 Tianyi Wu1†
Tianyi Wu1† Shanhe Wang1†
Shanhe Wang1† Lihong Wang1Weibo Zhang1
Lihong Wang1Weibo Zhang1 Weihao Chen1Xiaoyang Lv1Yue Li1Zahid Hussain1Wei Sun1,2,3*
Weihao Chen1Xiaoyang Lv1Yue Li1Zahid Hussain1Wei Sun1,2,3*- 1College of Animal Science and Technology, Yangzhou University, Yangzhou, China
- 2Joint International Research Laboratory of Agriculture and Agri-Product Safety of Ministry of Education of China, Yangzhou University, Yangzhou, China
- 3Jiangsu Co-innovation Center for Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou, China
Characterizing the factors that regulate the growth and development of muscle is central to animal production. Skeletal muscle satellite cells (SMSCs) provide an important material for simulating the proliferation and differentiation of muscle cells. YAP1, which can promote muscle growth, is closely related to the proliferation of SMSCs in Hu sheep (Ovis aries). In addition, some miRNAs, such as miR-541-3p, miR-142-5p, and miR-29a, can play critical roles in muscle growth by specifically binding with their target mRNAs. Meanwhile, lncRNA can competitively bind these miRNAs and reduce the regulatory effect of miRNAs on their target genes and thus play critical roles themselves in muscle growth. However, the regulatory molecular mechanism of miRNA and lncRNA on SMSC proliferation through YAP1 remains unclear. Here, we characterized the regulatory network among YAP1 and its targeted miRNAs and lncRNAs in Hu sheep SMSCs. The potential ncRNAs that regulate YAP1 (miR-29a and CTTN-IT1) were predicted through multilevel bioinformatics analysis. Dual-luciferase assays, RT-qPCR, and western blots revealed that miR-29a can significantly reduce the mRNA and protein expression level by binding to a specific 3′-UTR of YAP1 (P < 0.05), while CTTN-IT1 can restore the expression of YAP1 through competitive binding to miR-29a. Furthermore, the mRNA and protein expression levels of MyoG, MyoD, and MyHC showed that miR-29a can inhibit the expression of genes related to the differentiation of SMSCs, and CTTN-IT1 can increase the expression of these same genes. Thus, miR-29a may inhibit the differentiation of SMSCs and CTTN-IT1 can restore this inhibition. The EdU staining assay indicated that excessive miR-29a can significantly reduce the proliferation ability of SMSCs (P < 0.05), while overexpression of CTTN-IT1 can significantly increase the proliferation of SMSCs (P < 0.01). CTTN-IT1 is a novel lncRNA that is a competing endogenous RNA (ceRNA) of miR-29a and can promote SMSC proliferation and differentiation by restoring the expression of YAP1 when it is inhibited by miR-29a in Hu sheep. Overall, our findings construct a CTTN-IT1-miR-29a-YAP1 regulatory network that will help contribute new insight into improving the muscle development of Hu sheep.
Introduction
Hu sheep (Ovis aries), a well-known Chinese native sheep breed, has many advantages, including its reproductive performance, a high ratio of meat to bone, as well as tender and juicy meat, which customers often find appealing (Yue, 1996). However, the slow growth rates of Hu sheep have restricted the development and utilization of their excellent genetic resources. Thus, there is a need for more research to be conducted on the regulatory mechanisms underlying the muscle development of this local breed.
Mutton production is known to be closely tied to the muscle development of sheep. The process of mammalian skeletal muscle formation includes the proliferation, differentiation, and fusion of myogenic precursor cells into polynuclear muscle fibers (Buckingham, 2017). During the growth and regeneration of postpartum muscle fibers, the precursor cells to adult muscle that are located on the muscle fibers under the basal plate are called satellite cells (Mauro, 1961; Cossu and Biressi, 2005). These cells are responsible for the growth, hypertrophy, and regeneration of skeletal muscle after birth (Sun et al., 2017). When muscle is injured, activated satellite cells, which have been widely shown to be important adult stem cells, can proliferate and differentiate to form new muscle fibers (Beauchamp et al., 2000). Satellite cells of many animals have been successfully isolated, and they have become important materials for simulating the proliferation and differentiation of muscle cells in vitro (Nguyen et al., 2010). Therefore, Hu sheep skeletal muscle satellite cells (SMSCs) were used in this study to characterize the regulatory mechanism of genes related to muscle development and hypertrophy. An increasing number of genes regulating proliferation and differentiation have been found in SMSCs, such as Yes-associated protein 1 (YAP1) (Tremblay et al., 2014), MyoG (Jin et al., 2000), MyoD (Jin et al., 2000), MyHC (Chen et al., 2018), and CSRP3 (Han et al., 2019).
YAP1 is one of the key central regulators of the Hippo signaling pathway, is widely expressed in mammalian tissues (Overholtzer et al., 2006), and functions in the proliferation of muscle satellite cells (Tremblay et al., 2014; Mohamed et al., 2016). YAP can positively regulate skeletal muscle size via interaction with TEAD transcription factors (Watt et al., 2015). Ser127 phosphorylation of YAP1 is essential for the terminal differentiation of mice myoblast (C2C12) cells (Watt et al., 2010). In a previous study, we successfully established a line of Hu sheep SMSCs and showed that YAP1 in the Hippo channel can regulate the proliferation and differentiation processes of the SMSCs by participating in the regulation of TGF-β/Smad pathway activity (Su, 2015).
Many studies have reported that microRNA (miRNA) can regulate gene expression by regulating post-transcriptional translation (Lu and Rothenberg, 2018). The endogenous miRNA precursor containing a stem-ring structure can be cut into mature miRNA by the Dicer enzyme during its transfer to cytoplasm (Bak and Mikkelsen, 2014). It combines with the miRNA response element contained in the 3′-untranslated regions (3′-UTR) of mRNA in a fully or partially complementary way to promote the degradation or inhibit the translation of targeted mRNA (Salmena et al., 2011). A large number of miRNAs participate in muscle growth and development, such as miR-206 (Vergara et al., 2018), miR-125b-5p (Qiu et al., 2019), miR-17, and miR-19 (Kong et al., 2019). For example, the disruption of miR-29 leads to abnormal differentiation of vascular smooth muscle cells (Cushing et al., 2015) and contributes to dystrophic muscle pathogenesis (Wang et al., 2012). However, the overexpression of miR-29 can also lead to congenital muscular dystrophy (Liu C. et al., 2019). These findings suggest that miR-29 may play different roles in different species and tissues.
Long noncoding RNA (lncRNA) is defined as noncoding RNA with lengths greater than 200 nt. It can play a role in muscle growth and morphogenesis and can be reproduced in vitro (Buckingham and Vincent, 2009). The rapid development of information technology and increasingly sophisticated research techniques have revealed that lncRNA plays an important role in regulating muscle growth and development in animals. For example, lncRNA can participate in skeletal muscle growth and development as competing endogenous RNA (ceRNA). lncRNA can regulate the balance of muscle by affecting corresponding miRNA, and its expression level has an important influence on several muscle diseases, such as myocardial infarction and Duchenne muscular dystrophy (Eisenberg et al., 2007). lncRNA-AK017368 was found to be highly expressed in skeletal muscle cells and can function as a ceRNA of miR-30c to promote the proliferation of myoblasts by weakening the function of miR-30c (Liang et al., 2018). lncIRS1, a sponge of the miR-15 family, was shown to regulate the expression of IRS1 and promote skeletal muscle production (Li et al., 2019).
Research on muscle growth in Hu sheep, an important sheep breed native to China, is still in its incipient stages. Indeed, our understanding of the molecular mechanisms underlying the regulation of muscle growth, especially the regulation of lncRNA, is extremely limited. YAP1 plays an important role in skeletal muscle development; however, the underlying mechanism responsible for the beneficial effect of YAP1 in SMSCs remains unclear. This study aimed to explore the regulatory molecular pathway of lncRNA-miRNA-mRNA involved in mediating the function of YAP1. Our results will aid future studies of the breeding of Hu sheep as well as studies examining the molecular mechanisms underlying sheep muscle growth.
Materials and Methods
Ethics Statement
All experimental procedures were carried out in strict accordance with the guidelines for the care and use of laboratory animals in Jiangsu Province (License Number: 45) and the recommendations of the Animal Protection and Use Committee of the Ministry of Agriculture of China (License Number: 39). The protocol was approved by the Animal Care and Use Committee at Yangzhou University.
Sample Collection
After intravenous injection of 2% lidocaine hydrochloride (4 mg/kg) to anesthetize sheep, 2 g skeletal muscle tissues were sampled (by slaughter) from longissimus dorsi of three 1-month-old and three 6-month-old Hu sheep and were immediately stored in liquid nitrogen. Sheep were fed in Suzhou Sheep Farm (Suzhou, Jiangsu, China). After intravenous injection of 2% lidocaine hydrochloride (240 mg, 4 mg/kg) to anesthetize ewes, a 90-day-old Hu sheep fetus was removed from the mother’s body by surgery to separate the SMSCs. Sheep were provided by Hua Mai Sheep Farm (Yangzhou, Jiangsu, China).
