance ranged from 0.18 to 0.74. The standard errors of the variance components varied between 0.007 and 0.05.
Genetic and Phenotypic Correlations
The estimated genetic and phenotypic correlations between the measured body frame traits (Table 3) were positive and very high. Based on these high correlations, it would be recommended to reduce the number of measured traits in the scoring form, to make the whole procedure less time consuming. The ICAR guidelines for conformation recording (ICAR, 2018) recommend only rump height (called “stature”) to be scored. Likewise, Mazza et al. (2014) found high genetic (0.79) and phenotypic (above 0.53) correlations between body frame traits in first parity Rendena cows. On the other hand, lower positive genetic correlations between rump height and chest girth were estimated in Holstein (0.45), American Brown Swiss (0.34) and Red Holstein cows (0.54) (de Haas et al., 2007).
The estimated genetic correlations between the traits describing parts of the head (Table 4) were low to moderate, and mostly positive. The only slightly negative genetic correlation was found between head and horn length (−0.05), where SE was 0.06 and, therefore, the genetic correlation was not different from zero. The highest genetically correlated traits were head length and eyes (0.76). The phenotypic correlations were lower than the genetic ones, ranging from 0.09 (horn length - head length, horn length - eyes) to 0.54 (head nobility - head length).

TABLE 4. Estimated genetic correlations ± standard errors (above diagonal) and phenotypic correlations (bellow diagonal) for scored autochthonous traits from Group 2.
The estimated phenotypic and genetic correlation between the neck and dewlap (Table 5) was positive and moderate (0.53). Animals with thin skin on the neck had a less expressed dewlap and vice versa, which was expected.
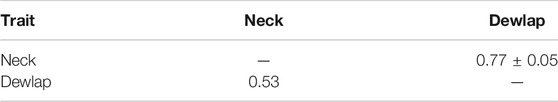
TABLE 5. Estimated genetic correlation ±standard error (above diagonal) and phenotypic correlation (bellow diagonal) for scored autochthonous traits from Group 3.
The estimated genetic and phenotypic correlations between the scored traits describing the coat color and white stripes patterns (Table 6) were positive, very low one the hand or very high on the other. The coat color was weakly correlated with all traits describing white stripe patterns, whereas the back stripe, the rear legs stripe, and the front legs stripe were highly correlated with each other. Likewise, the genetic correlation between the coat color and the front legs stripe (0.02) had a SE of 0.03 and, therefore, was not different from zero. First parity Cika cows with a wider white back stripe had wider white strips on the rear and front legs. Cows with a wider white stripe on the rear legs had a wider white stripe on the front legs as well.
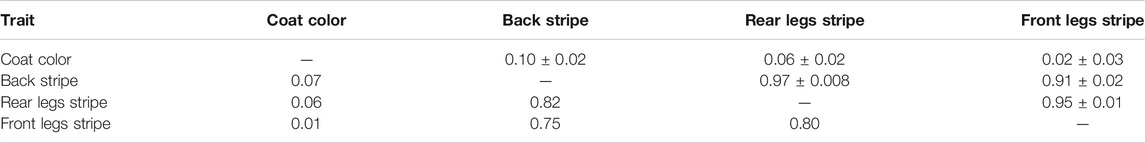
TABLE 6. Estimated genetic correlations ± standard errors (above diagonal) and phenotypic correlations (bellow diagonal) for scored autochthonous traits from Group 4.
The majority of the estimated correlations among the body frame traits (Table 7) were low to moderate. The lowest genetic correlation was found between rump angle and hock quality (−0.03), while the highest was between heel height and foot angle (0.89). However, the genetic correlation between rump angle and hock quality (−0.03) had a SE of 0.06 and was not different from zero. The highest phenotypic correlation (0.62) was also found between heel height and foot angle, whereas the lowest phenotypic correlation (−0.001) was between rear leg set and rump angle. Animals with a low foot angle had low heel height, while animals with a steep foot angle had high heel height. Moderate genetic correlations were estimated between the composite trait body frame and heel height as well as with foot angle (both 0.58). The composite trait body frame had the highest negative correlation with rear leg set (−0.59). Cows with high scores for the composite trait of body frame had relatively steep foot angles and high heel height, and a steep rear leg set. Špehar et al. (2012) estimated a similar genetic correlation between rear leg set and heel height (−0.33) in Slovenian Brown Swiss cows. Němcová et al. (2011) estimated lower genetic and the same phenotypic correlation between rump angle and heel height (−0.06; −0.04) in Czech Holstein-Friesian cows.
