 Josef Pánek
Josef Pánek Martin Modrák
Martin Modrák Marek Schwarz
Marek Schwarz
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 10 October 2017
Sec. Computational Genomics
Volume 8 - 2017 | https://doi.org/10.3389/fgene.2017.00147
While understanding the structure of RNA molecules is vital for deciphering their functions, determining RNA structures experimentally is exceptionally hard. At the same time, extant approaches to computational RNA structure prediction have limited applicability and reliability. In this paper we provide a method to solve a simpler yet still biologically relevant problem: prediction of secondary RNA structure using structure of different molecules as a template. Our method identifies conserved and unconserved subsequences within an RNA molecule. For conserved subsequences, the template structure is directly transferred into the generated structure and combined with de-novo predicted structure for the unconserved subsequences with low evolutionary conservation. The method also determines, when the generated structure is unreliable. The method is validated using experimentally identified structures. The accuracy of the method exceeds that of classical prediction algorithms and constrained prediction methods. This is demonstrated by comparison using large number of heterogeneous RNAs. The presented method is fast and robust, and useful for various applications requiring knowledge of secondary structures of individual RNA sequences.
Despite recent improvements [SHAPE-seq (Loughrey et al., 2014), PARS (Kertesz et al., 2010), and FragSeq (Underwood et al., 2010)], experimental identification of RNA structures is technically demanding and only a limited number of RNA structures has been resolved. Therefore, computational predictions of RNA secondary structures are frequently employed as proxies for native structures. There is plenty of heterogeneous prediction methods, that could be broadly categorized into (a) methods based on free energy and dynamic programming techniques, reviewed in Mathews et al. 2006 (Mathews, 2006) or (b) comparative methods as reviewed in Gardner et al. 2004 (Gardner and Giegerich, 2004). However, known methods in both categories are unreliable for longer sequences (~>150 nucleotides) and more complex structures, e.g., those that contain longer single-stranded segments. This is owing to the extreme theoretical complexity of the prediction.
Nevertheless, the number of experimentally identified RNA structures is growing in spite of the technical demands. These structures are available as potential templates to generate secondary structures of uncharacterized but related RNA sequences. In principle, template-based prediction can be treated as constrained prediction, which is supported by several methods, e.g., the RNA Vienna Package (Lorenz et al., 2011), RNAstructure (Mathews, 2004), and Locarna (Smith et al., 2010). However, the conversion of the template into a structural constraint for Locarna and RNAstructure is not trivial, as the template and the query sequence frequently have different lengths. Locarna produces a consensus structure different from an individual structure that cannot be mapped directly. RNAstructure requires multiple input sequences as input otherwise it predicts either MFE structure or a set of probable structures. The only directly applicable method is thus via RNA Vienna Package.
It includes a utility, refold.pl, that can be used for conversion of secondary structures into an RNAfold constraint through a sequence alignment. The constraint is then used for RNAfold constrained prediction.
A different approach to the template-based prediction was adopted for ribosomal RNAs (Gutell et al., 2002). The rRNA structure cannot be predicted by available methods due to their lengths and complexity. The method uses known rRNA structures as templates for comparative prediction of homologous sequences but it requires extensive manual input and is slow.
Our approach differs from both free energy and comparative methods: as we generate RNA secondary structure for the molecule under investigation directly from the template structure. The method uses a heuristic to determine which fragments of the structure can be transferred directly and employs free energy prediction algorithms to resolve the structure of the rest of the fragments. The reliability of the generated structure is then evaluated by a bootstrapping scheme. We show that the proposed method achieves high-quality predictions for sequences where a structure for a putative homolog is known, including a number of sequences that are intractable by current prediction software. The presented method is suitable for all RNAs that adopt a secondary structure.
An utility based on the method is available on request from the authors.
In this section, we first describe the proposed prediction method and then deal with the evaluation methodology. In the following text, we use the term “structure” to refer exclusively to secondary structure.
Input to the template-based prediction task consists of a template sequence, the corresponding template structure and a query sequence. The task is to predict the structure of the query sequence. The structure to be predicted is called the query structure.
In terms of folding space, we have a subspace of the complete folding space of the query sequence. The subspace contains all possible structures of evolutionarily unconserved segments of the query structure, while the structure of evolutionarily conserved segments is taken from the template and kept fixed. The solution over this subspace is in principle easier than over the complete space, and can be found by determining the optimal structure of the unconserved segments. An overview of the method is shown in Figure 1. A detailed description of the individual steps follows:
I. A pairwise alignment of the query and template sequences is computed with ClustalW2 (with default parameters except for GAPOPEN = 7 a GAPEXT = 0.5) (Thompson et al., 1994). The alignment can be treated as two functions: Aq maps positions in the query sequence to positions in the template sequence and At maps from template to query sequence. An example alignment is shown in Table S1.
II. The template structure is mapped onto the query sequence, producing intermediate structure. The intermediate structure preserves base pairs that the alignment maps to complementary nucleotides and marks all other bases as unpaired (see Figure 2). More precisely, for each position p in the query sequence, there are four possibilities:
1. Aq(p) is a gap,
2. Aq(p) is not paired in the template structure,
3. Aq(p) is paired to position r, but At(r) is either a gap or a non-canonical pair for p,
4. Aq(p) is paired to position r and At(r) is a canonical pair for p
In cases 1–3 the intermediate query structure marks p as unpaired, in case 4, p is paired with At(r). Further, in cases 2 and 4, the position p is considered to be consistent while in cases 1 and 3, p is considered to be inconsistent.
An example of an intermediate structure is shown in Table S1 and Figure 1B.
III. The intermediate structure is decomposed into basic structure elements: individual hairpins and stems (Figure 1C). Hairpins are identified first, then stems. Hairpins are identified by the following procedure:
1. The loops of the hairpins are identified first as base pairs with only unpaired nucleotides in between the pairing nucleotides.
