
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 04 December 2018
Sec. Epigenomics and Epigenetics
Volume 9 - 2018 | https://doi.org/10.3389/fgene.2018.00610
 Alexandre A. Lussier1,2*†
Alexandre A. Lussier1,2*† Tamara S. Bodnar1
Tamara S. Bodnar1 Matthew Mingay3
Matthew Mingay3 Alexandre M. Morin2Martin Hirst3,4
Alexandre M. Morin2Martin Hirst3,4 Michael S. Kobor2,5‡
Michael S. Kobor2,5‡ Joanne Weinberg1‡
Joanne Weinberg1‡Background: Prenatal alcohol exposure (PAE) can alter the development of neurobiological systems, leading to lasting neuroendocrine, neuroimmune, and neurobehavioral deficits. Although the etiology of this reprogramming remains unknown, emerging evidence suggests DNA methylation as a potential mediator and biomarker for the effects of PAE due to its responsiveness to environmental cues and relative stability over time. Here, we utilized a rat model of PAE to examine the DNA methylation profiles of rat hypothalami and leukocytes at four time points during early development to assess the genome-wide impact of PAE on the epigenome and identify potential biomarkers of PAE. Our model of PAE resulted in blood alcohol levels of ~80–150 mg/dl throughout the equivalent of the first two trimesters of human pregnancy. Hypothalami were analyzed on postnatal days (P) 1, 8, 15, 22 and leukocytes at P22 to compare central and peripheral markers. Genome-wide DNA methylation analysis was performed by methylated DNA immunoprecipitation followed by next-generation sequencing.
Results: PAE resulted in lasting changes to DNA methylation profiles across all four ages, with 118 differentially methylated regions (DMRs) displaying persistent alterations across the developmental period at a false-discovery rate (FDR) < 0.05. In addition, 299 DMRs showed the same direction of change in the hypothalamus and leukocytes of P22 pups at an FDR < 0.05, with some genes overlapping with the developmental profile findings. The majority of these DMRs were located in intergenic regions, which contained several computationally-predicted transcription factor binding sites. Differentially methylated genes were generally involved in immune function, epigenetic remodeling, metabolism, and hormonal signaling, as determined by gene ontology analyses.
Conclusions: Persistent DNA methylation changes in the hypothalamus may be associated with the long-term physiological and neurobehavioral alterations in observed in PAE. Furthermore, correlations between epigenetic alterations in peripheral tissues and those in the brain will provide a foundation for the development of biomarkers of fetal alcohol spectrum disorder (FASD). Finally, findings from studies of PAE provide important insight into the etiology of neurodevelopmental and mental health disorders, as they share numerous phenotypes and comorbidities.
Early-life environments influence the development of biological/neurobiological systems, leading to long-term consequences in offspring (Godfrey and Robinson, 1998; Hanson and Gluckman, 2008). In particular, prenatal alcohol exposure (PAE) can result in Fetal Alcohol Spectrum Disorders (FASD) in humans, which are associated with a wide variety of adverse effects. Exposure to alcohol at high levels throughout pregnancy can result in Fetal Alcohol Syndrome (FAS), characterized by growth retardation, a characteristic facial dysmorphology, and multiple central nervous system alterations. Exposure to alcohol at levels that do not produce FAS can result in either partial FAS (pFAS), where only some of the diagnostic features are observed, or in numerous alcohol-related neurobehavioral effects (alcohol-related neurodevelopmental disorder, ARND; Stratton et al., 1996). The degree to which alcohol affects development depends on a variety of factors such as timing, pattern, and level of alcohol exposure, overall maternal health and nutrition, and genetic background (Pollard, 2007), which may influence the broad range of effects of in utero alcohol exposure and the relatively high prevalence of FASD (Riley et al., 2011; May et al., 2018). Importantly, PAE can alter the development, function, and regulation of numerous neurobiological and physiological systems, giving rise to lasting deficits across the spectrum of FASD, including, but not limited to cognitive and behavioral deficits, impairment to self-regulation and adaptive functioning, immune dysregulation, and increased vulnerability to mental health problems across the lifespan (Zhang et al., 2005; Mattson et al., 2011; Pei et al., 2011).
The hypothalamus is highly susceptible to the programming effects of PAE (Eguchi, 1969; Matthews, 2002; Hellemans et al., 2008; Weinberg et al., 2008). The hypothalamus plays key roles in neuroendocrine regulation, autonomic regulation, and homeostatic control, regulating growth, sleep/wake behavior, circadian rhythms, metabolism, body temperature, and other vital functions (Card and Swanson, 2012). Data from both human clinical cohorts and animal models of FASD have identified alterations to physiological functions associated with the hypothalamus. For example, infants exposed to alcohol in utero show both elevated basal and post-stress levels of cortisol, and children with FASD who experience early life adversity exhibit dysregulation of the cortisol circadian rhythm (Keiver et al., 2015; McLachlan et al., 2016). Similarly, in animal models of PAE, exposed offspring exhibit hyperresponsiveness to stressors as well as altered central regulation of hypothalamic-pituitary-adrenal (HPA) axis activity (Ramsay et al., 1996; Jacobson et al., 1999; Haley et al., 2006; Weinberg et al., 2008). Furthermore, PAE also alters sleep patterns and circadian rhythms, results in deficits in thermoregulation, and is associated with inappropriate feeding behavior (Jones and Smith, 1973; Zimmerberg et al., 1987; Earnest et al., 2001; Sei et al., 2003; Chen et al., 2012; Werts et al., 2014). These deficits often persist across the life course, suggesting that alcohol exposure during prenatal life may alter developmental trajectories to increase the risk of adverse outcomes (Hellemans et al., 2010a). In the context of the fetal programming hypothesis, early environmental or non-genetic factors, including maternal undernutrition, stress, and exposure to drugs or other toxic agents, can permanently organize or imprint physiological and neurobiological systems and increase adverse cognitive, adaptive, and behavioral outcomes, as well as vulnerability to diseases or disorders later in life (Godfrey and Robinson, 1998; Hanson and Gluckman, 2008; Swanson et al., 2009). As the underlying mechanisms of these effects begin to emerge, it has become apparent that epigenetic mechanisms might be important candidates for the programming effects of PAE, linking early life environmental factors and neurobiological outcomes while influencing health and behavior well into adulthood (Yuen et al., 2011; Shulha et al., 2013). The term epigenetics broadly refers to the modifications of DNA and its packaging that alter DNA accessibility, which modulates gene expression and cell functions without changes to underlying genomic sequences (Bird, 2007). These epigenetic factors include direct modifications to DNA, post-translational modification of histones, and non-coding RNAs.
DNA methylation currently is the most studied epigenetic modification and involves the covalent attachment of a methyl group to the 5′ position of cytosine, typically occurring at cytosine-guanine dinucleotide (CpG) sites (Jones and Takai, 2001). Although closely linked to the regulation of gene expression, the association between DNA methylation and transcription depends on genomic context. Whereas, DNA methylation typically represses gene expression when located in promoter regions, its effects are more variable for CpGs residing in gene bodies and intergenic regions. DNA methylation can also directly control transcription factor binding to gene regulatory regions, such as enhancers, to modulate gene expression patterns (Tate and Bird, 1993). In addition to this role in transcriptional control, DNA methylation has been associated with altered mRNA splicing when located within introns, and its presence within certain exons may potentially regulate alternative transcriptional start sites (Maunakea et al., 2010, 2013; Shukla et al., 2011). Furthermore, DNA methylation is closely linked to several important developmental processes, including genomic imprinting, tissue specification, and differentiation, suggesting a role in the regulation of cellular functions and developmental trajectories (Smith and Meissner, 2013; Ziller et al., 2014). Perhaps most importantly, DNA methylation can be responsive to environmental influences and these changes can possibly be inherited through cell divisions to potentially persist throughout life (Hanson et al., 2011; Langevin et al., 2011; Yuen et al., 2011). An additional interesting aspect of DNA methylation is its emerging role as a potential biomarker of early-life exposures, as it is easily quantifiable, stable over time, and can be obtained from readily available peripheral tissues, such as buccal epithelial cells and white blood cells (Bock, 2009).
Given its role in the regulation of gene expression and cell function, as well as its responsiveness to environmental factors, DNA methylation provides an attractive mechanism for the biological embedding of the persistent deficits caused by PAE. Mounting evidence suggests a potential role in the etiology of PAE-induced deficits, as numerous studies have identified alterations to epigenetic programs in the central nervous system of animals exposed to alcohol in utero across various levels and stages of exposure (reviewed in Lussier et al., 2017). These range from differences in bulk levels of DNA methylation to genome-wide changes in DNA methylation patterns, supporting the hypothesis that PAE can alter the epigenome (Bekdash et al., 2013; Laufer et al., 2013). Although genome-wide studies have been performed on whole brains in mice, few studies have focused on targeted brain regions. Among those that have, PAE was shown to be associated with altered DNA methylation status of the POMC promoter in the rat hypothalamus (Bekdash et al., 2013; Ngai et al., 2015). As a key regulator of the stress response, alterations to this gene may reflect broader alterations to the regulatory functions of the hypothalamus. Studies from clinical cohorts of children with FASD have identified widespread changes to DNA methylation patterns in peripheral tissues (Laufer et al., 2015; Portales-Casamar et al., 2016; Lussier et al., 2018). However, alterations to central tissue are difficult if not impossible to directly assess in clinical populations, and while peripheral tissues are more easily accessible, changes in these cells may not fully reflect alterations in the brain (Berko et al., 2014). Furthermore, biological embedding of PAE's effects earlier in development could potentially lead to more systemic effects on the epigenome, which would be reflected by alterations present across a variety of tissues.
Currently, the genome-wide impact of PAE on DNA methylation within the hypothalamus remains unknown (Bekdash et al., 2013; Ngai et al., 2015). To address this gap, we utilized a well-established rat model of PAE to assess whether moderate to moderately high levels of first and second trimester-equivalent alcohol exposure alters DNA methylation profiles in the early postnatal period, and whether altered sites of DNA methylation could serve as biomarkers of gestational alcohol exposure if also identified in peripheral tissues. Using methylated DNA immunoprecipitation and next-generation sequencing (meDIP-seq), we identified statistically significant differentially methylated regions (DMRs) in female PAE animals that persisted across preweaning development of the hypothalamus. In parallel, we identified concordant DNA methylation alterations in white blood cells and the hypothalamus of female PAE animals compared to controls on postnatal day (P) 22. Our findings suggest that: (1) PAE alters DNA methylation patterns in both central and peripheral tissues, potentially reprogramming neurobiological/physiological systems and influencing the deficits observed in FASD; and (2) DNA methylation patterns in peripheral tissue reflect some changes in brain, which could serve as potential biomarkers for central alterations induced by PAE.
Details of the procedures for breeding and handling have been published previously (Bodnar et al., 2016). Briefly, outbred male and nulliparous female Sprague-Dawley rats were obtained from Charles River Laboratories (St. Constant, Quebec, Canada). Following a one-week habituation period, each female (n = 39) was pair-housed with a male and vaginal lavage samples were collected daily for estrous cycle staging and to check for the presence of sperm, indicating gestation day (GD) 1. Pregnant dams were singly housed and assigned to one of three prenatal treatment groups: Prenatal alcohol exposure (PAE)—ad libitum access to liquid ethanol diet, 36% ethanol-derived calories, 6.37% v/v, n = 13; Pair-fed (PF)—liquid-control diet, maltose-dextrin isocalorically substituted for ethanol, in the amount consumed by an PAE partner, g/kg body weight/GD, n = 14; or Control (C)—pelleted version of the liquid control diet, ad libitum, n = 12. All animals had ad libitum access to water. Experimental diets (Weinberg/Keiver Liquid Ethanol Diet #710324, Weinberg/Keiver Liquid Control Diet #710109, and Pelleted Control Diet #102698, Dyets Inc., Bethlehem, PA) were fed from gestation days 1–21, and then replaced with laboratory chow. Litters were weighed and culled at birth to 6 males and 6 females, when possible. Pregnancy outcomes and body weights can be found in Supplementary Table 1. Blood alcohol levels were measured as previously reported and ranged from 80 to 150 mg/dl in PAE females (Hellemans et al., 2010b; Uban et al., 2010; Bodnar et al., 2016).
We focused our investigation of PAE-induced epigenetic alterations on female animals, as sexually dimorphic effects of PAE are widely reported (Lee and Rivier, 1996; Weinberg et al., 2008) and females are generally underrepresented in molecular and genome-wide studies of FASD (Lussier et al., 2017). On postnatal day (P) 1, 8, 15, and 22 female offspring (max 1/litter) were decapitated, trunk blood collected (at P22 only), and brains removed and weighed; the hypothalamus was then quickly dissected and frozen on dry ice in RNAlater (n = 7–11/age/group; Figure 1; Qiagen, Hilden, Germany). WBCs were isolated using Ficoll-Paque (GE Healthcare, Uppsala, Sweden), which isolates peripheral blood mononuclear cells (PBMCs). All tissue collected was left at 4°C for 24 h to allow for complete permeation by RNAlater and then frozen at −80°C until DNA extraction, as per manufacturer instructions. WBCs were stored in RNAlater at −80°C until DNA extraction (Qiagen, Hilden, Germany). Due to the large number of animals associated with the experimental design of this study, animals were collected across four different cohorts (experimental breedings), spanning January 2012–December 2013.
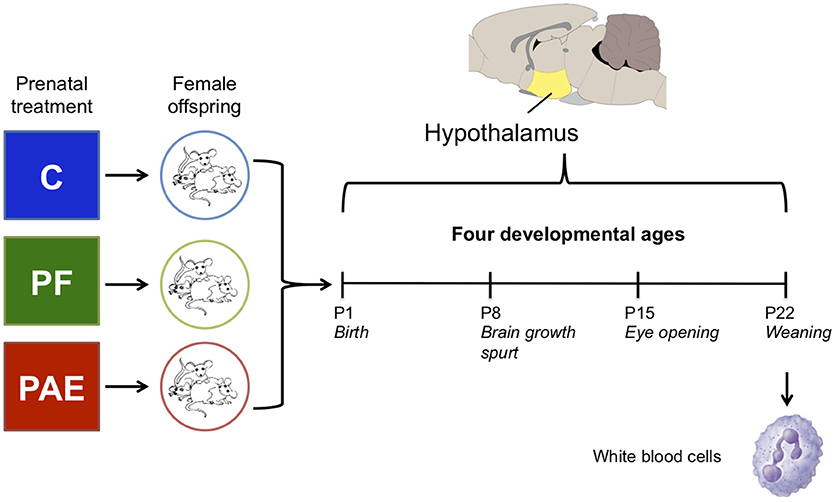
Figure 1. Overview of the experimental design. We collected the hypothalamus of female offspring from one of three prenatal treatment groups on postnatal days (P) 1, 8, 15, and 22. In parallel, white blood cells were collected on P22 from the same animals as the hypothalamus samples. Each group/age/tissue was composed of four samples for DNA methylation analysis by methylated DNA immunoprecipitated and next-generation sequencing (meDIP-seq).
Analysis of blood composition was done on samples from a separate cohort of animals, which were bred at a later time but under the same conditions as the main cohort. Briefly, on P22, trunk blood was collected from female offspring (C: n = 6; PF: n = 5; PAE: n = 5), and analyzed using the Advia120 hematology system, which assesses complete blood counts and differential WBC counts (CBC/Diff function). The reported values include counts for neutrophils, lymphocytes, monocytes, eosinophils, basophils, and large unclassified cells (Supplementary Table 2).
