 Muhammad Waheed1*
Muhammad Waheed1* Shiekh Marifatul Haq2
Shiekh Marifatul Haq2 Fahim Arshad1
Fahim Arshad1 Rainer W. Bussmann2,3
Rainer W. Bussmann2,3 Abeer Hashem4
Abeer Hashem4 Elsayed Fathi Abd_Allah5
Elsayed Fathi Abd_Allah5- 1Department of Botany, University of Okara, Okara, Pakistan
- 2Department of Ethnobotany, Institute of Botany, Ilia State University, Tbilisi, Georgia
- 3Department of Botany, State Museum of Natural History, Karlsruhe, Germany
- 4Botany and Microbiology Department, College of Sciences, King Saud University, Riyadh, Saudi Arabia
- 5Plant Production Department, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia
Forest vegetation is an important component of forest ecosystems, contributing to terrestrial plant diversity while also providing a variety of ecological services. In managed landscapes, plantations emerge as dominant kinds after stand-replacing disturbances. However, the dynamics of vegetation cover, diversity, and composition in plantation forests remains poorly understood in the subtropical region. Our study recorded a rich floral diversity with 173 angiosperm species, characterized by varying life forms and distinct flowering phenology. The uneven distribution of species across families demonstrated the complexity of the ecosystem, with Poaceae being dominant. Diversity patterns among different plantation types varied, with Dalbergia sissoo and Populus nigra plantations exhibiting higher species richness and diversity. Conversely, Eucalyptus camaldulensis and Morus alba plantations displayed lower diversity, emphasizing the influence of plantation type on biodiversity. Non-metric multidimensional scaling (nMDS) and PERMANOVA analyses revealed significant dissimilarity patterns of vegetation composition. Indicator species analysis identified unique compositions within each plantation type, emphasizing the importance of conserving specific types to protect indicator species and maintained ecological distinctiveness. Canonical Correspondence Analysis (CCA) demonstrated that road accessibility, stem cutting, and fire significantly influenced plant distribution patterns. The present research underscored the importance of considering plantation type in forest management for biodiversity conservation and highlighted the environmental variables’ influence on the formation of plant communities. These results provided major implications for sustainable forest management and conservation efforts in tropical regions.
1 Introduction
Forest management as an important ecological indicator produces a considerable influence on plant species diversity (Ćosović et al., 2020; Haq et al., 2024a,b). Understanding how different forest management approaches affect plant species diversity is essential for achieving ecologically sustainable forest management (Oettel and Lapin, 2021). However, majority of the forest clearing already takes place in subtropical forests, and future deforestation is predicted to be concentrated in the tropics (Wright, 2010). To maintain biodiversity, wise and rigorous forestry management practices are urgently required to address diversity loss and the resultant degradation of ecosystem performance (Shin et al., 2022). Reforestation activities are supported all across the Earth to accommodate the expanding requirement for forest products, particularly in developing countries (Haq et al., 2023a,b). Subtropical forests thus encourage the preservation of forest cover in the region, as well as the development of ecosystem services that support local populations’ livelihoods (Borma et al., 2022). Understanding vegetation dynamics and ecological aspects is crucial in managed forests, where human activity influences vegetation composition and structure (Ivanova et al., 2022).
The composition of vegetation within managed forests is influenced by a myriad of factors, ranging from climatic conditions and soil characteristics to human activities such as silvicultural practices and land management (Ameray et al., 2021). Different tree species have varying growth rates, competitive abilities, and interactions with understory vegetation, resulting in distinct ecological traits that influence ecosystem dynamics (Balandier et al., 2022). The success of these plantations is measured not only in terms of economic productivity but also in terms of their ability to maintain or enhance ecological integrity (Karr et al., 2022). For instance, fast-growing pioneer species may establish dominance in early successional stages, altering nutrient cycling and microclimate conditions (Poorter et al., 2021). On the other hand, slower-growing, shade-tolerant species may shape the later successional stages, influencing habitat complexity and biodiversity (Khoja et al., 2022; Forrester et al., 2023).
In subtropical regions, the subtleties of climatic variability, including seasonal changes in temperature and precipitation, further complicate the dynamics of managed forests (Devi et al., 2023). Consequently, different plantation types may display varying responses to these climatic fluctuations, which can subsequently influence the interactions between plantation species and native vegetation (Liu et al., 2023). Furthermore, the establishment of plantations may involve the removal of native vegetation or alteration of natural hydrological regimes, potentially leading modulations in soil characteristics, water availability, and nutrient flux (Zhou et al., 2020). Such modifications can have cascading impacts on the overarching ecosystem structure and function, necessitating a thorough understanding of how vegetation composition and ecological traits interact within these managed landscapes (Brown et al., 2023). Furthermore, the impacts of different plantation types extend beyond the vegetation composition alone. They encompass a spectrum of ecological processes that influence nutrient cycling, water availability, microclimate conditions, and habitat suitability (Piotto et al., 2021). Native species may respond differently to these alterations, potentially leading to shifts in community structure and diversity (Williams and Newbold, 2020).
The implications of these changes extend to various trophic levels, affecting wildlife habitat, pollinator interactions, and overall ecosystem resilience (Murphy et al., 2020; Theodorou, 2022). Therefore, evaluating the sustainability and multifunctionality of managed forests in subtropical temperatures necessitates a thorough examination of the complex connections between vegetation composition, ecological traits, and ecosystem processes. Remedying this scholarly gap, our investigation was undertaken. in the Chichawatni subtropical managed forests with the specific objectives (1) assessing vegetation composition, growth forms, life span, phenology, nativity, and taxonomic groups of major forest types in the region, (2) exploring conceivable variances in diversity and Spatial arrangements across these forests, (iii) how do anthropogenic factors influence the diversity patterns of vegetation composition and communities assemblage in different forest types? The results would directly affect management plans and decision-making procedures to improve cooperative effectiveness in achieving global goals for preserving biodiversity. Moreover, conservation strategies targeted at the species level played a pivotal role in enhancing the resilience and sustainability of ecosystems.
2 Materials and methods
2.1 Study area
The Chichawatni forest is situated in the southern zone of Punjab, Pakistan, encompassing a geographical range from approximately 30°-29′-32.91″N to 30°-33′-45.84″N in latitude and 72°36′00.25″E to 72°46′48.65″E in longitude (Figure 1). This forest plantation lies at an elevation that varies between 153.6 and 163.7 meters above sea level. Its establishment as a man-made forest dates back to 1913, with the initial allocation of 4,726.73 hectares officially designated as the Chichawatni Reserved Forest. As of the present day, the total area of this plantation stands at 4,666.8 hectares, with a net available planting stock area spanning 3,823.20 hectares. Originally, the Chichawatni plantation exemplified a typical dry tropical forest ecosystem, characterized by its indigenous flora, including species like Salvadora oleoides, Tamarix aphylla, Prosopis cineraria, and Capparis aphylla. Irrigation for this forest was facilitated by the Lower Bari Doab Canal, with an extensive network of water courses channeling water from canal to the forest. In its current state, the plantation primarily composed of Morus alba, Eucalyptus camaldulensis, Populus nigra, Bombax ceiba, and Dalbergia sissoo, both in pure stands and mixed forms. Additionally, there are naturally occurring Neltuma juliflora trees interspersed throughout. Other scattered species found within the plantation boundaries comprised of Capparis aphylla, Tamarix aphylla, Ziziphus mauritiana, Prosopis cineraria, Albizia lebbeck and Albizia procera.
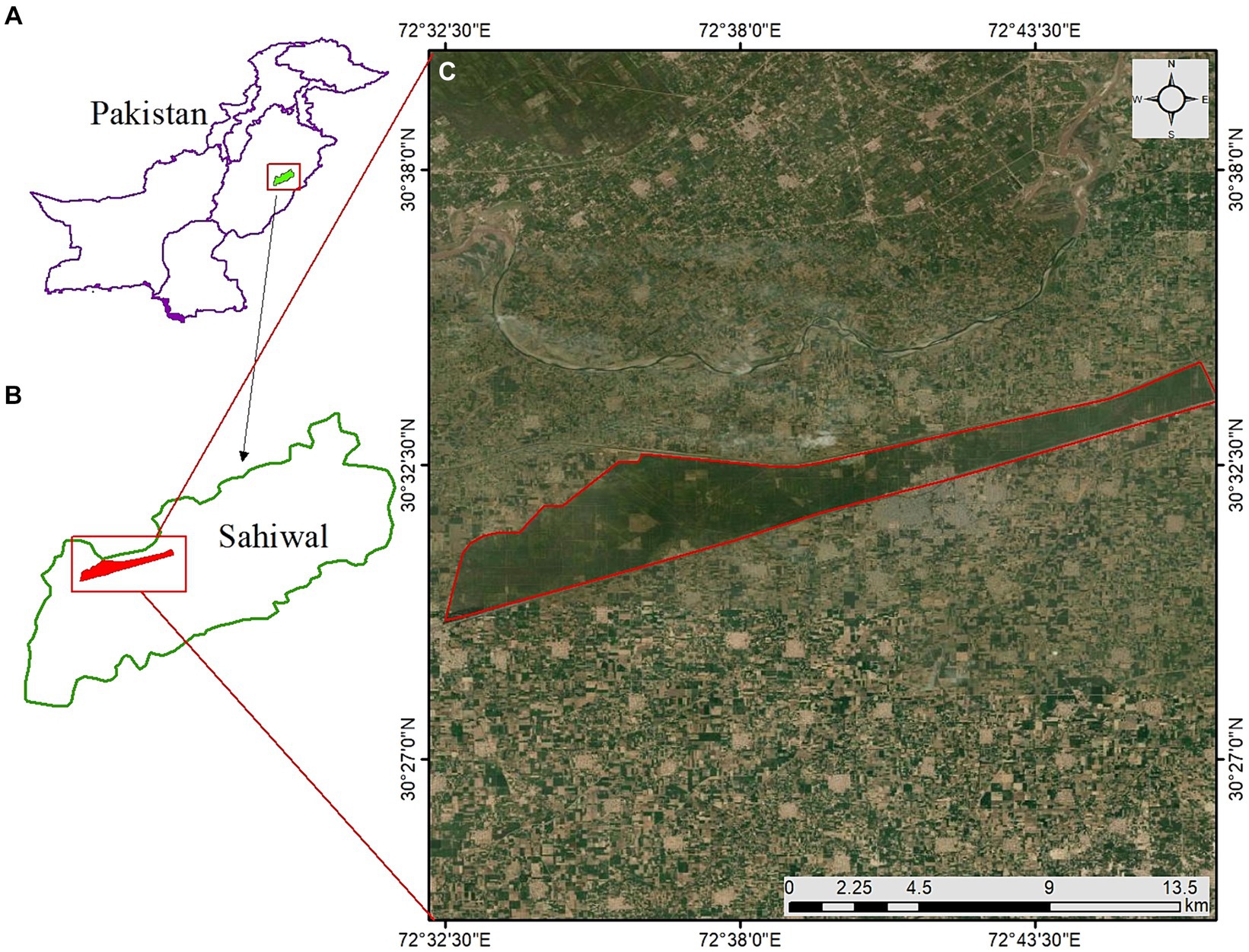
Figure 1. Study area map (A) Pakistan, (B) Sahiwal, (C) showing the location of Chichawatni forest.
