
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 14 October 2021
Sec. Behavioral and Psychiatric Genetics
Volume 12 - 2021 | https://doi.org/10.3389/fgene.2021.689756
 Cassandra C. Soo1,2*
Cassandra C. Soo1,2* Meagan T. Farrell3
Meagan T. Farrell3 Stephen Tollman4Lisa Berkman3,5
Stephen Tollman4Lisa Berkman3,5 Almut Nebel1,6†
Almut Nebel1,6† Michèle Ramsay1*†
Michèle Ramsay1*†Apolipoprotein E (APOE) 𝜀4 allele carrier status is well known for its association with an increased likelihood of developing Alzheimer’s disease, but its independent role in cognitive function is unclear. APOE genetic variation is understudied in African populations; hence, this cross-sectional study in a rural South African community examined allele and genotype frequencies, and their associations with cognitive function. Cognitive function was assessed using two different screening methods to produce a total cognition score and four domain-specific cognition scores for verbal episodic memory, executive function, language, and visuospatial ability. Cognitive phenotype and APOE genotype data were used to determine whether APOE variation was significantly associated with cognitive function in this population. Observed allele frequencies for 1776 participants from the HAALSI study [age 40–80years (mean=56.19); 58.2% female] were 58.1% (𝜀3), 25.4% (𝜀4) and 16.5% (𝜀2). Allele distributions were similar to the African super population, but different from all non-African super populations from the 1,000 Genomes Project. The 𝜀3 homozygous genotype was most common (34.9%) and used as the base genotype for comparison in regression models. Four models were tested for each of the five cognitive phenotypes to explore association of APOE variation with cognitive function. In the first model assessing association with all genotypes for all individuals, marginally significant associations were observed for 𝜀2 homozygotes where executive function scored higher by ~0.5 standard deviations (p=0.037, SE=0.23), and for 𝜀3/𝜀4 heterozygotes where visuospatial ability scores were lower (p=0.046, SE=0.14). These did not survive correction for multiple testing. Regional African population differences were observed at the APOE locus. Marginally, significant associations between APOE genotype, and executive function and visuospatial ability indicate the need for larger studies to better examine these associations in African populations. Furthermore, longitudinal data could shed light on APOE genetic association with rate of change, or decline, in cognitive function.
Studies conducted in different populations (mostly of European ancestry) have reported associations between apolipoprotein E (APOE) 𝜀4 and various detrimental outcomes. These include susceptibility to, and severity of cardiovascular diseases, HIV infection and its associated comorbidities, late onset Alzheimer’s disease, and recently COVID-19 (Hendrie et al., 1995; Savitz et al., 2006; Burt et al., 2008; Chang et al., 2011; Panos et al., 2013; Kotze et al., 2015; O’Donoghue et al., 2018; Kuo et al., 2020). Aside from its well-known involvement in disease pathogenesis, there is interest in understanding the role of APOE variation in normal cognitive function throughout lifespan. This is especially true as studies begin to identify associations across the globe in heterogeneous populations (Kotze et al., 2015). Furthermore, cognition is a complex multifactorial trait and studies examining normal cognitive ageing and brain morphology have observed associations with APOE (Small et al., 2004; Wisdom et al., 2011; Rebeck, 2017; O’Donoghue et al., 2018; Hays et al., 2019; Reas et al., 2019). These results have, however, been inconsistent, and larger meta-analyses have observed small, significant differences in cognitive performance and brain morphology in European populations between 𝜀4-carriers and non-𝜀4-carriers (Small et al., 2004; Wisdom et al., 2011; O’Donoghue et al., 2018). Studies focused on Alzheimer’s disease (AD) have shown that the presence of 𝜀4 is associated not only with increased susceptibility to AD, from 2-fold to 33-fold depending on the number of risk alleles and ethnicity, but also with an earlier age of onset of the disease (Wisdom et al., 2011; Liu et al., 2013; Wendelken et al., 2016; Rebeck, 2017). It has also been found to be associated with other neurocognitive phenotypes, including mild cognitive impairment (MCI), HIV-associated neurocognitive disorder (HAND), brain atrophy and structural pathologies, and the pace of age-related cognitive decline (Burt et al., 2008; Chang et al., 2015; Hendrie et al., 2015; Wendelken et al., 2016; Lipnicki et al., 2017; Espinosa et al., 2018; O’Donoghue et al., 2018; Reas et al., 2019). However, the results of these studies were mixed, as were the strengths of associations and effects for different ethnic groups (Wisdom et al., 2011; Panos et al., 2013; Lipnicki et al., 2019; Reas et al., 2019). This may be due to differences in study design, sample size, characterisation and collection of cognition data, and genetic diversity due to differences in ethnicity.
The APOE alleles 𝜀2, 𝜀3, and 𝜀4 encode structurally different protein isoforms with variable molecule-binding affinity (Corbo and Scacchp, 1999). APOE is involved in lipid metabolism, specifically of cholesterol, and is integral to cholesterol transport and clearance within the central nervous system (Rebeck, 2017). Global variation in APOE allele frequencies have been reported 185 between ethnic groups (Corbo and Scacchp, 1999; Demarchi et al. 2005; Singh et al., 2006. The 𝜀3 allele is the most common allele across all populations, and 𝜀4 frequencies are higher in African and aboriginal populations than in Europeans and Asians (Sandholzer et al., 1995; Corbo and Scacchp, 1999; Singh et al., 2006; Liu et al., 2013). Despite this, evidence for its association with AD and other neurocognitive phenotypes in populations of African ancestry is either lacking, or very weak (Osuntokun et al., 1995; Willis et al., 2003; Gureje et al., 2006; Chen et al., 2010; Liu et al., 2013; Morgan et al., 2013; Chang et al., 2015; Hendrie et al., 2015).
Genomic studies of cognitive function among Africans are limited by small sample sizes and low transferability of cognitive performance tests to regions where access to formal education and lower levels of literacy may affect reproducibility of results observed in largely European cohorts (Savitz et al., 2006; Humphreys et al., 2017; Kobayashi et al., 2019; Farrell et al., 2020). The Health and Aging in Africa: A Longitudinal Study of an INDEPTH Community in South Africa (HAALSI) was established to assess trends in sociodemographic determinants, morbidity, and mortality associated with population greying and the current epidemiological and behavioural transition underway in South Africa (SA) (Gómez-Olivé et al., 2018; Houle et al., 2019). Cognitive performance data were captured for over 5,000 individuals aged 40years and older from rural communities in Bushbuckridge, Mpumalanga, SA, a region known to have a high prevalence of HIV (Gómez-Olivé et al., 2018; Asiimwe et al., 2020; Rosenberg et al., 2020). Two screening tools were used to capture cognitive function: (1) a culturally adapted version of the United States Health and Retirement Study (US HRS) screening tool translated into the local language Shangaan and (2) the tablet-based Oxford Cognition Screen (OCS-Plus), which was adapted and validated in this study population (Humphreys et al., 2017; Gómez-Olivé et al., 2018; Kobayashi et al., 2019; Farrell et al., 2020). It provides domain-specific scores for cognitive function in episodic memory, executive function, language, and visuospatial ability without bias to levels of literacy or numeracy (Humphreys et al., 2017; Kobayashi et al., 2019; Farrell et al., 2020). Here, a subset (n=1776) of the larger HAALSI cohort was genotyped for the APOE locus. Our aim was to examine the associations of APOE with total cognition score and specific cognitive domains adjusting for known moderators, such as age, level of education and sex. HIV status was also included as a covariate, and we sought to explore a possible relationship between HIV status and APOE in this cohort.
Demographic, socioeconomic, health and cognitive function data were collected from 5,059 adults (men and women, aged 40years and older) recruited from Bushbuckridge, rural Mpumalanga, South Africa, from November 2014 to November 2015, as the baseline for the HAALSI longitudinal study (Gómez-Olivé et al., 2018). African participants who had been living in the area for at least 12months preceding data collection were selected from the Health and Demographic Surveillance System database and consented (Kahn et al., 2012). Cognitive function variables: HIV status, age, sex, and educational attainment expressed as level of education (none, primary, high and tertiary) were used in this population-based cross-sectional community study. Sample size differed per outcome due to missing data. Approximately 2000 of these same participants overlapped with the Africa Wits-INDEPTH Partnership for Genomic Studies (AWI-Gen) for which blood samples were collected for DNA extraction and genetic analyses (Ramsay et al., 2016; Ali et al., 2018). Data collection and informed consent templates for the HAALSI and AWI-Gen studies were approved by the University of the Witwatersrand, Johannesburg, Human Research Ethics Committee [Wits HREC (Medical); M141159, M121029], the Harvard T.H. Chan School of Public Health, Office of Human Research Administration (C13-1608-02), and the Mpumalanga Provincial Research and Ethics Committee (approved: 2014/10/22). Ethics approval for this study was independently granted by Wits HREC (Medical) (M170916).