2. From this base pair, both ends of the hairpin are extended until first base pair of a different hairpin is encountered on either end. The strands of the hairpin must contain the same number of pairing nucleotides. All single-strand nucleotides between pairing nucleotides are added to the hairpin as well.
3. If there are single-strand nucleotides following the last base pair of a hairpin, they are added to the hairpin while ensuring they are not shared by multiple neighboring hairpins.
Stems are identified in between hairpins. Stems have two strands, the 5′ strand and the 3′ strand, identified by the following procedure:
1. The strands of the stem start at the first nucleotide not occupied by hairpins or previously identified stems at 5′ and 3′ ends of the intermediate structure for the 5′ and 3′ strands, respectively.
2. The strands are extended in opposite directions, i.e., the 5′ strand in 5′->3′ and the 3′ strand in 3′->5′ for the same number of base pairs, until a base pair belonging already to a hairpin or a base paired to a non-neighboring part of the sequence is encountered.
Unlike hairpins, stems are not extended with neighboring single-strand nucleotides.
IV. Inconsistent elementary structure elements (Figure 1D) are identified. Structural elements are considered inconsistent, if their proportion of inconsistent positions identified in step II is over a given threshold. The threshold was set to 20% for hairpins and 10% for stems. The threshold values were identified based on optimization using both the cross-validation and large scale datasets.
V De novo prediction of the structure of the inconsistent elements (Figure 1E). RNAfold and RNAduplex (Hofacker et al., 2004) were used for hairpins and stems, respectively. The goal of this step is that the prediction corrects the wrong structure information at inconsistent positions. The advantage is that the inconsistent elements are small and therefore the prediction of their structure is highly reliable in contrast to the prediction of the whole structure.
VI The de novo predicted structures of the inconsistent elements are combined with the intermediate structure of the consistent elements (Figure 1F) to form the resulting structure.
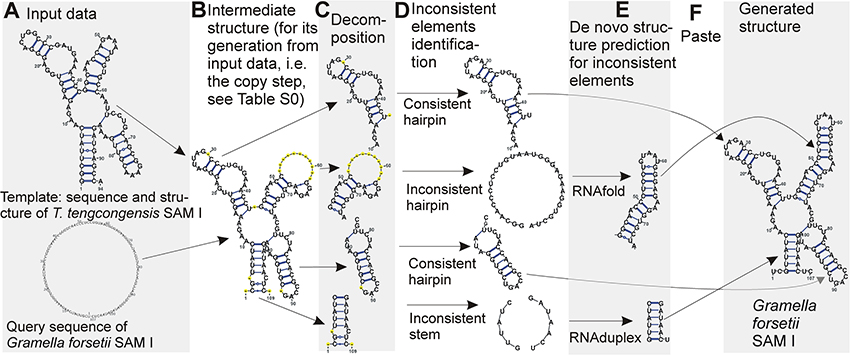
Figure 1. Demonstration of the method using SAM I structure. Starting with input data (A), an intermediate structure is built first (B), the structure is decomposed into individual hairpins and stems (C). If the structure of the individual elements is consistent, it is kept (D) while the structure of inconsistent elements is predicted de novo (E). The structures for the individual elements are then combined to form the final predicted structure (F). Structures are plotted by VARNA viewer (Darty et al., 2009). The sequence representation of the copy step is in Table S1. Note that VARNA interprets the signs for false non-canonical base pairs (“3”), gapped base pairs (“1”) and gaps (“–”) used in Table S1 as “–”signs in yellow circles.
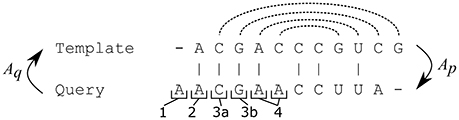
Figure 2. Possible situations when building the intermediate structure from the template structure. Dashed lines represent pairs in the template structure. A position in the query sequence can be mapped to a gap (1) or mapped to an unpaired position (2) or mapped to a position paired with a gap (3a) or a non-canonical base-pair (3b) in the query sequence or mapped to a position paired with a complementary nucleotide in the query sequence (4). Only the pairs from case 4 are preserved in the intermediate structure and positions from cases 1 and 3 are marked as inconsistent.
Since the presented method will generate a structure for any input, even if the template and query sequences are completely unrelated, it is important that we distinguish reliable results from spurious ones.
We compute the reliability using a bootstrapping scheme. We use the query sequence to generate N sequences with randomly shuffled dinucleotides. For the shuffled sequences, structures are generated with the same procedure as for the query sequence.
First we validated a criterion that is evaluated by the bootstrapping scheme. We chose between tree edit distances and free energy (FE). For the first, distances drnd = {drnd, 1,…,drnd, N} between the generated structures and the template structure are computed. For the later, FEs ernd = {ernd, 1,…,ernd, N} of the generated structures are computed.
Now drnd and ernd approximate the distributions of tree edit distances and FEs obtained from non-homologous, i.e., shuffled sequences with the same length and nucleotide composition. The quality of the generated structure is then assessed with a z-score (Kreyszig, 1979) relative to the population of non-homologous sequences:
The generated structure of the original sequence is considered reliable with a z-score ≥ 2 (corresponding to the limit of the statistical significance of p = 0.05). In our experiments, we used N = 100.
Unlike direct use of the tree edit distance, the z-scores are relevant also when the query sequence is only a fragment of the template sequence. The generated structure is then naturally dissimilar to the template structure and has a relatively large tree edit distance. But reliable structure can still be generated by transferring the relevant substructure of the template. The presented bootstrapping scheme correctly classifies such substructures as reliable.
For purpose of validation of the bootstrap metrics and evaluation of the variance of z-scores, we repeated the bootstrap 100 times (100 runs with 100 randomized sequences each) for the 52 generated structures of the cross-validation dataset.
