 Pandu Y. A. P. Wirabuana1
Pandu Y. A. P. Wirabuana1 Budi Mulyana2Himlal Baral3Rina L. Hendrati4
Budi Mulyana2Himlal Baral3Rina L. Hendrati4 I. L. G. Nurtjahjaningsih4Mashudi4Dedi Setiadi4Sugeng Pudjiono4Sumardi4
I. L. G. Nurtjahjaningsih4Mashudi4Dedi Setiadi4Sugeng Pudjiono4Sumardi4 Liliana Baskorowati4*
Liliana Baskorowati4*- 1Faculty of Forestry, Gadjah Mada University, Yogyakarta, Indonesia
- 2Faculty of Forestry, University of Sopron, Sopron, Hungary
- 3Center for International Forestry Research and World Agroforestry (CIFOR-ICRAF), Jl CIFOR Situ Gede, Bogor, Indonesia
- 4Research Center for Applied Botany, National Research and Innovation Agency, Bogor, Indonesia
Introduction: This study estimates community forest tree species’ energy using allometric models. Accurate tree-level energy stock estimation is needed to evaluate community forest tree species for renewable energy development. Three commercial species in community forests utilized as temporary energy sources, Falcataria moluccana, Swietenia macrophylla, and Tectona grandis were sampled by cutting 45 trees. Those tree types were investigated as co-firing materials supplied by the community while the region develops a strategy including finding the best species for continuous wood energy stock to power Java and Bali.
Methods: This study recorded data to compare with the potential determined fast-growing species established specifically for the next annual energy stock. Multiplying the biomass calorific value gave each tree component’s energy stock and adding all energy stock tree components result in an estimate for each tree. Regression analysis was used to make allometric models that use diameter at breast height (DBH) and tree height as predictors.
Results: The calorific value of components varied among species, with foliage having the highest (5,021.24 ± 39.37 kcal kg−1), followed by stem (4,352.8 ± 32.07 kcal kg−1) and branches (4,200.11 ± 33.54 kcal kg−1). Thus, leaves could be used for energy as trees are harvested for timber. All species stored most energy in stems (60.59 0.97%), branches (27.04 1.13%), and leaves (12.38 0.84%).
Discussion: As DBH classes grew, the energy stock of stems and branches improved but foliage fell. With adjusted R2adj of 0.84–0.99 and root mean square error (RMSE) of 0.03–0.64, allometric models accurately assessed species energy stocks in each component and the overall tree. Tree height does not necessarily improve model accuracy; therefore, each species had a different best-fit model. Generalized allometric models for all species might estimate component and tree energy storage and our study demonstrated that allometric models could measure community forest tree species energy stocks. These should also be estimated for the next species for co-firing wood biomass when coppice harvest is applied. The above alternative species and the proposed much faster-growing species for co-firing are compared.
1 Introduction
Renewable energy development is gaining great concern, along with the increasing impacts of climate change. This has caused an increase in the demand for alternative energy to minimize greenhouse gas emissions. Nowadays, several alternative renewable energy sources have been identified; one of them is forest wood biomass, especially from abundant trees grown in the community forest (Da Silva et al., 2019). Forest wood biomass is a good option for energy sources since it can be regenerated through reforestation. The demand for forest biomass also gradually increases over time and provides extraordinary market opportunities not only for commercial plantation forests but also for local people to get advantages from their community forest (Favero et al., 2023). Although the ideal use of most trees is for timber, people in the community often require immediate funding available for their needs, so they frequently sell them for lower prices. This includes the three species in this study, which are Falcataria mollucana, a fast-growing species, Swietenia macrophylla, and Tectona grandis, two slow-growing species.
Although some legume species have been used and developed for years for fuel wood in Indonesia, in the first place, it is the timber tree species that is mostly grown much more by the local people in this study (Hendrati et al., 2022; Wirabuana et al., 2022). Forest biomass from woody trees is an ideal raw material for producing charcoal and wood pellets, including those three species. Thus, it is also qualified to be used as co-fuel in power plants in accordance with the Indonesian government’s plans to achieve a mixture of 23% renewable energy in 2025 and 31% in 2050 (Ministry of Energy and Mineral Resources, Republic of Indonesia, 2024). Therefore, this study was undertaken in community forests at Srobyong Village, Jepara District, which is near to the power plant for supplying electricity for Java and Bali. It is also one of the priority zones for the biomass pool in supporting the national co-firing program. Indonesia Electricity Company, represented by PLN, is promoting, and empowering the people’s economy throughout Indonesia’s provinces by inviting the community to participate actively in the production of co-firing raw materials, from planting biomass plants to managing household waste in the region to be converted into pellets (Ministry of Energy and Mineral Resources, 2020). This includes the Central Java province, which receives provincial and government support. Because there is a minimal biomass plantation specifically established for co-firing raw materials, the community was charmed to seize this opportunity to earn money from their available plantations.
The community forest is substantially different from plantation forests, which are mainly monoculture, especially the short-rotation, fast-growing species. Meanwhile, community forests have multiple tree species with an irregular age distribution (Wulandari et al., 2018), in which the spacing between trees is not systematically arranged; thus, there is substantially high growth variation in this forest (Boedhihartono, 2017). The community forests are commonly sustainably managed using selective cutting, wherein the harvesting process is only conducted when the landowner needs urgent funding (Setiahadi, 2017; Setiahadi et al., 2020).
Managing community forests for renewable energy is challenging and quantifying the energy stock for those 3 alternative trees needs to be provided. That is essential information for opening people’s minds to the possibility of selecting new, much faster-growing and more profitable species. A continuous annual harvest from coppice from energy wood species is considered advantageous, such as the easy coppice of three legume species for energy: L. leucocephala, A. auriculiformis, and C. calothyrsus (Hendrati and Nurrohmah, 2018; Hendrati et al., 2022). This may inspire a more sustainable way to provide a long-term (15–20 year) annual harvest for wood energy which now is under consideration to be established by the local government.
This study was undertaken in the tropical country of Indonesia, which only has dry and wet seasons, specifically carried out in the heaviest populated island of Java, as the center of most government business. One of the regencies on the Java Island, Jepara is part of the Central Java Province. In Jepara regency, community forests accounted for 18,285.78 ha of the total forest area in 2014; by 2024, that amount had dropped to 13,975 ha. F. moluccana, T. grandis, and S. mahagony trees predominate in community forests, and they are frequently planted with agricultural products like tubers and spices. With 19,982 business units in the furniture sector, Jepara regency boasts a sizable furniture industry that uses wood as its primary raw material. The waste generated from the wood used in furniture production is utilized as firewood. However, in certain instances, communities make use of twigs, branches, and small logs as a source of firewood.
In this study, the assessment of energy stocks at the individual tree level is conducted by cutting the whole tree. This method is intended to achieve the highest level of accuracy (Simetti et al., 2018). However, this approach needs to be applied with sampling to minimize significant financial expenses, time consumption, but sufficiently avoiding and a lack of efficiency when used on a broad scale. This also avoiding a possible risk due to the occurrence of forest disturbances and a subsequent decline in regeneration capability (Kora et al., 2019). Therefore, this study practices allometric models that can be used to improve accuracy and reveal their worth for estimating the energy stock for different tree types in the community forests. This provides the approximate biomass production for energy for consideration against the next selected species.
The allometric model is an approximate method frequently used in the forestry sector, both at individual trees and at stand level. It is a reliable model to support the quantification of aboveground biomass (Altanzagas et al., 2019), carbon stock (Khan et al., 2018), and crown characteristics (Coombes et al., 2019). There is some previous research that has documented the applicability of this approach, including for estimating physiological tree properties such as leaf area index (Vyas et al., 2010). This model has also been formulated for estimating the energy stock of tree species in plantation forests, specifically Eucalyptus urophylla (Sadono et al., 2022). This suggests the potency of this model for estimating across many ecosystem types, including community forests. Nevertheless, this allometric model for the energy stock estimation in community forests has not yet been undertaken, especially because of the diverse array of tree species found within the community forest used for energy. The allometric model is expected to offer reliable predictions for individual or many tree species, applicable for future use. Because there has been no calculation before of those timber species by the local community when used as wood energy.
F. moluccana, S. macrophylla, and T. grandis are the three commercial species that locals typically choose and cultivate as alternatives for completing the co-firing program. This study aims to develop allometric models for estimating the energy stock of tree species mostly grown in the community forests that are used by people for supplying wood as co-firing in Jepara, Indonesia. It is one of the priority areas for the co-firing program in Indonesia, it is expected to become a good example of quantifying the total energy supply available. It is expected that this study can be used as a comparison in the future for establishing specific wood energy plantations from other faster-growing species.
Therefore, we hypothesize that: (i) the allometric models can accurately estimate the energy stock of those species in a community forest, wherein, more than 70% of the energy stock variation in each tree component and the total energy stock of each tree; (ii) the best allometric model for each species might be relatively different, and (iii) the general allometric models can be developed for all species, both in every component and for the total individual tree.
2 Materials and methods
2.1 Study area
This study was done in community woodlands at Srobyong village, Jepara district, Central Java province, Indonesia (Figure 1), one of the key zones for the biomass pool in supporting the National Co-Fairing. Its coordinates are 110°9′48.02″ and 110°58′37.40″ East and 5°43′20.67″ to 6°47′25.85″ N. It is near the power plants generating electricity for Java and Bali. The topography of the land consists primarily of flat regions characterized by a gradient ranging from 0 to 8% and an elevation between 70 and 150 meters above sea level. The average daily temperature is 29°C, accompanied by a mean air humidity of 84%. Annual rainfall is 2,446 mm year–1, with dry intervals for 4 months from May to August. The soil is categorized as alfisols, with a pH range of 5.5–6.0. Approximately 52% of the land cover in the research site was largely community forests, with an approximate size of 19.516 ha (Central Java Provincial Statistics Bureau, 2022).