Two different tools were utilised to collect data on cognitive function (Gómez-Olivé et al., 2018; Kobayashi et al., 2019; Farrell et al., 2020). The first tool was adapted from the US HRS cognition screening tool. It was translated into the local language, Shangaan, and then back-translated to ensure cultural appropriateness and transferability (Kobayashi et al., 2019; Farrell et al., 2020). It included questions assessing the following measures: orientation, immediate and delayed word recall, numeracy, self-rated memory, and literacy (Gómez-Olivé et al., 2018; Kobayashi et al., 2019; Farrell et al., 2020). A score from 0 to 24, defined here as, ‘total cognition score’, representing global cognition, was used as one outcome variable for this study. The OCS-Plus was developed and then validated in a subset of HAALSI participants to capture cognitive domain-specific data without literacy or numeracy bias (Humphreys et al., 2017). Episodic memory, language, executive function, and visuospatial cognition data were collected and transformed into z-scores for each individual per domain thus providing standardised domain-specific scores for this population using methods described by Seidlecki et al. (2008) (Farrell et al., 2020). The sample size was larger for total cognition score than for the domain outcomes.
DNA was extracted from buffy coat samples using the QIAsymphony SP (QIAGEN GmbH, QIAGEN Strasse 1, 40,724 Hilden, Germany) at the Sydney Brenner Institute for Molecular Bioscience (SBIMB). TaqMan™ single-nucleotide polymorphism (SNP) genotyping was performed using DNA from 1952 individuals for rs429358 and rs7412, of which 1776 were successfully genotyped at both loci. Genotyping was performed on the QuantStudio™ 5 Real-Time PCR System (Applied Biosystems, Thermo Fisher Scientific, Carlsbad, United States), and results were analysed using the QuantStudio™ Design & Analysis Software. APOE alleles were determined by using the genotypes for the SNPs rs429358 (T, C) and rs7412 (C, T). When the combined haplotype was TT, this represented the ε2 allele. When the haplotype was TC ε3, and when the haplotype was CC this was interpreted as ε4.
SNP data for 2,504 individuals from the 1000 Genomes Project (Phase 3) were downloaded from the Ensembl genome browser for the Genome Reference Consortium Human Genome build 37 (GRCh37; ftp://ftp.1000genomes.ebi.ac.uk/vol1/ftp/phase3/data; Accessed 11 Feb, 2021; Auton et al., 2015; 1,000 Genomes Project and AWS, RRID:SCR_008801). Samples were divided according to super population codes as representative of European (EUR), African (AFR), East Asian (EAS), South Asian (SAS), and Admixed American (AMR) ancestries (Auton et al., 2015; 1,000 Genomes Project and AWS, RRID:SCR_008801). APOE alleles were determined as above, and frequencies for each of these super populations were compared to our data.
Allele and genotype frequencies were calculated, and Hardy-Weinberg equilibrium (HWE) was tested using the Pearson’s Chi-squared test. Chi-squared tests were used to test for differences in APOE allele distribution and categorical measures of age, sex, level of education, HIV status, each of the 1,000 Genomes super populations and data from four previous African studies (Willis et al., 2003; Gureje et al., 2006; Chen et al., 2010 and Joska et al., 2010). The allele frequency data are represented as pie diagrams on the backdrop of a map of Africa created by MapChart1 (2021). In order to assess correlations between APOE distribution and age, age was categorised firstly as 10-year intervals, and then as younger (40–59years) vs. older (60years and older). Descriptive statistics was used to summarise continuous (age, total cognition score and domain-specific z-scores) and categorical variables (HIV status, level of education). Two-sample hypothesis testing was used to further tease out the observed association between APOE and HIV. This was followed by logistic regression to determine the effects of each allele on HIV status while adjusting for the presence or absence of one of the other alleles. Cognitive function, either as total cognition score or as one of the four above-mentioned domains, were used as outcome variables to test for associations. Linear regression models for each cognitive outcome were tested. Four models were tested as follows: (1) regression using six known genotypes (2) regression comparing 𝜀3 homozygotes to 𝜀4-carriers (3) regression comparing 𝜀4 homozygotes to the rest of the genotypes and (4) regression comparing 𝜀2 homozygotes to 𝜀4 homozygotes. Chi-squared tests, two-sample proportion testing and logistic regression analysis reporting McFadden’s R2 were performed using STATA 15.0.585 (StataCorp, 2017; Stata Statistical Software: Release 15, College Station, TX: StataCorp LLC; Stata, RRID:SCR_012763), and summary statistics, regression analyses and correction for multiple testing were performed using R (R Core Team, 2020. R: A language and environment for statistical computing; R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/; R Project for Statistical Computing, RRID:SCR_001905). Bonferroni correction was applied for multiple testing.
The phenotypic characteristics and covariates of the study population along with the number of participants (n) with data for each variable are shown in Table 1. The mean age was 56.19years, and there were more female participants (58.22%). Education levels in the community were low with the majority having no formal education or having only attended primary school (74.66%). HIV infection was high with 28.48% of the study population living with HIV. Total cognition score was normally distributed with a mean of 11.98. The four standardised domain scores showed considerable variability. The range was particularly wide for visuospatial cognition (−10.627 to 4.771) and executive function (−4.687 to 4.328).
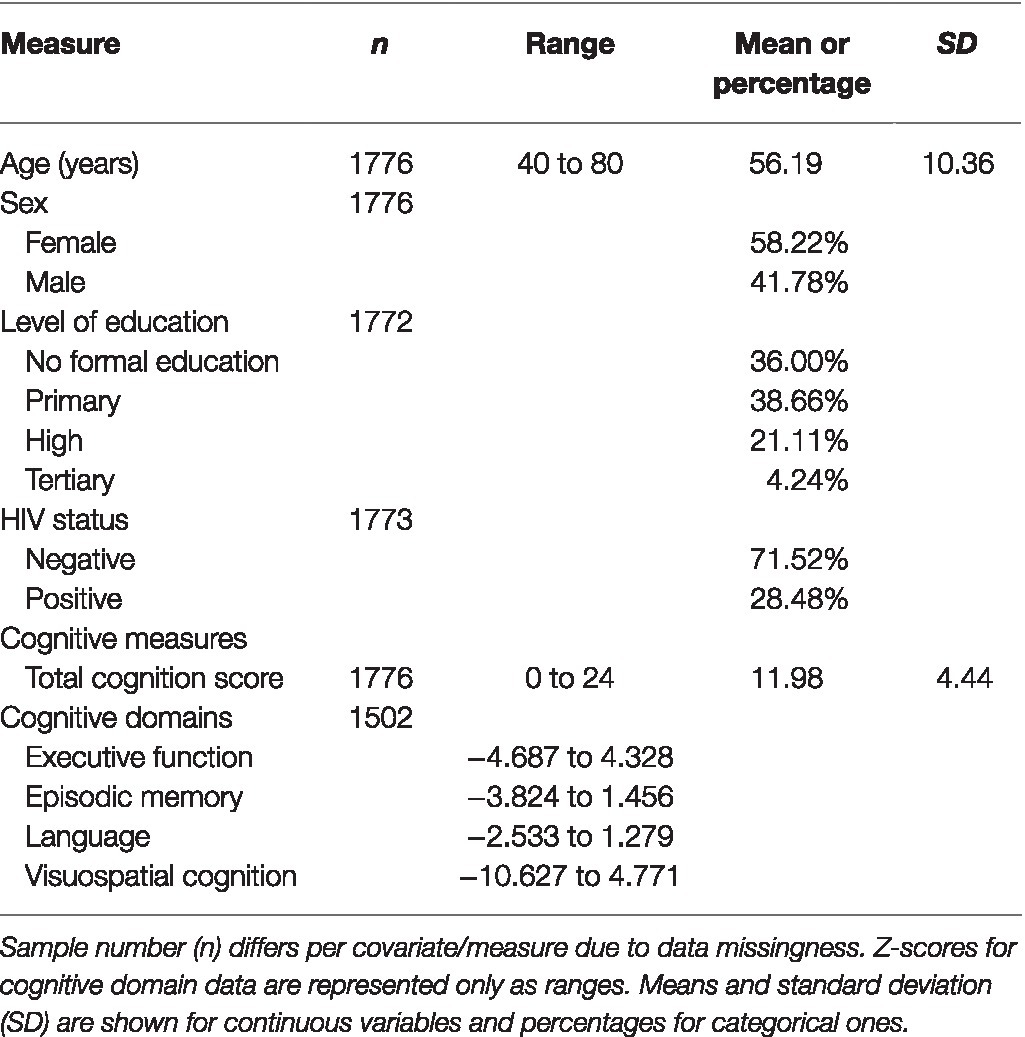
Table 1. Summary characteristics of study population.
