Corrigendum: poreCov - An Easy to Use, Fast, and Robust Workflow for SARS CoV-2 Genome Reconstruction via Nanopore Sequencing
 Christian Brandt1*
Christian Brandt1* Sebastian Krautwurst2
Sebastian Krautwurst2 Riccardo Spott1Mara Lohde1Mateusz Jundzill1Mike Marquet1Martin Hölzer3
Riccardo Spott1Mara Lohde1Mateusz Jundzill1Mike Marquet1Martin Hölzer3- 1Institute for Infectious Diseases and Infection Control, Jena University Hospital, Jena, Germany
- 2RNA Bioinformatics and High-Throughput Analysis, Friedrich Schiller University Jena, Jena, Germany
- 3Methodology and Research Infrastructure, MF1 Bioinformatics, Robert Koch Institute, Berlin, Germany
In response to the SARS-CoV-2 pandemic, a highly increased sequencing effort has been established worldwide to track and trace ongoing viral evolution. Technologies, such as nanopore sequencing via the ARTIC protocol are used to reliably generate genomes from raw sequencing data as a crucial base for molecular surveillance. However, for many labs that perform SARS-CoV-2 sequencing, bioinformatics is still a major bottleneck, especially if hundreds of samples need to be processed in a recurring fashion. Pipelines developed for short-read data cannot be applied to nanopore data. Therefore, specific long-read tools and parameter settings need to be orchestrated to enable accurate genotyping and robust reference-based genome reconstruction of SARS-CoV-2 genomes from nanopore data. Here we present poreCov, a highly parallel workflow written in Nextflow, using containers to wrap all the tools necessary for a routine SARS-CoV-2 sequencing lab into one program. The ease of installation, combined with concise summary reports that clearly highlight all relevant information, enables rapid and reliable analysis of hundreds of SARS-CoV-2 raw sequence data sets or genomes. poreCov is freely available on GitHub under the GNUv3 license: github.com/replikation/poreCov.
Introduction
It has been over a year since the WHO declared a pandemic related to SARS-CoV-2 on March 11, 2020. To date, this pandemic has caused over 112 million registered cases and more than 2.5 million deaths (March 2021). Over 190 countries have contributed to sequencing over 1.1 million publicly available SARS-CoV-2 genomes during this time, resulting in one of the greatest molecular surveillance efforts to date (data from GISAID; April 2021) (Elbe and Buckland-Merrett, 2017; Shu and McCauley, 2017). Thanks to global sequencing efforts, it is possible to track emerging variants, trace the evolution of the virus (Hodcroft et al., 2020; Brandt et al., 2021; Tang et al., 2021), and investigate potential escape mechanisms against vaccines at an early stage (Thompson et al., 2021; Zhou et al., 2021). In addition, the large amount of genomic data generated enables the development of simpler and faster rapid tests for the detection of variants of concern and characteristic mutations, e.g., for routine PCR screening in hospitals (Durner et al., 2021).
Illumina is still the most dominant approach for SARS-CoV-2 sequencing and genome reconstruction (Table 1). However, nanopore sequencing via tiled amplicon-based genome reconstruction (Quick et al., 2017; Yan et al., 2021) is also heavily used and of particular interest, as laboratories can enter sequencing with a comparably low initial investment. While many laboratories can implement nanopore sequencing techniques rapidly, bioinformatics know-how, on the other hand, is still very scarce in science (Hodcroft et al., 2021). The observed shortage of bioinformaticians can be explained by the necessary overlap in different areas of biology, bioinformatics, and computer science. For the latter, in particular, there is an enormous shortage of skilled workers, making the development of robust and widely applicable software tools necessary (Perkel, 2019). This bottleneck in bioinformatics expertise concerns accurate genotyping and subsequent genome reconstruction of SARS-CoV-2 with direct impact to accompanying downstream analyses, such as raw data and genome quality controlling, virus lineage determination, vaccine target surveillance, phylogenomics, and dynamic evolutionary observations, and the various metrics behind them (Brandt et al., 2021; Hodcroft et al., 2021; Hufsky et al., 2021). Bioinformatics tools or workflows for similar tasks (e.g., reference-based genome reconstruction) are constantly being reinvented and developed independently by many laboratories. A broad application combined with user-friendly handling is rarely in focus. One reason for that is the additional development time and necessary maintenance. However, the usage of workflow management systems and containerization also allow smaller labs to provide reproducibility and, due to modularization, easy to maintain software tools (Perkel, 2019). A few workflows targeting the analysis of SARS-CoV-2 amplicon-based nanopore sequencing data already exist (e.g., github.com/nf-core/viralrecon, github.com/connor-lab/ncov2019-artic-nf), which are primarily aimed at a bioinformatics audience, not intended for public use or are over-complicated for biologists to use due to high level of necessary parameterization. While such features enable important workflow flexibility to support different sequencing protocols, they can be challenging to apply for a non-expert audience. However, due to the high sequencing workload during the pandemic, many laboratories urgently require bioinformatics support. We developed poreCov to diminish the bioinformatic entry barriers as much as possible and to allow reliable genotyping and reference-based genome reconstruction of SARS-CoV-2 samples in a high-throughput manner. While poreCov is optimized to analyze amplicon-based nanopore sequencing data following the widely used ARTIC protocol, the pipeline can also be applied to other primer sets and protocols. Reference-based genome reconstruction is the central focus of our pipeline, complemented by a group of downstream analyses for virus lineage determination and a comprehensive summary of results.