Cell Acquisition and Culture
The 4th–5th generation SMSCs used in this study were isolated, identified, and cultured following a previously described method (Wu et al., 2012). Specifically, HEK293T cells kept in our lab and SMSCs were cultured in DMEM/F12 supplemented with 10% FBS and 1% penicillin-streptomycin (Gibco, Grand Island, NY, United States) at 37°C with 5% CO2. When the cells reached the logarithmic growth stage, they were seeded in different culture plates before transfection.
Plasmid Construction
All the primer sequences used in plasmid construction are listed in Table 1.

Table 1. Primers information for plasmid construction.
To overexpress lncRNA CTTN-IT1, the primers, which contained Hind III and BamH I restriction sites and vector homologous fragments, were used to amplify the full-length sequence of CTTN-IT1 (sequence can be found in Supp CTTN-IT1 Sequence). Next, the “lncRNA-overexpression plasmid” was constructed using the full-length sequence of CTTN-IT1 and the pcDNA3.1(+) vector (ThermoFisher, Shanghai, China) digested by Hind III and BamH I restriction enzymes (Takara, Kusatsu, Shiga, Japan).
For the dual-luciferase assay, wild and mutant 3′-UTRs (containing the miR-29a binding site) of YAP1 were amplified and linked downstream of the luciferase gene of the pmiRNA-REPORT Luciferase vectors (ThermoFisher, Shanghai, China) digested by Hind III and Mlu I restriction enzymes (Takara, Kusatsu, Shiga, Japan). The wild 3′-UTR of YAP1 was used as a template to construct the recombinant dual-luciferase plasmid of the “YAP1-wild” type and was copied by PCR with designed primers, which contained vector fragments and restriction endonuclease sites. Meanwhile the “YAP1-mutant” plasmid was modified from the “YAP1-wild” plasmid with a Fast Site-Directed Mutagenesis Kit (Tiangen, Beijing, China) per the manufacturer’s protocol.
The full-length sequence of CTTN-IT1 was amplified by PCR with primers containing Hind III and Mlu I restriction sites and vector homologous fragments to construct the recombinant dual-luciferase plasmid of the “lncRNA-wild” type. To explore the actual effects of the three potential binding sites predicted by software using the “lncRNA-wild” plasmid as a template, the base pairs involved in the miR-29a binding were modified using a Fast Site-Directed Mutagenesis Kit; and the plasmid was named the “lncRNA-mutant 1/2/3” plasmid. The sequences of wild and mutated fragments were shown in Supplementary Material 2.
Before constructing the plasmid, all of the DNA fragments were purified by gel cutting with a Universal DNA Purification Kit (Tiangen, Beijing, China). Next, the target segments were linked with the vector by a Trelief SoSoo Cloning Kit (Tsingke, Beijing, China) per the manufacturer’s protocol.
Cell Transfection
SMSCs at the exponential phase were digested by trypsin and seeded in 6, 24, or 96-well plates at a concentration of 3 × 104 cells/cm2. When cells were approximately 80% confluent, 100 nM of miRNA mimic/miRNA mimic-NC or 200 nM of miRNA inhibitor/miRNA inhibitor-NC and 50 nM of siRNA (All from RiboBio, Guangzhou, China) were transfected into the cells using Lipofectamine RNAiMAX Reagent (ThermoFisher, Shanghai, China) per the manufacturer’s protocol. Next, pcDNA3.1(+) vector was used as a Blank, and the lncRNA-overexpression plasmid was transfected or co-transfected with miRNA-mimics or other substances using jetPRIME (Polyplus-transfection, New York, NY, United States) per the manufacturer’s protocol.
siRNAs that can inhibit CTTN-IT1 were designed and produced by RiboBio Co. Ltd. (Guangzhou, China). To regulate the content of miRNA in cells, the miRNA mimic and mimic-NC as well as the miRNA inhibitor and inhibitor-NC, which were used for transfections into SMSCs or HEK293T cells, were synthesized by RiboBio Co. Ltd. as well.
EdU Staining Assay
A total of 1 × 104 cells/well of SMSCs were seeded in 96-well plates with five replicates per condition. When cells reached 60% confluence, 3 μg/mL of the CTTN-IT1 overexpression vector or pcDNA3.1(+) vector and 100 nM of miR-29a mimic, mimic-NC, or 200 nM miR-29a inhibitor were co-transfected into SMSCs. After 12 h, cells were treated with EdU reagent, which was contained in a Cell-Light EdU Apollo567 In Vitro Kit (RiboBio, Guangzhou, China), for 2 h and stained per the manufacturer’s instructions. Three groups of photos (400× magnification) per well of the stained cells were taken at random under a fluorescence microscope (Nikon, Tokyo, Japan). The ratio of EdU positive cells (Apollo567 staining cells/Hoechst staining cells) was then calculated.
RNA Isolation and RT-qPCR
To extract RNA, muscle tissue was ground into powder in liquid nitrogen, and the SMSCs were cultured for 48 h after transfection. Several tests were then conducted, and the steps for each are described below.
To detect the expression profile of miRNA, the miRNA of muscle tissue or cells was extracted using an miRcute miRNA Isolation Kit (Tiangen, Beijing, China) per the manufacturer’s instructions. The miRNA was reverse-transcribed into cDNAs by modifying the miRNA with a poly (A) tail using an miRcute Plus miRNA First-Strand cDNA Synthesis Kit (Tiangen, Beijing, China) per method A of the manufacturer’s instructions.
To measure the expression profiles of mRNA and lncRNA, the total RNA of the SMSCs was isolated using an RNAsimple Total RNA Kit. The cDNA was then synthesized using an RNA First-Strand cDNA Synthesis Kit (Tiangen, Beijing, China).
An Applied Biosystems 7900 Fast Real-Time PCR system (ThermoFisher, Shanghai, China) was used to conduct RT-qPCR. Primer information is shown in Table 2.

Table 2. Primer information for RT-Qpcr.
The RT-qPCR reaction solution system for miRNA using an miRcute Plus miRNA qPCR Kit (Tiangen, Beijing, China) is shown in Supplementary Table S1. The RT-qPCR program for miRNA was as follows: 95°C for 15 min, followed by 40 cycles of 94°C for 20 s and 60°C for 34 s.
The RT-qPCR reaction solution system for mRNA and lncRNA using SYBR Premix Ex Taq II (TaKaRa, Dalian, China) is shown in Supplementary Table S2. The RT-qPCR program for mRNA and lncRNA, which was the same as the method provided by the guide, was as follows: 95°C for 30 s, followed by 40 cycles of 95°C for 5 s and 60°C for 34 s and, finally, 95°C for 15 s and 60°C for 60 s.
There were three biological replicates for each condition. The results were analyzed using the 2–ΔΔCT method (Livak and Schmittgen, 2001) with either GAPDH or U6 as the internal reference genes.
Dual-Luciferase Assay
To elucidate the relationship between miR-29a and YAP1 and between miR-29a and CTTN-IT1, HEK293T cells were seeded in a 24-well plate and co-transfected with 500 ng/mL YAP1-wild, YAP1-mutant, lncRNA-wild, lncRNA-mutant, pRT-TK (RiboBio, Guangzhou, China) plasmid, and 100 nM of miR-29a mimic (and mimic-NC) or 200 nM of miR-29a inhibitor (and inhibitor-NC). The FuGENE HD Transfection Reagent (Promega, Fitchburg, WI, United States) was used for the transfection per the manufacturer’s protocol. The ratio of the transfection reagent to plasmid was 3:1. Forty-eight hours later, the dual-luciferase assay was conducted with a Promega Dual-Glo Luciferase-Assay System (Promega, Fitchburg, WI, United States) per the manufacturer’s protocol. The main steps were as follows:
1. The SMSCs from different treatments were first washed twice with PBS, 100 μL/well of PLB lysate was added, and the cells were fully lysed by shaking at room temperature for 15 min.
2. Twenty microliters of cell lysate was added into the 96-well enzyme plate, and 100 μL of Luciferase Reagent was added to test the luciferase activity of fireflies.
3. One hundred microliters of Stop & Glo Reagent was added into the well to detect the luciferase activity of Renilla.
4. Luciferase detection was run at the setting of 1–2 s delay and 5–10 s reading. The data were presented as relative luciferase activity (Firefly/Renilla, OD/OD) and were normalized by a control well. Each group consisted of three replicates.
Western Blot
The protein samples were extracted from SMSCs after the miR-29a mimic (and mimic-NC) or miR-29a inhibitor (and inhibitor-NC) and lncRNA-overexpression plasmid [and pcDNA3.1(+)] were co-transfected into the cells for 48 h. The concentration of protein samples was measured and standardized before the samples were compressed by 5% spacer gel (80 V, 20 min) and separated by 10% separation gel (100 V, 80 min) through electrophoresis. Next, the separated proteins were transferred from gel to a polyvinylidene difluoride (PVDF) membrane by electricity transfer (200 mA, 1 h). After being blocked with 5% skimmed milk for 1 h at room temperature, the PVDF membranes were kept in primary antibodies [against YAP1(ab205270, 1:1000), MyoG(ab187506, 1:1000), MyoD(ab126726, 1:1000), MyHC(ab91506, 1 μg/ml), and β-actin(ab6276, 1:5000)] (Abcam, Cambridge, United Kingdom) overnight at 4°C and then incubated with the HRP-conjugated secondary antibody goat-anti-mouse IgG (Zhongshan-Bio, Beijing, China) at room temperature for 2 h. The PVDF membranes were washed with PBST several times, 5 min per wash, after incubation in antibodies. Finally, the developer was added to the PVDF membrane and was placed at room temperature for 1 min. The membrane was then wrapped with plastic wrap (bubbles were avoided as much as possible) in a dark room, and the membrane was quickly pasted on the X-ray film for exposure. The image was developed and washed in the film-developing machine. The exposure time was adjusted until the best strip appeared.