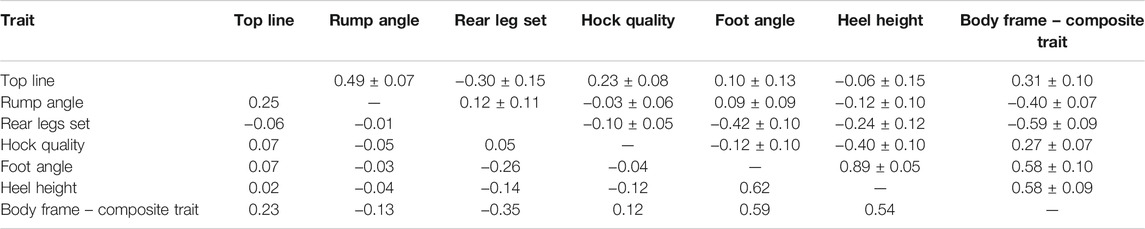
TABLE 7. Estimated genetic correlations ± standard errors (above diagonal) and phenotypic correlations (bellow diagonal) for scored and composite body frame traits from Group 5.
The estimated phenotypic correlations between udder traits (Table 8) were moderate and positive (up to 0.79 between fore udder attachment and the composite udder trait), and weak to moderate negative (up to −0.39 for front teat length and udder depth). Some estimated genetic correlations were high and positive, with the highest correlation between udder depth and the composite trait “udder” (0.94). The others were moderate and negative (e.g. −0.74 between udder depth and front teat length). The lowest genetic correlation was estimated between front teat length and fore udder attachment (−0.45). A functional udder should be extended under the abdomen, well attached, with thin and short teats.
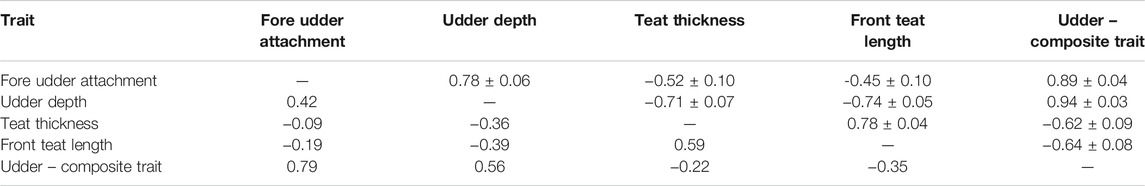
TABLE 8. Estimated genetic correlations ± standard errors (above diagonal) and phenotypic correlations (bellow diagonal) for scored udder traits and the composite trait udder from Group 6.
Fore udder attachment and udder depth were relatively strongly correlated (0.78), as well as front teat length and teat thickness (0.78). Cows with a genetic predisposition for weakly attached udders usually have deeper udders as well. Němcová et al. (2011) estimated higher phenotypic and lower genetic correlations between fore udder attachment and udder depth (0.44; 0.75) in Czech Holstein-Friesian cows. Mazza et al. (2014) estimated lower phenotypic and genetic correlations between fore udder attachment and the composite trait “udder” (0.68; 0.78) in Rendena cows.