In total, 1776 individuals were successfully genotyped, and all six genotypes were observed with 𝜀2 homozygotes having the lowest frequency (2.9%) and 𝜀3/𝜀3 the highest (34.9%; Table 2). The genotype distribution was in Hardy Weinberg equilibrium (p=0.28). Our sample had a similar allele frequency distribution to that of the AFR super population group of Phase 3 of the 1,000 Genomes Project (χ2=1.0983, df=2, p=0.35; Figure 1). The frequencies of 𝜀2 and 𝜀4 were higher in our sample than in the other populations (EUR, EAS, SAS and AMR representing South American ethnicities). When compared to other African studies (Figure 2), a significant difference in allele frequency distribution was observed between our data and that of Chen et al. (2010) in a study from Kenya (χ2=11.058, df=2, p=0.004), but due to the small sample size, this difference may not be true.

Table 2. APOE allele and genotype frequencies.
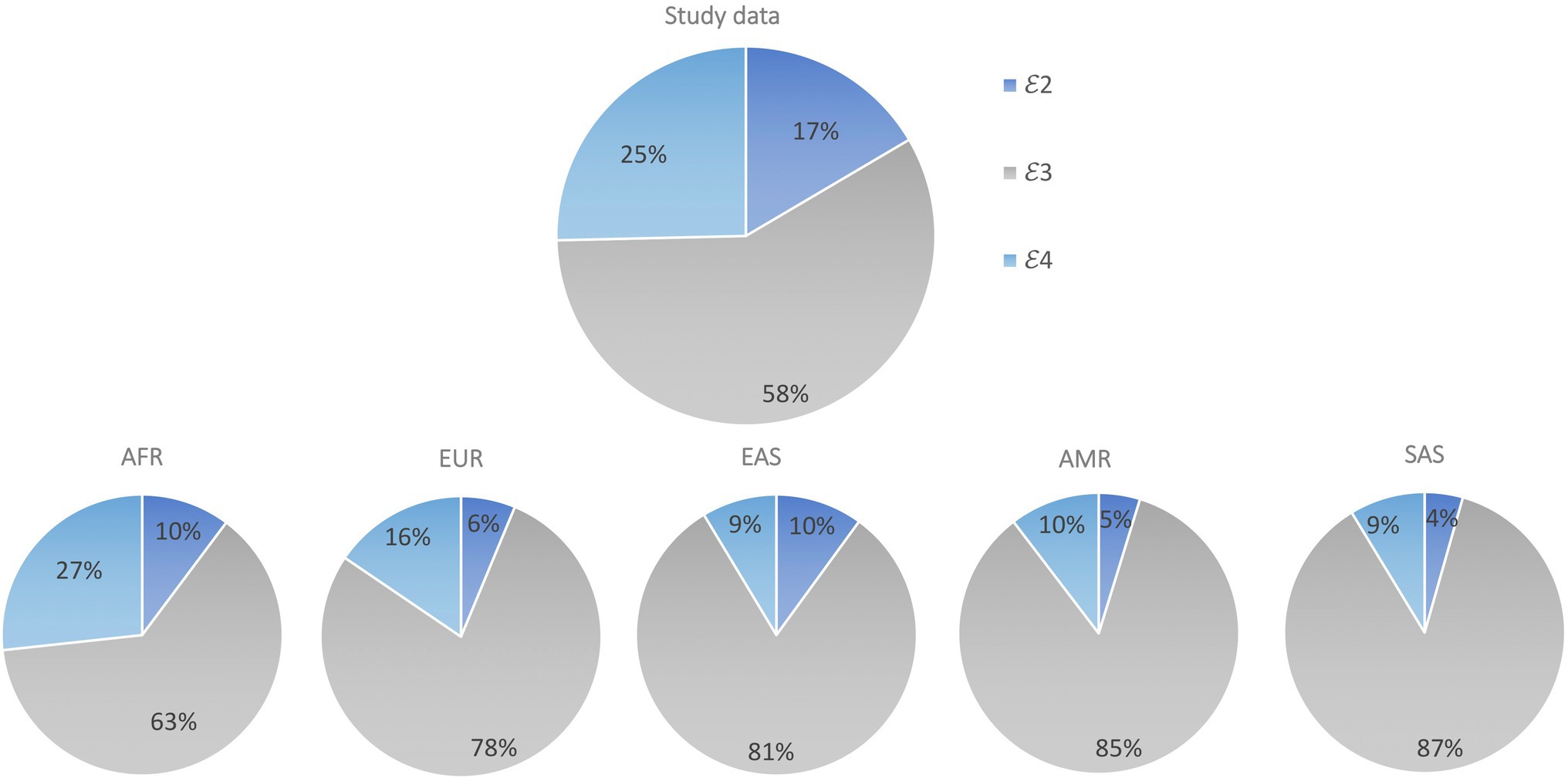
Figure 1. Comparison of APOE allele distribution in our data set (Study data) compared to 1,000 Genomes super population data sets AFR, EUR, EAS, SAS and AMR. Pearson’s Chi-squared test was used to assess differences in APOE allele frequency between sample populations from each of the 1,000 Genomes super populations and our study sample.
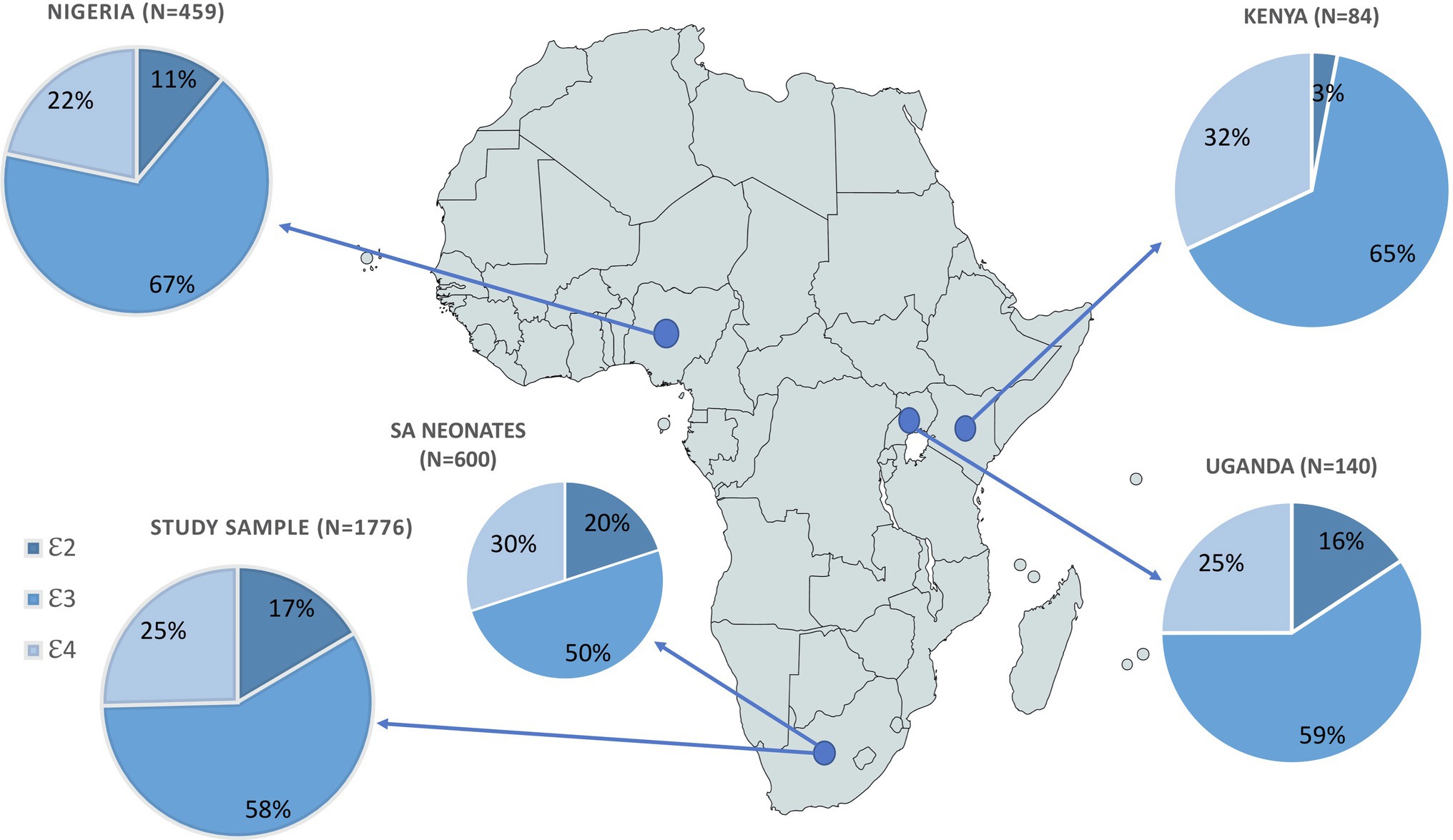
Figure 2. Comparison of APOE allele frequency distributions from four African studies, showing sample size for each study. Pearson’s Chi-squared test was used to assess differences in APOE allele frequency between sample populations from each of the African studies referenced by the country of origin/description: Uganda: Willis et al., 2003; Nigeria: Gureje et al., 2006; Kenya: Chen et al., 2010; SA neonates: Joska et al., 2010.