Software Prediction
To predict the potential target-related miRNAs in the 3′-UTR regions of YAP1 of Hu sheep, the potential miRNAs were selected by two online softwares: miRBase1 and miRanda2. Findtar33 and RNA224 were used to predict the existence of lncRNA that were potentially targeted by miR-29a.
Statistical Analysis
Excel 2013 (Microsoft, Redmond, WA, United States) was used for data analyses. SPSS 16.0 (IBM, Armonk, NY, United States) was used to calculate the significance of t-tests. In addition to using multiple comparisons (LSD) in Figure 4D, the significance for other figures was calculated using independent sample t-tests. All of the bar charts were drawn with GraphPad Prism 6 (GraphPad Software, San Diego, CA, United States). The marker “∗” represented a significant difference between the experimental group and the control group (P < 0.05), and “∗∗” represented an extremely significant difference between the experimental group and the control group (P < 0.01).
Results
miR-29a Regulates YAP1 Expression by Targeting Its 3′-UTR
To explore the miRNAs that target YAP1, online programs, such as Findtar3, miRBase, RNA22, and miRanda, were used to predict the miRNAs that could potentially target the 3′-UTR of YAP1 in sheep. Four miRNAs were found to potentially target YAP1 gene by binding to the 3′-UTR of YAP1 (Supplementary Table S3). Among these four miRNAs, miR-29a has already been shown to play an important role in skeletal muscle development of mice in several papers (Galimov et al., 2016; Zhou et al., 2016; Muluhngwi et al., 2017), consequently, the relationship between miR-29a and YAP1 was studied further.
Our previous study showed that the expression of YAP1 in slow-growing, 6-month-old Hu sheep was significantly lower than that in fast-growing, 1-month-old Hu sheep (Gao et al., 2016). RT-qPCR was then used to detect the expression of miR-29a in the longissimus dorsi muscle of 1-month-old and 6-month-old Hu sheep (Figure 1A). The relative expression level of miR-29a in the 6-month-old longissimus dorsi of Hu sheep was extremely significantly higher than those observed in 1-month-old longissimus dorsi (P < 0.01). The expression of miR-29a was negatively correlated with the expression of YAP1, indicating that YAP1 was a potential target gene of miR-29a.

Figure 1. Interaction between miR-29a and YAP1. (A) The expression of miR-29a in the longissimus dorsi muscle between 1-month-old and 6-month-old Hu sheep. (B) The relative expression level of miR-29a and YAP1 after miR-29a mimic or inhibitor was transfected into SMSCs. (C) Results of the dual-luciferase reporter assay to confirm the potential interaction between miR-29a and YAP1, after YAP1-wild or YAP1-mutant, recombined dual-luciferase reporter plasmid, and miR-29a mimic (and mimic-NC) or miR-29a inhibitor (and inhibitor-NC) were co-transfected into HEK293T cells. (D) The expression of YAP1 and β-actin was detected by western blot after miR-29a mimic and inhibitor were transfected into SMSCs. *P < 0.05 or **P < 0.01.
The effectiveness of the miR-29a mimic and inhibitor was detected by RT-qPCR after transfection of miR-29a mimic/mimic-NC and inhibitor/inhibitor NC into SMSCs (Figure 1B, right part). Transfection of both mimic and inhibitor was significantly different compared with the control (P < 0.05). Therefore, miR-29a mimics and inhibitors were used in subsequent experiments. The expression of YAP1 was also detected by RT-qPCR after transfection of miR-29a mimic/mimic-NC and inhibitor/inhibitor NC into SMSCs (Figure 1B, left part). Transfection of the miR-29a mimic resulted in a highly significant decrease in the expression of YAP1 (P < 0.01), and miR-29a inhibitor resulted in a highly significant increase in the expression of YAP1 (P < 0.01). Thus, miR-29a can negatively regulate YAP1.
The dual-luciferase assay and western blot were used to verify the relationship between YAP1 and miR-29a. YAP1-wild or YAP1-mutant was recombined with the dual-luciferase reporter plasmid, and miR-29a mimic/mimic-NC or miR-29a inhibitor/inhibitor-NC was co-transfected into HEK293T cells. After 48 h, the dual-luciferase reporter gene experiment was conducted. The relative activity of luciferase in the co-transfected miR-29a mimic and wild-type vector group was significantly lower than that in the control group (P < 0.05), while the relative luciferase activity in the co-transfected miR-29a mimic and mutated vector group was not significantly different from that in the control group (P > 0.05) (Figure 1C). Thus, the miR-29a seed region can bind to the target site of YAP1, consistent with our prediction. The western blot (Figure 1D) revealed that the miR-29a mimic could inhibit the expression of YAP1 protein and that the miR-29a inhibitor could increase its expression. Thus, miR-29a can reduce the expression level of YAP1 protein, suggesting that YAP1 is the target gene of miR-29a.
miR-29a Can Inhibit the Proliferation and Differentiation of SMSCs
YAP1 can regulate the proliferation and differentiation processes of the SMSCs. To determine whether miR-29a can regulate the proliferation and differentiation of SMSCs by regulating YAP1, the mRNA and protein expression of the marker genes of cell differentiation including MyoG, MyoD, and MyHC (Figures 2A,B) were detected after miR-29a mimic and inhibitor were transfected into SMSCs, and the cell proliferation ability was detected by the EdU staining assay (Figures 2C,D).
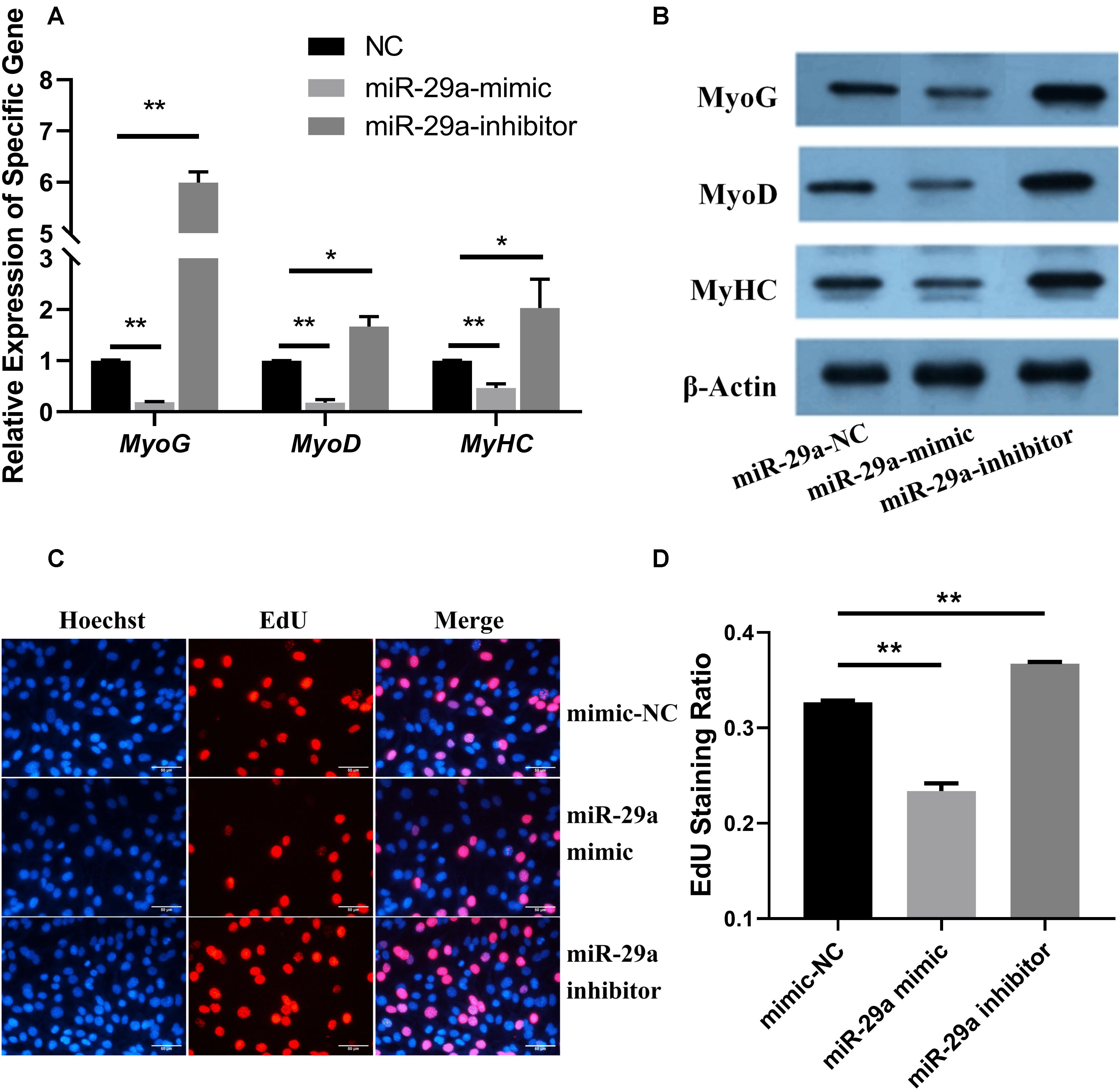
Figure 2. Functional analysis of miR-29a during the proliferation and differentiation of SMSCs. (A) The relative expression levels of MyoG, MyoD, and MyHC after the transfection of miR-29a mimic and inhibitor. (B) The protein expression of MyoG, MyoD, MyHC, and β-actin was detected by western blot after miR-29a mimic and inhibitor were transfected into SMSCs. (C) Images of the results of the EdU staining assay after miR-29a mimics, mimic-NC, and inhibitors were transfected into SMSCs. The short white lines in each picture represent 50 μm. (D) Data analysis of the EdU results. Higher values represent more rapid cell proliferation. *P < 0.05 or **P < 0.01.
