 Irene Valenti
Irene Valenti Luca Degradi
Luca Degradi Andrea Kunova
Andrea Kunova Paolo Cortesi
Paolo Cortesi Matias Pasquali
Matias Pasquali Marco Saracchi
Marco Saracchi
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Fungal Biol. , 09 February 2022
Sec. Fungal Genomics and Evolution
Volume 2 - 2021 | https://doi.org/10.3389/ffunb.2021.802511
This article is part of the Research Topic Mitochondrial Genomes and Mitochondrion Related Gene Insights to Fungal Evolution View all 11 articles
Ciborinia camelliae is the causal agent of camellia flower blight (CFB). It is a hemibiotrophic pathogen, inoperculate Discomycete of the family Sclerotiniaceae. It shows host and organ specificity infecting only flowers of species belonging to the genus Camellia, causing serious damage to the ornamental component of the plant. In this work, the first mitochondrial genome of Ciborinia camellia is reported. The mitogenome was obtained by combining Illumina short read and Nanopore long read technology. To resolve repetitive elements, specific primers were designed and used for Sanger sequencing. The manually curated mitochondrial DNA (mtDNA) of the Italian strain DSM 112729 is a circular sequence of 114,660 bp, with 29.6% of GC content. It contains two ribosomal RNA genes, 33 transfer RNAs, one RNase P gene, and 62 protein-coding genes. The latter include one gene coding for a ribosomal protein (rps3) and the 14 typical proteins involved in the oxidative metabolism. Moreover, a partial mtDNA assembled from a contig list was obtained from the deposited genome assembly of a New Zealand strain of C. camelliae. The present study contributes to understanding the mitogenome arrangement and the evolution of this phytopathogenic fungus in comparison to other Sclerotiniaceae species and confirms the usefulness of mitochondrial analysis to define phylogenetic positioning of this newly sequenced species.
The genus Ciborinia involves 23 different species of host-specific pathogens (https://eol.org/). Ciborinia spp. are members of the Sclerotiniaceae family (order Helotiales; phylum Ascomycota), which includes 14 genera of phytopathogenic fungi with a wide host range. The Sclerotiniaceae's ability to infect different hosts and adapt to various environments results in serious economic damage (Bolton et al., 2006). Unlike the other Sclerotiniaceae, Ciborinia camelliae Kohn infects only plants of the genus Camellia causing the camellia flower blight (CFB) (Taylor and Long, 2000; Saracchi et al., 2019). Sclerotia in fallen flowers lie dormant on the ground or in plant debris over summer, autumn, and winter. Toward the end of winter, the sclerotia begin to germinate, producing apothecia from which ascospores are released. The pathogen infects and colonizes only flowers, where brown spots appear and spread to the entire organ. Flowers tend to fall prematurely. This causes important damage to the camellia floriculture industry (Taylor and Long, 2000; Denton-Giles et al., 2013).
Studies on the diversity within the genus Ciborinia are scarce (Kohn, 1979). The C. camelliae variability was investigated using the UP-PCR analysis on strains from America and New Zealand, demonstrating a relatively low level of genetic variations within the two populations (van Toor et al., 2005). On the other hand, a recent study based on morpho-cultural characterization and ITS analysis of numerous Italian strains showed significant variability among the isolates (Saracchi et al., 2022).
The mitochondria are membrane-enclosed compartments that play a central role in providing energy to eukaryotic cells by oxidative phosphorylation (Newmeyer and Ferguson-Miller, 2003; Kriváková et al., 2005; Lv et al., 2017). They are also involved in the signal amplification leading to apoptosis (Goodsell, 2010), antifungal drug resistance as well as in virulence and pathogenicity (Shingu-Vazquez and Traven, 2011; Sandor et al., 2018; Kulik et al., 2020). The study of mitochondrial (mt) genomes reveals interesting features about the pathogen evolution and its relationships with the other related species (Ballard and Whitlock, 2004; Aguileta et al., 2014; Mardanov et al., 2014; Kulik et al., 2020). Even across distantly connected species, the mt genes are largely conserved due to their essential role in cell vitality (Medina et al., 2020). Nevertheless, intron numbers and secondary structures are highly variable and are prone to evolving rapidly. All these features make mitogenome a functional source for phylogenetic studies (Hamari et al., 2002; Burger et al., 2003; Galtier et al., 2009). The mitochondrial genome is often a circular double-stranded molecule with a condensed gene arrangement (Burger et al., 2003). Complete fungal mitogenomes vary notably in size. Typically, the fungal mitochondrial genome contains 14 conserved protein-coding genes involved in the oxidative metabolism (atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, and nad6) and one ribosomal protein S3 gene (rps3). Additionally, each mtDNA has two ribosomal RNA (rRNA) genes for the large and small rRNA subunit (rnl and rns) and a variable number of tRNAs (Bullerwell and Lang, 2005).
In this study, we present a comprehensive analysis of the first C. camelliae mitochondrial DNA. The complete sequence of an Italian strain was obtained with a hybrid strategy combining Illumina Hiseq paired reads and MinIon Nanopore long reads sequencing. The mitochondrial sequences were identified and assembled together constructing the complete and circular mitogenome. The results were compared with data from the available Sclerotiniaceae mitogenomes. This study allows to position the C. camelliae pathogen in the Sclerotiniaceae family and offers some insight into the specificities of mtDNA of C. camelliae.
The investigated strain was isolated from pieces of sclerotium collected in Oggebbio (Verbania, Italy) (45°59′47.5782″ N, 08°39′05.9659″ E) and showing C. camelliae characteristics. The pathogen was grown on Potato Dextrose Agar medium (PDA: 800 mL/L of potato extract; 20 g/L glucose, BioFROXX, Germany; 15 g/L agar, Applichem, Germany). The isolate was identified according to colony morphology and microscopic analysis of conidia and ascospore traits, as well as ITS sequence (Saracchi et al., 2022). The strain ITAC2 is maintained in the laboratory of Plant Pathology at the Department of Food, Environmental and Nutritional Sciences (University of Milan, Italy), and it is also deposited in a public collection (DSMZ-German Collection of Microorganisms and Cell Cultures GmbH) with the accession number DSM 112729.
High-molecular-weight genomic DNA was extracted from conidia (109 conidia/mL) using DNeasy PowerSoil Pro Kit (Qiagen, Hilden, Germany). DNA was quantified using the Qbit Fluorometer (Invitrogen, Thermo Fisher Scientific—USA) with the Qbit® dsDNA HS Assay Kit (Invitrogen, Thermo Fisher Scientific—USA). The genome quality was checked by spectrophotometer and 1% agarose gel electrophoresis. The gel was visualized using UV-transilluminator Gel Doc 2000 (BIO-RAD laboratories, USA) and Quantity One software (BIO-RAD laboratories, USA) to verify possible smearing of the DNA band.
Two different platforms were used to sequence the genomic DNA: Oxford Nanopore Technologies (ONT) and Illumina Hiseq, executed by Eurofins Genomics (Ebersberg, Germany). The ONT sequencing was performed by the MinION system (FLO-MIN-106 R9.4 flow-cell) using EXP-NBD104, EXPNBD114 in conjunction with the SQK-LSK109 kit and also SQK-RAD004 sequencing kit. A complete mitochondrial genome was obtained by combining long Nanopore reads and short Illumina reads. Sequencing data were analyzed using the European Galaxy web platform (https://usegalaxy.eu/) tools (Afgan et al., 2018) and Geneious Prime software, version 2021.1.1 (Biomatters, Auckland, New Zealand).
