 Saveena Sangry
Saveena Sangry Praveen Kumar
Praveen Kumar D.R. Bhardwaj
D.R. Bhardwaj K.S. Dogra2
K.S. Dogra2- 1Department of Silviculture and Agroforestry, College of Forestry, Dr Y S Parmar University of Horticulture and Forestry, Nauni-Solan, Himachal Pradesh, India
- 2Botanical Survey of India (Ministry of Environment, Forest & Climate Change, Govt. of India), High Altitude Western Himalayan Regional Centre, UHF, Nauni Campus, Solan, India
- 3Faculty of Applied Sciences and Biotechnology, Shoolini University, Solan, Himachal Pradesh, India
Introduction: The Indian Himalayan forests are remarkable landforms experiencing tremendous climatic variation, constituting complex and diversified ecosystems with prominent vegetation zones. Despite their global significance and substantial research efforts focused on plant diversity in the temperate Himalayan region, only a few studies have explicitly assessed the distribution patterns of understory vegetation in relation to forest compositional types along altitudinal gradients.
Methods: To cover a wide range of altitudes and diverse overstory compositions, stands were sampled across four altitudinal ranges from 1500 to 3500 meters above mean sea level with increments of 500 meters in elevation steps. The overstory compositions were classified on the basis of dominant tree species in each stand on the similar sites. Vegetation in the shrub and ground layers was surveyed by visually estimating the percentage coverage within circular plots.
Results: A total of 99 understory species including 37 species each in the shrub layer, 62 species in the herb layer vegetation were recorded. The abundance, species diversity and composition of understory vegetation differed significantly along the altitudinal gradient and dominant overstory composition types. Moreover, distinct understory vegetation communities were observed at lower elevations compared to higher elevations, with middle elevations exhibiting intermediate vegetation characteristics. The study also highlighted the importance of dominant overstory composition types in shaping the pattern of understory vegetation abundance, species diversity and composition in the temperate Himalayan region. The higher resource conditions associated with broadleaved stands supported higher understory species abundance at lower elevations, while the heterogeneous conditions induced by the mixedwood stands promoted higher understory species diversity.
Conclusion: The hump shaped pattern along the altitudinal gradient appeared to be the most dominant pattern of plant abundance and species diversity and call for more conservation concern towards the middle elevation zones in the temperate Himalayan region. Furthermore, the management interventions should aim at maintaining diverse range of overstory composition types for conserving biodiversity and their ecological functions in the temperate Himalayan region.
1 Introduction
Forest ecosystems are one of the major categories of land forms which constitute the most important natural resources of the world and a template for biodiversity and ecosystem functions. In India, forest constitute around 21.71 percent of country’s geographical area (Forest Survey of India, 2021), ranging from Himalayan temperate forest to Dry zone forests, and representing one of the 12 mega biodiversity regions of the world (Chitale et al., 2014; Sankaran and Dinesh, 2020). Among all, Himalayan temperate forests are the most remarkable landforms on earth, experiencing tremendous variation in climatic conditions (Rawal et al., 2018; White et al., 2019), constituting complex and diversified ecosystems with prominent vegetation zones and are one of the youngest and richest ecosystems on the earth. These forests contribute nearly 36% of the total forest area in the country with the variety of species, forest types having high level of endemism and are recognized as one of the hotspots for biodiversity (Maletha et al., 2020; Forest Survey of India, 2021; Maletha et al., 2022). Despite its global significance, the biodiversity of the Himalayan region is inadequately studied with a considerable portion of research published in low-impact-factor journals and is data and information deficient (Rana et al., 2021). This is primarily due to the heterogeneous landscapes together with accessibility challenges along wide altitudinal gradients. The lack of knowledge hampers our ability to understand the patterns of plant diversity and composition along altitudinal gradients and under different dominant overstory compositions in the temperate region of Himachal Pradesh-India.
In the temperate Himalayan region, the majority of plant diversity is in the understory layer (Roberts and Gilliam, 2003; Whigham, 2004). The understory layer mainly consist of herbs and shrubs together with a mixture of seedlings and saplings of canopy trees and other non-vascular plants (Roberts and Gilliam, 2003; Bartels and Chen, 2013; Kumar et al., 2017a). Understorey vegetation accounts for more than 80% of vascular species in temperate forests (Gilliam, 2007), in contrast to tropical forests, which generally have 14–40 percent of all forest vascular species (Costa et al., 2005). They represent the vital stratum of forest ecosystem and play a crucial role in the functioning of forest ecosystems (Augusto et al., 2003; Hart and Chen, 2006; Gilliam, 2007; Barbier et al., 2008; Cardinale et al., 2012; Kumar et al., 2018). While understory vegetation in temperate forests may not contribute significantly to biomass compared to trees, their contribution in terms of regulating ecosystem processes, both in short term by affecting tree seedling regeneration and in long term by driving soil bio-geochemical cycles, are way more than their biomass (Hart and Chen, 2006; Barbier et al., 2008; Haq et al., 2024). However, the distribution pattern of understory vegetation in the temperate Himalayan region largely remains understudied and is one of least explored areas compared to the trees. Moreover, few studies have explicitly assessed the distribution patterns of understory vegetation abundance, diversity and composition in relation to forest compositional types along the altitudinal gradient in the Himalayan region. Therefore, understanding the distribution pattern of understory vegetation can be a major aspect in the biodiversity studies in Himalayan region.
The diversity in composition of plant communities and their distribution patterns in forest ecosystems are primarily influenced by factors such as the altitudinal gradient, distinct forest types, species composition, topographical, geographical and climatic variations etc (Grytnes and Beaman, 2006; McCain, 2007; Dar and Sundarapandian, 2016; Bhat et al., 2020; Bisht et al., 2022; Wani et al., 2022; Rawat et al., 2023; Sekar et al., 2023). However, altitude, among others, is considered to be the most important factor affecting the species distribution, structure and composition of the forest vegetation in the mountainous region (Wang et al., 2007; Sinha et al., 2018; Maletha et al., 2022). As altitude itself represents a complex combination of factors influencing solar radiation, water, and nutrient distribution, resulting in climatic, edaphic, and disturbance variations that significantly affect plant community structure and composition in mountainous temperate ecosystems (Körner, 2007; McVicar and Korner, 2013; Ping et al., 2013; Dar and Sundarapandian, 2016; Xu et al., 2017; Rawal et al., 2018). These ecosystems provide large altitudinal gradient and variations in the characteristics of vegetation with altitude effectively characterize the vegetation in a simple yet powerful way (Baniya et al., 2010). Although, various studies have attempted to understand the pattern of species diversity and richness along the altitudinal gradient in the temperate Himalayan region, but there is still no consensus on a universal pattern of plant diversity (Liang et al., 2020). Some studies have reported a mid-elevation peak (Chawla et al., 2008; Acharya et al., 2011; Rawat et al., 2021; Liang et al., 2023), while others have reported a monotonic decrease with elevation (Sharma et al., 2009a; Yang et al., 2014; Zhang et al., 2016; Geberhiwot et al., 2019). However, very few studies have exclusively focused on the patterns of understory vegetation abundance, species diversity, and composition along the altitudinal gradient in the temperate Himalayan Region. As a result, data on the abundance, diversity and composition of understory vegetation in the temperate Himalayan region remains scarce. Furthermore, much of the research on plant diversity in the Himalayan region dates back to the 19th century (Rana et al., 2021), creating a knowledge gap about the dynamics of the Himalayan region which is extremely susceptible to climatic and anthropogenic disturbances (Sinha et al., 2018). Therefore, there is an urgent need for new research and exploration of Himalayan biodiversity along the altitudinal gradient to restore the ecological balance and sustainable development in Himalayan ecosystems.
In the forest ecosystems, the interaction between the dominant overstory tree and understory plants plays a critical role in driving the community composition and structure in the understory vegetation (Bartels and Chen, 2013; Kumar et al., 2017b). The distribution and composition of plant communities are strongly influenced by competitive and facilitative effect from the overstory trees (Gracia et al., 2007; Barbier et al., 2008; Cavard et al., 2011; Zhang et al., 2016; Kumar et al., 2018). The structure and composition of overstory tree, i.e., the uppermost layer of foliage in a forest canopy can control the understory vegetation species diversity and composition by modifying the availability of resources such as light, water, and soil nutrients, and by competing with both above and below ground resources (Pelt and Franklin, 2000; Van Oijen et al., 2005; Kumar et al., 2017b). The interception of solar radiation by the overstory trees is a major factor affecting the understory vegetation (Hardwick et al., 2015; Tonteri et al., 2016), depending upon the overstory tree species properties, such as spatial arrangement of leaves, leaf area index, and varies considerably among different forest types (Ishii et al., 2013; Tinya and Ódor, 2016; Tonteri et al., 2016). On the contrary, understory vegetation inhabiting the tree canopy may benefit from increased organic matter due to litter fall production, enhanced moisture levels, and reduced air and soil temperatures by the overstory trees (Nilsson and Wardle, 2005; Valladares et al., 2016; Kumar et al., 2020).
In the temperate Himalayan region, the overstory species composition of various broadleaved and coniferous trees transition along the altitudinal gradient where broadleaved stands transition from Quercus leucotricophora at lower elevations to Quercus semecarpifolia at higher elevations, coniferous from Cedrus deodara at lower elevations to Picea samithaina and Abies pindrow at higher elevations, with mixedwood co-dominated by oaks and coniferous species along the elevation gradients (Thakur et al., 2024). Therefore, distinct plant communities can be found under the stands dominated by broadleaved and coniferous species at distinct elevation zones as a result of different resource conditions in the understory layers (Hart and Chen, 2008; Kumar et al., 2017b). For example, broadleaf canopies support nutrient-demanding and shade-intolerant vascular plants due to the high resource availability, such as light and soil nutrients, for the understory plants (Bartels and Chen, 2013). In contrast, coniferous trees facilitate the growth and establishment of shade-tolerant species due to lower light availability and a recalcitrant litter layer (Hart and Chen, 2008; Chavez and Macdonald, 2010). Mixedwood stands, on the other hand, create heterogeneous resource conditions (both spatially and temporally) in the understory which can support more diverse understory plant communities than the pure forests (Hart and Chen, 2006; Barbier et al., 2008; Chavez and Macdonald, 2010). However, few studies have explicitly assessed the distribution patterns of plant diversity in relation to forest compositional types along the altitudinal gradient in the temperate Himalayan region. The purpose of this study is to examine the cumulative effect of the altitudinal gradient and dominant overstory types on the abundance, species diversity and composition of understory plant communities in the temperate region of Himachal Pradesh. We specifically address the following questions: (1) how does the abundance; diversity and composition of vascular plants vary along the wide altitudinal gradient in the temperate Himalayan region? (2) Do the elevation related changes in abundance, diversity and composition of understory plants differ among the dominant forest composition types? Given the known effect of altitudinal gradients and overstory composition types on understory diversity and composition, we hypothesize that the understory vegetation abundance and diversity will follow a hump-shaped relationship along the altitudinal gradients which will increase linearly from lower to middle altitudes and decreased thereafter as a result of optimal growth conditions for the growth and establishment of vascular plants (Zhang et al., 2016; Liang et al., 2020). Additionally, the intermediate location of this zone, serving as a transition between lower and higher elevations, supports the presence of additional species with distinct niche preferences (Scheiner et al., 2011; Zhang et al., 2016; Das et al., 2020). Furthermore, we also hypothesized that independent of altitudinal gradients; the stands dominated by mixedwood species (both broadleaved and conifers) would support the higher understory vegetation abundance and diversity as compared to pure stands because of heterogeneous conditions for growth and development of diverse plant communities in the understory (Hart and Chen, 2006; Barbier et al., 2008; Cavard et al., 2011).