The relative expression levels of MyoG, MyoD, and MyHC were significantly lower in the control group (P < 0.01) after overexpression of miR-29a by transfection of the miR-29a mimic, but they were significantly higher than the control group after inhibiting endogenous miR-29a (P < 0.05; Figure 2A). Furthermore, western blot was used to detect the protein levels of these genes related to muscle differentiation after transfection of miR-29a mimic or inhibitor (Figure 2B). After overexpression of miR-29a in SMSCs, the protein expression levels of these genes were lower than those in the negative control group; in contrast, after inhibition of miR-29a, the protein expression levels of these genes in muscle were higher than those in the negative control group. These results thus suggested that miR-29a might inhibit the differentiation of SMSCs.
After miR-29a mimics, mimic-NC, and inhibitors were transfected into SMSCs, the proliferation of Hu sheep SMSCs was detected by the EdU staining assay (Figures 2C,D). Compared with the control group, the transfection of miR-29a mimic could significantly reduce cell proliferation, while the transfection of miR-29a-inhibitor could significantly increase cell proliferation (P < 0.05). These results indicated that miR-29a could inhibit the proliferation of Hu sheep SMSCs by silencing YAP1 mRNA expression.
lncRNA CTTN-IT1 Upregulates YAP1 by Acting as a ceRNA Through the Absorption of miR-29a
Two online programs, RNA22 and Findtar3, were used to predict the potential lncRNA-containing target sites binding to miR-29a. Eventually, the CTTN-IT1 and its three binding sites potentially targeted by miR-29a were identified (Table 3).
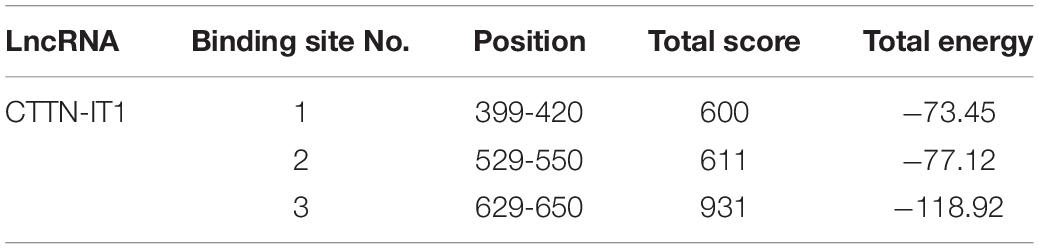
Table 3. Target prediction of CTTN-IT1 and oar-miR-29a.
First, the validity of the plasmid used for transfection was tested. The relative expression level of CTTN-IT1 was detected by RT-qPCR after the lncRNA-overexpression plasmid, lncRNA-siRNA, or its negative control were transfected into Hu sheep SMSCs. The relative expression level of CTTN-IT1 in the overexpression group was significantly higher than that in the negative control group (P < 0.01), and its expression level in the interference group was significantly lower than that in the negative control group (P < 0.05) (Figure 3A). Thus, the lncRNA-overexpression plasmid was constructed successfully, and siRNA was effective enough for use in the subsequent experiment.
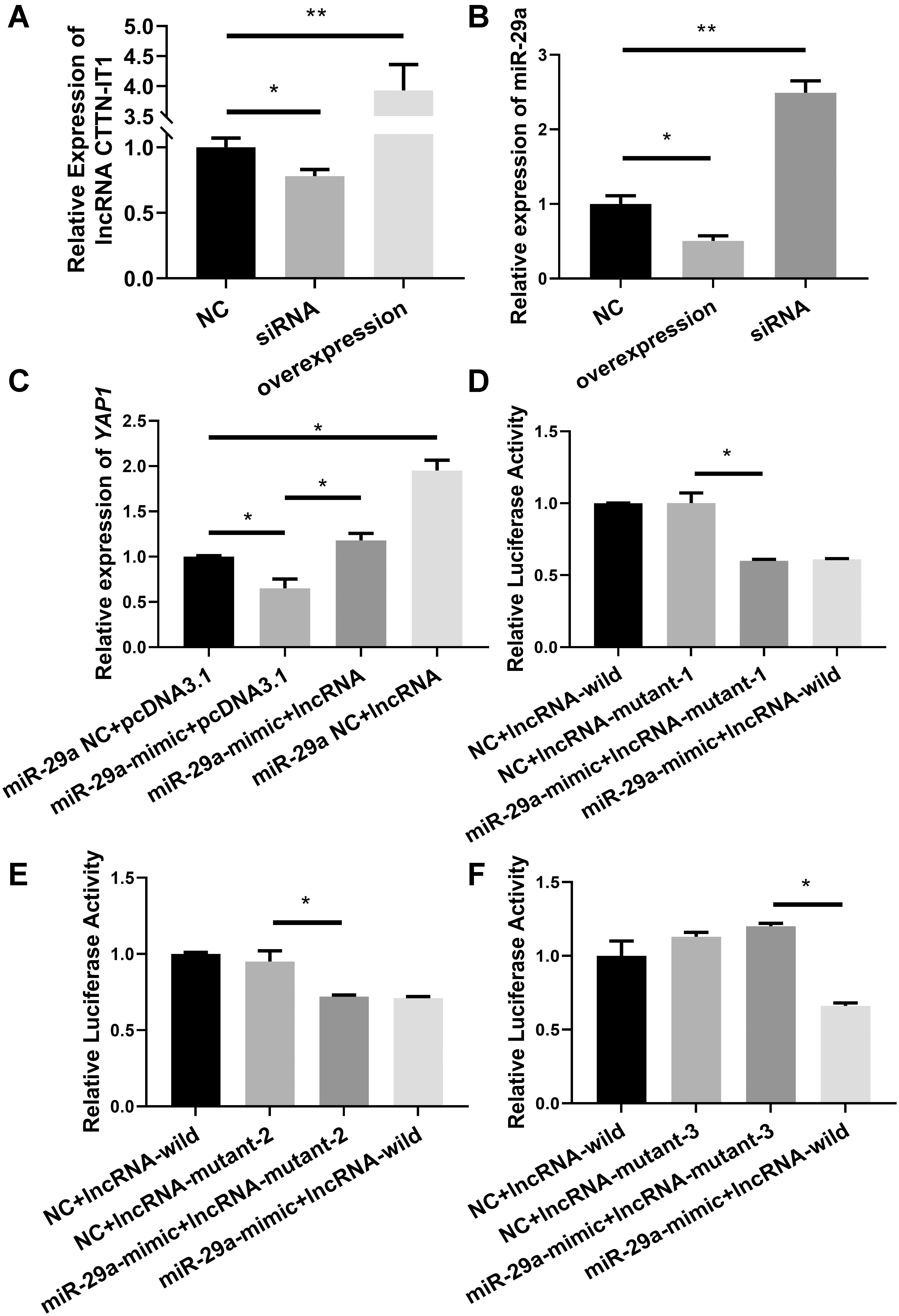
Figure 3. Verification of target regulation relationship between miR-29a and CTTN-IT1. (A) The relative expression level of CTTN-IT1 after overexpression or interference of CTTN-IT1 in SMSCs. (B) The relative expression level of miR-29a after overexpression or interference of CTTN-IT1 in SMSCs. (C) The relative expression level of YAP1 after lncRNA-overexpression plasmid [pcDNA3.1(+) used as Blank] was co-transfected with miR-29a mimic or miR-29a mimic-NC into SMSCs. (D–F) These three figures are the results of dual-luciferase reporter assay. Relative luciferase activity was assessed after co-transfecting the miR-29a mimic (miR-29a mimic-NC used as Blank) with the wild or the mutant plasmids of CTTN-IT1 into HEK293T cells. Mutant plasmids of three different binding sites listed in Table 3 were used in figures (D–F), respectively. The “lncRNA” in C means the transfection of lncRNA-overexpression plasmid. *P < 0.05 or **P < 0.01.
