 Jingwen Yin1,2,3,4†
Jingwen Yin1,2,3,4† Xudong Luo1†
Xudong Luo1† Qian Peng1†
Qian Peng1† Susu Xiong1
Susu Xiong1 Dong Lv1
Dong Lv1 Zhun Dai1
Zhun Dai1 Jiawu Fu5
Jiawu Fu5 Ying Wang1Yaxue Wei1Chunmei Liang5
Ying Wang1Yaxue Wei1Chunmei Liang5 Xusan Xu5Dandan Zhang1Lulu Wang1
Xusan Xu5Dandan Zhang1Lulu Wang1 Dongjian Zhu1
Dongjian Zhu1 Xia Wen5Xiaoqing Ye1
Xia Wen5Xiaoqing Ye1 Zhixiong Lin1
Zhixiong Lin1 Juda Lin1You Li5
Juda Lin1You Li5 Jiafeng Wang6
Jiafeng Wang6 Guoda Ma5,7*
Guoda Ma5,7* Keshen Li5,7*
Keshen Li5,7* Yajun Wang7*
Yajun Wang7*- 1Department of Psychiatry, Affiliated Hospital of Guangdong Medical University, Zhanjiang, China
- 2Center for Cognitive and Brain Sciences, Institute of Collaborative Innovation, University of Macau, Taipa, China
- 3Department of Psychology, Faculty of Social Sciences, University of Macau, Taipa, China
- 4Maternal and Children’s Health Research Institute, Shunde Maternal and Children’s Hospital, Guangdong Medical University, Foshan, China
- 5Institute of Neurology, Guangdong Medical University, Zhanjiang, China
- 6Stem Cell Research and Cellular Therapy Center, Affiliated Hospital of Guangdong Medical University, Zhanjiang, China
- 7Guangdong Key Laboratory of Age-Related Cardiac and Cerebral Diseases, Affiliated Hospital of Guangdong Medical University, Zhanjiang, China
Objective: To investigate the effects of microRNA-137 (MIR137) polymorphisms (rs1198588 and rs2660304) on the risk of schizophrenia in a Han Chinese population.
Methods: Schizophrenia was diagnosed according to the DSM-5. Clinical symptoms and cognitive functions were assessed with the Positive and Negative Symptom Scale (PANSS) and Brief Assessment of Cognition in Schizophrenia (BACS), respectively. The polymorphisms were genotyped by improved multiplex ligation detection reaction (iMLDR) technology in 1,116 patients with schizophrenia and 1,039 healthy controls.
Results: Significant associations were found between schizophrenia and MIR137 in the distributions of genotypes (p = 0.037 for rs1198588; p = 0.037 for rs2660304, FDR corrected) and alleles (p = 0.043 for rs1198588; p = 0.043 for rs2660304, FDR corrected) of two SNPs. When the population was stratified by sex, we found female-specific associations between MIR137 and schizophrenia in terms of genotype and allele distributions of rs1198588 (χ2 = 4.41, p = 0.036 and χ2 = 4.86, p = 0.029, respectively, FDR corrected) and rs2660304 (χ2 = 4.74, p=0.036 and χ2 = 4.80, p = 0.029, respectively, FDR corrected). Analysis of the MIR137 haplotype rs1198588-rs2660304 showed a significant association with schizophrenia in haplotype T-T [χ2 = 4.60, p = 0.032, OR = 1.32, 95% CI (1.02–1.70)]. Then, significant female-specific associations were found with the haplotypes T-T and G-A [χ2 = 4.92, p = 0.027, OR = 1.62, 95% CI (1.05–2.50); χ2 = 4.42, p = 0.035, OR = 0.62, 95% CI (0.39–0.97), respectively]. When the TT genotype of rs1198588 was compared to the GT+GG genotype, a clinical characteristics analysis also showed a female-specific association in category instances (t = 2.76, p = 0.042, FDR corrected).
Conclusion: The polymorphisms within the MIR137 gene are associated with susceptibility to schizophrenia, and a female-specific association of MIR137 with schizophrenia was reported in a Han Chinese population.