Total RNA and DNA were simultaneously extracted from the hypothalamus and white blood cells (n = 4/group/age/tissue; Qiagen AllPrep DNA/RNA Mini kit, Hilden, Germany). Frozen tissue was thawed on ice, quickly weight, and placed in lysis buffer for 5 min. Homogenization was performed by 5 strokes of an 18 G needle, 10 strokes of a 20 G needle, and 10 strokes of a 23 G needle. The resulting homogenate was centrifuged at 21,000 g for 3 min and the supernatant was collected for DNA and RNA extraction. White blood cells were thawed on ice and then centrifuged at 10,000 g for 10 min. RNAlater was carefully removed without disturbing the cell pellet and cells were resuspended in lysis buffer. The cells were then frozen at −80°C to disrupt cell membranes and thawed on ice. The resulting homogenate was used for DNA and RNA extraction. DNA concentration was assessed using Qubit Fluorometric Quantitation (Life Technologies, Carlsbad, USA). Full developmental data on the animals can be found in Supplementary Table 3.
Our methylated DNA immunoprecipitation followed by next-generation sequencing (meDIP-seq) procedures were adapted from a previously published protocol and are outlined in detail below (Taiwo et al., 2012). Importantly, meDIP-seq provides qualitative measures of methylated CpG levels genome-wide and, unlike bisulfite dependent assays, is specific for methyl-cytosine. As previously noted, meDIP-seq coverage is highly reproducible with reduced coverage in regions of sparse CpG density, showing >99% concordance with bisulfite sequencing methods (Harris et al., 2010). However, our experimental design is based on treatment vs. controls and thus only queries genomic regions that are addressable by meDIP-seq reads. As such, we cannot eliminate the possibility that additional DMRs are present within genomic regions with sparse CpG coverage.
For each sample, 500 ng of DNA were diluted in a total volume of 60 μL of EB buffer (Qiagen, Hilden, Germany). DNA was then transferred to a 96-well plate and sheared for 1 h using the Covaris Focused-ultrasonicator. DNA was purified using Ampure XP in 20% polyethylene glycol (PEG) beads to obtain fragments sized from 200 to 500 basepairs (Beckman-Coulter, Brea, USA). Library preparation was performed on the Bravo Automated Liquid Handling Platform (Agilent, Santa Clara, USA) using the TruSeq DNA PCR-Free Sample Preparation Kit (Illumina, San Diego, USA). Following end-repair and A-tailing, adapters were ligated overnight at room temperature. PCR-free library preparation allowed for the conservation of methylated cytosines for subsequent methylated DNA immunoprecipitation. Finally, DNA was resuspended in 35 μL of EB buffer (Qiagen, Hilden, Germany). DNA was quality controlled using Qubit Fluorometric Quantitation and the DNA 1000 Bioanalyzer 2100 kit (Agilent, Santa Clara, USA) to verify DNA concentration and fragment size (250–550 bp).
For each sample, 400 ng of the sequencing library DNA were diluted in a total volume of 50 μL of IP Buffer (10 mM sodium phosphate buffer, pH 7.0, 140 mM NaCl, 0.05% triton). DNA was denatured by incubation at 95°C for 10 min, followed by the addition of 48 μL ice-cold IP buffer and incubation on ice for 10 min. Two microliters of anti-5-methylcytosine antibody (Eurogenetec, Liège, Belgium), diluted to 1/50 in IP buffer (1 μL of antibody per 1 μg of DNA ratio), were added to each sample. Immunoprecipitation reactions were incubated for 16 h at 4°C with overhead rotation. Following two 5 min washes with 150 μL of 0.1% BSA/PBS, 50 μL of Dynabeads Protein G were incubated with 5 μL of secondary antibody (rabbit anti-mouse IgG; Jackson Immunoresearch, West Grove, USA) in 45 μL ice-cold IP buffer for 15 min at room temperature with overhead rotation. Beads were washed twice with IP buffer to remove unbound secondary antibody and resuspended in 50 μL IP buffer. The antibody-bound beads were added to the immunoprecipitation reactions and incubated for 2 h at 4°C with overhead rotation. Beads were then washed 6 times with 150 μL of ice-cold IP buffer and resuspended in 98.97 μL of Proteinase K digestion buffer (TE with 0.5% SDS). Following the addition of 1.25 μL Proteinase K (20 mg/mL; Qiagen, Hilden, Germany), samples were incubated in a thermomixer for 2 h at 55°C with a rotation speed of 1,250 rpm. The reaction was then allowed to cool at room temperature for 15 min. Supernatant was collected and bead cleanup was performed using equal volume SeraMag beads with 30% PEG. DNA was resuspended in 35 μL of EB buffer (Qiagen, Hilden, Germany).
To reduce PCR amplification bias, two separate reactions of the same meDIP sample were run in parallel using the following PCR amplification cycle conditions. The reaction mixes were as follows: 15 μL DNA, 27 μL H2O, 12 μL 5X HF buffer, 1.5 μL DMSO, 1.0 μL paired-end primer (Illumina), 0.5 μL Phusion High-Fidelity DNA polymerase (New England Biolabs), 2 μL indexing primer (Illumina—specific to each sample). The amplification cycle was as follows: 98°C for 1 min, 12X (98°C for 15 s, 65°C for 30 s, 72°C for 30 s), 72°C for 5 min. Reactions from the same sample were pooled and bead cleanup was performed using SeraMag beads in 20% PEG (102 μL of beads per 120 μL of reaction). DNA was resuspended in a final volume of 35 μL of EB buffer.
Indexed meDIP libraries were combined in 3 pools of 20 samples each, distributing samples evenly by tissue, age, and prenatal treatment across all three sets. Next-generation sequencing was performed on the pools by the Genome Sciences Centre in Vancouver, BC, Canada. Each sample pool was run on two HiSeq lanes, which produced approximately 600,000,000 paired-end reads of 125 bases per lane. Fastq files were aligned to the most current rat genome (Rn6, July 2014) using the Burrows-Wheeler Transform (BWA) tool to obtain.bam files (Li and Durbin, 2009). Bam files were filtered using samtools to remove duplicate reads, unpaired reads, and reads with a minimum quality score below 10. Following alignment and filtering, each the two runs for each sample were merged using samtools to obtain a single bam file for each sample (Li et al., 2009). Supplementary Table 11 shows sequencing related information: sample pool, sample index, number of raw reads, number of filtered reads, and total number of reads/sample.
Model-based analysis of ChIP-seq (MACS2; version 2.1.0.20140616) was used to identify enriched regions of DNA methylation across the genome (Zhang et al., 2008). The peak calling to identify peak regions (DNA methylation windows) was performed using the “callpeaks” function on paired end bam files with no control input and the following options: –f BAMPE–m 5 50–bw 300–g 2.9e9–q 0.05. Each sample was modeled individually, generating 60 total peaksets. These were imported into R using the DiffBind package (Stark and Brown, 2011). As all samples had slightly different predicted peaks, peaksets were combined into common regions using the dba.count function in DiffBind, which removed peaks found in < 3 samples across the entire dataset and provided the total number of reads within each peak/sample. This created a final dataset of 469,162 peaks and 48 samples from the developmental profile of the hypothalamus, and a final dataset of 350,960 peaks and 24 samples in the P22 hypothalamus and WBC (tissue-concordant) peakset.
First, the total reads within each peak were adjusted to reads/kilobase by dividing the number of reads within each region by their length. In turn, these were converted to reads per kilobase per million (RPKM) by dividing the reads/kilobase by the total number of reads found in the predicted peaks to account for differences in sequencing depth between samples. The samples in the developmental dataset were highly correlated (r > 0.95 for all samples), with samples clustering most closely with animals of the same age (Supplementary Figure 1). No outliers were detected in this first pass analysis.
Principal component analysis of the normalized RPKM data revealed significant levels of variation associated with batch effects. Notably, meDIP and DNA extraction rounds were associated with a large proportion of variation within the dataset. However, both these factors were highly confounded with age, as samples for each age were extracted as a batch and all P22 samples were immunoprecipitated in a separate batch (Supplementary Figure 2). To account for these effects, ComBat correction, from the SVA package, was performed on the RPKM data from the hypothalamic samples to correct the effects of meDIP round and DNA extraction round in the dataset (Leek et al., 2012). Age was also slightly confounded with the breeding from which animals were collected, as not all ages were sampled from each of the different cohorts. Interestingly, some partial effects of breeding remained in the dataset following ComBat correction, suggesting that this covariate was not fully confounded with age. Furthermore, prenatal treatment accounted for a larger proportion of variance within the dataset following ComBat correction, suggesting that the removal of batch effects might allow for the identification of more subtle effects of PAE. The corrected and normalized RPKM values obtained from ComBat were used for plotting purposes, but were converted back to reads/kilobase for downstream statistical analyses.
The tissue-concordant dataset was preprocessed and normalized as described above. Samples were highly correlated within tissue (r > 0.96), the main driver of DNA methylation patterns, and well correlated within the same animals (r > 0.92). However, one PF WBC sample clustered with the hypothalamus samples, suggesting that it may have been mislabeled during processing. As such, this sample was removed from the dataset, resulting in a dataset of 23 samples (Supplementary Figure 3). Principal component analysis of the normalized tissue-concordant RPKM data revealed significant levels of variation associated with DNA extraction round batch effects (Supplementary Figure 4). Tissue type was the covariate most strongly associated with variance in the dataset, although it was slightly confounded with extraction round. While ComBat correction was used to account for the effects of DNA extraction round in the tissue-concordant dataset, this approach limited our ability to identify tissue-specific differences, as it removed the majority of tissue-associated variance from the dataset. Again, prenatal treatment was associated with a larger proportion of variance within the dataset following ComBat correction. Interestingly, breeding once again remained a major contributor to variability within the dataset, suggesting that differences between cohorts may have an important influence on epigenetic patterns.
Using previously characterized transcriptomic profiles from mouse neurons, oligodendrocytes, and astrocytes, we identified DNA methylation peaks within genes that are specifically expressed in each different cellular subtype (1.5-fold expression difference compared to other cell types; Cahoy et al., 2008). Given the relationship between gene expression and epigenetic patterns, it is possible that alterations to the DNA methylation levels of these genes could reflect changes in the cell-type proportions within this dataset. However, the majority of the peaks in the dataset were located within intergenic regions, with no annotated associations with these genes, reducing our ability to capture cell-type related differences. As such, only regions directly located within neuron-, oligodendrocyte-, or astrocyte-specific genes were removed from further analyses to reduce the potential confounding factor of cell type, resulting in a dataset of 451,112 peaks for downstream analyses of the hypothalamus.
Linear modeling was performed using edgeR, which is typically used to analyze RNA-seq count data and includes a factor to account for the number of reads in each sample (Robinson et al., 2010a; Nikolayeva and Robinson, 2014). This method was used to identify differentially methylated regions (DMRs) that were consistently different between PAE animals and both control groups across the different ages and tissues. For both analyses, the model accounted for the effects of collection during different breedings, and p-values were corrected for multiple-testing using the Benjamini-Hochberg method. Statistically significant DMRS at a false discovery rate (FDR) < 0.05 were obtained for the following contrasts: PAE vs. C, PAE vs. PF, and C vs. PF. The final PAE-specific DMRs were statistically significant in both PAEvC and PAEvPF, and were not found in the PFvC contrasts.
Custom annotations were built for each peakset using the UCSC genome browser gene annotations. Briefly, genomic coordinates of all CpG islands, exons, introns, promoters (TSS-200 bp and TSS-1500 bp), 3′ untranslated regions (UTR), 5′ UTRs for the rn6 genome were obtained as bed files from the table browser. In parallel, MeDIP-seq peaks were converted to the bed file format and the overlap of genomic features with MeDIP-seq peaks was computed iteratively using the intersectBed function from bedtools, retaining only the peaks that contained the assessed genomic feature (Quinlan and Hall, 2010). The overlaps were concatenated into a single annotation set in R, where individual peaks contained information for each potential genomic feature. Of note, regions spanning both introns and exons were deemed intron/exons boundaries. P-values for genomic feature enrichment analyses were calculated by computing background levels of genomic features on 1,000 random subsets of DMRs, using the same number of PAE-specific DMRs.
Enrichment of different transcription factors binding sites (TFBS) in PAE-specific DMRs was assessed using the motifEnrichment function of the PWMEnrich package (Stojnic and Diez, 2013). DMR DNA sequences were obtained from the UCSC genome browser (Rn6 genome). As no binding motifs were available for the Rattus norvegicus genome, motifs from the Mus musculus genome were obtained from the PWMEnrich.Mmusculus.background. Motifs were summarized using the groupReport function. P-values were calculated by performing enrichment analysis on 1,000 random subsets of DMRs, using the same number of PAE-specific DMRs for each analysis to assess background levels of each TFBS in the different peaksets.
The gene-score resampling (GSR) tool of ErmineJ (version 3.0.2) was used to identify gene function enrichment in the differentially methylated genes including the Gene Ontology (GO) annotations molecular function, biological process, and cellular component (Lee et al., 2005; Gillis et al., 2010). The ermineJ GSR tool was set with the following parameters: max gene set size = 2,000; min gene set size = 2; iterations = 10,000. Once again, statistically significant associations (p < 0.05 and multifunctionality score < 0.05) were obtained for the following contrasts: PAE vs. C, PAE vs. PF, and C vs. PF. The final PAE-specific GO terms were statistically significant in both PAEvC and PAEvPF, and were not found in the PFvC contrasts. Importantly, ermineJ accounts for the multiple functions assigned to many genes, generating a multifunctionality p-value to reduce the bias of gene ontology analyses toward pathways with numerous common genes (Gillis and Pavlidis, 2011; Ballouz et al., 2017).
DNA from the same samples as above were subjected to bisulfite conversion using the Zymo EZ DNA Methylation Kit (Zymo Research, Irvine, California), which converts DNA methylation information into sequence base differences by deaminating unmethylated cytosines to uracil while leaving methylated cytosines unchanged. Bisulfite pyrosequencing assays were designed with PyroMark Assay Design 2.0 (Qiagen, Hilden, Germany; Supplementary Table 4). The regions of interest were amplified by PCR using the HotstarTaq DNA polymerase kit (Qiagen, Hilden, Germany) as follows: 15 min at 95°C, 45 cycles of 95°C for 30 s, 58°C for 30 s, and 72°C for 30 s, and a 5 min 72°C final extension step. For pyrosequencing, single-stranded DNA was prepared from the PCR product with the Pyromark™ Vacuum Prep Workstation (Qiagen, Hilden, Germany) and the sequencing was performed using sequencing primers on a Pyromark™ Q96 MD pyrosequencer (Qiagen, Hilden, Germany). The quantitative levels of methylation for each CpG dinucleotide were calculated with Pyro Q-CpG software (Qiagen, Hilden, Germany). Of note, only PAE and C animals were assessed by bisulfite pyrosequencing. We selected several DMRs for verification by bisulfite pyrosequencing based on their potential role in PAE-induced deficits, mainly focusing on their associated gene.
Our initial analysis of this dataset aimed to identify persistent alterations to DNA methylation patterns in the female rat hypothalamus across early development (P1, 8, 15, and 22) using methylated DNA immunoprecipitation (meDIP-seq). These ages were selected as they represent key developmental time points, including birth (P1), the brain growth spurt (P8), eye opening (P15), and weaning (P22; Dobbing and Sands, 1979; McCormick and Mathews, 2010).
As cell type proportions are a major driver of DNA methylation patterns, we first removed peaks that were located within genes specifically expressed in neurons, astrocytes, or oligodendrocytes, resulting in a dataset of 48 samples and 451,112 peaks. We assessed the cell-type associated peaks independently by linear modeling (18,050 peaks), identifying few differences among prenatal groups (Supplementary Figure 5). Although this filter was limited by the use of genic and promoter regions, as well as gene expression patterns from isolated neurons, glia, and oligodendrocytes, this partial analysis suggested that few differences in the proportions of the three major brain cell types were present in the dataset. In addition to cell type differences, highly-methylated regions associated with age could have also introduced additional variation into our data. As such, we included age as a covariate in our statistical models, which may partially account for these age-induced effects through statistical means.