The whole genome assembly was performed using Flye v.2.8.3+galaxy0 (Lin et al., 2016) with a default setting. Medaka tool v.1.3.2 +galaxy0 was employed for autopolishing, and mapping of short reads on the draft assembly was performed by minimap2 on the same platform (Li, 2018). The final assembly was obtained using Pilon v.1.20.1 (Walker et al., 2014).
Contigs including the mitochondrial DNA were identified within the whole genome assembly using NCBI data as reference, and BLAST or minimap2 (Li, 2018) as a tool. The detected sequences were extracted and assembled using Geneious Prime software. Single uncalled bases (Ns) and small gaps, resulting from the genome assembly, were fixed by mapping raw reads on the draft assembly and performing manual corrections. Ns and gaps were replaced with the matching bases from illumina reads. Re-mapping was performed to validate the corrections. The mitogenome was circularized by searching the terminal sequences using the minimap2 tool.
After the assembly, the mitochondrial DNA exhibited some unsure repeated regions and sets of uncalled bases (Ns). These mismatches were solved by PCR analysis, using newly designed primers. Primer3web (Untergasser et al., 2012) program was used to design and validate our primers. The new primer pairs were synthesized by Eurofins Genomics (Srl Vimodrone, Italy) (Table 1). The primer pairs 1–2 and 6–7 were used to validate the same sequence amplifying regions with different lengths. The Primer3 was paired with both Primer4 and Primer5. Two PCR rounds were conducted. The first round, using primers 1–2, was performed with 32 cycles in a total 25 μL reaction volume containing: 12.5 μL of Q5® Hot Start High-Fidelity 2X Master Mix (Biolabs, New England), 0.5 μM of forward and reverse primer, 1 μL of DNA sample and the remaining volume of sterile distilled water. The PCR program involved the initial denaturation at 98°C for 2′, 32 cycles of denaturation at 98°C for 20″, annealing at 66°C for 20″, extension at 72°C for 2.5′, followed by the final extension at 72°C for 7′. The second PCR round, using the primer pairs 3–4, 3–5 and 6–7, was performed in a total volume of 30 μL which contained: 0.18 μL of GoTaq® DNA Polymerase 5 U/μL (Promega, Madison, WI, USA), 6 μL of GoTaq® Reaction Buffer 5X (Promega, Madison, WI, USA), 1.2 μL of 10 mM dNTP (Promega, Madison, WI, USA), 1.5 μL of 50 μM forward primer, 1.5 μL of 50 μM reverse primer, 18.62 μL of sterile distilled water and 1 μL of DNA sample. The PCR reaction was carried out with the following parameters: 94°C for 2′; 32 cycles at 95°C for 30″, 55°C for 20″ and 72°C for 45″; 5′ at 72°C. Thermal Cyclers (iCycler-BIO-RAD laboratories, USA and VeritiProTM96-Well Thermal Cycler, Applied Biosystems by Thermo Fisher Scientific) were used to amplify the DNA regions. The reaction products were visualized by electrophoresis on a 1% agarose gel containing ethidium bromide. All PCR products were sequenced using the Sanger technology with the same primers (Eurofins Genomics, Ebersberg, Germany). The sequencing data were analyzed by Geneious Prime software. These sequences were compared to the draft of mitochondrial DNA.
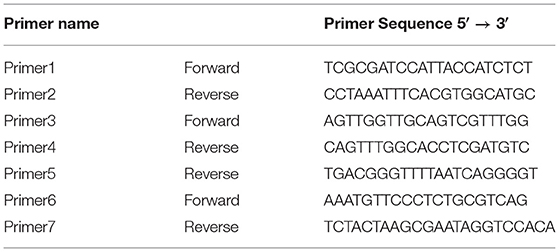
Table 1. Primer sequences table.
The mitochondrial genes were annotated using MFannot (http://megasun.bch.umontreal.ca/cgi-bin/mfannot/mfannotInterface.pl) (Beck and Lang, 2010) and RNAweasel (https://megasun.bch.umontreal.ca/cgi-bin/RNAweasel/RNAweaselInterface.pl) (Lang et al., 2007) tools and MITOS WebServer (http://mitos.bioinf.uni-leipzig.de/index.py) (Bernt et al., 2013). Manual corrections were required. To verify the annotation results, sequences and encoded proteins were compared with related species using BLASTp searches against the NCBI database. For each open reading frame (ORF), the protein with the highest similarity was found among biological sequences in the NCBI database. These results were further verified by BLASTn analysis.
The mitogenome features were compared among some closely related species in the Sclerotiniaceae family, order Helotiales (Botryotinia fuckeliana (KC 832409), Ciboria shiraiana (CM 017871.1), C. camelliae (GCA_001247705.1), Monilinia fructicola (NC_056195.1), Monilinia laxa (NC_051483.1), Monilinia polystroma (GCA_002909645.1), Sclerotinia borealis (KJ434027) and Sclerotinia sclerotiorum (KT283062). Glarea lozoyensis (Order Helotiales; Family Helotiaceae) was used as an outgroup. For all investigated species, data were downloaded from the NCBI database. In species with no mitochondrial genome available, we used BLAST searches to identify the mtDNA in the total genome assembly and subsequently, each detected mitochondrial sequence was annotated according to the procedure described earlier. When possible, these additional results were used in comparative analyses. Based on data availability, seven Sclerotiniaceae species (Botryotinia fuckeliana, Ciboria shiraiana, C. camelliae strain ITAC2, M. fructicola, M. laxa, S. borealis, and S. sclerotiorum) were investigated in terms of gene arrangement. Monilinia polystroma and C. camelliae ICMP 19812 strain were excluded from this analysis due to the lack of the whole mitochondrial sequence. Multiple mitogenome alignment was performed within the previously cited Sclerotiniaceae using the MAUVE tool of Geneious Prime software and setting nad4L gene as starting point. MAUVE alignment identifies homologous regions shared by two or more mitogenomes. These regions are denominated locally collinear blocks (LCBs) (Darling et al., 2004). Tandem Repeat Finder was employed to identify tandem repeats within the mitochondrial genomes (Benson, 1999). The codon usage was estimated using both Sequence Manipulation Suite webserver (Stothard, 2000) and cusp tool of the European Galaxy web platform (https://galaxy-iuc.github.io/emboss-5.0-docs/cusp.html).
The phylogenetic study was performed on the multiple alignments of 14 concatenated mitochondrial proteins using amino acid sequences obtained from each mtDNA. A maximum-likelihood phylogenetic tree was executed by IQ-TREE (Trifinopoulos et al., 2016) web server selecting default setting, and performed by MEGAX software (Kumar et al., 2018).
The circular mitochondrial genome of C. camelliae strain ITAC2, submitted to the GenBank database with the accession number OK326902, has a total length of 114,660 bp. The nucleotide composition is the following: 34.7% of A, 13.2% of C, 16.5% of G, 35.7% of T with a GC content of 29.6%.
This mitogenome contains all 14 typical genes encoding the subunits of ATP synthase (atp6, atp8, and atp9), NADH dehydrogenase (nad1, nad2, nad3, nad4, nad4L, nad5, and nad6), apocytochrome b (cob), and cytochrome c oxidase (cox1, cox2, and cox3). Additionally, the conserved ribosomal protein-coding gene S3 (rps3), untranslated genes of the large and small ribosomal RNAs (rnl and rns), the ribonuclease P RNA (rnpB) gene, and 33 transfer RNAs (tRNA) genes were detected. Overall, the RNA region accounted for 9.26% of the whole mitochondrial genome. The length of individual tRNAs ranged from 69 to 85 bp and most of these genes were placed around the rnl region (Figure 1). The total tRNAs are related to 18 essential amino acids.