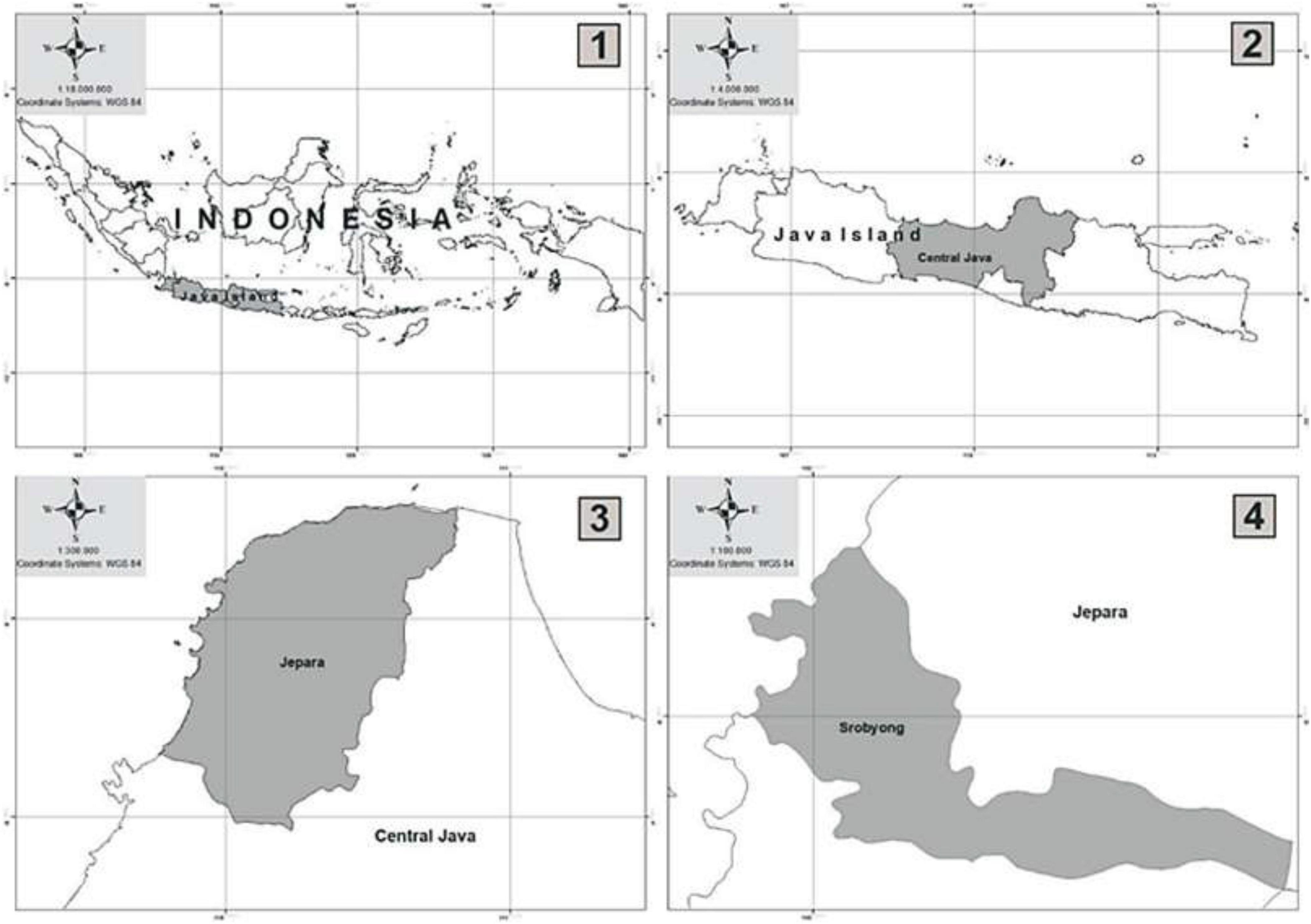
Figure 1. Location of the study site in community forests at Srobyong District.
2.2 Tree sampling
The forest inventory was conducted utilizing the quadrat method, employing a sampling plot size of 25 m × 25 m (Wirabuana et al., 2021). A total of 64 units of permanent plots were randomly dispersed over the study site for the inventory, as depicted in Figure 2. Thus, this inquiry covered a sampling area of 4 hectares. Data was collected from each plot, encompassing the landowner’s name, tree species, diameter at breast height (DBH), and tree height (H). The DBH was measured at a height of 1.3 meters above the ground using a phi band, while the H was measured from the ground to the top of the tree crown using a spiegel. Each tree within every sampling plot was assigned a unique numerical tag to simplify the identification of specific tree samples for the purpose of sampling through complete tree removal. Upon completion of the forest inventory, all the data were merged into a single DBH, which was divided into four distinct groups: 0-10 cm, 11-20 cm, 21-30 cm, and ≥ 31 cm (Altanzagas et al., 2019; Wirabuana et al., 2020; Setiahadi, 2021). The goal was to determine the abundance of trees spanning from small number to abundance. Previous studies frequently utilized this approach to get tree samples that precisely represented the variation in growth (Chave et al., 2014; Mulyana et al., 2020; Sadono et al., 2021). The number of tree samples in our study was limited to 45 due to resource restrictions and the necessity for permission from landowners. However, an equal number of samples were assigned to each species. Previous research, with much smaller sample sizes (Youkhana and Idol, 2011; Stas et al., 2017; Sadono et al., 2022) have shown that our modest sample sizes were enough for generating the allometric models.
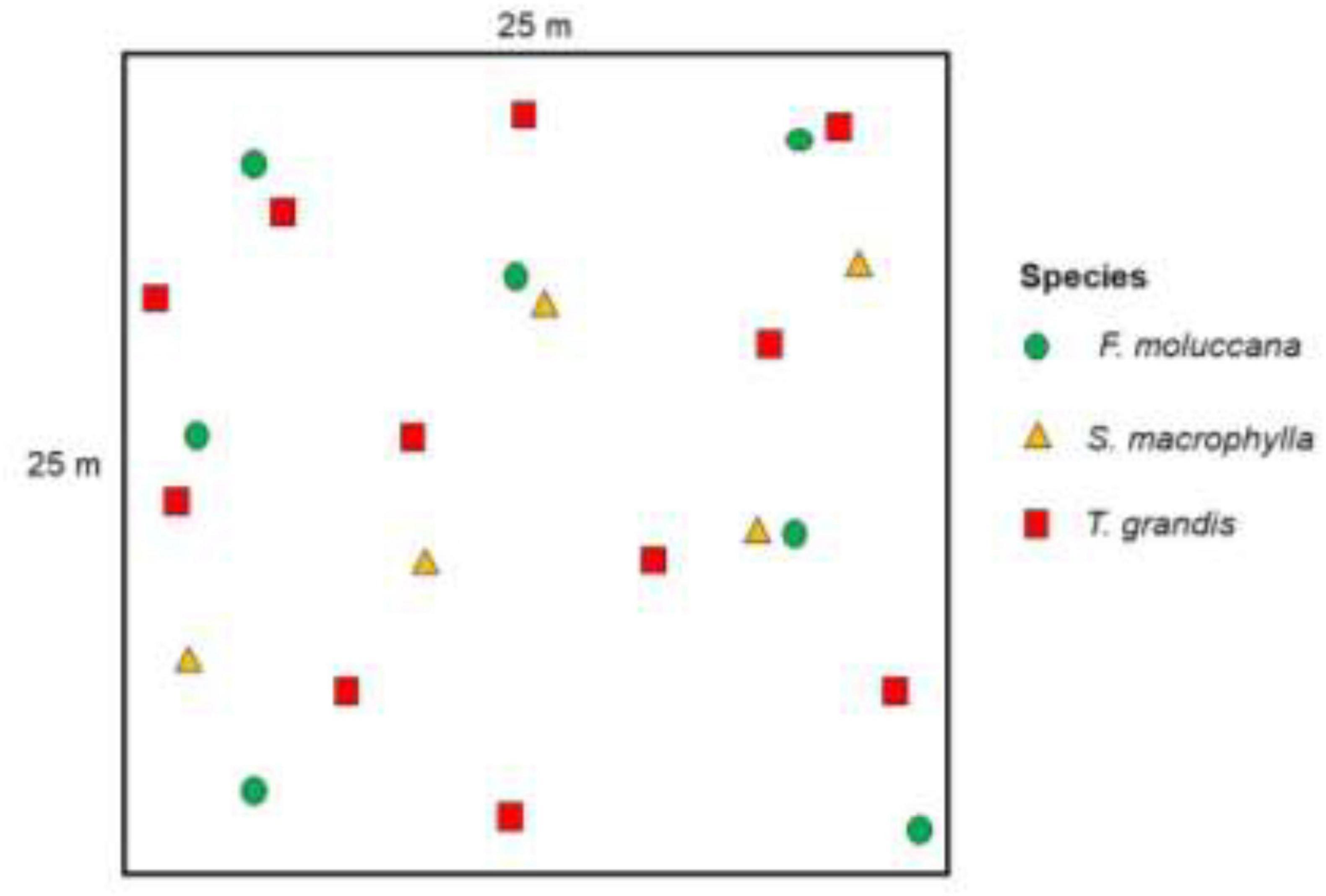
Figure 2. Unit plot illustration in community forest as Srobyong District.
The process of cutting the whole tree was conducted and the GPS was used to record the coordinates of each tree sample (Figure 2). Once the chosen tree was cut down, its parts were divided into trunks, branches, and leaves. The weight of each component of the tree was measured using a suspended scale. Subsequently, around 500 g of subsample was obtained from each component and subjected to drying in the laboratory using a forced-air over at 70°C for 48 h for biomass determination. The total biomass in every component (Equation 1) and at the individual tree level (Equation 2) were calculated following these equations:
where Wc is the biomass of tree component (kg), DWs is a dry weight of subsample (g), FWs is a fresh weight of subsample (g), FWc is the total fresh weight of tree component (kg), Wt is the total biomass at the individual tree level (kg), Ws is the stem biomass (kg), Wb is the branches biomass (kg), and Wf is the foliage biomass (kg).
2.3 Determination of energy stock
The desiccated subsamples were examined utilizing the standard of the American Society for Testing and Material (ASTM E-711-87) to measure the calorific value of each individual tree component. The energy stock of each component (Equation 3) and the total energy stock (Equation 4) were calculated using these formulas:
where ESc is an energy stock of tree component (MJ), Wt is the total biomass at the individual tree level (kg), HHVc is a calorific value of tree component (kcal kg–1), f is a constant of 4.184 × 10–3, ESt is the total energy stock at the individual tree level (MJ), ESs is an energy stock in stem (MJ), ESb is an energy stock in branches, and ESf is an energy stock in foliage.
This study also calculated the energy expansion factor (EEF) and energy harvesting index (EHI). EEF is the ratio between the total energy stock at the individual tree level and the energy stock in stem (Equation 5). EEF can be used as a constant to convert the energy stock in the stem component to the total individual tree. Meanwhile, EHI is the ratio between the allowable total energy harvested and the total energy produced from the individual tree (Equation 6). Woody components like stem and branches are generally classified as the allowable harvested. Both parameters were calculated using the following equations:
where EEF is an energy expansion factor of species, ESt is the total energy stock at the individual tree level (MJ), ESs is an energy stock in stem (MJ), ESb is an energy stock in branches (MJ) and EHI is an energy harvesting index of species.
2.4 Allometric model development
Data screening was conducted using the Boxplot method to identify the existence of outliers in the dataset. Then, the normality of the data was examined using the Shapiro–Wilk test. Scatter diagrams were also constructed to recognize the pattern of relationships between independent and dependent variables. The DBH and H were selected as the independent variables, while the ES in each tree component and in total were considered as the dependent variables. The scatter plot method was frequently used in previous studies before developing the allometric models (Vega-Nieva et al., 2015; Lisboa et al., 2018; Ogana and Ercanli, 2022).
Two allometric models were examined in this study. Both models differed in the number of independent variables. The first model used DBH as the predictor (Equation 7), while the second applied DBH and H as the predictors (Equation 8). The allometric models were developed in two steps, namely specific species, and the combination of all species. For details, both formulas are shown below:
where Y is the energy stock in tree component or in total (MJ), DBH is the diameter at breast height (cm), H is tree height (m), and a, b, and c are fit coefficients.