Introduction
Based on converging evidence from several research disciplines, it has been generally accepted that schizophrenia is a disease characterized by a nervous system dysfunction caused by the “accumulation” of dysregulation events in multiple candidate genes (Zhu et al., 2016). These susceptible genes can act on several levels, from the gene product itself through molecular, cellular, and tissue levels, all the way to complex phenotypes such as psychotic symptoms and cognitive deficits (Dobbyn et al., 2018). Systematically understanding how genes act to confer risk for schizophrenia is an obvious strategy in biological psychiatry.
miR-137 is highly expressed in the brain and has important roles in neurogenesis, neuronal maturation, and signal transduction (Guella et al., 2012; Wang et al., 2020). The dysregulation of miR-137 usually causes micro-abnormalities in neural structure and function, which might lead to brain function disorders, including schizophrenia. Clinical research further provided evidence of 2.63-fold higher miR-137 expression in the peripheral blood of schizophrenia patients compared with healthy controls (Wu et al., 2016). Transgenic mouse overexpression of MIR137 in the whole brain induces several phenotypes that are relevant to aspects of psychiatric disorders, including sensory gating deficits and social and cognitive deficits (Arakawa et al., 2019). In addition, MIR137 has also been identified as specifically regulating genes with replicated evidence for a role in schizophrenia, most notably N-methyl-D-aspartate (NMDA) receptor (Strazisar et al., 2014), glutamate ionotropic receptor NMDA type subunit 2A (Grin2A; Zhao et al., 2013), GluA1 subunit of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors (AMPARs; Olde Loohuis et al., 2015), zinc finger binding protein 804A (ZNF804A; Kim et al., 2012), transcription factor 4 (TCF4; Wright et al., 2013), brain-derived neurotrophic factor (BDNF; Hill et al., 2014; Thomas et al., 2017), and glyoxalase 1 (Hambsch, 2011; Bangel et al., 2015; Lv et al., 2018). Overall, MIR137 appears to be an integral part of the biological pathways of schizophrenia and a component of the genetic architecture of the complex phenotypes of schizophrenia.
Genome-wide association studies (GWAS) have suggested that rs1625579 of MIR137 was the strongest locus associated with schizophrenia (Ripke et al., 2011). In our previous study, rs1625579 was reported as a risk SNP in the southern Chinese population (Ma et al., 2014). SNP rs2660304, located in the promoter (4.0 kb upstream) of the primary transcript sequence of the MIR137 host gene, has strong linkage disequilibrium (LD) with rs1625579. Siegert (2015) and Warburton et al. (2016) observed increased transcriptional and expression levels of miR-137 in the minor allele-carrying cells of rs2660304 compared to the major allele. Rs1198588 is also a GWAS-suggested locus that lies within a noncoding genomic but open chromatin region and can alter miR-137 expression in an epigenetic manner (Ripke et al., 2013; Siegert et al., 2015; Forrest et al., 2017). To further assess the associations of MIR137 SNPs with the risk of schizophrenia, we conducted a case-control study in the Han Chinese population.
Subjects and Methods
Subjects
A total of 1,116 schizophrenic patients and 1,039 healthy controls were recruited from the Affiliated Hospital of Guangdong Medical University, and all patients were from the Han population. All patients underwent a series of standardized examinations, including family history, an evaluation regarding a wide range of drugs and alcohol, a physical and neurological examination, and laboratory testing, to eliminate those with a drug-induced psychotic disorder or psychosis resulting from general medical conditions. The diagnosis was made independently by two well-trained senior psychiatrists based on the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5). Psychiatric symptoms were assessed using a structured interview by the Positive and Negative Symptom Scale (PANSS). Evaluations of the cognitive function of patients were performed by the Brief Assessment of Cognition in Schizophrenia (BACS; Keefe et al., 2004). The BACS includes the following six parts: working memory measured by the digit sequencing task, verbal memory measured by list learning, motor speed measured by the token motor task, reasoning and problem solving measured by the Tower of London, attention and processing speed, and verbal fluency measured by symbol coding. None of the healthy controls had a personal or family history of major mental disease, severe physical illness, or substance abuse. The study protocol was approved by the ethics committee of the Affiliated Hospital of Guangdong Medical University, and written informed consent was obtained from subjects or immediate family members.
DNA Extraction and Genotyping
Genomic DNA from EDTA-anticoagulated peripheral blood was extracted using the TIANamp Blood DNA Kit (Tiangen Biotech, Beijing, China). The rs1198588 and rs2660304 SNPs were genotyped using the improved multiplex ligation detection reaction (iMLDR) method (Genesky Biotechnologies Inc., Shanghai, China) as described previously (Xu et al., 2018; Yin et al., 2019). The primer information for the multiplex polymerase chain reaction and ligation is described in Supplementary Table S1.