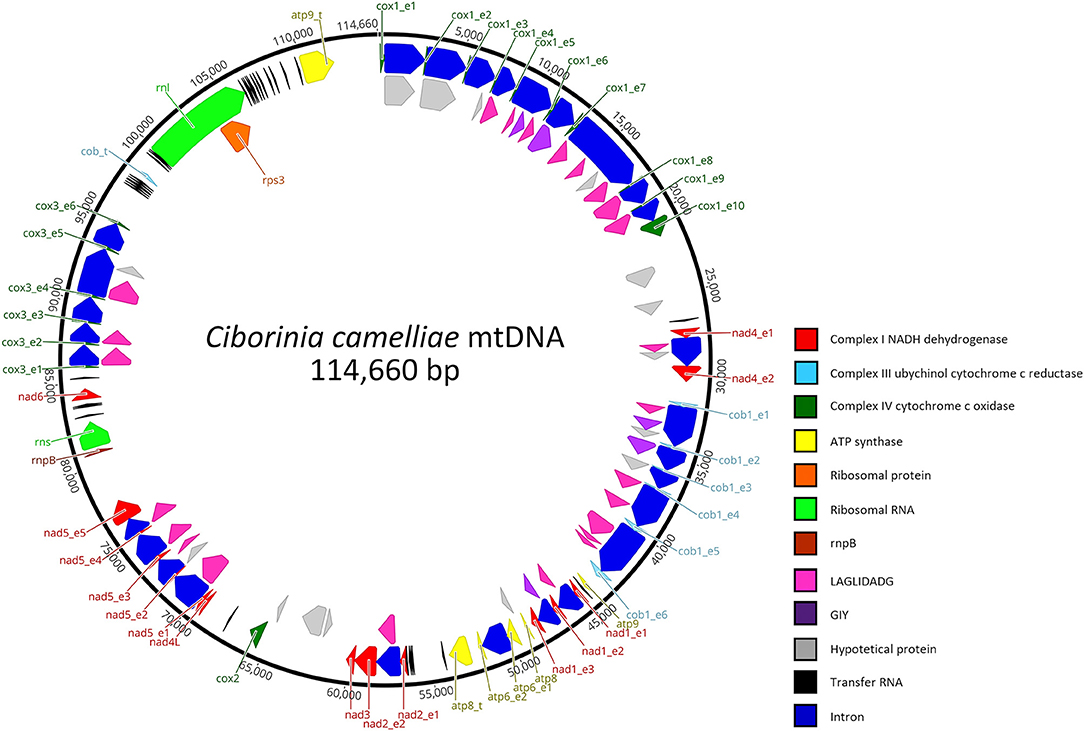
Figure 1. Genetic organization of Ciborinia camelliae mitogenome. Circular map is generated by Geneious Prime software, version 2021.1.1 (Biomatters, Auckland, New Zealand). Genes are represented by different colored blocks.
In total, the mitogenome contains 62 putative genes, including 44 open reading frames (ORFs) for hypothetical proteins (15) and homing endonucleases of the LAGLIDADG (24) and GIY-YIG (5) families. BLASTp analysis was conducted for these non-conserved ORFs. The results (Supplementary Data Sheet 1), also verified by BLASTn analysis indicate that 22 ORFs have high identities with mitochondrial ORFs of the genus Monilinia. On the other hand, a high similarity with the Sclerotiniaceae species was not observed in 14 % of the ORFs. These exhibit the highest amino acid identity (ranging from 63.41 to 90.75%) with fungal species belonging to the family of Nectriaceae, Ceratocystidaceae, Cryphonectriaceae, Erysiphaceae, and Orbiliaceae. The last two are respectively evolutionary the closest and the most distant from the Sclerotiniaceae family. A LAGLIDADG of Golovinomyces cichoracearum and one of Fusarium tricinctum are the most representative ORFs with the highest amino acid identity: 85 and 90.75% with 100 and 92 % of coverage, respectively.
The coding sequences (CDS) of the 14 core mitochondrial proteins and rps3 were used in the codon usage analysis. This investigation was carried out among the selected Sclerotiniaceae species and F. oxysporum as external control (Supplementary Data Sheet 2). The results did not show significant variability in the use of the codons. All the investigated species showed the same preferential codons, encoding the same amino acid, except for Valine and Threonine (Table 2). The most used codons for Threonine are ACA and ACT. In C. camelliae strain ITAC2, S. sclerotiorum, M. laxa, C. shiraiana, and B. fuckeliana the preferential codon is ACA, while in M. fructicola, S. borealis, and G. lozoyensis is ACT. The preferential codon for Valine is GTT for all investigated species, except Monilia fructicola, which uses more frequently GTA. Nevertheless, the GTA codon represents the second choice to encode Valine also in the other analyzed species.
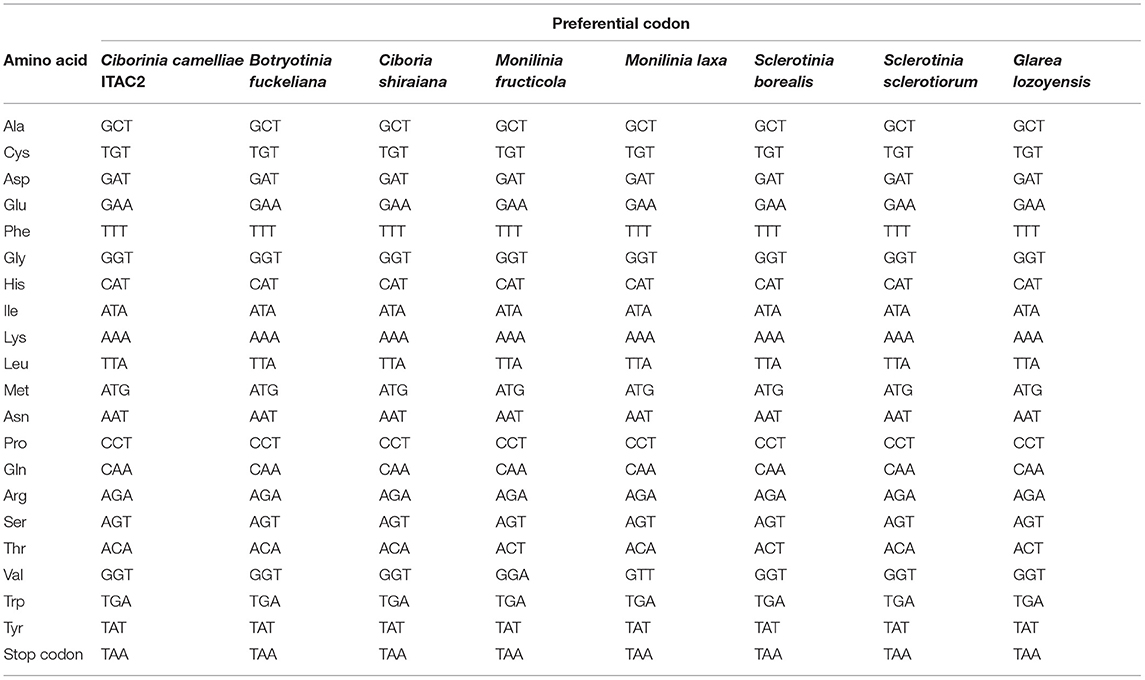
Table 2. Codon usage patterns for the 14 core mitochondrial protein coding-genes and rps3 in Sclerotiniaceae species (Ciborinia camelliae strain ITAC2, Botryotinia fuckeliana, Ciboria shiraiana, Monilinia fructicola, Monilinia laxa, Sclerotinia borealis, Sclerotinia sclerotiorum) and Glarea lozoyensis.