2 Methods
2.1 Study area
The study area covered Solan and Shimla districts of Himachal Pradesh-India extended along the wide altitudinal range from 1,500–3,500 m above mean sea level (amsl), lying between 30°90′74′′N to 31°24′12′′N latitude and 77°12′61′′E to 77°50′19′′E longitude, and have distinct elevational zones that were further divided into broadleaved, mixedwood and coniferous forest composition types (Supplementary Figure S1). The study location experiences temperate climatic conditions characterized by distinct seasonal variations. Summers (April to mid-June) are generally warm and dry, while the rainy season (mid-June to mid-September) is slightly warm and humid. Winters (October to March) are harsh, with freezing temperatures and frequent snowfalls. March and April are generally sunny, but snowfall can still occur in elevated areas. The temperature starts rising rapidly in late May or early June. April to June is relatively dry months, with occasional severe hail and thunderstorms. The monsoon season can also extend from the end of August to the middle of September, bringing the majority of the area’s rainfall. October and November are mostly dry but cold, with the possibility of snowfall in higher elevations from October onwards. The frost, particularly at higher altitudes, is also common throughout from November to February; however, the intensity and duration of the conditions vary with the altitude.1
2.2 Stand and site selection
In the present study, a factorial experiment was conducted focusing on two primary factors: altitude and overstory composition types, each with fixed levels. To ensure comprehensive sampling, a stratified random sampling technique was employed. This approach allowed selection of stands across a broad range of altitudes and diverse overstory compositions. Specifically, stands were sampled at four altitudinal ranges, ranging from 1,500 to 3,500 meters above mean sea level, with increments of 500 meters in elevation steps. The overstory composition was selected from relatively pure coniferous stands, both broadleaf and conifer mixtures, and pure broadleaf stands on the basis of dominant tree species in each stand on the similar sites. Pure stands were defined as those with an overstory composed of more than 65% broadleaf and coniferous species by stand basal area. Mixedwood stands were defined as those where neither broadleaf nor conifer tree species comprised more than 65% of the composition by stem density or stand basal area (Kumar et al., 2017b). As broadleaf and coniferous species transition along the altitudinal gradient in temperate Himalayan region, broadleaved stands dominated by pure oaks species (transitioning along the altitude from Quercus leucotricophora to Quercus semecarpifolia), coniferous stands ranging from Cedrus deodara at lower elevations to Picea samithaina and Abies pindrow at higher elevations, and mixedwood stands co-dominated by oaks and coniferous species were selected. Each altitudinal range and overstory type combinations were replicated minimum of three times resulting in a total of 36 sampled plots. The selected stands were spatially interspersed across large area in order to minimize spatial autocorrelation. The selected stand was >1 ha in area and visually homogeneous in structure and composition. To minimize the effects of edaphic variability, selected stands were located on the flat or mid-slope positions.
2.3 Field data collection
The field survey was conducted during peak vegetation cover period (May–August), 2022. In each forest stand, a 400m2 circular plot (radius = 11.28 m) located at least 100 m from the forest edges, was established, within which all sampling was done. Overstory and understory were classified based on their crown position in a stand utilizing the classification method proposed by Avery and Burkhart (2002). Within the plot, all live trees with diameters at breast height (DBH, 1.3 m above the root collar) ≥ 5 cm were tallied by species and their DBH were measured and recorded. DBH measurements were used to calculate the basal areas of sampled tree species.
The understory plants survey including both shrub and herb or ground layer plants were carried out during the peak vegetation cover period from July to August in 2022. Vegetation in the shrub layer and ground layer were surveyed by visually estimating the percentage of coverage within circular plots by using the method described by Mueller-Dombois and Ellenberg (1974). The shrub layer was defined as any species, including tree saplings of height between 1.3 and 4.0 m (Kumar et al., 2017b); as such, species found in the shrub layer could also be present in the ground layer. All the plant species <1.3 m in the height including small tree saplings, dwarf shrub and shrubs and herbaceous vascular plants (gramminoids, herbs/forbs and ferns) were sampled as a part of herb or ground layer (Su et al., 2021). The shrub layer was sampled separately because of large spatial requirements in sampling. The shrub layer species was sampled by visually estimating the percent cover of each species with four sub-plots of size 5 m × 5 m (25 m2) plots (Kumar et al., 2017b). These sub-plots were located at random distance from the plot center. The ground layer vegetation was sampled by visually estimating the percent cover of all vascular plant species within 10 randomly located 1 m × 1 m sub-plots. The quadrates were established by random throws from the plot center at random distances and directions. Every effort was made to identify all the vascular plants to the species level, and unidentified plant species were identified with the help of expert at the High-Altitude Regional Center-Solan of the Botanical Survey of India.2 Species-specific sub-plot level percent cover data were averaged to represent the sample stand. We used species richness and Shannon Index as a measure of plant diversity because each species contributes uniquely to ecosystem functions according to the singular hypothesis (Naeem, 2002).
2.4 Data analysis
Total species abundance (percent cover) was calculated by summing the percent covers of all understory species within each plot, including those present in both the shrub and herb or ground layers. The species richness was calculated as the total number of unique species recorded in each sub-plot and Shannon’s index was calculated by utilizing the percent cover proportions of the constituent species within each sample plot. These calculations were performed separately for all data pooled (total), the shrub and the herb layers. Since different ecological processes may regulate abundance and species diversity (Grace, 1999), we analyzed separate models for understory cover and species diversity. In order to determine the effects of altitudinal gradient and dominant overstory composition type on total, shrub and herb layer abundance, diversity indices, following general linear model was used:
Where, Yijk is the understory plant species cover, richness, or Shannon Index (separately analyzed by total, shrub and herb layers), μ is the overall mean, Ai is the altitudinal gradient (i = 1, 2, 3, 4), Cj is the dominant overstory composition types (j = 1, 2, 3), ɛk(ij) is sampling error among replicates within altitude and overstory combination type. The assumption of normality was assessed by Shapiro–Wilk test and the homogeneity of variance with Bartlett’s test. To mitigate the violation to the normality assumption and to improve coefficient estimates, we bootstrapped the fitted coefficients of linear models by using “ggplot2” (Wickham, 2009). We bootstrapped the 95% confidence intervals and considered estimates to be significantly different if their confidence intervals did not overlap others’ means by using the boot package in R (Canty and Ripley, 2017).
We used Permutation multivariate analysis of variance (perMANOVA) to examine the effect of altitudinal gradient and overstory composition types on the understory species composition. PerMANOVA is a nonparametric, multivariate method that uses permutation techniques to test for compositional differences between more than one variable (Anderson, 2005; Warton et al., 2012). To summarize variation in understory species composition, we used NMDS (non-metric multidimensional scaling) which is a robust ordination technique well suited for community data, because it preserves the rank order of dissimilarities among samples and avoids assumptions of normality and homogeneity of variance (Kruskal, 1964; McCune and Grace, 2002). All calculations were performed in the R statistical program using the adonis function in the “vegan” package (Oksanen et al., 2020). All the statistical analyses were performed in R statistical software (R Development Core Team, 2023).
3 Results
3.1 Understory vegetation abundance
A total of 99 understory species were recorded in the 36 sampled plots, which included 37 species in shrub layer and 62 species in the herb layer. Some species, present in the shrub layer were also present in the herb layer. Total abundance (percent cover) of understory vegetation and its components showed significant variation along the altitude and dominant overstory types along with the significant interaction between the two factors (Table 1). Altitude was found to be the primary factor influencing both the abundance and species diversity of understory vegetation. Total understory vegetation abundance, consisting of shrub and herb species, increased linearly from altitude A1 (1500-2000m) to altitude A2 (2000-2500m), peaked at altitude A2 and decreased thereafter at altitude A3 (2500-3000m) and altitude A4 (3000-3500m) (Figure 1). Among different overstory composition types, broadleaved stands had significantly higher total understory abundance at altitude A1 (1500-2000m) and altitude A2 (2000-2500m). Mixedwood stands, on the other hand had higher total understory abundance at altitude A3 (2500-3000m) and altitude A4 (3000-3500m). In contrast, coniferous stands consistently had the lowest total understory vegetation abundance across all altitudes (Table 1; Figure 1). The shrub abundance also followed the similar pattern to total understory abundance along the altitudinal gradient and dominant overstory composition types (Table 1; Figure 1). The shrub abundance showed an increase from altitude A1 (1500-2000m) to altitude A2 (2000-2500m), peaked at altitude A2 and declined thereafter at altitude A3 (2500-3000m) and altitude A4 (3000-3500m). Among the different overstory composition types, shrub abundance was maximum under the broadleaved stands at altitude A1 (1500-2000m) and altitude A2 (2000-2500m). However, at altitude A3 (2500-3000m) and altitude A4 (3000-3500m), shrub abundance was highest under mixedwood stands, followed by broadleaved and coniferous stands (Figure 1). The influence of altitudinal gradient and dominant overstory types on herb abundance mirrored the patterns observed for total and shrub abundance (Table 1; Figure 1). Herb abundance gradually increased from A1 (1500-2000m) to A2 (2000-2500m), peaked at A2, and then declined at A3 (2500-3000m) and A4 (3000-3500m), and broadleaved stands had higher herb abundance at altitude A1 (1500-2000m) to A2 (2000-2500m), while at altitude A3 (2500-3000m) and altitude A4 (3000-3500m), mixedwood showed higher herb abundance. However, coniferous stands consistently had the lowest herb abundance among the dominant overstory types along the altitudinal gradient (Figure 1).