To verify the effect of CTTN-IT1 on miR-29a, the relative expression level of miR-29a was detected by RT-qPCR after the lncRNA-overexpression plasmid, lncRNA-siRNA, or its negative control were transfected into Hu sheep SMSCs. Compared with the negative control group, the relative expression level of miR-29a in the CTTN-IT1 overexpression group was significantly lower (P < 0.05). Transfection of lncRNA-siRNA showed that the relative expression level of miR-29a in the lncRNA-siRNA group was significantly higher than that in the negative control group (P < 0.01) (Figure 3B). Thus, CTTN-IT1 could effectively reduce mature miR-29a in SMSCs.
To characterize the effect of CTTN-IT1 on YAP1 through its reduction of miR-29a, lncRNA-overexpression plasmid, or pcDNA3.1 were co-transfected with miR-29a mimic or miR-29a mimic-NC into Hu sheep SMSCs. The co-transfected pcDNA3.1, and miR-29a mimic-NC was the control group. RT-qPCR was used to detect the relative expression level of YAP1 (Figure 3C). Co-transfection of the miR-29a mimic and pcDNA3.1 could significantly reduce YAP1 content in cells (P < 0.05). However, overexpression of miR-29a and CTTN-IT1 significantly increased the expression of YAP1 (P < 0.05) relative to the overexpression of miR-29a alone. In general, overexpression of CTTN-IT1 could significantly increase the expression of YAP1 (P < 0.05). Overall, CTTN-IT1 can positively regulate YAP1 expression, and overexpression of CTTN-IT1 can restore the expression of YAP1 from inhibition by miR-29a.
To identify the key binding site of CTTN-IT1 during this regulatory process, different plasmids (lncRNA-wild/lncRNA-mutant) were co-transfected with miR-29a mimic/mimic-NC into HEK293T cells. The co-transfected lncRNA-wild, and miR-29a mimic-NC was the control group. After 48 h, the dual-luciferase assay revealed that the activity of luciferase in the co-transfected miR-29a mimic and the lncRNA-wild group in all three histograms was significantly lower than that in the control group (P < 0.05). Compared with binding sites 1 and 2, only binding site 3 showed no significant difference in relative luciferase activity between the co-transfected miR-29a mimic and lncRNA-mutant group compared with the control group (P > 0.05) (Figures 3D–F). Thus, binding site 3 (located from 629 to 650 bp on CTTN-IT1) was the binding site of miR-29a and CTTN-IT1.
lncRNA CTTN-IT1 Affects the Differentiation and Proliferation of SMSCs
The mRNA expression levels of YAP1, MyoG, MyoD, and MyHC (Figure 4A) were assessed after lncRNA overexpression or lncRNA-siRNA [pcDNA3.1(+) as the control group] were transfected into SMSCs. Overexpression of CTTN-IT1 significantly increased the expression level of YAP1 (P < 0.01), and the interference of CTTN-IT1 significantly decreased the expression level of YAP1 (P < 0.05). Thus, CTTN-IT1 can regulate the expression of YAP1.
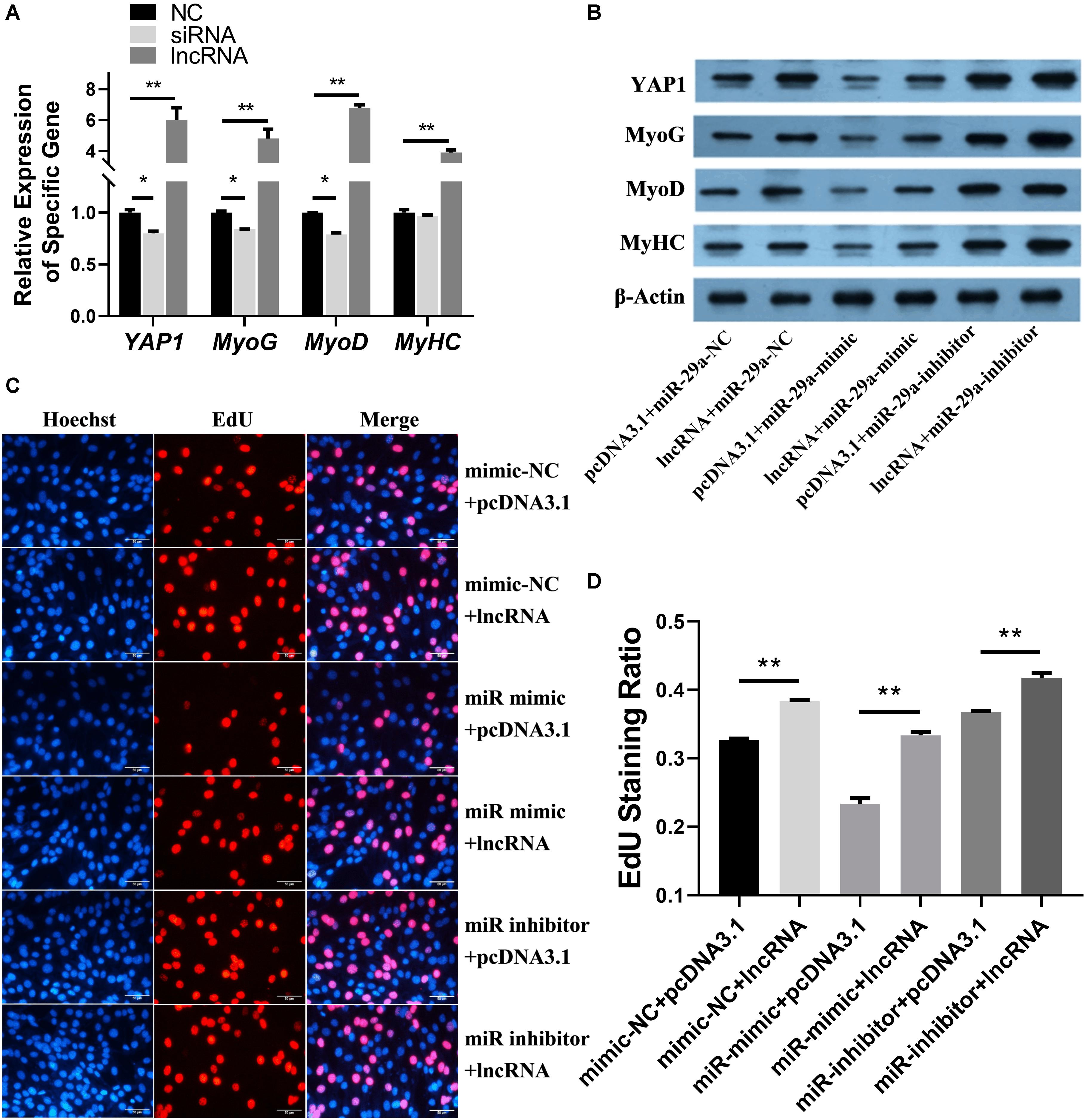
Figure 4. Functional analysis of miR-29a and CTTN-IT1 during the proliferation and differentiation of SMSCs. (A) The relative expression levels of YAP1, MyoG, MyoD and MyHC after lncRNA-overexpression or LncRNA-siRNA were transfected into SMSCs [pcDNA3.1(+) used as NC]. (B) The protein expression of YAP1, MyoG, MyoD, MyHC and β-actin was assessed by western blot after the transfection of lncRNA-overexpression plasmid [pcDNA3.1(+) used as Blank], miR-29a mimic or miR-29a inhibitor (miR-29a mimic-NC used as Blank) in SMSCs. (C) Images of the results of the EdU staining assay after miR-29a mimics, mimic-NC and inhibitors were co-transfected with LncRNA-overexpression plasmid into SMSCs. The short white lines in each picture represent 50 μm. (D) Data analysis of the EdU results. Higher values represent faster cell proliferation. All the “lncRNA” this figure means the transfection of lncRNA-overexpression plasmid. *P < 0.05 or **P < 0.01.
The relative expression levels of MyoG, MyoD, and MyHC were significantly higher than those of the control group (P < 0.01) after overexpression of CTTN-IT1. The transfection of siRNA could significantly down-regulate the relative expression levels of MyoG and MyoD (P < 0.05). However, siRNA transfection had no significant effect on the expression of MyHC (P > 0.05). Thus, CTTN-IT1 can promote the expression of genes related to the differentiation of SMSCs.
After lncRNA-overexpression plasmid [pcDNA3.1(+) was used as a blank] and miR-29a mimic or miR-29a inhibitor (miR-29a mimic-NC used as Blank) were co-transfected into SMSCs, the proteins expression of YAP1, MyoG, MyoD, MyHC, and β-actin was detected by western blot (Figure 4B). The protein expression level in the CTTN-IT1 overexpressed group (lncRNA+miR-29a-NC) was higher than that in the negative control group (pcDNA3.1+miR-29a-NC). CTTN-IT1 can restore the protein expression of YAP1, MyoG, MyoD, and MyHC from inhibition by miR-29a in the lncRNA+miR-29a mimic group relative to the pcDNA3.1+miR-29a mimic group and pcDNA3.1+miR-29a-NC groups. The co-transfection of lncRNA-overexpression plasmid and miR-29a inhibitor had the highest protein expression levels. Thus, CTTN-IT1 can regulate the expression of YAP1 at the protein level and can increase the expression levels of MyoG, MyoD, and MyHC, proteins related to the differentiation of SMSCs.