2.2 Data collection
The comprehensive survey of the entire forest was conducted during the period spanning from 2022 to 2023. Based on a range of factors including plantation type, physiognomy, levels of disturbance, grazing pressure, stem cutting, and the age of the plantation, the study area was categorized into five distinct plantation types to analyze vegetation. A systematic sampling approach was employed to survey the vegetation within each of these designated plantation types. The investigation into vegetation composition was targeted at delineating five distinct plantation types, comprising four monoculture plantations and a singular plantation characterized by a mixed assembly of tree species, denoted as the Mixed Tree Plantation (MTP). The monoculture plantations under scrutiny encompassed a diverse array of tree species, namely the Populus nigra plantation (PNP), Dalbergia sissoo Plantation (DSP), Eucalyptus camaldulensis plantation (ECP), and Morus alba plantation (MAP). Within each plantation type, 10 plots measuring 30 m × 30 m were randomly allocated based on vegetation abundance. These plots were then further subdivided into five 5 m × 5 m quadrats for the collection of phytosociological data about shrub species. In addition, for herbaceous plants, 10 quadrats of 1 m × 1 m size were systematically placed within the selected plots. Primary vegetation data, encompassing Density, Frequency, and Cover, were systematically documented for each species at every site, by established standard methodologies outlined by Mueller-Dombois and Ellenberg (1974). To accurately identify the collected plant specimens, we relied on a combination of available literature and flora, including references such as the Flora of Pakistan, e-flora of China, Flowers of India, as well as works by Nasir and Ali (1970–1985), Stewart (1972), Ali And Qaiser (1986), Qazi et al. (2023) and online resources such as Kew online.1 To conduct a thorough assessment of the biological spectrum of plants, incorporating diverse life forms and leaf characteristics, Raunkiaer’s (1934) classification methodology was adopted. By employing such systematic framework, we were able to classify and examined the wide range of life forms and leaf characteristics displayed by the plant species under investigation. Additionally, to quantify the ecological importance of each species, we computed the Importance Value Index. The index was calculated by aggregating the relative values of Relative Density, Relative Frequency, and Relative Canopy Cover, following the approach outlined by Böcher and Bentzon (1958). Furthermore, the assessment of human disturbance was visually conducted for each sample plot. Degrees of grazing, stem cutting, fire, road accessibility, and biological invasion were appraised on a four-tiered scale. These visual assessments categorized the intensities of disturbance as follows: 0 for absent, 1 for low, 2 for moderate, and 3 for strong. These categories were established following methodologies outlined in prior studies by Abbas et al. (2017) and Nowak-Olejnik et al. (2020).
2.3 Data analysis
To scrutinize significant variations in species composition between different plantation types, a robust PERMANOVA analysis was conducted, bolstered by 999 permutations. Subsequently, SIMPER was employed to pinpoint the key species responsible for discriminating between the plantation types, elucidating the drivers behind diversity differences. All these analyses were executed using PAST software (version 4.12), as previously documented (Waheed et al., 2022a,b; Haq et al., 2024a). The study systematically analyzed four key diversity metrics in the study area, included the Shannon index (H), measuring species richness and evenness, and the Simpson index (1-D) calculating species dominance. The study compared such measures to determine whether vegetation diversity was due to a few dominant species (Simpson index) or many equally abundant species (Shannon diversity). The Dominance Index and Evenness were added to the analysis to better understand community structure. Canonical Correspondence Analysis (CCA), performed with CANOCO version 5.0 software, was employed to explore the complex interplay between plantation and type of disturbance. Alongside CCA, we strategically applied the Non-Metric Multi-Dimensional Scaling (NMDS) technique to address the high dimensionality of the dataset, a commonly utilized approach for assessing plant community similarity. The NMDS and chord diagrams were created using the Origin Pro software version 10.
3 Results
3.1 Floristic composition and ecological traits
The Chichawatni Forest boasted a rich floral diversity, harboring a total of 173 angiosperm species belonging to 42 different families, as exhibited in Supplementary Table S1. However, such distribution across families was not uniform, with six families representing half of the total species count, while the remaining 37 families shared the other half. Eighteen families contained just a single species each. The Chichawatni Forest stands out as a unique ecosystem within the broader irrigated region, characterized by its fertile terrain. That particular environment offered a stable range of climatic conditions and other variables, which in turn supported a remarkable diversity of species within a relatively compact area. Among the families, Poaceae occupied the largest, with 25 species (14.4%) of the total species count, followed by the Fabaceae family (12%), Asteraceae (9%), and Amaranthaceae (7.5%) of species (Supplementary Table S1).
Of the total number of species recorded, 49% (N = 85) were forbs, 19% (N = 33) trees, 7% (N = 24) trees, 14% (N = 25) graminoids, 8% (N = 14) shrubs, 5% (N = 9) climbers, and 4% (N = 7) sedges (Table 1). Most species were annuals (68%, N = 117) followed by perennials (27%, N = 47) and biennials (5%, N = 9) (Supplementary Table S1). The predominant tree species inhabiting the forest were deciduous (e.g., Dalbergia sissoo, Populus nigra, Bauhinia variegata, Morus alba, Morus nigra, Bombax ceiba, Cassia fistula, and Prosopis cineraria). The evergreen trees and shrubs were also frequent including Terminalia arjuna, Nerium oleander, Eucalyptus camaldulensis, Callistemon lanceolatus, Corymbia citriodora, and Salix tetrasperma, etc. The Raunkiaer biological spectrum analysis revealed that therophytes comprised the dominant life form, accounting for 44% (N = 76), followed by hemicryptophytes at 31% (N = 54), Phanerophytes 12% (N = 21), Chamaephytes 8% (N = 14), and Geophytes 5% (N = 8) (Figure 2). The most common leaf spectrum was microphyll (48%, N = 83) followed by mesophyll (23%, N = 40), nanophyll (20%, N = 35), leptophyll (5%, N = 9), macrophyll (9%, N = 29) megaphyll (2%, N = 4), and aphyllous (1%, N = 2) (Figure 3).
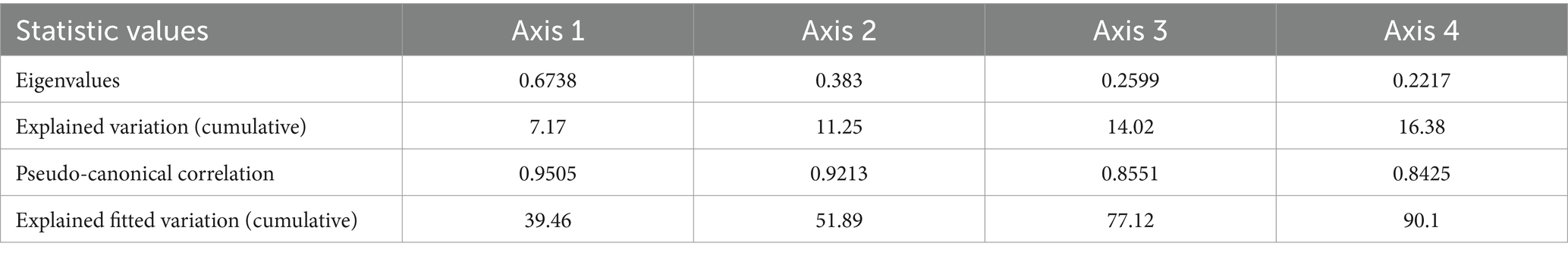
Table 1. Eigenvalues and explained variation of species distribution and environmental variables on the first four canonical correspondence analysis (CCA) ordination axes.
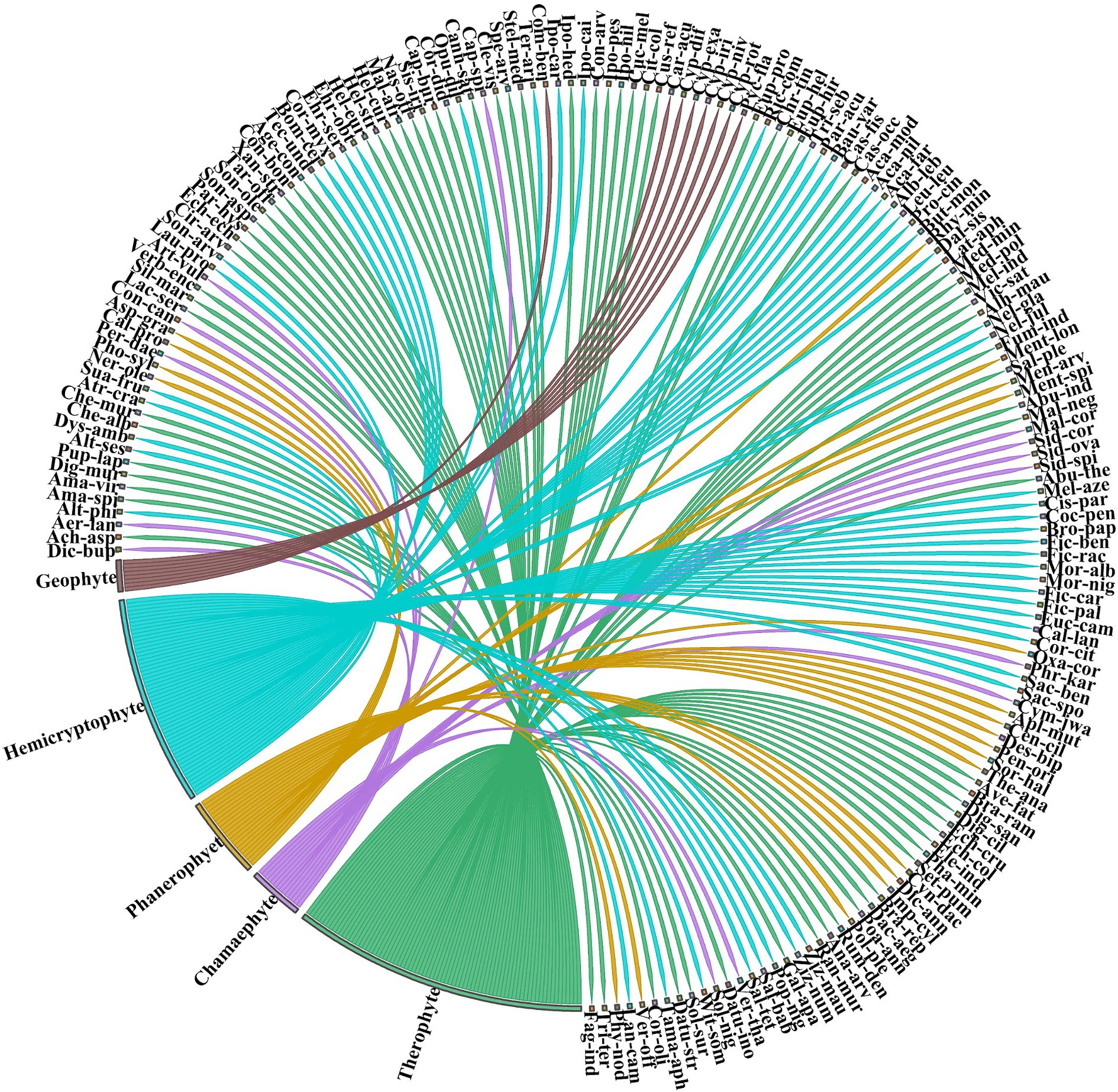
Figure 2. Distribution of species in Chichawatni Forest is classified using the Raunkiaer life form categorization. The direction of the lines in a graphical depiction reflects the association of each species with certain life form categories, while the thickness of each bar represents the number of species within each individual life form category. Table 1 contains a complete list of species names related to their life form types. For full species names see Supplementary Table S1.