The presented method was compared to a classical prediction method (RNAfold), constrained prediction with refold.pl and RNAfold, and constrained prediction using Rsearch/infernal alignments and RNAfold. The Vienna RNA package ver. 2.3.3 and the infernal package ver. 1.1.2 (July 2016) were used. The first method represents state-of-the-art of de novo prediction and is included mainly to put the improvements made by our method into proper scale. The latter two tools should in theory perform the same task as our method and use the same input information and thus are a more fair comparison.
The refold.pl script takes as input an alignment and a consensus structure. To perform template-based prediction we pass it a pairwise alignment between the subject sequence and the sequence of the template and the template structure extended to have the same length as the alignment by introducing the gaps identified by the alignment into it. The constraints were then used with RNAfold −C, as described in the Vienna RNA package user guide.
To perform template-based prediction using Rsearch/infernal method (Nawrocki and Eddy, 2013), we build a CM-model using the sequence of the template and the template structure in the Stockholm format using the command “cmbuild -F –rsearch RIBOSUM85.mat/RIBOSUM65.mat CM_model input_stockholm” and aligned the CM model with query sequences using cmalign. Non-canonical base pairs were removed from the alignment and the remaining base pairs were used as constraints for RNAfold constrained prediction.
The comparison consists of two steps: first we validate the proposed method on a small dataset of experimentally identified RNA structures and then we perform a large scale evaluation on sequences without known structure.
For the cross-validation, RNA families with at least two homologs with experimentally identified structures were identified and used. They let us to generate structure of one homolog using structure of other homolog of the same family as a template/constraint and vice versa. The generated structures were then compared to their experimentally identified counterparts.
The sequences and structures of the experimentally identified RNAs used for the cross-validation are shown in Supplementary File S1.fasta. We collected 34 families with at least two experimentally identified structures per one family mainly from PDB, allowing for 52 predictions. The sources of the structures including databases and/or related papers are included in Table S4.
Accuracy of the generated/predicted structures was evaluated using two criteria: (1) percent of nucleotide positions with correctly predicted structural information, (2) tree edit distances (computed by RNAdistance; Hofacker, 2004) to the experimentally identified structures. Ideally, the generated/predicted structures should have 100% of nucleotide positions with correctly predicted structural information and their tree edit distance to the experimentally identified structures should be zero.
While the cross validation dataset provides exact results since true structures of the query sequences are known, it covers only a small portion of the variability of known RNA species in terms of both structure and sequence. We therefore performed a larger comparison using RNA families where a structure of at least one possible template is available, but using query sequences without experimentally determined structures. This forces us to use a proxy metric for evaluation of the quality of the structures, but permits testing our method on a much more variable dataset and understanding its robustness.
The comparison was carried out using a reference dataset. Its characteristics are summarized in Tables S2, S3. The sequences are included in Supplementary File S2.fasta. Templates including their sequences and structures are included in Supplementary File S2a.fasta. The dataset was created from the test dataset of CentroidHomfold (Kiryu et al., 2007; Hamada et al., 2009) and extended with other RNAs to get more sequence/structure variability. The dataset consisted of 32 RNA families where at least one experimentally identified structure is known. In total, the dataset contains 3,192 sequences with pairwise sequence similarity within families ranging from 43 to 95% and sequence similarity to the templates of 20–93%. The sequences were downloaded from Rfam (Gardner et al., 2009) and SRPDB (Andersen et al., 2006) databases, or when unavailable in the databases, identified using corresponding papers cited in Table S3 and downloaded from Genbank. As templates, the experimentally identified structures were used, downloaded together with their sequences from databases (mostly PDB; Sussman et al., 1998) or acquired using corresponding papers (Table S3).
The results of the cross-validation are summarized in Table S4 and Figure 3. The methodology of the cross-validation is explained in details in Methods. The proposed method generated more accurate structures than RNAfold and the refold method for 49 of total 52 predictions (94%), and was more accurate than the Rsearch-based method for 42 of total 52 predictions (81%). The result was the same, when the accuracy was evaluated by tree edit distance and percentage of nucleotide positions with correctly predicted structural information.
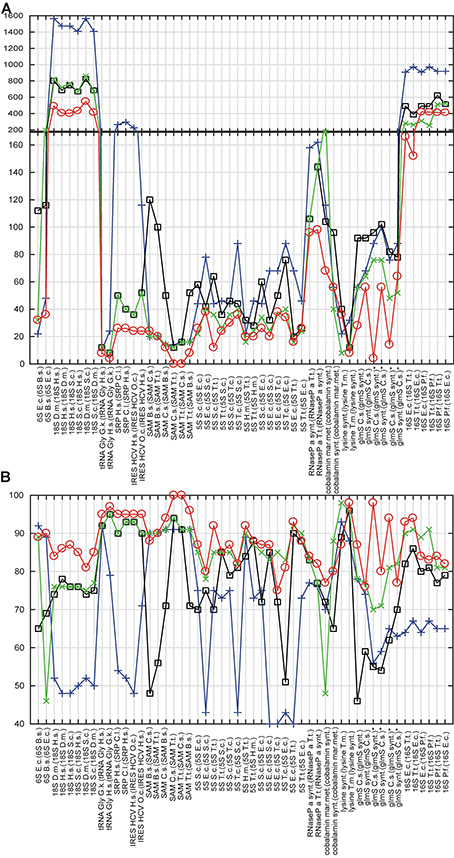
Figure 3. Cross-validation. In (A,B), x-axis shows RNAs, whose structure is predicted and (in parenthesis) the RNAs, whose structures were used either for constraints or as templates. Y-axis shows tree edit distances (A) and percentage of nucleotide positions with correctly predicted structural information (B). Circles, squares, crosses, and x's show values for the proposed method, the refold method, RNAfold and the Rsearch-based method, respectively. For (A), the lesser the distance, the higher the structural similarity to experimentally identified structure; 0 for identical structures. For (B), the maximum of structural similarity to experimentally identified structure is indicated by 100%. For predictions marked with *, three structural versions were obtained by removing pseudo knots. Organism names are abbreviated: E.c., E. coli; H.m., H. marismortui; B.s., B. subtilis; D.m., D. melanogaster; H.s., H. sapiens; O.c., O. cuniculus; T.t., T. tencogensis; C.s., C. subterraneus; P.f., P. falciparum; T.m., T. maritime; S.c., S. coelicolor.