TABLE 1
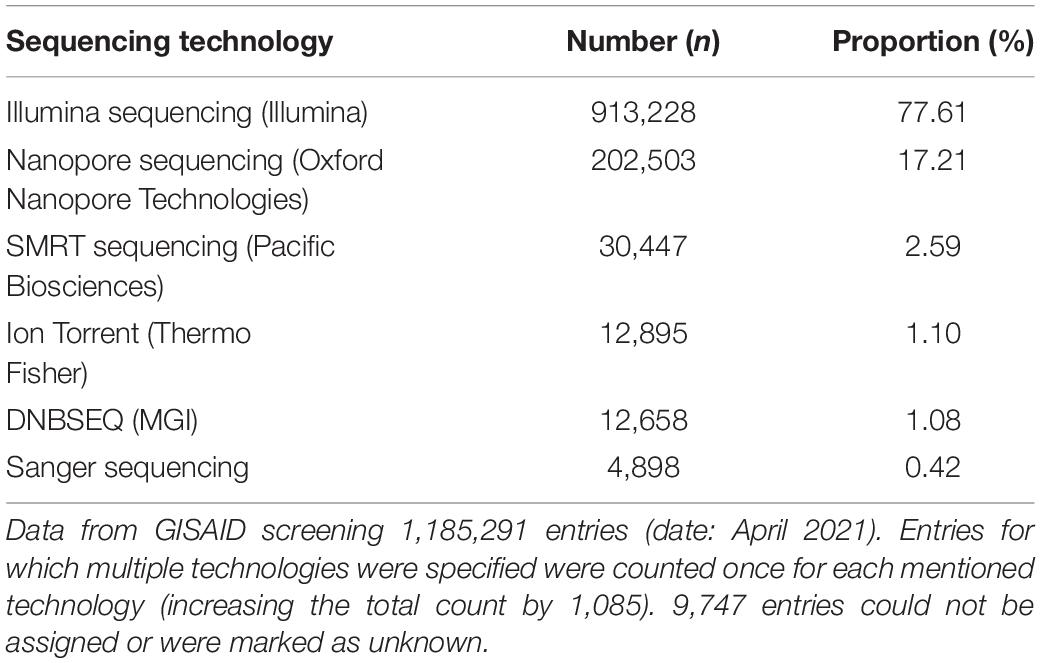
Table 1. Sequencing technology used for SARS-CoV-2 surveillance.
Materials and Methods
General Design
One of the main aspects of poreCov is to keep the bioinformatics part as simplistic as possible while still comprehensive and robust. Our development focused on easy installation, configuration, and execution on various computer systems with clear results and a minimal set of necessary input parameters. We developed customized scripts to collect and visualize all relevant results in a comprehensive summary report that comes as an intuitive HTML report as well as an Excel sheet (XLSX) and a tab-separated table (TSV) for direct programmatic access. To start an analysis with poreCov, only the bare necessity of inputs is required (where is the data located, what kind of input data is it, which primers are used). Other parameters are set to recommended default values but can still be changed by experienced users if necessary.
We use Nextflow (Di Tommaso et al., 2017) as a workflow manager giving poreCov the capabilities to be executed on most computational infrastructures from a laptop up to a compute cluster and cloud computing solutions. The native container support (Docker, Singularity) allows for comfortable, stable, and install-free execution of the various included tools without software dependency conflicts. Our containers are prebuilt, stored on docker hub,1 and automatically downloaded by poreCov if not already available on the system. This leaves Nextflow and either Docker or Singularity as the only dependencies needed to be installed for poreCov. If these dependencies are available, an initial installation and further updates of the workflow and the included tools can be performed with a single simple command (“nextflow pull replikation/poreCov”). When poreCov runs on a high-performance computing (HPC) cluster or cloud services, all resources (CPUs, RAM) for all processes and associated containers are pre-configured but can be adjusted via a user-specific configuration file when needed.
We use complete version control for poreCov from the workflow itself to every tool to achieve reproducible results. For this purpose, each of the more than 15 containers used is version-controlled based on their associated tool. Besides, each poreCov version can be accessed and executed individually, and the tool versions used during genome reconstruction and analysis are listed in the final report.
Inputs
poreCov automatically adjusts the necessary workflow steps based on the provided input. As a result, a more flexible workflow execution is possible depending on the available data (Figure 1). As each sequencing lab has its own way for processing nanopore sequencing data, we allow for three different sequence inputs to start the genome reconstruction and variant calling steps: (1) a directory containing fast5 raw files (not basecalled), (2) an already basecalled directory containing fastq sequence files (e.g., the output from a MinIT or GridION). Additionally, (3) combined, single fastq file (one fastq file per sample) can be provided to simplify further the reanalysis of read data sets (e.g., from public repositories like ENA). Moreover, the genome reconstruction step is automatically skipped if already reconstructed genomes (fasta files) are provided as a direct input. By that, poreCov also allows to quickly and conveniently re-analyze previous genomes, e.g., to check for new clade or lineage assignments or to simply generate quality metrics and a summary report for a set of SARS-CoV-2 genomes. Optionally, a sample sheet can be provided to rename barcodes automatically and to simplify data analysis and file management post-analysis.