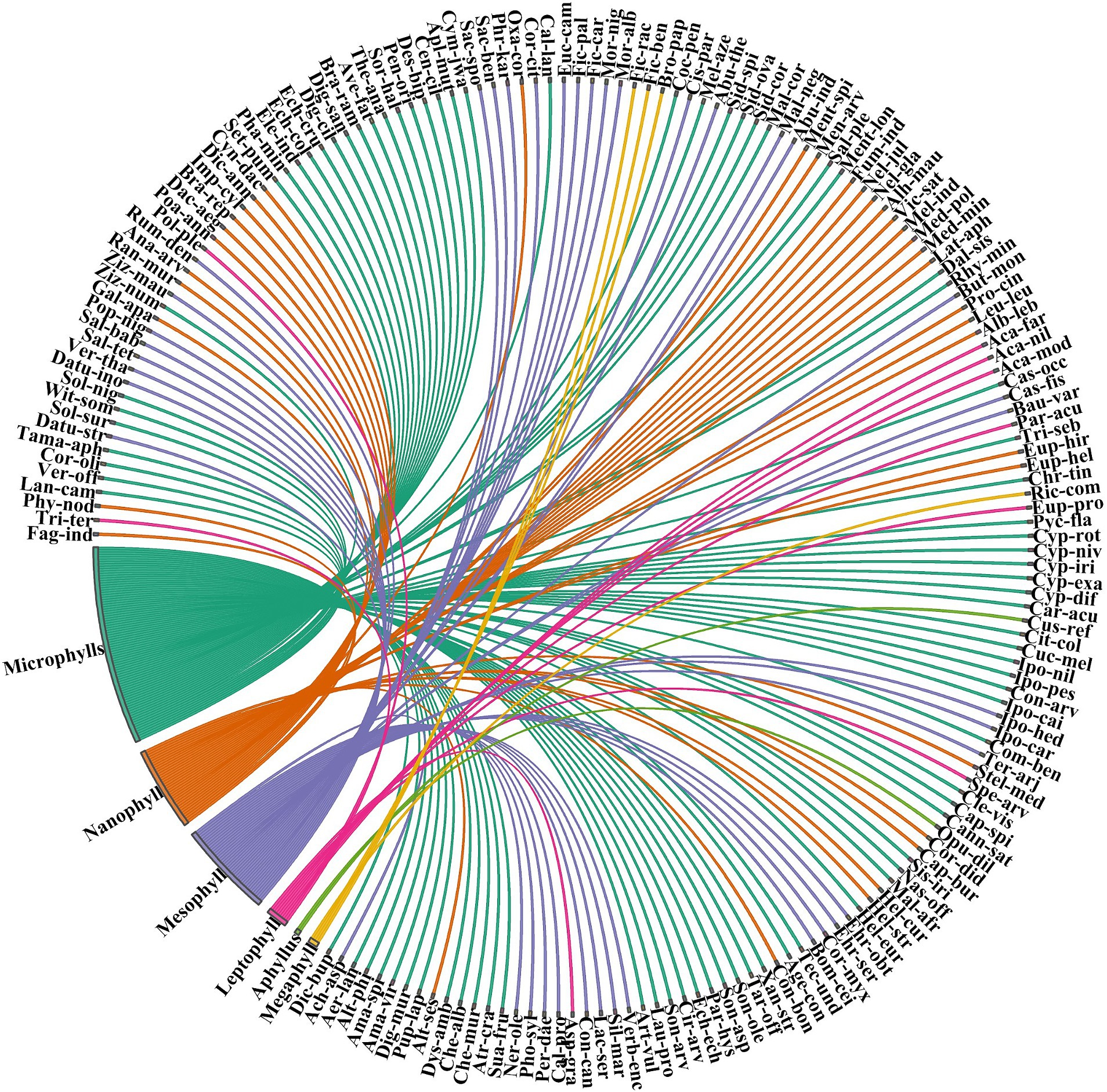
Figure 3. Distribution of species in Chichawatni Forest is classified using the leaf spectra categorization. The direction of the lines in a graphical depiction reflects the association of each species with certain leaf spectra categories, while the thickness of each bar represents the number of species within each individual life form category. Supplementary Table S1 contains a complete list of species names related to their leaf spectrum types. For full species names see Supplementary Table S1.
3.2 Pattern of alien species
Out of the 173 species, 48 species were alien species in all forest types, the ECP (41 species), followed by the PNP (36 species), MAP (25 species), DSP (23 species), and MTF (17 species) (Figure 4). Lantana camara, Parthenium hysterophorus, Neltuma glandulosa, Malvastrum coromandelianum, Abutilon theophrasti, Imperata cylindrica, and Datura inoxia were discovered in five forest types examined. Neltuma juliflora was spread widely over the ECP and MAP. In MTP and DSP, Ipomoea hederacea and Ageratum conyzoides were widely distributed.

Figure 4. Status of native and alien species in different forest types of the study area.
3.3 Pattern of flowering phenology
Plants were stratified into four clusters, while months were categorized into three groups according to their flowering phenology (Figure 5). The phenological spectrum of flora primarily encompasses the flowering duration of each species. Notably, the current investigation delineated two principal flowering periods within the flora. The primary flowering period extended from May to August in which about (N = 93, 54%) of plant species like (Poa annua, Euphorbia helioscopia, Achyranthes aspera, Digera muricata, Chenopodium album, Lactuca serriola, Cirsium arvense, Heliotropium strigosum, Cleome viscosa, Ipomoea carnea, Xanthium strumarium, and Citrullus colocynthis) were noted during the flowering stage. The subsequent flowering phase was lasted from March–April, in which a total of (N- 58, 34%) plant species (e.g., Cocculus pendulus, Conyza canadensis, Oxalis corniculata, Verbascum thapsus, Mentha longifolia, Galium aparine, Withania somnifera, Lantana camara, and Polygonum plebejum) appeared in full bloom. A significant portion of the variability in flower timing was linked to the growth habit, deciduous nature, and size of the plant species. Additionally, certain species initiated flowering even during the winter season. Notably, species such as Sida spinosa, Malva neglecta, and Sisymbrium irio exhibited flowering activity in February.
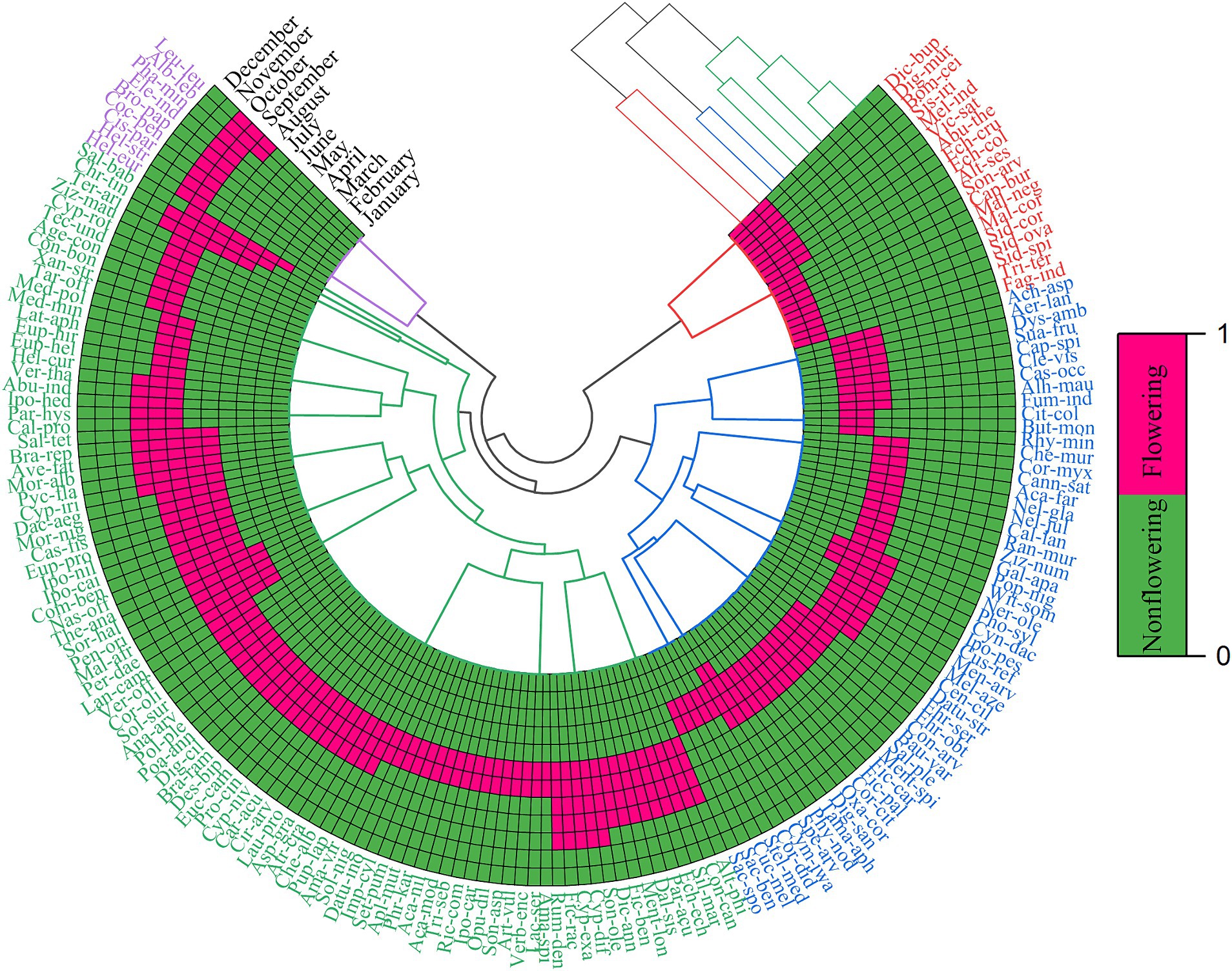
Figure 5. Polar cluster heat map showing the species and moth blue color indicates non-flowering and yellow for flowing stage. For full species names see Supplementary Table S1.
3.4 Diversity pattern
During examination of various plantation types, the species richness typically in a range of 50 to 72, with 61 in the Dalbergia sissoo plantation (DSP) and 59 in the Populus nigra plantation (PNP) was observed. The Shannon diversity indices, provided insights into the diversity of species within these plantations, displayed significant variation, ranging from a maximum of 2.4 at the Dalbergia sissoo plantation (DSP) to a minimum of 0.61 at the Morus alba plantation (MAP). These diversity indices exhibited noteworthy differences between different plantation types, as depicted in Figure 6. The highest levels of Shannon and Simpson index values were recorded in the Dalbergia sissoo plantation (DSP) and Populus nigra plantation (PNP), suggesting a greater diversity of species in these areas. Conversely, the Eucalyptus camaldulensis plantation (ECP) and Morus alba plantation (MAP) exhibited significantly lower Shannon index values. Furthermore, when it came to Pielou’s evenness, the Dalbergia sissoo plantation (DSP) stood out with the highest value, while the mix trees plantation (MTP) had the lowest. In terms of dominance index, it was noteworthy that the Morus alba plantation (MAP) and mix trees plantation (MTP) displayed the highest levels of dominance, indicating a prevalence of certain species within these plantations. Conversely, the Populus nigra plantation (PNP) exhibited the lowest dominance index, signifying a more balanced distribution of species. The findings underscored the varying ecological dynamics and biodiversity levels among different plantation types.
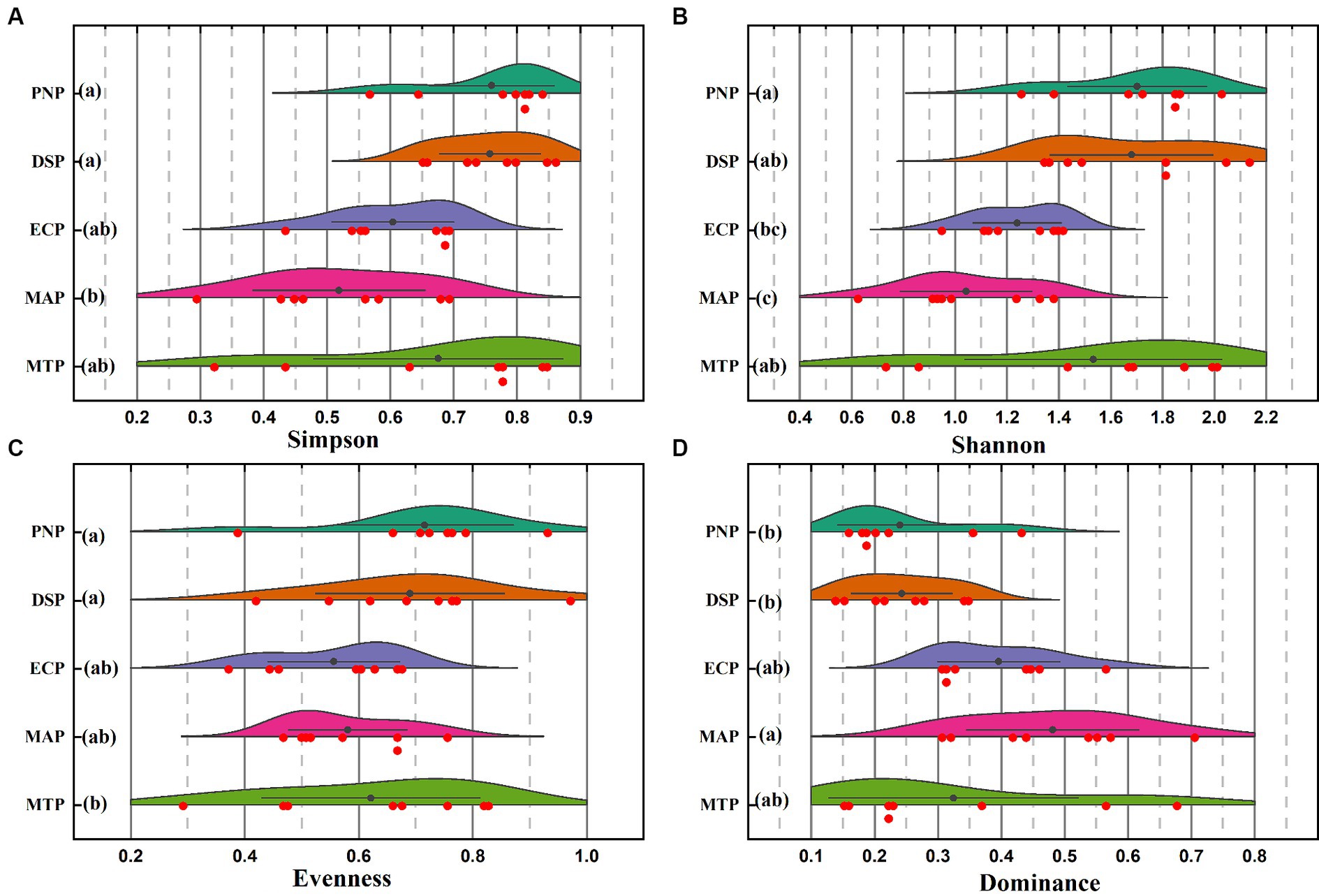
Figure 6. Diversity profiles of the plantation type within the Chichawatni Forest in Pakistan are illustrated in the ridgeline diagram. In this diagram, the sampling sites are represented, and lower-case letters (a, b) are used to denote significant differences between various plantation types, as determined by the Tukey test. (A) Simpson index, (B) Shannon index, (C) Evenness, and (D) Dominance Index.