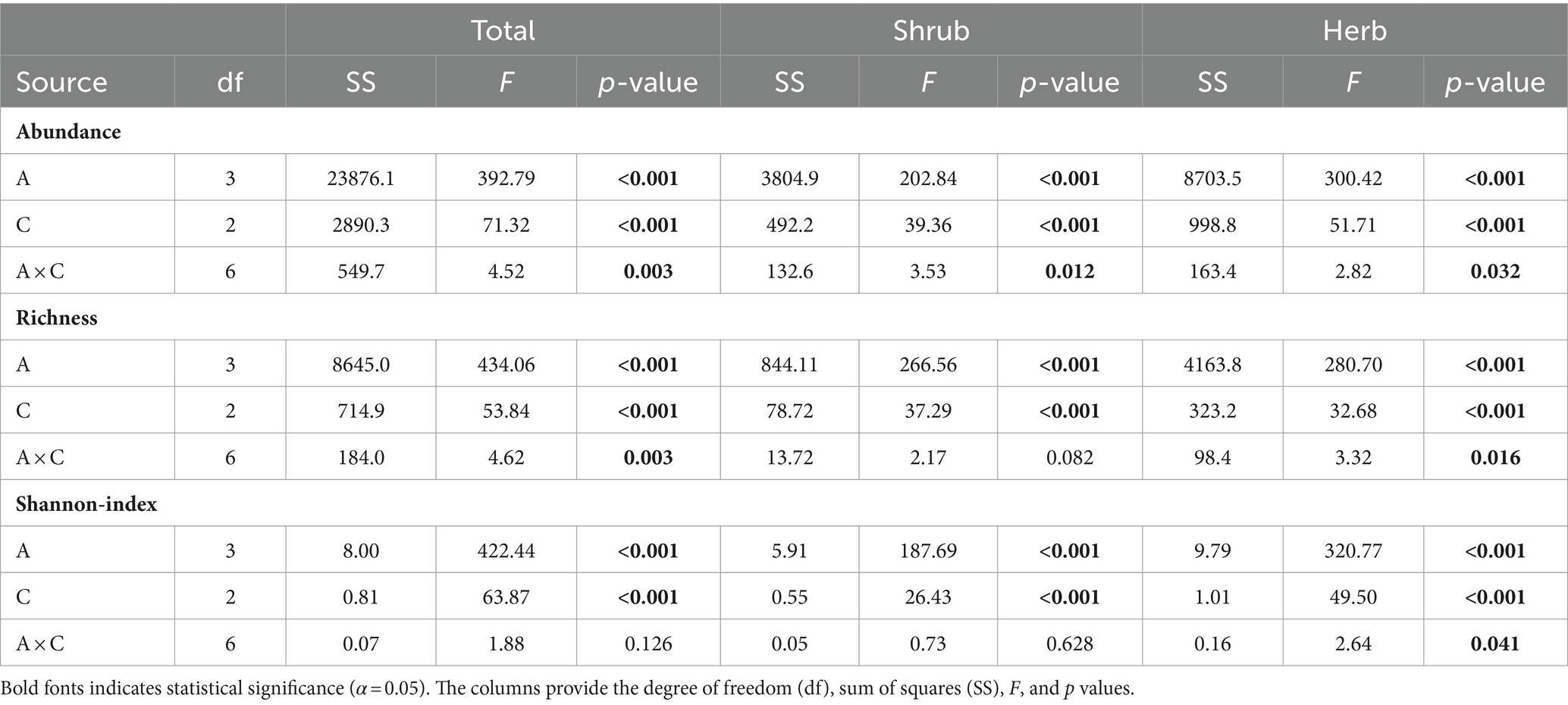
Table 1. Results of analysis of variance showing the effects of altitudinal gradients (A) and dominant overstory composition types (C) and their interactions (A × C) on understory plants abundance, species richness and diversity, separately analyzed by total, shrub and herb layers.
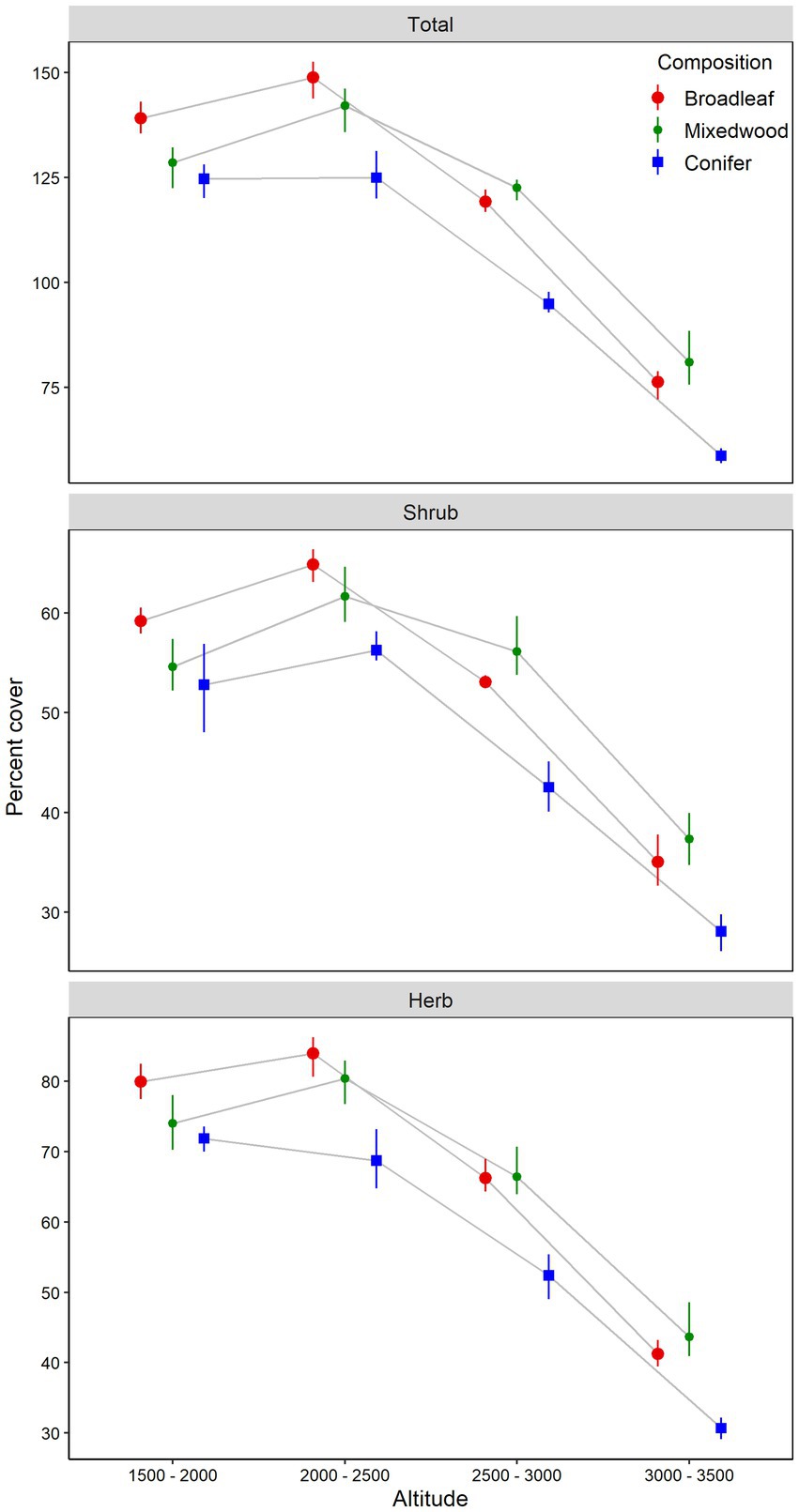
Figure 1. Understory plants total, shrub and herb layers cover (mean and bootstrapped 95% confident intervals) in relation to altitudinal gradients and overstory composition types. The difference between the altitudinal gradients or forest composition types is significant at α = 0.05 if their confidence intervals do not overlap the other’s mean.
3.2 Understory vegetation richness and diversity
The total understory vegetation richness exhibited the significant variation along the altitudinal gradient and dominant overstory types along with the significant interaction effects between the two factors (Table 1). Total understory vegetation richness increased linearly from altitude A1 (1500-2000m) to altitude A2 (2000-2500m), peaked at altitude A2 (2000-2500m) and decreased thereafter at altitude A3 (2500-3000m) to altitude A4 (3000-3500m). Among distinct overstory composition types, mixedwood stands had the higher total understory vegetation richness as compared to broadleaved and coniferous stands across all altitudinal gradients (Figure 2). The shrub richness also displayed the similar pattern to that of total richness (Table 1; Figure 2), showed gradual increase from altitude A1 (1500-2000m) to altitude A2 (2000-2500m), peaked and decreased thereafter at altitude A3 (2500-3000m) and altitude A4 (3000-3500m). Among the different dominant overstory types, shrub richness was found to be highest under mixedwood stands followed by broadleaved and coniferous stands (Figure 2). The herb species richness also followed the similar pattern to that of total and shrub richness along the altitudinal gradients and among different overstory composition types (Table 1; Figure 2).
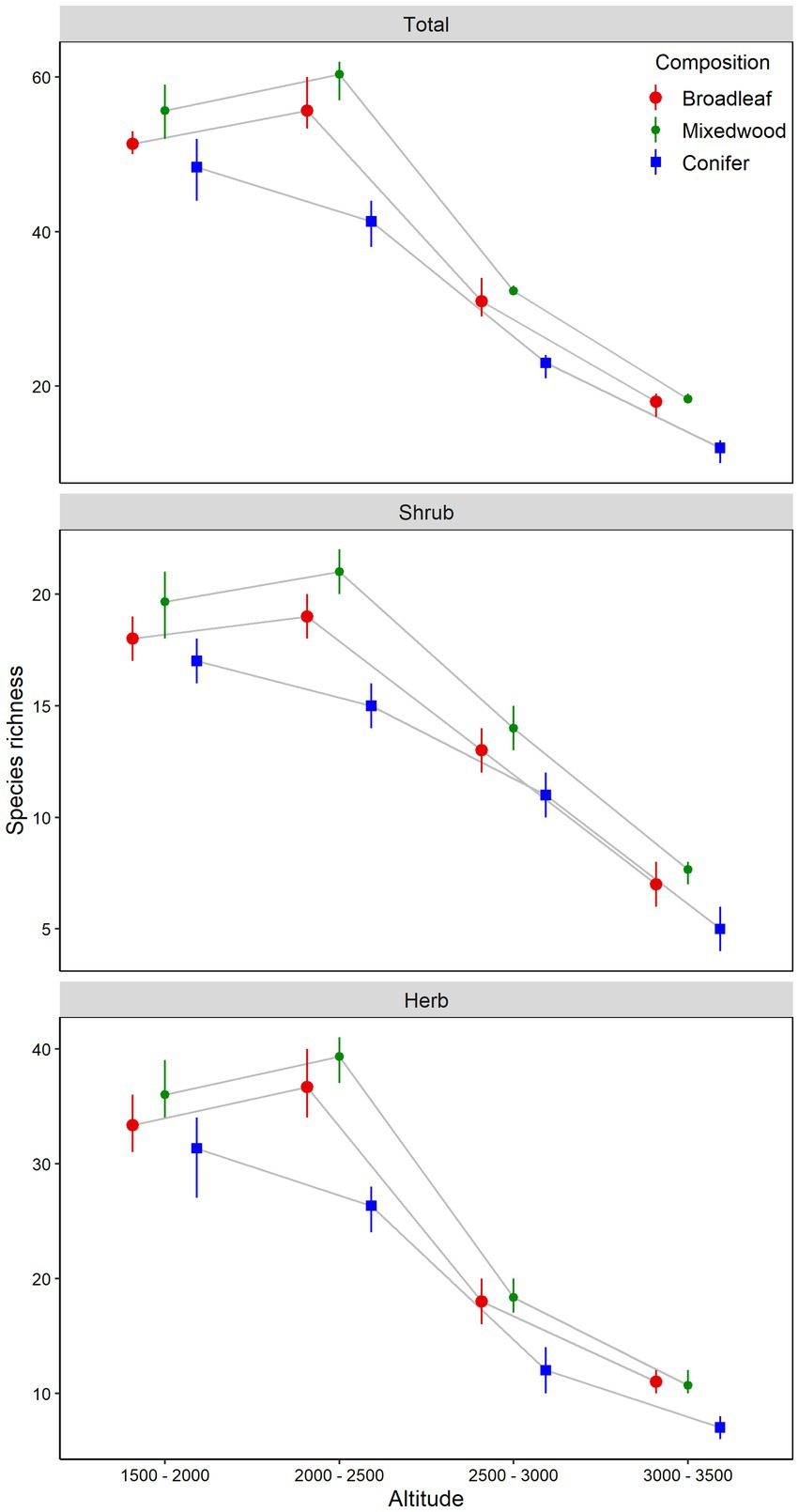
Figure 2. Understory plants total, shrub and herb layers species richness (mean and bootstrapped 95% confident intervals) in relation to altitudinal gradients and overstory composition types. The difference between the altitudinal gradients or forest composition types is significant at α = 0.05 if their confidence intervals do not overlap the other’s mean.