FIGURE 1
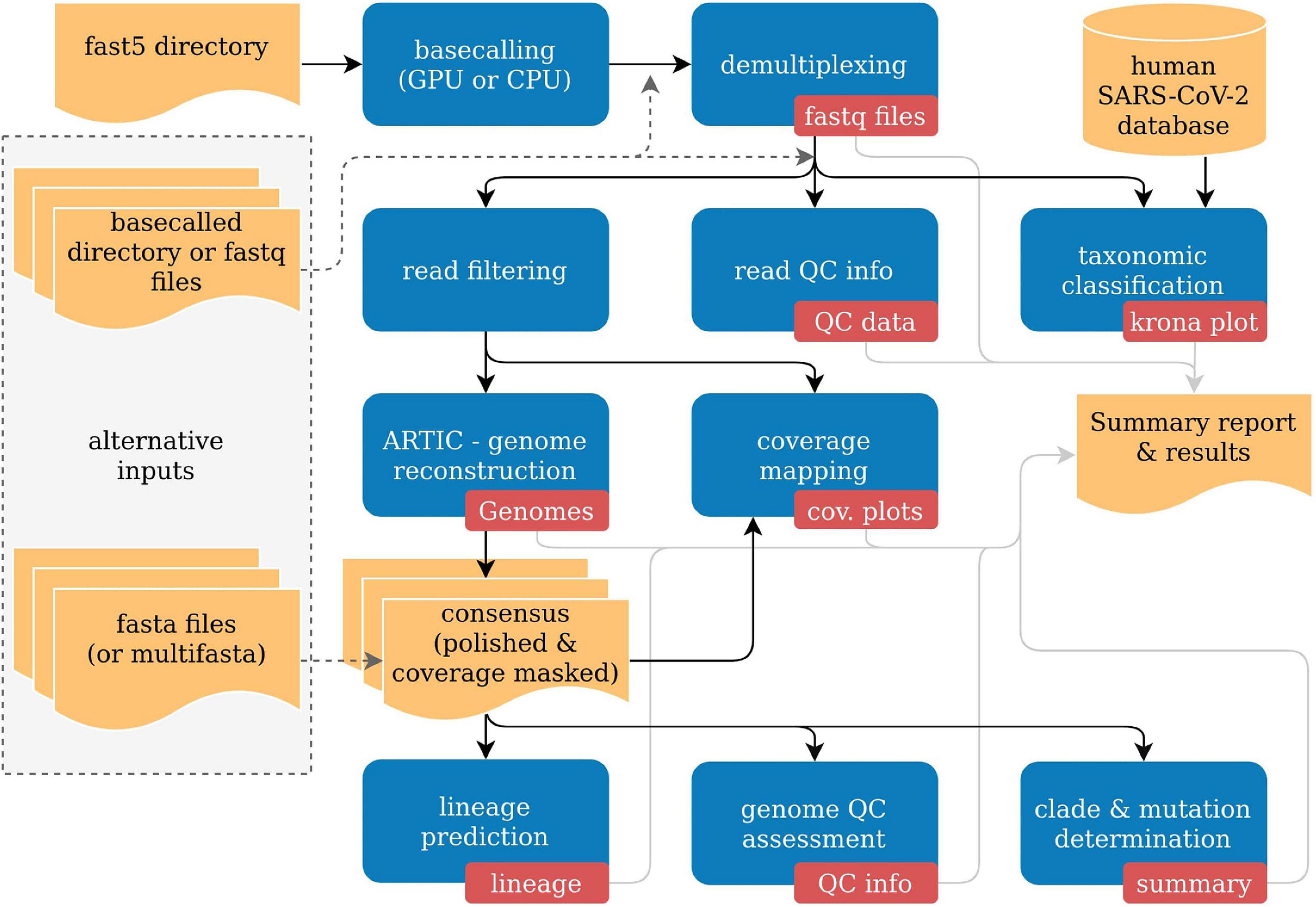
Figure 1. Simplified overview of the poreCov workflow. Results and inputs are colored in yellow. Workflow processes are colored blue, and the resulting information of a process is red.
Included Analysis Steps and Tools
Read Processing and Quality Control
For the nanopore raw data (fast5) as an input, poreCov provides some necessary flexibility as the GPU basecaller Guppy (Oxford Nanopore Technologies) and the dependencies can be difficult to install. By default, poreCov assumes that either Docker or Singularity with GPU support can be used. However, users can switch to a local installation of Guppy or CPU-based basecalling or skip basecalling automatically by providing already basecalled reads (fastq). All reads are taxonomically classified via Kraken2 (Wood et al., 2019) against a pre-computed database comprised of the human GRCH38.p13 genome and all GISAID SARS-CoV-2 genomes as of February 2021 (available at doi.org/10.5281/zenodo.4534746) to unveil possible contamination or isolation issues from the wet lab. Krona plots (Ondov et al., 2011) are used to visualize the results. Reads are quality-controlled (read length distribution, quality scores, etc.) either via NanoPlot (De Coster et al., 2018) for fastq files or pycoQC (Leger and Leonardi, 2019) for fast5 files. All reads are prefiltered based on their length (default: 400–700 bp for version 3 of the ARTIC primers) and these reads are mapped back against the Wuhan reference genome (Accession: NC_045512.2) using minimap2 (Li, 2018) to provide visual feedback of amplicon dropouts for each sample, which are a common issue during the multiplex PCR step of the tiled amplicon sequencing protocol.