Genetic Trends
All 26 individual type traits as well as the four composite traits were already introduced in the genetic evaluation of first parity Cika cows in the year 2016. The composite traits, which include all individual traits, are widely used. There is a plan to decrease the number of traits in the future since some of the traits within each group are highly correlated. Nevertheless, Figure 1 includes the genetic trends of all 26 individual type traits as well as the four composite traits for first parity Cika cows born from 2000 to 2019. The genetic trends of all four measured body frame traits are negative, which is desirable and expected since the breeders prefer Cika animals with a smaller body frame that were not admixed with Pinzgauer in the past. Moreover, potential Cika sires are genotyped each year to select those from the group suitable for artificial insemination that do not have genes of Pinzgauer or other foreign breeds. The autochthonous characteristics of the cows are represented by 12 individual traits divided into traits of the head, neck and coat color as well as coat color patterns. The genetic trends for the traits of the head are all positive. First parity Cika cows born in the last years had shorter and finer heads with larger eyes and thinner, shorter horns with a more forward direction compared to Cika cows born before 2010, which is in accordance with the breeding goals of the breeding program. A very similar situation is reflected by the positive genetic trends of the neck and dewlap, which became finer in younger cows. Likewise, heavier necks and dewlaps are typical for Cika cows that were admixed with Pinzgauer in the past. According to the genetic trends for coat color and its patterns, the coat color is becoming brighter, while white stripes on the back as well as on the rear and front legs have not changed very much in animals born during the last years. Among the scored body frame traits, only hock quality had a positive genetic trend, while rump angle, foot angle and heel height had negative genetic trends. Consequently, cows born in the last years have more sloping rumps, as well as cleaner and drier hocks with steeper angles of the foot and taller heels. In the past, a lot of cows had high pins and hocks with a lot of fluid, which was very undesirable for the breeders, i.e. the genetic trends are in accordance with the breeding goals now. Among the scored udder traits, only fore udder attachment and udder depth had slightly positive trends, which means that cows have more strongly attached and shallower udders than in the past. Finally, three (autochthonous characteristics, body frame, udder) of four composite traits had positive genetic trends. All in all, first parity Cika cows born in the last years showed more typical autochthonous type traits of purebred Cika cows, which distinguished them from Cika cows admixed with Pinzgauer in the past. Likewise, first parity Cika cows nowadays have more excellent body frames and udders.
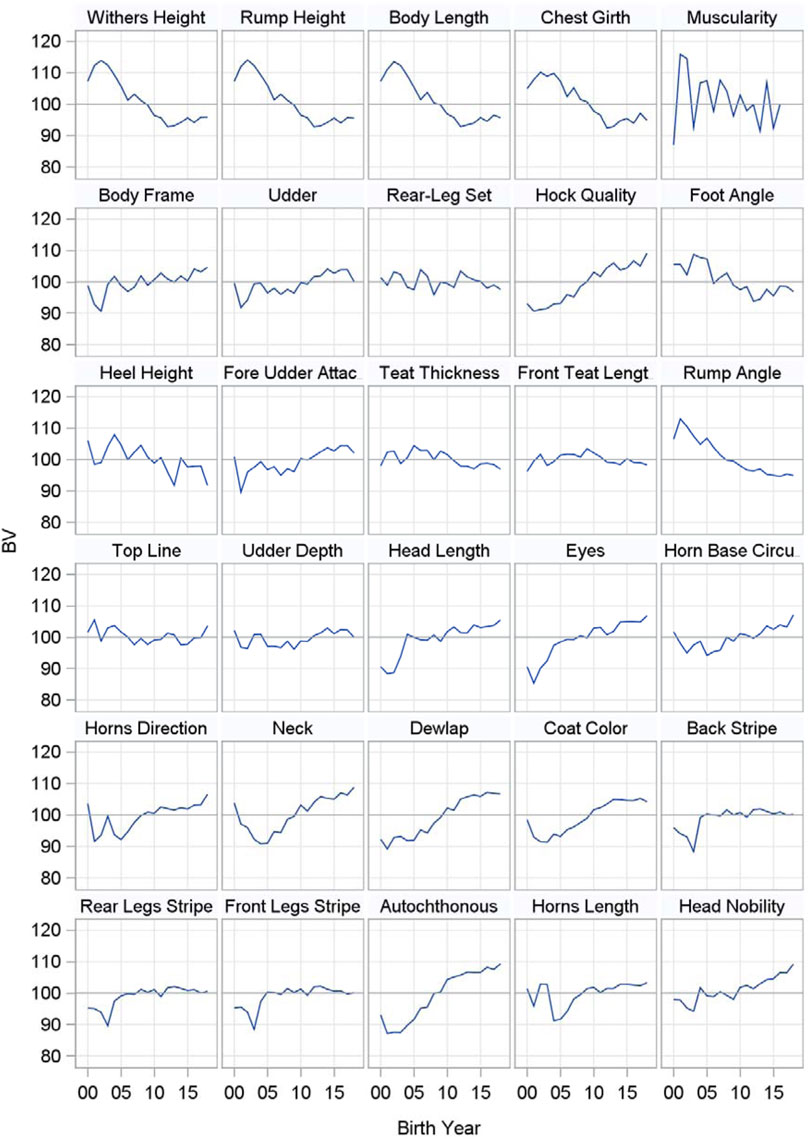
FIGURE 1. Genetic trends for type traits in first parity Cika cows born in the years 2000–2019.