APOE allele distribution showed no correlation with sex (χ2=1.3329, df=2, p=0.514) or level of education (χ2=5.7393, df=6, p=0.453). No significant difference in allele frequency was observed between age categories per decade nor for younger vs. older individuals (χ2=2.0264, df=6, p=0.917, and χ2=0.4175, df=2, p=0.812, respectively). There was evidence of a relationship between HIV status and APOE allele frequency (χ2=6.9266, df=2, p=0.031). Results of hypothesis testing indicated that the proportion of ɛ3 alleles was higher in HIV individuals than in those without HIV (p=0.001) and the opposite was observed for ɛ4 (p=0.007). Logistic regression results showed the size and direction of the effects of each allele on HIV status, but these associations were not significant as our model had little predictive value (McFadden’s pseudo R2=0.0023; Table 3).
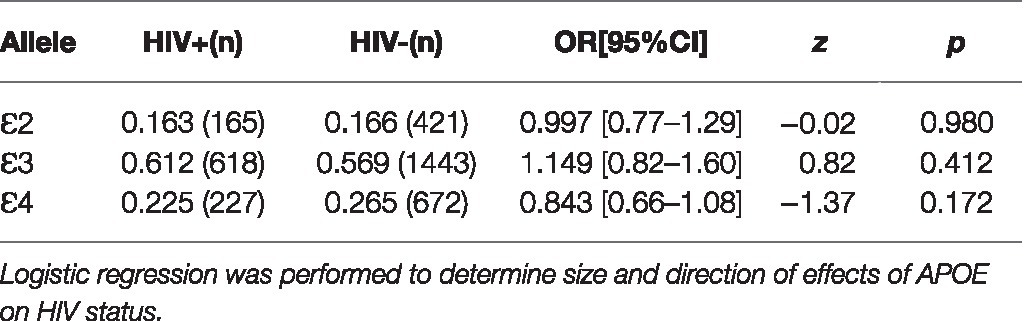
Table 3. APOE allele association with HIV status.
Four multiple regression models were applied, to allow for comparative analyses with other studies, to each of the five cognitive phenotypes. Each model was representative of the comparison between APOE genotypes (Model 1: all genotypes; Model 2: 𝜀3 homozygotes vs. 𝜀4-carriers; Model 3: 𝜀4 homozygotes vs. the rest of the genotypes; and Model 4: 𝜀2 homozygotes vs. 𝜀4 homozygotes) and their effect on cognition while adjusting for covariates (age, sex, level of education and HIV status). Age and level of education were significantly associated with cognitive function in all of the models, and sex was significant for executive function, language and visuospatial cognition for models 1–3 where being male increased the standard estimates for each domain (Table 4 and Supplementary Table S1). Significant associations were observed in Model 1 (Table 4) for APOE genotypes, where 𝜀2/𝜀2 was associated with higher levels of executive function by approximately half a standard deviation (β=0.489, SE=0.57, p=0.037) and lower levels (but not significantly) when participants were heterozygous 𝜀2-carriers, and 𝜀3/𝜀4 was associated with lower levels of visuospatial cognition (β=−0.286, SE=0.14, p=0.046). These effects (f2=0.395 and f2=0.597, respectively) were not significant after Bonferroni correction for multiple testing (p=0.337 and 0.416, respectively).
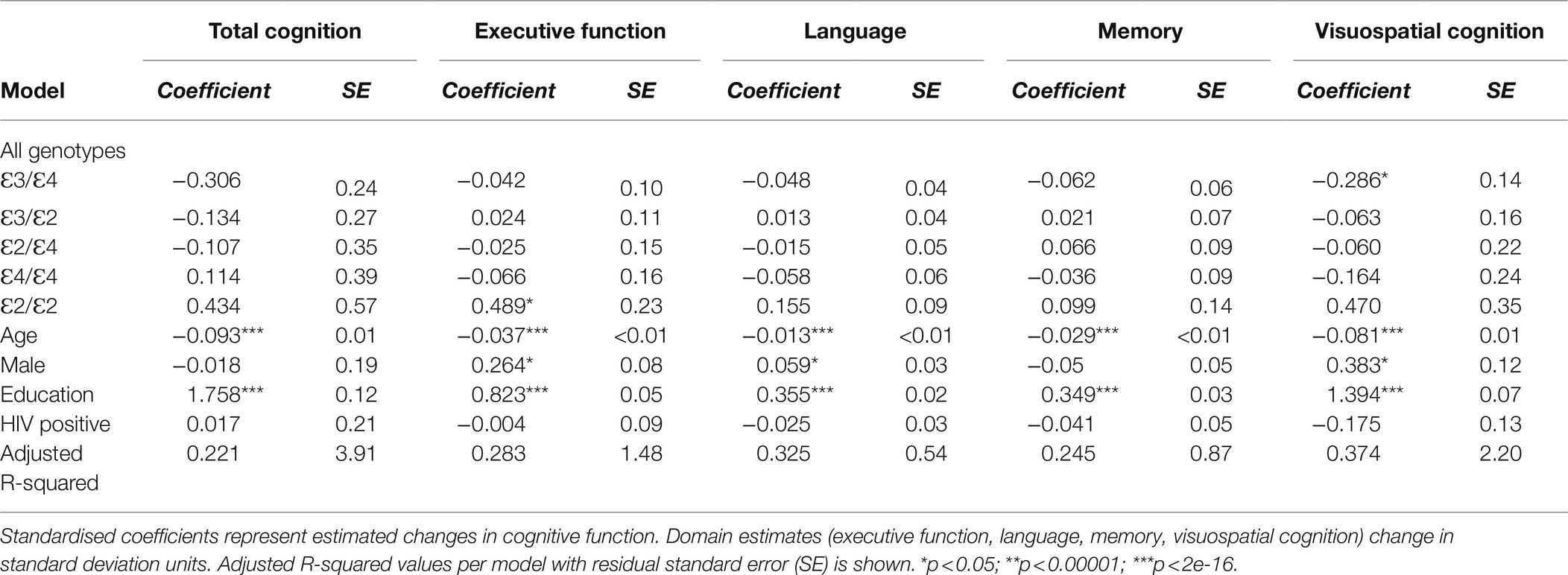
Table 4. Linear regression model 1 using all genotypes to estimate the effects of APOE and covariates on cognitive function.
This cross-sectional study assessed the potential effect of APOE genetic variation on five measures of cognition in a rural-dwelling SA population. Participants were adults of African ethnicity aged 40years and older. We aimed to address the lack of African data for large cohorts that can be used to examine the genetics of cognition and cognitive decline, specifically in rural areas where vulnerability to cognitive impairment and dementia is becoming a major public health concern (Kobayashi et al., 2019). Association studies have observed genetic variation in neurocognitive traits between different ethnicities (Savitz et al., 2006; Fitzgerald et al., 2020). Here, we determined APOE allele frequency distribution for our dataset and then compared this to existing data from other population groups. The genetic contribution to cognitive function is complex, and the tests used to define levels of cognition have made equitable comparisons between studies difficult (Savitz et al., 2006; Goriounova and Mansvelder, 2019; Fitzgerald et al., 2020). This is especially challenging in a low literacy setting despite the development of tests to assess cognition regardless of levels of literacy. Nonetheless, both literacy and level of education are still associated with cognition in this study cohort (Kobayashi et al., 2019; Farrell et al., 2020). Traditionally, intelligence has been measured using the intelligence quotient (IQ) or Spearman’s g (used in meta-analyses), which are biased towards access to formal education (as they focus mostly on aspects related to literacy and numeracy) (Goriounova and Mansvelder, 2019; Fitzgerald et al., 2020). Here, we focused on latent cognitive ability which encompasses domain-specific and global cognitive function outside of educational attainment (Goriounova and Mansvelder, 2019; Fitzgerald et al., 2020). Collecting cognition data is further complicated by using screening tools which have not been validated for transferability across populations; therefore, the OCS-Plus tool was developed to provide an alternative method for capturing cognition data in low-income populations that was neither reliant on literacy nor numeracy (Humphreys et al., 2017). The HAALSI baseline study provided an appropriate setting to validate this tablet-based tool, providing population-specific cognitive function data for an understudied and socially homogeneous population (Humphreys et al., 2017; Gómez-Olivé et al., 2018). Leveraging these data and further capitalising on the overlap of AWI-Gen participants for which DNA was available, we sought to explore genetic effects of APOE on cognitive function.