After lncRNA-overexpression plasmid [pcDNA3.1(+) used as Blank] and miR-29a mimic or miR-29a inhibitor (miR-29a mimic-NC used as a blank) were co-transfected into SMSCs, the cell proliferation ability was assessed by the EdU staining assay (Figures 4C,D). Our results showed that overexpression of CTTN-IT1 could significantly increase the proliferation of SMSCs in each comparison (P < 0.01). Thus, CTTN-IT1 can promote SMSC proliferation, which was probably achieved by reducing the content of miR-29a to restore the function of YAP1.
In sum, miR-29a can inhibit the proliferation and differentiation of Hu sheep SMSCs by silencing YAP1 and indirectly reducing the expression of MyoG, MyoD, and MyHC. CTTN-IT1 can promote the proliferation and differentiation of SMSCs by reducing the content of endogenous miR-29a.
Discussion
YAP/TAZ act downstream in the Hippo pathway and have been extensively studied given their important functions (Piccolo et al., 2014). YAP1 can regulate cell proliferation, migration, and apoptosis (Lapi et al., 2008; Wang et al., 2009). Knockdown of YAP1 can decreased the viability of vascular smooth muscle cells, and overexpression of YAP1-5SA, which lacks five serine phosphorylation sites, found that YAP1 can inhibit the apoptosis induced by endoplasmic reticulum stress (Takaguri et al., 2017). Overexpression of YAP1 can lead to the hypertrophy of cardiac muscle cells (Del Re et al., 2013). Our previous study also showed that YAP1 is closely related to the proliferation and differentiation of Hu sheep muscle (Su, 2015). Research on YAP1 regulation has gradually deepened yearly. The expression of YAP1 protein can been down regulated by tunicamycin in vascular smooth muscle cells (Takaguri et al., 2017). The mRNA expression of YAP1 can be targeted and reduced by some miRNAs such as miR-375 (Zhao et al., 2020) and miR-9 (Zheng et al., 2019) in cell line models of human diseases. However, the regulatory molecular mechanism of miRNA on the proliferation and differentiation of Hu sheep muscle through YAP1 remains unclear.
MiRNAs, miR-29a included, were involved in a variety of physiological processes like differentiation, disease, and development. The miR-29a not only participates in the formation of omental neovascularization (Han et al., 2018), diabetes (Zhang et al., 2017; Huang et al., 2019), and tumors (Liu et al., 2018) but also in the growth and development of skeletal muscle (Galimov et al., 2016; Zhou et al., 2016; Muluhngwi et al., 2017). The growth rate of Hu sheep peaks at the age of 1 month and gradually tends to flatten at the age of 6 months (Chen et al., 2014). This difference in growth rate allowed us to compare the differences in the expression of miRNAs between 1- and 6-month-old sheep and to identify the miRNAs associated with YAP1. The relative expression level of miR-29a was significantly higher at the age of 6 months than at the age of 1 month, indicating that the expression of miR-29a may increase as the proliferation rate of Hu sheep muscle cells decreases. We therefore speculated that miR-29a plays a negative role in the proliferation of Hu sheep satellite cells. The expression profile of miR-29a in skeletal muscle documented in fetal and mature goats (Wang et al., 2014) and pigs (Siengdee et al., 2015) was similar to the profile observed in our study. Then we found that miR-29a can inhibit the proliferation and differentiation of SMSCs in Hu Sheep by targeting the 3′-UTR of YAP1.
Furthermore, RT-qPCR, EdU, and western blot technology were used to reveal the function of miR-29a. Considering the crucial roles of MyoD, MyoG, and MyHC in the differentiation of muscle satellite cells (Tedesco et al., 2010), these genes were selected as the marker genes of myocyte differentiation in this study. Both RT-qPCR and western blot revealed that miR-29a may play a role in inhibiting SMSC differentiation. Previous studies have also suggested that YAP1 can facilitate the proliferation of SMSCs (Watt et al., 2010; Liu et al., 2017) but inhibit their differentiation (Sun et al., 2017). Previous studies have also suggested that miR-29a can inhibit the growth of skeletal muscle (Wang et al., 2011; Liu C. et al., 2019) and the proliferation of satellite cells and promote the differentiation of satellite cells (Kapinas et al., 2010; Wei et al., 2013). Our results also showed that miR-29a can inhibit the proliferation of SMSCs, which is consistent with previous studies. However, our preliminary results showed that miR-29a can inhibit the genes related to SMSC differentiation, indicating that miR-29a might inhibit both proliferation and differentiation in Hu sheep SMSCs. One potential explanation is that the functions of miR-29a differ between species. In addition, several studies have revealed similar expression profiles between MyoG (Lv et al., 2015), MyHC (Gao et al., 2016), and YAP1 (decreases with age), indicating that miR-29a may inhibit both YAP1 and genes related to differentiation in Hu sheep SMSCs. Another potential explanation is that, in addition to YAP1, there might be other genes targeted by miR-29a involved in the proliferation and differentiation of SMSCs. All of these possibilities require further study.
LncRNA CTTN-IT1, is located within the intron between exon 10–11 of CTTN on chromosome 21 (49352559 bp to 49353268 bp) of sheep. Some researchers believe that lncRNAs regulate cell functions through competitively binding miRNAs (Geng and Tan, 2016; Marchese et al., 2017). In line with this idea, after verifying that CTTN-IT1 can regulate the expression of miR-29a, a dual-luciferase vector and siRNA were designed for three potential miR-29a target sites of CTTN-IT1 to complete a series of experiments. CTTN-IT1 was found to play a positive role in YAP1 regulation through the third binding site by reducing the natural content of miR-29a. It means that CTTN-IT1 can act as a ceRNA and function as a sponge that absorbs miR-29a. Thus, overexpression of CTTN-IT1 can up-regulate the expression of YAP1 and genes related to muscle differentiation, and when the miR-29a mimic was transfected, the opposite pattern was observed. The result of the EdU staining assay also verified that CTTN-IT1 can promote the proliferation of SMSCs. CTTN-IT1 in this experiment has the same effect in promoting the proliferation of muscle cells as ANRIL (Tan et al., 2019), lncRNA-MEG3 (Bai et al., 2019), and Linc00299 (Liu Y. et al., 2019). There is another lncRNA has been reported can also regulate YAP1. LncRNA PFAR can positive regulate YAP1 by acts as a ceRNA of miR-138 to promote lung fibroblast activation and fibrosis (Zhao et al., 2018). Other lncRNAs have been reported to be involved in the differentiation or proliferation of SMSC. LncR-125b can promotes the differentiation of goat SMSC by absorbing miR-125b (Zhan et al., 2019). Lnc133b can regulate bovine SMSC proliferation and differentiation by mediating miR-133b (Jin et al., 2017).
In sum, the results of the present work identify a lncRNA CTTN-IT1 witch can function as a ceRNA to regulate YAP1 by competitively binding to miR-29a. In addition, CTTN-IT1 can promote the proliferation and differentiation of SMSCs. CTTN-IT1 can be used as a new molecular marker for studying the proliferation of Hu sheep muscle cells and contribute to improving the muscle development of Hu sheep.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
All experimental procedures were carried out in strict accordance with the guidelines for the care and use of laboratory animals in Jiangsu Province (License Number: 45) and the recommendations of the Animal Protection and Use Committee of the Ministry of Agriculture of China (License Number: 39). The protocol was approved by the Animal Care and Use Committee at Yangzhou University.
Author Contributions
TW and SW completed the EdU assay and wrote the draft. YL and WZ constructed the vectors and cultured cells. TW and WC analyzed the data. LW, WS, and SW designed the experiment and performed the RT-qPCR and Western blot. XL and SW gave several suggestions for manuscript modification. TW, WZ, LW, YL, XL, WC, and SW collected the sample. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by National Natural Science Foundation of China, grant number (31872333). The Projects of Domesticated Animals Platform of the Ministry of Science, Key Research and Development Plan (modern agriculture) in Jiangsu Province (BE2018354), Major new varieties of agricultural projects in Jiangsu Province (PZCZ201739), Jiangsu Agricultural Science and Technology Innovation Fund (CX (18) 2003), The Priority Academic Program Development of Jiangsu Higher Education Institutions, Major projects of Natural Science Research of colleges and universities in Jiangsu Province (17KJA230001), The Project of six peak of talents of Jiangsu Province of China and Postgraduate Innovation Project of Yangzhou University (XKYCX17_060, SJCX18_0804).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all the enterprises that have provided help for this study, especially Suzhou Hu Sheep Farm. At the same time, we also thank the editors and reviewers for their valuable comments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2020.00843/full#supplementary-material
Footnotes
- ^ http://www.mirbase.org/
- ^ http://www.microrna.org/microrna/home.do
- ^ http://bio.sz.tsinghua.edu.cn/
- ^ https://cm.jefferson.edu/rna22/Interactive/
References
Bai, Y., Zhang, Q., Su, Y., Pu, Z., and Li, K. (2019). Modulation of the proliferation/apoptosis balance of vascular smooth muscle cells in atherosclerosis by lncRNA-MEG3 via regulation of miR-26a/Smad1 axis. Int. Heart J. 60, 444–450. doi: 10.1536/ihj.18-195
Bak, R. O., and Mikkelsen, J. G. (2014). miRNA sponges: soaking up miRNAs for regulation of gene expression. Wiley Interdiscipl. Rev. RNA 5, 317–333. doi: 10.1002/wrna.1213
Beauchamp, J. R., Heslop, L., Yu, D. S. W., Tajbakhsh, S., Kelly, R. G., Wernig, A., et al. (2000). Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J. Cell Biol. 151, 1221–1233.