The total Shannon index also exhibited significant variation along the altitudinal gradient and dominant overstory types, with a significant interaction between the two factors (Table 1). The total Shannon index for understory vegetation displayed a linearly increasing pattern from altitude A1 (1500-2000m) to A2 (2000-2500m), peaking at A2, and then declining from altitude A3 (2500-3000m) to A4 (3000-3500m). Mixedwood stands showed a significantly higher total Shannon index than broadleaved stands, while coniferous stands showed the lowest total Shannon index value (Figure 3). Similarly, the shrub Shannon index also increased linearly from altitude A1 (1500-2000m) to altitude A2 (2000-2500m), peaked and subsequently decreased from altitude A3 (2500-2500m) to altitude A4 (3000-3500m). The shrub Shannon index was significantly higher under mixedwood stands compared to broadleaved and coniferous stands (Figure 3). The herb Shannon index also exhibited the similar pattern along the altitudinal gradient to those of total and shrub Shannon index and was consistently higher under mixedwood stands followed by broadleaved and coniferous stands (Figure 3).
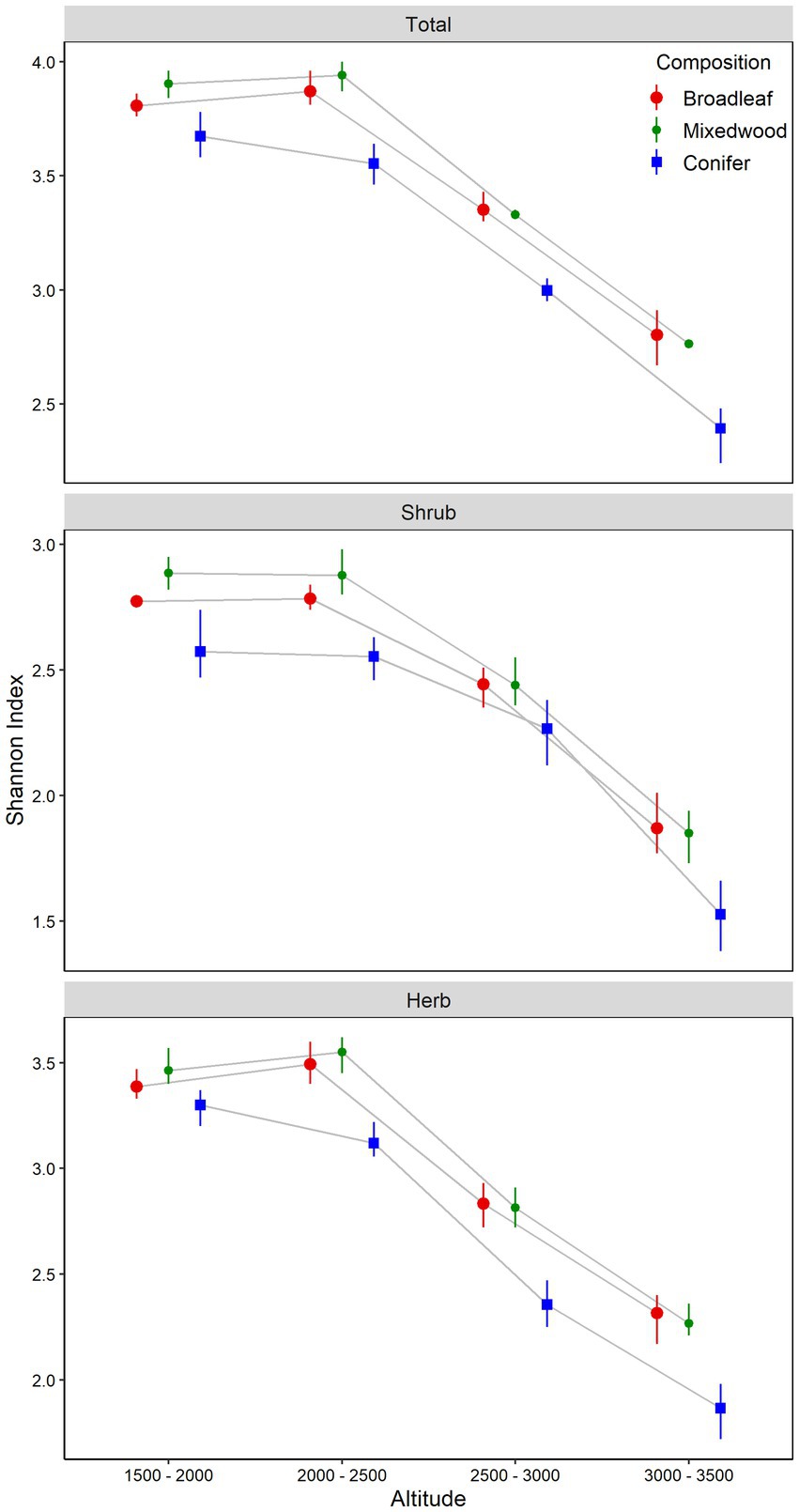
Figure 3. Understory plants total, shrub and herb layers Shannon Index (mean and bootstrapped 95% confident intervals) in relation to altitudinal gradients and overstory composition types. The difference between the altitudinal gradients or forest composition types is significant at α = 0.05 if their confidence intervals do not overlap the other’s mean.
3.3 Understory vegetation species composition
The results from PerMANOVA showed that the species composition of understory vegetation differed significantly along the altitudinal gradients and dominant overstory composition types (Table 2). Moreover, the effect of altitudinal gradient differed significantly with overstory composition types, as indicated by a significant interaction effect of altitudinal gradients and overstory composition types (Table 2). The NMDS ordination revealed distinct differentiation of understory species composition along altitudinal gradients (Figure 4). The understory vegetation communities were well-separated in the ordination space. Most notably, the understory vegetation communities at lower elevations showed distinct separation from those at higher elevations, while the understory vegetation communities at middle elevations exhibited intermediate vegetation characteristics to lower and higher elevations (Figure 4).
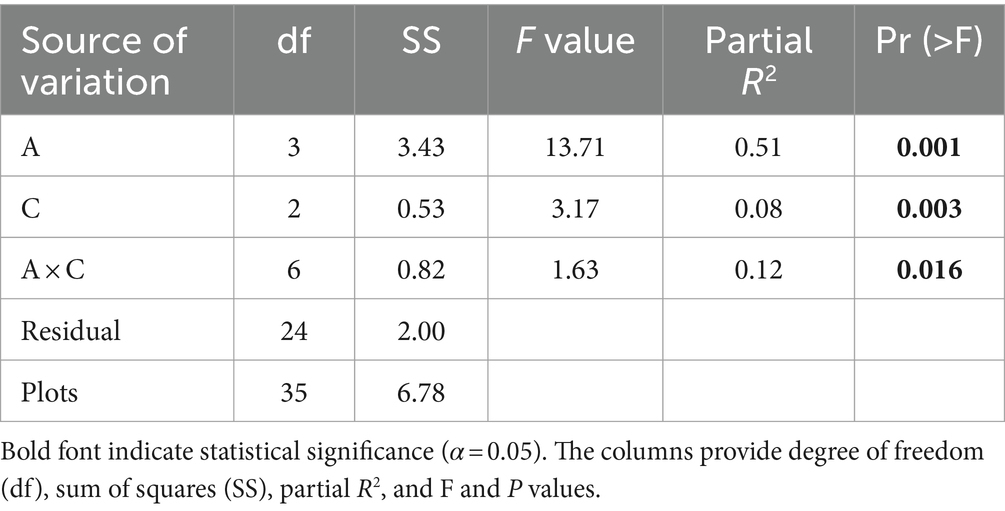
Table 2. Results of permutation multivariate analysis of variance (perMANOVA) testing the effects of altitudinal gradients (A) and overstory compositions (C) and their interactions (A × C) on understory species composition.
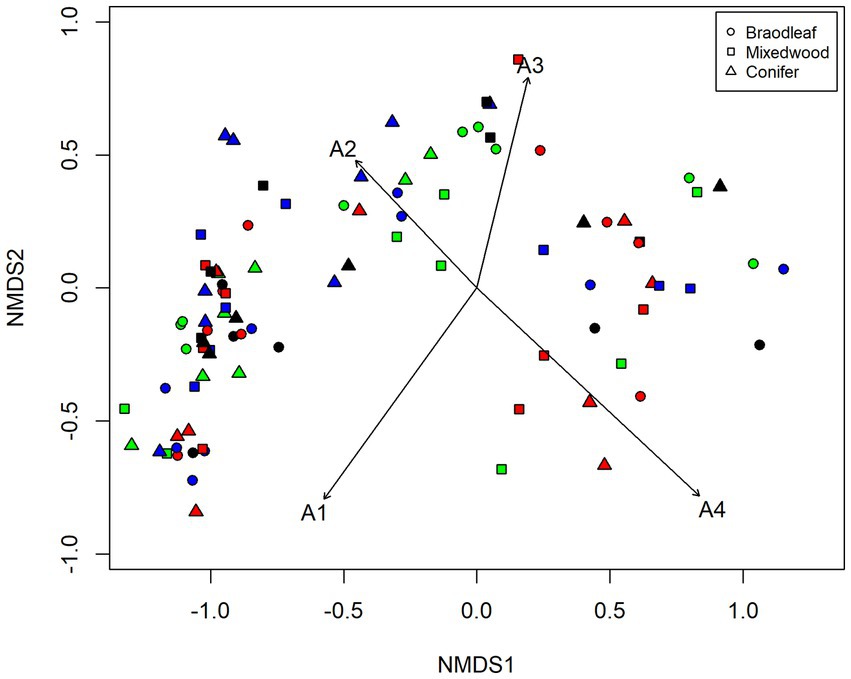
Figure 4. Two-dimensional non-metric multidimensional scaling ordination showing: the difference in understory plant communities along the altitudinal gradients and dominant overstory composition types. Altitudinal gradients are A1 (green), A2 (blue), A3 (red), A4 (black) and symbol shapes on legends differentiate dominant overstory types. Points nearest each other in ordination space have similar floristic assemblages, whereas those located farther apart are less similar. Vectors (arrows) indicate the significant (p < 0.05) joints axis correlation with altitude and overstory, and length of the vector represents the strength of the correlation.