Statistical Analysis
Statistical analysis was performed by SPSS 21.0 software. Descriptive data are presented as the mean ± standard deviation (SD) and frequencies (%) as appropriate. Pearson’s chi-square test was used to assess Hardy-Weinberg equilibrium (HWE) and the differences in genotypic and allelic distributions between patients and controls. Then, generalized odds ratios (ORs) with 95% confidence intervals (CIs) of the alleles were calculated. In the association analysis between genotypes and clinical characteristics, categorical variables were compared by Pearson’s chi-square tests, and continuous variables were compared by Student’s t test between two independent groups. The normalized linkage disequilibrium coefficient (D') and squared correlation coefficient (r2) were used to measure LD. The LD status and haplotype analysis were determined using Haploview 4.2 software. Only those haplotypes with frequencies greater than 3% were further analyzed. Multiple comparisons were performed with false discovery rate (FDR) correction in R and R studio software. Power calculations were performed using QUANTO 1.2 software. A value of 0.05 was used as a threshold for statistical significance after correction for multiple comparisons.
Results
There were no statistically significant differences in the age and sex distribution of the MIR137 polymorphisms between the patient group and the control group, as shown in Supplementary Table S2. The distributions of the two loci in the controls were consistent with Hardy-Weinberg equilibrium, as shown in Table 1. Significant differences were observed in the genotype and allele distributions of rs1198588 (χ2 = 4.37, p = 0.037 and χ2 = 4.57, p = 0.043, respectively, FDR corrected) and rs2660304 (χ2 = 4.42, p = 0.037 and χ2 = 4.10, p = 0.043, respectively, FDR corrected) of MIR137, as shown in Table 1.
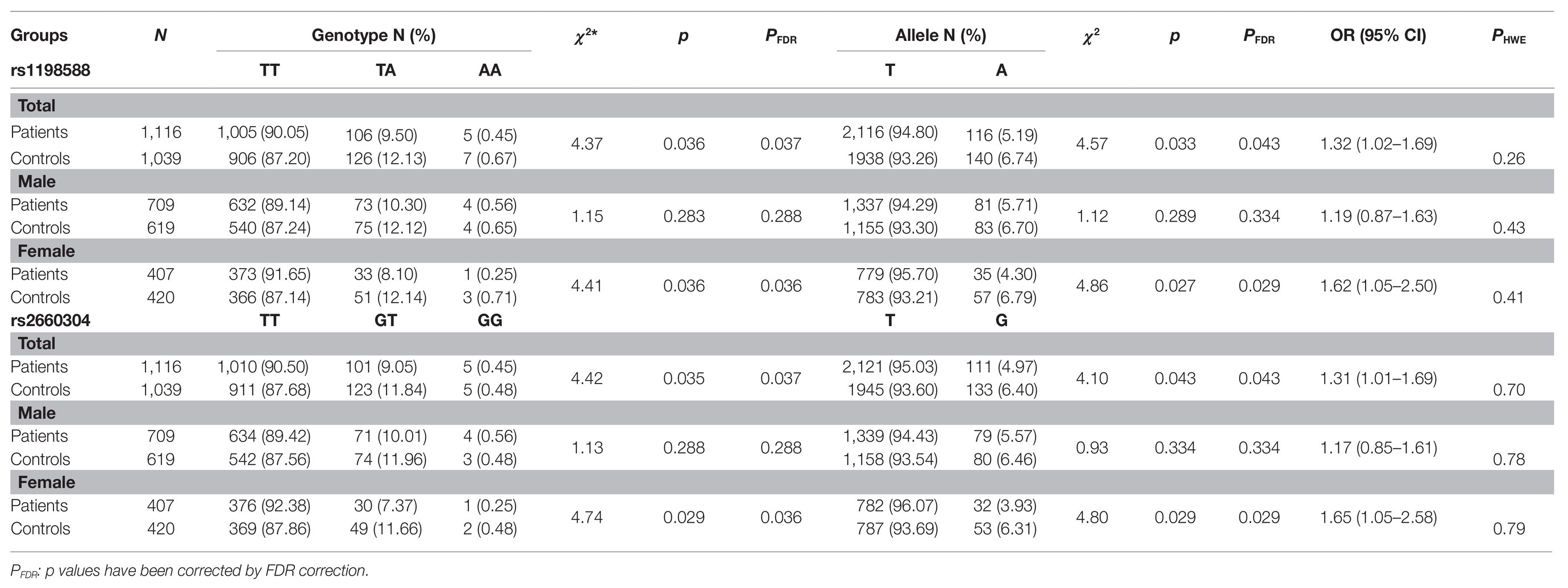
Table 1. Genotype and allele frequencies of SNPs in MIR137 in schizophrenic patients and controls.