Figure 3 indicates that the proposed method was capable to generate more accurate structures than both classical prediction represented by RNAfold and the principally same methods represented by the refold method and the Rsearch–based method. The main improvement is the ability to generate both large structures of long sequences and structures with long single-strand segments that are notoriously hard to be predicted with available prediction methods. Such typical structures here are ribosomal RNAs and bacterial RNaseP. For some species of shorter and highly paired structures, as the E. coli and T. tencogensis 5S RNAs and lysine riboswitches, the proposed method, the refold method and the Rsearch–based method provided similar accuracy (Figures 3A,B).
With respect to the structure similarity metrics, both tree edit distance and percentage of nucleotide position with correctly predicted structural information, were similarly efficient, thus cross-validating each other. In the remainder of the evaluation, we use tree edit distance. The other metrics depends on the sequence alignment method. Note that the value of tree edit distance depends on the size of structures. As it is a distance, the higher the similarity, the lower the score, and the value of zero indicates structural identity.
The bootstrap procedure for evaluation of the reliability of the generated structure and its metrics (see section Materials and Methods) was validated using the cross-validation dataset, i.e., experimentally identified structures. The FE-based z-scores obtained by the repeated bootstrap (100 runs with 100 randomized sequences each) evaluated 18 of total 52 generated structures as unreliable. Nevertheless, 15 of these 18 unreliable structures were false negatives (FNs). An example of a FN is shown in Figure 4. It is the secondary structure of C. subterraneus glmS ribozyme generated using synthetic glmS ribozyme as the template. The generated structure is obviously accurate (cf. Figures 4B,D), but its ze = 1.85, marking it as unreliable (ze < 2). For comparison, we generated true negative (TN) structure of the same sequence using the proposed method with different values of inconsistency thresholds (30% for hairpins and 20% for stems; Figure 4C). Its ze = 2.7, i.e., evaluated as reliable (>2), though it is obviously unreliable (cf. Figures 4B,C) and therefore a false positive. In spite of the z-scores, the accuracy of both the structures was documented well by their tree edit distances (d = 4 and d = 44, respectively) to their experimentally identified counterpart. Analogous situations occurred for the other 15 generated structures that were evaluated as unreliable by the FE-based z-scores.
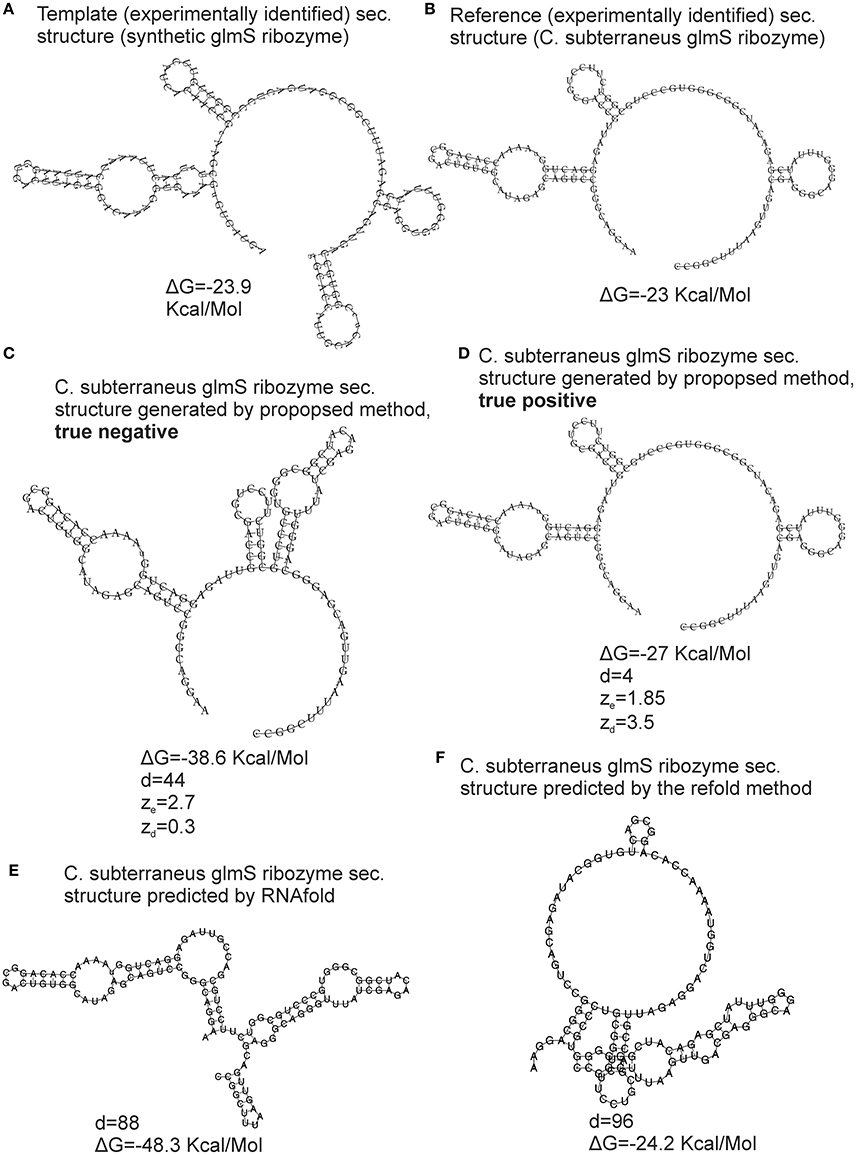
Figure 4. Validation of the structural similarity metrics. An example of glmS ribozyme is used for demonstration. The template was the synthetic glmS ribozyme with secondary structure derived from PDB ID 3l3c (A), the reference RNA was C. subterraneus glmS ribozyme with secondary structure derived from PDB ID 3b4c (B). True negative and true positive structures generated by the proposed method using different values of parameters are shown in (C,D), respectively. The structures predicted by RNAfold and the refold method are shown in (E,F), respectively. The proposed method and the refold method used the secondary structure of synthetic glmS ribozyme as template and constraint, respectively. ΔG—free energy, d—tree edit distance, ze and zd are z-scores based on FE and tree edit distances, respectively.