3.5 Similarities between plantation types
The nMDS ordination provided compelling evidence of distinct dissimilarity patterns among different plantation types. The analysis involved the examination of data collected from 173 plant species sampled across various plantation types (Figure 7). The nMDS plot revealed that groups positioned closely or along the same axis exhibited a positive correlation, while those situated far apart or on different axes displayed a negative correlation. This spatial representation of the data visually conveyed the variations in plant composition based on altitude.
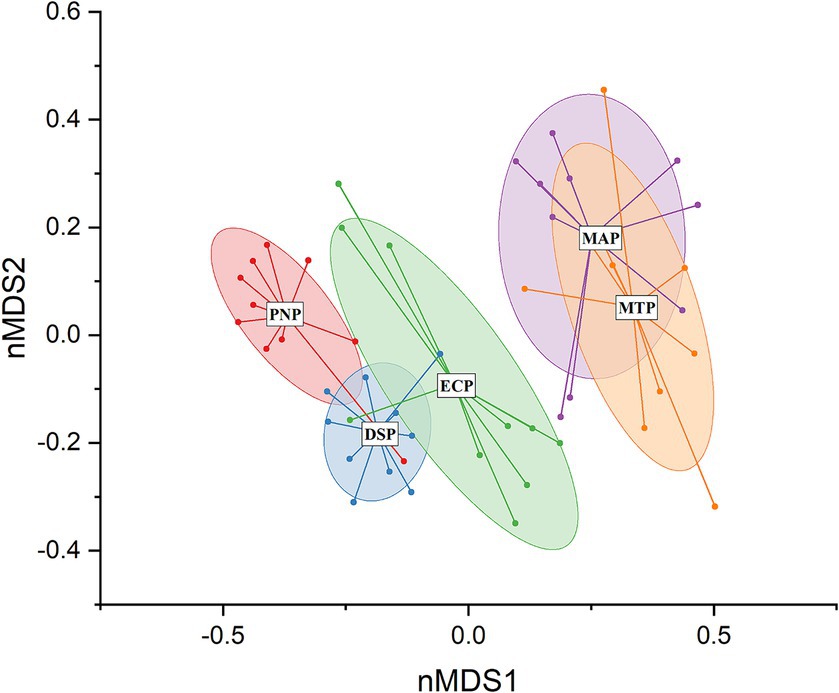
Figure 7. Representation of sampling sites at the alpine region, concerning the plantation types [Populus nigra plantation (PNP), Dalbergia sissoo plantation (DSP), Morus alba plantation (MAP), Eucalyptus camaldulensis plantation (ECP), and mix trees plantation (MTP)], was achieved using non-metric multidimensional scaling (NMDS). The 95% confidence intervals are displayed through ellipses.
Furthermore, the PERMANOVA analysis yielded significant results, indicating marked differences in plant composition between sites within the Populus nigra plantation (PNP) and Eucalyptus camaldulensis plantation (ECP) in comparison to the Morus alba plantation (MAP) and mix trees plantation (MTP). Bray–Curtis dissimilarities were the main factor influencing the observed differences. Additionally, the study highlighted substantial differences in species composition across the different kinds of plantations, as shown by significant findings for the Jaccard (F = 17.59, p < 0.01) and Bray–Curtis (F = 27.74; p < 0.01) indexes. These outcomes emphasized the influence of plantation type on species richness and relative abundances, contributing significantly to variations in species composition among the plantation types under investigation. The study unveiled a substantial divergence in plant species composition among the PNP, ECP, DSP, MAP, and MTP, with all five plantation types displaying noteworthy distinctions in their species makeup. Out of the 173 recorded species, 86 were recognized as noteworthy contributors to the differences in plant composition seen in the various forest plantations. Remarkably, the key species collectively accounted for a substantial 75% of the overall contribution, highlighting their pivotal contribution to the formation of plant communities at varying plantation types. The close association of the Populus nigra plantation (PNP) and Dalbergia sissoo plantation (DSP) groups in the nMDS plot suggests a high degree of species composition similarity between these two types of plantations. This spatial clustering suggests that the ecological characteristics and environmental conditions of DSP and PNP foster similar species assemblages.
3.6 Indicator species
Through a detailed SIMPER analysis, the investigation unveiled that a specific set of 86 plant species played a pivotal role in accounting for a significant 75% of the dissimilarity observed among the five plantation types under study, namely the Populus nigra plantation (PNP), Dalbergia sissoo plantation (DSP), Morus alba plantation (MAP), Eucalyptus camaldulensis plantation (ECP), and mix trees plantation (MTP). To further elucidate the distinctiveness of each plantation type, an indicator species approach was applied, which led to the identification of 24 indicator species distributed across these plantations.
The results from the indicator species analysis painted a clear picture of the unique compositions within each plantation types. In the Populus nigra plantation (PNP), the significant indicator species included Dicliptera bupleuroides, Xanthium strumarium, Bombax ceiba, Capsella bursa-pastoris, Cuscuta reflexa, and Ziziphus nummularia, each displaying a noteworthy indicator value (p ≤ 0.05). Meanwhile, in Dalbergia sissoo plantation (DSP), the indicator species were comprised of Acacia farnesiana, Chenopodium album, Broussonetia papyrifera, and Xanthium strumarium, all marked by significant p-values. In the Eucalyptus camaldulensis plantation (ECP), the indicator plants emerged were Acacia nilotica, Ehretia serrata, Cleome viscosa, Euphorbia helioscopia, Melilotus indica, and Neltuma glandulosa, once again featuring significant indicator values. In Morus alba plantation (MAP), the significant indicator species were Cannabis sativa, Cirsium arvense, Lantana camara, and Saccharum bengalense while in mix trees plantation (MTP), the species possessing significant indicator values were Neltuma juliflora, Euphorbia hirta, Parthenium hysterophorus, and Ehretia serrata (Figure 8). It’s noteworthy that Xanthium strumarium emerged as a common indicator species in Populus nigra plantation (PNP) and Dalbergia sissoo plantation (DSP), while Lantana camara is was common indicator species in Morus alba plantation (MAP) and Eucalyptus camaldulensis plantation (ECP).
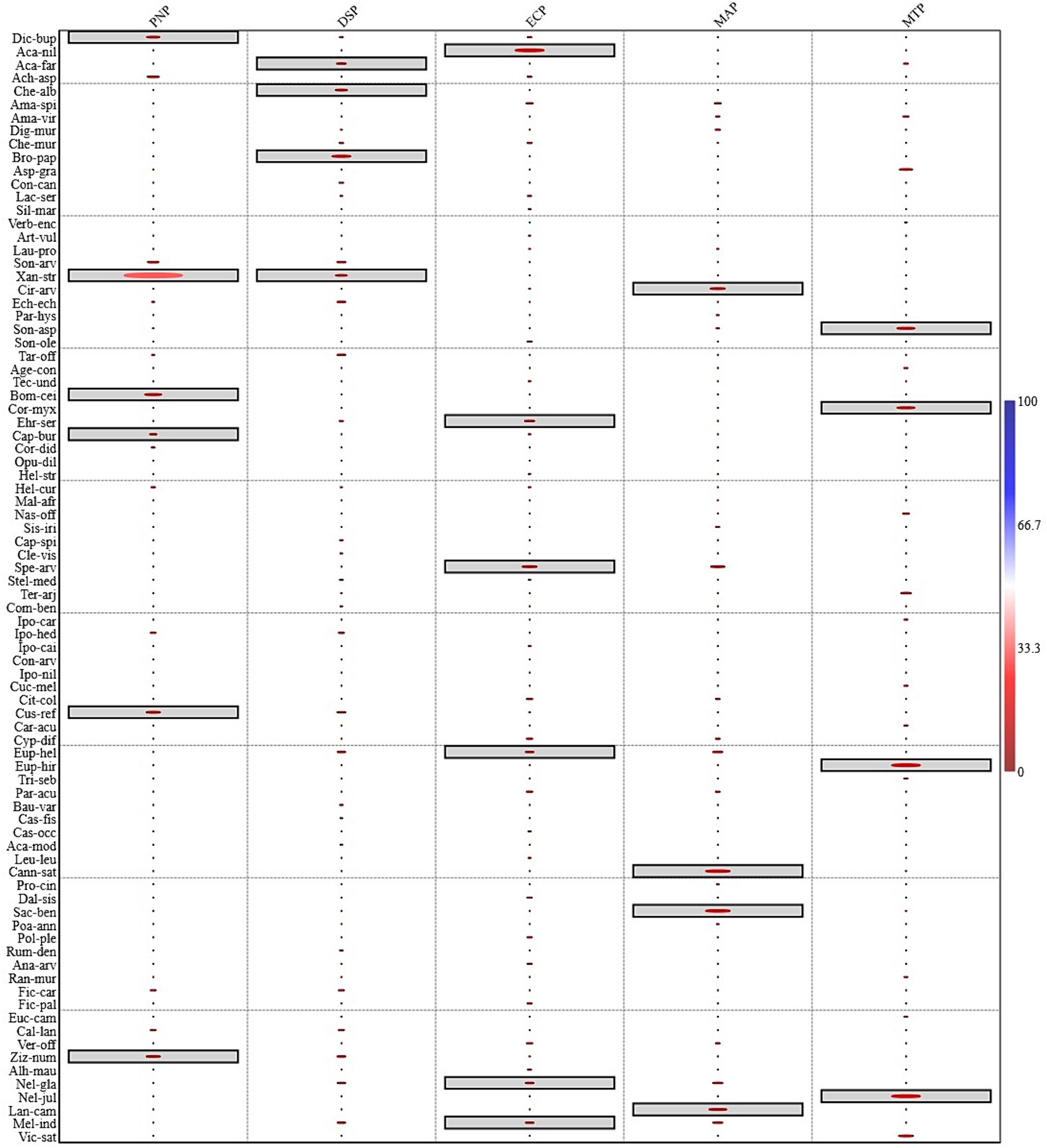
Figure 8. Assessment of indicator species among plants from different plantation types in the Chichawatni Forest to enlighten the noteworthy species (p ≤ 0.05) specific to each plantation types. These significant species were identified and specially marked within their respective boxes. For full species names see Supplementary Table S1.