The allometric models were evaluated in several stages, i.e., (1) assessing the model fitting; (2) testing the model validation; and (3) checking the significance of predictor addition. There are five things that were used to judge how well the model fit: the ANOVA test result, the fitted parameters (a, b, and c) result, the adjusted coefficient of determination (R2adj) (Equation 9), the residual standard error (RSE) (Equation 10), and the Akaike information criterion (AIC) (Equation 11). Meanwhile, two criteria were selected to examine the validation, such as mean absolute error (MAE) (Equation 12) and root mean square error (RMSE) (Equation 13). We used the leave-one-out cross-validation (LOOCV) method for validating the model due to the small sample size. Previous studies also applied LOOCV when developing models with a small sample size (Altanzagas et al., 2019; Tetemke et al., 2019; Wirabuana et al., 2020; Sadono et al., 2021). High R2adj, small RSE, AIC, MAB, MAE, and RMSE indicate high model precision (Altanzagas et al., 2019). Detailed formula for calculating these criteria is presented below:
where Yi is the observed energy stock, is the average observed energy stock, is the estimated energy stock from the fitted model, R2 is the coefficient of determination, n is the sample size, RSS is the residual sum of squares from the fitted model, k is the number of parameters, and p is the number of terms in the model.
We used the Extra Sums of Square (ESS) to verify the significant contribution of additional H in the allometric models. This method is reliable to check whether the addition of other predictors provides a substantial effect on improving the model’s reliability (Hector et al., 2016; Wirabuana et al., 2021; Sadono et al., 2022). When the result is insignificant, using a single predictor in the allometric equation is sufficient to obtain a good estimation.
3 Results
3.1 Distribution of sample tree
The number of sample trees differed for each species within the DBH class (Table 1). Fifteen trees classified most tree samples into the DBH class of 0–10 cm. There was a total of nine trees in the DBH class, ranging from 11 to 20 cm and 21 to 30 cm. F. moluccana and T. grandis had the most tree samples in the smallest DBH class (0–10 cm), whereas S. macrophylla had a majority in the greatest DBH class (31 cm). Our investigation observed that each species had an equal number of tree samples within the DBH range of 21–30 cm, specifically three trees. Meanwhile, T. grandis had the smallest sample size in DBH class of 11–20 cm, with only two trees. The restrictions imposed by landowners’ permission are the main cause for the variation in the distribution of tree samples among various species and the DBH classes. Owners of trees only permitted destructive sampling of trees that met the criteria.
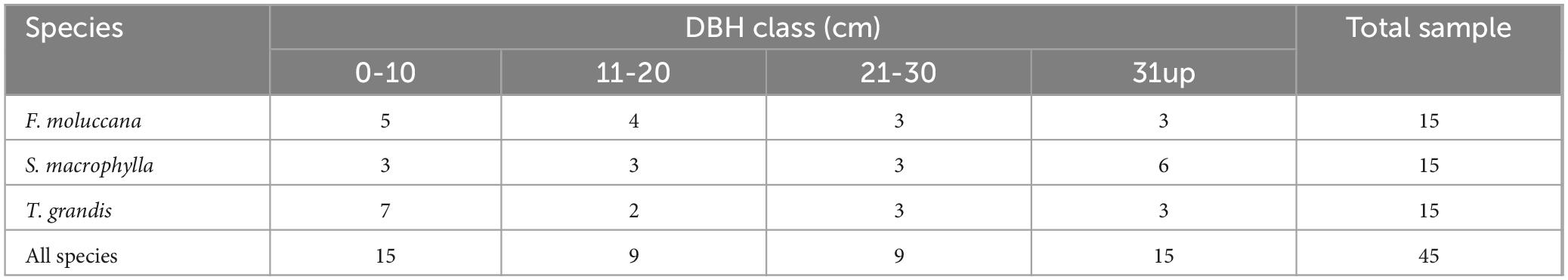
Table 1. The distribution of selected tree samples in every DBH class.
3.2 Calorific value, biomass accumulation, and energy distribution
The calorific value varied significantly among each component of the tree (Table 2). The mean calorific value for all species was highest in foliage (5,021.24 ± 39.37 kcal kg–1), then stems (4,352.8 ± 32.07 kcal kg–1), and finally branches (4,200.11 ± 33.54 kcal kg–1). Across all species, it was consistently observed that the foliage had a larger calorific value compared to other components. Additionally, this study found that the calorific value of T. grandis was comparatively greater than that of F. moluccana and S. macrophylla for every component of the tree. Meanwhile, S. macrophylla exhibited the lowest calorific value among all the species’ components.
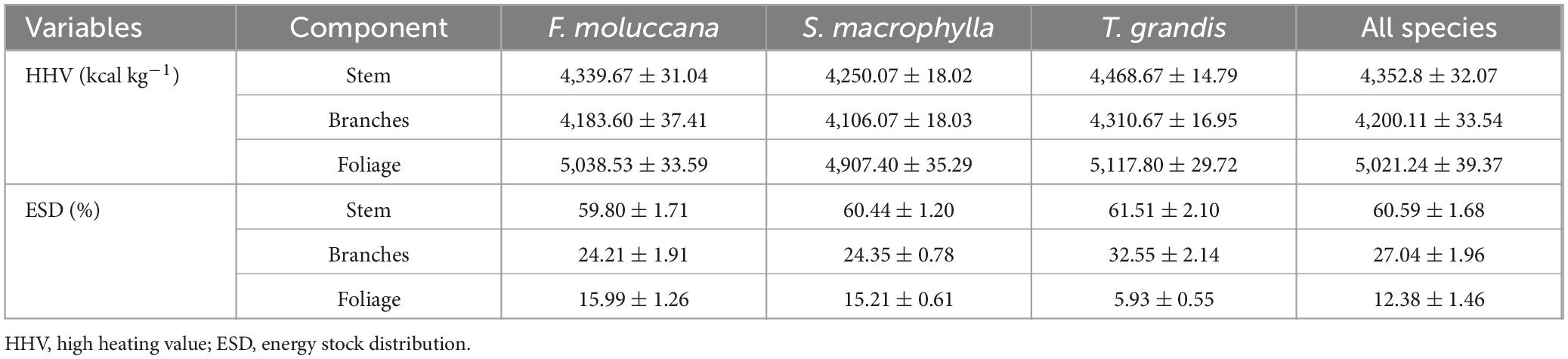
Table 2. Calorific value and energy stock distribution in every species.
Our analysis revealed that while the leaves had the highest calorific value, the stem of each species contained the maximum energy store (Table 2). The energy allocation in stocks showed a consistent decrease from the stem to the foliage in all species, as shown in Table 2. The stem of the individual tree had almost fifty percent of the energy supply since it had the highest biomass accumulation compared to other tree components (Table 3).
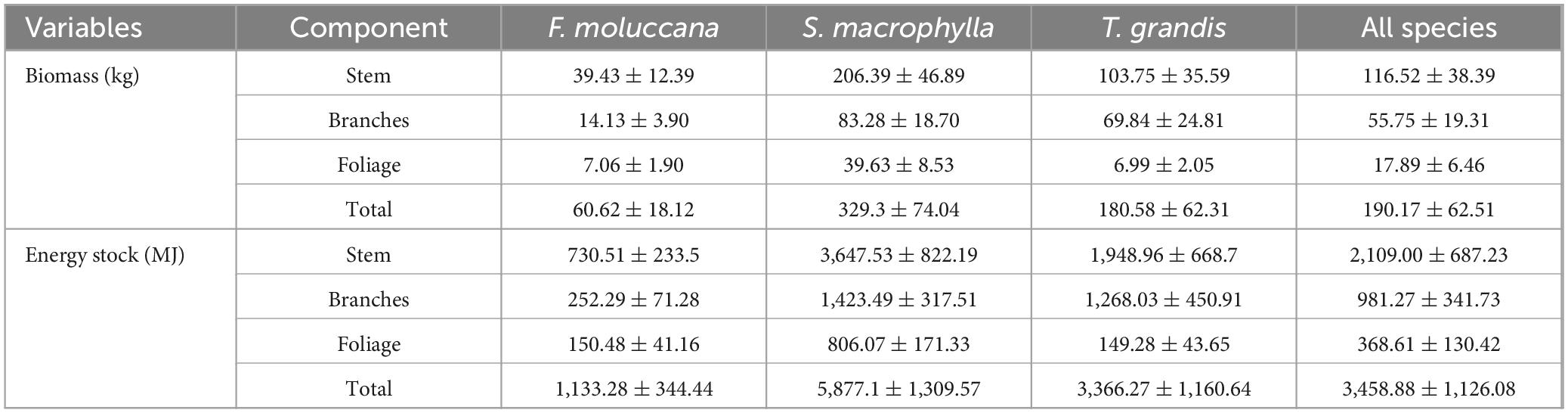
Table 3. Biomass accumulation and energy stock in every species.
Notably, each tree component of all species exhibited a distinct pattern of energy distribution as the DBH class increased (Figure 3). The energy distribution in the stems of F. moluccana and S. macrophylla improved as the DBH class increased. Conversely, it demonstrated a decreasing pattern in T. grandis. The stem exhibited a greater dispersion of energy, but the foliage showed a lesser concentration of energy.