We therefore used tree edit distance between generated structures and templates instead of FE. The distance-based z-scores evaluated 3 of 52 generated structures as unreliable with more than 50% of their z-scores obtained by the repeated bootstrap (100 runs with 100 randomized sequences each) <2. Reliability of the remaining structures with their zd > 2 is demonstrated by the example in Figure 4. The z-scores were zd = 3.5 and 0.3 for the accurate and inaccurate structures, respectively (Figures 4C,D, respectively). Such z-scores better corresponded to the reliability of the structures.
Our z-scores are however not definitive proofs of quality of the structure and zd > 2 should be interpreted only as with a high likelihood the structure is reasonable. The main reason is the z-scores's variance with respect to the randomized structures used in the bootstrap. The variance was estimated using the repeated bootstrap (100 runs with 100 randomized sequences each) for the 52 generated structures of the cross-validation dataset and counting how many z-scores were <2 and >2 for each generated structure. Besides the 3 unreliable structures, 39 of total 52 structures had 100% z-scores >2. Remaining 10 structures had their z-scores >2 from 90.6% in average (for individual values, see the black curve in Figure S1).
The reason why FE was inadequate for our task was most likely its position independency. Two dissimilar structures with similar base pairs, though at different position on a sequence, can have similar FEs. As a result, an inaccurate structure can have correct FE, as demonstrated by the example in Figure 4. This fact is further demonstrated by the structure predicted by the refold method (Figure 4F) that is relatively dissimilar to its experimentally identified counterpart (Figure 4B). Nevertheless, the difference in FE between the predicted and experimentally identified structure is 1.2 Kcal/Molecule (−23 to −24.2 Kcal/Mol). The structure generated by the proposed method (Figure 4D) is fairly similar to the experimentally identified structure, but the difference in FE is 3.8 Kcal/Mol (−23 to −27 Kcal/Mol), i.e., higher, indicating stronger dissimilarity than for the structure predicted by the refold method.
As shown above, tree edit distance is biologically more relevant for comparison than free energy. In the following section we thus treat tree edit distance as our primary metric.
Results of the large-scale evaluation are summarized by Table S5 and Figure 5 and Figure S2. They document higher accuracy of the presented method when compared to RNAfold, the refold-based method and the Rsearch-based method. In the first case, the higher accuracy was achieved due to the extra information used by the presented method. For the latter two methods, that use the same input information, the higher accuracy was due to the active search for inconsistent structural elements and correction of their structure.
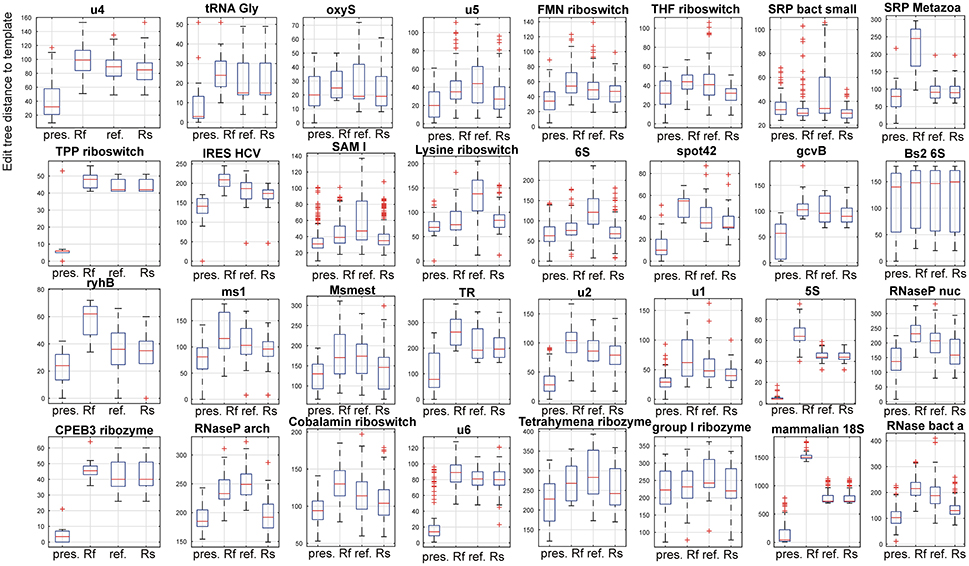
Figure 5. Comparison of the presented method. The compared methods were RNAfold as a representative of classical, single sequence secondary RNA structure prediction, a refold.pl-based method and the Rsearch method that both allow for the principally same type of prediction as the presented method. In the figure, 32 panels show results for 32 families of the reference dataset. In each panel, four box plots for the presented method, RNAfold, the refold method and the Rsearch-based method, are shown (x-axis). Individual box plots show the median (red line), the 25th and 75th percentiles (the tops and bottoms of the boxes, respectively) and outliers (the whiskers) of the edit tree distances of the predicted structures of a single family to templates. The distances between the tops and bottoms of the boxes are the interquartile ranges. The families are indicated by titles of the plots.