To assess persistent alterations to DNA methylation patterns caused by PAE, we performed linear modeling on the hypothalamic samples across all ages with a model that also accounted for differences across breeding cohorts. Using contrast analyses, we successfully identified 118 PAE-specific DMRs at an FDR < 0.05 that persisted across all four developmental ages and showed consistently different DNA methylation levels between PAE animals and controls (Figure 2; Supplementary Table 5). Of these, 47 were up-methylated and 75 were down-methylated in PAE animals vs. control groups, and their sizes ranged from 316 to 1027 bp (median = 494.5 bp).
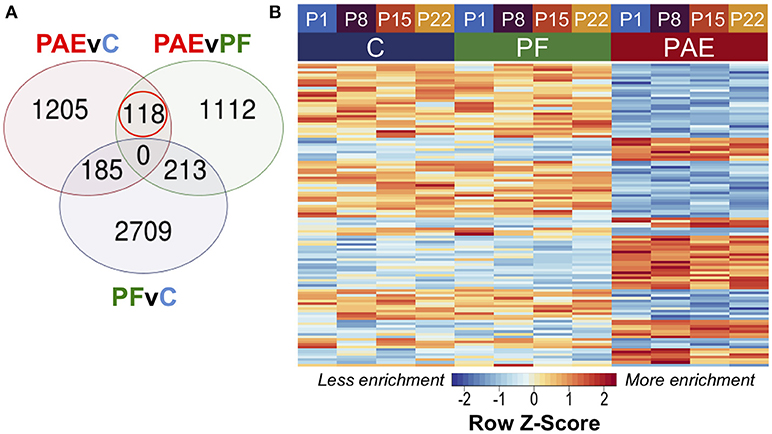
Figure 2. PAE-specific DMRs across pre-weaning development of the hypothalamus. (A) Contrast analysis revealed 118 PAE-specific differentially methylated regions (DMR), which were significantly different in PAE vs. C animals and PAE vs. PF animals, but not significantly different between PF vs. C. (B) The DMRs showed consistent difference between PAE animals and controls across ages. Each row represents a different DMR, while each column shows the mean for all animals within that group/age (n = 4). Reads per kilobase per million (RPKM) data were scaled and centered to produce a Z-score for each DMR, where those in blue showed less DNA methylation enrichment and those in red showed more enrichment.
Overall, 34 DMRs were located in genes, particularly within those involved in dopamine signaling (Drd4), the immune response (Ifih1, Ccrl2, Il20ra), and blood-brain barrier function (Plvap). Of note, two overlapping genes, Golga4 and Ctdspl, contained two separate DMRs, and were the only genes with multiple DMRs. Although the entire DMR set did not show any significant differences in genomic location enrichment compared to the background of the dataset, the up-methylated DMRs displayed significantly more enrichment in CGI and exons compared to random chance (p < 0.05; Figure 3). Furthermore, the majority of DMRs were located in intergenic regions, and while these were not significantly enriched compared to the entire dataset, these results suggested that intergenic regions may be more responsive to the influence of PAE on the epigenome, and may contain important regulatory regions that are not yet annotated in the rat genome.
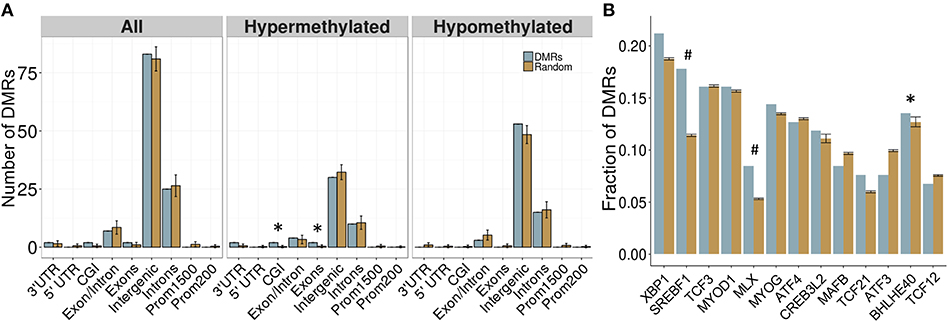
Figure 3. Enrichment patterns of the developmental DMRs. (A) Genomic feature enrichment profile of all, up-methylated, and down-methylated DMRs. The probe counts for each feature (blue) were compared to the results from permutation analyses of 118 random regions (orange), which were used to compute the p-value. The majority of DMRs were located in intergenic regions or introns. Up-methylated regions in PAE animals contained more CpG islands (CGI) and exons than expected by chance (p < 0.05). (B) Overrepresentation analysis of transcription factor binding sites in the DMRs. Only BHLHE40 showed higher enrichment in the PAE-specific DMRs (blue) than by random chance (orange; p < 0.05), although SREBF1 and MLX trended toward significance (p < 0.1). *p < 0.05, #p < 0.1.
Of note, meDIP-seq provides relative levels of DNA methylation based on enrichment scores, and thus, the magnitude of change (i.e., % methylation) was not assessed using this method. Nevertheless, 38 of the DMRs showed at least 1.5-fold change in DNA methylation levels in PAE animals vs. controls, including those in Drd4, Plvap, and Cntnap5c (Supplementary Table 6), suggesting that PAE could induce robust alterations to DNA methylation patterns in the hypothalamus.
To follow up on the large proportion of intergenic regions in the PAE-specific DMRs, we assessed the enrichment of transcription factor binding sites (TFBS) within these regions using binding motifs from the mouse genomes. Although the overlap between the rat and mouse genomes is not perfect (~70%), the rodent family shares many genomic characteristics and this analysis provided an important first pass analysis of potential regulatory factors within these regions. Following multiple-test correction (FDR < 0.05), few TFBS were enriched within these regions compared to background levels. However, the BHLHE40 binding motif was significantly enriched within the PAE-specific DMRs (p < 0.05), while the SREBF1 and MLX motifs trended toward significance (p < 0.10; Figure 3B). These results suggest that certain transcription factors may play a role in the long-term reprogramming of hypothalamic functions by PAE and may act in concert with other factors to sculpt the epigenome and downstream phenotypes.
We performed GO analysis to ascertain the broad functional impact of PAE-induced changes in DNA methylation patterns of the hypothalamus across early development. This analysis revealed 20 PAE-specific biological processes (both p-values and multifunctionality p-value < 0.05 in PAEvC and PAEvPF, >0.05 in PFvC; Table 1). Although these findings failed to reach significance after multiple-test correction, they were potentially reflective of the broader effects of PAE on the epigenome. Of note, the top GO terms were associated with steroid receptor signaling (GO:0042921, GO:0030518, GO:0031958, GO:0030520), a key function of the hypothalamus. Several processes associated with epigenetic regulation (GO:0016577, GO:0006482, GO:0070932) were also enriched in the PAE-specific DMRs, as were processes involved in immune function (GO:0030885, GO:0030886, GO:0002314), and cellular metabolism (GO:0050812).
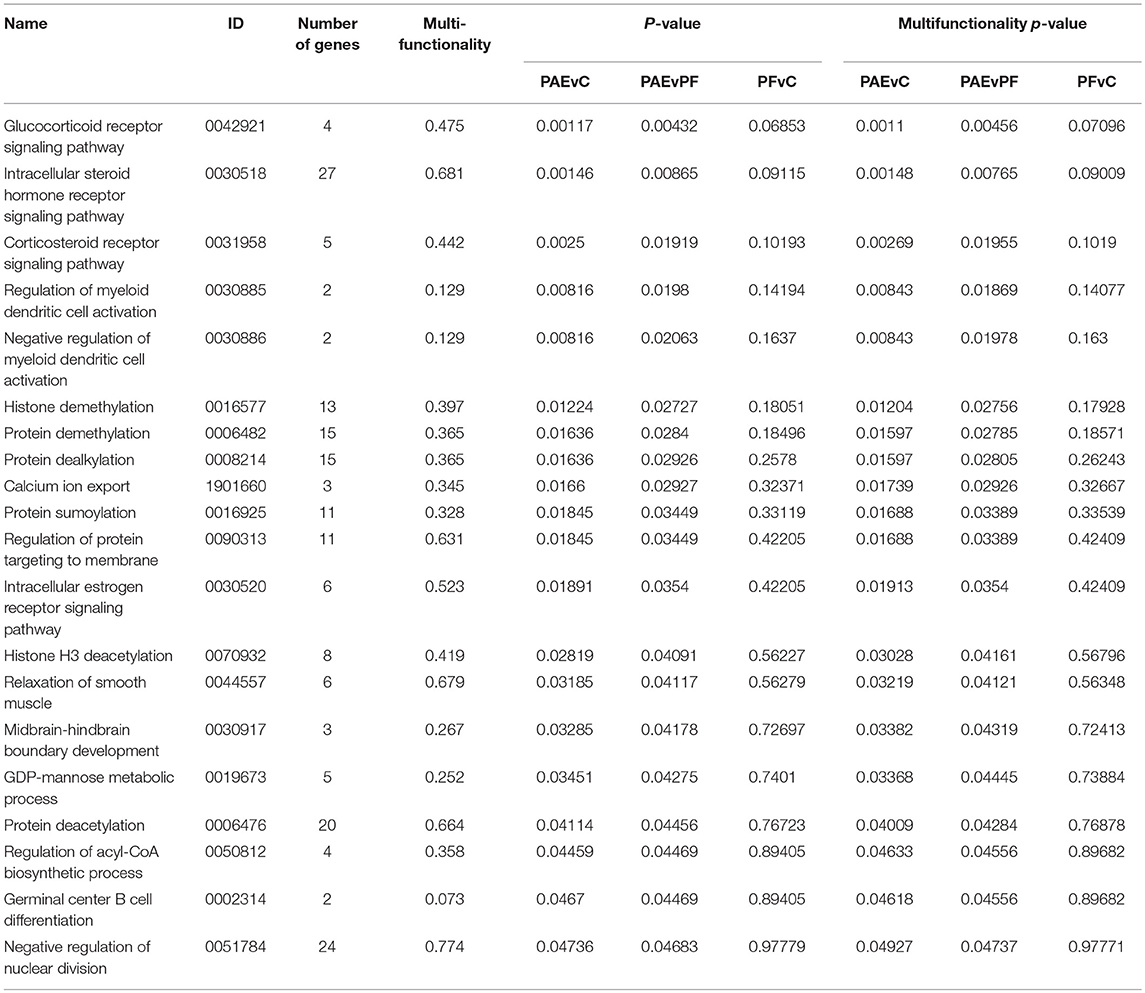
Table 1. Biological processes enriched in the developmental profile DMRs.
Given that meDIP-seq provides a relative signal of DNA methylation levels, we verified the PAE-specific DMRs using bisulfite pyrosequencing, a highly quantitative measure of DNA methylation, to test whether meDIP-seq could accurately detect alterations in DNA methylation patterns. Importantly, this technique also detects DNA hydroxymethylation, but cannot differentiate between the different cytosine modifications, while meDIP-seq is specific to DNA methylation due to the nature of the antibody used. We assessed four different DMRs, based on their potential role in the etiology of PAE-induced deficits. Of note, only a portion of CpGs within each DMR were assessed by bisulfite pyrosequencing due to limitations in read length, and as such, additional CpGs within the DMR may partially drive some of the differential DNA methylation enrichment identified by meDIP-seq.
We first assayed 16 CpGs within the 3′ UTR of the Drd4 DMR (chr1:214,281,174-214,281,640) in the same samples as the meDIP-seq analysis (Figure 4; Supplementary Table 7). DNA methylation at all CpGs assayed was significantly associated with PAE when correcting for age (p < 0.05). Moreover, this analysis detected a >5% change in DNA methylation across the entire DMR on P1 and P22 in PAE compared to C animals (p < 0.05). A 5% DNA methylation change is often used as an arbitrary quantitative cutoff for potential biological significance in epigenome-wide association studies, although we note that many studies report lower effect sizes (Mill and Heijmans, 2013; Breton et al., 2017). At older ages, several of the CpGs remained significantly different between PAE and controls, with several remaining present on P22. Overall, bisulfite pyrosequencing showed the same direction of change as the meDIP-seq analysis in this DMR and generally displayed good concordance between the two technologies (Table 2).
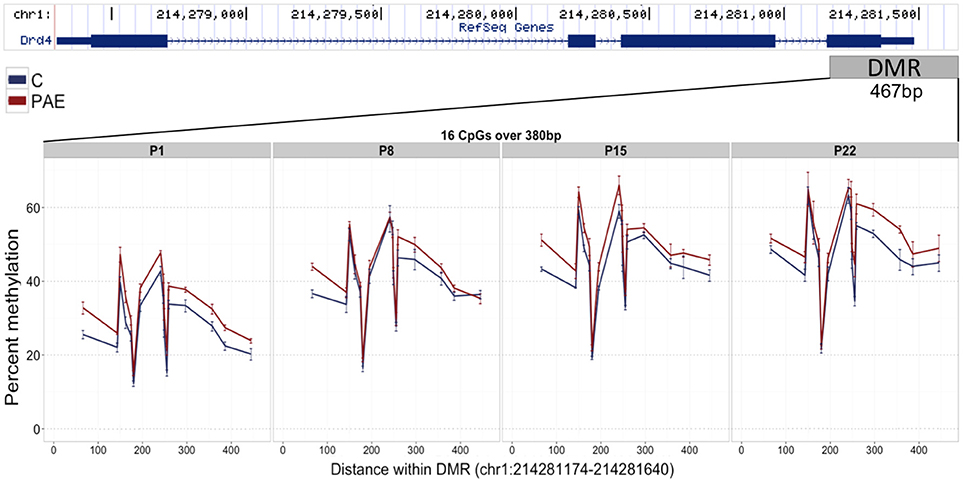
Figure 4. Bisulfite pyrosequencing verification of the Drd4 DMR. 16 CpGs (#7–22) spanning 380 base pairs (bp) of the DMR located in the 3′ UTR of Drd4 were verified by pyrosequencing in the same animals as the meDIP-seq analysis. All CpGs on P1 displayed >5% change in DNA methylation levels between PAE (red) and controls (blue). Of these, several were consistently different across all ages and a number persisted until P22. The total levels of DNA methylation in the DMR also increased with age across all groups.
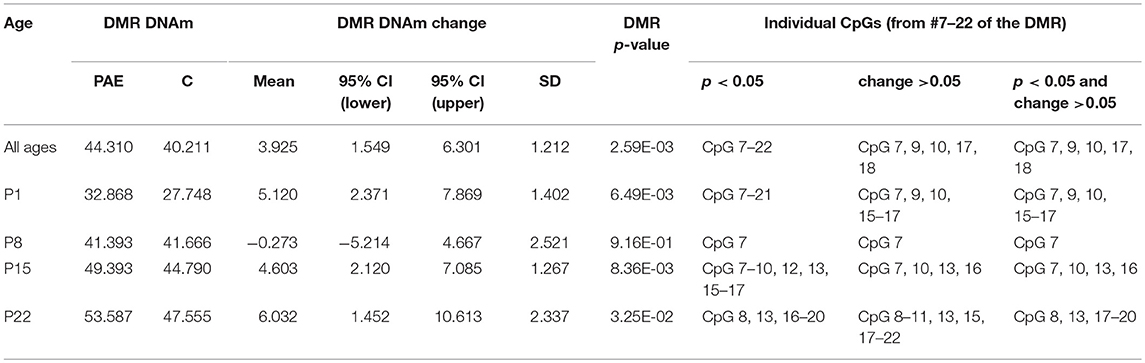
Table 2. Summary of the pyrosequencing results for the Drd4 DMR.