APOE genotyping revealed that ɛ2 and ɛ4 are generally more common in African populations, including in our own, with SA and Uganda having the highest frequencies of ɛ2 among African populations (Figure 2). Although these allele frequencies differed significantly from the Chen et al. (2010) cross-sectional study of a population in Kenya, the small sample size of this Kenyan study was insufficient for accurate comparison of intra-continental variation of APOE. The higher ɛ4 frequency in Africans has been attributed to selective advantage as protection against infectious disease (Corbo and Scacchp, 1999; Fujioka et al., 2014; Van Exel et al., 2017; Smith et al., 2019). APOE ɛ4 has been suggested to be protective against hepatitis C infection as well as carriers being observed to have a better prognosis post infection (Kuhlmann et al., 2010; Mueller et al., 2016; Smith et al., 2019). There is also evidence that ɛ4 inhibits growth of Plasmodium falciparum conveying a protective effect against malaria (Fujioka et al., 2014; Van Exel et al., 2017; Smith et al., 2019). Higher observed ɛ4 frequency in Africans has also been attributed to the hypothesis that it is beneficial for fertility and early infant survival (Joska et al., 2010; Van Exel et al., 2017; Smith et al., 2019). Despite these proposed early life advantages, it has also been associated with increased morbidity and mortality later in life (Hendrie et al., 1995; Savitz et al., 2006; Burt et al., 2008; Chang et al., 2011; Panos et al., 2013; Kotze et al., 2015; Van Exel et al., 2017; O’Donoghue et al., 2018; Smith et al., 2019; Kuo et al., 2020). Smaller African studies mainly assessing APOE association with AD showed no significant associations (Willis et al., 2003; Gureje et al., 2006; Chen et al., 2010; Joska et al., 2010). The genetic associations between ɛ4 and neurocognitive phenotypes (mostly AD due to its highly replicated association with increased susceptibility) have observed weaker effects in those of admixed African ancestry (AA and Brazilians) and lack of association in Africans (Osuntokun et al., 1995; Willis et al., 2003; Small et al., 2004; Wisdom et al., 2011; Hendrie et al., 2015; O’Donoghue et al., 2018; Gouveia et al., 2019).
Previous epidemiological studies on the HAALSI cohort have reported the effects of age, sex, level of education and HIV status on these same cognitive phenotypes (Kobayashi et al., 2019; Asiimwe et al., 2020; Farrell et al., 2020). Higher educational attainment and younger age are known to be associated with better cognitive performance, and we observed these same education-related effects across all cognitive phenotypes in our study and in our regression models. In this study population, we observed better performance in the domains of executive function and language in male participants, a finding that had already been published by Farrell et al. (2020) who attributed this sex difference to socio-cultural differences and access to education (Kobayashi et al., 2019). Other studies have suggested that older women generally have better cognitive performance than their male counterparts possibly due to the effects of oestrogen or differences in cognitive reserve (Wu et al., 2012; Hayat et al., 2014; Clifford et al., 2015; Sundermann et al., 2016; Lipnicki et al., 2019). Sex effects appear to be domain-specific, and this has been observed in studies where women performed better in tests assessing memory than men irrespective of educational background (Clifford et al., 2015; Sundermann et al., 2016).
For this study, we used HIV status as a covariate because of the high prevalence of HIV and its known contribution to cognitive impairment through HAND (Andres, 2011; Panos et al., 2013; Chang et al., 2015; Rebeck, 2017; Geffin and McCarthy, 2018; Hulgan et al., 2018). Upon assessment of the APOE distributions for each of the covariates, we observed statistical differences between allele distribution according to HIV status. When modelling the effects of each allele on HIV status, logistic regression indicated that our model consisting of HIV status conditional on APOE allele was inadequate, which could be due to small sample size. Although the proportion of ɛ3 alleles was significantly higher in HIV positive individuals, the relative effect of this allele on HIV status compared to the effects of the other alleles was not significant. This is contrary to another SA study reporting lower ɛ3 allele frequency in HIV positive adults vs. newborn controls (Joska et al., 2010). The opposite relationship for ɛ4, with a higher proportion observed in those without HIV, was also not significant, but our results suggested that ɛ3 is the risk allele for HIV infection. APOE ɛ4 has been linked to AIDS severity and increased mortality especially in ɛ4 homozygotes (Valcour et al., 2004; Burt et al., 2008; Chang et al., 2015; Wendelken et al., 2016). It has been associated with faster disease progression and higher viral load in seropositive individuals due to enhanced entry of the virus into T cells (Burt et al., 2008; Kuhlmann et al., 2010). This may explain why we see a higher proportion of ɛ3 in our HIV positive sample as faster disease progression and poorer prognosis may have resulted in earlier HIV-related mortality in ɛ4-carriers. The mortality profile of the community from which this cohort was recruited was subject to high levels of HIV-associated death, and increased all cause death in age groups from 15 to 64years in the years preceding data collection for this study (Kabudula et al., 2014). HIV progression is accompanied by domain-specific cognitive deficits characterised as HAND, where ɛ4 is associated with reduced performance in these domains (Chang et al., 2011; Panos et al., 2013; Chang et al., 2015; Wendelken et al., 2016; Geffin and McCarthy, 2018). Some studies have failed to replicate these findings and have reported either no association between ɛ4 and HIV, or lack of association between ɛ4 and HAND (Joska et al., 2010; Morgan et al., 2013; Becker et al., 2015; Cysique et al., 2015). HIV status had no significant effect on cognitive function in any of our models. This result may be attributed to widespread use of antiretroviral therapy in this community and also an indication that people living with HIV may have better access to healthcare and thus are better able to manage both age-related and HIV-associated morbidity (Kabudula et al., 2014; Asiimwe et al., 2020; Rosenberg et al., 2020). There is evidence that this sample population may have a high prevalence of HAND within the HIV positive group, but further evidence for trends in the development of accurately diagnosed HAND and seropositivity in the context of APOE have not been explored (Asiimwe et al., 2020; Rosenberg et al., 2020).
Although we observed possible associations of APOE genetic variants with executive function and visuospatial cognition, these associations did not survive correction for multiple testing. APOE ɛ2 homozygosity was linked to increased executive function scores and ɛ3/ɛ4 heterozygosity to lower visuospatial performance. Other studies comparing performance on cognition tests between ɛ3 homozygotes and ɛ4-carriers observed impaired executive function, episodic memory, global cognition and processing speed in ɛ4-carriers which was exacerbated in ɛ4 homozygotes (Lipnicki et al., 2017; O’Donoghue et al., 2018; Hays et al., 2019). Replication of these findings has proven challenging (Savitz et al., 2006; O’Donoghue et al., 2018; Fitzgerald et al., 2020). We observed an increase of ~0.5 standard deviations in the domain of executive function for ɛ2/ɛ2 individuals and~0.3 standard deviations reduction in visuospatial ability of ɛ3/ɛ4 individuals, larger effects than those of a large meta-analysis by Small et al. (2004). They observed reduced performance of <0.1 standard deviations for global cognition, episodic memory and executive function in ɛ4 carriers, with significantly poorer cognitive performance associated with ɛ4 homozygosity (Small et al., 2004). A later meta-analysis of 77 studies totalling 40,942 individuals observed similar results when assessing the effects of APOE on normal cognitive function (Wisdom et al., 2011). Contrary to the findings of Wisdom et al. (2011), we did not observe significantly higher memory scores in ɛ2 homozygotes compared to ɛ4 homozygotes.
We report on baseline genetic and domain-specific cognitive function data for a large African study cohort. However, sample size was a limitation for observing small genetic effects for associations between APOE genotypes and cognitive function. Meta-analyses have estimated the effect sizes of APOE on different cognitive domains to vary between approximately 0.07 and 0.002 (Small et al., 2004; Wisdom et al., 2011). But to detect the effect size of 0.01 estimated by Wisdom et al. (2011) for visuospatial ability, we would need to double our sample size. Our models explained modest variation of cognitive function mostly due to highly significant associations observed with age (cognition score decreasing with increasing age) and level of education (cognition score increasing as higher levels of education are achieved), which are known to influence normal cognitive function. It has been suggested that cognition changes throughout the lifespan and that ɛ4-carriers perform better at cognition tests when they are younger due to antagonistic pleiotropy and the compensation of other pathways which improve cognition and cognitive reserve at younger ages, but become detrimental over the age of 60years (Chang et al., 2011; Panos et al., 2013; Rebeck, 2017; O’Donoghue et al., 2018). We were unable to confirm this in our study, but longitudinal data from HAALSI’s second wave of data collection will enable the assessment of the effects of APOE on age-related cognitive decline.