For 3 families (SRP bact small, Bs2 6S and group I ribozyme) the compared methods performed nearly the same. These families include densely and unambiguously paired structures that are convenient for the classical prediction, represented by RNAfold.
In the following, the presented method is demonstrated in details using selected RNAs from the reference dataset. The examples are intended to illustrate situations when the proposed method is advantageous. Additional state-of-the-art prediction algorithms, principally different from the proposed method, were included in this demonstration to cover a broader spectrum of available prediction methods.
The first example is gcvB RNA, whose structure, experimentally identified in Sharma et al. (2007) for S. typhimurium (Figure 6A), is difficult to predict as it includes relatively long single-strand segments. For this example we predict structures of gcvB homologs identified in the above cited paper. The sequence and structure of the template and the sequences of gcvB homologs and their predicted/generated structures are in Supplementary File S3.fasta in sections (a) and (b), respectively.
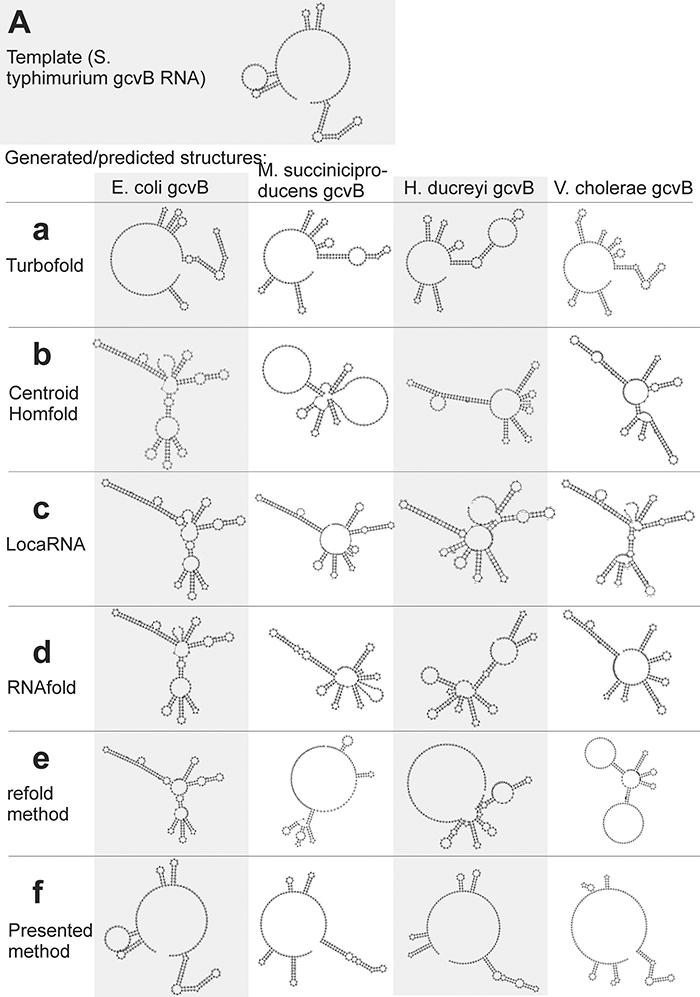
Figure 6. Individual secondary structures of gcvB RNA homologs predicted by available methods (a–e) and generated by the presented method (f). Structures are organized in rows and columns according to the method and species, respectively. The experimentally identified template structure of S. typhimurium gcvB RNA structure is shown at the top (A).
RNAfold and CentroidHomfold (used for single sequence prediction) tended to pair the sequences of long single-strand segments (Figures 6b,d). More accurate was Turbofold thanks to all the sequences of the homologs used as input (Figure 6a). Locarna and the refold.pl-based method that used the experimentally identified structure as constraint did not predict plausible structures of homologs (Figures 6c,e). Tree edit distances that quantify the similarity of the generated/predicted structures are shown in Table S6.
In contrast, the presented method was capable to generate structures that were similar to the experimentally identified structure (cf. Figures 6A,f). The similarity is measured by tree edit distances (see Table S6). It was for two reasons: (i) the wrong, excessive pairing was prevented by the information of single-stranded segments copied from the template. This made the proposed method more accurate than the single-sequence prediction methods. (ii) The proposed method actively searched for inconsistent structural elements after copy step and predicted their structure de novo. This made it more accurate than the other methods that use the same information as input. Comparison of the accuracy using tree edit distances is in Table S6.
The improved accuracy can help to recognize non-homologous sequences. It is demonstrated here with the sequence of E. coli gcvB with randomly shuffled dinucleotides. Structure of this shuffled sequence, which represented an RNA not homologous to gcvB, could be distinguished from the gcvB homologs by its edit tree distance to the experimentally identified gcvB structure, when generated by the proposed method. The distance was twice longer than those of the structures of the gcvB homologs (Table S6). Recognition of this non-homologous RNA was not clear by the available methods as its tree edit distance was not unambiguously higher than the distances of the gcvB homologs (Table S6). The predicted structures of the shuffled RNA are in Supplementary File S4.fasta.
The non-homologous RNA with the shuffled sequence could also be recognized by z-score of its generated structure. It was −3.6, which indicated strong unreliability. In contrast, z-scores for the gcvB homologs were all higher than 2 (namely 9.7, 3.6, 2.9, and 4.4 for E. coli, V. cholera, H. ducreyi, and M. succiniciproducens, respectively). The usefulness of z-scores of the generated structures is further demonstrated in the next example.
Large structures with long sequences are another class of sequences, when the classical prediction is often inaccurate. This is demonstrated here by the structure of mammalian 18S rRNAs. The methods of classical prediction that use either a single input sequence (RNAfold) or multiple input sequences (CentroidHomfold, Turbofold), and also methods that use a homologous experimentally identified structure (H. sapiens 18S rRNA) as a constraint (Locarna and the refold method) were largely inaccurate. This is demonstrated visually by Figures 7b,c (for RNAfold and the refold method only from technical reasons due to the large size of the 18S rRNA structures). The predicted structures were included in Supplementary File S5.fasta.