3.7 Impact of anthropogenic disturbance on vegetation
Collectively, the first and second axes in the Canonical Correspondence Analysis (CCA) jointly elucidated 18.42% of the total variation in the data, and impressively, they accounted for a substantial 91.35% of the fitted variation. More specifically, CCA axis 1 unveiled 7.17% of the total variation, corresponding to 39.46% of the fitted variance, while axis 2 contributed 11.25% of the total variation, capturing 51.89% of the fitted variance (as detailed in Table 1). The impact of various parameters on the composition of plant species was found to be highly significant (p < 0.01). The CCA results underscored that certain factors, notably road accessibility, stem cutting, and fire, played pivotal roles in explaining the variance observed in plant distribution patterns (p < 0.05), while factors such as biological invasion and grazing contributed less to this observed variation. Notably, the percentage contribution was statistically significant for only three parameters in elucidating the variance in vegetation patterns (p < 0.05). Road accessibility emerged as the most influential, accounting for 34.9% of the observed variation, followed by stem cutting at 20.5%, and fire at 18% among these key factors (as presented in Table 2). Road accessibility, stem cutting, and fire emerged as the dominant drivers behind the observed variation in vegetation patterns, shedding light on their crucial roles in shaping the plant composition and distribution within the studied plantations (Figure 9).
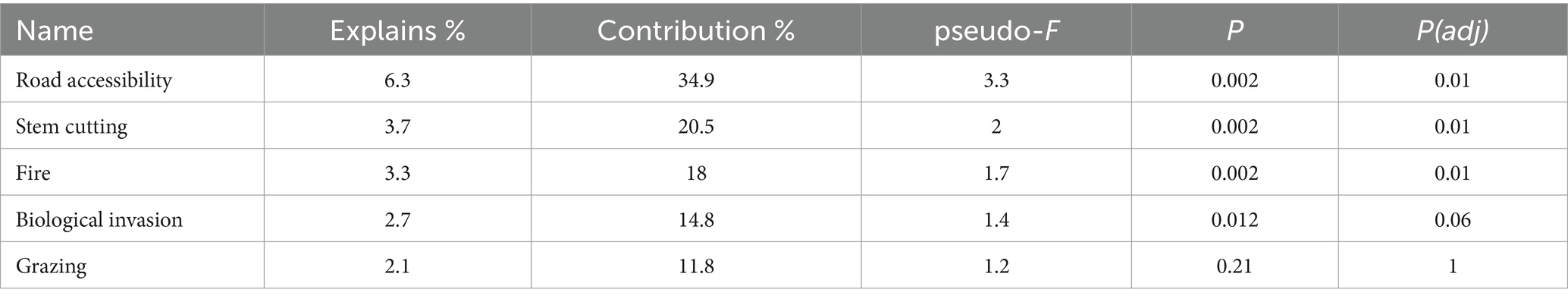
Table 2. Effects of variables on plantation types.
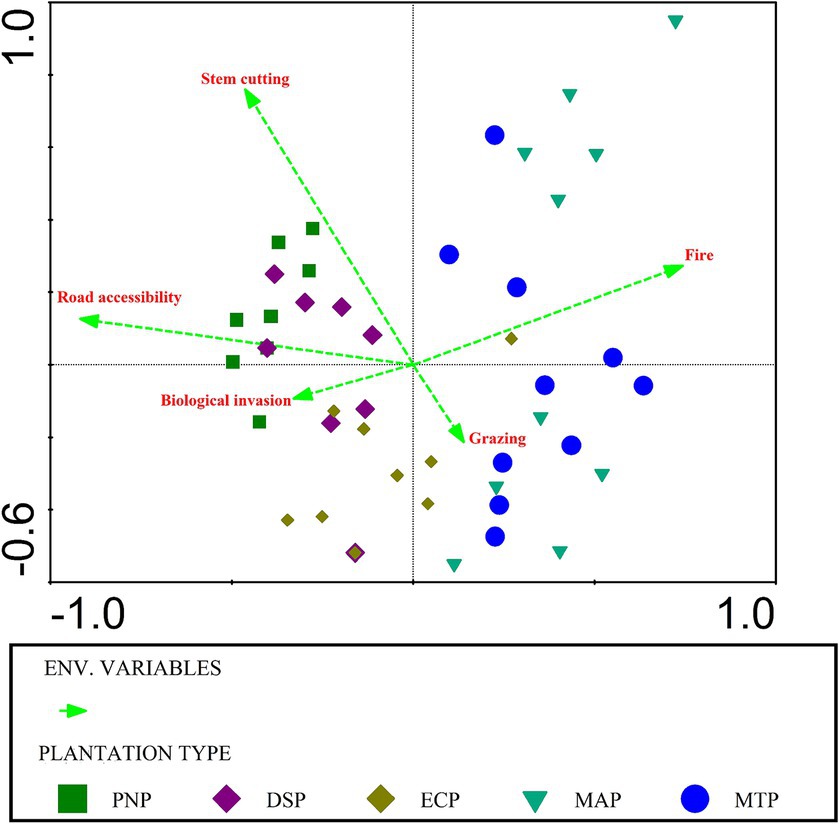
Figure 9. Species ordination using CCA (canonical correspondence analysis) and five disturbance variables. The direction and length of the arrow reflect the link between the factor and the ordination axes, while the various shapes and color symbolizes the plantation types.
4 Discussion
Vegetation structural complexity as an essential ecological characteristic referred to the three-dimensional distribution of plants within an ecosystem (LaRue et al., 2023). One certainty in the modern anthropogenic epoch biological communities exhibited dynamic instability and underwent fluctuations over both short and long temporal scales (Smith et al., 2009). Predicting the effects of ongoing habitat and biodiversity declined on ecosystem functions and services required an understanding of the role diversity plays in ecosystem stability (Dolezal et al., 2020). One of the primary objectives of the study was to assess the biodiversity and ecological traits of plant species within the Chichawatni forest plantation. The results revealed a remarkable diversity of angiosperm species, with 173 species belonging to 42 different families. This high diversity is a testament to the unique ecological conditions prevailing in the subtropical region of the India–Pakistan subcontinent. The dominance of the Poaceae family, with 25 species, is noteworthy. This family’s prevalence is indicative of the ecological importance of grasses in subtropical ecosystems, which often play vital roles in soil stabilization, nutrient cycling, and as habitat for various wildlife (Altaf et al., 2022; Waheed et al., 2022a,b, 2023a; de Oliveira et al., 2023; Joshi et al., 2023). Other prominent families, such as Fabaceae, Asteraceae, and Amaranthaceae, also contributed significantly to the overall species count. Comparable findings were documented by Arshad et al. (2023) and Waheed et al. (2023a,b) from the subtropical region of Pakistan. Fabaceae was recorded as the dominant family by Haq et al. (2023a,b) from Shawilks Mountain Range, Western Himalayas, while Mehraj et al. (2018) recorded Asteraceae as the dominant family from Himalayas. The research insights of Haq et al. (2019, 2021) demonstrated the remarkable adaptive capacity of the Asteraceae family across various regions and climates. The abundance of graminoids, forbs, and deciduous tree species, including Dalbergia sissoo and Morus alba, indicated the adaptability of these plants to the region’s distinct climate, characterized by seasonal changes in temperature and precipitation. This diversity mirrored the results of studies in other subtropical forests such as the Western Ghats in India (Subashree et al., 2021) and the Kabal valley, Swat, Pakistan (Ilyas et al., 2018). However, while the number of species is impressive, the uneven distribution across families is a common trend in forest ecosystems globally (Singh et al., 2020). The prevalence of the Poaceae family is consistent with its ubiquitous presence in many forest ecosystems (Behera and Misra, 2006; Joshi et al., 2021).
The presence of therophytes as the dominant life form aligns with the region’s semi-arid climate, where these annual plants have evolved strategies to complete their life cycles within the limited wet period (Moro et al., 2015; Rashid et al., 2021; Manan et al., 2022). Hemicryptophytes, with their capacity to persist through unfavorable conditions, also exhibited a substantial role. These life forms reflected the resilience of plant species in subtropical climates, where they must cope with both dry periods and monsoon rains (Pfadenhauer and Klötzli, 2020). The prevalence of microphyll and mesophyll leaf spectrum types revealed the need for efficient water use in subtropical climates, where water availability could be a limiting factor (Amjad et al., 2017; Rahman et al., 2021). Understanding the prevalence of these ecological traits is crucial for predicting how plant communities may respond to future climate fluctuations in the region (Alexander et al., 2018). These traits are crucial for the survival of plant species during extended dry spells and highlighted the adaptability of the Chichawatni forest flora. The identification of two major flowering periods in the Chichawatni Forest is a noteworthy finding, as it showcases the adaptability of plant species to seasonal changes. Such phenological variations are consistent with the findings of studies in other subtropical and temperate forests, where species exhibited distinct flowering periods in response to local climatic conditions (Pei et al., 2015; Du et al., 2019; Song et al., 2021). However, the study also reported flowering during the winter months, a phenomenon observed less frequently in subtropical forests but documented in temperate ecosystems (Negi et al., 2022). This suggests that the Chichawatni Forest may be subjected to unique climatic influences, further highlighting the subtropical region’s ecological complexity.
The study indicated the variations in plant richness and diversity metrics across various plantation types within the forest. Such variations in diversity are common in managed forests globally (Goded et al., 2019; Poudyal et al., 2019). The higher diversity observed in Dalbergia sissoo and Populus nigra plantations were aligned with findings in various plantation studies, where the species-rich plantations tended to support a more diverse range of understory species (Ramovs and Roberts, 2003; Bremer and Farley, 2010). Conversely, Eucalyptus camaldulensis and Morus alba plantations exhibited lower diversity, a pattern often associated with Eucalyptus camaldulensis plantations (Alem and Pavlis, 2012; Bekele and Abebe, 2018). The findings underscored the importance of considering plantation type in forest management decisions to promote biodiversity (Liu et al., 2019; Blondeel et al., 2021). Such finding implied that particular tree species exerted a more favorable impact on the diversity of the understory vegetation. The outcomes aligned with prior research had shown the importance of tree species selection in plantation design (Barlow et al., 2007). The significance of plantation type was extended beyond species richness and diversity. The study also unraveled varying levels of evenness and dominance within different plantation types. The Dalbergia sissoo plantation displayed the highest evenness, indicating a more balanced distribution of species, while the mixed trees plantation had the lowest evenness, suggesting that certain species dominated the plantation. Understanding such patterns of evenness and dominance could have practical implications for forest managers. For instance, enhancing evenness within plantations could contribute more stable and resilient ecosystems, less susceptible to the dominance of a few species (Sanderson et al., 2004). The findings had direct implications for forest management strategies in subtropical regions like India and Pakistan. Sustainable forestry practices should consider not only the economic value of timber but also the ecological consequences of tree species selection. The positive relationship between certain plantation types and higher biodiversity suggested that careful species selection could promote both economic goals and the preservation of native biodiversity.
The nMDS ordination results offer compelling evidence of distinct dissimilarity patterns among different plantation types within the Chichawatni forest. The analysis involved data collected from 173 plant species sampled across various plantation types, namely the Populus nigra plantation (PNP), Dalbergia sissoo plantation (DSP), Morus alba plantation (MAP), Eucalyptus camaldulensis plantation (ECP), and mix trees plantation (MTP). The nMDS plot visually represented these dissimilarity patterns by placing groups of plant communities closer or farther apart based on their similarity in species composition (Clarke et al., 2006). Conversely, groups situated far apart or on different axes display a negative correlation, indicating greater dissimilarity in species composition. The dissimilarity patterns revealed by the nMDS analysis were not just abstract representations but carried significant ecological implications (Legendre and Anderson, 1999). The patterns highlighted the varying ecological dynamics and species compositions among different plantation types, which were influenced by factors such as tree species selection, silvicultural practices, and disturbance regimes (Paillet et al., 2010). For example, the proximity of the Dalbergia sissoo plantation (DSP) and Populus nigra plantation (PNP) groups in the nMDS plot suggested a higher degree of similarity in species composition between these two plantation types. Such observation aligned with the previously discussed findings of higher species richness and diversity indices in such plantations (Wang et al., 2019). It indicated that certain species within these plantations might share ecological niches or exhibited similar responses to environmental conditions, leading to a more homogeneous plant community (Hamann and Wang, 2006). Conversely, the Morus alba plantation (MAP) and mix trees plantation (MTP) groups appeared farther apart in the nMDS plot, indicating greater dissimilarity in species composition. Such divergence revealed the distinct ecological dynamics within the plantation types. Factors such as tree growth patterns, competitive interactions, and responses to disturbances might contribute to the dissimilarities (Lloret et al., 2011). Understanding differences is crucial for tailoring management strategies that promote both economic objectives and ecological health (Moldan et al., 2012). The PERMANOVA analysis showed significant plant composition differences between plantation types. Notable dissimilarities were identified between the Populus nigra plantation (PNP) and the Eucalyptus camaldulensis plantation (ECP) compared to the Morus alba plantation (MAP) and the mixed tree plantation (MTP). This emphasizes the crucial significance of individual species in generating different plant communities across various plantation types (Waheed et al., 2020, 2022b; Haq et al., 2023a).