There is little data on cognitive function and domain-specific genetic associations in African populations. Research in European and Asian populations has shown that APOE genetic variation is associated with cognition and differences in susceptibility to several diseases. Despite previous associations between ɛ4 and cognition, we were unable to replicate these findings. Instead, we observed a trend towards higher executive function in ɛ2 homozygotes and lower visuospatial cognition in ɛ3/ɛ4 heterozygotes. These associations were strongly moderated by education and age, and there was minimal indication for the independent influence of APOE on latent cognitive function. Cognition screening tools which are culturally adapted and unbiased in terms of literacy and numeracy are essential for the accurate interpretation of cognition in populations with low levels of education. The inconclusive results of this study may further catalyse African exploratory studies of the complex genetics behind cognition, and the role of known environmental confounders that are becoming increasingly important in the prediction, prevention, and treatment of dementia in low- to middle-income countries.
The HAALSI baseline data are publicly available at the Harvard Center for Population and Development Studies (HCPDS) programme website [www.haalsi.org]. Data are also accessible through the MRC/Wits Agincourt Research Unit’s data repository [https://data.agincourt.co.za/index.php/catalog/18], the Inter-university Consortium for Political and Social Research (ICPSR) at the University of Michigan [www.icpsr.umich.edu], and the INDEPTH Data Repository [https://www.indepth-ishare.org/index.php/catalog/113]. The linked genetic and phenotypic data is available through the Harvard Center for Population and Development Studies (HCPDS) programme website [https://dataverse.harvard.edu/dataset.xhtml?persistentId=doi:10.7910/DVN/QWSQXR]. Requests for these data through the website are made to the principal investigators of the HAALSI study.
The studies involving human participants were reviewed and approved by University of the Witwatersrand, Johannesburg, Human Research Ethics Committee [Wits HREC (Medical)] (M141159, M121029, and M170916), the Harvard T.H. Chan School of Public Health, Office of Human Research Administration (C13-1608-02), and the Mpumalanga Provincial Research and Ethics Committee (approved: 2014/10/22). The patients/participants provided their written informed consent to participate in this study.
The study was developed by CS together with MR, AN, ST and LB. MF transformed and analysed the raw cognition data and provided information on cognitive phenotypes. CS performed the analyses and drafted the manuscript, and all other authors critically revised and approved the manuscript.
The HAALSI study was funded by the National Institute on Aging (P01 AG041710) and is nested within the Agincourt Health and Demographic Surveillance System site, supported by the University of the Witwatersrand and Medical Research Council, South Africa, and the Wellcome Trust, United Kingdom (grants no. 058893/Z/99/A; 069683/Z/02/Z; 085477/Z/08/Z; and 085477/B/08/Z). It has been carried out through a collaboration between the Harvard Center for Population and Development Studies from Harvard T.H. Chan School of Public Health, MRC/Wits Rural Public Health and Health Transitions Research Unit from School of Public Health at the University of the Witwatersrand in South Africa, and the INDEPTH Network in Accra, Ghana. The content is solely the responsibility of the authors and does not necessarily represent the official views of National Institute on Aging or the Wellcome Trust.
The AWI-Gen Study was funded by the National Human Genome Research Institute (NHGRI), Office of the Director (OD), Eunice Kennedy Shriver National Institute Of Child Health & Human Development (NICHD), the National Institute of Environmental Health Sciences (NIEHS), the Office of AIDS research (OAR) and the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK), of the National Institutes of Health (NIH) under award number U54HG006938 and its supplements, as part of the H3Africa Consortium. The study design, conclusions, and opinions expressed in this paper do not necessarily represent the official views of the National Institutes of Health.
CS was supported by the NIH grant U54HG006938. CS was funded by the National Research Foundation (NRF) Thuthuka funding instrument grant no. TTK160602167377, and the NIH Fogarty International Centre SEED funding (D43TW008330) under the umbrella of the Wits Non-Communicable Disease Research Leadership Program. The views expressed herein are those of the authors and do not necessarily reflect those of the NIH, the NRF, or the NIH Fogarty International Centre.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Busisiwe Mthembu performed the APOE genotyping under the supervision of CS. Andrew May and Innocent Maposa provided guidance and advice on the statistical methods and using R.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2021.689756/full#supplementary-material
AA, African American; AD, Alzheimer’s Disease; APOE, Apolipoprotein E Gene; AFR, 1,000 Genomes African super population group; AMR, 1,000 Genomes Admixed American super population group; AWI-Gen, Africa Wits-INDEPTH Partnership for Genomic Studies; CVDs, Cardiovascular Diseases; EAS, 1,000 Genomes East Asian super population group; EUR, 1,000 Genomes European super population group; GRCh37, Genome Reference Consortium Human genome build 37; HAALSI, The Health and Aging in Africa: A Longitudinal Study of an INDEPTH Community in South Africa; HAND, HIV-Associated Neurocognitive Disorder; HIV, Human Immunodeficiency Virus; HWE, Hardy Weinberg Equilibrium; LOAD, Late-onset Alzheimer’s Disease; MCI, Mild Cognitive Impairment; OCS-Plus, Oxford Cognition Screen Plus; SAS, 1,000 Genomes South Asian super population group; SBIMB, Sydney Brenner Institute for Molecular Bioscience; SNPs, Single-nucleotide polymorphisms; US HRS, United States Health and Retirement Study.
Ali, S. A., Soo, C., Agongo, G., Alberts, M., Amenga-Etego, L., Boua, R. P., et al. (2018). Genomic and environmental risk factors for Cardiometabolic diseases in Africa: methods used for phase 1 of the AWI-gen population cross-sectional study. Glob. Health Action 11:150733. doi: 10.1080/16549716.2018.1507133
Andres, M. A. (2011). HHS public access. J. Neuroimmune Pharmacol. 6, 389–398. doi: 10.1007/s11481-010-9254-3
Asiimwe, S. B., Farrell, M., Kobayashi, L. C., Manne-Goehler, J., Kahn, K., Tollman, S. M., et al. (2020). Cognitive differences associated with HIV Serostatus and antiretroviral therapy use in a population-based sample of older adults in South Africa. Sci. Rep. 10:16625. doi: 10.1038/s41598-020-73689-7
Auton, A., Abecasis, G. R., Altshuler, D. M., Durbin, R. M., Bentley, D. R., Chakravarti, A., et al. (2015). A global reference for human genetic variation. Nature 526, 68–74. doi: 10.1038/nature15393
Becker, J. T., Martinson, J. J., Penugonda, S., Kingsley, L., Molsberry, S., Reynolds, S., et al. (2015). No association between Apoε 4 alleles, HIV infection, age, neuropsychological outcome, or death. J. Neuro Virology 21, 24–31. doi: 10.1007/s13365-014-0290-2
Burt, T. D., Agan, B. K., Marconi, V. C., He, W., Kulkarni, H., Mold, J. E., et al. (2008). Apolipoprotein (Apo) E4 enhances HIV-1 cell entry in vitro, and the APOE Ε4/Ε4 genotype accelerates HIV disease progression. Proc. Natl. Acad. Sci. U. S. A. 105, 8718–8723. doi: 10.1073/pnas.0803526105