At altitude A1, the dominant species in the shrub layer were Sarcococca coriacea, Rubus ellipticus, Punica granatum, Berberis lycium, Murraya koenigii, Pyrus pashia, Duranta repens, Zanthoxylum armatum, Vitex negundo, Desmodium tiliaefolium; and in the herb layer were Cheilanthes bicolor, Jasminum officinale, Cystopteris fragilis, Cynodon dactylon, Adiantum caudatum, Rubus ellipticus (Table 3). However, the altitude A2, in the shrub layer was characterized by Sarcococca coriacea, Spiraea canescens, Murraya koenigii, Jasminum officinale; while in the herb layer was Dryopteris filix, Pteris vittata, Thelypteris arida, Asplenium dalhousieae, Selaginella Kraussiana, Jasminum officinale, Murraya koenigii, Dicanthium annulatum etc. (Table 3). At altitude A3, in the shrub layer Rosa moschata, Rhododendron arboretum, Sarcococca coriacea, Spiraea canescens were dominant and in the herb layer Pteris vittata, Oxalis corniculata, Asplenium dalhousieae, Thelypteris arida, Fragaria nubicola, Cynodon dactylon were the dominant species (Table 3). The altitude A4 was characterized by Sarcococca coriacea, Rubus ellipticus, Rhododendron arbreum, Berberis lycium in the shrub layer, whereas Selaginella Kraussiana, Adiantum caudatum, Pteris vittata, Dryopteris filix were most dominant in the herb layer (Table 3). Pertaining to the three distinct overstory composition types, no distinct grouping was found among the different overstory types as they were showing species composition common to broadleaf, mixedwood and coniferous stands. The broadleaf stands were characterized by Sarcococca coriacea, Rubus ellipticus, Berberis lycium, Rosa moschata and Pyrus pashia in the shrub layer, while Asplenium dalhousieae, Pteris vittata, Thelypteris arida, Andropogon munroi, Dicanthium annulatum were dominant species in the herb layer (Table 3). In the mixedwood forests, Sarcococca coriacea, Berberis lycium, Rubus ellipticus and Pyrus pashia were the most frequent species in the shrub layer, while Adiantum caudatum, Onychium japonicum, Athyrium filix-femina, Asplenium dalhousieae, Polystichum aculeatum, Cystopteris fragilis were in the herb layer (Table 3). Whereas, in the coniferous stands, the dominant species in the shrub layer were Rubus ellipticus, Pyrus pashia, Sarcococca coriacea, and Rosa moschata, while in the herb layer were Selaginella Kraussiana, Dryopteris filix, Adiantum caudatum, Thelypteris arida, Cystopteris fragilis, and Pteris vittata (Table 3).
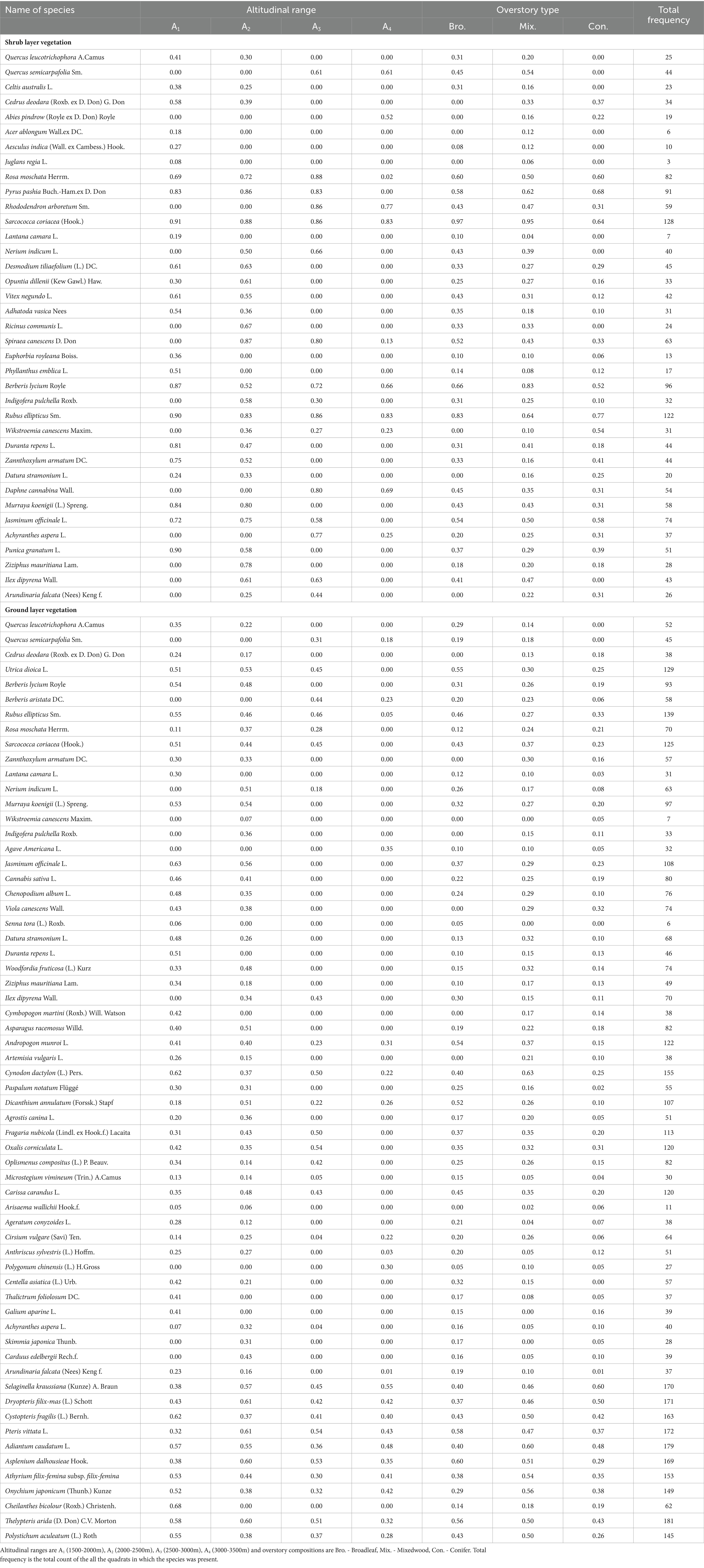
Table 3. The relative frequency of understory plants including shrub and herb layers vegetation at each altitudinal gradient and overstory composition type in the studied area.
4 Discussion
Our study provides a comprehensive assessment of the distribution patterns of understory vegetation abundance, plant diversity and species composition along the altitudinal gradient and in relation to forest compositional types in the temperate region of Himachal Pradesh-India. The results demonstrated significant variations in understory vegetation abundance, diversity, and composition based on altitude and dominant overstory composition types. The total, shrub layer and herb layer abundance and species diversity followed a hump shaped relationship along the altitudinal gradients which increased linearly from altitude A1 to A2, and decreased thereafter from altitude A3 to altitude A4 and were found to be highest at middle altitudes. These findings are consistent with the other studies in the Himalayan region along the elevation gradients (Grytnes and Vetaas, 2002; Grytnes and Beaman, 2006; Singh et al., 2007; Chawla et al., 2008; Sharma et al., 2009b; Acharya et al., 2011; Dar and Sundarapandian, 2016; Manish and Pandit, 2018; Sharma et al., 2019; Bhat et al., 2020; Rawat et al., 2021; Liang et al., 2023). However, the total number of species recorded in the present study is higher compared to those reported by other studies in the Western Himalayas (Sherman et al., 2008; Singh and Gupta, 2009; Gairola et al., 2012). The highest abundance and species diversity at the mid-altitude are likely the results of optimal climatic conditions, including temperature, rainfall, solar radiation, and humidity, favoring the growth and establishment of most vascular plants (Grytnes and Vetaas, 2002; Bhattarai and Vetaas, 2003; Zhang et al., 2016; Liang et al., 2020; Rawat et al., 2023). Although environmental factors were not explored in the present study, Liang et al. (2020) found that climatic factors, specifically temperature and precipitation, played a predominant role compared to other factors in shaping the richness of vascular plants along elevational gradients. Specifically, regions characterized by warm and wet climates generally support higher species diversity (Weins and Donoghue, 2004; Weins and Graham, 2005; Liang et al., 2020), highlighting the critical influence of climatic factors on species diversity along elevation gradients. Notably, both herbaceous and woody plants exhibited similar responses to these climatic conditions across elevation gradients. Furthermore, the water-energy dynamic hypothesis, proposed by Liang et al. (2020), suggests that the combined effects of energy (temperature) and water availability play a crucial role in shaping the diversity patterns of vascular plants along elevation gradients as water availability supports organismal activities, while energy regulates water states. This hypothesis provides additional insights into understanding the primary drivers shaping vascular plant richness along elevational gradients. Moreover, variations in environmental conditions along the altitudinal gradients significantly influence soil characteristics such as organic matter content, pH, and soil moisture, which in turn affect soil nutrient availability and have likely favored the growth and establishment of both herbaceous and woody plants in the understory (Decker and Boerner 2003; Kewlani et al., 2021). In addition to this, the higher abundance and species diversity at middle altitude can also be attributed to its intermediate location, forming a transition zone between low and higher elevations. This intermediate position increases immigration from both directions and enhances environmental heterogeneity, leading to additional species with distinct niche preferences (Grytnes and Vetaas, 2002; Singh et al., 2007; Scheiner et al., 2011; Zhang et al., 2016; Das et al., 2020).
However, the decrease in understory vegetation abundance and species diversity at higher altitudes can be attributed to the harsh environmental conditions, rugged topography, prolonged winter snow cover, and reduced growing season (Gómez-Díaz et al., 2017; Negi et al., 2018; Kewlani et al., 2021). These factors might support the dominance of site-specific endemic species that can potentially employ various strategies to overcome nutrient limitations and access necessary nutrients for growth under extreme environmental conditions, thereby forming species ‘fundamental niches’ (Lang et al., 2011; Graham et al., 2014; Wang et al., 2014; Coelho et al., 2018). Additionally, the slow rate of organic matter decomposition in harsh environmental conditions may have resulted in the accumulation of humus in the soil. This accumulation can limit soil production and reduce the absorption of nutrients by many herbaceous and woody plants (Prescott et al., 2000; Sanchez-Gonzalez and Lopez-Mata, 2005). On the other hand, lower understory vegetation abundance and species diversity at lower altitudes can be as a result of higher anthropogenic disturbances such as overgrazing, deforestation, trampling and camping etc. (Zhang et al., 2016; Rawal et al., 2018), which reduced species richness (Koh et al., 2006; Paudel and Sipos, 2014) at lower elevations.