SARS-CoV-2 Genome Reconstruction
All Reads that passed the length filtering are passed to the genome reconstruction via the ARTIC pipeline.2 The ARTIC pipeline comprises tools and custom scripts for working with viral nanopore sequencing data generated from tiling amplicon schemes. General mappings are performed with minimap2 (Li, 2018), and intermediate file transformations are performed with SAMtools and BCFtools (Danecek et al., 2021). By default, genotyping is performed via medaka3 and Longshot (Edge and Bansal, 2019) but can also be done via nanopolish4 if fast5 data is provided. Regions with a low read coverage of 20× or less are masked by default.
SARS-CoV-2 Genome Analysis
After the genome reconstruction via ARTIC, each reconstructed and polished genome is further analyzed via pangolin (lineage determination5), nextstrain (nextstrain clades, mutations, and deletions) (Hadfield et al., 2018), and president (calculating various genome quality metrics6). Finally, all results are summarized in an HTML, TSV, and XLSX report to allow fast detection of low-performing samples, evaluation of negative controls, and an overview of virus lineages and detected mutations.
Runtime and Resources
The execution of poreCov requires only a lightweight computing device with four threads (two CPUs) and 8 GB of RAM. However, the analysis will take longer for lower configurations. poreCov shows its strengths on larger workstations or HPCs with full parallelization through the used workflow management system. By default, poreCov utilizes all available threads and tries to run at least four jobs in parallel for the analysis. If necessary, this behavior can be individually controlled and fine-tuned via the cores option flag (Table 2).
TABLE 2

Table 2. poreCov time to results (runtime of the workflow starting from fastq files to the final report) for different hardware configurations using 10 read files as input (fastq files).
Results and Discussion
Result Reporting
Quick and easy-to-understand result interpretation is a crucial challenge for labs that want to analyze hundreds of SARS-CoV-2 genomes or more in short periods. To achieve this, poreCov summarizes all the necessary results besides the reconstructed genomes in a final report file (see Figure 2).
FIGURE 2
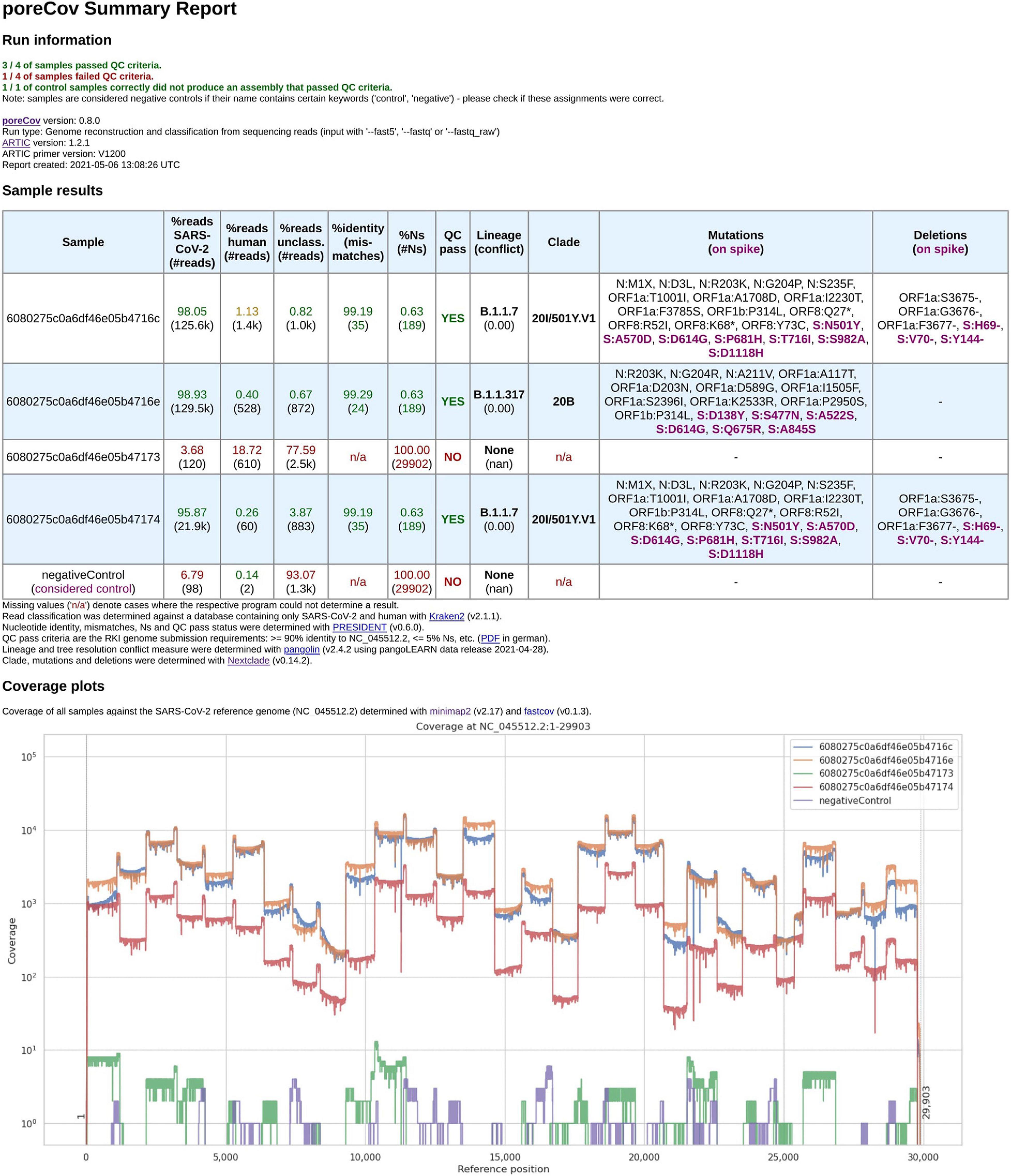
Figure 2. Final report generated by poreCov after each run. Four samples and one negative control were analyzed using 1,200 bp amplicon primers. The first part of the report, the run information, summarizes the overall performance of the sequencing run in a short form across all samples. The second section, the sample results, summarizes all relevant information for each sample in tabular form. The coverage plots are presented in batches of six samples, and additional charts are automatically added for larger runs. The negative control contained 98 reads classified as SARS-CoV-2 which might be attributed to lab contamination or remaining SARS-CoV-2 primers that were sequenced.