Buckingham, M. (2017). Gene regulatory networks and cell lineages that underlie the formation of skeletal muscle. Proc. Natl. Acad. Sci. U.S.A. 114, 5830–5837.
Buckingham, M., and Vincent, S. D. (2009). Distinct and dynamic myogenic populations in the vertebrate embryo. Curr. Opin. Genet. Dev. 19, 444–453. doi: 10.1016/j.gde.2009.08.001
Chen, L., Lyu, X., Wang, Q., Wang, X., Su, R., Yin, J., et al. (2014). Prediction and trend analysis of growth curve model of Hu sheep. Chin. Anim. Husbandry Vet. Med. 41, 239–243.
Chen, X., Guo, Y., Jia, G., Zhao, H., Liu, G., and Huang, Z. (2018). Arginine promotes slow myosin heavy chain expression via Akirin2 and the AMP-activated protein kinase signaling pathway in porcine skeletal muscle satellite cells. J. Agricult. Food Chem. 66, 4734–4740. doi: 10.1021/acs.jafc.8b00775
Cossu, G., and Biressi, S. (2005). Satellite cells, myoblasts and other occasional myogenic progenitors: possible origin, phenotypic features and role in muscle regeneration. Semin. Cell Dev. Biol. 16, 623–631. doi: 10.1016/j.semcdb.2005.07.003
Cushing, L., Costinean, S., Xu, W., Jiang, Z., Madden, L., Kuang, P., et al. (2015). Disruption of miR-29 leads to aberrant differentiation of smooth muscle cells selectively associated with distal lung vasculature. PLoS Genet. 11:e1005238. doi: 10.1371/journal.pgen.1005238
Del Re, D. P., Yang, Y., Nakano, N., Cho, J., Zhai, P., Yamamoto, T., et al. (2013). Yes-associated protein Isoform 1 (Yap1) promotes cardiomyocyte survival and growth to protect against myocardial ischemic injury. J. Biol. Chem. 288, 3977–3988. doi: 10.1074/jbc.m112.436311
Eisenberg, I., Eran, A., Nishino, I., Moggio, M., Lamperti, C., Arnato, A. A., et al. (2007). Distinctive patterns of microRNA expression in primary muscular disorders. Proc. Natl. Acad. Sci. U.S.A. 104, 17016–17021. doi: 10.1073/pnas.0708115104
Galimov, A., Merry, T. L., Luca, E., Rushing, E. J., Mizbani, A., Turcekova, K., et al. (2016). MicroRNA-29a in adult muscle stem cells controls skeletal muscle regeneration during injury and exercise downstream of fibroblast growth factor-2. Stem Cells 34, 768–780. doi: 10.1002/stem.2281
Gao, W., Sun, W., Su, R., Lv, X. Y., Wang, Q. Z., Li, D., et al. (2016). Sheep YAP1 temporal and spatial expression trend and its relation with MyHCs expression. Genet. Mol. Res. 15:gmr7260.
Geng, H., and Tan, X.-D. (2016). Functional diversity of long non-coding RNAs in immune regulation. Genes Diseases 3, 72–81. doi: 10.1016/j.gendis.2016.01.004
Han, S., Cui, C., Wang, Y., He, H., Liu, Z., Shen, X., et al. (2019). Knockdown of CSRP3 inhibits differentiation of chicken satellite cells by promoting TGF-beta/Smad3 signaling. Gene 707, 36–43. doi: 10.1016/j.gene.2019.03.064
Han, X., Zheng, J., Wang, Y., and Gao, Z. (2018). miRNA-29a inhibits colon cancer growth by regulation of the PTEN/Akt/GSK3 beta and Wnt/beta-catenin signaling pathways. Oncol. Lett. 16, 2638–2644.
Huang, Z., Li, N., Shan, Y., and Liang, C. (2019). Hsa-miRNA-29a protects against high glucose-induced damage in human umbilical vein endothelial cells. J. Cell. Biochem. 120, 5860–5868. doi: 10.1002/jcb.27871
Jin, C. F., Li, Y., Ding, X. B., Li, X., Zhang, L. L., Liu, X. F., et al. (2017). lncl33b, a novel, long non-coding RNA, regulates bovine skeletal muscle satellite cell proliferation and differentiation by mediating miR-133b. Gene 630, 35–43. doi: 10.1016/j.gene.2017.07.066
Jin, Y., Murakami, N., Saito, Y., Goto, Y., Koishi, K., and Nonaka, I. (2000). Expression of MyoD and myogenin in dystrophic mice, mdx and dy, during regeneration. Acta Neuropathol. 99, 619–627. doi: 10.1007/s004010051172
Kapinas, K., Kessler, C., Ricks, T., Gronowicz, G., and Delany, A. M. (2010). miR-29 modulates Wnt signaling in human osteoblasts through a positive feedback loop. J. Biol. Chem. 285, 25221–25231. doi: 10.1074/jbc.m110.116137
Kong, D., He, M., Yang, L., Zhou, R., Yan, Y.-Q., Liang, Y., et al. (2019). MiR-17 and miR-19 cooperatively promote skeletal muscle cell differentiation. Cell. Mol. Life Sci. 76, 5041–5054. doi: 10.1007/s00018-019-03165-7
Lapi, E., Di Agostino, S., Donzelli, S., Gal, H., Domany, E., Rechavi, G., et al. (2008). PML, YAP, and p73 are components of a proapoptotic autoregulatory feedback loop. Mol. Cell 32, 803–814. doi: 10.1016/j.molcel.2008.11.019
Li, Z., Cai, B., Abdalla, B. A., Zhu, X., Zheng, M., Han, P., et al. (2019). LncIRS1 controls muscle atrophy via sponging miR-15 family to activate IGF1-PI3K/AKT pathway. J. Cachexia Sarcop. Muscle 10, 391–410. doi: 10.1002/jcsm.12374
Liang, T., Zhou, B., Shi, L., Wang, H., Chu, Q., Xu, F., et al. (2018). lncRNA AK017368 promotes proliferation and suppresses differentiation of myoblasts in skeletal muscle development by attenuating the function of miR-30c. FASEB J. 32, 377–389. doi: 10.1096/fj.201700560rr
Liu, C., Li, L., Ge, M., Gu, L., Wang, M., Zhang, K., et al. (2019). Overexpression of miR-29 leads to myopathy that resemble pathology of ullrich congenital muscular dystrophy. Cells 8:459. doi: 10.3390/cells8050459
Liu, M., Zeng, X., Lu, Y. X., Mo, Y. J., Liao, T. H., Gan, C., et al. (2018). Study on molecular mechanism of MiRNA-29a in promoting proliferation and invasion of non-small-cell lung cancer by inhibiting MTSS1. Eur. Rev. Med. Pharmacol. Sci. 22, 5531–5538.
Liu, T., Xu, J., Guo, J. L., Lin, C. Y., Luo, W. M., Yuan, Y., et al. (2017). YAP1 up-regulation inhibits apoptosis of aortic dissection vascular smooth muscle cells. Eur. Rev. Med. Pharmacol. Sci. 21, 4632–4639.
Liu, Y., Chen, Y., Tan, L., Zhao, H., and Xiao, N. (2019). Linc00299/miR-490-3p/AURKA axis regulates cell growth and migration in atherosclerosis. Heart Vessels 34, 1370–1380. doi: 10.1007/s00380-019-01356-7
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lv, X. Y., Sun, W., Su, R., Li, D., Wang, Q. Z., Musa, H. H., et al. (2015). Correlation between sheep YAP1 temporal and spatial expression trends and MSTN and MyoG gene expression. Genet. Mol. Res. 14, 3244–3256. doi: 10.4238/2015.april.13.3
Marchese, F. P., Raimondi, I., and Huarte, M. (2017). The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 18:206.