Focusing on gene-specific codon usage, only in the nad2 gene do all the investigated Sclerotiniaceae use the same preferential codons, suggesting that the nad2 is the most conserved gene, followed by the atp8 and rps3 genes showing differences in only two amino acids. On the other hand, cox1, atp9, nad3, cob, and nad4L are the genes with the greatest differences in codons usage. They exhibited dissimilarity from 9 to 12 out of 21 amino acids (Supplementary Data Sheet 3).
Only for the C. camelliae ITAC2 strain, the codon usage of non-conserved ORFs was investigated and compared with that of conserved genes. The ORFs and the conserved regions exhibited a similar codon usage, excluding Isoleucine (ATA in conserved genes, ATT in ORFs) and Threonine amino acids (ACA and ACT in conserved regions and ORFs, respectively) (Supplementary Data Sheet 4).
The mitochondrial genome size is associated with a high content of introns (Deng et al., 2018). In C. camelliae mtDNA we detected 28 introns, which are located in eight out of 14 genes: atp6 (1), cob (5), cox1 (9), cox3 (5), nad1 (2), nad2 (1), nad4 (1) and nad5 (4) (Table 3). Introns exhibit a variable length ranging from 1,015 bp to 4,750 bp. The cox3, cox1, and cob genes showed the highest fraction of intronic sequences: 91.2, 90.6, and 90.1%, respectively. For each gene containing introns, the exon protein-coding sequence (CDS) was <54%. In the 14 mitochondrial genes, the portion of non-coding DNA ranged from 0 to 68% (Table 3). The total content of non-coding DNA in the ITAC2 strain mitogenome is ~48%. The sequences encoding the fourteen core mitochondrial proteins and rps3 protein represent only 13.3% of the entire mitogenome.
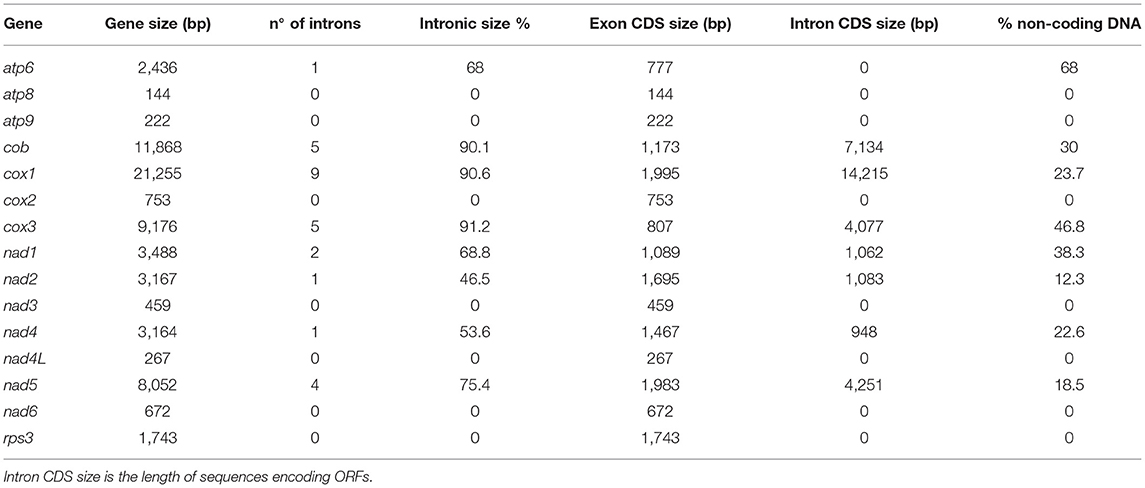
Table 3. List of the 15 mt genes with their: total length, introns, protein coding sequence (CDS) size and the portion of non-coding DNA.
The mtDNA of C. camelliae contains incomplete duplicated copies of atp8, atp9, and cob genes. All of these extra copies appeared truncated. The atp8-like ORF is located between atp6 and nad2 genes. The first 33 amino acids (aa) exhibit 75 % of similarity with the atp8 protein. Instead, no significant result was found for the remaining 385 aa. The atp9 copy is placed between cox1 and rnl region. The first 59 amino acids show 100% identity with the atp9 protein. This similarity decreases for what concerns the rest of the protein. The cob-like ORF was detected between cox3 and rnl region and consists of only 68 aa with an identity of 80% compared to the cob gene. All three extra copies were observed also in C. camelliae ICMP 19812 strain from New Zealand (Supplementary Data Sheet 5). The extra genes similarity between the two C. camelliae strains ranges from 87.9 to 100%. The variability between the two strains was especially due to the lack of sequences or uncalled bases (Ns) in the New Zealand whole genome assembly.
A total of 36 tandem repeats were found within the C. camelliae mitogenome. The two longest repeat sequences measure 218 and 101 bp, respectively. The first is located in an intergenic region among tRNA genes. The second is located in the third intron of cox1. In total, the pair-wise nucleotide similarities ranged from 71 to 100%. Many of these regions were repeated approximately twice. The highest repetition was 7. Repetitive sequences were found also in the other investigated species (Table 4).
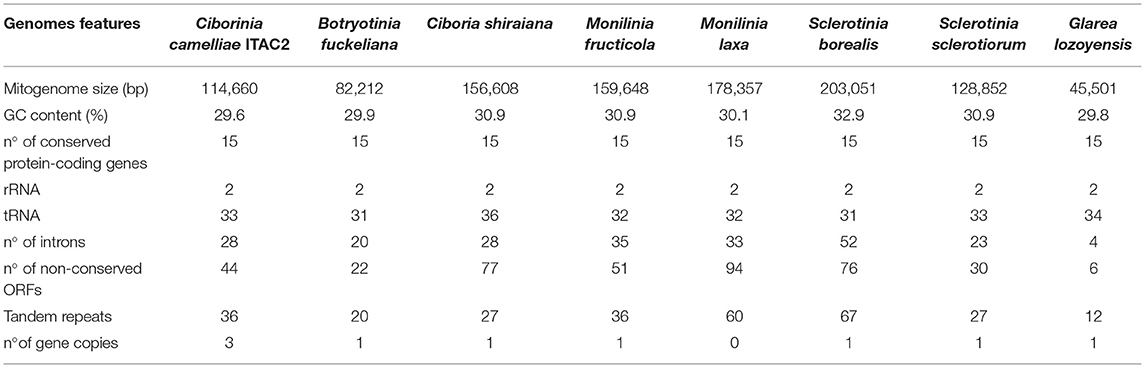
Table 4. Comparison of Ciborinia camelliae strain ITAC2 mitogenome (OK326902) with closely related species: Botryotinia fuckeliana (KC 832409), Ciboria shiraiana (CM 017871.1), Monilinia fructicola (NC_056195.1), Monilinia laxa (NC_051483.1), Sclerotinia borealis (KJ434027), Sclerotinia sclerotiorum (KT283062) and Glarea lozoyensis (NC_031375.1) as outgroup.
Pearson's correlation analysis was performed in species reported in Table 4. The association between the mitogenome size and the number of introns, non-conserved ORFs and tandem repeats was supported by statistical data (Supplementary Data Sheet 6). Each independent variable exhibited a high correlation coefficient with the mitogenome size, ranging from 0.929 to 0.869 (p-value < 0.05).
The arrangements of the 15 protein-coding genes were compared among some related species (Figure 2). Gene order in C. camelliae is identical to most of the Sclerotiniaceae species, except for minor differences due to replication events of atp8, atp9, and cob genes. Among Sclerotiniaceae, gene arrangement in M. fructicola appears with more evident differences. Nevertheless, all investigated species preserved four synteny units: nad5-nad4L, nad3-nad2, atp6-atp8, and cox3-nad6. The location of atp9 and cob truncated genes of C. camelliae could be connected to the gene order of Glarea lozoyensis, even if reversed. The position of the atp8 truncated gene between nad2 and atp6 was found also in Aspergillus flavus (NC_026920.1).