Furthermore, to examine the possible potential effect of sex on the associations between the SNPs and schizophrenia, we stratified the participants into male and female subgroups. Significant differences were maintained in the genotype and allele distributions of rs1198588 (χ2 = 4.41, p = 0.036 and χ2 = 4.86, p = 0.029, respectively, FDR corrected) and rs2660304 (χ2 = 4.74, p=0.036 and χ2 = 4.80, p = 0.029, respectively, FDR corrected) in the female subgroup but vanished in the male subgroup (p > 0.05; Table 1).
LD analysis was performed in two loci of MIR137. Strong LD was observed in rs1198588 and rs2660304 (D' = 0.996, r2 = 0.94). The association of risk SNPs with schizophrenia was further supported by the results of the haplotype analysis, and the corresponding results are shown in Table 2. The frequency of the T-T haplotype (rs2660304-rs1198588) was significantly higher in the schizophrenia patients than in the controls [χ2 = 4.60, p = 0.032, OR = 1.32, 95% CI (1.02–1.70)]. When the population was stratified by sex, a significant association was also evidenced only in the female subgroup, wherein the overall association was significant [χ2 = 4.92, p = 0.027, OR = 1.62, 95% CI (1.05–2.50)], and no such association was observed in the male subgroup [χ2 = 1.12, p = 0.289, OR = 1.19, 95% CI (0.87–1.63)]. Thus, the haplotype was shown to be a female-specific risk factor for schizophrenia. Furthermore, our results showed that the G-A haplotype was a protective factor against schizophrenia only in the female group and not in the male group [χ2 = 4.42, p = 0.035, OR = 0.62, 95% CI (0.39–0.97); Table 2].
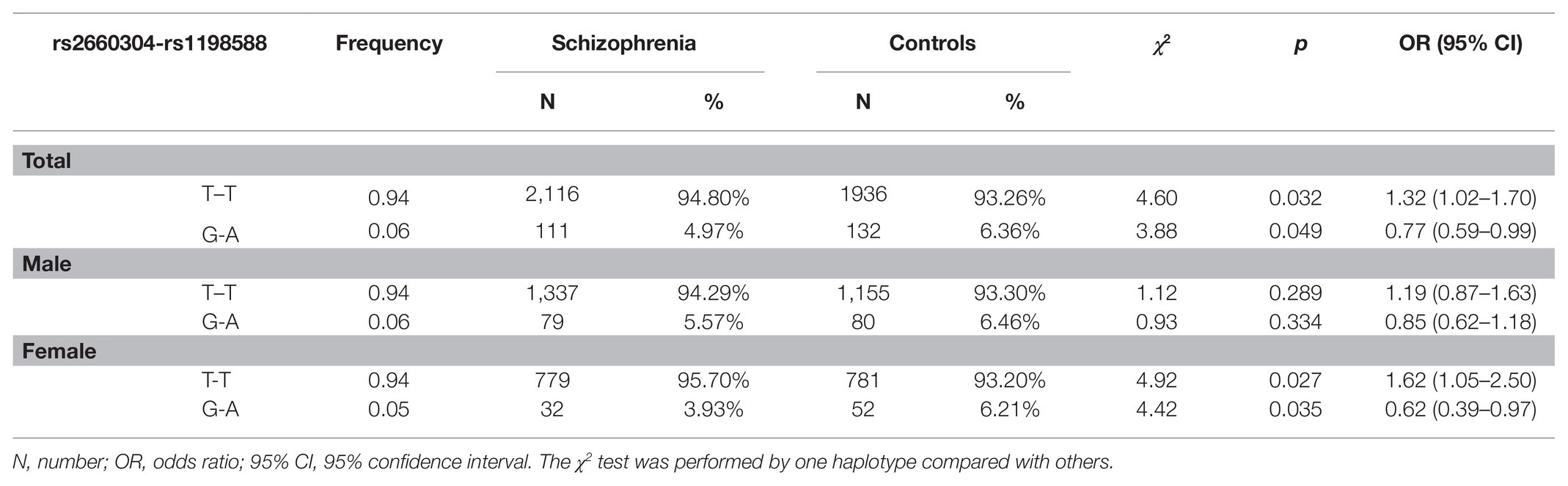
Table 2. Haplotype analysis of SNPs in MIR137 in schizophrenic patients and controls.