We also used this method to verify three additional DMRs, located within Ifih1 (chr3: 48,561,559-48,561,925), Mycbp (chr5:141,565,784-141,566,172), and Plvap (chr16: 19,912,813-19,913,185) (Supplementary Figure 6; Supplementary Table 7). These showed less consistent changes in DNA methylation between the two methods, as some ages appeared to drive DNA methylation patterns more than others and some CpGs showed opposite direction of change between meDIP-seq and pyrosequencing. For instance, the Ifih1 locus only showed significant differential DNA methylation at P15 only (p = 0.044; change = 1.21%), while the Mycbp and Plvap loci displayed a subset of significant CpGs at various ages. Although the small differences identified among groups suggest that meDIP may be sensitive enough to detect small changes in DNA methylation levels, the direction of change was not always consistent with the meDIP-seq results. These findings also raised the possibility that DNA hydromethylation differences may also be in play within these loci, since this DNA modification shows high prevalence in the brain and that could potentially explain the discrepancies between meDIP-seq and pyrosequencing (Lister et al., 2013).
In parallel to the analysis of DNA methylation in the hypothalamus during early development, we used meDIP-seq to assay DNA methylation in the hypothalamus and WBC of the same P22 females. This analysis aimed to identify tissue-concordant alterations present in both the central nervous system (CNS) and peripheral tissue in response to PAE.
As noted, cell type proportions are a major driver of epigenetic variability. However, the volume of blood collected from P22 animals was too small to perform both epigenetic and blood composition analyses on the same animals. For this reason, we collected samples from P22 animals from an independent cohort (i.e., bred at a later time, but under the same conditions as those in the present study) to determine whether PAE altered the proportions of WBCs that would be collected using the Ficoll-Paque method. Composition analysis of whole blood indicated the proportions of lymphocytes, neutrophils, monocytes, basophils, eosinophils, and large unclassified cells. Linear modeling revealed no significant differences among prenatal treatment groups, suggesting that PAE does not alter the proportion of the major WBC subtypes (Supplementary Figure 7). These findings suggest that WBC proportions likely did not influence differences in DNA methylation patterns between groups in the present dataset.
To identify tissue-concordant alterations to DNA methylation patterns associated with PAE, we performed linear modeling on the tissue-concordant dataset with a model that also accounted for differences across breeding cohorts. This method identified 299 PAE-specific DMRs at an FDR < 0.05 that were present in both tissues and showed the same direction of change in PAE vs. control animals (Figure 5; Supplementary Table 8). In contrast to the developmental profile, these DMRs showed smaller fold changes, with only 7/299 displaying a 2-fold change in PAE animals vs. controls, suggesting that subtle, but potentially important alterations are observed across tissues (Supplementary Table 9).
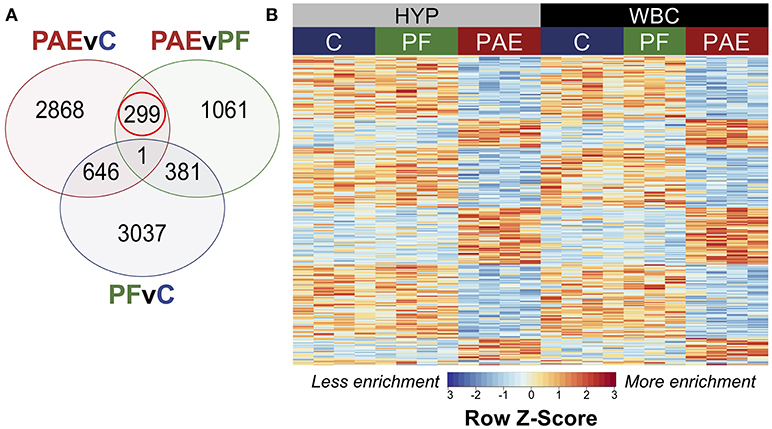
Figure 5. PAE-specific DMRs concordant across the hypothalamus and white blood cells. (A) Contrast analysis revealed 299 PAE-specific differentially methylated regions (DMR) between both tissues, which were significantly different in PAE vs. C animals and PAE vs. PF animals, but not significantly different between PF vs. C. (B) Heatmap of the DMRs. Each row represents a different DMR, while each column shows the meDIP-seq data for each animal (n = 4, except PF WBC: n = 3). Reads per kilobase per million (RPKM) data were scaled and centered to produce a Z-score for each DMR, where those in blue showed less DNA methylation enrichment and those in red showed more enrichment. PAE-specific DMRs showed the same direction of change in both tissues, with some graded effects of tissue type.
Of the significant DMRs, 105 were up-methylated and 194 were down-methylated in PAE animals, and their size ranged from 355 to 2,038 bp (median = 574 bp). The majority of DMRs also displayed small tissue-specific effects in the relative enrichment of DNA methylation, although the magnitude of change was similar between PAE and controls across both tissues (Figure 5).
Again, a majority of DMRs were located in intergenic regions, and were not associated with any gene (Figure 6A). However, the DMRs showed increased enrichment in intergenic regions compared to backgorund levels and less enrichment in intron/exon boundaries, which was driven mainly by the down-methylated regions. These results may reflect the role of DNA methylation in the regulation of splice variants, which could potentially be affected by PAE. Overall, 75 DMRs were located in genes, although the majority of these were once again located in intronic regions. Several DMRs were located in genes involved in immune function (Fgf9, Il18r1) and alcohol metabolism (Adh4). Of note, one DMR spanned 9 different isoforms of the Utg1a family of genes, which could be related to alternative splicing, while Caln1 and Cntnap5c each contained three separate DMRs.
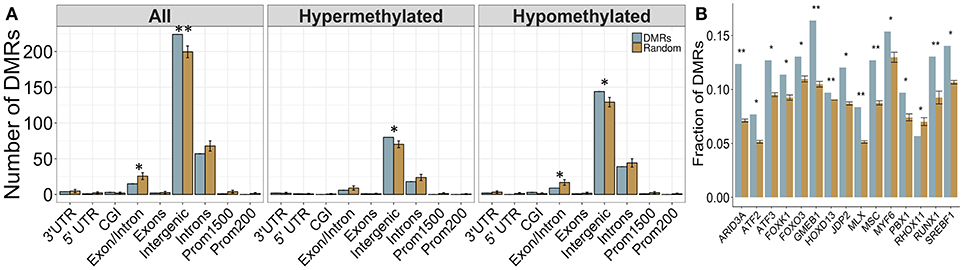
Figure 6. Enrichment patterns of the tissue-concordant DMRs. (A) Genomic feature enrichment profile of all, up-methylated, and down-methylated DMRs. The probe counts for each feature (blue) were compared to the results from permutation analyses of 299 random regions (orange), which were used to compute the p-value. While the majority of DMRs were located in intergenic regions, they showed a higher proportion than expected by random change (p < 0.01). By contrast, exon/intron boundaries were underrepresented in the DMRs, particularly within the regions that were down-methylated in PAE animals. (B) Overrepresentation analysis of transcription factor binding sites in the DMRs. Several TFBS showed higher enrichment in the tissue-concordant DMRs (blue) than expected by random chance (orange), with GMEB1 showing the highest enrichment at 16% of all DMRs. *p < 0.05, **p < 0.01.
We assessed the enrichment of TFBS within these cross-tissue PAE-specific DMRs to examine potential regulatory regions. Following multiple-test correction (FDR < 0.05), we identified 16 TFBS enriched within these regions compared to background levels (Figure 6B). The most frequent motif belonged to GMEB1, which was found in 16% of all DMRs. Several binding sites for the forkhead box (FOX) family of transcription factors were also enriched in these regions. Of note, the enrichment of MLX and SREBF1 motifs in the cross-tissue DMRs overlapped with the results from the developmental profile.
We performed GO analysis to ascertain the broad functional impact of PAE-induced changes in DNA methylation patterns across the hypothalamus and WBC. Through this analysis, we identified 35 PAE-specific biological processes (both p-values and multifunctionality p-value < 0.05 in PAEvC and PAEvPF, >0.05 in PFvC; Table 3). However, with multiple-test correction, these findings failed to reach significance, suggesting that these may represent more subtle effects on the global epigenome. Of note, the top GO terms were associated with metabolic processes, including aldehyde metabolism (GO:0006081). Several processes were also associated with immune function (GO:0045063, GO:0071351, GO:0032733, GO:0070673, GO:2674), chromatin remodeling (GO:6338, GO:90239), and the stress response (GO:42320).
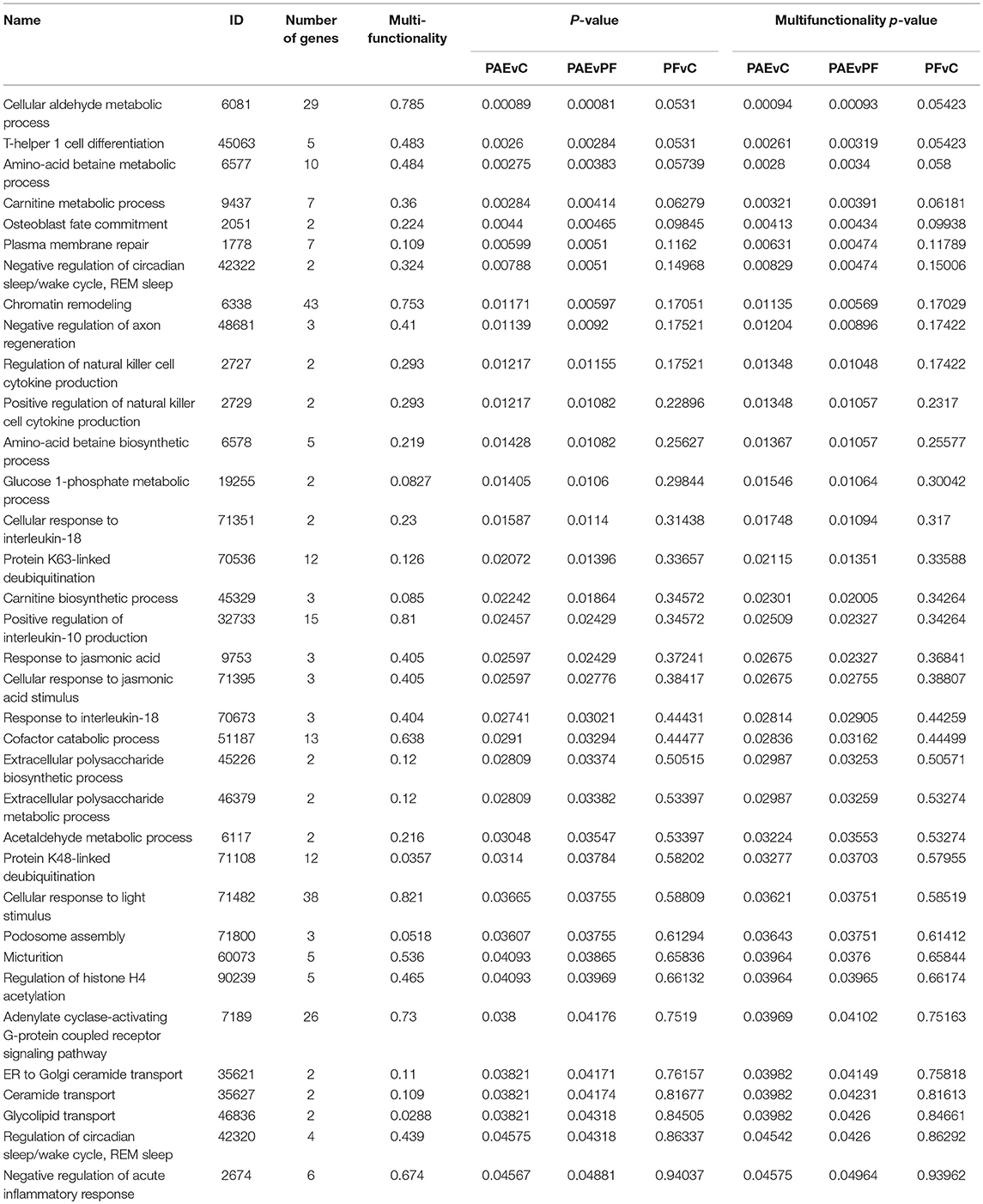
Table 3. Biological processes enriched in the tissue-concordant DMRs.
We used bisulfite pyrosequencing to compare quantitative levels of DNA methylation between PAE and Control animals in three cross-tissue DMRs. More specifically, we analyzed DNA methylation in the final exon and 3′ UTR of Adh4 (chr2:243,719,416-243,720,233), the first exon and 5′ UTR of Ctnnbip1 (chr5:166,485,057-166,485,637), and the first intron of Ffg9 (chr15:38,377,629-38,378,027; Supplementary Figure 8; Supplementary Table 10). The main differences in DNA methylation levels were identified between tissues, which sometimes showed different directions of change between PAE and controls. In particular, a single CpG within the Adh4 DMR showed ~5% methylation difference in the hypothalamus of PAE animals (p = 0.011; change = 5.94%), but this effect was not present in WBC or both tissues combined and was in the opposite direction of the meDIP-seq results. Another CpG within the Adh4 locus showed small, but not statistically significant, changes that were consistent between tissues. This pattern was also observed in the Fgf9 locus, which suggests subtle but potential systemic effects of PAE. Furthermore, DNA methylation in the Fgf9 DMR was significantly associated with PAE in the hypothalamus (p = 0.031; change = 0.94%), but not both tissues combined or WBC alone. By contrast, the Ctnnbip1 locus showed opposite, but non-significant, effects between tissues (decreased in the hypothalamus; increased in WBC), with only CpG showing significant differential methylation in WBC alone (p = 0.036; change = 5.50%) suggesting that other factors may come into play, such as DNA hydroxymethylation or genetic influences. Moreover, as we did not assess quantitative DNA methylation level across the entire DMR due to pyrosequencing limitations, other CpGs may drive the enrichment patterns previously identified by meDIP-seq.
Alcohol exposure in utero has been shown to reprogram physiological and neurobiological systems, increasing the risk of adverse developmental outcomes across the lifespan (Zhang et al., 2005; Mattson et al., 2011; Pei et al., 2011). Given the potential role of epigenetic mechanisms in mediating the long-term effects of PAE, the present study aimed to extend previous work on the influence of in utero alcohol exposure on the epigenome, using an animal model of PAE to assess genome-wide DNA methylation patterns in the hypothalamus and WBCs during early postnatal development in females, a group largely underrepresented in neurobiological, molecular and genetic studies. We identified 118 differentially methylated regions (DMRs) that were altered in the hypothalamus of PAE vs. control animals across the pre-weaning period. In parallel, we found 299 DMRs displaying concordant DNA methylation alterations between the hypothalamus and WBCs of PAE animals at weaning. Several differentially methylated genes were functionally related to PAE-induced deficits, having roles in the immune response, neurobiological function, and mental health. Additionally, functional enrichment revealed several PAE-specific biological processes, including those related to immune function, the stress response, and epigenetic regulation. In addition, we identified several transcription factor binding sites (TFBS) that were enriched in the DMRs, which may potentially reflect broader programming effects of PAE on the epigenome. Overall, this study is among the first to compare central and peripheral effects of PAE on DNA methylation patterns in addition to characterizing genome-wide changes in females prenatally exposed to alcohol. Our findings suggest that PAE results in widespread alterations to epigenomic programs in both the CNS and peripheral tissues, with a potential impact on both neurobiological and physiological systems. In addition, we demonstrated that peripheral changes in DNA methylation profiles could serve as a potential biomarker of PAE's effects on the CNS. These findings can also provide insight into other neurodevelopmental and mental health disorders, such as autism, ADHD, depression, and many more, as they share numerous outcomes and comorbidities.