Figure 2. Mitogenome organization of the seven Sclerotiniaceae investigated [Botryotinia fuckeliana (KC 832409), Ciboria shiraiana (CM 017871.1), Ciborinia camelliae strain ITAC2 (OK326902), Monilinia fructicola (NC_056195.1), Monilinia laxa (NC_051483.1), Sclerotinia borealis (KJ434027) and Sclerotinia sclerotiorum (KT283062)] and Glarea lozoyensis (NC_031375.1) as outgroup.
The Mauve alignment was performed only considering Sclerotiniaceae species. This investigation revealed the presence of 19 homologous regions between the seven species (Figure 3). The length of homologous sites ranged from 128 bp to 115,310 bp. The two largest regions contain respectively the cox1 gene and nad4L, nad5, rns, nad6, and cox3 gene. A homologous region (5 Kbp), containing the truncated atp9 gene, was found only in C. camelliae and S. borealis mitogenomes.
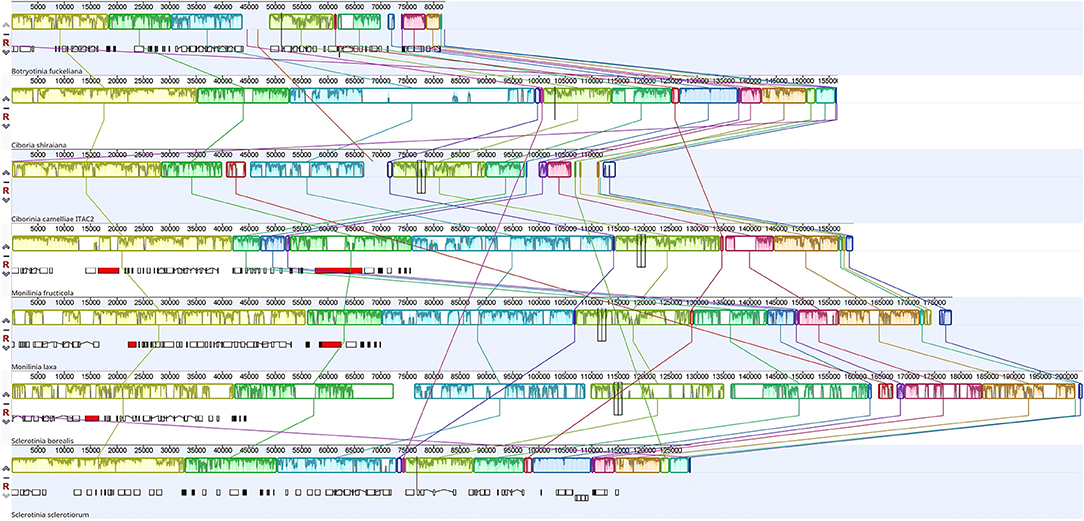
Figure 3. Mauve alignment of Sclerotiniaceae mitochondrial genomes (Botryotinia fuckeliana, Ciboria shiraiana, Ciborinia camelliae strain ITAC2, Monilinia fructicola, Monilinia laxa, Sclerotinia borealis and Sclerotinia sclerotiorum). The alignment was performed setting nad4L gene as origin. The colored blocks reveal homologous regions between mitogenomes. Each block exhibits nucleotide similarity profile. Homologous sites are connected by lines.
Phylogenetic analysis was carried out using the amino acid sequences of 14 protein-coding mitochondrial genes. Nine Sclerotiniaceae species with publicly available data were investigated. To obtain a second mitochondrial set of genes from another C. camelliae, a partial mitochondrial genome composed of a contig list was obtained from the whole genome assembly of C. camelliae ICMP 19812 strain from New Zealand (Supplementary Data Sheet 5). Glarea lozoyensis was considered as an outgroup. The phylogenetic tree (Figure 4), executed according to the model VT+F+R3, demonstrated a close connection between C. camelliae ITAC2 strain and the members of the Sclerotiniaceae family. The two C. camelliae strains group as an independent clade, distinct from the other Sclerotiniaceae species. They exhibit 95.4 and 98.3% of similarity in nucleotide and amino acid sequence, respectively.
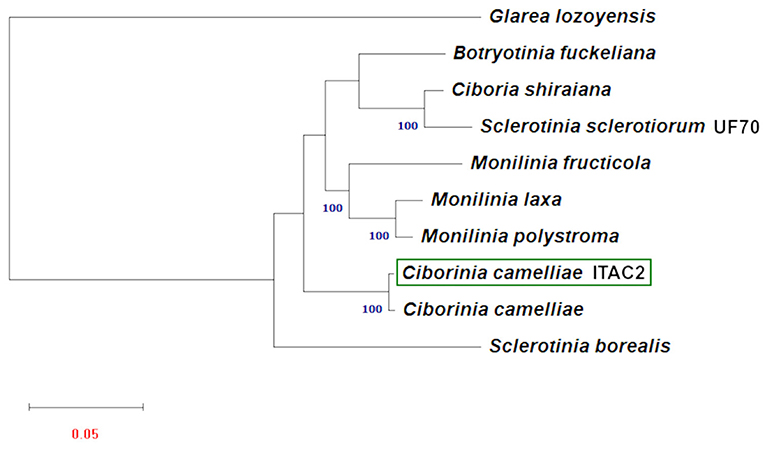
Figure 4. Maximum Likelihood phylogenetic tree based on 14 encoded mitochondrial genome. Blue numbers are bootstrap support values (>90). Red number denotes branch length. The GeneBank accession numbers are listed as follow: Botryotinia fuckeliana (KC 832409), Ciboria shiraiana (CM 017871.1), Ciborinia camelliae (GCA_001247705.1), Ciborinia camelliae strain ITAC2 (OK326902) Glarea lozoyensis (NC_031375.1), Monilinia fructicola (NC_056195.1), Monilinia laxa (NC_051483.1), Monilinia polystroma (GCA_002909645.1), Sclerotinia borealis (KJ434027) and Sclerotinia sclerotiorum (KT283062).
Nowadays, the growth of high-throughput sequencing technologies has improved phylogenetic works using mitogenomes (Zardoya, 2020). In this study, the first mitochondrial genome of C. camelliae was described demonstrating the phylogenetic position of the species in the Sclerotiniaceae family. The mitochondrial genome of C. camelliae was obtained by combining Illumina and Nanopore reads, complemented with Sanger sequencing of specific PCR products to verify the assembly. Experimentally, the combination of long and short reads allowed us to obtain a good quality mitochondrial genome as observed for other species (Degradi et al., 2021). Employing different sequencing technologies was useful to verify the existence of repetitive elements and to define their length and position in the mitogenome assembly (Kinkar et al., 2021). The Sclerotiniaceae mitogenomes were already investigated previously (Mardanov et al., 2014; Yildiz and Ozkilinc, 2020, 2021). Our work contributes to improving the knowledge on mitogenomes of the Sclerotiniaceae family by performing comparative analysis. According to the NCBI data, the C. camelliae strain ITAC2 mitogenome with its 114,660 bp is confirmed in the range from 82 to 203 kbp of the Sclerotiniaceae species (Table 4). Within this family, B. fuckeliana and C. camelliae exhibit the most compact mitochondrial genomes. Among the investigated species, C. camelliae mitogenome contains the highest number of truncated gene copies (atp8, atp9, and cob). Intermediate values in the number of non-conserved open reading frames (ORFs), introns, and tandem repeats were detected. According to Pearson's correlation analysis, the distribution of these elements, due to the evolutionary process, contributes to the high variability in mitogenome size among closely related species (Jung et al., 2010; Xiao et al., 2017; Chen et al., 2021). The intronic regions resulted the greatest promoters of variations in mitogenomes size (Li Q. et al., 2021).