The study also observed higher total, shrub and herb layer abundance under broadleaved stands at lower elevations compared to mixedwood and coniferous stands. The higher total, shrub, and herb layer cover under broadleaved stands could be attributed to higher light availability and the positive effect of broadleaf litter on soil quality, as broadleaf litter decomposes quickly and providing rich nutrients and higher pH (Hart and Chen, 2006; Chytrý et al., 2010; Bartels and Chen, 2013; Augusto et al., 2015; Kumar et al., 2017b), supporting specific species dominance in the ecosystem (Sundriyal and Sharma, 1996). Notably, the broadleaved stands in our study at lower elevations had a dense cover of tall shrubs such as, Sarcococca coriacea, Murraya koenigii, Zanthoxylum armatum, and invasive species Lantana camara as compared to mixedwood and conifer stands as these species flourish well in the warm and humid climatic conditions under broadleaved stands. Additionally, higher anthropogenic activities, such as deforestation and selective lopping of oak trees for fodder at lower elevations (Paudel and Sasaki, 2020; Joshi et al., 2021), might have led to canopy gaps, allowing greater light penetration to the forest floor, which in turn have facilitated the growth of tall shrubs and establishment of invasive species in the understory (Duncan et al., 1998; Arellano-Cataldo and Smith-Ramίrez, 2016).
However, higher total, shrub and herb layer abundance at higher elevations along with the higher species diversity along wide altitudinal gradients in the mixedwood stands could be attributed to the fact that mixedwood stands are characterized by greater resource heterogeneity in the understory (Messier et al., 1998; Hart and Chen, 2006; Barbier et al., 2008; Cavard et al., 2011) which allows for growth, development and facilitation of both herbaceous and woody vegetation (Kumar et al., 2017b). Furthermore, mixedwood stands are functionally and compositionally intermediate between conifer and broadleaved stands and have intermediate light penetration to the understory layer, thus facilitating the co-existence of both shade tolerant and intolerant plant species (Messier et al., 1998; Hart and Chen, 2006; Barbier et al., 2008; Cavard et al., 2011; Chavez and Macdonald, 2012; Mestre et al., 2017; Zhang et al., 2017). However, the lowest total abundance and species diversity in the shrub and herb layers under coniferous stands can be attributed to the negative effect of acidic and recalcitrant litter of conifers and the inability of the vascular species to tolerate low light and nutrient poor conditions under conifer canopies (Barbier et al., 2008; Hart and Chen, 2008; Chavez and Macdonald, 2010).
5 Conclusion
Our study demonstrated that the abundance (percent cover) and species diversity of understory vegetation differed along the altitudinal gradient and dominant overstory composition types. The hump shaped pattern along the altitudinal gradient appeared to be the most dominant pattern of understory vegetation abundance and species diversity and thus call for more conservation concern toward the middle elevation zones in the temperate Himalayan region. The species composition of understory vegetation also differed along the altitudinal gradients and dominant overstory composition types as well as with their interactions. Moreover, distinct understory vegetation communities were found at lower elevations than those at higher elevations, and middle elevations exhibited intermediate vegetation. The study also highlighted the importance of dominant overstory composition types in driving the abundance, species diversity and composition of understory vegetation in the Temperate Himalayan region. The higher resource conditions associated with broadleaved stands coupled with increased disturbance intensity, supported greater herbaceous abundance and growth of tall shrubs at lower elevations, while the heterogeneous resource conditions induced by the mixedwood stands have supported the higher understory species diversity and abundance at higher elevations. Therefore, management interventions should aim at maintaining diverse range of overstory composition types for conserving biodiversity and their ecological functions in the temperate Himalayan region. While the mechanisms proposed in this study provide a sufficient explanation for the understory vegetation pattern, further verification is required to understand how these hypothesized mechanisms, particularly climatic and edaphic factors, shape the abundance, diversity, and composition of understory vegetation in the temperate Himalayan region. Looking forward, future studies should investigate the mechanisms driving understory vegetation community patterns along altitudinal gradients and assess the influence of climate change on plant diversity patterns and their ecological functions in the temperate Himalayan region.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SS: Conceptualization, Formal analysis, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing, Data curation, Investigation. PK: Writing – review & editing, Validation, Supervision, Resources, Writing – original draft, Visualization, Software, Methodology, Formal analysis, Conceptualization. DB: Writing – review & editing, Validation, Supervision, Resources. KD: Writing – review & editing, Validation, Methodology, Formal analysis. PP: Software, Visualization, Writing – review & editing, Formal analysis, Data curation.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
The authors express their gratitude to the Head of the Department of Silviculture and Agroforestry at Y. S. Parmar University of Horticulture and Forestry for providing essential facilities during the study. Special thanks are extended to Geetanjali, Sashwat, and Jatin for their valuable assistance during the fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1420855/full#supplementary-material
Footnotes
References
Acharya, B. K., Sanders, N. J., Vijayan, L., and Chettri, B. (2011). Distribution patterns of trees along an elevation gradient of eastern Himalayas, India. Acta Oecol. 37, 329–336. doi: 10.1016/j.actao.2011.03.005
Anderson, M. J. (2005). Permutational multivariate analysis of variance. Department of Statistics. Auckland, New Zealand: University of Auckland.
Arellano-Cataldo, G., and Smith-Ramίrez, C. (2016). Establishment of invasive plant species in canopy gaps on Robinson Crusoe Island. Plant Ecol. 217, 289–302. doi: 10.1007/s11258-016-0570-4
Augusto, L., De Schrijver, A., Vesterdal, L., Smolander, A., Prescott, C., and Ranger, J. (2015). Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal species. Biol. Rev. 90, 444–466. doi: 10.1111/brv.12119
Baniya, C. B., Solhoy, T., Gauslaa, Y., and Palmer, M. W. (2010). The elevation gradient of lichen species richness in Nepal. The Lichnenologist. 42, 83–96.
Augusto, L., Dupouey, J. L., and Ranger, J. (2003). Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 60, 823–831. doi: 10.1051/forest:2003077
Barbier, S., Gosselin, F., and Balandier, P. (2008). Influence of tree species on understory vegetation diversity and mechanisms involved – a critical review for temperate and boreal forests. For. Ecol. Manag. 254, 1–15. doi: 10.1016/j.foreco.2007.09.038
Bartels, S. F., and Chen, H. Y. H. (2013). Interactions between overstory and understory vegetation along an overstory compositional gradient. Veg. Sci. 24, 543–552. doi: 10.1111/j.1654-1103.2012.01479.x
Bhat, J. A., Kumar, M., Negi, A. K., Todaria, N. P., Malik, Z. A., Pala, N. A., et al. (2020). Species diversity of woody vegetation along altitudinal gradient of the Western Himalayas. Glob. Ecol. Conserv. 24, e01302–e09894. doi: 10.1016/j.gecco.2020.e01302
Bhattarai, K. R., and Vetaas, O. R. (2003). Variation in plant species richness of different life forms along a subtropical elevation gradient in the Himalayas, East Nepal. Glob. Ecol. Biogeogr. 12, 327–340. doi: 10.1046/j.1466-822X.2003.00044.x
Bisht, M., Sekar, K. C., Mukherjee, S., Thapliyal, N., Bahukhandi, A., Singh, D., et al. (2022). Influence of anthropogenic pressure on the plant species richness and diversity along the elevation gradient of Indian Himalayan high-altitude protected areas. Front. Ecol. Evol. 10, 2296–701X. doi: 10.3389/fevo.2022.751989
Canty, A., and Ripley, B. (2017). Package ‘boot’. Available at: http://cran.r-project.org/web/packages/boot/index.html (Accessed July 03, 2024).
Cardinale, B., Duffy, J., Gonzalez, A., Hopper, D. U., Perrings, C., Venail, P., et al. (2012). Biodiversity loss and its impact on humanity. Nature 486, 59–67. doi: 10.1038/nature11148
Cavard, X., Bergeron, Y., Chen, H. Y. H., and Paré, D. (2011). Effect of forest canopy composition on soil nutrients and dynamics of understory: mixed canopies serve neither vascular nor bryophytes strata. Veg. Sci. 22, 1105–1119. doi: 10.1111/j.1654-1103.2011.01311.x
Chavez, V., and Macdonald, S. E. (2010). The influence of canopy patch mosaics on understory plant community composition in boreal mixedwood forest. For. Ecol. Manag. 259, 1067–1075. doi: 10.1016/j.foreco.2009.12.013
Chavez, V., and Macdonald, S. E. (2012). Partitioning vascular understory diversity in mixed wood boreal forests the importance of mixed wood canopies for diversity conservation. For. Ecol. Manag. 271, 19–26. doi: 10.1016/j.foreco.2011.12.038
Chawla, A., Rajkumar, S., Singh, K. N., Lal, B., Singh, R. D., and Thukral, A. K. (2008). Plant species diversity along an altitudinal gradient of Bhabha valley in the Western Himalayas. J. Mt. Sci. 5, 157–177. doi: 10.1007/s11629-008-0079-y
Chitale, V. S., Behera, M. D., and Roy, P. S. (2014). Future of endemic Flora of biodiversity hotpots in India. PLoS One 9:e115264. doi: 10.1371/journal.pone.0115264
Chytrý, M., Danihelka, J., Horsák, M., Kočí, M., Kubešová, S., Lososová, Z., et al. (2010). Modern analogues from the southern Urals provide insights into biodiversity change in the early Holocene forests of Central Europe. J. Biogeogr. 37, 767–780. doi: 10.1111/j.1365-2699.2009.02256.x
Coelho, M. S., Carneiro, M. A., Branco, C. A., Borges, R. A. X., and Fernandes, G. W. (2018). Species turnover drives β-diversity patterns across multiple spatial scales of plant-galling interactions in mountaintop grasslands. PLoS One 13:e0195565. doi: 10.1371/journal.pone.0195565
Costa, F. R. C., Magnusson, W. E., and Luizao, R. C. (2005). Mesoscale distribution patterns of Amazonian understory herbs in relation to topography, soil and watersheds. J. Ecol. 93, 863–878. doi: 10.1111/j.1365-2745.2005.01020.x
Dar, J., and Sundarapandian, S. (2016). Patterns of plant diversity in seven temperate forest types of Western Himalaya, India. J. Asia Pac. Biodivers 9, 280–292. doi: 10.1016/j.japb.2016.03.018
Das, D. S., Rawat, D. S., Maity, D., Dash, S. S., and Sinha, B. K. (2020). Species richness patterns of different life-forms along altitudinal gradients in the great Himalayan National Park, Western Himalaya, India. Taiwania 65, 154–162.
Decker, K. L. M., and Boerner, R. E. J. (2003). Elevation and vegetation influences on soil properties in Chilean Nothofagus forests. Revista Chilena de Historia Natural. 76, 371–381.