Conclusion
Comparing the estimated heritabilities for the majority of included type traits with heritabilities from the literature, it was found that they were within the expected range and similar to those reported for other breeds. Among the measured body frame traits, the highest heritability was estimated for rump height. The heritabilites of scored body frame traits in Cika were similar to those of other breeds as well. In the scoring form for Cika, only four scored udder traits are included (fore udder attachment, udder depth, teat thickness, front teat length). Nevertheless, their heritabilities were very similar to those reported for cows of dairy breeds. The estimated heritabilities of the scored autochthonous traits, which describe the breed characteristics, were moderate to high. This suggests the possibility to breed first parity Cika cows with unique type traits known for autochthonous Cika cattle. Unfortunately, according to the best of our knowledge, no autochthonous type traits like those included in the analysis have been investigated before. Consequently, comparison with the literature was not possible.
All above estimated genetic parameters are already used for breeding value prediction in the Cika cattle population. According to the genetic trends, first parity Cika cows born in the last years showed more typical autochthonous type traits of purebred Cika cows, which distinguished them from Cika cows admixed with Pinzgauer in the past. Likewise, first parity Cika cows nowadays have more excellent body frames and udders.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
MS wrote the manuscript and gave the main idea, BL wrote the manuscript and prepared a plan of analysis, MŠ prepared all analysis, BL prepared the data, KP checked the analysis and manuscript in detail.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Comunod, S., Guida, S., Vigo, D., Beretti, V., Munari, E., Colombani, C., et al. (2013). Body Measures and Milk Production, Milk Fat Globules Granulometry and Milk Fatty Acid Content in Cabanina Cattle Breed. Ital. J. Anim. Sci. 12 (1), e18. doi:10.4081/ijas.2013.e18
Dal Zotto, R., De Marchi, M., Dalvit, C., Cassandro, M., Gallo, L., Carnier, P., et al. (2007). Heritabilities and Genetic Correlations of Body Condition Score and Calving Interval with Yield, Somatic Cell Score, and Linear Type Traits in Brown Swiss Cattle. J. Dairy Sci. 90, 5737–5743. doi:10.3168/jds.2007-0280
de Haas, Y., Janss, L. L. G., and Kadarmideen, H. N. (2007). Genetic and Phenotypic Parameters for Conformation and Yield Traits in Three Swiss Dairy Cattle Breeds. J. Anim. Breed. Genet. 124, 12–19. doi:10.1111/j.1439-0388.2007.00630.x
Felius, M., Koolmees, P. A., Theunissen, B., and Lenstra, J. A. (2011). On the Breeds of Cattle-Historic and Current Classifications. Diversity 3, 660–692. doi:10.3390/d3040660
Forabosco, F., Groen, A. F., Bozzi, R., Van Arendonk, J. A. M., Filippini, F., Boettcher, P., et al. (2004). Phenotypic Relationships between Longevity, Type Traits, and Production in Chianina Beef Cattle1. J. Anim. Sci. 82, 1572–1580. doi:10.2527/2004.8261572x
Frigo, E., Samorè, A. B., Vicario, D., Bagnato, A., and Pedron, O. (2013). Heritabilities and Genetic Correlations of Body Condition Score and Muscularity with Productive Traits and Their Trend Functions in Italian Simmental Cattle. Ital. J. Anim. Sci. 12, e40. doi:10.4081/ijas.2013.e40
Gengler, N., Seutin, C., Boonen, F., and Van Vleck, L. D. (1995). Estimation of Genetic Parameters for Growth, Feed Consumption, and Conformation Traits for Double-Muscled Belgian Blue Bulls Performance-Tested in Belgium. J. Anim. Sci. 73, 3269–3273. doi:10.2527/1995.73113269x
Groeneveld, E., Kovač, M., and Mielenz, N. (2010). VCE User’s Guide and Reference Manual Version 6.0. Neustadt, Germany: Institute of Farm Animal Genetics. Available at: https://www.pdffiller.com/303612605-vce6-manual-31-A4pdf-VCE-bUser39s-Guideb-and-Reference-bManualb-Version-60-ftp-tzv-fal-.