Excluding the events of gene duplication, the C. camelliae gene arrangement is identical to most of the Sclerotiniaceae. All the examined species exhibited four syntenic units: nad5-nad4L, nad3-nad2, atp6-atp8, and cox3-nad6. These clusters were preserved also in the MAUVE alignment. One of the largest LCBs contains the nad4L-nad5 and nad6-cox3 pairs. The homologous regions measuring 22 and 38 Kbp include the nad2-nad3 and atp6-atp8 genes. These conserved clusters could have originated from a common ancestral gene order and later assumed a different organization. Some Helotiales species, such as Phialocephala subalpine, Rhynchosporium orthosporum, or Rhynchosporium secalis, evolutionary close to Sclerotiniaceae, exhibit the unit nad4-nad1, also found in some Sordariomycetes and Eurotiomycetes (Pantou et al., 2006). This cluster was instead split by atp9 and cob genes in Sclerotiniaceae. In all investigated species, the rps3 gene is located in the rnl region and is not interrupted by introns.
Codon usage analysis showed similar use of the optimal codons among the related species (C. camelliae, B. fuckeliana, C. shiraiana, M. fructicola, M. laxa, S. borealis, and S. sclerotiorum). The main differences were observed in the use of codons encoding Valine and Threonine amino acids. The same results were obtained considering less related species such as Glarea lozoyensis (NC_031375.1) and Fusarium oxysporum (NC_017930.1), suggesting the preferential use of codons is conserved among more evolutionarily distant species of fungi. Our study indicates the presence of more conserved mitochondrial regions (nad2, atp8, and rps3) and others prone to more variability (cox1, atp9, nad3, cob, and nad4L). For example, all the investigated Sclerotiniaceae showed the same codon usage in the nad2 gene, while atp9 and nad3 exhibited dissimilarities in 12 amino acids. This variability may be due to weak natural selection toward synonymous substitutions not resulting in amino acid modifications (Stewart et al., 2011). Also the ORFs and the conserved regions of C. camelliae mtDNA showed a similar codon usage suggesting the same selective pressure for translational efficiency (Whittle et al., 2012). Nevertheless, some differences between conserved and non-conserved ORFs were observed in the differential use of synonymous codons, namely codon usage bias (CUB). The codon bias has been reported in many organisms (Duret, 2002; Sharp et al., 2005; Xiang et al., 2015; Gupta and Singh, 2021) including mitochondrial genomes (Wei et al., 2014; Yildiz and Ozkilinc, 2021). Highly-expressed genes exhibit a strong bias to conserve their translational efficiency. On the other hand, CUB decreases in non-conserved genes due to weak selection, thus they can afford to use different codons having low expression (Bulmer, 1991). On the contrary, a low CUB in weakly expressed ORFs is a mechanism for preserving a low level of expression (Grosjean and Fiers, 1982). The influence of selective pressure on the codon bias makes CUB an interesting source of species evolution force leading to their environmental adaptation (Angellotti et al., 2007; Li X. et al., 2021), which include host-pathogen relationships (Gupta and Singh, 2021). Further studies may investigate the functional role of CUB in fungal mitochondrial genomes.
As observed in other fungi (Mardanov et al., 2014; Yildiz and Ozkilinc, 2021), only the Methionine amino acid ends in G, instead, the last base of all the other preferential codons is A or T confirming the high AT content of fungal mitogenomes (Chen et al., 2019; Zhang et al., 2020).
The BLASTp analysis of the non-conserved ORFs was useful for revealing phylogenetic relationships among species. All ORFs exhibiting a significant hit in BLAST analysis, showed the best similarity with an Ascomycota species. Sixty-six percent of ORFs demonstrated the highest similarity with ORFs from the Sclerotiniaceae species. According to BLAST results, six of C. camelliae ORFs could have origins distinct from the Sclerotiniaceae family. Two interesting LAGLIDADG, located in nad5 and cox1 introns, showed 90.75 and 85% of amino acid identity with F. tricinctum and G. cichoracearum, respectively. These regions may represent events of horizontal transmission from distant fungal species, given the ability of intron I and homing endonuclease genes to move and integrate into diverse genomes (Beaudet et al., 2013; Celis et al., 2017).
The phylogenetic study based on multiple alignment of 14 concatenated mtDNA-encoded protein-related sequences further confirmed that C. camelliae belongs to the family of Sclerotiniaceae and its relationships among strictly related species. Phylogenetic results showed a significant difference between the two Sclerotinia species. Our results are partially confirmed by other works on mitochondrial DNA, where the dissimilarity between S. borealis and S. sclerotiorum was reported (Mardanov et al., 2014; Ma et al., 2019; Yildiz and Ozkilinc, 2021). The nucleotide and amino acid similarity between the two Sclerotinia species are 36.4 and 77.7%, respectively. Discrepancies between mitochondrial and nuclear phylogeny shall be further investigated in this genus. This phylogenetic analysis proved the closest match between the strain ITAC2 and the strain ICMP 19812 of C. camelliae from New Zealand, with which they constitute an independent monophyletic group. Nevertheless, some differences between the two strains at the nucleotide and amino acid sequence level could be detected in the mitochondrial genome (95.4 and 98.3% of similarity, respectively). The major level contributor to the diversity in numbers is the cox1 gene. The Italian and New Zealand cox1 genes exhibit 88.3% of amino acid identity, due to the deletion of some amino acids located at the beginning of cox1 in the strain from New Zealand. According to BLAST search, the first exon of the Italian strain results lacking in the New Zealand strain. Excluding this gene, the amino acid identity between the two strains would be 99.8%. According to Mardanov et al. (2014), cox1 is the most common target of intron insertions. This means that cox1 represents a useful marker in phylogenetic studies, as it is evolving more rapidly than the other mitochondrial genes. The significant difference between the two C. camelliae strains may be due to a misassembly of the New Zealand strain genome or could represent the beginning of a speciation event. The Italian and New Zealand population are separated by geographic barriers and they may diverge in complete independence under different selection pressures (Dettman et al., 2008; Stukenbrock, 2013). Moreover, the impact of globalization on the diffusion of the phytopathogens influences the evolutionary process resulting in more variability. One day, the subsequent differentiation of populations may evolve into two different species. Large sequencing of C. camelliae population is warranted to further investigate the worldwide diversity of the species.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: GenBank [accession: OK326902].
MP, MS, and PC designed the work. IV, LD, and AK performed the experiments and acquired data. MP, MS, IV, and LD analyzed and interpreted the data for the work. IV, MP, MS, and AK drafted the manuscript. All authors revised critically the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors acknowledge support from the University of Milan through the APC initiative.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffunb.2021.802511/full#supplementary-material
Afgan, E., Baker, D., Batut, B., van den Beek, M., Bouvier, D., Cech, M., et al. (2018). The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 46, W537–W544. doi: 10.1093/nar/gky379
Aguileta, G., de Vienne, D. M., Ross, O. N., Hood, M. E., Giraud, T., Petit, E., et al. (2014). High variability of mitochondrial gene order among fungi. Genome Biol. Evol. 6, 451–465. doi: 10.1093/gbe/evu028
Angellotti, M. C., Bhuiyan, S. B., Chen, G., and Wan, X. F. (2007). CodonO: codon usage bias analysis within and across genomes. Nucleic Acids Res. 35, W132–W136. doi: 10.1093/nar/gkm392
Ballard, J. W., and Whitlock, M. C. (2004). The incomplete natural history of mitochondria. Mol. Ecol. 13, 729–744. doi: 10.1046/j.1365-294X.2003.02063.x
Beaudet, D., Nadimi, M., Iffis, B., and Hijri, M. (2013). Rapid mitochondrial genome evolution through invasion of mobile elements in two closely related species of arbuscular mycorrhizal fungi. PLoS One 8:e60768. doi: 10.1371/journal.pone.0060768
Beck, N., and Lang, B. (2010). MFannot, Organelle Genome Annotation Websever. Montreal, QC: Université de Montréal.