Using the BACS, we analyzed the neurocognitive function of the patients, and significant differences were found in category instances when the TT genotypes of rs1198588 and rs2660304 were compared to the GT+GG genotype in the female group, but only the comparison in rs1198588 survived after FDR correction (t = 2.76, p = 0.042). Otherwise, differences were shown in token motor task scores in males when the TT genotype was compared to G carriers in both rs1198588 and rs2660304, but these differences did not survive multiple comparison correction (shown in Supplementary Table S3). Our study further analyzed the distribution of other clinical characteristics among different genotypes in the patients, but there were no significant differences in age of onset, family history, or PANSS scores (p > 0.05).
Discussion
The MIR137 host gene was most strongly associated with schizophrenia in GWAS (Ripke et al., 2011). MIR137 is highly and specifically expressed in the brain (Supplementary Figure S1; data were downloaded from the GTEx database) and regulates neural structure and function (Yin et al., 2014; Mahmoudi and Cairns, 2017). Increasing evidence has shown that MIR137 and its gene regulatory network may be involved in the genetic and biological basis of schizophrenia and have been associated with phenotypes in schizophrenia, such as age of onset (Lett et al., 2013) and brain structure and function (Whalley et al., 2012; Lett et al., 2013; Vogel et al., 2018; Zhang et al., 2018). Rs1198588 and rs2660304 are located at 41.1 and 4.0 kb, respectively, upstream of the MIR137 host gene region and are related to the transcript regulation and expression of miR-137 (Warburton et al., 2016; Forrest et al., 2017). Rs2660304 resulted in downregulation of reporter gene expression in a tissue culture model (Warburton et al., 2016; Tian et al., 2019). The LD analysis indicated that rs1198588 and rs2660304 might be proxies of rs1625579. In our sample, rs1625579 also showed LD with rs1198588 (D' = 0.99, r2 = 0.94) and rs2660304 (D' = 1, r2 = 0.99; calculated by combining the data set with previous results). We performed a case-control study to primarily investigate rs1198588 and rs2660304 in the risk of schizophrenia in a Han population from southern China. The results indicated that both are associated with the risk of schizophrenia. At present, the biological mechanisms of MIR137 rs1198588 and rs2660304 polymorphisms are still unclear, but the results of our case-control study provide convincing evidence for the association between MIR137 polymorphisms and schizophrenia. Power analysis showed that the sample has 95% power to detect relative risk with an assumed odds ratio of 1.5 at the 0.05 level.
Interesting results in our study were sex differences in the MIR137 genotype and haplotype, which were relatively pronounced in females but not significant in males. It is perhaps not surprising, considering the substantial evidence for sex differences in the etiology and pathogenesis of schizophrenia (Aleman et al., 2003; Kulkarni et al., 2012). Previous studies have presented female-specific effects on MIR137 and target genes of MIR137. Kandratsenka et al. (2018) reported female-specific effects on negative symptom scores and the total PANSS scores of the rs1625579 polymorphism of MIR137 in a Belarusian population. Considering the high LD in rs1625579 and rs1198588, the sex-specific differences with rs1198588 failed to be repeated in the PANSS scores in our sample, but differences were found in cognitive function. A possible explanation might be the heterogeneity in symptoms and genetic distribution related to ethnicity. Interestingly, Zhang et al. (2011) provided evidence that ZNF804A, a target gene of MIR137, also showed a female-specific association with schizophrenia. In addition, Glo-1, another target gene of MIR137, has also been reported to be involved in the regulation of sex hormones. This evidence suggests a possible pathway by which sex hormones modulate the expression of miR-137 and its targets, which may account for the observed sex-specific effects. Functional and expression studies are necessary to further clarify this hypothesis, and the result needs to be replicated in different independent samples, since there is a possibility of population bias. The limited sample size for the individual sexes was not large enough, and the sex-genotype interaction has not been formally tested.