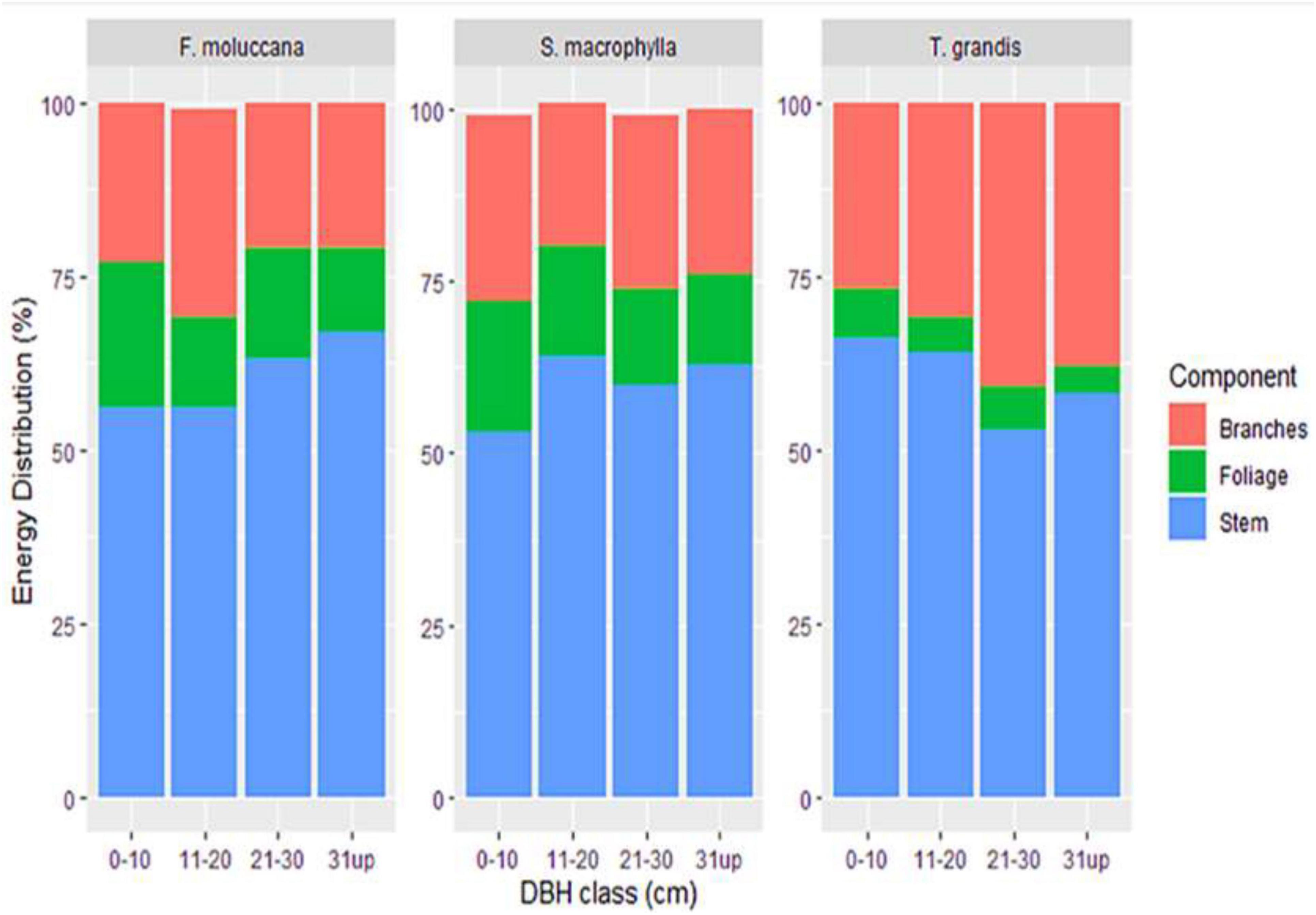
Figure 3. Energy stock distribution in every tree component across the DBH classes.
The findings also revealed a significant variation in the total energy stock among species, with S. macrophylla exhibiting the highest average of 5,877.1 ± 1,309.57 MJ, followed by T. grandis with 3,366.27 ± 1,160.64 MJ, and F. moluccana with 1,133.28 ± 344.44 MJ (Table 3). The species exhibited the highest average energy of 2,109.00 ± 687.23 MJ in the year. Further, the branches were with an average energy of 981.27 ± 341.73 MJ and leaves with an average energy of 368.61 ± 130.42 MJ. So that the wood constituents amassed a greater amount of energy compared to the non-woody components.
The study found that the EEF was almost the same across different species, ranging from 1.66 to 1.69 (Figure 4). Concurrently, the EHI for all species varied between 0.88 and 0.94. This shows that over 85 percent of the energy generated at the level of each individual tree in every species may be collected to facilitate the advancement of renewable energy. T. grandis had a greater EHI compared to F. moluccana and S. macrophylla, with a difference of 0.94 ± 0.01; which demonstrated that T. grandis had a higher energy stock yield compared to other species.
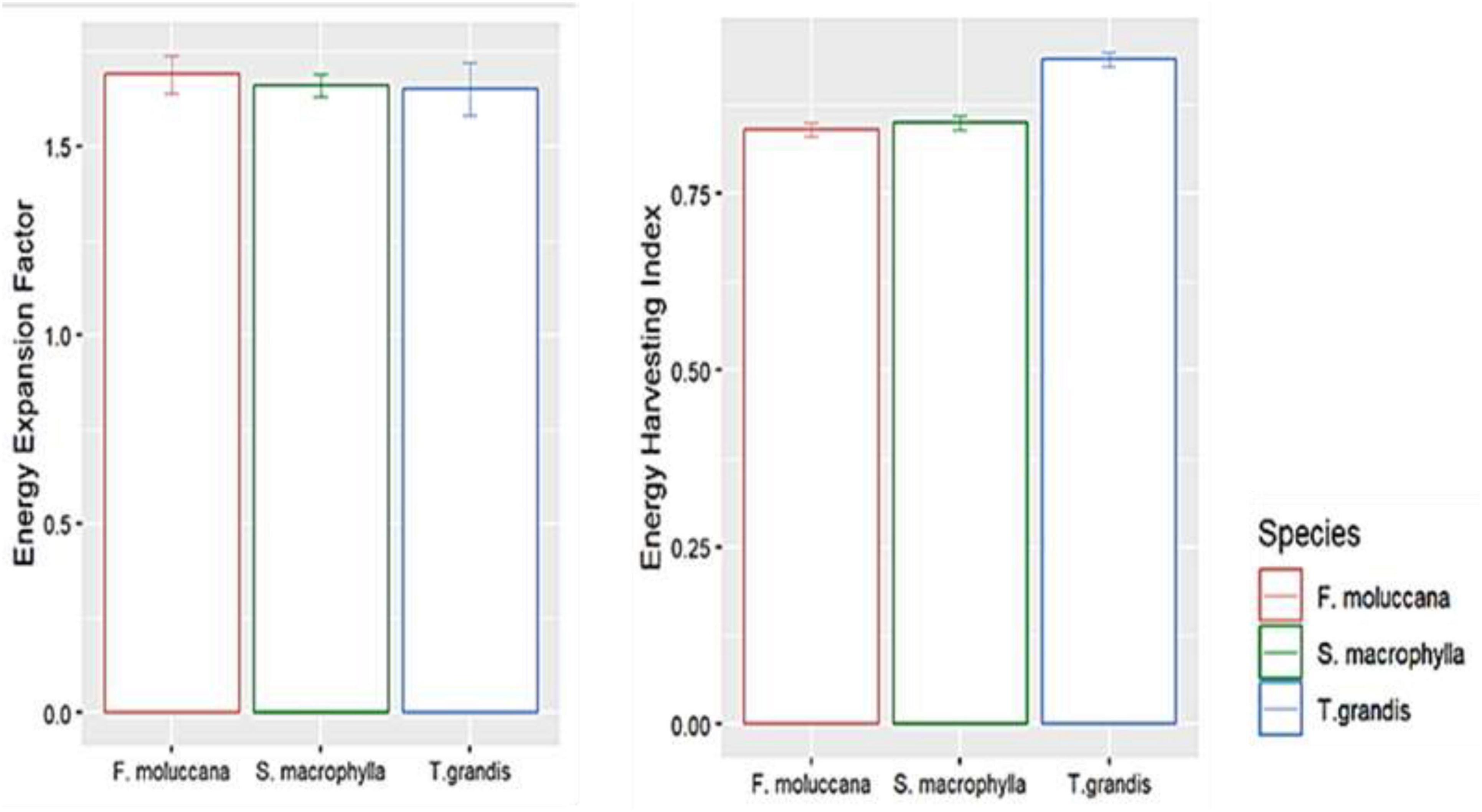
Figure 4. Energy expansion factor and harvesting index in every species.
3.3 Allometric models
The cumulative energy reserves of each species exhibited a progressive increase as the DBH and H increased (Figure 5). These results highlighted that DBH and H have the capability to serve as predictor factors for evaluating the energy stock. The outcomes of constructing allometric models also demonstrated a strong fit for each individual model (Table 4). Furthermore, allometric models were used to explain about 80% of the variations in energy stocks across different parts of the tree and the entire tree for each species. General allometric models can be developed for all species. As shown in Table 4, this model estimates high degree of the energy stock for each individual component of a tree with a precision of up to 90%.
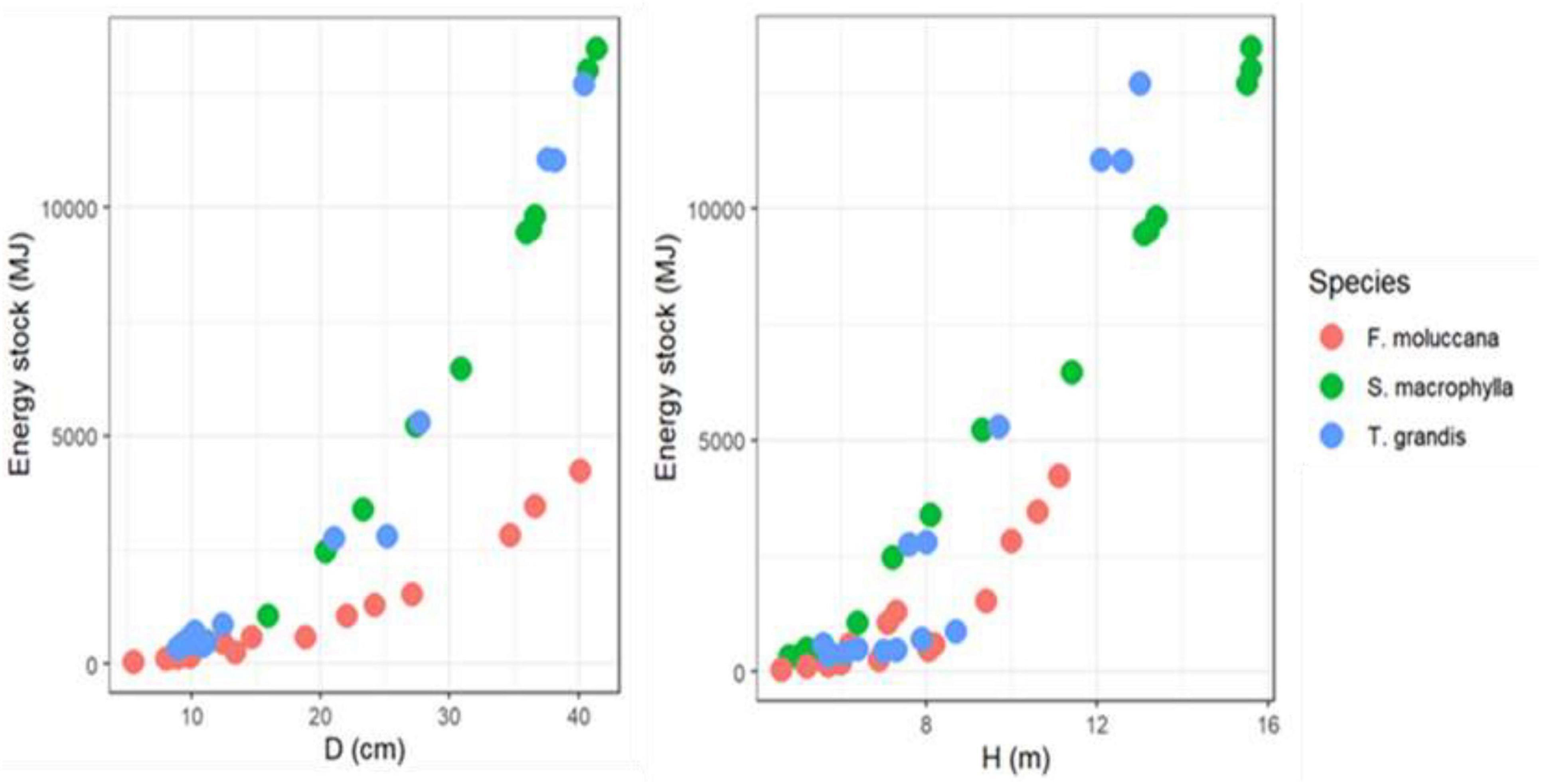
Figure 5. Relationship between independent variables (DBH, H) and dependent variables (ES).