Duncan, R. P., Buckley, H. L., Urlich, S. C., Stewart, G. H., and Geritzlehner, J. (1998). Small-scale species richness in forest canopy gaps: the role of niche limitation versus the size of the species pool. J. Veg. Sci. 9, 455–460. doi: 10.2307/3237109
Gairola, S., Sharma, C. M., and Ghildiyal, S. K. (2012). Chemical properties of soils in relation to forest composition in moist temperate valley slopes of Garhwal Himalays, India. Environmentalist 32, 512–523. doi: 10.1007/s10669-012-9420-7
Geberhiwot, K., Demissew, S., Woldu, Z., Fekadu, M., Desalegn, T., and Teferi, E. (2019). Altitudinal changes in vascular plants richness, diversity, and distribution pattern in Abune Yosef mountain range, northern Ethiopia. Plant Divers. 41, 220–228. doi: 10.1016/j.pld.2019.06.005
Gilliam, F. S. (2007). The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57, 845–858. doi: 10.1641/B571007
Gómez-Díaz, J. A., Krömer, T., Kreft, H., Greold, G., Carvajal-Hernández, C. I., and Heitkamp, F. (2017). Diversity and composition of herbaceous angiosperms along gradient of elevation and forest-use intensity. PLoS One 12:e0182893. doi: 10.1371/journal.pone.0182893
Grace, J. B. (1999). The factors controlling species density in herbaceous plant communities: an assessment. Perspect. Plant Ecol. Evol. Syst. 2, 1–28. doi: 10.1078/1433-8319-00063
Gracia, M., Montane, F., and Pique, J. (2007). Overstory structure and topographic gradient determining diversity and abundance of understory shrub species in temperate forests in Central Pyrenees (NE Spain). For. Ecol. Manag. 242, 391–397. doi: 10.1016/j.foreco.2007.01.056
Graham, C. H., Carnaval, A. C., Cadena, C. D., Zamudio, K. R., Roberts, T. E., Parra, J. L., et al. (2014). The origin and maintenance of the montane diversity: integrating evolutionary and ecological processes. Ecography 37, 711–719. doi: 10.1111/ecog.00578
Grytnes, A., and Beaman, J. H. (2006). Altitudinal species richness patterns for vascular plants on mount Kinabalu, Borneo. J. Biogeogr. 33, 1838–1849. doi: 10.1111/j.1365-2699.2006.01554.x
Grytnes, J. A., and Vetaas, O. R. (2002). Species richness and altitude: a comparison between simulation models and interpolated plant species richness along the Himalayan altitude gradient. Nepal. Am. Nat. 159, 294–304. doi: 10.1086/338542
Haq, S. M., Waheed, M., Darwish, M., Siddiqui, M. H., Goursi, U. H., Kumar, M., et al. (2024). Biodiversity and carbin stocks of the understory vegetation as indicators for the forest health in the Zabarwan Mountain range, Indian Western Himalaya. Ecol. Indic. 159, 1470–1160.
Hardwick, S. R., Toumi, R., Pfeifer, M., Turner, E. C., Nilus, R., and Ewers, R. M. (2015). The relationship between leaf area index and microclimate in tropical forest and oil palm plantation: forest disturbance drives changes in mircoclimate. Agric. For. Meteorol. 201, 187–195. doi: 10.1016/j.agrformet.2014.11.010
Hart, S. A., and Chen, H. Y. H. (2006). Understory vegetation dynamics of north American boreal forests. Crit. Revi. Plant Sci. 25, 381–397. doi: 10.1080/07352680600819286
Hart, S. A., and Chen, H. Y. H. (2008). Fire, logging, and overstory effect understory abundance, diversity, and composition in boreal forests. Ecol. Monogr. 78, 123–140. doi: 10.1890/06-2140.1
Ishii, H., Azuma, W., and Nabeshima, E. (2013). The need for a canopy perspective to understand the importance of phenotypic plasticity for promoting species coexistence and light-use complementarity in forest ecosystem. Ecol. Res. 28, 191–198. doi: 10.1007/s11284-012-1025-6
Joshi, V. C., Sundriyal, R. C., and Arya, D. (2021). Forest floor diversity, distribution and biomass pattern of oak and Chir-pine forest in the Indian Western Himalaya. Indian J. Ecol. 48, 232–237.
Kewlani, P., Negi, V. S., Bhatt, I. D., Rawal, R. S., and Nandi, S. K. (2021). Soil nutrients concentration along altitudinal gradient in Indian Western Himalayas. Scand. J. For. Res. 36, 98–104. doi: 10.1080/02827581.2020.1871065
Koh, P., Lee, R., and Lin, R. (2006). Bird species richness patterns of northern Taiwan: primary productivity, human population density, and habitat heterogeneity. Divers. Distrib. 12, 546–554. doi: 10.1111/j.1366-9516.2006.00238.x
Körner, C. (2007). The use of altitude in the ecological research. Trends Ecol. Evol. 1, 569–574. doi: 10.1016/j.tree.2007.09.006
Kruskal, J. B. (1964). Nonmetric multidimensional scaling: a numerical method. Psychometrika 29, 115–129. doi: 10.1007/BF02289694
Kumar, P., Chen, H. Y. H., Searle, E. B., and Shahi, C. (2018). Dynamics of understory biomass, production and turnover associated with long-term overstory succession in boreal forest of Canada. For. Ecol. Manag. 427, 152–161. doi: 10.1016/j.foreco.2018.05.066
Kumar, P., Chen, H. Y. H., Thomas, S. C., and Shahi, C. (2017a). Effects of coarse woody debris on plant and lichen species composition in boreal forests. J. Veg. Sci. 28, 389–400. doi: 10.1111/jvs.12485
Kumar, P., Chen, H. Y. H., Thomas, S. C., and Shahi, C. (2017b). Linking resource availability and heterogeneity to understory species diversity through succession in boreal forest of Canada. J. Ecol. 106, 1266–1276. doi: 10.1111/1365-2745.12861
Kumar, M., Verma, A. K., and Garkoti, S. C. (2020). Lantana camara and Ageratina adenophora invasion alter the understory species composition and diversity of chir pine forest in Central Himalayas, India. Acta Oecol. 109:103642. doi: 10.1016/j.actao.2020.103642
Lang, C., Seven, J., and Polle, A. (2011). Host preferences and differential contribution of deciduous tree species shape mycorrhizal species richness in a mixed central European forest. Mycorrhiza 21, 297–308. doi: 10.1007/s00572-010-0338-y
Liang, J., Ding, Z., Lie, G., Zhau, Z., Zhang, Z., and Hu, H. (2023). Pattern and drivers of phylogenetic diversity of seed plants along an altitudinal gradient in Central Himalayas. Glob. Ecol. Conserv. 47, 2351–9894.
Liang, J., Ding, Z., Lie, G., Zhou, Z., Singh, P. B., Zhang, Z., et al. (2020). Species richness patterns of vascular plants and their drivers along an altitudinal gradient in the Central Himalayas. Glob. Ecol. Conserv. 24:e01279.
Maletha, A., Maikhuri, R., and Bargali, S. (2020). Forest structure, diversity and regeneration pattern along altitudinal gradient in temperate zone of Nanda Devi biosphere reserve (a world heritage site). India: Western Himalaya.
Maletha, A., Maikhuri, R. K., Bargali, S. S., Sharma, A., Negi, V. S., and Rawat, L. S. (2022). Vegetation dynamics and soil nutrient availability in a temperate forest along altitudinal gradient of Nanda Devi biosphere reserve, Western Himalaya, India. PLoS One 17:e0275051. doi: 10.1371/journal.pone.0275051
Manish, K., and Pandit, M. K. (2018). Phylogenetic diversity, structure and diversification patterns of endemic plants along the elevationl gradient in the eastern Himalayas. Plant Ecol. Divers. 11, 501–513. doi: 10.1080/17550874.2018.1534147
McCain, C. M. (2007). Could temperature and water availability drive altitudinal species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 16, 1–13. doi: 10.1111/j.1466-8238.2006.00263.x
McCune, B., and Grace, J. B. (2002). Analysis of ecological communities. Gleneden Beach, Oregon: MjM Software Design.
McVicar, T. R., and Korner, C. (2013). On the use of elevation, altitude and height in the ecological and climatological literature. Oecologia 171, 335–337. doi: 10.1007/s00442-012-2416-7
Messier, C., Parent, S., and Bergeron, Y. (1998). Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests. J. Veg. Sci. 9, 511–520. doi: 10.2307/3237266
Mestre, L., Toro-Manríquez, M., Soler, R., Huertas-Herrera, A., Martínez-Pastur, G., and Lencinas, M. V. (2017). The influence of canopy-layer composition on understory plant diversity in southern temperate forests. For. Ecosyst. 4:6. doi: 10.1186/s40663-017-0093-z
Mueller-Dombois, D., and Ellenberg, H. (1974). Aims and methods of vegetation ecology. New York, United States: John Wiley and Sons.
Naeem, S. (2002). Ecosystem consequences of biodiversity loss: the evolution of a paradigm. Ecology 83, 1537–1552. doi: 10.1890/0012-9658(2002)083[1537:ECOBLT]2.0.CO;2
Negi, V. S., Kewlani, P., Pathak, R., Bhatt, D., Bhatt, I. D., Rawal, R. S., et al. (2018). Criteria and indicators for promoting cultivation and conservation of medicinal and aromatic plants in Western Himlaya, India. Ecol. Indic. 93, 434–446. doi: 10.1016/j.ecolind.2018.03.032
Nilsson, M. C., and Wardle, D. A. (2005). Understory vegetation as forest ecosystem drivers: evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 3, 421–428. doi: 10.1890/1540-9295(2005)003[0421:UVAAFE]2.0.CO;2
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P., O’Hara, R., et al. (2012). Vegan: Community ecology package. R Package Version 2.0–2. 2012.
Oksanen, J., Blanchett, F. G., Kindt, R., Legender, P., Minchin, P. R., O’Hara, R. B., et al. (2020). Vegan: Community Ecology Package. R package version 2. 5–7. Available at: https://www.researchgate.net/publication/282247686_Vegan_Community_Ecology_Package_R_package_version_20–2 (Accessed July 3, 2024).
Paudel, A., and Sasaki, N. (2020). Assessment of carbon stocks in oak forests along the altitudinal gradient: A case study in Punchase conservation area in Nepal, p. 23.
Paudel, P. K., and Sipos, J. (2014). Conservation status affects elevational gradient in bird diversity in the Himalaya: a new prespective. Glob. Ecol. Conserv. 2, 338–348. doi: 10.1016/j.gecco.2014.10.012
Pelt, R. V., and Franklin, J. F. (2000). Influence of canopy structure on the understory environment in tall, old-growth, conifer forests. Can. J. For. Res. 30, 1231–1245. doi: 10.1139/x00-050
Ping, C. L., Michaelson, G. J., Stiles, C. A., and González, G. (2013). Soil characterstics, carbon stores and nutrient distribution in eight forests types along an altitudinal gradient, eastern Puerto Rico. Ecol. Sci. 54, 67–86.