Chang, L., Andres, M., Sadino, J., Jiang, C. S., Nakama, H., Miller, E., et al. (2011). Impact of Apolipoprotein E Ε4 and HIV on cognition and brain atrophy: antagonistic pleiotropy and premature brain aging. NeuroImage 58, 1017–1027. doi: 10.1016/j.neuroimage.2011.07.010
Chang, L., Jiang, C., Cunningham, E., Buchthal, S., Douet, V., Andres, M., et al. (2015). Effects of Apoe4, age, and HIV on glial Metabolits and cog deficits. Neurology 82, 2213–2222. doi: 10.1212/WNL.0000000000000526
Chen, C.-H., Mizuno, T., Elston, R., Kariuki, M. M., Hall, K., Unverzagt, F., et al. (2010). A comparative study to screen dementia and APOE genotypes in an ageing east African population. Neurobiol. Aging 31, 732–740. doi: 10.1016/j.neurobiolaging.2008.06.014
Clifford, R., Wiste, H. J., Weigand, S. D., Knopman, D. S., Vemuri, P., Mielke, M. M., et al. (2015). Age, sex, and APOE Ε4 effects on memory, brain structure, and β-amyloid Across the adult life span. JAMA Neurol. 72, 511–519. doi: 10.1001/jamaneurol.2014.4821
Corbo, R. M., and Scacchp, R. (1999). Apolipoprotein E (APOE) allele distribution in the world. Is APOE*4 a “thrifty” allele? Ann. Hum. Genet. 63, 301–310. doi: 10.1046/j.1469-1809.1999.6340301.x
Cysique, L. A., Hewitt, T., Croitoru-Lamoury, J., Taddei, K., Martins, R. N., Chew, C. S. N., et al. (2015). APOE Ε4 moderates abnormal CSF-Abeta-42 levels, While neurocognitive impairment is associated with abnormal CSF tau levels in HIV+ individuals - A cross-sectional observational study. BMC Neurol. 15, 4–11. doi: 10.1186/s12883-015-0298-0
Demarchi, D. A., Salzano, M. E., Altuna, M., Fiegenbaum, M., Hill, K., Hurtado, A. M., et al. (2005). APOE Polymorphism Distribution among Native Americans and Related Populations. Ann Hum Biol. 32, 351–65. doi: 10.1080/03014460500097237
Espinosa, A., Hernández-Olasagarre, B., Moreno-Grau, S., Kleineidam, L., Heilmann-Heimbach, S., Hernández, I., et al. (2018). Exploring genetic associations of Alzheimer’s disease loci With mild cognitive impairment neurocognitive Endophenotypes. Front. Aging Neurosci. 10:340. doi: 10.3389/fnagi.2018.00340
Farrell, M. T., Kobayashi, L. C., Montana, L., Wagner, R. G., Demeyere, N., and Berkman, L. (2020). Disparity in educational attainment partially explains cognitive gender differences in older rural south Africans. J. Gerontol. B Psychol. Sci. Soc. Sci. 75, E161–E173. doi: 10.1093/geronb/gbaa035
Fitzgerald, J., Morris, D. W., and Donohoe, G. (2020). Cognitive genomics: recent advances and current challenges. Cur. Psychiatry Rep. 22:2. doi: 10.1007/s11920-019-1125-x
Fujioka, H., Phelix, C. F., Friedland, R. P., Zhu, X., Perry, E. A., Castellani, R. J., et al. (2014). Apolipoprotein E4 prevents growth of malaria at the Intraerythrocyte stage: implications For diff Erences in racial susceptibility to Alzheimer’s disease. J. Health Care Poor Underserved 24, 70–78. doi: 10.1353/hpu.2014.0009
Geffin, R., and McCarthy, M. (2018). Aging and Apolipoprotein E in HIV infection. J. Neuro Virology 24, 529–548. doi: 10.1007/s13365-018-0660-2
Gómez-Olivé, F., Montana, L., Wagner, R. G., Kabudula, C. W., Rohr, J. K., Kahn, K., et al. (2018). Cohort profile: health and ageing in Africa: A longitudinal study of an Indepth Community in South Africa (HAALSI). Int. J. Epidemiol. 47, 689–690J. doi: 10.1093/ije/dyx247
Goriounova, N. A., and Mansvelder, H. D. (2019). Genes, cells and brain areas of intelligence. Fron. Hum. Neurosci. 13:44. doi: 10.3389/fnhum.2019.00044
Gouveia, M. H., Cesar, C. C., Santolalla, M. L., Sant, H. P., Scliar, A. M. O., Leal, T. P., et al. (2019). Genetics of cognitive trajectory in Brazilians: 15 years of follow-up from the Bambuí-Epigen cohort study of aging. Sci. Rep. 9:18085. doi: 10.1038/s41598-019-53988-4
Gureje, O., Ogunniyi, A., Baiyewu, O., Price, B., Unverzagt, F. W., Evans, R. M., et al. (2006). APOE Ɛ4 is not associated with Alzheimer’s disease in elderly Nigerians. Ann. Neurol. 59, 182–185. doi: 10.1002/ana.20694
Hayat, S. A., Luben, R., Moore, S., Dalzell, N., Bhaniani, A., Anuj, S., et al. (2014). Cognitive function in a general population of men and women: A cross sectional study in the European investigation of cancer-Norfolk cohort (EPIC-Norfolk). BMC Geriatr. 14:142. doi: 10.1186/1471-2318-14-142
Hays, C. C., Zlatar, Z. Z., Meloy, M. J., Bondi, M. W., Gilbert, P. E., Liu, T. T., et al. (2019). APOE modifies the interaction of entorhinal cerebral blood flow and cortical thickness on memory function in cognitively Normal older adults. NeuroImage 202:116162. doi: 10.1016/j.neuroimage.2019.116162
Hendrie, H. C., Hall, K. S., Hui, S., Unverzagt, F. W., Yu, C. E., Lahiri, D. K., et al. (1995). Apolipoprotein E genotypes and Alzheimer’s disease in a community study of elderly African Americans. Ann. Neurol. 37, 118–120. doi: 10.1002/ana.410370123
Hendrie, H. C., Murrell, J., Baiyewu, O., Lane, K. A., Ogunniyi, A., Unverzagt, F. W., et al. (2015). APOE ε4 and the risk for Alzheimer disease and cognitive decline in African Americans and Yoruba. Int.Psychogeriatr. 26, 977–985. doi: 10.1017/S1041610214000167.APOE
Houle, B., Gaziano, T., Meagan, F., Xavier Gómez-Olivé, F., Kobayashi, L. C., Crowther, N. J., et al. (2019). Cognitive function and Cardiometabolic disease risk factors in rural South Africa: baseline evidence from the HAALSI study. BMC Public Health 19:1579. doi: 10.1186/s12889-019-7938-z
Hulgan, T., Kallianpur, A. R., Guo, Y., Barnholtz-Sloan, J. S., Gittleman, H., Brown, T. T., et al. (2018). Peripheral blood mitochondrial DNA copy number obtained From genome-wide genotype data is associated With neurocognitive impairment in persons With chronic HIV infection. J. Acquir. Immune Defic. Syndr. 80, e95–e102. doi: 10.1097/QAI.0000000000001930
Humphreys, G. W., Duta, M. D., Montana, L., Demeyere, N., McCrory, C., Rohr, J., et al. (2017). Cognitive function in low-income and low-literacy settings: validation of the tablet-based Oxford cognitive screen in the health and aging in Africa: A longitudinal study of an INDEPTH Community in South Africa (HAALSI). J. Gerontology. B Psychol. Scie. Soc. Sci. 72, 38–50. doi: 10.1093/geronb/gbw139
Joska, J. A., Combrinck, M., Valcour, V. G., Hoare, J., Leisegang, F., Mahne, A. C., et al. (2010). Association between Apolipoprotein E4 genotype and human immunodeficiency virus-associated dementia in younger adults starting antiretroviral therapy in South Africa. J. Neurovirol. 16, 377–383. doi: 10.3109/13550284.2010.513365
Kabudula, C. W., Tollman, S., Mee, P., Ngobeni, S., Bernard, S., Xavier Gómez-Olivé, F., et al. (2014). Two decades of mortality change in rural Northeast South Africa. Glob. Health Action 7:25596. doi: 10.3402/gha.v7.25596
Kahn, K., Collinson, M. A., Xavier Gómez-olivé, F., Mokoena, O., Twine, R., Mee, P., et al. (2012). Profile: Agincourt health and socio-demographic surveillance system. Int. J. Epidemiol. 41, 988–1001. doi: 10.1093/ije/dys115
Kobayashi, L. C., Mateen, F. J., Montana, L., Wagner, R. G., Kahn, K., Tollman, S. M., et al. (2019). Cognitive function and impairment in older, rural south African adults: evidence from “health and aging in Africa: A longitudinal study of an INDEPTH Community in Rural South Africa”. Neuroepidemiology 52, 32–40. doi: 10.1159/000493483
Kotze, M., Brand, T., Pretorius, J., van Rensburg, S. J., and Luckhoff, H. K. (2015). Apolipoprotein E ε-4 as a genetic determinant of Alzheimer’s disease heterogeneity. Degenerative Neurol. Neuromuscular Dis. 9, 9–18. doi: 10.2147/dnnd.s41721