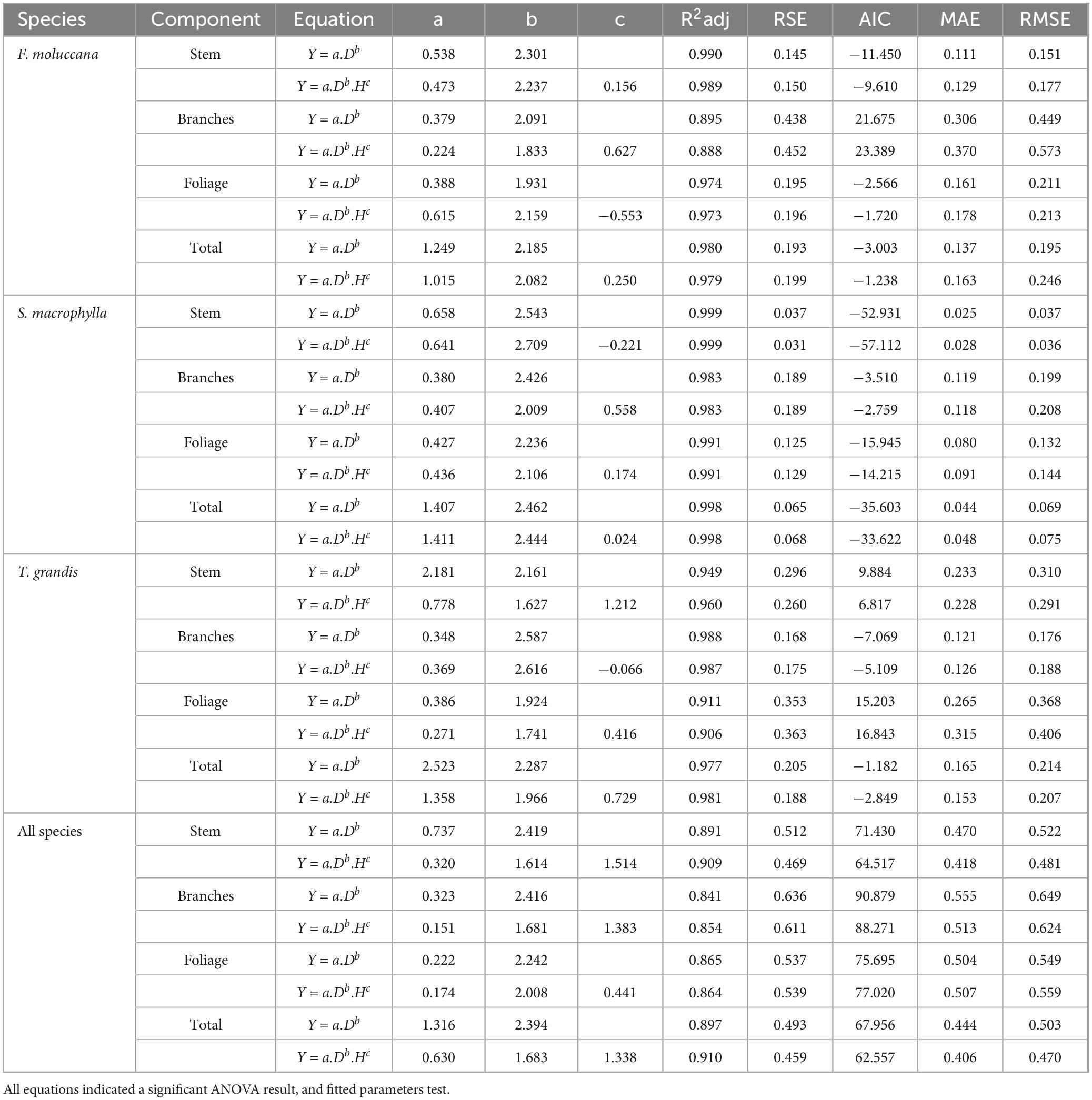
Table 4. Summary evaluation of statistics of every allometric models for estimating energy.
The estimation of the total energy stored at the level of individual trees was determined to be dependable. It was noted that incorporating H as a separate variable in the regression model did not consistently yield a statistically significant enhancement in the model’s predictive precision, as demonstrated in Table 5. The addition of H has significantly improved the precision of the model in determining the energy content in the stems of S. macrophylla and T. grandis, encompassing all species. Additionally, the incorporation of this variable was essential in improving the reliability of the overall allometric models used to calculate the total energy storage. Therefore, the most efficient models for each individual part within each species displayed significant differences.
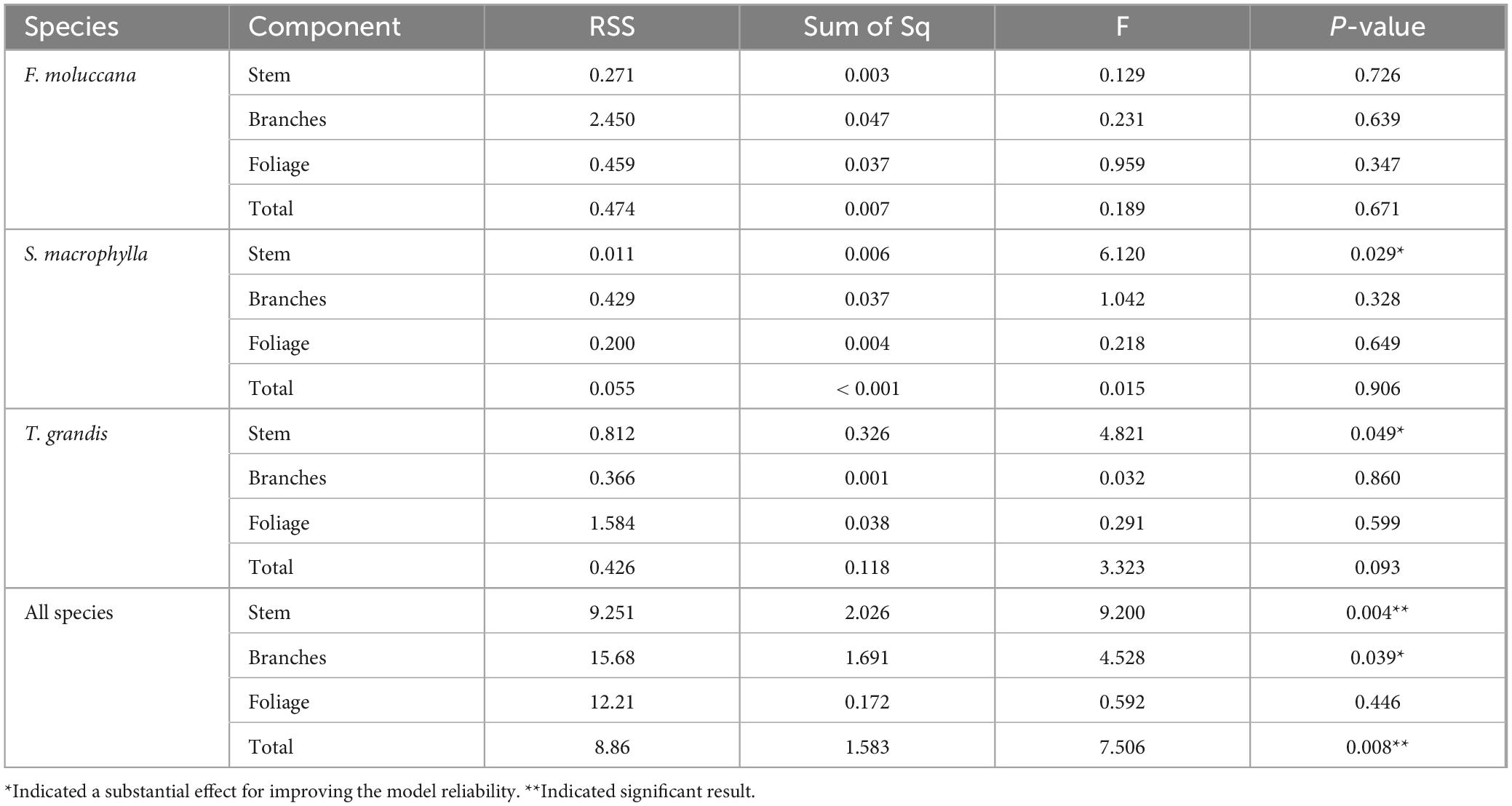
Table 5. Summary results of the ESS test for the allometric equations.
The allometric models underestimated the energy stock value for S. macrophylla and T. grandis compared to the real value. Conversely, it yielded a greater estimate when assessing the energy reserves for F. moluccana (Figure 6). While F. moluccana and T. grandis, the biggest error in estimation was seen in the DBH category of 31 cm, and in S. macrophylla, it was seen in the DBH category of 21–30 cm (Figure 7). However, the overall allometric model remains adequate for estimating energy storage in all species.
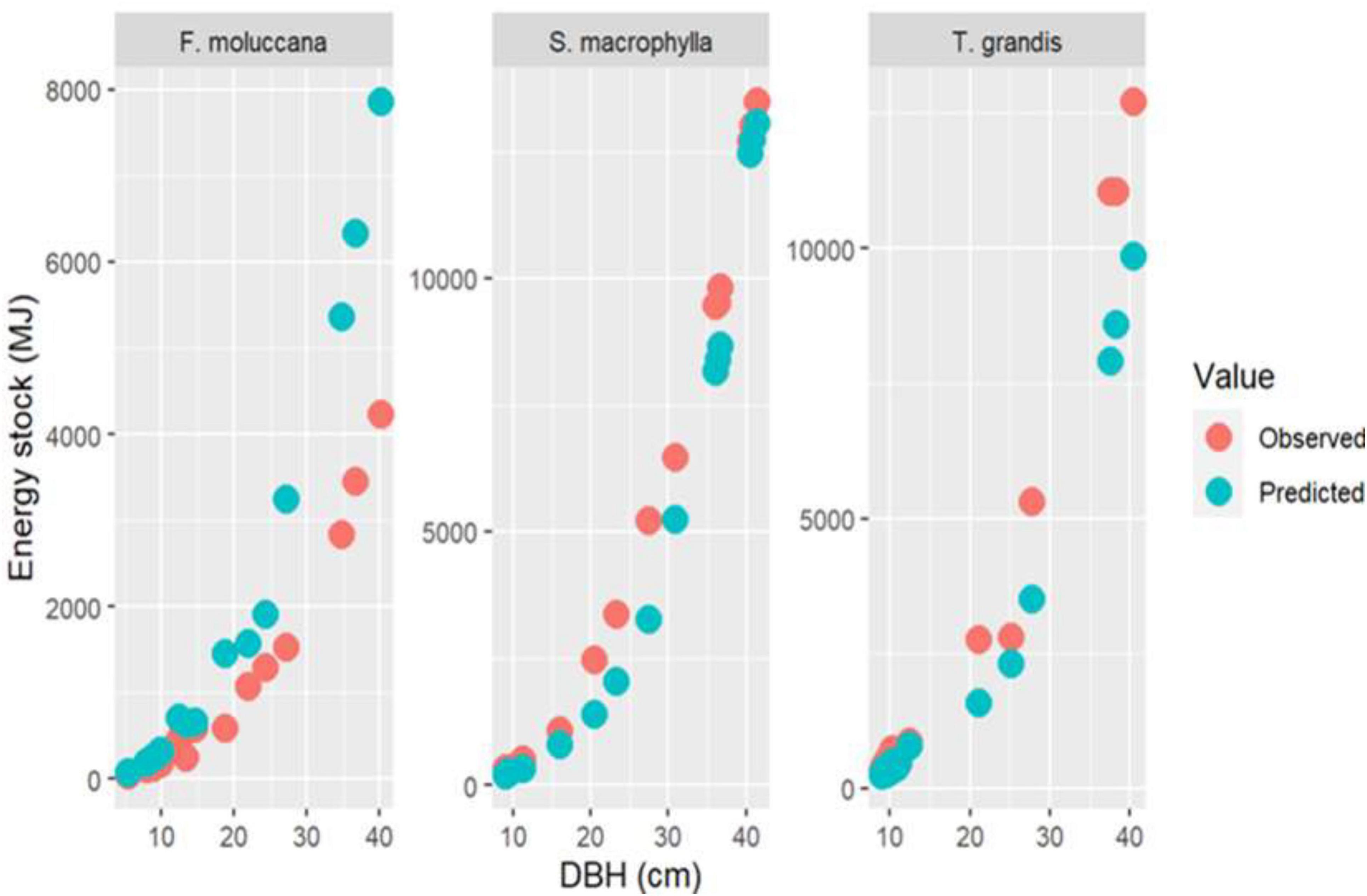
Figure 6. Comparison between observed and predicted energy stock in every species.