Our initial analysis of the DMRs revealed several differentially methylated genes that could be relevant to PAE-induced deficits. In particular, the dopamine receptor D4 (Drd4) gene contained a DMR that persisted across the early developmental period. Given its crucial role in dopaminergic function, as well as interactions among dopaminergic, neuroendocrine, and immune systems, alterations to this gene could reflect broader alterations to signaling in the brain. Interestingly, differential DNA methylation patterns of Drd4 are also present in the buccal epithelial cells of individuals with FASD, suggesting that this may constitute an association of PAE with the epigenome that replicates across organisms (Fransquet et al., 2016; Portales-Casamar et al., 2016; Lussier et al., 2018). Although these were not identified in central tissues, buccal epithelial cells may act as a passable surrogate tissue for brain in human DNA methylation studies, as they originate from the same germ layer (Lowe et al., 2013; Portales-Casamar et al., 2016). Importantly, in addition to this association with FASD, genetic and epigenetic variation in Drd4 has been linked to attention deficit hyperactivity disorder (ADHD), schizophrenia, bipolar disorder, substance-use disorders, and several other neurobiological disorders (Bau et al., 2001; Chen et al., 2011; Ptácek et al., 2011; Docherty et al., 2012; Kordi-Tamandani et al., 2013; Zhang et al., 2013; Cheng et al., 2014; Faraone et al., 2014; Dadds et al., 2016; Ji et al., 2016). Golga4 also contained two PAE-specific DMRs across hypothalamic development, and is overexpressed in the prefrontal cortex of individuals with bipolar disorder (Iwamoto et al., 2004). As a member of the Golgi secretory pathway, it could also potentially influence the secretion of neuropeptides by cells of the hypothalamus, possibly playing a role in altered function or responsiveness following PAE (Wong and Munro, 2014). Similarly, increased Plvap expression increases the breakdown and permeability of the blood-brain barrier (BBB; Shue et al., 2008). Given that ethanol increases the permeability of the BBB in adult mice, slight alterations in DNA methylation of Plvap could reflect broader effects on the BBB, which could, in turn, could affect downstream neurobiological functions (Alfonso-Loeches et al., 2016).
The tissue-concordant DMRs also contained several genes previously associated with mental health disorders. Although the same temporal stability could not be assessed here, as both tissues originated from the same age, these findings may point to more systemic effects of PAE on the developing organism. In particular, Adh4 was differentially methylated across the hypothalamus and WBCs of PAE animals, and has been previously associated with alcohol dependence and substance abuse (Luo et al., 2005). Importantly, it is a key component of alcohol metabolism pathways, and could reflect increased susceptibility to the effects of alcohol during development. Furthermore, Caln1 contained 3 separate DMRs; as it contains a risk allele for schizophrenia in some human populations, it could also play a role in some of the neuropsychiatric deficits observed in individuals with FASD (Li et al., 2015).
Of note, two genes displayed differential DNA methylation patterns in both the developmental profile and tissue-concordance analysis, Cntnap5c and Ush2a, which may reflect persistent alterations to DNA methylation patterns across both age and tissue types. In humans, genetic variation in Cntnap5 is associated with risk for Alzheimer's disease and bipolar disorder, while its deletion is associated with autism and dyslexia, suggesting the possibility that common pathways may come into play among different neurobiological disorders (Pagnamenta et al., 2010; Xu et al., 2014; Schott et al., 2016). By contrast, mutations in Ush2a cause Usher syndrome II, which is associated with hearing deficiencies, deficits also commonly found in individuals with FASD (Church and Gerkin, 1988). Importantly, our animal model is based on an outbred population of Sprague-Dawley rats, which display fairly broad genetic diversity. Although genetic background can also influence DNA methylation patterns throughout the genome, the greater variability both within and between our treatment groups may reduce the likelihood of genetic variation having a major impact on our results and suggests that our results may represent more robust associations between PAE and DNA methylation patterns (Fraser et al., 2012; Heyn et al., 2013; Moen et al., 2013).
Finally, several DMRs in both datasets were located in genes associated with immune function and response. In particular, Ifih1 was identified across all ages in the hypothalamus; as a receptor for double stranded RNA that responds to viral infections, it could be associated with vulnerability to immunological deficits (Rice et al., 2014). In addition, Fgf9, a key factor in embryonic and glial cell development, was differentially methylated in both the hypothalamus and WBCs (Thisse and Thisse, 2005). This growth factor promotes pro-inflammatory environments through Ccl2 and Ccl7 chemokine secretion, consistent with several DMRs that were located in genes associated with pro-inflammatory cytokine and chemokine signaling (Lindner et al., 2015). These included Il20ra and Ccrl2 in the developmental profile, and Il18r1 in the tissue-concordance analysis, suggesting that PAE could influence inflammatory pathways through epigenetic mechanisms, and ultimately, potentially alter brain development and the neuroimmune response.
We also assessed the functional enrichment of genes located within PAE-specific DMRs, identifying a number of biological processes associated with differential DNA methylation patterns in PAE animals compared to controls. In the DMRs identified across hypothalamic development, a large number of GO processes were associated with functions related to steroid receptor signaling. The hypothalamus is central to numerous physiological systems that function through steroid hormones, many of which are dysregulated by PAE (Weinberg et al., 2008). As such, this enrichment pattern suggests that DNA methylation may play a role in the reprogramming of hormonal systems during early development, potentially priming physiological systems to new set-points. In addition, several processes in both the developmental and tissue-concordant DMRs were associated with histone modifications, which may reflect the complex interplay between different layers of the epigenetic machinery. Several studies have identified alterations to histone modifications in the brain following developmental alcohol exposure, further highlighting their potential role in FASD (Guo et al., 2011; Govorko et al., 2012; Bekdash et al., 2013; Subbanna et al., 2013, 2014; Goldowitz et al., 2014; Veazey et al., 2015; Zhang et al., 2015; Chater-Diehl et al., 2016; Lussier et al., 2017). A large number of immune-related biological processes were also identified through this analysis, which is particularly relevant to individuals with FASD, who may be more susceptible to infections and immune deficits, a phenotype recapitulated across multiple animal models of PAE, and with some preliminary evidence in humans (Bodnar and Weinberg, 2013; Bodnar et al., 2016). Given the role of the hypothalamus in modulating the immune response, altered epigenetic programs could potentially contribute to altering baseline function and/or responsiveness to immune challenge of the hypothalamus, limiting the organism's ability to defend against disease or infection. In addition, the top GO term associated with PAE in the tissue-concordant DMRs was “cellular aldehyde metabolic process,” which may reflect lasting effects of PAE on the organism's ability to metabolize and tolerate alcohol's metabolic byproducts and possibly modulate susceptibility to substance abuse later in life. While no overlaps were identified between the specific biological processes identified in the developmental profile and tissue-concordance analyses, both contained a high proportion of processes associated with immune, endocrine, or epigenetic functions. These findings suggest that PAE may cause systemic effects on the epigenome across multiple tissue types, which may, in turn, influence downstream neurobiological and physiological processes.
Previous studies have identified subtle effects of PAE on gene expression programs and epigenomic patterns, which is consistent with the effects of other prenatal exposures (Rakyan et al., 2011; Zhou et al., 2011; Ladd-Acosta et al., 2013; Laufer et al., 2013; Berko et al., 2014; Lussier et al., 2015; Chater-Diehl et al., 2016). Regions containing lower CpG density appear to be more responsive to environmental exposures, highlighting the importance of selecting a method that covers a large portion of the epigenome when analyzing environmental exposures (Irizarry et al., 2009). We analyzed genome-wide DNA methylation using meDIP-seq, which reduces the complexity of the dataset by omitting unmethylated regions while interrogating a majority of genome (Harris et al., 2010). However, one limitation of meDIP-seq is that it is highly dependent on DNA methylation levels, local CpG density, and CpG position, which can introduce biases in the data (Pelizzola et al., 2008; Robinson et al., 2010b; Lentini et al., 2018). However, by performing pairwise comparisons between treatment and control groups, such biases are significantly reduced or eliminated (Harris et al., 2010). The enrichment patterns observed across our datasets are consistent with the fact that the majority of the mammalian genome is CpG-depleted and, with the exception of active regulatory regions, the remaining CpGs are methylated. By contrast, the vast majority of CpG-rich regions (i.e., promoters, CpG islands) are unmethylated and are less likely to bind to the 5-methylcytosine antibody, leading to depletion in meDIP-seq data, as observed in our own datasets. Consistent with the fact that promoter associated CpG islands are largely insensitive to environmental stimuli, few DMRs across our analyses were identified in promoters and CpG islands. Indeed, the majority of DMRs were located in intergenic regions and introns. Although these findings will require additional validation, these intergenic regions are critical to further explore, as they are potentially more responsive to the effects of PAE and may contain important regulatory regions not yet annotated in the rat genome. In addition, several DMRs were located in intron/exon boundaries. Given that DNA methylation plays a role in regulating alternative splice variants, these findings may reflect alterations to the balance of different isoforms within the cell, which could influence downstream cellular profiles and phenotypes (Maunakea et al., 2010, 2013; Shukla et al., 2011). Although isoform balance has not been investigated in the context of PAE, alcohol consumption can influence the proportions of different splice variants in the brain, supporting a potential role in early-life exposures as well (MacKay et al., 2011; Lee et al., 2014; Farris et al., 2015; Mathew et al., 2016). Interestingly, a larger proportion of down-methylated DMRs were identified in both analyses, which is consistent with several studies showing that PAE decreases bulk DNA methylation levels (Otero et al., 2012; Chen et al., 2013; Mukhopadhyay et al., 2013; Perkins et al., 2013; Liyanage et al., 2015; Nagre et al., 2015). These findings provide important insight into the various outcomes from different paradigms of alcohol exposure and suggest that similar upstream mechanisms may impact DNA methylation across models. In particular, alcohol is known to impact one-carbon metabolism through multiple pathways, including inhibition of folate-dependent methylation pathways and AdoMet synthesis (Medici and Halsted, 2013; Ngai et al., 2015). Furthermore, PAE can influence both the expression and activity of DNA methyltransferases, which are essential for the establishment and maintenance of DNA methylation profiles (reviewed in Lussier et al., 2017). These well-known mechanisms of alcohol-induced DNA methylation alterations likely contribute to the differential DNA methylation patterns observed across models and organisms, with specific effects potentially dictated by the timing and dosage of alcohol exposure, or other environmental factors (Pollard, 2007; Rogic et al., 2016; Lussier et al., 2017). For example, our animal model uses a constant level of alcohol exposure, resulting in blood alcohol levels of 80–150 mg/dl across the equivalent of the first two trimesters of human pregnancy. With this moderate to moderately high level of exposure, equivalent to 1–2 times the legal driving limit in most jurisdictions, we do not observe dysmorphologies or other severe growth and functional deficits. However, this paradigm does cause a broad range of behavioral and functional abnormalities, which are typically present across the entire spectrum of FASD. As such, the differential DNA methylation profiles observed in the present study potentially reflect a broader portion of the FASD spectrum in humans, and likely point to some of the common pathways affected by PAE across different exposure patterns.
The large proportion of DMRs located in intergenic regions suggests that these could contain regulatory regions susceptible to the influence of PAE. Given that the rat genome is poorly annotated for regulatory features, we assessed the enrichment profiles of different transcription factor binding sites in the DMRs, which could be influenced by DNA methylation levels within specific loci. While only the binding site for the BHLHE40 transcription factor was significantly enriched in PAE-specific DMRs across early development, we previously identified this gene as differentially expressed in the brain of PAE adult animals (Lussier et al., 2015). This gene negatively regulates the circadian rhythm, a key function of the hypothalamus that is dysregulated in individuals with FASD (Nakashima et al., 2008). The BHLHE40 transcription factor could potentially play a role in early programming effects of PAE on neurobiological systems, with persistent expression and downstream effect into later life. By contrast, the tissue-concordant DMRs contained a high proportion of significantly enriched TFBS, including SREBF1, which trended toward significance in the developmental profile DMRs. SREBF1 is associated with key metabolic processes for hormonal signaling, as it plays a role in the regulation of cholesterol production (Osborne, 2001). It is also associated with Smith–Magenis syndrome, which is characterized by intellectual disability, disordered sleep, and behavioral problems (Smith et al., 2002). Furthermore, additional TFBS enriched in the tissue-concordant dataset included two members of the forkhead box family of genes, including FOXK1 and FOXO3. In particular, FOXO3 was identified as a hub gene in the brain PAE animals following an immune challenge, suggesting that it may prime biological systems from early in life (Lussier et al., 2015). Finally, the highest represented TFBS in the tissue-concordant dataset was GMEB1, which is involved in signal transduction of the glucocorticoid response (Zeng et al., 2000). Importantly, recent evidence suggests that DNA methylation patterns can influence transcription factor occupancy, modulating the use of enhancer elements and gene expression levels (Maurano et al., 2015; Yin et al., 2017). Taken together, these findings suggest that the DMRs identified in both the developmental and tissue-concordant analysis may contain key regulatory regions, and that various transcription factors likely act in concert with DNA methylation to mediate the effects of PAE on physiological functions.
Although meDIP-seq allows for the investigation of more variable regions of the epigenome, it presents a particular caveat when assessing DNA methylation levels, as it provides relative levels of DNA methylation across broad regions of the genome, rather than quantitative and granular data. While a relative method was reasonable for the purpose of our study, which was to identify differences between animals exposed to alcohol and controls, we also attempted to verify our findings from the meDIP-seq analysis through bisulfite pyrosequencing, the gold standard for targeted DNA methylation analyses. However, the concordance between the two methods was not as strong as expected, with only the Drd4 locus showing significant changes across the entire DMR. By contrast, other verified regions each showed a subset of CpGs that could be driving the associations. This discordance may potentially be due to a number of factors, including DNA hydroxymethylation, a bias of meDIP toward regions of high DNA methylation and CpG density, or sample size. A major limitation of the standard bisulfite pyrosequencing methods used here as a verification method for meDIP-seq is that it detects both methylated and hydroxymethylated cytosines, and there is no way to distinguish the two modifications using solely bisulfite conversion, resulting in a mixed signal. Although oxidative bisulfite conversion can distinguish DNA methylation and hydroxymethylation when used in parallel with bisulfite conversion, we could not perform this analysis on our samples due to DNA input limitations (Booth et al., 2012). By contrast, meDIP-seq specifically enriches DNA methylation, as the antibody is highly specific to 5-methylcytosine (Taiwo et al., 2012). Given that neuronal cells contain a high proportion of DNA hydroxymethylation compared to other cell types, it is possible that the observed differences in outcomes from the two methodologies are due to the confound of additional epigenetic patterns not assessed in the meDIP-seq analysis (Wang et al., 2012; Lister et al., 2013). Indeed, a number of studies have shown that developmental alcohol exposure can alter DNA hydroxymethylation programs in neuronal cells, suggesting that it may also play a role in the etiology of FASD and may have biased our verification analyses (Chen et al., 2013; Öztürk et al., 2017). In addition, the lack of verification could potentially be due to the relatively low number of animals used in the present study, as well as increased variability in the enrichment profiles obtained from meDIP-seq, given the broader regions assessed. Nevertheless, the Drd4 locus identified in the developmental profile of the hypothalamus displayed consistent DNA methylation alterations in both methods, suggesting that meDIP-seq can indeed capture differences in DNA methylation patterns, regardless of the influence of DNA hydroxymethylation, and also highlighting the importance of the dopaminergic system in PAE-induced alterations. Additional studies are required to fully validate these findings and assess their relationship to the deficits observed following PAE. Given the high prevalence of DNA hydromethylation in the brain, future studies should endeavor to tease out the role of DNA methylation and DNA hydroxymethylation in the context of PAE, using methods that can quantitatively parse the two modifications (Wang et al., 2012; Lister et al., 2013).