The first part of the report is a table containing the relevant information regarding contaminations (amount of human, viral and unclassified reads), the overall quality of the genomes (percent identity to the Wuhan genome, amount of ambiguous nucleotides). All these features are summarized into “QC pass” if genomes have ≥ 90% identity to the Wuhan strain (NC_045512.2) and ≤ 5% ambiguous nucleotides (Ns). The last columns of the table highlight the determined lineages based on the pangolin and nextstrain nomenclatures and include information about substitutions and deletions based on the nextclade alignments (Hadfield et al., 2018). Any substitutions or deletions in the spike protein are highlighted. Furthermore, clear color codings (green, yellow, red) are used to quickly allow the user to visually screen for issues or problems that might have arisen throughout the genome reconstruction and analysis steps. The last part of the report are coverage plots shown in batches of six samples to rapidly assess why a certain sample did not yield a genome that meets the QC criteria or in which regions the coverage was low (e.g., amplicon dropouts due to primer dimerization; Itokawa et al., 2020). Here, we use the length-filtered reads mapped to the reconstructed consensus sequences (minimap2) to estimate how well the genomes are covered by the reads. In addition, we also implemented CoVarPlot (Ferguson et al., 2021) to provide more concise amplicon coverage plots per sample. Here, we directly use the BAM files that are later on also used for variant calling as an input to visualize amplicon coverage based on the length-filtered and primer-clipped reads mapped to the reference genome.
So far, poreCov was used and evaluated on more than 600 SARS-CoV-2-PCR-positive samples analyzed at the Jena University Hospital, Thuringia (Germany) and over 400 samples at the University Hospital Regensburg, Bavaria (Germany). In addition, the pipeline is routinely used for genome reconstruction at Germany’s Public Health Institute and analyzed between February 23 and May 5, 2021, 1,012 samples sequenced on MinION or GridION machines.
SARS-CoV-2 Genomic Surveillance via Nanopore Sequencing: Challenges and Pitfalls
During our work on the poreCov pipeline, we identified several common challenges and pitfalls that arise, particularly during library preparation and sequencing, which can significantly impact downstream bioinformatics analyses. Here, we share such common issues in the context of SARS-CoV-2 nanopore sequencing and how poreCov can help to identify and subsequently avoid these.
Sample-Barcode Bleeding
As nanopore sequencing is a molecule-based approach, it is easy to “over- or underload” cDNA molecules for library preparation. This overloading often happens if the majority (>50%) of all fragments are “super small” (<100 nucleotides) in the sample. Molecule-overloading usually leads to unspecific barcode ligation during the adapter ligation step after barcode pooling. Free barcodes from samples with a low amount of DNA molecules might ligate to non-barcoded DNA from samples with a highly overloaded number of molecules. A negative control can visualize this issue (poreCov highlights negative controls in the report if specified). Furthermore, using only reads with barcodes present on both ends mitigates the problem (the default setting for poreCov). If sample bleeding occurred, a false-positive negative control usually inherits a similar SNP proportion pattern as the barcoded samples used on the same run (see Figure 3). This can be visually inspected by opening the .bam file in the result dir “2.Genomes” via, e.g., Unipro UGENE (Okonechnikov et al., 2012) or IGV viewer (Robinson et al., 2011).