Mauro, A. (1961). Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 9, 493–495. doi: 10.1083/jcb.9.2.493
Mohamed, A., Sun, C., De Mello, V., Selfe, J., Missiaglia, E., Shipley, J., et al. (2016). The Hippo effector TAZ (WWTR1) transforms myoblasts and TAZ abundance is associated with reduced survival in embryonal rhabdomyosarcoma. J. Pathol. 240, 3–14. doi: 10.1002/path.4745
Muluhngwi, P., Krishna, A., Vittitow, S. L., Napier, J. T., Richardson, K. M., Ellis, M., et al. (2017). Tamoxifen differentially regulates miR-29b-1 and miR-29a expression depending on endocrine-sensitivity in breast cancer cells. Cancer Lett. 388, 230–238. doi: 10.1016/j.canlet.2016.12.007
Nguyen, T. H. N., Bertrand, M. J. M., Sterpin, C., Achouri, Y., and De Backer, O. R. Y. (2010). Maged1, a new regulator of skeletal myogenic differentiation and muscle regeneration. BMC Cell Biol. 11:57. doi: 10.1186/1471-2121-11-57
Overholtzer, M., Zhang, J., Smolen, G. A., Muir, B., Li, W., Sgroi, D. C., et al. (2006). Transforming properties of YAP, a candidate oncogene on the chromosome 11q22 amplicon. Proc. Natl. Acad. Sci. U.S.A. 103, 12405–12410. doi: 10.1073/pnas.0605579103
Piccolo, S., Dupont, S., and Cordenonsi, M. (2014). The biology of YAP/TAZ: hippo signaling and beyond. Physiol. Rev. 94, 1287–1312. doi: 10.1152/physrev.00005.2014
Qiu, J., Zhu, J., Zhang, R., Liang, W., Ma, W., Zhang, Q., et al. (2019). miR-125b-5p targeting TRAF6 relieves skeletal muscle atrophy induced by fasting or denervation. Ann. Transl. Med. 7:456. doi: 10.21037/atm.2019.08.39
Salmena, L., Poliseno, L., Tay, Y., Kats, L., and Pandolfi, P. P. (2011). A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell 146, 353–358.
Siengdee, P., Trakooljul, N., Murani, E., Brand, B., Schwerin, M., Wimmers, K., et al. (2015). Pre- and post-natal muscle microRNA expression profiles of two pig breeds differing in muscularity. Gene 561, 190–198. doi: 10.1016/j.gene.2015.02.035
Su, R. (2015). Study on Hippo-YAP1 Pathway Involved in the Regulation of Hu Sheep Muscle Growth and Development. Ph. D. Thesis, Yangzhou University, Jiangsu.
Sun, C., De Mello, V., Mohamed, A., Quiroga, H. P. O., Garcia-Munoz, A., Al Bloshi, A., et al. (2017). Common and distinctive functions of the hippo effectors Taz and Yap in skeletal muscle stem cell function. Stem Cells 35, 1958–1972. doi: 10.1002/stem.2652
Takaguri, A., Kubo, T., Mori, M., and Satoh, K. (2017). The protective role of YAP1 on ER stress-induced cell death in vascular smooth muscle cells. Eur. J. Pharmacol. 815, 470–477. doi: 10.1016/j.ejphar.2017.09.033
Tan, P., Guo, Y.-H., Zhan, J.-K., Long, L.-M., Xu, M.-L., Ye, L., et al. (2019). LncRNA-ANRIL inhibits cell senescence of vascular smooth muscle cells by regulating miR-181a/Sirt1. Biochem. Cell Biol. 97, 571–580. doi: 10.1139/bcb-2018-0126
Tedesco, F. S., Dellavalle, A., Diaz-Manera, J., Messina, G., and Cossu, G. (2010). Repairing skeletal muscle: regenerative potential of skeletal muscle stem cells. J. Clin. Invest. 120, 11–19. doi: 10.1172/jci40373
Tremblay, A. M., Missiaglia, E., Galli, G. G., Hettmer, S., Urcia, R., Carrara, M., et al. (2014). The hippo transducer YAP1 transforms activated satellite cells and is a potent effector of embryonal rhabdomyosarcoma formation. Cancer Cell 26, 273–287. doi: 10.1016/j.ccr.2014.05.029
Vergara, H. M., Ramirez, J., Rosing, T., Nave, C., Blandino, R., Saw, D., et al. (2018). miR-206 is required for changes in cell adhesion that drive muscle cell morphogenesis in Xenopus laevis. Dev. Biol. 438, 94–110. doi: 10.1016/j.ydbio.2018.03.021
Wang, K., Degerny, C., Xu, M., and Yang, X.-J. (2009). YAP, TAZ, and Yorkie: a conserved family of signal-responsive transcriptional coregulators in animal development and human disease. Biochem. Cell Biol. Biochim. Biol. Cell. 87, 77–91. doi: 10.1139/o08-114
Wang, L., Zhou, L., Jiang, P., Lu, L., Chen, X., Lan, H., et al. (2012). Loss of miR-29 in myoblasts contributes to dystrophic muscle pathogenesis. Mol. Ther. 20, 1222–1233. doi: 10.1038/mt.2012.35
Wang, X. H., Hu, Z., Klein, J. D., Zhang, L., Fang, F., and Mitch, W. E. (2011). Decreased miR-29 Suppresses Myogenesis in CKD. J. Am. Soc. Nephrol. 22, 2068–2076. doi: 10.1681/asn.2010121278
Wang, Y., Zhang, C., Fang, X., Zhao, Y., Chen, X., Sun, J., et al. (2014). Identification and profiling of microRNAs and their target genes from developing caprine skeletal muscle. PLoS One 9:e96857. doi: 10.1371/journal.pone.0096857
Watt, K. I., Turner, B. J., Hagg, A., Zhang, X., Davey, J. R., Qian, H., et al. (2015). The Hippo pathway effector YAP is a critical regulator of skeletal muscle fibre size. Nat. Commun. 6:6048.
Watt, K. I., Judson, R., Medlow, P., Reid, K., Kurth, T. B., Burniston, J. G., et al. (2010). Yap is a novel regulator of C2C12 myogenesis. Biochem. Biophys. Res. Commun. 393, 619–624. doi: 10.1016/j.bbrc.2010.02.034
Wei, W., He, H. B., Zhang, W. Y., Zhang, H. X., Bai, J. B., Liu, H. Z., et al. (2013). miR-29 targets Akt3 to reduce proliferation and facilitate differentiation of myoblasts in skeletal muscle development. Cell Death Dis. 4:e668. doi: 10.1038/cddis.2013.184
Wu, H., Ren, Y., Li, S., Wang, W., Yuan, J., Guo, X., et al. (2012). In vitro culture and induced differentiation of sheep skeletal muscle satellite cells. Cell Biol. Int. 36, 579–587. doi: 10.1042/cbi20110487
Yue, G. H. (1996). Reproductive characteristics of Chinese Hu sheep. Anim. Reprod. Sci. 44, 223–230. doi: 10.1016/0378-4320(96)01562-x
Zhan, S., Qin, C., Li, D., Zhao, W., Nie, L., Cao, J., et al. (2019). A novel long noncoding RNA, lncR-125b, promotes the differentiation of goat skeletal muscle satellite cells by sponging miR-125b. Front. Genet. 10:1171. doi: 10.3389/fgene.2019.01171
Zhang, L.-Q., Cui, H., Wang, L., Fang, X., and Su, S. (2017). Role of microRNA-29a in the development of diabetic retinopathy by targeting AGT gene in a rat model. Exp. Mol. Pathol. 102, 296–302. doi: 10.1016/j.yexmp.2017.02.004
Zhao, L., Shi, X., Wang, N., Liu, C., and Wang, J. (2020). YAP1, targeted by miR-375, enhanced the pro-angiogenesis of airway smooth muscle cells in asthma via STAT3 activation. Cell Cycle 19, 1275–1284. doi: 10.1080/15384101.2020.1746874
Zhao, X., Sun, J., Chen, Y., Su, W., Shan, H., Li, Y., et al. (2018). lncRNA PFAR promotes lung fibroblast activation and fibrosis by targeting mir-138 to regulate the YAP1-twist axis. Mol. Ther. 26, 2206–2217. doi: 10.1016/j.ymthe.2018.06.020
Zheng, J., Peng, B., Zhang, Y., Ai, F., and Hu, X. (2019). miR-9 knockdown inhibits hypoxia-induced cardiomyocyte apoptosis by targeting Yap1. Life Sci. 219, 129–135. doi: 10.1016/j.lfs.2019.01.014
Keywords: YAP1, miR-29a, lncRNA, skeletal muscle, satellite cell, Hu sheep
Citation: Wu T, Wang S, Wang L, Zhang W, Chen W, Lv X, Li Y, Hussain Z and Sun W (2020) Long Noncoding RNA (lncRNA) CTTN-IT1 Elevates Skeletal Muscle Satellite Cell Proliferation and Differentiation by Acting as ceRNA for YAP1 Through Absorbing miR-29a in Hu Sheep. Front. Genet. 11:843. doi: 10.3389/fgene.2020.00843
Received: 13 May 2020; Accepted: 13 July 2020;
Published: 07 August 2020.
Edited by:
Naoyuki Kataoka, The University of Tokyo, JapanReviewed by:
Zhenhui Li, South China Agricultural University, ChinaTao Zhong, Sichuan Agricultural University, China
Siyuan Zhan, Sichuan Agricultural University, China
Copyright © 2020 Wu, Wang, Wang, Zhang, Chen, Lv, Li, Hussain and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Sun, ZGt4bXN1bndlaUAxNjMuY29t
†These authors share first authorship