Gutiérrez, J. P., and Goyache, F. (2002). Estimation of genetic parameters of type traits in Asturiana de los Valles beef cattle breed. J. Anim. Breed. Genet. 119, 93–100. doi:10.1046/j.1439-0388.2002.00324.x
ICAR (2018). ICAR Guidelines for Conformation Recording of Dairy Cattle, Beef Cattle, Dual Purpose Cattle and Dairy Goats – Section 5. New Delhi: ICAR – International Committee for Animal Recording, 76. Available at: https://www.icar.org/Guidelines/05-Conformation-Recording.pdf.
Larroque, H., and Ducrocq, V. (2001). Relationships between Type and Longevity in the Holstein Breed. Genet. Selection Evol. 33, 39–59. doi:10.1186/1297-9686-33-1-39
Mancin, E., Sartori, C., Guzzo, N., Tuliozi, B., and Mantovani, R. (2021). Selection Response Due to Different Combination of Antagonistic Milk, Beef, and Morphological Traits in the Alpine Grey Cattle Breed. Animals 11, 1340. doi:10.3390/ani11051340
Mantovani, R., Cassandro, M., Contiero, B., Albera, A., and Bittante, G. (2010). Genetic Evaluation of Type Traits in Hypertrophic Piemontese Cows. J. Anim. Sci. 88, 3504–3512. doi:10.2527/jas.2009-2667
Mazza, S., Guzzo, N., Sartori, C., Berry, D. P., and Mantovani, R. (2014). Genetic Parameters for Linear Type Traits in the Rendena Dual-Purpose Breed. J. Anim. Breed. Genet. 131 (1), 27–35. doi:10.1111/jbg.12049
Mazza, S., and Mantovani, R. (2012). Heritability of Linear Type Traits in the Autochthonous Rendena Dual Purpose Breed. Acta Agric. Slov., Suppl. 3, 161–165.
Mazza, S., Sartori, C., and Mantovani, R. (2015). Genetic Parameters of Type Traits in Two Strains of Dual Purpose Autochthonous Valdostana Cattle. Livestock Sci. 178, 35–42. doi:10.1016/j.livsci.2015.06.008
Němcová, E., Štrípková, M., and Zavadilová, L. (2011). Genetic Parameters for Linear Type Traits in Czech Holstein Cattle. Czech J. Anim. Sci. 56 (4), 157–162.