The indicator species identified played critical ecological roles within their respective plantation types in the subtropical forest. The reported species possessed specific ecological traits that made them well-suited to their environments, and understanding their traits enhanced our comprehension of the broader ecosystem dynamics (Dufrêne and Legendre, 1997). For instance, in the Populus nigra plantation, species like Dicliptera bupleuroides and Xanthium strumarium, characterized by high indicator values, are often associated with open habitats and pioneer vegetation. Those might serve as early colonizers, contributing to the initial stages of ecological succession (McLane et al., 2012). Bombax ceiba, a prominent indicator species, was a deciduous tree known for its ability to tolerate seasonal water fluctuations, potentially influencing the hydrological regime of the plantation (Maiti et al., 2021). Capsella bursa-pastoris, a common indicator, as an annual herb thrived in disturbed areas, reflecting the ecological conditions within the Populus nigra plantation (Petersen et al., 2021). In the Dalbergia sissoo plantation (DSP), indicator species like Acacia farnesiana and Chenopodium album were often associated with arid and semi-arid regions (Waheed et al., 2023a,b), reflecting the specific moisture and soil conditions of the DSP. Broussonetia papyrifera, another DSP indicator, was valued for its role in stabilizing soils and providing shade, highlighting its importance in shaping the microenvironment (Agyeman et al., 2016). Xanthium strumarium, shared with the PNP, was an adaptive species with broad ecological tolerance, indicative of its capacity to thrive in different environments (Ullah et al., 2022). In the Eucalyptus camaldulensis plantation (ECP), species like Acacia nilotica and Lantana camara, identified as indicators, often found in disturbed areas and were known for their allelopathic effects, which could influence plant interactions (Zhou et al., 2019).
The species might be significantly involved in shaping the competitive dynamics and understory composition. Neltuma juliflora, another indicator, was adapted to arid environments, reflecting the specific ecological conditions of the ECP. Understanding these ecological traits of indicator species provides insights into the interactions and processes shaping the Eucalyptus camaldulensis plantation (ECP) (Dondofema et al., 2023). In the Morus alba plantation (MAP) and mix trees plantation (MTP), indicator species viz.; Cannabis sativa and Cirsium arvense were often associated with ruderal habitats and might thrive in areas subjected to disturbances like fire and grazing (Vakhlamova et al., 2022). Lantana camara, shared with the ECP, was known for its invasive potential, influencing plant community composition. Understanding these ecological roles is essential for managing the dynamics of these plantation types and addressing issues related to disturbance and invasiveness (Meyer et al., 2021; Haq et al., 2022; Rai, 2022).
The Canonical Correspondence Analysis (CCA) elucidates the significant influence of factors like road accessibility, stem cutting, and fire on plant distribution patterns within various plantation types, aligned with a broad body of literature documenting the impacts of disturbance on vegetation and species composition in managed and natural forest ecosystems (Haq et al., 2019; Kutnar et al., 2019; Alam et al., 2023). Understanding such impacts was vital for informed forest management and conservation strategies (Haq et al., 2021). The impact of anthropogenic disturbances, such as road accessibility, stem cutting, and fire, on plant distribution patterns reaffirmed their significance in shaping forest ecosystems. Those findings held significant implications for forest management and conservation strategies in subtropical regions, emphasizing the necessity for sustainable practices aimed at alleviating the adverse effects of disturbances while promoting biodiversity. Future research in the Chichawatni Forest and similar subtropical ecosystems should focus on long-term monitoring to assess the resilience of plant communities for ongoing environmental changes and disturbances. Road accessibility emerged as a significant driver of vegetation change, with road construction fragmenting habitats and potentially facilitating the spread of invasive plants (Ramprasad et al., 2020). Stem cutting, often associated with resource extraction, had discernible impacts on specific sites within the plantation, highlighting the need for sustainable harvesting practices (Vogel et al., 2021). Fire, another disturbance factor, can disrupt ecosystem processes and negatively affect plant communities (Miller and Safford, 2020). These findings underscore the importance of managing those disturbances carefully to maintain the ecological integrity of managed forest ecosystems.
The Chichawatni forest plantation serves as a microcosm of subtropical managed forests in the broader India-Pakistan region. These forests are invaluable not only for timber production but also for their role in regulating water resources, conserving biodiversity, and providing ecosystem services to local communities (Ghafoor et al., 2022). As such, the results of the present investigation inferred regional significance. Such findings regionally provided valuable insights into how plantation design, ecological traits, and disturbances intersect in subtropical ecosystems. The study’s results had implications for conservation and forest management in the subtropical region. Forest managers could use the insights gained to inform tree species selection and plantation design, aiming for a balance between economic productivity and ecological health. Conservation efforts could be significant from a better understanding of how subtropical ecosystems respond to human activities and climate change, facilitating the preservation of these vital ecosystems (Haq et al., 2024a,b). The research provided a comprehensive understanding of the intricate relationships between biodiversity, ecological traits, plantation types, and disturbances in a subtropical managed forest in the region. The findings contributed to our knowledge of subtropical ecosystems and offered practical guidance for sustainable forest management and conservation in the ecologically diverse and economically significant area.
4.1 Implications for forest management
The Forest ecosystem management is hailed as the answer to conservation issues. Ecologists are inferring that natural reserves alone would not be sufficient to effectively preserve biological diversity. Because of the current reserve network’s limitations, the likelihood of a significant extension, and movement constraints, species found in reserves are particularly sensitive to the effects of climate change. Thus, present studies devised a mechanism of supplementing the natural forest reserves with a managed forest matrix that utilized ecological principles to manage for both commodity production and species variety conservation. Forty-eight species as alien species from various forest types, and the maximum (41 species) in the Eucalyptus camaldulensis plantation, and minimum of (17 alien species) in the Mix trees plantation were reported. Moreover, alien species posed a significant challenge to vegetation management due to their invasive nature. Hence, considering the ecological traits of species within each forest type was imperative. The current investigation brought attention to the phenomenon observed in Populus and Eucalyptus forest stands characterized by a prevalence of alien species. The findings highlighted that the introduction of exotic plantations significantly facilitated the expansion of those alien species when contrasted with their native counterparts. Thus, it was vehemently opposed the introduction of non-native species into the managed forests, and further suggested that the natural forest should be used as reference system to offer guidance on the plantation in the management forests that would be dedicated to both wood production and conservation of biodiversity.
Present findings indicated that native Dalbergia sissoo plantations and mixed tree plantations harbor greater richness in vascular plant species, particularly featuring a higher proportion of native taxa compared to exotic tree plantations. That highlighted the distinct effects of exotic tree plantations on understory plant communities and biodiversity. The study underscored the importance of selecting native tree species for plantation in biodiversity preservation efforts, which had often been overlooked in large-scale forest restoration projects worldwide. Given the increasing prevalence of exotic plantations in Pakistan and other regions, urgent modifications to reforestation management practices were needed to enhance stand structure, understory vegetation, and biodiversity conservation. The present analysis recommended prioritizing native or mixed tree plantation strategies for clear-cut areas in Pakistan subtropical region to better preserve indigenous plant diversity in managed forests. Finally, it was suggested that the lessons learnt from the current study would be useful for design and effectively manage such multipurpose landscape.
5 Conclusion
The present study provides valuable insights into the intricate relationships between vegetation composition, ecological traits, plantation types, and disturbances within the Chichawatni forest plantation, a subtropical managed forest type. The Chichawatni forest is enriched with a diverse floral composition comprising of 173 angiosperm species from 42 families. Such diversity reflected the region’s subtropical climate, fertile terrain, and varied ecological niches. The prevalence of therophytes, deciduous trees, and specific leaf spectrum types underscored the adaptability of plant species to the region’s seasonal variations. Different plantation types within the forest exhibited varying impacts on biodiversity and vegetation composition. Notably, Dalbergia sissoo and Populus nigra plantations displayed higher species richness and diversity indices, highlighting the role of plantation type in shaping ecological dynamics. Those findings had important implications for sustainable forest management practices tailored to specific plantation types. Disturbances, such as road accessibility, stem cutting, and fire, significantly influenced plant distribution patterns within the forest. Road construction could fragment habitats, stem cutting impacted certain sites, and fire could disrupt ecosystem processes. Understanding the impacts of those disturbances was critical for effective forest conservation and management. The study’s findings aligned with broader ecological patterns observed from the subtropical region. Subtropical ecosystems from the area were characterized by unique climatic conditions and rich biodiversity. The study contributed to the understanding of how subtropical forests responded to various ecological factors, providing insights that could monitor and create conservation and management strategies for the ecologically diverse region. Forest managers, policymakers, and conservationists in the subtropical region could be benefited from the study’s insights. Tailoring forest management approaches to different plantation types, considering ecological traits and responses to disturbances, also helped to strike a balance between economic interests and ecological health in managed forests.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MW: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. SH: Data curation, Project administration, Validation, Writing – review & editing. FA: Project administration, Supervision, Validation, Writing – review & editing. RB: Project administration, Writing – review & editing. AH: Funding acquisition, Writing – review & editing. EFA: Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2024R356), King Saud University, Riyadh, Saudi Arabia. This manuscript is part of the Ph.D. thesis of MW. All authors are deeply indebted to the University of Okara for providing facilities for the field expeditions to accomplish the research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1406075/full#supplementary-material