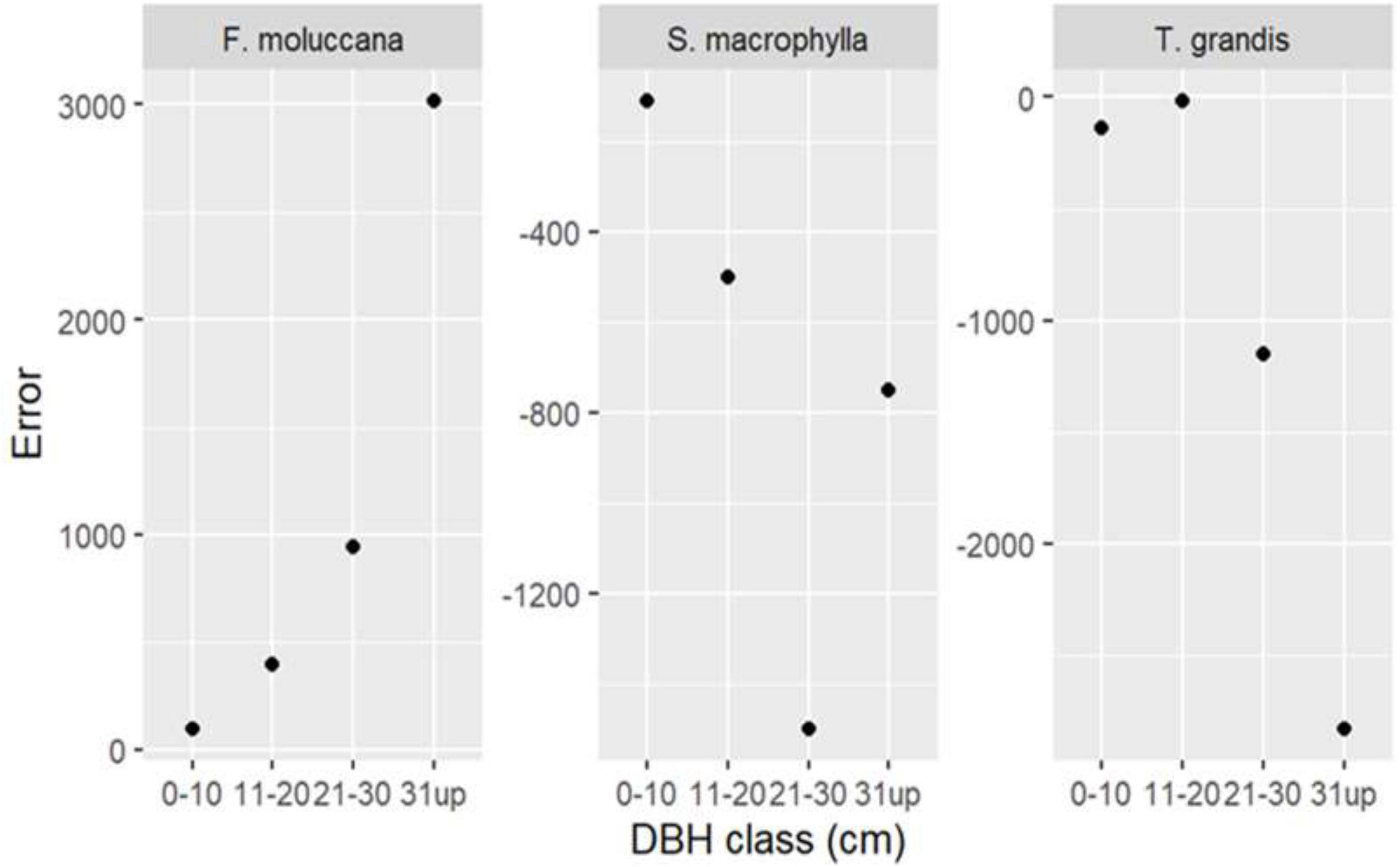
Figure 7. Trend of error in energy stock estimation from every species across DBH classes.
4 Discussion
4.1 Calorific value, energy distribution, and harvesting index
The key factor contributing to the greater energy content of foliage compared to stems and branches is outlined in Table 2. Trees typically harness solar energy through the process of photosynthesis, with foliage playing a crucial part in controlling this function. It regulates the transfer of energy between trees and the atmosphere (de Mattos et al., 2020). The energy is stored by foliage through the process of photosynthesis (Ellison et al., 2017). Furthermore, the size of the leaf’s surface area has an impact on the quantity of solar radiation that is absorbed by the foliage (Lee et al., 2019). Trees possessing greater leaf surface areas will assimilate a bigger amount of solar radiation compared to those with smaller leaf surface areas. Due to its larger leaf size compared to S. macrophylla and F. moluccana, T. grandis possessed a higher caloric content.
The fact that the foliage of the three species had a higher calorific value than the other components suggests that the abundant species’ leaves available as waste could be used for co-firing biomass. Further, because community people commonly collect teak leaves from the ground during the dry season for traditional food packages, they can be used as a biomass replacement for co-firing. So, once appropriate plantations of faster growing species for co-firing wood biomass are established, the species remain beneficial, especially when the trees are returned to their original use as a supply of lumber. Given that T. grandis had a higher calorific value than F. moluccana and S. macrophylla for all components of the tree, this important information reinforces the importance of planting this easily planted teak tree in any less fertile soil between unused community lands. This is especially helpful since teak can be coppiced for harvesting without having to replant.
Trees, along with other autotrophic organisms, have the natural capacity to transform solar energy into biomass through photosynthesis (Lee et al., 2019; de Mattos et al., 2020). This process contributes to the net primary productivity. Thus, an increased quantity of tree biomass indicates a better capacity for energy storage. The energy distribution pattern inside a single tree has resemblances to biomass allocation, with the stem containing the highest proportion of leaves and branches. This phenomenon can occur because the stem serves a vital function in maintaining the stability of trees and facilitating the process of translocation (Wirabuana et al., 2020). Therefore, it can be deduced that the stem has a larger store of energy in comparison to other components. Pruning and leaf abscission can also lead to a reduction in the formation of energy reserves in the branches and leaves (Suchocka et al., 2021). These occurrences occur at regular intervals over an organism’s existence and result in a decrease in the total amount of living matter. Although the stem of all species contains the most energy, for co-firing wood biomass energy that does not require a beautiful shape, the ability to be harvested from the coppice on a regular basis is required, which T. grandis and F. moluccana possess that coppicing ability but S. macrophylla does not. Meanwhile, F. moluccana, which grows relatively quickly, may be favored over S. macrophylla, which has the lowest calorific value of all the species’ components (Table 2).
The correlation between energy levels and tree age is evident through the growth of energy reserves in all species and the enhancement of DBH class. The DBH normally rises over time due to the gradual growth of biomass (Wirabuana et al., 2022). Trees having a larger DBH typically indicate a more advanced level of maturity in comparison to trees with smaller DBH measurements. Prior studies have also recorded comparable results, suggesting that a larger DBH is a reliable indicator of a more advanced age of the tree (Kirongo et al., 2012; Sumida et al., 2013). With these findings, an assessment of the diameter and allometric models for coppice, particularly for T. grandis and F. moluccana, is required, especially when both species are included as selected species for the next specific co-firing plantations.
The energy harvesting index in T. grandis demonstrated a superior level in comparison to other species, as illustrated in Figure 4. As stated by reference Gutiérrez-Soto et al. (2008), the plant consistently sheds its leaves during the dry season as a tactic to minimize transpiration. At this point, the focus will be on the allocation of energy toward the woody constituents. T. grandis had a less dense canopy compared to F. moluccana and S. macrophylla. As a result, this species shows a greater distribution of biomass toward its stem and branches to ensure its survival. The reason for exclusively harvesting stems and branches for renewable energy materials was confirmed.
4.2 Reliability of allometric models
DBH and H are frequently employed as the independent variables for constructing allometric models. In addition to their ease of measurement in the field, the variables are strongly correlated with the plant development mechanism, specifically photosynthesis (Zhang et al., 2016). The increase in DBH and H is a direct consequence of greater biomass storage in the stems (Köhl et al., 2017). Hence, using allometric models that incorporate both variables ensures a high degree of accuracy in estimating the energy store of each tree component as well as the overall total for each species (R2adj = 0.88–0.99; Table 4). Therefore, this is in line with the first hypothesis that the allometric models can accurately estimate the energy stock of those species in a community forest. It also confirmed that the best models are expected to predict more than 70% of the energy stock variation in each tree component and the total energy stock of each tree. Furthermore, the unique patterns of allometric models for each species were confirmed.
Furthermore, other research investigations also utilize these factors of DBH and H as independent variables when creating allometric models for other tree parameters, including biomass, carbon storage, and crown features (Chave et al., 2005; Zahabu et al., 2018; Pretzsch, 2019; Shi et al., 2019). For species in this study, adding H only showed a significant role in improving the model’s reliability for estimating the energy stock in the stem of S. macrophylla and T. grandis. Meanwhile, this variable of H was essential to increase the reliability of general allometric models for estimating the total energy stock. Therefore, the best-fit models for every component in each species were relatively different, which is in accordance with our second hypothesis.