Benson, G. (1999). Tandem repeats finder: a program to analyze DNA sequences. Nucleic Acids Res. 27, 573–580. doi: 10.1093/nar/27.2.573
Bernt, M., Donath, A., Jühling, F., Externbrink, F., Florentz, C., Fritzsch, G., et al. (2013). MITOS: improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 69, 313–319. doi: 10.1016/j.ympev.2012.08.023
Bolton, M. D., Thomma, B. P., and Nelson, B. D. (2006). Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 7, 1–16. doi: 10.1111/j.1364-3703.2005.00316.x
Bullerwell, C. E., and Lang, B. F. (2005). Fungal evolution: the case of the vanishing mitochondrion. Curr. Opin. Microbiol. 8, 362–369. doi: 10.1016/j.mib.2005.06.009
Bulmer, M. (1991). The selection-mutation-drift theory of synonymous codon usage. Genetics 129, 897–907. doi: 10.1093/genetics/129.3.897
Burger, G., Gray, M. W., and Lang, B. F. (2003). Mitochondrial genomes: anything goes. Trends Genet. 19, 709–716. doi: 10.1016/j.tig.2003.10.012
Celis, J. S., Edgell, D. R., Stelbrink, B., Wibberg, D., Hauffe, T., Blom, J., et al. (2017). Evolutionary and biogeographical implications of degraded LAGLIDADG endonuclease functionality and group I intron occurrence in stony corals (Scleractinia) and mushroom corals (Corallimorpharia). PLoS ONE 12:e0173734. doi: 10.1371/journal.pone.0173734
Chen, C., Li, Q., Fu, R., Wang, J., Deng, G., Chen, X., et al. (2021). Comparative mitochondrial genome analysis reveals intron dynamics and gene rearrangements in two Trametes species. Sci. Rep. 11:2569. doi: 10.1038/s41598-021-82040-7
Chen, C., Li, Q., Fu, R., Wang, J., Xiong, C., Fan, Z., et al. (2019). Characterization of the mitochondrial genome of the pathogenic fungus Scytalidium auriculariicola (Leotiomycetes) and insights into its phylogenetics. Sci. Rep. 9:17447. doi: 10.1038/s41598-019-53941-5
Darling, A. C., Mau, B., Blattner, F. R., and Perna, N. T. (2004). Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 14, 1394–1403. doi: 10.1101/gr.2289704
Degradi, L., Tava, V., Kunova, A., Cortesi, P., Saracchi, M., and Pasquali, M. (2021). Telomere to telomere genome assembly of Fusarium musae F31, causal agent of crown rot disease of banana. Mol. Plant Microbe Interact. 34, 1455–1457. doi: 10.1094/MPMI-05-21-0127-A
Deng, Y., Hsiang, T., Li, S., Lin, L., Wang, Q., Chen, Q., et al. (2018). Comparison of the mitochondrial genome sequences of six Annulohypoxylon stygium isolates suggests short fragment insertions as a potential factor leading to larger genomic size. Front. Microbiol. 9:2079. doi: 10.3389/fmicb.2018.02079
Denton-Giles, M., Bradshaw, R. E., and Dijkwel, P. P. (2013). Ciborinia camelliae (Sclerotiniaceae) induces variable plant resistance responses in selected species of Camellia. Phytopathology 103, 725–732. doi: 10.1094/PHYTO-11-12-0289-R
Dettman, J. R., Anderson, J. B., and Kohn, L. M. (2008). Divergent adaptation promotes reproductive isolation among experimental populations of the filamentous fungus Neurospora. BMC Evol. Biol. 8:35. doi: 10.1186/1471-2148-8-35
Duret, L. (2002). Evolution of synonymous codon usage in metazoans. Curr. Opin. Genet. Dev. 12, 640–649. doi: 10.1016/S0959-437X(02)00353-2
Galtier, N., Nabholz, B., Glémin, S., and Hurst, G. D. (2009). Mitochondrial DNA as a marker of molecular diversity: a reappraisal. Mol. Ecol. 18, 4541–4550. doi: 10.1111/j.1365-294X.2009.04380.x
Goodsell, D. S. (2010). Mitochondrion. Biochem. Mol. Biol. Educ. 38, 134–140. doi: 10.1002/bmb.20406
Grosjean, H., and Fiers, W. (1982). Preferential codon usage in prokaryotic genes: the optimal codon-anticodon interaction energy and the selective codon usage in efficiently expressed genes. Gene 18, 199–209. doi: 10.1016/0378-1119(82)90157-3
Gupta, S., and Singh, R. (2021). Comparative study of codon usage profiles of Zingiber officinale and its associated fungal pathogens. Mol. Genet. Genomics 296, 1121–1134. doi: 10.1007/s00438-021-01808-8
Hamari, Z., Juhász, A., and Kevei, F. (2002). Role of mobile introns in mitochondrial genome diversity of fungi (a mini review). Acta Microbiol. Immunol. Hung. 49, 331–335. doi: 10.1556/amicr.49.2002.2-3.22
Jung, P. P., Friedrich, A., Souciet, J. L., Louis, V., Potier, S., de Montigny, J., et al. (2010). Complete mitochondrial genome sequence of the yeast Pichia farinosa and comparative analysis of closely related species. Curr. Genet. 56, 507–515. doi: 10.1007/s00294-010-0318-y
Kinkar, L., Gasser, R. B., Webster, B. L., Rollinson, D., Littlewood, D., Chang, B., et al. (2021). Nanopore sequencing resolves elusive long tandem-repeat regions in mitochondrial genomes. Int. J. Mol. Sci. 22:1811. doi: 10.3390/ijms22041811
Kriváková, P., Cervinková, Z., Lotková, H., Kucera, O., and Rousar, T. (2005). Mitochondrie a jejich úloha v bunecném metabolismu [Mitochondria and their role in cell metabolism]. Acta medica (Hradec Kralove) 48, 57–67. doi: 10.14712/18059694.2018.34
Kulik, T., Brankovics, B., van Diepeningen, A. D., Bilska, K., Zelechowski, M., Myszczyński, K., et al. (2020). Diversity of mobile genetic elements in the mitogenomes of closely related Fusarium culmorum and F. graminearum sensu stricto strains and its implication for diagnostic purposes. Front. Microbiol. 11:1002. doi: 10.3389/fmicb.2020.01002
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Lang, B. F., Laforest, M. J., and Burger, G. (2007). Mitochondrial introns: a critical view. Trends Genet. 23, 119–125. doi: 10.1016/j.tig.2007.01.006
Li, H. (2018). Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics (Oxford, England) 34, 3094–3100. doi: 10.1093/bioinformatics/bty191
Li, Q., Li, L., Feng, H., Tu, W., Bao, Z., Xiong, C., et al. (2021). Characterization of the complete mitochondrial genome of basidiomycete yeast Hannaella oryzae: intron evolution, gene rearrangement, and its phylogeny. Front. Microbiol. 12:646567. doi: 10.3389/fmicb.2021.646567
Li, X., Wang, X., Gong, P., Zhang, N., Zhang, X., and Li, J. (2021). Analysis of codon usage patterns in Giardia duodenalis based on transcriptome data from GiardiaDB. Genes 12:1169. doi: 10.3390/genes12081169
Lin, Y., Yuan, J., Kolmogorov, M., Shen, M. W., Chaisson, M., and Pevzner, P. A. (2016). Assembly of long error-prone reads using de Bruijn graphs. Proc. Natl. Acad. Sci. 113, E8396–E8405. doi: 10.1073/pnas.1604560113