The present study has several potential limitations. First, the sample in this study was limited to the Han Chinese population. Further investigations of the association between MIR137 polymorphisms and schizophrenia need to be carried out in different ethnic populations. Second, we observed that differences between genotypes and haplotype frequency distributions were common, specifically among females in our study groups. Thus, a further functional analysis is needed to clarify how variants in MIR137 play a female-specific role in the pathogenesis of schizophrenia.
In conclusion, to our knowledge, this study is the first to show that MIR137 SNPs are significantly associated with schizophrenia in the Han Chinese population. Moreover, our study provides further evidence for a female-specific influence of the gene on the development of schizophrenia.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of Guangdong Medical University. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by the National Nature Science Foundation of China (81670252, 81770034) and the Guangdong Basic and Applied Basic Research Foundation (2019A1515011306).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2021.627874/full#supplementary-material
References
Aleman, A., Kahn, R. S., and Selten, J. P. (2003). Sex differences in the risk of schizophrenia: evidence from meta-analysis. Arch. Gen. Psychiatry 60, 565–571. doi: 10.1001/archpsyc.60.6.565
Arakawa, Y., Yokoyama, K., Tasaki, S., Kato, J., Nakashima, K., Takeyama, M., et al. (2019). Transgenic mice overexpressing miR-137 in the brain show schizophrenia-associated behavioral deficits and transcriptome profiles. PLoS One 14:e0220389. doi: 10.1371/journal.pone.0220389
Bangel, F. N., Yamada, K., Arai, M., Iwayama, Y., Balan, S., Toyota, T., et al. (2015). Genetic analysis of the glyoxalase system in schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 59, 105–110. doi: 10.1016/j.pnpbp.2015.01.014
Dobbyn, A., Huckins, L. M., Boocock, J., Sloofman, L. G., Glicksberg, B. S., Giambartolomei, C., et al. (2018). Landscape of conditional eQTL in dorsolateral prefrontal cortex and co-localization with schizophrenia GWAS. Am. J. Hum. Genet. 102, 1169–1184. doi: 10.1016/j.ajhg.2018.04.011
Forrest, M. P., Zhang, H., Moy, W., McGowan, H., Leites, C., Dionisio, L. E., et al. (2017). Open chromatin profiling in hiPSC-derived neurons prioritizes functional noncoding psychiatric risk variants and highlights neurodevelopmental loci. Cell Stem Cell 21, 305–318. doi: 10.1016/j.stem.2017.07.008
Guella, I., Sequeira, A., Rollins, B., Torri, F., Vawter, M. P., and MacCiardi, F. (2012). miR-137 expression in schizophrenia and bipolar disorder brains. Neuroscience Meeting Planner in Program No. 223.10.
Hambsch, B. (2011). Altered glyoxalase 1 expression in psychiatric disorders: cause or consequence? Semin. Cell Dev. Biol. 22, 302–308. doi: 10.1016/j.semcdb.2011.02.005
Hill, M. J., Donocik, J. G., Nuamah, R. A., Mein, C. A., Sainz-Fuertes, R., and Bray, N. J. (2014). Transcriptional consequences of schizophrenia candidate miR-137 manipulation in human neural progenitor cells. Schizophr. Res. 153, 225–230. doi: 10.1016/j.schres.2014.01.034
Kandratsenka, H., Nestsiarovich, A., Goloenko, I., Danilenko, N., Makarevich, A., Obyedkov, V., et al. (2018). Association of MIR137 with symptom severity and cognitive functioning in Belarusian schizophrenia patients. Front. Psych. 9:295. doi: 10.3389/fpsyt.2018.00295