A critical strength of animal models derives from their ability to directly compare central and peripheral tissues to ascertain potential correlations between the two, which may identify potential biomarkers reflective of brain function in a tissue that is available for study in human populations. Although several studies have assessed the concordance of DNA methylation patterns between different tissues in humans, the correlation between tissues tends to be rather low, highlighting the importance of analyzing both tissues in parallel (Farré et al., 2015; Edgar et al., 2017). Other than tissue identity, cell type heterogeneity within a tissue is a major driver of DNA methylation patterns. Therefore, we attempted to correct, at least partially, for cellular heterogeneity among groups by removing regions that were associated with the gene expression patterns of major cell types in the brain—neurons, astrocytes, and oligodendrocytes (Cahoy et al., 2008). However, we were limited by the use of regions located within genes, as well as the indirect usage of transcriptomes, and thus could not correct for important regulatory regions that may be associated with cell type or be poised for activation, such as intergenic enhancers or insulators. Furthermore, additional cellular subtypes, such as glia, are also present in the hypothalamus and could have influenced the results obtained through differences in cell-type proportions. Although we did not detect any significant PAE-specific differences in the DNA methylation levels of cell-type associated peaks, it is possible that the regions not captured by this peak-based filter could drive the differences in neuronal and glial function previously observed following PAE (Wilhelm and Guizzetti, 2015; Noor and Milligan, 2018). As several protocols now exist to isolate cell type-specific nuclei from both frozen and fresh tissue samples, future studies should attempt to replicate these findings in specific cellular subtypes to tease apart the role of DNA methylation in PAE-induced deficits (Habib et al., 2016; Milani et al., 2016).
By contrast, we measured the proportion of different WBC subtypes in an independent cohort of animals. The fact that we did not identify any significant differences in WBC composition of whole blood among groups suggests that blood composition does not drive the DMRs identified in the tissue-concordant analysis. There is also a small possibility that, as Ficoll-Paque is a highly technical procedure, differences between WBC extractions could have influenced the proportions of cells analyzed in the present study. Additionally, it is possible that group differences might be uncovered if cell subtypes are further subdivided through more sophisticated methods such as fluorescence-activated cell sorting. In contrast to human studies of DNA methylation, no bioinformatic tools exist to predict the proportion of different cell types using epigenomic profiles in rats, and future studies should take this into consideration. Nevertheless, we successfully identified several PAE-specific DMRs that showed the same direction of change between the two tissues, suggesting that these regions may be responsive to ethanol across multiple tissues and may represent more stable biomarkers of PAE.
Finally, female animals were the main focus of this epigenome-wide study, partially due to the broad sexual dimorphisms observed across studies of PAE and FASD (Lee and Rivier, 1996; Weinberg et al., 2008), as well as the widespread underrepresentation of females in neurobiological, molecular, and genome-wide studies of FASD (Lussier et al., 2017). However, as in the majority of studies utilizing only male subjects, this approach presents an important caveat in the interpretation of our results, as males and females often show sexually dimorphic responses to stress, disease, and other environmental factors, displaying differences in behavioral patterns, HPA axis function/activation, immune system activity, neurogenesis, and other physiological and cellular functions (Oldehinkel and Bouma, 2011; Bale and Epperson, 2015; Panzica and Melcangi, 2016; Yagi and Galea, 2018). In particular, the effects of PAE on the developing organism show marked sex-specific differences spanning cognitive and behavioral phenotypes, as well as in the differential susceptibilities to stressors and mental health disorders across the lifespan (Hellemans et al., 2008; Weinberg et al., 2008). Given that genetic and epigenetic patterns are highly associated with sex, research on females is vital to our understanding of the biological mechanisms underlying PAE (Zhang et al., 2011). Nevertheless, our findings should also be validated in male animals to fully assess the effects of PAE on the transcriptome and DNA methylome and elucidate the sexually dimorphic effects that may exist.
Our results support and significantly extend previous studies indicating a role for DNA methylation in the early-life reprogramming of hypothalamic functions by PAE, and suggest that DNA methylation patterns in WBC could potentially be used as a surrogate for alterations in the central nervous system. We identified persistent PAE-induced alterations to the DNA methylome of the hypothalamus, including several DMRs that could, at least in part, underlie some of the deficits observed in FASD. Although PAE-induced alterations to DNA methylation profiles at any of these development ages may not persist into adulthood, changes early in development could alter developmental trajectories and induce lasting alterations in brain structure, connectivity, and function, and/or prime physiological systems to different set-points. Of note, we demonstrate for the first time that PAE-specific DMRs can occur concordantly across central and peripheral tissues, which potentially represent systemic effects of PAE on the epigenome and could serve as an epigenetic biomarker or signature of FASD. Taken together, these findings provide insight into the important role of epigenetic alterations in the short and long-term deficits observed in FASD and provide a foundation for the development of robust biomarkers of PAE.
The datasets generated for this study can be found in the Gene Expression Omnibus (GEO) database (GSE121582).
All animal protocols were approved by the University of British Columbia Animal Care Committee and are consistent with the NIH Guide for the Care and Use of Laboratory Animals (certificates: A06-0017, A07-0381, A10-0136, A10-0016, A12-0032).
AL designed and performed all experiments and wrote the manuscript. TB helped with experimental design, data collection, and interpretations of results. MM helped with the meDIP-seq experiments and analyses. AM performed part of the pyrosequencing analyses. MH helped with design and analysis of meDIP-seq data. MK and JW contributed to all stages, including design, analysis, interpretation, and writing.
This research is supported by grants from the US National Institutes of Health/National Institute on Alcohol Abuse and Alcoholism (R37 AA007789 and RO1 AA022460); the Kid's Brain Health Network (formerly NeuroDevNet); and the Canadian Foundation for Fetal Alcohol Research. AL was supported by a Developmental Neurosciences Research Training award from Brain Canada & the Kid's Brain Health Network. MK is the Canada Research Chair in Social Epigenetics.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the Serrano lab (UBC) for allowing us to use their Advia120 hematology system. We also thank members of the Weinberg and Kobor labs for their assistance with this project.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2018.00610/full#supplementary-material
Alfonso-Loeches, S., Ureña-Peralta, J., Morillo-Bargues, M. J., Gómez-Pinedo, U., and Guerri, C. (2016). Ethanol-induced TLR4/NLRP3 neuroinflammatory response in microglial cells promotes leukocyte infiltration across the BBB. Neurochem. Res. 41, 193–209. doi: 10.1007/s11064-015-1760-5
Bale, T. L., and Epperson, C. N. (2015). Sex differences and stress across the lifespan. Nat. Neurosci. 18, 1413–1420. doi: 10.1038/nn.4112
Ballouz, S., Pavlidis, P., and Gillis, J. (2017). Using predictive specificity to determine when gene set analysis is biologically meaningful. Nucleic Acids Res. 45:e20e20. doi: 10.1093/nar/gkw957
Bau, C. H., Almeida, S., Costa, F. T., Garcia, C. E., Elias, E. P., Ponso, A. C., et al. (2001). DRD4 and DAT1 as modifying genes in alcoholism: interaction with novelty seeking on level of alcohol consumption. Mol. Psychiatry 6, 7–9. doi: 10.1038/sj.mp.4000819
Bekdash, R. A., Zhang, C., and Sarkar, D. K. (2013). Gestational choline supplementation normalized fetal alcohol-induced alterations in histone modifications, DNA methylation, and proopiomelanocortin (POMC) gene expression in beta-endorphin-producing POMC neurons of the hypothalamus. Alcohol. Clin. Exp. Res. 37, 1133–1142. doi: 10.1111/acer.12082
Berko, E. R., Suzuki, M., Beren, F., Lemetre, C., Alaimo, C. M., Calder, R. B., et al. (2014). Mosaic epigenetic dysregulation of ectodermal cells in autism spectrum disorder. PLoS Genet. 10:e1004402. doi: 10.1371/journal.pgen.1004402
Bodnar, T. S., Hill, L. A., and Weinberg, J. (2016). Evidence for an immune signature of prenatal alcohol exposure in female rats. Brain. Behav. Immun. 58, 130–141. doi: 10.1016/j.bbi.2016.05.022
Bodnar, T. S., and Weinberg, J. (2013). “Prenatal alcohol exposure: impact on neuroendocrine–neuroimmune networks BT,” in Neural-Immune Interactions in Brain Function and Alcohol Related Disorders, eds C. Cui, L. Grandison, and A. Noronha (Boston, MA: Springer US), 307–357. doi: 10.1007/978-1-4614-4729-0_10
Booth, M. J., Branco, M. R., Ficz, G., Oxley, D., Krueger, F., and Reik, W. (2012). Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution. Science 80:336. doi: 10.1126/science.1220671
Breton, C. V., Marsit, C. J., Faustman, E., Nadeau, K., Goodrich, J. M., Dolinoy, D. C., et al. (2017). Small-magnitude effect sizes in epigenetic end points are important in children's environmental health studies: the children's environmental health and disease prevention research center's epigenetics working group. Environ. Health Perspect. 125, 511–526. doi: 10.1289/EHP595
Cahoy, J. D., Emery, B., Kaushal, A., Foo, L. C., Zamanian, J. L., Christopherson, K. S., et al. (2008). A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J. Neurosci. 28, 264–278. doi: 10.1523/JNEUROSCI.4178-07.2008
Card, J. P., and Swanson, L. W. (2012). “The hypothalamus: an overview of regulatory systems,” in Fundamental Neuroscience, 4th Edn. 717–727. doi: 10.1016/B978-0-12-385870-2.00033-0
Chater-Diehl, E. J., Laufer, B. I., Castellani, C. A., Alberry, B. L., and Singh, S. M. (2016). Alteration of gene expression, DNA methylation, and histone methylation in free radical scavenging networks in adult mouse hippocampus following fetal alcohol exposure. PLoS ONE 11:e0154836. doi: 10.1371/journal.pone.0154836
Chen, D., Liu, F., Shang, Q., Song, X., Miao, X., and Wang, Z. (2011). Association between polymorphisms of DRD2 and DRD4 and opioid dependence: Evidence from the current studies. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 156, 661–670. doi: 10.1002/ajmg.b.31208
Chen, M., Olson, H., Picciano, J., Starr, J., and Owens, J. (2012). Sleep problems in children with fetal alcohol spectrum disorders. J. Clin. Sleep Med. 8, 421–429. doi: 10.5664/jcsm.2038
Chen, Y., Ozturk, N. C., and Zhou, F. C. (2013). DNA methylation program in developing hippocampus and its alteration by alcohol. PLoS ONE 8, 1–11. doi: 10.1371/journal.pone.0060503
Cheng, J., Wang, Y., Zhou, K., Wang, L., Li, J., Zhuang, Q., et al. (2014). Male-specific association between dopamine receptor D4 gene methylation and schizophrenia. PLoS ONE 9:e89128. doi: 10.1371/journal.pone.0089128
Church, M. W., and Gerkin, K. P. (1988). Hearing disorders in children with fetal alcohol syndrome: findings from case reports. Pediatrics 82, 147–154.
Dadds, M. R., Schollar-Root, O., Lenroot, R., Moul, C., and Hawes, D. J. (2016). Epigenetic regulation of the DRD4 gene and dimensions of attention-deficit/hyperactivity disorder in children. Eur. Child Adolesc. Psychiatry 25, 1081–1089. doi: 10.1007/s00787-016-0828-3
Dobbing, J., and Sands, J. (1979). Comparative aspects of the brain growth spurt. Early Hum. Dev. 3, 79–83.
Docherty, S. J., Davis, O. S. P., Haworth, C. M. A., Plomin, R., D'Souza, U., and Mill, J. (2012). A genetic association study of DNA methylation levels in the DRD4 gene region finds associations with nearby SNPs. Behav. Brain Funct. 8:31. doi: 10.1186/1744-9081-8-31
Earnest, D. J., Chen, W. J., and West, J. R. (2001). Developmental alcohol and circadian clock function. Alcohol Res. Health 25, 136–140. Available online at: https://pubs.niaaa.nih.gov/publications/arh25-2/136-140.htm
Edgar, R. D., Jones, M. J., Meaney, M. J., Turecki, G., and Kobor, M. S. (2017). BECon: a tool for interpreting DNA methylation findings from blood in the context of brain. bioRxiv[Preprint]. doi: 10.1101/111609
Eguchi, Y. (1969). “Interrelationships between the foetal and maternal hypophysial adrenal axes in rats and mice,” in Physiology and Pathology of Adaptation Mechanisms, ed E. Bajusz (New York, NY: Perganon Press), 3–27.