Footnotes
References
Abbas, Z., Khan, S. M., Alam, J., Khan, S. W., and Abbasi, A. M. (2017). Medicinal plants used by inhabitants of the Shigar Valley, Baltistan region of Karakorum range-Pakistan. J. Ethnobiol. Ethnomed. 13, 1–15. doi: 10.1186/s13002-017-0172-9
Agyeman, V. K., Addo-Danso, S. D., Kyereh, B., and Abebrese, I. K. (2016). Vegetation assessment of native tree species in Broussonetia papyrifera-dominated degraded forest landscape in southern Ghana. Appl. Veg. Sci. 19, 498–507. doi: 10.1111/avsc.12241
Alam, N., Bohnett, E., Zafar, M., Sher, H., Ahmad, B., Ramadan, M. F., et al. (2023). Impact of anthropogenic threats on species diversity: a case study of the sub-Himalayan tropical dry deciduous forests of Pakistan. Sustain. For. 15:2829. doi: 10.3390/su15032829
Alem, S., and Pavlis, J. (2012). Native woody plants diversity and density under Eucalyptus camaldulensis plantation, in Gibie Valley, South Western Ethiopia. Open J. For. 2:232. doi: 10.4236/ojf.2012.2402
Alexander, J. M., Chalmandrier, L., Lenoir, J., Burgess, T. I., Essl, F., Haider, S., et al. (2018). Lags in the response of mountain plant communities to climate change. Glob. Change Biol. 24, 563–579. doi: 10.1111/gcb.13976
Ali, S. I., and Qaiser, M. (1986). A phytogeographical analysis of the phanerogams of Pakistan and Kashmir. Proc. Royal Soc. Edinburgh Sect. B Biol. Sci. 89, 89–101. doi: 10.1017/S0269727000008939
Altaf, A., Haq, S. M., Shabnum, N., and Jan, H. A. (2022). Comparative assessment of phyto diversity in Tangmarg Forest division in Kashmir Himalaya. India. Acta Ecologica Sinica 42, 609–615. doi: 10.1016/j.chnaes.2021.04.009
Ameray, A., Bergeron, Y., Valeria, O., Montoro Girona, M., and Cavard, X. (2021). Forest carbon management: a review of silvicultural practices and management strategies across boreal, temperate and tropical forests. Curr. For. Rep. 7, 245–266. doi: 10.1007/s40725-021-00151-w
Amjad, M. S., Arshad, M., Page, S., Qureshi, R., and Mirza, S. N. (2017). Floristic composition, biological spectrum and phenological pattern of vegetation in the subtropical forest of Kotli District, AJK, Pakistan. PAB 6, 426–447. doi: 10.19045/bspab.2017.60043
Arshad, F., Haq, S. M., Waheed, M., Jameel, M. A., and Bussmann, R. W. (2023). Environmental variables drive medicinal plant composition and distribution in various forest types of subtropical region in Pakistan. Acta Eco. Sin 44, 234–246. doi: 10.1016/j.chnaes.2023.05.004
Balandier, P., Mårell, A., Prévosto, B., and Vincenot, L. (2022). Tamm review: Forest understorey and overstorey interactions: so much more than just light interception by trees. For. Eco. Mansag. 526:120584. doi: 10.1016/j.foreco.2022.120584
Barlow, J., Gardner, T. A., Araújo, I. S., Ávila-Pires, T. C., Bonaldo, A. B., Costa, J. E., et al. (2007). Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. PNAS 104, 18555–18560. doi: 10.1073/pnas.0703333104
Behera, S. K., and Misra, M. K. (2006). Floristic and structure of the herbaceous vegetation of four recovering forest stands in the eastern Ghats of India. Biodivers. Conserv. 15, 2263–2285. doi: 10.1007/s10531-004-8215-7
Bekele, T., and Abebe, W. (2018). Indigenous woody species regeneration under the canopies of exotic tree plantations at Tore Forest, Gelana District, southern Oromia, Ethiopia. Biodivers. Int. J. 2, 1–6. doi: 10.15406/bij.2018.02.00034
Blondeel, H., Landuyt, D., Vangansbeke, P., De Frenne, P., Verheyen, K., and Perring, M. P. (2021). The need for an understory decision support system for temperate deciduous forest management. For. Ecol. Manag. 480:118634. doi: 10.1016/j.foreco.2020.118634
Böcher, T. W., and Bentzon, M. W. (1958). Density determination in plant communities. Oikos 9, 35–56. doi: 10.2307/3564896
Borma, L. S., Costa, M. H., da Rocha, H. R., Arieira, J., Nascimento, N. C. C., Jaramillo-Giraldo, C., et al. (2022). Beyond carbon: the contributions of south American tropical humid and subhumid forests to ecosystem services. Rev. Geophys. 60:e2021RG000766. doi: 10.1029/2021RG000766
Bremer, L. L., and Farley, K. A. (2010). Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 19, 3893–3915. doi: 10.1007/s10531-010-9936-4
Brown, K. A., Bunting, M. J., Carvalho, F., de Bello, F., Mander, L., Marcisz, K., et al. (2023). Trait-based approaches as ecological time machines: developing tools for reconstructing long-term variation in ecosystems. Funct. Ecol. 37, 2552–2569. doi: 10.1111/1365-2435.14415
Clarke, K. R., Somerfield, P. J., and Chapman, M. G. (2006). On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Bio. Ecol. 330, 55–80. doi: 10.1016/j.jembe.2005.12.017
Ćosović, M., Bugalho, M. N., Thom, D., and Borges, J. G. (2020). Stand structural characteristics are the most practical biodiversity indicators for forest management planning in Europe. Forests 11:343. doi: 10.3390/f11030343
de Oliveira, N. D., Diniz, E. S., and Avila, R. S. D. (2023). Composition and phylogenetic structure of Pampean grasslands under distinct land use and presence of alien species. Community Ecol. 24, 73–86. doi: 10.1007/s42974-023-00136-8
Devi, N. L., Brearley, F. Q., and Tripathi, S. K. (2023). Phenological diversity among sub-tropical moist forest trees of North-Eastern India. J. Trop. Ecol. 39:e29. doi: 10.1017/S0266467423000184
Dolezal, J., Fibich, P., Altman, J., Leps, J., Uemura, S., Takahashi, K., et al. (2020). Determinants of ecosystem stability in a diverse temperate forest. Oikos 129, 1692–1703. doi: 10.1111/oik.07379
Dondofema, F., Nethengwe, N., Taylor, P., and Ramoelo, A. (2023). Comparison of satellite platform for mapping the distribution of Mauritius thorn (Caesalpinia decapetala) and river red gum (Eucalyptus camaldulensis) in the Vhembe biosphere reserve. Rem. Sens. 15:2753. doi: 10.3390/rs15112753
Du, Y., Yang, B., Chen, S. C., and Ma, K. (2019). Diverging shifts in spring phenology in response to biodiversity loss in a subtropical forest. J. Veg. Sci. 30, 1175–1183. doi: 10.1111/jvs.12806
Dufrêne, M., and Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–366. doi: 10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Ellenberg, D., and Mueller-Dombois, D. (1974). Aims and methods of vegetation ecology. New York: Wiley, 547.
Forrester, J. A., Keyser, T. L., and Schnake, D. K. (2023). “Ecological Silviculture for southern Appalachian hardwood forests” in Ecological Silvicultural Systems: exemplary Models for Sustainable Forest Management. Eds. Franklin, J. F., Johnson, K. N., and Johnson, D. L., (Waveland Press). 98.
Ghafoor, G. Z., Sharif, F., Shahid, M. G., Shahzad, L., Rasheed, R., and Khan, A. U. H. (2022). Assessing the impact of land use land cover change on regulatory ecosystem services of subtropical scrub forest, Soan Valley Pakistan. Sci. Rep. 12, 1–12. doi: 10.1038/s41598-022-14333-4
Goded, S., Ekroos, J., Domínguez, J., Azcárate, J. G., Guitián, J. A., and Smith, H. G. (2019). Effects of eucalyptus plantations on avian and herb species richness and composition in north-West Spain. Glob. Ecol. Conser. 19:e00690. doi: 10.1016/j.gecco.2019.e00690
Hamann, A., and Wang, T. (2006). Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecol. 87, 2773–2786. doi: 10.1890/0012-9658(2006)87[2773:PEOCCO]2.0.CO;2
Haq, S. M., Amjad, M. S., Waheed, M., Bussmann, R. W., and Proćków, J. (2022). The floristic quality assessment index as ecological health indicator for forest vegetation: a case study from Zabarwan Mountain range, Himalayas. Ecol. Indic. 145:109670. doi: 10.1016/j.ecolind.2022.109670
Haq, S. M., Lone, F. A., Waheed, M., Siddiqui, M. H., Arshad, F., and Bussmann, R. W. (2024a). Analyzing anthropogenic determinants of vegetation distributions pattern in high-elevation forests of the Himalayas. Pak. J. Bot. 56, 1441–1450. doi: 10.30848/PJB2024-4(42)
Haq, S. M., Rashid, I., Khuroo, A. A., Malik, Z. A., and Malik, A. H. (2019). Anthropogenic disturbances alter community structure in the forests of Kashmir Himalaya. Trop. Ecol. 60, 6–15. doi: 10.1007/s42965-019-00001-8
Haq, S. M., Rashid, I., Waheed, M., and Khuroo, A. A. (2023a). From forest floor to tree top: partitioning of biomass and carbon stock in multiple strata of forest vegetation in Western Himalaya. Envir. Monitor. Asses. 195:812. doi: 10.1007/s10661-023-11376-6
Haq, S. M., Waheed, M., Bussmann, R. W., and Arshad, F. (2023b). Vegetation composition and ecological characteristics of the forest in the Shawilks Mountain range from Western Himalayas. Acta Ecol. Sin. 43, 798–809. doi: 10.1016/j.chnaes.2022.10.008
Haq, S. M., Waheed, M., Darwish, M., Siddiqui, M. H., Goursi, U. H., Kumar, M., et al. (2024b). Biodiversity and carbon stocks of the understory vegetation as indicators for forest health in the Zabarwan Mountain range, Indian Western Himalaya. Ecol. Indic. 159:111685. doi: 10.1016/j.ecolind.2024.111685
Haq, S. M., Yaqoob, U., Calixto, E. S., Kumar, M., Rahman, I. U., Hashem, A., et al. (2021). Long-term impact of transhumance pastoralism and associated disturbances in high-altitude forests of Indian Western Himalaya. Sustain. For. 13:12497. doi: 10.3390/su132212497
Ilyas, M., Qureshi, R., Akhtar, N., Ziaul-Haq, M. K. A., and Khan, A. M. (2018). Floristic diversity and vegetation structure of the remnant subtropical broad leaved forests from Kabal valley, swat, Pakistan. Pak. J. Bot. 50, 217–230.
Ivanova, N., Fomin, V., and Kusbach, A. (2022). Experience of forest ecological classification in assessment of vegetation dynamics. Sustain. For. 14:3384. doi: 10.3390/su14063384
Joshi, V. C., Bisht, D., Sundriyal, R. C., and Pant, H. (2023). Species richness, diversity, structure, and distribution patterns across dominating forest communities of low and mid-hills in the central Himalaya. Geol. Ecol. Landsc. 7, 329–339. doi: 10.1080/24749508.2021.2022424
Joshi, V. C., Sundriyal, R. C., and Arya, D. (2021). Forest floor diversity, distribution and biomass pattern of oak and Chir-pine forest in the Indian Western Himalaya. Indian J. Ecol. 48, 232–237.
Karr, J. R., Larson, E. R., and Chu, E. W. (2022). Ecological integrity is both real and valuable. Conserv. Sci. Pract. 4:e583. doi: 10.1111/csp2.583
Khoja, A. A., Haq, S. M., Majeed, M., Hassan, M., Waheed, M., Yaqoob, U., et al. (2022). Diversity, ecological and traditional knowledge of pteridophytes in the western Himalayas. Diversity 14:628. doi: 10.3390/d14080628
Kutnar, L., Nagel, T. A., and Kermavnar, J. (2019). Effects of disturbance on understory vegetation across Slovenian forest ecosystems. Forests 10:1048. doi: 10.3390/f10111048
LaRue, E. A., Fahey, R. T., Alveshere, B. C., Atkins, J. W., Bhatt, P., Buma, B., et al. (2023). A theoretical framework for the ecological role of three-dimensional structural diversity. Front. Ecol. Environ. 21, 4–13. doi: 10.1002/fee.2587
Legendre, P., and Anderson, M. J. (1999). Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 69, 1–24. doi: 10.1890/0012-9615(1999)069[0001:DBRATM]2.0.CO;2
Liu, J., Coomes, D. A., Gibson, L., Hu, G., Liu, J., Luo, Y., et al. (2019). Forest fragmentation in China and its effect on biodiversity. Biol. Rev. 94, 1636–1657. doi: 10.1111/brv.12519
Liu, D., Semenchuk, P., Essl, F., Lenzner, B., Moser, D., Blackburn, T. M., et al. (2023). The impact of land use on non-native species incidence and number in local assemblages worldwide. Nat. Commun. 14:2090. doi: 10.1038/s41467-023-37571-0
Lloret, F., Keeling, E. G., and Sala, A. (2011). Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos 120, 1909–1920. doi: 10.1111/j.1600-0706.2011.19372.x
Maiti, S. K., Bandyopadhyay, S., and Mukhopadhyay, S. (2021). “Importance of selection of plant species for successful ecological restoration program in coal mine degraded land” in: Phytorestoration of abandoned mining. (Elsevier), 325–357.