Keefe, R. S., Goldberg, T. E., Harvey, P. D., Gold, J. M., Poe, M. P., and Coughenour, L. (2004). The brief assessment of cognition in schizophrenia: reliability, sensitivity, and comparison with a standard neurocognitive battery. Schizophr. Res. 68, 283–297. doi: 10.1016/j.schres.2003.09.011
Kim, A. H., Parker, E. K., Williamson, V., McMichael, G. O., Fanous, A. H., and Vladimirov, V. I. (2012). Experimental validation of candidate schizophrenia gene ZNF804A as target for hsa-miR-137. Schizophr. Res. 141, 60–64. doi: 10.1016/j.schres.2012.06.038
Kulkarni, J., Hayes, E., and Gavrilidis, E. (2012). Hormones and schizophrenia. Curr. Opin. Psychiatry 25, 89–95. doi: 10.1097/YCO.0b013e328350360e
Lett, T. A., Chakavarty, M. M., Felsky, D., Brandl, E. J., Tiwari, A. K., Goncalves, V. F., et al. (2013). The genome-wide supported microRNA-137 variant predicts phenotypic heterogeneity within schizophrenia. Mol. Psychiatry 18, 443–450. doi: 10.1038/mp.2013.17
Lv, N., Hao, S., Luo, C., Abukiwan, A., Hao, Y., Gai, F., et al. (2018). miR-137 inhibits melanoma cell proliferation through downregulation of GLO1. Sci. China Life Sci. 61, 541–549. doi: 10.1007/s11427-017-9138-9
Ma, G., Yin, J., Fu, J., Luo, X., Zhou, H., Tao, H., et al. (2014). Association of a miRNA-137 polymorphism with schizophrenia in a Southern Chinese Han population. Biomed. Res. Int. 2014:751267. doi: 10.1155/2014/751267
Mahmoudi, E., and Cairns, M. J. (2017). MiR-137: an important player in neural development and neoplastic transformation. Mol. Psychiatry 22, 44–55. doi: 10.1038/mp.2016.150
Olde Loohuis, N. F., Ba, W., Stoerchel, P. H., Kos, A., Jager, A., Schratt, G., et al. (2015). MicroRNA-137 controls AMPA-receptor-mediated transmission and mGluR-dependent LTD. Cell Rep. 11, 1876–1884. doi: 10.1016/j.celrep.2015.05.040
Ripke, S., O’Dushlaine, C., Chambert, K., Moran, J. L., Kähler, A. K., Akterin, S., et al. (2013). Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat. Genet. 45, 1150–1159. doi: 10.1038/ng.2742
Ripke, S., Sanders, A., Kendler, K., Levinson, D., Sklar, P., and Holmans, P. (2011). Genome-wide association study identifies five new schizophrenia loci. Nat. Genet. 43, 969–976. doi: 10.1038/ng.940
Siegert, S., Seo, J., Kwon, E. J., Rudenko, A., Cho, S., Wang, W., et al. (2015). The schizophrenia risk gene product miR-137 alters presynaptic plasticity. Nat. Neurosci. 18, 1008–1016. doi: 10.1038/nn.4023
Strazisar, M., Cammaerts, S., van der Ven, K., Forero, D. A., Lenaerts, A. S., Nordin, A., et al. (2014). MIR137 variants identified in psychiatric patients affect synaptogenesis and neuronal transmission gene sets. Mol. Psychiatry 20, 472–481. doi: 10.1038/mp.2014.53
Thomas, K. T., Anderson, B. R., Shah, N., Zimmer, S. E., Hawkins, D., Valdez, A. N., et al. (2017). Inhibition of the schizophrenia-associated MicroRNA miR-137 disrupts Nrg1α neurodevelopmental signal transduction. Cell Rep. 20, 1–12. doi: 10.1016/j.celrep.2017.06.038
Tian, Z., Yang, Y., Feng, Z., Wu, D., Yang, W., and Liu, D. (2019). Genetic variant in the promoter region of microRNA-137 reduces the warfarin maintenance dose in patients with atrial fibrillation. Mol. Med. Rep. 19, 5361–5367. doi: 10.3892/mmr.2019.10205
Vogel, B. O., Lett, T. A., Erk, S., Mohnke, S., Wackerhagen, C., Brandl, E. J., et al. (2018). The influence of MIR137 on white matter fractional anisotropy and cortical surface area in individuals with familial risk for psychosis. Schizophr. Res. 195, 190–196. doi: 10.1016/j.schres.2017.09.030