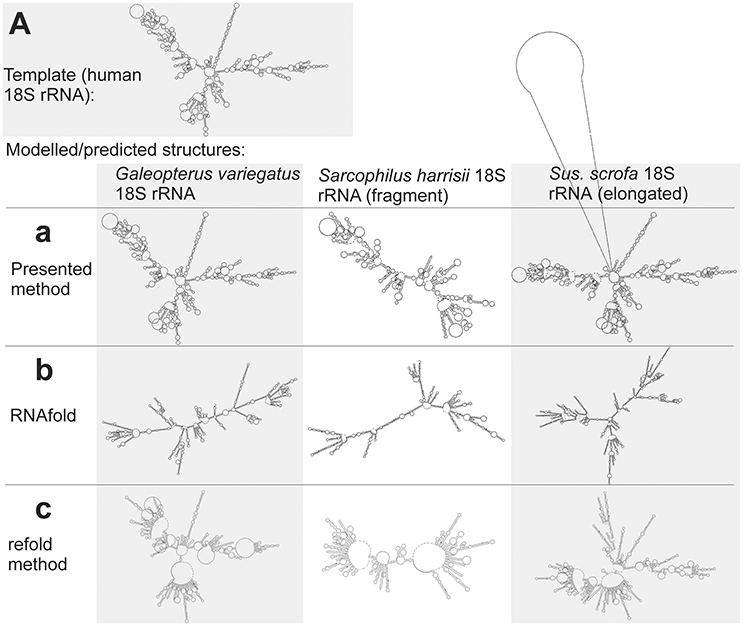
Figure 7. Individual secondary structures of 18S rRNA homologs predicted by available methods (b,c) and generated by the presented method (a). Structures are organized in rows and columns according to the method and species, respectively. The experimentally identified template structure of H. sapiens 18S rRNA structure is shown at the top (A).
The presented method was more accurate as its accuracy is largely independent of sequence length (Figure 7a). The improved accuracy was demonstrated by shorter tree edit distances of the generated structures to the experimental identified template of H. sapiens 18S rRNA (Table S7).
Interesting is the identification of the 18S rRNA fragment of S. harrisii 18S rRNA and the elongated 18s sequence of S. scrofa 18S rRNA. The proposed method identified correctly that the S. harrisii fragment contained only the expansion segments 3 and 6 of the whole 18S rRNA structure (Figure 7a). In contrast, the S. scrofa sequence included, beside the entire 18S rRNA structure, an additional ~700 nucleotides flanking the regular 18S rRNA structure (Figure 7a). Tree edit distances of these two structures were relatively high, when compared to the G. variegatus 18S rRNA structure that is complete (Table S7). This was due to the natural dissimilarity of either structural fragments or elongated structures to regular structures. However, z-scores were far greater than 2 (6.6, 12.1, and 19.2 for S. harrisii, S. scrofa, and G. variegatus, respectively) indicating that these sequences are genuine 18S rRNAs, yet fragmented/elongated.
An interesting experiment and also validation of the above identification was to use an RNA that was non-homologous to both the fragment and the elongated sequence. To that end, we deployed E. coli 16S rRNA as a template. Now the template and the query sequences were no longer homologous, yet still related (all were rRNAs), and the z-scores should indicate unreliability of the generated structures. Indeed, the z-scores were −0.2, −6, and 0.2 for S. harrisii, S. scrofa, and G. variegatus, respectively, indicating that the query sequences were not homologous to the 16S rRNA. In general, this procedure makes it possible to recognize, when the template and query sequences are not homologous, in other words, when the transfer of the template structure is biologically irrelevant producing wrong structures. What is important is that this bootstrap-based procedure is independent of the fact that query sequences are fragmented or elongated, as demonstrated in both this example and the previous example.
A method for template–based prediction/generation of single-sequence secondary RNA structure is presented. As demonstrated on examples, it is useful for determining whether an RNA molecule under investigation can conform to a secondary structure taken from a different molecule. This is useful for both obtaining RNA secondary structures and estimating ability of sequences to adopt the investigated structure. The method provides a solution in situations when available methods for secondary RNA structure prediction cannot be used or are inaccurate. It is applicable to all RNAs that adopt a secondary structure (Gorodkin and Ruzzo, 2014).
It does not mean that a new de novo RNA secondary structure prediction algorithm/method is devised. It should be stressed that the presented method requires sequences and structures as input in contrast to the prediction methods that usually work only with sequences. However, when experimentally derived structures of homologous sequences are available, our method is able to correctly predict true biological structures, as shown by our cross validation study. We have also performed a large scale comparison of the results provided by our method for sequences where the ground truth is unknown, where our method also performed favorably. The large scale comparison is not used to benchmark the method, but merely for establishing a basis for evaluating the efficiency of the presented method.
The quality of structures generated by our method depends on the choice of the template. There are no strict guidelines for choosing the template—in principle, any RNA secondary structure either experimentally identified or predicted can be used as a template. It is expected that the user leverages their expertise to find a template that is homologous or contains homologous subsequences. Naturally, reliability of the generated structure depends on the biological similarity between the template and the query. The z-score produced by the bootstrapping step is a proxy for this similarity and low z-scores should reveal situations when the template was not chosen appropriately.
Our method fills a gap resulting from the poor performance of the available methods of constrained prediction using known structures. Such prediction is increasingly useful as number of experimentally identified RNA structures (and thus the number of available templates) grows. The presented method is useful in various situations as demonstrated in this work. The method does not depend on length of sequences neither on the type of structure. It is fast and robust and it can be used for characterization of large numbers of sequences including fragments by structures of other RNAs. It produces z-scores based on bootstrapping of generated secondary structures that indicate whether the generated structures are relevant for the sequences.