Faraone, S. V., Bonvicini, C., and Scassellati, C. (2014). Biomarkers in the diagnosis of ADHD – promising directions. Curr. Psychiatry Rep. 16:497. doi: 10.1007/s11920-014-0497-1
Farré, P., Jones, M. J., Meaney, M. J., Emberly, E., Turecki, G., and Kobor, M. S. (2015). Concordant and discordant DNA methylation signatures of aging in human blood and brain. Epigenet. Chromatin 8:19. doi: 10.1186/s13072-015-0011-y
Farris, S. P., Arasappan, D., Hunicke-Smith, S., Harris, R. A., and Mayfield, R. D. (2015). Transcriptome organization for chronic alcohol abuse in human brain. Mol. Psychiatry 20, 1438–1447. doi: 10.1038/mp.2014.159
Fransquet, P. D., Hutchinson, D., Olsson, C. A., Wilson, J., Allsop, S., Najman, J., et al. (2016). Perinatal maternal alcohol consumption and methylation of the dopamine receptor DRD4 in the offspring: the Triple B study. Environ. Epigenetics 2:dvw023-dvw023. doi: 10.1093/eep/dvw023
Fraser, H. B., Lam, L. L., Neumann, S. M., and Kobor, M. S. (2012). Population-specificity of human DNA methylation. Genome Biol. 13:R8. doi: 10.1186/gb-2012-13-2-r8
Gillis, J., Mistry, M., and Pavlidis, P. (2010). Gene function analysis in complex data sets using ErmineJ. Nat. Protoc. 5, 1148–1159. doi: 10.1038/nprot.2010.78
Gillis, J., and Pavlidis, P. (2011). The impact of multifunctional genes on “guilt by association” analysis. PLoS ONE 6:e17258. doi: 10.1371/journal.pone.0017258
Godfrey, K. M., and Robinson, S. (1998). Maternal nutrition, placental growth and fetal programming. Proc. Nutr. Soc. 57, 105–111. doi: 10.1079/PNS19980016
Goldowitz, D., Lussier, A. A., Boyle, J. K., Wong, K., Lattimer, S. L., Dubose, C., et al. (2014). Molecular pathways underpinning ethanol-induced neurodegeneration. Front. Genet. 5:203. doi: 10.3389/fgene.2014.00203
Govorko, D., Bekdash, R. A., Zhang, C., and Sarkar, D. K. (2012). Male germline transmits fetal alcohol adverse effect on hypothalamic proopiomelanocortin gene across generations. Biol. Psychiatry 72, 378–388. doi: 10.1016/j.biopsych.2012.04.006
Guo, W., Crossey, E. L., Zhang, L., Zucca, S., George, O. L., Valenzuela, C. F., et al. (2011). Alcohol exposure decreases CREB binding protein expression and histone acetylation in the developing cerebellum. PLoS ONE 6:e19351. doi: 10.1371/journal.pone.0019351
Habib, N., Li, Y., Heidenreich, M., Swiech, L., Avraham-Davidi, I., Trombetta, J. J., et al. (2016). Div-seq: single nucleus RNA-Seq reveals dynamics of rare adult newborn neurons. Science 353, 925–928. doi: 10.1126/science.aad7038
Haley, D. W., Handmaker, N. S., and Lowe, J. (2006). Infant stress reactivity and prenatal alcohol exposure. Alcohol. Clin. Exp. Res. 30, 2055–2064. doi: 10.1111/j.1530-0277.2006.00251.x
Hanson, M. A., and Gluckman, P. D. (2008). Developmental origins of health and disease: new insights. Basic Clin. Pharmacol. Toxicol. 102, 90–93. doi: 10.1111/j.1742-7843.2007.00186.x
Hanson, M. A., Godfrey, K. M., Lillycrop, K. A., Burdge, G. C., and Gluckman, P. D. (2011). Developmental plasticity and developmental origins of non-communicable disease: theoretical considerations and epigenetic mechanisms. Prog. Biophys. Mol. Biol. 106, 272–280. doi: 10.1016/j.pbiomolbio.2010.12.008
Harris, R. A., Wang, T., Coarfa, C., Nagarajan, R. P., Hong, C., Downey, S. L., et al. (2010). Comparison of sequencing-based methods to profile DNA methylation and identification of monoallelic epigenetic modifications. Nat. Biotechnol. 28, 1097–1105. doi: 10.1038/nbt.1682
Hellemans, K. G. C., Sliwowska, J. H., Verma, P., and Weinberg, J. (2010a). Prenatal alcohol exposure: fetal programming and later life vulnerability to stress, depression and anxiety disorders. Neurosci. Biobehav. Rev. 34, 791–807. doi: 10.1016/j.neubiorev.2009.06.004
Hellemans, K. G. C., Verma, P., Yoon, E., Yu, W., and Weinberg, J. (2008). Prenatal alcohol exposure increases vulnerability to stress and anxiety-like disorders in adulthood. Ann. N. Y. Acad. Sci. 1144, 154–175. doi: 10.1196/annals.1418.016
Hellemans, K. G. C., Verma, P., Yoon, E., Yu, W. K., Young, A. H., and Weinberg, J. (2010b). Prenatal alcohol exposure and chronic mild stress differentially alter depressive- and anxiety-like behaviors in male and female offspring. Alcohol. Clin. Exp. Res. 34, 633–645. doi: 10.1111/j.1530-0277.2009.01132.x
Heyn, H., Moran, S., Hernando-Herraez, I., Sayols, S., Gomez, A., Sandoval, J., et al. (2013). DNA methylation contributes to natural human variation. Genome Res. 23, 1363–1372. doi: 10.1101/gr.154187.112
Irizarry, R. A., Ladd-Acosta, C., Wen, B., Wu, Z., Montano, C., Onyango, P., et al. (2009). The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores. Nat. Genet. 41, 178–186. doi: 10.1038/ng.298
Iwamoto, K., Kakiuchi, C., Bundo, M., Ikeda, K., and Kato, T. (2004). Molecular characterization of bipolar disorder by comparing gene expression profiles of postmortem brains of major mental disorders. Mol. Psychiatry 9, 406–416. doi: 10.1038/sj.mp.4001437
Jacobson, S. W., Bihun, J. T., and Chiodo, L. M. (1999). Effects of prenatal alcohol and cocaine exposure on infant cortisol levels. Dev. Psychopathol. 11, 195–208. doi: 10.1017/s0954579499002011
Ji, H., Wang, Y., Jiang, D., Liu, G., Xu, X., Dai, D., et al. (2016). Elevated DRD4 promoter methylation increases the risk of Alzheimer's disease in males. Mol. Med. Rep. 14, 2732–2738. doi: 10.3892/mmr.2016.5560
Jones, K. L., and Smith, D. W. (1973). Recognition of the fetal alcohol syndrome in early infancy. Lancet 302, 999–1001. doi: 10.1016/S0140-6736(73)91092-1
Jones, P. A., and Takai, D. (2001). The role of DNA methylation in mammalian epigenetics. Science 293, 1068–1070. doi: 10.1126/science.1063852
Keiver, K., Bertram, C. P., Orr, A. P., and Clarren, S. (2015). Salivary cortisol levels are elevated in the afternoon and at bedtime in children with prenatal alcohol exposure. Alcohol 49, 79–87. doi: 10.1016/j.alcohol.2014.11.004
Kordi-Tamandani, D. M., Sahranavard, R., and Torkamanzehi, A. (2013). Analysis of association between dopamine receptor genes' methylation and their expression profile with the risk of schizophrenia. Psychiatr. Genet. 23, 183–7. doi: 10.1097/YPG.0b013e328363d6e1
Ladd-Acosta, C., Hansen, K. D., Briem, E., Fallin, M. D., Kaufmann, W. E., and Feinberg, A. P. (2013). Common DNA methylation alterations in multiple brain regions in autism. Mol. Psychiatry 19:862. doi: 10.1038/mp.2013.114
Langevin, S. M., Houseman, E. A., Christensen, B. C., Wiencke, J. K., Nelson, H. H., Karagas, M. R., et al. (2011). The influence of aging, environmental exposures and local sequence features on the variation of DNA methylation in blood. Epigenetics 6, 908–919. doi: 10.4161/epi.6.7.16431
Laufer, B. I., Kapalanga, J., Castellani, C. A., Diehl, E. J., Yan, L., and Singh, S. M. (2015). Associative DNA methylation changes in children with prenatal alcohol exposure. Epigenomics 7, 1–16. doi: 10.2217/epi.15.60
Laufer, B. I., Mantha, K., Kleiber, M. L., Diehl, E. J., Addison, S. M. F., and Singh, S. M. (2013). Long-lasting alterations to DNA methylation and ncRNAs could underlie the effects of fetal alcohol exposure in mice. Dis. Model. Mech. 6, 977–992. doi: 10.1242/dmm.010975
Lee, C., Mayfield, R. D., and Harris, R. A. (2014). Altered gamma-aminobutyric acid type B receptor subunit 1 splicing in alcoholics. Biol. Psychiatry 75, 765–773. doi: 10.1016/j.biopsych.2013.08.028
Lee, H. K., Braynen, W., Keshav, K., and Pavlidis, P. (2005). ErmineJ: tool for functional analysis of gene expression data sets. BMC Bioinformatics 6:269. doi: 10.1186/1471-2105-6-269
Lee, S., and Rivier, C. (1996). Gender differences in the effect of prenatal alcohol exposure on the hypothalamic-pituitary-adrenal axis response to immune signals. Psychoneuroendocrinology 21, 145–155. doi: 10.1016/0306-4530(95)00038-0
Leek, J. T., Johnson, W. E., Parker, H. S., Jaffe, A. E., and Storey, J. D. (2012). The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 28, 882–883. doi: 10.1093/bioinformatics/bts034
Lentini, A., Lagerwall, C., Vikingsson, S., Mjoseng, H. K., Douvlataniotis, K., Vogt, H., et al. (2018). A reassessment of DNA-immunoprecipitation-based genomic profiling. Nat. Methods 15, 499–504. doi: 10.1038/s41592-018-0038-7
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Li, Z., Xiang, Y., Chen, J., Li, Q., Shen, J., Liu, Y., et al. (2015). Loci with genome-wide associations with schizophrenia in the Han Chinese population. Br. J. Psychiatry 207, 490–494. doi: 10.1192/bjp.bp.114.150490
Lindner, M., Thümmler, K., Arthur, A., Brunner, S., Elliott, C., McElroy, D., et al. (2015). Fibroblast growth factor signalling in multiple sclerosis: inhibition of myelination and induction of pro-inflammatory environment by FGF9. Brain 138, 1875–1893. doi: 10.1093/brain/awv102
Lister, R., Mukamel, E. A., Nery, J. R., Urich, M., Puddifoot, C. A., Johnson, N. D., et al. (2013). Global epigenomic reconfiguration during mammalian brain development. Science 341:1237905. doi: 10.1126/science.1237905
Liyanage, V. R. B., Zachariah, R. M., Davie, J. R., and Rastegar, M. (2015). Ethanol deregulates Mecp2/MeCP2 in differentiating neural stem cells via interplay between 5-methylcytosine and 5-hydroxymethylcytosine at the Mecp2 regulatory elements. Exp. Neurol. 265, 102–117. doi: 10.1016/j.expneurol.2015.01.006
Lowe, R., Gemma, C., Beyan, H., Hawa, M. I., Bazeos, A., Leslie, R. D., et al. (2013). Buccals are likely to be a more informative surrogate tissue than blood for epigenome-wide association studies. Epigenetics 8, 445–454. doi: 10.4161/epi.24362
Luo, X., Kranzler, H. R., Zuo, L., Lappalainen, J., Yang, B., and Gelernter, J. (2005). ADH4 gene variation is associated with alcohol dependence and drug dependence in european Americans: results from HWD tests and case-control association studies. Neuropsychopharmacology 31, 1085–1095. doi: 10.1038/sj.npp.1300925
Lussier, A. A., Morin, A. M., MacIsaac, J. L., Salmon, J., Weinberg, J., Reynolds, J. N., et al. (2018). DNA methylation as a predictor of fetal alcohol spectrum disorder. Clin. Epigenet. 10:5. doi: 10.1186/s13148-018-0439-6
Lussier, A. A., Stepien, K. A., Neumann, S. M., Pavlidis, P., Kobor, M. S., and Weinberg, J. (2015). Prenatal alcohol exposure alters steady-state and activated gene expression in the adult rat brain. Alcohol. Clin. Exp. Res. 39, 251–261. doi: 10.1111/acer.12622
Lussier, A. A., Weinberg, J., and Kobor, M. S. (2017). Epigenetics studies of fetal alcohol spectrum disorder: where are we now? Epigenomics 9, 291–311. doi: 10.2217/epi-2016-0163
MacKay, R. K., Colson, N. J., Dodd, P. R., and Lewohl, J. M. (2011). Differential expression of 14-3-3 isoforms in human alcoholic brain. Alcohol. Clin. Exp. Res. 35, 1041–1049. doi: 10.1111/j.1530-0277.2011.01436.x
Mathew, D. E., Larsen, K., Janeczek, P., and Lewohl, J. M. (2016). Expression of 14-3-3 transcript isoforms in response to ethanol exposure and their regulation by miRNAs. Mol. Cell. Neurosci. 75, 44–49. doi: 10.1016/j.mcn.2016.06.006
Matthews, S. G. (2002). Early programming of the hypothalamo-pituitary-adrenal axis. Trends Endocrinol. Metab. 13, 373–380. doi: 10.1016/S1043-2760(02)00690-2
Mattson, S. N., Crocker, N., and Nguyen, T. T. (2011). Fetal alcohol spectrum disorders: neuropsychological and behavioral features. Neuropsychol. Rev. 21, 81–101. doi: 10.1007/s11065-011-9167-9
Maunakea, A. K., Chepelev, I., Cui, K., and Zhao, K. (2013). Intragenic DNA methylation modulates alternative splicing by recruiting MeCP2 to promote exon recognition. Cell Res. 23, 1256–1269. doi: 10.1038/cr.2013.110
Maunakea, A. K., Nagarajan, R. P., Bilenky, M., Ballinger, T. J., D'Souza, C., Fouse, S. D., et al. (2010). Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 466, 253–257. doi: 10.1038/nature09165
Maurano, M. T., Wang, H., John, S., Shafer, A., Canfield, T., Lee, K., et al. (2015). Role of DNA methylation in modulating transcription factor occupancy. CellReports 12, 1184–1195. doi: 10.1016/j.celrep.2015.07.024
May, P. A., Chambers, C. D., Kalberg, W. O., Zellner, J., Feldman, H., Buckley, D., et al. (2018). Prevalence of fetal alcohol spectrum disorders in 4 US communities. JAMA 319, 474–482. doi: 10.1001/jama.2017.21896
McCormick, C. M., and Mathews, I. Z. (2010). Adolescent development, hypothalamic-pituitary-adrenal function, and programming of adult learning and memory. Prog. Neuro-Psychopharmacology Biol. Psychiatry 34, 756–765. doi: 10.1016/j.pnpbp.2009.09.019
McLachlan, K., Rasmussen, C., Oberlander, T. F., Loock, C., Pei, J., Andrew, G., et al. (2016). Dysregulation of the cortisol diurnal rhythm following prenatal alcohol exposure and early life adversity. Alcohol 53, 9–18. doi: 10.1016/j.alcohol.2016.03.003
Medici, V., and Halsted, C. H. (2013). Folate, Alcohol, and Liver Disease. Mol. Nutr. Food Res. 57, 596–606. doi: 10.1002/mnfr.201200077
Milani, P., Escalante-chong, R., Shelley, B. C., Patel-murray, N. L., Xin, X., Adam, M., et al. (2016). Cell freezing protocol suitable for ATAC-Seq on motor neurons derived from human induced pluripotent stem cells. Nat. Publ. Gr. 2016, 1–10. doi: 10.1038/srep25474
Mill, J., and Heijmans, B. T. (2013). From promises to practical strategies in epigenetic epidemiology. Nat. Rev. Genet. 14, 585–594. doi: 10.1038/nrg3405
Moen, E. L., Zhang, X., Mu, W., Delaney, S. M., Wing, C., McQuade, J., et al. (2013). Genome-wide variation of cytosine modifications between European and African populations and the implications for complex traits. Genetics 194, 987–996. doi: 10.1534/genetics.113.151381
Mukhopadhyay, P., Rezzoug, F., Kaikaus, J., Greene, R. M., and Pisano, M. M. (2013). Alcohol modulates expression of DNA methyltranferases and methyl CpG-/CpG domain-binding proteins in murine embryonic fibroblasts. Reprod. Toxicol. 37, 40–48. doi: 10.1016/j.reprotox.2013.01.003
Nagre, N. N., Subbanna, S., Shivakumar, M., Psychoyos, D., and Basavarajappa, B. S. (2015). CB1-receptor knockout neonatal mice are protected against ethanol-induced impairments of DNMT1, DNMT3A, and DNA methylation. J. Neurochem. 132, 429–442. doi: 10.1111/jnc.13006
Nakashima, A., Kawamoto, T., Honda, K. K., Ueshima, T., Noshiro, M., Iwata, T., et al. (2008). DEC1 modulates the circadian phase of clock gene expression. Mol. Cell. Biol. 28, 4080–4092. doi: 10.1128/MCB.02168-07