Kuhlmann, I., Minihane, A. M., Huebbe, P., Nebel, A., and Rimbach, G. (2010). Apolipoprotein E genotype and hepatitis C, HIV and herpes simplex disease risk: A literature review. Lipids Health Dis. 9:8. doi: 10.1186/1476-511X-9-8
Kuo, C. L., Pilling, L. C., Atkins, J. L., Masoli, J. A. H., Delgado, J., Kuchel, G. A., et al. (2020). ApoE E4e4 genotype and mortality with COVID-19 in UK biobank. J. Gerontol A Biol. Sci.Med. Sci. 75, 1801–1803. doi: 10.1093/gerona/glaa169
Lipnicki, D. M., Crawford, J. D., Dutta, R., Thalamuthu, A., Kochan, N. A., Gavin, A., et al. (2017). Age-related cognitive decline and associations with sex, education and Apolipoprotein E genotype across Ethnocultural groups and geographic regions: A collaborative cohort study. PLoS Med. 14:e1002261. doi: 10.1371/journal.pmed.1002261
Lipnicki, D. M., Makkar, S. R., Crawford, J. D., Thalamuthu, A., Kochan, N. A., Lima-Costa, M. F., et al. (2019). Determinants of cognitive performance and decline in 20 diverse ethno-regional groups: A COSMIC collaboration cohort study. PLoS Med. 16:e1002853. doi: 10.1371/journal.pmed.1002853
Liu, C.-C., Kanekiyo, T., Xu, H., and Guojun, B. (2013). Apolipoprotein E and Alzheimer disease: risk, mechanisms and therapy. Nat. Rev. Neurol. 9, 106–118. doi: 10.1038/nrneurol.2012.263
Morgan, E. E., Woods, S. P., Letendre, S. L., Franklin, D. R., Bloss, C., Goate, A., et al. (2013). Apolipoprotein E4 genotype does not increase risk of HIV-associated neurocognitive disorders. J. NeuroVirology 19, 150–156. doi: 10.1007/s13365-013-0152-3
Mueller, T., Fischer, J., Gessner, R., Rosendahl, J., Böhm, S., van Bömmel, F., et al. (2016). Apolipoprotein E allele frequencies in chronic and self-limited hepatitis C suggest a protective effect of APOE4 in the course of hepatitis C virus infection. Liver Int. 36, 1267–1274. doi: 10.1111/liv.13094
O’Donoghue, M. C., Murphy, S. E., Zamboni, G., Nobre, A. C., and Mackay, C. E. (2018). APOE genotype and cognition in healthy individuals at risk of Alzheimer’s disease: A review. Cortex 104, 103–123. doi: 10.1016/j.cortex.2018.03.025
Osuntokun, B. O., Sahota, A., Ogunniyi, A. O., Gureje, O., Baiyewu, O., Adeyinka, A., et al. (1995). Lack of an association between Apolipoprotein E ε 4 and Alzheimer’s disease in elderly Nigerians. Ann. Neurol. 38, 463–465. doi: 10.1002/ana.410380319
Panos, S. E., Hinkin, C. H., Singer, E. J., Thames, A. D., Patel, S. M., Sinsheimer, J. S., et al. (2013). Apolipoprotein-E genotype and human immunodeficiency virus-associated neurocognitive disorder: The modulating effects of older age and disease severity. Neurobehav. HIV Med. 5, 11–22. doi: 10.2147/NBHIV.S39573
R Core Team (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. Available at: https://www.R-project.org/
Ramsay, M., Crowther, N., Tambo, E., Agongo, G., Baloyi, V., Dikotope, S., et al. (2016). H3Africa AWI-gen collaborative Centre: A resource to study the interplay between genomic and environmental risk factors for Cardiometabolic diseases in four sub-Saharan African countries. Glob. Health, Epidemiol.Genom. 1:e20. doi: 10.1017/gheg.2016.17
Reas, E. T., Laughlin, G. A., Bergstrom, J., Kritz-Silverstein, D., Barrett-Connor, E., and McEvoy, L. K. (2019). Effects of APOE on cognitive aging in community-dwelling older adults. Neuropsychology 33, 406–416. doi: 10.1037/neu0000501
Rebeck, G. W. (2017). The role of APOE on lipid homeostasis and inflammation in Normal brains. J. Lipid Res. 58, 1493–1499. doi: 10.1194/jlr.R075408
Rosenberg, M., Xavier Gómez-Olivé, F., Wagner, R. G., Rohr, J., Payne, C. F., Berkman, L., et al. (2020). The relationships between cognitive function, literacy and HIV status knowledge among older adults in rural South Africa. J. Int. AIDS Soc. 23:e25457. doi: 10.1002/jia2.25457
Sandholzer, C., Delport, R., Vermaak, H., and Utermann, G. (1995). High Frequency of the Apo Ɛ4 Allele in Khoi San from South Africa. Hum. Genet. 95, 46–48. doi: 10.1007/BF00225073
Savitz, J., Solms, M., and Ramesar, R. (2006). Apolipoprotein E variants and cognition in healthy individuals: A critical opinion. Brain Res. Rev. 51, 125–135. doi: 10.1016/j.brainresrev.2005.10.006
Siedlecki, K. L., Honig, L. S., and Stern, Y. (2008). Exploring the structure of a neuropsychological battery across healthy elders and Those with questionable dementia and Alzheimer’s disease. Neuropsychology 22, 400–411. doi: 10.1037/0894-4105.22.3.400
Singh, P. P., Singh, M., and Mastana, S. S. (2006). APOE Distribution in World Populations with New Data from India and the UK. Ann. Hum. Biol. 33, 279–308. doi: 10.1080/03014460600594513
Small, B. J., Rosnick, C. B., Fratiglioni, L., and Bäckman, L. (2004). Apolipoprotein E and cognitive performance: A meta-analysis. Psychol. Aging 19, 592–600. doi: 10.1037/0882-7974.19.4.592
Smith, C. J., Wesson Ashford, J., and Perfetti, T. A. (2019). Putative survival advantages in young Apolipoprotein Ɛ4 carriers are associated with increased neural stress. J. Alzheimers Dis. 68, 885–923. doi: 10.3233/JAD-181089
Sundermann, E. E., Maki, P. M., Rubin, L. H., Lipton, R. B., Landau, S., Biegon, A., et al. (2016). Female advantage in verbal memory: evidence of sex-specific cognitive reserve. Neurology 87, 1916–1924. doi: 10.1212/WNL.0000000000003288
Valcour, V., Shikuma, C., Shiramizu, B., Watters, M., Poff, P., Selnes, O. A., et al. (2004). Age, Apolipoprotein E4, and the risk of HIV dementia: The Hawaii aging with HIV cohort. J. Neuroimmunology 157, 197–202. doi: 10.1016/j.jneuroim.2004.08.029
van Exel, E., Koopman, J. J. E., Van Bodegom, D., Meij, J. J., De Knijff, P., Ziem, J. B., et al. (2017). Effect of APOE Ε4 allele on survival and fertility in an adverse environment. PLoS One 12:e0179497. doi: 10.1371/journal.pone.0179497
Wendelken, L. A., Jahanshad, N., Rosen, H. J., Busovaca, E., Allen, I., Coppola, G., et al. (2016). ApoE Ε4 is associated with cognition, brain integrity, and atrophy in HIV Over age 60. J. Acquir. Immune Defic. Syndr. 73, 426–432. doi: 10.1097/QAI.0000000000001091
Willis, F., Graff-Radford, N., Pinto, M., Lawson, L. S., Adamson, J., Epstein, D., et al. (2003). Apolipoprotein ∈4 allele frequency in young Africans of Ugandan descent versus African Americans. J. Natl. Med. Assoc. 95, 71–76.
Wisdom, N. M., Callahan, J. L., and Hawkins, K. A. (2011). The effects of Apolipoprotein E on non-impaired cognitive functioning: A meta-analysis. Neurobiol. Aging 32, 63–74. doi: 10.1016/j.neurobiolaging.2009.02.003
Keywords: apolipoprotein E 𝝴4, cognitive function, African population, educational attainment, memory, executive function, visuospatial ability, language
Citation: Soo CC, Farrell MT, Tollman S, Berkman L, Nebel A and Ramsay M (2021) Apolipoprotein E Genetic Variation and Its Association With Cognitive Function in Rural-Dwelling Older South Africans. Front. Genet. 12:689756. doi: 10.3389/fgene.2021.689756
Edited by:
Brad Verhulst, Texas A&M University, United StatesReviewed by:
Karen Anne Mather, University of New South Wales, AustraliaCopyright © 2021 Soo, Farrell, Tollman, Berkman, Nebel and Ramsay. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cassandra C. Soo, Y2Fzc2FuZHJhLnNvb0B3aXRzLmFjLnph; YXJpbW9tby5zYW5AZ21haWwuY29t; c29vLmNhc3NhbmRyYS5jQGdtYWlsLmNvbQ==; Michèle Ramsay, bWljaGVsZS5yYW1zYXlAd2l0cy5hYy56YQ==
†These authors have contributed equally to this work and share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.