FIGURE 3
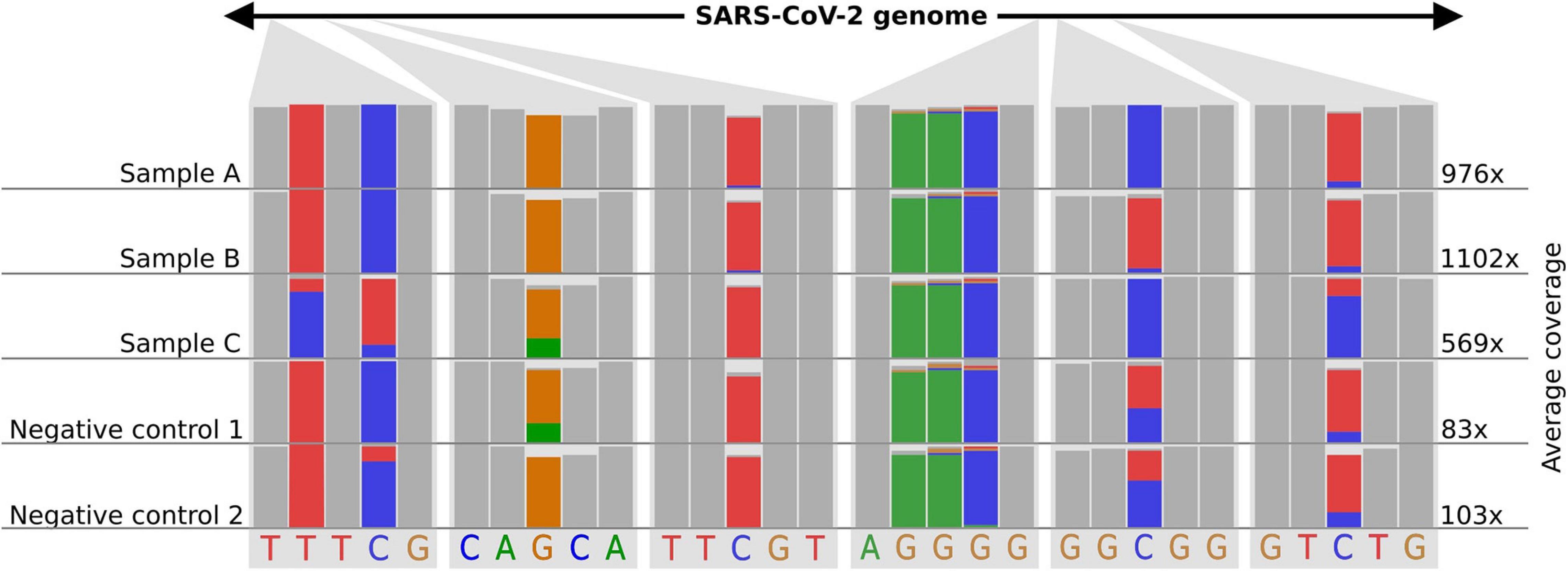
Figure 3. Example of how sample-barcode bleeding can be identified by inspecting SNP proportions of false-positive negative controls in mapping (.bam) files. Three samples and two negative controls are shown. Each column shows the proportions of the bases (A-green, T-red, G-yellow, C-blue) that were found in all mapped reads for the respective position. A position where all the bases of the reads are the same is shown as a gray bar. The read coverage is noted on the right side of the figure. Library preparation for samples (A–C) was performed with too much DNA. Negative controls follow the same library preparation steps as the samples (A–C) but with pure water instead of viral amplicon DNA. Additionally, the library preparation for the negative controls was performed in a separate lab and with new reagents up until the barcode pooling. Negative controls indicate SNP proportions that roughly reflect sample (A–C) considering their average coverage. The schematic figure is adapted from IGV.
Missing Amplicons
A common problem after SARS-CoV-2 genomes reconstruction are long stretches of ambiguous nucleotides due to amplicon dropouts because of the high number of primers during the multiplex PCR. An alternative to the commonly used ARTIC V3 amplicon protocol uses longer 1,200 bp amplicons to decrease the overall number of primers used. Longer amplicons usually lead to higher overall throughput (total bases) due to the longer read median. poreCov supports both primer sets (the standard 400 bp amplicons and longer 1,200 bp amplicons) and visualizes each sample’s coverage plot to identify errors during the multiplex PCR step (see Figure 2). A possible solution is to increase primer concentration for these specific “low coverage” regions. Additionally, it is possible to just re-amplify the missing regions via a cheap Flongle flowcell, combine both read sets and reconstruct and re-analyze via poreCov. For this, poreCov screens each genome and reports back which minimal set of primers would be needed to amplify only regions with ambiguous bases.
Many Ambiguous Bases
Besides missing amplicons (see above), the genome may contain many masked regions even though DNA concentrations and the number of sequenced reads were high. Most labs use direct patient material from swabs to sequence SARS-CoV-2 genomes to quickly return results without delay. Therefore, the sampling quality directly influences the sequencing run as any given sample could still have more human RNA/DNA than viral after isolation or if some of the isolation steps went wrong. This issue can be easily identified via poreCov as the amount of viral and human reads for each sample are summarized. Typical samples contain 90% or more viral reads after sequencing, and decreasing proportions might fail the genome reconstruction step or result in low-quality genomes (see Figure 2, columns: “%reads”).
Many Unclassified Barcodes
When using only reads with barcodes on both ends, this might lead to many reads without a barcode determination if something went wrong during the barcoding step in the wet lab. The number of unclassified barcodes can be visually inspected via the pycoQC report in the “1.Read_quality” dir. A possible solution is to deactivate both end requirements via poreCov’s “− −one_end’’ option flag. However, this can lead to the ‘‘strain bleeding’’ problem described above and is also not recommended.7 Therefore, it is strongly recommended to adjust the wet lab procedures as longer ligation times and fewer DNA molecules (mol) usually resolve this.
Staying Up to Date With Lineage Classifications
Lineage classification via pangolin is widely used and continuously updated and corrected based on the current pandemics’ prevalent virus variants. Due to this, the pangolin lineage assignment step in poreCov needs to be updated regularly. We ensure this via automatic, version-controlled, and tested container creation. Valid containers are automatically pushed and stored on hub.docker.com/r/nanozoo/pangolin. poreCov can check what is the latest available version for lineage classification before each run. If up-to-date lineage assignments are crucial, this feature can be manually activated and is deactivated by default to ensure the stability of the pipeline and stable offline usage. The pangolin version and the pangoLEARN version (the trained model for classification) are both noted on the final report to ensure reproducibility.