Prescott, C. E., Zabek, L. M., Stanley, C. L., and Kabzema, R. (2000). Decomposition of broadleaf and needle litter in forest of British Columbia: influences of litter type, forest types and litter mixture. Can. J. For. Res. 30, 1742–1750. doi: 10.1139/x00-097
R Development Core Team . (2023). R studio: Integrated development environment for R. Posit Software, PBC, Boston, MA. Available at: http://www.posit.co/ (Accessed July 03, 2024).
Rana, S. K., Rawal, R. S., Dangwal, B., Bhatt, I. D., and Price, T. D. (2021). 200 years of research on Himalayan biodiversity: trends, gaps, and policy implications. Front. Ecol. Evol. 8:603422. doi: 10.3389/fevo.2020.603422
Rawal, R. S., Rawal, R., Rawat, B., Negi, V. S., and Pathak, R. (2018). Plant species diversity and rarity patterns along altitude range covering treeline ecotone in Uttrakhand: conservation implications. Trop. Ecol. 59, 225–239.
Rawat, D. S., Bagri, A. S., Parveen, M., Nautiyal, M., Tiwari, P., and Tiwari, J. K. (2021). Pattern of species richness and floristic spectrum along the elevation gradient: a case study from Western Himalaya, India. Acta Ecol. Sin. 2021, 545–551. doi: 10.1016/j.chnaes.2021.03.012
Rawat, B., and Negi, A. S. (2021). Plant diversity patterns along environmental gradient in Nanda Devi biosphere reserve, west Himalaya. Trop. Ecol. 62, 61–70. doi: 10.1007/s42965-020-00122-5
Rawat, Y. S., Negi, V. S., Moussa, I. M., Zaman, W., and Elansary, H. O. (2023). Diversity, distribution and vegetation assessment of woody plant species in the cold desert environment, North-Western Himalaya, India. Sustain. For. 15:10429. doi: 10.3390/su151310429
Roberts, M. R., and Gilliam, F. S. (2003). “Response of the herbaceous layer to disturbance in eastern forests” in The herbaceous layer in forests of eastern North America. eds. F. S. Gilliam and M. R. Roberts (New York: Oxford University Press), 302–320.
Sanchez-Gonzalez, A., and Lopez-Mata, L. (2005). Plant species richness and diversity along the altitudinal gradient in the Sierra Nevada, Mexico. Divers. Distrib. 11, 567–575. doi: 10.1111/j.1366-9516.2005.00186.x
Sankaran, M., and Dinesh, M. R. (2020). Biodiversity of tropical fruits and their conservation in India. J. Hortic. Sci. 15, 107–126. doi: 10.24154/jhs.v15i2.894
Scheiner, S. M., Chairucci Fox, G. A., Helmus, M. R., McGlinn, D. J., and Willing, M. R. (2011). The underpinnings of the relationship of species richness with space and time. Ecol. Monogr. 81, 195–213. doi: 10.1890/10-1426.1
Sekar, K. C., Thapliyal, N., Pandey, A., Joshi, B., Mukherjee, S., Bhojak, P., et al. (2023). Plant species diversity and density patterns along altitude gradient covering high-altitude alpine region of West Himalayas, India. Geol. Ecol. Landsc. 1, 1–15. doi: 10.1080/24749508.2022.2163606
Sharma, N., Behera, M. D., Das, A. P., and Panda, R. M. (2019). Plant species richness in an elevation gradient in the eastern Himalaya. Biodivers. Conserv. 28, 2085–2104. doi: 10.1007/s10531-019-01699-7
Sharma, C. M., Ghildiyal, S. K., Gairola, S., and Suyal, S. (2009a). Vegetation structure, composition and diversity in relation to the soil characteristics of temperate mixed broad-leaved forest along the altitudinal gradient in the Garhwal Himalaya. Indian J. Sci. Technol. 2, 39–45. doi: 10.17485/ijst/2009/v2i7.11
Sharma, C. M., Suyal, S., Gairola, S., and Ghildiyal, S. K. (2009b). Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal Himalaya. Am. J. Sci. 5, 119–128.
Sherman, R., Mullen, R., and Haomin, L. (2008). Spatial patterns of the plant diversity and communities in alpine ecosystems of the Hengduan Mountains, Northwest Yunnan, China. J. Plant Ecol. 1, 117–136. doi: 10.1093/jpe/rtn012
Singh, A., and Gupta, N. K. (2009). Assessment of floristic diversity and regeneration status of Cedrus deodara (Roxb.) loud. Stands under forest management systems in Western Himachal Himalayas: a case study of Shimla district. Indian J. For. 32, 45–54. doi: 10.54207/bsmps1000-2009-H2E642
Singh, K. N., Lal, B., Singh, R. D., Todaria, N. P., and Ahuja, P. S. (2007). Species richness, distribution pattern and conservation status of higher plants in the Spiti cold desert of trans-Himalayan, India. Int. J. Biodivers. Sci. Manag. 3, 223–233. doi: 10.1080/17451590709618176
Sinha, S., Badola, H. K., Chhetri, B., Gaira, K. S., Lepcha, J., and Dhyani, P. P. (2018). Effect of altitude and climate in shaping the forest compositions of Singalila National Park in Khangchendzonga landscape, eastern Himalaya, India. J. Asia Pac. Biodivers 11, 267–275. doi: 10.1016/j.japb.2018.01.012
Su, X., Li, S., Wan, X., Huang, Z., Liu, B., Fu, S., et al. (2021). Understory dynamics of Chinese fir plantations and natural secondary forests of subtropical China. For. Ecol. Manag. 483, 378–1127.
Sundriyal, R. C., and Sharma, E. (1996). Anthropogenic pressures on tree structure and biomass in the temperate forest of Mamlay watershed in Sikkim. For. Ecol. Manag. 81, 113–134. doi: 10.1016/0378-1127(95)03657-1
Thakur, G., Kumar, P., Bhardwaj, D. R., Prakash, P., and Kanwar, P. (2024). Dynamics of aboveground vegetation biomass and carbon stocks along the altitudinal gradients and overstory composition types in the temperate Himalayan region. Trees For. People. 2024:100553. doi: 10.1016/j.tfp.2024.100553
Tinya, F., and Ódor, P. (2016). Congruence of the spatial pattern of light and understory vegetation in an old-growth, temperate mixed forest. For. Ecol. Manag. 381, 84–92. doi: 10.1016/j.foreco.2016.09.027
Tonteri, T., Salemaa, M., Rautio, P., Hallikainen, V., Korpela, L., and Merilä, P. (2016). Forest management regulates temporal change in the cover of boreal plant species. For. Ecol. Manag. 381, 115–124. doi: 10.1016/j.foreco.2016.09.015
Valladares, F., Laanisto, L., Niinemets, Ü., and Zavala, M. A. (2016). Shedding light on shade: ecological perspectives of understory plant life. Plant Ecol. Divers. 9, 237–251. doi: 10.1080/17550874.2016.1210262
Van Oijen, D., Feijen, M., Hommel, P., Den Ouden, J., and De Waal, R. (2005). Effects of tree species composition on within-forest distribution of understory species. Appl. Veg. Sci. 8, 155–166.
Wang, S. X., Guo, H., Wang, X. A., Fan, W. Y., and Duan, R. Y. (2014). Multiple-scale assessment of the understory herb species diversity in pine plantations after long-term restoration. Nord. J. Bot. 32, 680–688. doi: 10.1111/j.1756-1051.2013.00268.x
Wang, C. T., Long, R. J., Ding, L. M., Wang, Q. J., and Wang, M. P. (2007). Effects of altitude on plant-species diversity and productivity in an alpine meadow, Qinghai-Tibetan plateau. Aust. J. Bot. 55, 110–117. doi: 10.1071/BT04070
Wani, Z. A., Khan, S., Bhat, J. A., Malik, A. H., Alyas, T., Pant, S., et al. (2022). Patterns of β-diversity and plant species richness along vertical gradient in northwest Himalaya, India. Biology 11:1064. doi: 10.3390/biology11071064
Warton, D. I., Wright, S. T., and Wang, Y. (2012). Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 3, 89–101. doi: 10.1111/j.2041-210X.2011.00127.x
Weins, J. J., and Donoghue, M. J. (2004). Historical biogeography, ecology and species richness. Trends Ecol. Evol. 19, 639–644. doi: 10.1016/j.tree.2004.09.011
Weins, J. J., and Graham, C. (2005). Niche conservatism: integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 36, 519–539. doi: 10.1146/annurev.ecolsys.36.102803.095431
Whigham, D. E. (2004). Ecology of woodland herbs in temperate deciduous forests. Annu. Rev. Ecol. Evol. Syst. 35, 583–621. doi: 10.1146/annurev.ecolsys.35.021103.105708
White, A. E., Dey, K. K., Mohan, D., Stephens, M., and Price, T. D. (2019). Regional influences on community structure across the tropical-temperate divide. Nat. Commun. 14, 10(1):2646. doi: 10.1038/s41467-019-10253-6
Xu, M., Ma, L., Jia, Y., and Liu, M. (2017). Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS One 12:e0174231. doi: 10.1371/journal.pone.0174231
Yang, J. C., Hwang, H. S., Lee, H. J., Jung, S. Y., Ji, S. J., Oh, S. H., et al. (2014). Distribution of vascular plants along the altitudinal gradient of Gyebangsan (Mt.) in Korea. J. Asia Pac. Biodivers. 7, e40–e71. doi: 10.1016/j.japb.2014.03.008
Zhang, Y., Chen, H. Y. H., and Taylor, A. R. (2016). Aboveground biomass of understory vegetation has a negligible or negative association with overstory tree species diversity in natural forests. Glob. Ecol. Biogeogr. 25, 141–150. doi: 10.1111/geb.12392
Zhang, Y., Chen, H. Y. H., and Taylor, A. R. (2017). Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstory trees. Funct. Ecol. 31, 419–426. doi: 10.1111/1365-2435.12699
Keywords: abundance, altitudinal gradient, overstory composition, species diversity, temperate Himalayan region, understory vegetation, forest management
Citation: Sangry S, Kumar P, Bhardwaj DR, Dogra KS and (2024) Abundance, diversity and composition of understory plants along the altitudinal gradient and dominant overstory composition types in the temperate Himalayan region. Front. For. Glob. Change. 7:1420855. doi: 10.3389/ffgc.2024.1420855
Edited by:
Yashwant Singh Rawat, Federal Technical and Vocational Education and Training Institute, EthiopiaReviewed by:
Muhammad Waheed, University of Okara, PakistanZeeshan Ahmad, Chinese Academy of Sciences, China
Copyright © 2024 Sangry, Kumar, Bhardwaj, Dogra and Poonam. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Praveen Kumar, cGt1bWFydWhmQGdtYWlsLmNvbQ==; cGt1bWFydWhmQHlzcHVuaXZlcnNpdHkuYWMuaW4=