Ngai, Y. F., Sulistyoningrum, D. C., O'Neill, R., Innis, S. M., Weinberg, J., and Devlin, A. M. (2015). Prenatal alcohol exposure alters methyl metabolism and programs serotonin transporter and glucocorticoid receptor expression in brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 309, R613–R622. doi: 10.1152/ajpregu.00075.2015
Nikolayeva, O., and Robinson, M. D. (2014). edgeR for Differential RNA-seq and ChIP-seq analysis: an application to stem cell biology. Methods Mol. Biol. 1150, 45–79. doi: 10.1007/978-1-4939-0512-6_3
Noor, S., and Milligan, E. D. (2018). Lifelong impacts of moderate prenatal alcohol exposure on neuroimmune function. Front. Immunol. 9:1107. doi: 10.3389/fimmu.2018.01107
Oldehinkel, A. J., and Bouma, E. M. C. (2011). Sensitivity to the depressogenic effect of stress and HPA-axis reactivity in adolescence: a review of gender differences. Neurosci. Biobehav. Rev. 35, 1757–1770. doi: 10.1016/j.neubiorev.2010.10.013
Osborne, T. F. (2001). CREating a SCAP-less liver keeps SREBPs pinned in the ER membrane and prevents increased lipid synthesis in response to low cholesterol and high insulin. Genes Dev. 15, 1873–1878. doi: 10.1101/gad.916601
Otero, N. K. H., Thomas, J. D., Saski, C. A., Xia, X., and Kelly, S. J. (2012). Choline supplementation and DNA methylation in the hippocampus and prefrontal cortex of rats exposed to alcohol during development. Alcohol. Clin. Exp. Res. 36, 1701–1709. doi: 10.1111/j.1530-0277.2012.01784.x
Öztürk, N. C., Resendiz, M., Öztürk, H., and Zhou, F. C. (2017). DNA Methylation program in normal and alcohol-induced thinning cortex. Alcohol 60, 135–147. doi: 10.1016/j.alcohol.2017.01.006
Pagnamenta, A. T., Bacchelli, E., de Jonge, M. V., Mirza, G., Scerri, T. S., Minopoli, F., et al. (2010). Characterization of a family with rare deletions in CNTNAP5 and DOCK4 suggests novel risk loci for autism and dyslexia. Biol. Psychiatry 68, 320–328. doi: 10.1016/j.biopsych.2010.02.002
Panzica, G., and Melcangi, R. C. (2016). Structural and molecular brain sexual differences: a tool to understand sex differences in health and disease. Neurosci. Biobehav. Rev. 67, 2–8. doi: 10.1016/j.neubiorev.2016.04.017
Pei, J., Denys, K., Hughes, J., and Rasmussen, C. (2011). Mental health issues in fetal alcohol spectrum disorder. J. Ment. Heal. 20, 473–483. doi: 10.3109/09638237.2011.577113
Pelizzola, M., Koga, Y., Urban, A. E., Krauthammer, M., Weissman, S., Halaban, R., et al. (2008). MEDME: an experimental and analytical methodology for the estimation of DNA methylation levels based on microarray derived MeDIP-enrichment. Genome Res. 18, 1652–1659. doi: 10.1101/gr.080721.108
Perkins, A., Lehmann, C., Lawrence, R. C., and Kelly, S. J. (2013). Alcohol exposure during development: Impact on the epigenome. Int. J. Dev. Neurosci. 31, 391–397. doi: 10.1016/j.ijdevneu.2013.03.010
Pollard, I. (2007). Neuropharmacology of drugs and alcohol in mother and fetus. Semin. Fetal Neonatal Med. 12, 106–113. doi: 10.1016/j.siny.2006.12.001
Portales-Casamar, E., Lussier, A. A., Jones, M. J., MacIsaac, J. L., Edgar, R. D., Mah, S. M., et al. (2016). DNA methylation signature of human fetal alcohol spectrum disorder. Epigenet. Chromatin 9, 81–101. doi: 10.1186/S13072-016-0074-4
Ptácek, R., KuŽelová, H., and Stefano, G. B. (2011). Dopamine D4 receptor gene DRD4 and its association with psychiatric disorders. Med. Sci. Monit. 17, RA215–RA220. doi: 10.12659/MSM.881925
Quinlan, A. R., and Hall, I. M. (2010). BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842. doi: 10.1093/bioinformatics/btq033
Rakyan, V. K., Down, T. A., Balding, D. J., and Beck, S. (2011). Epigenome-wide association studies for common human diseases. Nat. Rev. Genet. 12, 529–541. doi: 10.1038/nrg3000
Ramsay, D. S., Bendersky, M. I., and Lewis, M. (1996). Effect of prenatal alcohol and cigarette exposure on two- and six-month-old infants' adrenocortical reactivity to stress. J. Pediatr. Psychol. 21, 833–840. doi: 10.1093/jpepsy/21.6.833
Rice, G. I., del Toro Duany, Y., Jenkinson, E. M., Forte, G. M. A., Anderson, B. H., Ariaudo, G., et al. (2014). Gain-of-function mutations in IFIH1 cause a spectrum of human disease phenotypes associated with upregulated type I interferon signaling. Nat. Genet. 46, 503–509. doi: 10.1038/ng.2933
Riley, E. P., Infante, M. A., and Warren, K. R. (2011). Fetal alcohol spectrum disorders: an overview. Neuropsychol. Rev. 21, 73–80. doi: 10.1007/s11065-011-9166-x
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010a). EdgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Robinson, M. D., Stirzaker, C., Statham, A. L., Coolen, M. W., Song, J. Z., Nair, S. S., et al. (2010b). Evaluation of affinity-based genome-wide DNA methylation data: effects of CpG density, amplification bias, and copy number variation. Genome Res. 20, 1719–1729. doi: 10.1101/gr.110601.110
Rogic, S., Wong, A., and Pavlidis, P. (2016). Meta-analysis of gene expression patterns in animal models of prenatal alcohol exposure suggests role for protein synthesis inhibition and chromatin remodeling. Alcohol. Clin. Exp. Res. 40, 717–727. doi: 10.1111/acer.13007
Schott, J. M., Crutch, S. J., Carrasquillo, M. M., Uphill, J., Shakespeare, T. J., Ryan, N. S., et al. (2016). Genetic risk factors for the posterior cortical atrophy variant of Alzheimer's disease. Alzheimer's Dement. 12, 862–871. doi: 10.1016/j.jalz.2016.01.010
Sei, H., Sakata-Haga, H., Ohta, K., Sawada, K., Morita, Y., and Fukui, Y. (2003). Prenatal exposure to alcohol alters the light response in postnatal circadian rhythm. Brain Res. 987, 131–134. doi: 10.1016/S0006-8993(03)03329-8
Shue, E. H., Carson-Walter, E. B., Liu, Y., Winans, B. N., Ali, Z. S., Chen, J., et al. (2008). Plasmalemmal Vesicle Associated Protein-1 (PV-1) is a marker of blood-brain barrier disruption in rodent models. BMC Neurosci. 9:29. doi: 10.1186/1471-2202-9-29
Shukla, S., Kavak, E., Gregory, M., Imashimizu, M., Shutinoski, B., Kashlev, M., et al. (2011). CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature 479, 74–79. doi: 10.1038/nature10442
Shulha, H. P., Cheung, I., Guo, Y., Akbarian, S., and Weng, Z. (2013). Coordinated cell type-specific epigenetic remodeling in prefrontal cortex begins before birth and continues into early adulthood. PLoS Genet. 9:e1003433. doi: 10.1371/journal.pgen.1003433
Smith, A. C. M., Gropman, A. L., Bailey-Wilson, J. E., Goker-Alpan, O., Elsea, S. H., Blancato, J., et al. (2002). Hypercholesterolemia in children with Smith-Magenis syndrome: del (17)(p11.2p11.2). Genet. Med. 4, 118–125. doi: 10.1097/00125817-200205000-00004
Smith, Z. D., and Meissner, A. (2013). DNA methylation: roles in mammalian development. Nat. Rev. Genet. 14, 204–220. doi: 10.1038/nrg3354
Stark, R., and Brown, G. (2011). DiffBind: Differential Binding Analysis of ChIP-Seq Peak Data. Available online at: https://bioconductor.org/packages/release/bioc/html/DiffBind.html
Stratton, K., Howe, C., and Battaglia, F. (1996). Fetal Alcohol Syndrome: Diagnosis, Epidemiology, Prevention and Treatment. Washington, DC: National Academy Press.
Subbanna, S., Nagre, N. N., Shivakumar, M., Umapathy, N. S., Psychoyos, D., and Basavarajappa, B. S. (2014). Ethanol induced acetylation of histone at G9a exon1 and G9a-mediated histone H3 dimethylation leads to neurodegeneration in neonatal mice. Neuroscience 258, 422–432. doi: 10.1016/j.neuroscience.2013.11.043
Subbanna, S., Shivakumar, M., Umapathy, N. S., Saito, M., Mohan, P. S., Kumar, A., et al. (2013). G9a-mediated histone methylation regulates ethanol-induced neurodegeneration in the neonatal mouse brain. Neurobiol. Dis. 54, 475–485. doi: 10.1016/j.nbd.2013.01.022
Swanson, J. M., Entringer, S., Buss, C., and Wadhwa, P. D. (2009). Developmental origins of health and disease: environmental exposures. Semin. Reprod. Med. 27, 391–402. doi: 10.1055/s-0029-1237427
Taiwo, O., Wilson, G., Morris, T., Seisenberger, S., Reik, W., Pearce, D., et al. (2012). Methylome analysis using MeDIP-seq with low DNA concentrations. Nat. Protoc. 7, 617–636. doi: 10.1038/nprot.2012.012
Tate, P. H., and Bird, A. P. (1993). Effects of DNA methylation on DNA-binding proteins and gene expression. Curr. Opin. Genet. Dev. 3, 226–231. doi: 10.1016/0959-437X(93)90027-M
Thisse, B., and Thisse, C. (2005). Functions and regulations of fibroblast growth factor signaling during embryonic development. Dev. Biol. 287, 390–402. doi: 10.1016/j.ydbio.2005.09.011
Uban, K. A., Sliwowska, J. H., Lieblich, S., Ellis, L. A., Yu, W. K., Weinberg, J., et al. (2010). Prenatal alcohol exposure reduces the proportion of newly produced neurons and glia in the dentate gyrus of the hippocampus in female rats. Horm. Behav. 58, 835–843. doi: 10.1016/j.yhbeh.2010.08.007
Veazey, K. J., Parnell, S. E., Miranda, R. C., and Golding, M. C. (2015). Dose-dependent alcohol-induced alterations in chromatin structure persist beyond the window of exposure and correlate with fetal alcohol syndrome birth defects. Epigenetics Chromatin 8:39. doi: 10.1186/s13072-015-0031-7
Wang, T., Pan, Q., Lin, L., Szulwach, K. E., Song, C. X., He, C., et al. (2012). Genome-wide DNA hydroxymethylation changes are associated with neurodevelopmental genes in the developing human cerebellum. Hum. Mol. Genet. 21, 5500–5510. doi: 10.1093/hmg/dds394
Weinberg, J., Sliwowska, J. H., Lan, N., and Hellemans, K. G. C. (2008). Prenatal alcohol exposure: foetal programming, the hypothalamic-pituitary-adrenal axis and sex differences in outcome. J. Neuroendocrinol. 20, 470–488. doi: 10.1111/j.1365-2826.2008.01669.x
Werts, R. L., Van Calcar, S. C., Wargowski, D. S., and Smith, S. M. (2014). Inappropriate feeding behaviors and dietary intakes in children with fetal alcohol spectrum disorder or probable prenatal alcohol exposure. Alcohol. Clin. Exp. Res. 38, 871–878. doi: 10.1111/acer.12284
Wilhelm, C. J., and Guizzetti, M. (2015). Fetal alcohol spectrum disorders: an overview from the glia perspective. Front. Integr. Neurosci. 9:65. doi: 10.3389/fnint.2015.00065
Wong, M., and Munro, S. (2014). The specificity of vesicle traffic to the Golgi is encoded in the golgin coiled-coil proteins. Science 346:1256898. doi: 10.1126/science.1256898
Xu, W., Cohen-Woods, S., Chen, Q., Noor, A., Knight, J., Hosang, G., et al. (2014). Genome-wide association study of bipolar disorder in Canadian and UK populations corroborates disease loci including SYNE1 and CSMD1. BMC Med. Genet. 15:2. doi: 10.1186/1471-2350-15-2
Yagi, S., and Galea, L. A. M. (2018). Sex differences in hippocampal cognition and neurogenesis. Neuropsychopharmacology 44, 200–213. doi: 10.1038/s41386-018-0208-4
Yin, Y., Morgunova, E., Jolma, A., Kaasinen, E., Sahu, B., Khund-Sayeed, S., et al. (2017). Impact of cytosine methylation on DNA binding specificities of human transcription factors. Science 356:eaaj2239. doi: 10.1126/science.aaj2239.
Yuen, R. K. C., Neumann, S. M. A., Fok, A. K., Peñaherrera, M. S., McFadden, D. E., Robinson, W. P., et al. (2011). Extensive epigenetic reprogramming in human somatic tissues between fetus and adult. Epigenet. Chromatin 4:7. doi: 10.1186/1756-8935-4-7
Zeng, H., Kaul, S., and Simons, S. S. (2000). Genomic organization of human GMEB-1 and rat GMEB-2: structural conservation of two multifunctional proteins. Nucleic Acids Res. 28, 1819–1829. doi: 10.1093/nar/28.8.1819
Zhang, C. R., Ho, M.-F., Vega, M. C. S., Burne, T. H. J., and Chong, S. (2015). Prenatal ethanol exposure alters adult hippocampal VGLUT2 expression with concomitant changes in promoter DNA methylation, H3K4 trimethylation and miR-467b-5p levels. Epigenet. Chromat. 8:40. doi: 10.1186/s13072-015-0032-6
Zhang, F. F., Cardarelli, R., Carroll, J., Fulda, K. G., Kaur, M., Gonzalez, K., et al. (2011). Significant differences in global genomic DNA methylation by gender and race/ethnicity in peripheral blood. Epigenetics 6, 623–629. Available online at: http://eutils.ncbi.nlm.nih.gov/entrez/eutils/elink.fcgi?dbfrom=pubmedandid=21739720andretmode=refandcmd=prlinks
Zhang, H., Herman, A. I., Kranzler, H. R., Anton, R. F., Zhao, H., Zheng, W., et al. (2013). Array-based profiling of DNA methylation changes associated with alcohol dependence. Alcohol. Clin. Exp. Res. 37 Suppl 1, E108–115. doi: 10.1111/j.1530-0277.2012.01928.x
Zhang, X., Sliwowska, J. H., and Weinberg, J. (2005). Prenatal alcohol exposure and fetal programming: effects on neuroendocrine and immune function. Exp. Biol. Med. (Maywood). 230, 376–388. doi: 10.1177/15353702-0323006-05
Zhang, Y., Liu, T., Meyer, C. A., Eeckhoute, J., Johnson, D. S., Bernstein, B. E., et al. (2008). Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9:R137. doi: 10.1186/gb-2008-9-9-r137
Zhou, F. C., Balaraman, Y., Teng, M., Liu, Y., Singh, R. P., and Nephew, K. P. (2011). Alcohol Alters DNA Methylation Patterns and Inhibits Neural Stem Cell Differentiation. Alcohol. Clin. Exp. Res. 35, 735–746. doi: 10.1111/j.1530-0277.2010.01391.x
Ziller, M. J., Gu, H., Müller, F., Donaghey, J., Tsai, L. T.-Y., Kohlbacher, O., et al. (2014). Charting a dynamic DNA methylation landscape of the human genome. Nature 500, 477–481. doi: 10.1038/nature12433
Keywords: prenatal alcohol exposure, fetal alcohol spectrum disorder, brain, development, epigenetics, DNA methylation, immune
Citation: Lussier AA, Bodnar TS, Mingay M, Morin AM, Hirst M, Kobor MS and Weinberg J (2018) Prenatal Alcohol Exposure: Profiling Developmental DNA Methylation Patterns in Central and Peripheral Tissues. Front. Genet. 9:610. doi: 10.3389/fgene.2018.00610
Received: 05 September 2018; Accepted: 19 November 2018;
Published: 04 December 2018.
Edited by:
Mojgan Rastegar, University of Manitoba, CanadaReviewed by:
Kazuhiko Nakabayashi, National Center for Child Health and Development (NCCHD), JapanCopyright © 2018 Lussier, Bodnar, Mingay, Morin, Hirst, Kobor and Weinberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexandre A. Lussier, YWx1c3NpZXJAY21tdC51YmMuY2E=
†Present Address: Alexandre A. Lussier, Department of Biological Statistics and Computational Biology, Cornell University, Ithaca, NY, United States
‡These authors have contributed equally to this work and are senior authors
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.