While the allometric models show satisfactory reliability, it is important to consider many factors before employing this model. For instance, the utilization of the species-specific model in forest inventory may yield the most precise estimation, but at the cost of increased time consumption. The technician should modify the equation whenever they encounter distinct species. However, utilizing allometric models with two predictors is likewise wasteful in terms of time and cost, albeit providing improved precision. When gathering data, measuring tree height presents a greater level of difficulty compared to measuring tree diameter due to the limitation imposed by canopy overlap between trees, which can potentially introduce errors (Magalhães, 2017; Dey et al., 2021; Kafuti et al., 2022). Hence, employing a universal model with a solitary predictor (DBH) is more advantageous for facilitating the estimation of energy stock in tree species within the community forest in the study site. This predictor can provide accurate estimation approximately more than 80% (Table 4). Prior research has also demonstrated that a straightforward using DBH as a solitary predictor for facilitating forest inventory is more effective when it is used to estimated energy stock for many species in large area (Lumbres et al., 2015; Forrester et al., 2017; Romero et al., 2020).
Our observations indicate that the overall model, using DBH as the sole predictor, has demonstrated sufficient dependability. Therefore, we can confidently suggest it as a tool for landowners or local governments to assess the energy stock of tree species in community forests. Therefore, using a general model with a single predictor (DBH) is better to facilitate the energy stock estimation supports of community forest tree species in the study site. This also support our third hypothesis, that the general allometric models can be developed for all species, using DBH as the sole predictor both in every component and for the total individual tree.
Furthermore, it is crucial to recognize the constraints of our research, which encompass the utilization of a rather small sample size including only 45 trees, representing merely three unique species. The investigation was constrained to a singular location, hence precluding the assessment of the equation’s dependability across several sites and forest ecosystems. Further investigation is required to validate the reliability of allometric equations in accurately estimating energy reserves, especially when applied to a broader array of species and bigger sample numbers. Moreover, it is preferable that future research endeavors to include many sites to evaluate the overall validity of this approach.
4.3 The next strategy
The current local government is discussing about constructing a specific biomass plantation for co-firing raw materials. To reduce the setup costs, it is worthwhile to investigate species that yield high-quality biomass for energy while also producing coppices without the need to plant new trees. T. grandis and F. moluccana are two species that may be included, as well as many other fast-growing species that have been genetically improved for high-volume and quality wood for energy (Hendrati and Nurrohmah, 2018; Hendrati et al., 2022). F. moluccana has also been genetically improved for high biomass and gall tolerance, making it an appealing alternative for inclusion (Baskorowati et al., 2017). These species are legumes, and they can not only be harvested annually from close-spacing plantations, but they also improve soil and land quality and prevent erosion. T. grandis with a MAI (Mean Annual Increment) of 6.9 m3ha–1year–1 and S. macrophylla 19.71 m3ha–1year–1 (Hiratsuka et al., 2005; Krisnawati et al., 2011), which were used in this study, may produce much less wood biomass for co-firing, compared to the above-mentioned legume trees with a range MAI of 43–65 m3ha–1year–1. In 2021, the approximate requirement of biomass for generating electricity was estimated to be around 570,000 metric tons (Economic Research Institute for ASEAN and East Asia, 2022), while there are 19,516 ha of community forests in Jepara. Of these, half can still be planted with trees for timber for fulfilling the community needs of lumber, and the other half, or about 10,000 ha, can produce 430,000 m3ha–1year–1 (258,000 metric tons) of wood biomass when genetically improved legume species are grown for co-firing plantations. With a year of rotation, this only area in Jepara is estimated to be adequate to meet 25% of the requirement wood biomass for co-firing.
5 Conclusion
The findings of our study indicate that the allometric models demonstrated a high level of reliability in facilitating the estimation of energy stock for 3 alternative trees’ components within the community forest tree species. This is also applicable as well as for the entire individual tree. Allometric models also have the potential to estimate about eighty percent of the variation in energy stocks. Nevertheless, there were notable variations in the best-fit model for each species, as the inclusion of tree height as a predictor variable did not consistently enhance the dependability of allometric models. Furthermore, the application of generalized allometric models can further enhance the assessment of energy stocks for all species under observation. Later, this must be contrasted to the faster-growing species that will be expressly selected for plantations to provide co-firing wood biomass, notably from the coppice mode of harvesting. Additionally, we suggest conducting additional research to assess the dependability of allometric models in calculating energy stock across various species and site conditions. We suggest that in Jepara district, half of the community forest still can be used for fulfilling the community needs of lumber, and the other half, can produce of wood biomass for energy by coppice system using genetically improved legume species as co-firing plantations.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
PW: Conceptualization, Data curation, Formal analysis, Methodology, Software, Writing – original draft, Writing – review & editing. BM: Funding acquisition, Project administration, Software, Writing – original draft, Writing – review & editing. HB: Funding acquisition, Writing – original draft, Writing – review & editing. RH: Data curation, Methodology, Validation, Writing – original draft, Writing – review & editing. IN: Data curation, Methodology, Validation, Writing – original draft, Writing – review & editing. Mashudi: Data curation, Formal analysis, Investigation, Resources, Writing – original draft. DS: Data curation, Formal analysis, Investigation, Resources, Writing – original draft. SP: Data curation, Formal analysis, Investigation, Resources, Writing – original draft. Sumardi: Data curation, Investigation, Resources, Visualization, Writing – original draft, Writing – review & editing. LB: Conceptualization, Data curation, Methodology, Resources, Supervision, Writing – original draft, Writing – review & editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This study was financially supported by the Tropical Forest Mensuration and Silviculture Technology (TROFSIT) through the community forests research fellowship by grant number 05/24/TRF/P-JPR/CF-08 for data collection and article processing charge was also supported by the Center for International Forestry Research and World Agroforestry (CIFOR-ICRAF).
Acknowledgments
We express our gratitude to the Government of Srobyong Village, who allowed us to conduct this study. We also thank the organization of Tropical Forest Mensuration and Silviculture Technology (TROFSIT), which provides financial funds to harvest tree samples and laboratory analysis, and to the Center for International Forestry Research and World Agroforestry (CIFOR-ICRAF), for publication fee. High appreciation is also addressed to the Surveyor Team, who help the data collection process. We also thank two reviewers for their suggestions to improve manuscript quality.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Altanzagas, B., Luo, Y., Altansukh, B., Dorjsuren, C., Fang, J., and Hu, H. (2019). Allometric equations for estimating the above-ground biomass of five forest tree species in Khangai, Mongolia. Forests 10:661. doi: 10.3390/f10080661
Baskorowati, L., Bush, D., Setiadi, D., and Susanto, M. (2017). Genetic variation of growth and disease resistance traits in open-pollinated provenance-progeny trials of Falcataria moluccana growing on two rust-affected sites at age-18 months. J. Trop. For. Manag. 23, 1–7. doi: 10.7226/jtfm.23.1.1
Boedhihartono, A. (2017). Can community forests be compatible with biodiversity conservation in indonesia? Land 6:21. doi: 10.3390/land6010021
Central Java Provincial Statistics Bureau (2022). Land use and forest area by regency/city in central java province. Statistical report (Indonesia). Available online at: https://jateng.bps.go.id/statictable/2022/03/15/2547/luas-penggunaan-lahan-dan-luas-kawasan-hutan-menurut-kabupaten-kota-di-provinsi-jawa-tengah-km2-2021.html (accessed October 20, 2023).
Chave, J., Andalo, C., Brown, S., Cairns, M., Chambers, J., Eamus, D., et al. (2005). Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145, 87–99. doi: 10.1007/s00442-005-0100-x
Chave, J., Réjou-Méchain, M., Búrquez, A., Chidumayo, E., Colgan, M. S., Delitti, W. B. C., et al. (2014). Improved allometric models to estimate the aboveground biomass of tropical trees. Glob Chang. Biol. 20, 3177–3190. doi: 10.1111/gcb.12629
Coombes, A., Martin, J., and Slater, D. (2019). Defining the allometry of stem and crown diameter of urban trees. Urban For. Urban Green. 44:126421. doi: 10.1016/j.ufug.2019.126421
Da Silva, D. A., Eloy, E., Caron, B. O., and Trugilho, P. F. (2019). Elemental chemical composition of forest biomass at different ages for energy purposes. Flor. Ambient. 26:4. doi: 10.1590/2179-8087.020116
de Mattos, E., Binkley, D., Campoe, O., Alvares, C., and Stape, J. (2020). Variation in canopy structure, leaf area, light interception, and light use efficiency among Eucalyptus clones. For. Ecol. Manag. 463:118038. doi: 10.1016/j.foreco.2020.118038
Dey, T., Ahmed, S., and Islam, M. (2021). Relationships of tree height-diameter at breast height (DBH) and crown diameter-DBH of Acacia auriculiformis plantation. Asian J. For. 5:203. doi: 10.13057/asianjfor/r050203
Economic Research Institute for ASEAN and East Asia (2022). Forecast of biomass demand potential in Indonesia: Seeking a business model for wood pellets. ERIA research project report 2022 No. 01. Jakarta: Economic Research Institute for ASEAN and East Asia.
Ellison, D., Morris, C., Locatelli, B., Sheil, D., Cohen, J., Murdiyarso, D., et al. (2017). Trees, forests, and water: Cool insights for a hot world. Glob. Environ. Change 43, 51–61. doi: 10.1016/j.gloenvcha.2017.01.002
Favero, A., Daigneault, A., Sohngen, B., and Baker, J. (2023). A system-wide assessment of forest biomass production, markets, and carbon. GCB Bioenergy 15, 154–165. doi: 10.1111/gcbb.13013
Forrester, D., Tachauer, I., Annighoefer, P., Barbeito, I., Pretzsch, H., Ruiz-Peinado, R., et al. (2017). Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 396, 160–175. doi: 10.1016/j.foreco.2017.04.011
Gutiérrez-Soto, M. V., Pacheco, A., and Holbrook, N. (2008). Leaf age and the timing of leaf abscission in two tropical dry forest trees. Trees 22, 393–401. doi: 10.1007/s00468-007-0200-9
Hector, A., Felten, S., and Schmid, B. (2016). Analysis of variance with unbalanced data: An update for ecology & evolution. J. Anim. Ecol. 79, 308–316. doi: 10.1111/j.1365-2656.2009.01634.x
Hendrati, R., and Nurrohmah, S. (2018). Quality of genetically-improved Acacia auriculiformis for renewable short-rotation wood-energy. J. Trop For. Manag. 3, 136–143. doi: 10.7226/jtfm.24.3.136
Hendrati, R., Hidayati, N., Hardiyanto, E., Susanto, M., Baskorowati, L., Mashudi, M., et al. (2022). Genetic parameters of growth and biomass in Leucaena leucocephala for wood energy. Trop. Grasslands Forrajes Trop. 31:10. doi: 10.17138/tgft(10)15-21
Hiratsuka, M., Chingchai, V., Kantinan, P., Sirirat, J., Sato, A., Nakayama, Y., et al. (2005). Tree biomass and soil carbon in 17- and 22-year-old stands of teak (Tectona grandis L.f.) in northern Thailand. Tropics 4, 377–382.