Manan, F., Khan, S. M., Muhammad, Z., Ahmad, Z., Abdullah, A., Han, H., et al. (2022). Floristic composition, biological spectrum, and phytogeographic distribution of the bin Dara Dir, in the western boundary of Pakistan. Front. For. Glob. Change. 5:1019139. doi: 10.3389/ffgc.2022.1019139
McLane, C. R., Battaglia, L. L., Gibson, D. J., and Groninger, J. W. (2012). Succession of exotic and native species assemblages within restored floodplain forests: a test of the parallel dynamics hypothesis. Restor. Ecol. 20, 202–210. doi: 10.1111/j.1526-100X.2010.00763.x
Mehraj, G., Khuroo, A. A., Qureshi, S., Muzafar, I., Friedman, C. R., and Rashid, I. (2018). Patterns of alien plant diversity in the urban landscapes of global biodiversity hotspots: a case study from the Himalayas. Biodivers. Conserv. 27, 1055–1072. doi: 10.1007/s10531-017-1478-6
Meyer, S. E., Callaham, M. A., Stewart, J. E., and Warren, S. D. (2021). “Invasive species response to natural and anthropogenic disturbance” in: Invasive species in forests and rangelands of the United States: A comprehensive science synthesis for the United States forest sector, (Springer Nature). 85–110.
Miller, J. E., and Safford, H. D. (2020). Are plant community responses to wildfire contingent upon historical disturbance regimes? Glob. Ecol. Biogeogr. 29, 1621–1633. doi: 10.1111/geb.13115
Moldan, B., Janoušková, S., and Hák, T. (2012). How to understand and measure environmental sustainability: indicators and targets. Ecol. Indic. 17, 4–13. doi: 10.1016/j.ecolind.2011.04.033
Moro, M. F., Silva, I. A., Araújo, F. S. D., Nic Lughadha, E., Meagher, T. R., and Martins, F. R. (2015). The role of edaphic environment and climate in structuring phylogenetic pattern in seasonally dry tropical plant communities. PLoS One 10:e0119166. doi: 10.1371/journal.pone.0119166
Mueller-Dombois, D., and Ellenberg, H. (1974). Vegetation types: a consideration of available methods and their suitability for various purposes. Available at: http://hdl.handle.net/10125/29130
Murphy, S. M., Richards, L. A., and Wimp, G. M. (2020). Arthropod interactions and responses to disturbance in a changing world. Front. Ecol. Evol. 8:93. doi: 10.3389/fevo.2020.00093
Nasir, E., and Ali, S. I. (1970–1985). “Flora of Pakistan”. nos. 1–167 (1970–1985). Islamabad: University of Karachi Press. 1–167.
Negi, G. C. S., Joshi, S., Singh, P., and Joshi, R. (2022). Phenological response patterns of forest communities to annual weather variability at long-term ecological monitoring sites in Western Himalaya. Trees For. People. 8:100237. doi: 10.1016/j.tfp.2022.100237
Nowak-Olejnik, A., Mocior, E., Hibner, J., and Tokarczyk, N. (2020). Human perceptions of cultural ecosystem services of semi-natural grasslands: the influence of plant communities. Ecosys. Serv. 46:101208. doi: 10.1016/j.ecoser.2020.101208
Oettel, J., and Lapin, K. (2021). Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 122:107275. doi: 10.1016/j.ecolind.2020.107275
Paillet, Y., Bergès, L., Hjältén, J., Ódor, P., Avon, C., Bernhardt-Römermann, M., et al. (2010). Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 24, 101–112. doi: 10.1111/j.1523-1739.2009.01399.x
Pei, N. C., Kress, W. J., Chen, B. F., Erickson, D. L., Wong, K. M., Zhang, J. L., et al. (2015). Phylogenetic and climatic constraints drive flowering phenological patterns in a subtropical nature reserve. J. Plant Ecol. 8, 187–196. doi: 10.1093/jpe/rtv009
Petersen, T. K., Speed, J. D., Grøtan, V., and Austrheim, G. (2021). Competitors and ruderals go to town: plant community composition and function along an urbanization gradient. Nord. J. Bot. 39. doi: 10.1111/njb.03026
Pfadenhauer, J. S., and Klötzli, F. A. (2020). Global vegetation: fundamentals, ecology and distribution : Springer Nature.
Piotto, D., Magnago, L. F. S., Montagnini, F., Ashton, M. S., Oliver, C., and Thomas, W. W. (2021). Nearby mature forest distance and regenerating forest age influence tree species composition in the Atlantic forest of southern Bahia, Brazil. Biodivers. Conser. 30, 2165–2180. doi: 10.1007/s10531-021-02192-w
Poorter, L., Rozendaal, D. M., Bongers, F., Almeida, D. J. S., Álvarez, F. S., Andrade, J. L., et al. (2021). Functional recovery of secondary tropical forests. PNAS 118:e2003405118. doi: 10.1073/pnas.2003405118
Poudyal, B. H., Maraseni, T., and Cockfield, G. (2019). Impacts of forest management on tree species richness and composition: assessment of forest management regimes in Tarai landscape Nepal. Appl. Geogr. 111:102078. doi: 10.1016/j.apgeog.2019.102078
Qazi, A. W., Saqib, Z., and Zaman-ul-Haq, M. (2023). Diversity and distribution of endemic flora in Pakistan. Proc. Pak. Acad. Sci.: B. 60, 165–179. doi: 10.53560/PPASB(60-2)777
Rahman, I. U., Afzal, A., Iqbal, Z., Alzain, M. N., al-Arjani, A. B. F., Alqarawi, A. A., et al. (2021). Classification and characterization of the Manoor Valley’s (lesser Himalaya) vegetation from the subtropical-temperate ecotonal forests to the alpine pastures along ecological variables. Plan. Theory 11:87. doi: 10.3390/plants11010087
Rai, P. K. (2022). Environmental degradation by invasive alien plants in the anthropocene: challenges and prospects for sustainable restoration. Anthrop. Sci. 1, 5–28. doi: 10.1007/s44177-021-00004-y
Ramovs, B. V., and Roberts, M. R. (2003). Understory vegetation and environment responses to tillage, forest harvesting, and conifer plantation development. Ecol. Appl. 13, 1682–1700. doi: 10.1890/02-5237
Ramprasad, V., Joglekar, A., and Fleischman, F. (2020). Plantations and pastoralists: afforestation activities make pastoralists in the Indian Himalaya vulnerable. Ecol. Soc. 25:1. doi: 10.5751/ES-11810-250401
Rashid, I., Haq, S. M., Lembrechts, J. J., Khuroo, A. A., Pauchard, A., and Dukes, J. S. (2021). Railways redistribute plant species in mountain landscapes. J. Appl. Ecol. 58, 1967–1980. doi: 10.1111/1365-2664.13961
Raunkiaer, C. (ed.) (1934). The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiær. 58, 1967–1980.
Sanderson, M. A., Skinner, R. H., Barker, D. J., Edwards, G. R., Tracy, B. F., and Wedin, D. A. (2004). Plant species diversity and management of temperate forage and grazing land ecosystems. Crop Sci. 44, 1132–1144. doi: 10.2135/cropsci2004.1132
Shin, Y. J., Midgley, G. F., Archer, E. R., Arneth, A., Barnes, D. K., Chan, L., et al. (2022). Actions to halt biodiversity loss generally benefit the climate. Glob. Change Biol. 28, 2846–2874. doi: 10.1111/gcb.16109
Singh, S. P., Gumber, S., Singh, R. D., and Singh, G. (2020). How many tree species are in the Himalayan treelines and how are they distributed? Trop. Ecol. 61, 317–327. doi: 10.1007/s42965-020-00093-7
Smith, M. D., Knapp, A. K., and Collins, S. L. (2009). A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecol. 90, 3279–3289. doi: 10.1890/08-1815.1
Song, Z., Du, Y., Primack, R. B., Miller-Rushing, A. J., Ye, W., and Huang, Z. (2021). Surprising roles of climate in regulating flowering phenology in a subtropical ecosystem. Ecography 44, 1379–1390. doi: 10.1111/ecog.05629
Subashree, K., Dar, J. A., Karuppusamy, S., and Sundarapandian, S. (2021). Plant diversity, structure and regeneration potential in tropical forests of Western Ghats, India. Acta Ecol. Sin. 41, 259–284. doi: 10.1016/j.chnaes.2020.02.004
Theodorou, P. (2022). The effects of urbanisation on ecological interactions. Curr. Opin. Insect Sci. 52:100922. doi: 10.1016/j.cois.2022.100922
Ullah, R., Khan, N., and Shaukat, S. S. (2022). Biomass allocation of Xanthium strumarium L.: a key factor for invasive success at high elevation. Acta Botan. Brasi. 36:e2021abb0272. doi: 10.1590/0102-33062021abb0272
Vakhlamova, T., Wagner, V., Padulles Cubino, J., Chytrý, M., and Lososová, Z. (2022). Urban plant diversity in Kazakhstan: effects of habitat type, city size and macroclimate. Appl. Veget. Sci. 25:e12679. doi: 10.1111/avsc.12679
Vogel, J. G., Bracho, R., Akers, M., Amateis, R., Bacon, A., Burkhart, H. E., et al. (2021). Regional assessment of carbon pool response to intensive silvicultural practices in loblolly pine plantations. Forests 13:36. doi: 10.3390/f13010036
Waheed, M., Arshad, F., Iqbal, M., Fatima, K., and Fatima, K. (2020). Ethnobotanical assessment of woody flora of district Kasur (Punjab), Pakistan. Ethnobot. Res. Appl. 20:33.
Waheed, M., Haq, S. M., Arshad, F., Bussmann, R. W., Ali, H. M., and Siddiqui, M. H. (2023a). Phyto-ecological distribution patterns and identification of alien invasive indicator species in relation to edaphic factors from semi-arid region. Ecol. Indic. 148:110053. doi: 10.1016/j.ecolind.2023.110053
Waheed, M., Haq, S. M., Arshad, F., Bussmann, R. W., Iqbal, M., Bukhari, N. A., et al. (2022a). Grasses in semi-arid lowlands-community composition and spatial dynamics with special regard to the influence of edaphic factors. Sustain. For. 14:14964. doi: 10.3390/su142214964
Waheed, M., Haq, S. M., Fatima, K., Arshad, F., Bussmann, R. W., Masood, F. R., et al. (2022b). Ecological distribution patterns and indicator species analysis of climber plants in Changa manga Forest plantation. Diversity 14:988. doi: 10.3390/d14110988
Waheed, M., Haq, S. M., Jameel, M. A., Arshad, F., and Bussmann, R. W. (2023b). Documentation of ethnomedicinal plants used by the people living in reserved forests of semi-arid region Punjab Pakistan. Ethnobot. Res. Appl. 26:39. doi: 10.32859/era.26.39.1-17
Wang, X., Hua, F., Wang, L., Wilcove, D. S., and Yu, D. W. (2019). The biodiversity benefit of native forests and mixed-species plantations over monoculture plantations. Divers. Distrib. 25, 1721–1735. doi: 10.1111/ddi.12972
Williams, J. J., and Newbold, T. (2020). Local climatic changes affect biodiversity responses to land use: a review. Divers. Distrib. 26, 76–92. doi: 10.1111/ddi.12999
Wright, S. J. (2010). The future of tropical forests. Ann. N. Y. Acad. Sci. 1195, 1–27. doi: 10.1111/j.1749-6632.2010.05455.x
Zhou, Y., Su, Y., Zhong, Y., Xie, P., Xu, M., and Su, Z. (2019). Community attributes predict the relationship between habitat invasibility and land use types in an agricultural and forest landscape. Forests 10:867. doi: 10.3390/f10100867
Keywords: species diversity, plantations, forest management, subtropical, Pakistan
Citation: Waheed M, Haq SM, Arshad F, Bussmann RW, Hashem A and Abd_Allah EF (2024) Plant distribution, ecological traits and diversity patterns of vegetation in subtropical managed forests as guidelines for forest management policy. Front. For. Glob. Change. 7:1406075. doi: 10.3389/ffgc.2024.1406075
Edited by:
Gopal Shukla, North Eastern Hill University, IndiaReviewed by:
Shivani Dobhal, Central Agricultural University, IndiaKaushal Singh, Banda University of Agriculture and Technology, India
Copyright © 2024 Waheed, Haq, Arshad, Bussmann, Hashem and Abd_Allah. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Waheed, ZjE5LXBoZC1ib3QtNTAxM0B1by5lZHUucGs=