Lv, J., Bhatia, M., and Wang, X. (2017). Roles of mitochondrial DNA in energy metabolism. Adv. Exp. Med. Biol. 1038, 71–83. doi: 10.1007/978-981-10-6674-0_6
Ma, Y., Huang, L., Abuduaini, A., Zhou, H., Wang, Y., and Suo, F. (2019). Complete mitochondrial genome of plant pathogen Monilinia fructicola (Sclerotiniaceae, Helotiales). Mitochondrial DNA B. 4, 791–792. doi: 10.1080/23802359.2019.1567282
Mardanov, A. V., Beletsky, A. V., Kadnikov, V. V., Ignatov, A. N., and Ravin, N. V. (2014). The 203 kbp mitochondrial genome of the phytopathogenic fungus Sclerotinia borealis reveals multiple invasions of introns and genomic duplications. PLoS ONE 9:e107536. doi: 10.1371/journal.pone.0107536
Medina, R., Franco, M., Bartel, L. C., Martinez Alcántara, V., Saparrat, M., and Balatti, P. A. (2020). Fungal mitogenomes: relevant features to planning plant disease management. Front. Microbiol. 11:978. doi: 10.3389/fmicb.2020.00978
Newmeyer, D. D., and Ferguson-Miller, S. (2003). Mitochondria: releasing power for life and unleashing the machineries of death. Cell 112, 481–490. doi: 10.1016/S0092-8674(03)00116-8
Pantou, M. P., Kouvelis, V. N., and Typas, M. A. (2006). The complete mitochondrial genome of the vascular wilt fungus Verticillium dahliae: a novel gene order for Verticillium and a diagnostic tool for species identification. Curr. Genet. 50, 125–136. doi: 10.1007/s00294-006-0079-9
Sandor, S., Zhang, Y., and Xu, J. (2018). Fungal mitochondrial genomes and genetic polymorphisms. Appl. Microbiol. Biotechnol. 102, 9433–9448. doi: 10.1007/s00253-018-9350-5
Saracchi, M., Colombo, E. M., Locati, D., Valenti, I., Corneo, A., Cortesi, P., et al. (2022). Morphotypes of Ciborinia camelliae Kohn infecting camellias in Italy. J. Plant Pathol. 104. doi: 10.1007/s42161-022-01040-2
Saracchi, S., Locati, D., Colombo, E. M., and Pasquali, M. (2019). Updates on Ciborinia camelliae, the causal agent of camellia flower blight. J. Plant Pathol. doi: 10.1007/s42161-018-0173-0
Sharp, P. M., Bailes, E., Grocock, R. J., Peden, J. F., and Sockett, R. E. (2005). Variation in the strength of selected codon usage bias among bacteria. Nucleic Acids Res. 33, 1141–1153. doi: 10.1093/nar/gki242
Shingu-Vazquez, M., and Traven, A. (2011). Mitochondria and fungal pathogenesis: drug tolerance, virulence, and potential for antifungal therapy. Eukaryotic Cell 10, 1376–1383. doi: 10.1128/EC.05184-11
Stewart, J. E., Kawabe, M., Abdo, Z., Arie, T., and Peever, T. L. (2011). Contrasting codon usage patterns and purifying selection at the mating locus in putatively asexual Alternaria fungal species. PLoS ONE 6:e20083. doi: 10.1371/journal.pone.0020083
Stothard, P. (2000). The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques 28, 1102–1104. doi: 10.2144/00286ir01
Stukenbrock, E. H. (2013). Evolution, selection and isolation: a genomic view of speciation in fungal plant pathogens. New Phytol. 199, 895–907. doi: 10.1111/nph.12374
Taylor, C. H., and Long, P. G. (2000). Review of literature on camellia flower blight caused by Ciborinia camelliae. N. Z. J. Crop Horticult. Sci. 28, 123–138. doi: 10.1080/01140671.2000.9514132
Trifinopoulos, J., Nguyen, L. T., von Haeseler, A., and Minh, B. Q. (2016). W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, W232–W235. doi: 10.1093/nar/gkw256
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3–new capabilities and interfaces. Nucleic Acids Res. 40:e115. doi: 10.1093/nar/gks596
van Toor, R. F., Ridgway, H. J., Butler, R. C., Jaspers, M. V., and Stewart, A. (2005). Assessment of genetic diversity in isolates of Ciborinia camelliae Kohn from New Zealand and the United States of America. Australas Plant Path 34, 319–325. doi: 10.1071/AP05040
Walker, B. J., Abeel, T., Shea, T., Priest, M., Abouelliel, A., Sakthikumar, S., et al. (2014). Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 9:e112963. doi: 10.1371/journal.pone.0112963
Wei, L., He, J., Jia, X., Qi, Q., Liang, Z., Zheng, H., et al. (2014). Analysis of codon usage bias of mitochondrial genome in Bombyx mori and its relation to evolution. BMC Evol. Biol. 14:262. doi: 10.1186/s12862-014-0262-4
Whittle, C. A., Sun, Y., and Johannesson, H. (2012). Genome-wide selection on codon usage at the population level in the fungal model organism Neurospora crassa. Mol. Biol. Evol. 29, 1975–1986. doi: 10.1093/molbev/mss065
Xiang, H., Zhang, R., Butler, R. R. 3rd, Liu, T., Zhang, L., Pombert, J. F., and Zhou, Z. (2015). Comparative analysis of codon usage bias patterns in microsporidian genomes. PLoS ONE 10:e0129223. doi: 10.1371/journal.pone.0129223
Xiao, S., Nguyen, D. T., Wu, B., and Hao, W. (2017). Genetic drift and indel mutation in the evolution of yeast mitochondrial genome size. Genome Biol. Evol. 9, 3088–3099. doi: 10.1093/gbe/evx232
Yildiz, G., and Ozkilinc, H. (2020). First characterization of the complete mitochondrial genome of fungal plant-pathogen Monilinia laxa which represents the mobile intron rich structure. Sci. Rep. 10:13644. doi: 10.1038/s41598-020-70611-z
Yildiz, G., and Ozkilinc, H. (2021). Pan-mitogenomics approach discovers diversity and dynamism in the prominent brown rot fungal pathogens. Front. Microbiol. 12:647989. doi: 10.3389/fmicb.2021.647989
Zardoya, R. (2020). Recent advances in understanding mitochondrial genome diversity. F1000Research 9, 1–19. doi: 10.12688/f1000research.21490.1
Keywords: fungal diversity, pathogen of Camellia, mitochondrial assembly and annotation, short and long read sequencing, mitochondrial diversity
Citation: Valenti I, Degradi L, Kunova A, Cortesi P, Pasquali M and Saracchi M (2022) The First Mitochondrial Genome of Ciborinia camelliae and Its Position in the Sclerotiniaceae Family. Front. Fungal Biol. 2:802511. doi: 10.3389/ffunb.2021.802511
Received: 26 October 2021; Accepted: 06 December 2021;
Published: 09 February 2022.
Edited by:
Georg Hausner, University of Manitoba, CanadaReviewed by:
Daniel S. Araújo, Loyola University Chicago, United StatesCopyright © 2022 Valenti, Degradi, Kunova, Cortesi, Pasquali and Saracchi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matias Pasquali, bWF0aWFzLnBhc3F1YWxpQHVuaW1pLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.