JP conceived, designed, coded, and tested the method, and wrote the manuscript. MC designed the test of the method and participated on writing the manuscript. MS carried out the comparison to RSearch/infernal.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The work was supported by the Grant Agency of the Czech Republic [GA15-00885S] and ELIXIR CZ research infrastructure project (MEYS Grant No: LM2015047).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2017.00147/full#supplementary-material
Andersen, E. S., Rosenblad, M. A., Larsen, N., Westergaard, J. C., Burks, J., Wower, I. K., et al. (2006). The tmRDB and SRPDB resources. Nucleic Acids Res. 34, D163–D168. doi: 10.1093/nar/gkj142
Darty, K., Denise, A., and Ponty, Y. (2009). VARNA: interactive drawing and editing of the RNA secondary structure. Bioinformatics 25, 1974–1975. doi: 10.1093/bioinformatics/btp250
Gardner, P. P., Daub, J., Tate, J. G., Nawrocki, E. P., Kolbe, D. L., Lindgreen, S., et al. (2009). Rfam: updates to the RNA families database. Nucleic Acids Res. 37, D136–D140. doi: 10.1093/nar/gkn766
Gardner, P. P., and Giegerich, R. (2004). A comprehensive comparison of comparative RNA structure prediction approaches. BMC Bioinformatics 5:140. doi: 10.1186/1471-2105-5-140
Gorodkin, J., and Ruzzo, W. L. (2014). RNA Sequence, Structure, and Function Computational and Bioinformatic Methods. New York, NY: Humana Press.
Gutell, R. R., Lee, J. C., and Cannone, J. J. (2002). The accuracy of ribosomal RNA comparative structure models. Curr. Opin. Struct. Biol. 12, 301–310. doi: 10.1016/S0959-440X(02)00339-1
Hamada, M., Sato, K., Kiryu, H., Mituyama, T., and Asai, K. (2009). Predictions of RNA secondary structure by combining homologous sequence information. Bioinformatics 25, i330–i338. doi: 10.1093/bioinformatics/btp228
Hofacker, L. (2004). RNA secondary structure analysis using the Vienna RNA package. Curr. Protoc. Bioinformatics Chapter 12, Unit 12 2. doi: 10.1002/0471250953.bi1202s04
Hofacker, I. L., Bernhart, S. H., and Stadler, P. F. (2004). Alignment of RNA base pairing probability matrices. Bioinformatics 20, 2222–2227. doi: 10.1093/bioinformatics/bth229
Kertesz, M., Wan, Y., Mazor, E., Rinn, J. L., Nutter, R. C., Chang, H. Y., et al. (2010). Genome-wide measurement of RNA secondary structure in yeast. Nature 467, 103–107. doi: 10.1038/nature09322
Kiryu, H., Kin, T., and Asai, K. (2007). Robust prediction of consensus secondary structures using averaged base pairing probability matrices. Bioinformatics 23, 434–441. doi: 10.1093/bioinformatics/btl636
Lorenz, R., Bernhart, S. H., Honer Zu Siederdissen, C., Tafer, H., Flamm, C., and Stadler, P. F. (2011). ViennaRNA Package 2.0. Algorithms Mol. Biol. 6:26. doi: 10.1186/1748-7188-6-26
Loughrey, D., Watters, K. E., Settle, A. H., and Lucks, J. B. (2014). SHAPE-Seq 2.0: systematic optimization and extension of high-throughput chemical probing of RNA secondary structure with next generation sequencing. Nucleic Acids Res. 42:e165. doi: 10.1093/nar/gku909
Mathews, D. H. (2004). Using an RNA secondary structure partition function to determine confidence in base pairs predicted by free energy minimization. RNA 10, 1178–1190. doi: 10.1261/rna.7650904
Mathews, D. H. (2006). Revolutions in RNA secondary structure prediction. J. Mol. Biol. 359, 526–532. doi: 10.1016/j.jmb.2006.01.067
Nawrocki, E. P., and Eddy, S. R. (2013). Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 29, 2933–2935. doi: 10.1093/bioinformatics/btt509
Sharma, C. M., Darfeuille, F., Plantinga, T. H., and Vogel, J. (2007). A small RNA regulates multiple ABC transporter mRNAs by targeting C/A-rich elements inside and upstream of ribosome-binding sites. Genes Dev. 21, 2804–2817. doi: 10.1101/gad.447207
Smith, C., Heyne, S., Richter, A. S., Will, S., and Backofen, R. (2010). Freiburg RNA Tools: a web server integrating INTARNA, EXPARNA and LOCARNA. Nucleic Acids Res. 38, W373–W377. doi: 10.1093/nar/gkq316
Sussman, J. L., Lin, D., Jiang, J., Manning, N. O., Prilusky, J., Ritter, O., et al. (1998). Protein Data Bank (PDB): database of three-dimensional structural information of biological macromolecules. Acta Crystallogr. D Biol. Crystallogr. 54, 1078–1084. doi: 10.1107/S0907444998009378
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680.
Keywords: RNA, secondary structure, homology, prediction, template structure
Citation: Pánek J, Modrák M and Schwarz M (2017) An Algorithm for Template-Based Prediction of Secondary Structures of Individual RNA Sequences. Front. Genet. 8:147. doi: 10.3389/fgene.2017.00147
Received: 24 July 2017; Accepted: 25 September 2017;
Published: 10 October 2017.
Edited by:
Alessandro Laganà, Icahn School of Medicine at Mount Sinai, United StatesReviewed by:
Zhi-Ping Liu, Shandong University, ChinaCopyright © 2017 Pánek, Modrák and Schwarz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josef Pánek, cGFuZWtAYmlvbWVkLmNhcy5jeg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.