Conclusion
Nanopore sequencing represents an important pillar in the sequencing of SARS-CoV-2 genomes and thus in the molecular surveillance of novel viral variants and outbreak events of the global pandemic. However, due to its novelty and rapid development, nanopore sequencing also poses downstream challenges and pitfalls for accurate bioinformatic analysis and reproducibility. Many laboratories have ramped up nanopore sequencing but are in dire need of bioinformatics support given the high volume of global sequencing during the SARS-CoV-2 pandemic, especially when hundreds of samples need to be processed recurrently. Here we present poreCov, a high-throughput software pipeline for reliable genotyping and reference-based genome reconstruction of SARS-CoV-2 samples based on nanopore tiled amplicon sequencing. The pipeline is built around the widely used ARTIC protocol and integrates a comprehensive summary report with a clear result structure, allowing a quick assessment of the genome reconstruction performance. Clear color highlighting allows straightforward result interpretation even for large sequencing batches. Typical pitfalls during routine sequencing can be, therefore, quickly spotted and solved. In addition, the user can access all of the intermediate data via a well-structured result directory for further analysis or troubleshooting. We ensure a high degree of reproducibility and stability by utilizing Nextflow and virtualizing each tool through containers. We made sure that poreCov can run with the minimal required inputs and robust default parameters to diminish the bioinformatic entry barriers and enhance SARS-CoV-2 genome reconstruction and analysis during the pandemic.
Availability and Future Directions
The poreCov workflow is fully modularized to allow straightforward integration of additional processes, update tools, and add, if necessary, new primer schemes. As the primary framework utilizes the ARTIC protocol for genome reconstruction (as before also applied for, e.g., ZIKA and EBOV) (Quick et al., 2017), it can be easily adjusted to work for other viruses and potential further outbreaks. By that, “sub-workflows” of the pipeline can be automatically deactivated or conditionally activated based on the input data to avoid convoluting the workflow with parameters that need to be set by the user. Because poreCov is productively used at the public health institute in Germany, the SARS-CoV-2 side of the workflow will be continuously updated and extended if the necessity arises.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://github.com/replikation/poreCov.
Author Contributions
CB performed the conceptualization, outlined the overall manuscript and the workflow, and performed the project administration and funding. CB, SK, RS, and MH did the overall result validations and code contributions. RS, ML, MJ, and MM did the stability testing and issued reports. CB, RS, and MH created the figures. All authors actively participated in the writing and editing of the manuscript, read, and agreed to the published version of the manuscript.
Funding
This work received financial support from the Ministry for Economics, Sciences and Digital Society of Thuringia (TMWWDG), under the framework of the Landesprogramm ProDigital (DigLeben-5575/10-9) and by grants from the Federal Ministry of Education and Research (Grant No. 01KX2021).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Stephan Fuchs and Oliver Drechsel for creating and providing the Kraken2 database, used in gitlab.com/RKIBioinformaticsPipelines/ncov_minipipe, and Kathrin Trappe for feedback on running and testing the pipeline continuously. We thank all the people on GitHub that wrote poreCov issues and helped to identify and resolve bugs. We thank the GISAID team for providing metadata about the sequencing technologies used for SARS-CoV-2 sequencing.
Footnotes
- ^ hub.docker.com
- ^ github.com/artic-network/fieldbioinformatics
- ^ github.com/nanoporetech/medaka
- ^ github.com/jts/nanopolish
- ^ github.com/cov-lineages/pangolin
- ^ gitlab.com/RKIBioinformaticsPipelines/president
- ^ artic.network/quick-guide-to-tiling-amplicon-sequencing-bioinformatics.html
References
Brandt, C., Spott, R., Hölzer, M., Kühnert, D., Fuchs, S., Lohde, M., et al. (2021). Molecular epidemiology of SARS-CoV-2 - a regional to global perspective. medRxiv [Preprint]. doi: 10.1101/2021.01.25.21250447
Danecek, P., Bonfield, J. K., Liddle, J., Marshall, J., Ohan, V., Pollard, M. O., et al. (2021). Twelve years of SAMtools and BCFtools. Gigascience 10:giab008. doi: 10.1093/gigascience/giab008
De Coster, W., D’Hert, S., Schultz, D. T., Cruts, M., and Van Broeckhoven, C. (2018). NanoPack: visualizing and processing long-read sequencing data. Bioinformatics 34, 2666–2669. doi: 10.1093/bioinformatics/bty149
Di Tommaso, P., Chatzou, M., Floden, E. W., Barja, P. P., Palumbo, E., and Notredame, C. (2017). Nextflow enables reproducible computational workflows. Nat. Biotechnol. 35, 316–319. doi: 10.1038/nbt.3820