Wang, Y., Chen, R., Zhou, X., Guo, R., Yin, J., Li, Y., et al. (2020). miR-137: a novel therapeutic target for human glioma. Mol. Ther. Nucleic Acids 21, 614–622. doi: 10.1016/j.omtn.2020.06.028
Warburton, A., Breen, G., Bubb, V. J., and Quinn, J. P. (2016). A GWAS SNP for schizophrenia is linked to the internal MIR137 promoter and supports differential allele-specific expression. Schizophr. Bull. 42, 1003–1008. doi: 10.1093/schbul/sbv144
Whalley, H. C., Papmeyer, M., Romaniuk, L., Sprooten, E., Johnstone, E. C., Hall, J., et al. (2012). Impact of a microRNA MIR137 susceptibility variant on brain function in people at high genetic risk of schizophrenia or bipolar disorder. Neuropsychopharmacology 37, 2720–2729. doi: 10.1038/npp.2012.137
Wright, C., Turner, J. A., Calhoun, V. D., and Perrone-Bizzozero, N. (2013). Potential impact of miR-137 and its targets in schizophrenia. Front. Genet. 4:58. doi: 10.3389/fgene.2013.00058
Wu, S., Zhang, R., Nie, F., Wang, X., Jiang, C., Liu, M., et al. (2016). MicroRNA-137 inhibits EFNB2 expression affected by a genetic variant and is expressed aberrantly in peripheral blood of schizophrenia patients. EBioMedicine 12, 133–142. doi: 10.1016/j.ebiom.2016.09.012
Xu, X., Liang, C., Lv, D., Yin, J., Luo, X., Fu, J., et al. (2018). Association of the Synapse-Associated Protein 97 (SAP97) gene polymorphism with neurocognitive function in schizophrenic patients. Front. Psych. 9:458. doi: 10.3389/fpsyt.2018.00458
Yin, J., Lin, J., Luo, X., Chen, Y., Li, Z., Ma, G., et al. (2014). miR-137: a new player in schizophrenia. Int. J. Mol. Sci. 15, 3262–3271. doi: 10.3390/ijms15023262
Yin, J., Zhu, D., Li, Y., Lv, D., Yu, H., Liang, C., et al. (2019). Genetic variability of TCF4 in schizophrenia of Southern Chinese Han population: a case-control study. Front. Genet. 10:513. doi: 10.3389/fgene.2019.00513
Zhang, F., Chen, Q., Ye, T., Lipska, B. K., Straub, R. E., Vakkalanka, R., et al. (2011). Evidence of sex-modulated association of ZNF804A with schizophrenia. Biol. Psychiatry 69, 914–917. doi: 10.1016/j.biopsych.2011.01.003
Zhang, Z., Yan, T., Wang, Y., Zhang, Q., Zhao, W., Chen, X., et al. (2018). Polymorphism in schizophrenia risk gene MIR137 is associated with the posterior cingulate Cortex’s activation and functional and structural connectivity in healthy controls. Neuroimage Clin. 19, 160–166. doi: 10.1016/j.nicl.2018.03.039
Zhao, L., Li, H., Guo, R., Ma, T., Hou, R., Ma, X., et al. (2013). miR-137, a new target for post-stroke depression? Neural Regen. Res. 8, 2441–2448. doi: 10.3969/j.issn.1673-5374.2013.26.005
Keywords:MIR137, polymorphism, rs2660304, rs1198588, schizophrenia
Citation: Yin J, Luo X, Peng Q, Xiong S, Lv D, Dai Z, Fu J, Wang Y, Wei Y, Liang C, Xu X, Zhang D, Wang L, Zhu D, Wen X, Ye X, Lin Z, Lin J, Li Y, Wang J, Ma G, Li K and Wang Y (2021) Sex-Specific Associations of MIR137 Polymorphisms With Schizophrenia in a Han Chinese Cohort. Front. Genet. 12:627874. doi: 10.3389/fgene.2021.627874
Edited by:
Cunyou Zhao, Southern Medical University, ChinaReviewed by:
Zhongju Wang, Southern Medical University, ChinaJian-Huan Chen, Jiangnan University, China
Fuquan Zhang, Nanjing Brain Hospital Affiliated to Nanjing Medical University, China
Copyright © 2021 Yin, Luo, Peng, Xiong, Lv, Dai, Fu, Wang, Wei, Liang, Xu, Zhang, Wang, Zhu, Wen, Ye, Lin, Lin, Li, Wang, Ma, Li and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guoda Ma, c2loYW4xMTA3QDEyNi5jb20=; Keshen Li, bGlrZXNoZW4xOTcxQDE2My5jb20=; Yajun Wang, d2FuZ3lhanVueTE5NzdAYWxpeXVuLmNvbQ==
†These authors have contributed equally to this work