Kafuti, C., Van den Bulcke, J., Beeckman, H., Van Acker, J., Hubau, W., De Mil, T., et al. (2022). Height-diameter allometric equations of an emergent tree species from the Congo Basin. For .Ecol. Manag. 504:119822. doi: 10.1016/j.foreco.2021.119822
Khan, M., Shil, M., Azad, M., Sadath, M., Feroz, S., and Mollick, A. (2018). Allometric relationships of stem volume and stand level carbon stocks at varying stand density in Swietenia macrophylla King plantations. Bangladesh. For. EC 430, 639–648. doi: 10.1016/j.foreco.2018.09.002
Kirongo, B., Mbelase, A., Senelwa, K., Hitimana, J., and Etiegni, L. (2012). Spacing and genotype on height and diameter growth of four eucalyptus under short rotation. J. Trop For. Manag. 18, 1–9. doi: 10.7226/jtfm.18.1.1
Köhl, M., Neupane, P., and Lotfiomran, N. (2017). The impact of tree age on biomass growth and carbon accumulation capacity: A retrospective analysis using tree ring data of three tropical tree species grown in natural forests of Suriname. PLoS One 12:e0181187. doi: 10.1371/journal.pone.0181187
Kora, S., Guendehou, G., Goussanou, C., Assogbadjo, A., and Sinsin, B. (2019). Allometric equations from a non-destructive approach for biomass prediction in natural forest and plantation in West Africa. South. For. J. For Sci. 81, 111–122. doi: 10.2989/20702620.2018.1512795
Krisnawati, H., Varis, E., Kallio, M., and Kanninen, M. (2011). Swietenia macrophylla: Ecology, silviculture, and productivity. Bogor: CIFOR.
Lee, H., Park, J., Cho, S., Lee, M., and Kim, H. (2019). Impact of leaf area index from various sources on estimating gross primary production in temperate forests using the JULES land surface model. Agric. For. Meteorol. 27:107614. doi: 10.1016/j.agrformet.2019.107614
Lisboa, S., Guedes, B., Ribeiro, N., and Sitoe, A. (2018). Biomass allometric equation and expansion factor for a mountain moist evergreen forest in Mozambique. Carbon. Balance Manag. 13:23. doi: 10.1186/s13021-018-0111-7
Lumbres, R., Lee, Y., Yun, C., Koo, C., Kim, S., and Bin, et al. (2015). DBH-height modeling and validation for Acacia mangium and Eucalyptus pellita in Korintiga Hutani Plantation, Kalimantan, Indonesia. Forest Sci. Technol. 11, 119–125. doi: 10.1080/21580103.2014.957356
Magalhães, T. (2017). Site-specific height-diameter and stem volume equations for Lebombo-ironwood. Ann. For. Res. 60:2. doi: 10.15287/afr.2017.838
Ministry of Energy and Mineral Resources (2020). Apply co-firing method in PLTU, the potential of biomass to substitute coal. Article (Indonesia). Available online at: https://ebtke.esdm.go.id/post/2020/02/28/2490/terapkan.metode.co-firing.di.pltu.ini.potensi.biomassa.untuk.subtitusi.batubara (accessed October 20, 2023).
Ministry of Energy and Mineral Resources, Republic of Indonesia. (2024). Capaian Program Cofiring EBTKE di 43 PLTU, Kurangi Emisi Karbon 1,1 Juta Ton. Available online at: https://www.esdm.go.id/id/media-center/arsip-berita/capaian-program-cofiring-ebtke-di-43-pltu-kurangi-emisi-karbon-11-juta-ton
Mulyana, B., Soeprijadi, D., and Purwanto, R. (2020). Allometric model of wood biomass and carbon for Gliricidia (Gliricidia sepium (Jacq.) Kunth ex Walp.) at bioenergy plantation in Indonesia. For. Ideas 26, 153–164.
Ogana, F., and Ercanli, I. (2022). Modelling height-diameter relationships in complex tropical rain forest ecosystems using deep learning algorithm. J For Res. 33, 883–898. doi: 10.1007/s11676-021-01373-1
Pretzsch, H. (2019). The effect of tree crown allometry on community dynamics in mixed-species stands versus monocultures. A review and perspectives for modeling and silvicultural regulation. Forests 10:810. doi: 10.3390/f10090810
Romero, F., Jacovine, L., Ribeiro, S., Torres, C., Silva, L., and da, et al. (2020). Allometric equations for volume, biomass, and carbon in commercial stems harvested in a managed forest in the southwestern amazon: A case study. Forests 11:874. doi: 10.3390/f11080874
Sadono, R., Wahyu, W., and Idris, F. (2021). Allometric equations for estimating aboveground biomass of Eucalyptus urophylla S.T. Blake in East Nusa Tenggara. J. Trop For. Manag. 27, 24–31. doi: 10.7226/jtfm.27.1.24
Sadono, R., Wardhana, W., Idris, F., and Wirabuana, P. (2022). Allometric equation for estimating energy production of Eucalyptus urophylla in dryland ecosystems at east nusa tenggara. J. Trop For. Manag. 28, 32–39. doi: 10.7226/jtfm.28.1.32
Setiahadi, R. (2017). How significant is the existence of forest community contribution in GHG emissions reduction? J. Eng. Appl. Sci. 12, 4826–4830.
Setiahadi, R. (2021). Comparison of individual tree aboveground biomass estimation in community forests using allometric equation and expansion factor in Magetan, East Java, Indonesia. Biodiversitas J. Biol. Divers. 22:936. doi: 10.13057/biodiv/d220936
Setiahadi, R., Sari, S., Maryudi, A., Kalmirah, J., and Baskorowati, L. (2020). Monitoring implementation impact of the EU-Indonesia’s VPA on SME livelihood. Int. J. For. Res. 2020, 1–9. doi: 10.1155/2020/4327802
Shi, P., Liu, M., Ratkowsky, D., Gielis, J., Su, J., Yu, X., et al. (2019). Leaf area–length allometry and its implications in leaf shape evolution. Trees 33, 1073–1085. doi: 10.1007/s00468-019-01843-4
Simetti, R., Bonduelle, G., Silva, D., da, Mayer, S., Souza, H., et al. (2018). Production of biomass and energy stock for five Eucalyptus species. Rev. Ciência Mad 9, 30–36. doi: 10.12953/2177-6830/rcm.v9n1p30-36
Stas, S., Rutishauser, E., Chave, J., Anten, N., and Laumonier, Y. (2017). Estimating the aboveground biomass in an old secondary forest on limestone in the Moluccas, Indonesia: Comparing locally developed versus existing allometric models. For. Ecol. Manag. 389, 27–34. doi: 10.1016/j.foreco.2016.12.010
Suchocka, M., Swoczyna, T., Kosno-Joñczy, J., and Kalaji, H. (2021). Impact of heavy pruning on development and photosynthesis of Tilia cordata Mill. trees. PLoS One 16:e0256465. doi: 10.1371/journal.pone.0256465
Sumida, A., Miyaura, T., and Torii, H. (2013). Relationships of tree height and diameter at breast height revisited: Analyses of stem growth using 20-year data of an even-aged Chamaecyparis obtusa stand. Tree Physiol. 33, 106–118. doi: 10.1093/treephys/tps127
Tetemke, B., Birhane, E., Rannestad, M., and Eid, T. (2019). Allometric models for predicting aboveground biomass of trees in the dry afromontane forests of Northern Ethiopia. Forests 10:1114. doi: 10.3390/f10121114
Vega-Nieva, D. J., Valero, E., Picos, J., and Jiménez, E. (2015). Modeling the above and belowground biomass of planted and coppiced Eucalyptus globulus stands in NW Spain. Ann. For. Sci. 72, 967–980. doi: 10.1007/s13595-015-0493-6
Vyas, D., Mehta, N., Dinakaran, J., and Krishnayya, N. (2010). Allometric equations for estimating leaf area index (LAI) of two important tropical species (Tectona grandis and Dendrocalamus strictus). J. For. Res. 21, 197–200. doi: 10.1007/s11676-010-0032-0
Wirabuana, P. Y., Hendrati, R. L., Baskorowati, L., Susanto, M., Mashudi, B. S., Sulistiadi, H., et al. (2022). Growth performance, biomass accumulation, and energy production in age series of clonal teak plantation. Forest Sci. Technol. 18, 2. doi: 10.1080/21580103.2022.2063952
Wirabuana, P., Setiahadi, R., Sadono, R., Lukito, M., and Martono, D. (2021). The influence of stand densityand species diversity into timber production and carbon stock in community forest. Indones J. For. Res. 8, 13–22. doi: 10.20886/ijfr.2021.8.1.13-22
Wirabuana, P., Setiahadi, R., Sadono, R., Lukito, M., Martono, D., and Matatula, J. (2020). Allometric equations for estimating biomass of community forest tree species in Madiun, Indonesia. Biodivers. J. Biol. Divers. 21:947. doi: 10.13057/biodiv/d210947
Wulandari, C., Bintoro, A., Rusita, R., Santoso, T., Duryat, D., Kaskoyo, H., et al. (2018). Community forestry adoption based on multipurpose tree species diversity towards to sustainable forest management in ICEF of University of Lampung, Indonesia. Biodivers. J. Biol. Divers. 19, 1102–1109. doi: 10.13057/biodiv/d190344
Youkhana, A., and Idol, T. (2011). Allometric models for predicting above- and belowground biomass of Leucaena-KX2 in a shaded coffee agroecosystem in Hawaii. Agrofor. Syst. 83, 331–345. doi: 10.1007/s10457-011-9403-6
Zahabu, E., Mugasha, W., Katani, J., Malimbwi, R., Mwangi, J., and Chamshama, S. (2018). “Allometric tree biomass and volume models for Tectona grandis plantations,” in Allometric tree biomass and volume models in Tanzania, 2nd Edn, Vol. 1, eds R. Malimbwi, T. Eid, and S. Chamshama (Dar es Salaam: E&D Vision Publishing).
Keywords: biomass, calorific value, allometric model, renewable energy, community forest
Citation: Wirabuana YAPP, Mulyana B, Baral H, Hendrati LR, Nurtjahjaningsih ILG, Mashudi, Setiadi D, Pudjiono S, Sumardi and Baskorowati L (2024) Energy storage of Indonesian community forest tree species employing allometry equations for designing the next strategies. Front. For. Glob. Change 7:1373683. doi: 10.3389/ffgc.2024.1373683
Received: 20 January 2024; Accepted: 08 August 2024;
Published: 29 August 2024.
Edited by:
Erkki Tomppo, University of Helsinki, FinlandReviewed by:
Iulian Alexandru Bratu, Lucian Blaga University of Sibiu, RomaniaAzra Čabaravdić, University of Sarajevo, Bosnia and Herzegovina
Copyright © 2024 Wirabuana, Mulyana, Baral, Hendrati, Nurtjahjaningsih, Mashudi, Setiadi, Pudjiono, Sumardi and Baskorowati. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liliana Baskorowati, bGlsaWFuYS5iYXNrb3Jvd2F0aUBicmluLmdvLmlk