Durner, J., Burggraf, S., Czibere, L., Tehrani, A., Watts, D. C., and Becker, M. (2021). Fast and cost-effective screening for SARS-CoV-2 variants in a routine diagnostic setting. Dent. Mater. Off. Publ. Acad. Dent. Mater. 37, e95–e97. doi: 10.1016/j.dental.2021.01.015
Edge, P., and Bansal, V. (2019). Longshot enables accurate variant calling in diploid genomes from single-molecule long read sequencing. Nat. Commun. 10:4660. doi: 10.1038/s41467-019-12493-y
Elbe, S., and Buckland-Merrett, G. (2017). Data, disease and diplomacy: GISAID’s innovative contribution to global health. Glob. Chall. 1, 33–46. doi: 10.1002/gch2.1018
Ferguson, J. M., Gamaarachchi, H., Nguyen, T., Gollon, A., Tong, S., Aquilina-Reid, C., et al. (2021). InterARTIC: an interactive web application for whole-genome nanopore sequencing analysis of SARS-CoV-2 and other viruses. bioRxiv [Preprint]. doi: 10.1101/2021.04.21.440861
Hadfield, J., Megill, C., Bell, S. M., Huddleston, J., Potter, B., Callender, C., et al. (2018). Nextstrain: real-time tracking of pathogen evolution. Bioinformatics 34, 4121–4123. doi: 10.1093/bioinformatics/bty407
Hodcroft, E. B., Maio, N. D., Lanfear, R., MacCannell, D. R., Minh, B. Q., Schmidt, H. A., et al. (2021). Want to track pandemic variants faster? Fix the bioinformatics bottleneck. Nature 591, 30–33. doi: 10.1038/d41586-021-00525-x
Hodcroft, E. B., Zuber, M., Nadeau, S., Crawford, K. H. D., Bloom, J. D., Veesler, D., et al. (2020). Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. medRxiv [Preprint]. doi: 10.1101/2020.10.25.20219063
Hufsky, F., Lamkiewicz, K., Almeida, A., Aouacheria, A., Arighi, C., Bateman, A., et al. (2021). Computational strategies to combat COVID-19: useful tools to accelerate SARS-CoV-2 and coronavirus research. Brief Bioinform. 22, 642–663. doi: 10.1093/bib/bbaa232
Itokawa, K., Sekizuka, T., Hashino, M., Tanaka, R., and Kuroda, M. (2020). Disentangling primer interactions improves SARS-CoV-2 genome sequencing by multiplex tiling PCR. PLoS One 15:e0239403. doi: 10.1371/journal.pone.0239403
Leger, A., and Leonardi, T. (2019). pycoQC, interactive quality control for Oxford Nanopore sequencing. J. Open Source Softw. 4:1236. doi: 10.21105/joss.01236
Li, H. (2018). Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34, 3094–3100. doi: 10.1093/bioinformatics/bty191
Okonechnikov, K. Golosova, O., and Fursov, M. UGENE Team (2012). Unipro UGENE: a unified bioinformatics toolkit. Bioinform. Oxf. Engl. 28, 1166–1167. doi: 10.1093/bioinformatics/bts091
Ondov, B. D., Bergman, N. H., and Phillippy, A. M. (2011). Interactive metagenomic visualization in a Web browser. BMC Bioinform. 12:385. doi: 10.1186/1471-2105-12-385
Perkel, J. M. (2019). Workflow systems turn raw data into scientific knowledge. Nature 573, 149–150. doi: 10.1038/d41586-019-02619-z
Quick, J., Grubaugh, N. D., Pullan, S. T., Claro, I. M., Smith, A. D., Gangavarapu, K., et al. (2017). Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat. Protoc. 12, 1261–1276. doi: 10.1038/nprot.2017.066
Robinson, J. T., Thorvaldsdóttir, H., Winckler, W., Guttman, M., Lander, E. S., Getz, G., et al. (2011). Integrative genomics viewer. Nat. Biotechnol. 29, 24–26. doi: 10.1038/nbt.1754
Shu, Y., and McCauley, J. (2017). GISAID: global initiative on sharing all influenza data - from vision to reality. Euro Surveill. 22:30494. doi: 10.2807/1560-7917.ES.2017.22.13.30494
Tang, J. W., Tambyah, P. A., and Hui, D. S. (2021). Emergence of a new SARS-CoV-2 variant in the UK. J. Infect. 82, e27–e28. doi: 10.1016/j.jinf.2020.12.024
Thompson, R. N., Hill, E. M., and Gog, J. R. (2021). SARS-CoV-2 incidence and vaccine escape. Lancet Infect. Dis. 21, 913–914. doi: 10.1016/S1473-3099(21)00202-4
Wood, D. E., Lu, J., and Langmead, B. (2019). Improved metagenomic analysis with Kraken 2. Genome Biol. 20:257. doi: 10.1186/s13059-019-1891-0
Yan, Y., Wu, K., Chen, J., Liu, H., Huang, Y., Zhang, Y., et al. (2021). Rapid acquisition of high-quality SARS-CoV-2 genome via amplicon-oxford nanopore sequencing. Virol. Sin. doi: 10.1007/s12250-021-00378-8
Keywords: Nextflow, bioinformatics, coronavirus – COVID-19, lineages, docker, SARS-CoV-2, pipeline, nanopore sequencing
Citation: Brandt C, Krautwurst S, Spott R, Lohde M, Jundzill M, Marquet M and Hölzer M (2021) poreCov-An Easy to Use, Fast, and Robust Workflow for SARS-CoV-2 Genome Reconstruction via Nanopore Sequencing. Front. Genet. 12:711437. doi: 10.3389/fgene.2021.711437
Received: 18 May 2021; Accepted: 09 July 2021;
Published: 28 July 2021.
Edited by:
Giovanni Bacci, University of Florence, ItalyReviewed by:
Ahmed Moustafa, American University in Cairo, EgyptLuca Ermini, King Abdullah University of Science and Technology, Saudi Arabia
Christopher Riccardi, University of Florence, Italy
Copyright © 2021 Brandt, Krautwurst, Spott, Lohde, Jundzill, Marquet and Hölzer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Brandt, christian.brandt@med.uni-jena.de