Neuenschwander, T., Kadarmideen, H. N., Wegmann, S., and de Haas, Y. (2005). Genetics of Parity-Dependant Production Increase and its Relationship with Health, Fertility, Longevity, and Conformation in Swiss Holsteins. J. Dairy Sci. 88, 1540–1551. doi:10.3168/jds.s0022-0302(05)72823-x
Norman, H. D., Powell, R. L., Wright, J. R., and Cassell, B. G. (1988). Phenotypic and Genetic Relationship between Linear Functional Type Traits and Milk Yield for Five Breeds. J. Dairy Sci. 71, 1880–1896. doi:10.3168/jds.s0022-0302(88)79758-1
Rogers, G. W., Hargrove, G. L., Cooper, J. B., and Barton, E. P. (1991). Relationships Among Survival and Linear Type Traits in Jerseys. J. Dairy Sci. 74, 286–291. doi:10.3168/jds.s0022-0302(91)78172-1
Rupp, R., and Boichard, D. (1999). Genetic Parameters for Clinical Mastitis, Somatic Cell Score, Production, Udder Type Traits, and Milking Ease in First Lactation Holsteins. J. Dairy Sci. 82, 2198–2204. doi:10.3168/jds.s0022-0302(99)75465-2
Sambraus, H. H. (1999). Gefährdete Nutztierrassen Ihre Zuchtgeschichte, Nutzung und Bewahrung [Endangered breeds, their breeding history, use and preservation]. Ulmer, Stuttgart.
Simčič, M., Lenstra, J. A., Baumung, R., Dovč, P., Čepon, M., and Kompan, D. (2013). On the Origin of the Slovenian Cika Cattle. J. Anim. Breed. Genet. 130, 487–495. doi:10.1111/jbg.12034
Simčič, M., Smetko, A., Sölkner, J., Seichter, D., Gorjanc, G., Kompan, D., et al. (2015a). Recovery of Native Genetic Background in Admixed Populations Using Haplotypes, Phenotypes, and Pedigree Information-Uusing Cika Cattle as a Case Breed. Plos One 10 (4), e0123253. doi:10.1371/journal.pone.0123253
Simčič, M., Sölkner, J., Kompan, D., and Medugorac, I. (2015b). “Exclusion of Admixed Animals from Cika Cattle In Situ Conservation Program Using SNP Haplotypes,” in Book of Abstracts of the 66th Annual Meeting of the European Federation of Animal Science (Wageningen,Netherlands: Wageningen Academic Publishers), 101.
Simčič, M., Štepec, M., Logar, B., Potočnik, K., and Terčič, D. (2016). Analysis of Type Traits of Cika First Calving Cows. Acta Agric. Slov. 108 (2), 93–102. doi:10.14720/aas.2016.108.2.4
Špehar, M., Štepec, M., and Potočnik, K. (2012). Variance Components Estimation for Type Traits in Slovenian Brown Swiss Cattle. Acta Agric. Slov. 100 (2), 107–115.
Thomas, C. L., Vinson, W. E., Pearson, R. E., and Norman, H. D. (1985). Components of Genetic Variance and Covariance for Linear Type Traits in Jersey Cattle. J. Dairy Sci. 68, 2989–2994. doi:10.3168/jds.s0022-0302(85)81194-2
Van Dorp, T. E., Dekkers, J. C. M., Martin, S. W., and Noordhuizen, J. P. T. M. (1998). Genetic Parameters of Health Disorders, and Relationships with 305-day Milk Yield and Conformation Traits of Registered Holstein Cows. J. Dairy Sci. 81, 2264–2270. doi:10.3168/jds.s0022-0302(98)75806-0
Keywords: Cika cattle, first parity cow, type traits, genetic parameters, variance components
Citation: Simčič M, Luštrek B, Štepec M, Logar B and Potočnik K (2021) Estimation of Genetic Parameters of Type Traits in First Parity Cows of the Autochthonous Cika Cattle in Slovenia. Front. Genet. 12:724058. doi: 10.3389/fgene.2021.724058
Received: 11 June 2021; Accepted: 27 October 2021;
Published: 22 November 2021.
Edited by:
Mohammed Ali Al Abri, Sultan Qaboos University, OmanReviewed by:
Hugo Oswaldo Toledo-Alvarado, National Autonomous University of Mexico, MexicoCristina Sartori, University of Padua, Italy
Copyright © 2021 Simčič, Luštrek, Štepec, Logar and Potočnik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mojca Simčič, bW9qY2Euc2ltY2ljQGJmLnVuaS1sai5zaQ==