 Haoze Yu†
Haoze Yu† Hui Zhang
Hui Zhang- Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, School of Tropical Agriculture and Forest, Hainan University, Haikou, China
Introduction: Asian tropical rainforests have the highest rates of degradation in the world. Consequently, a large decline in Chinese Vatica mangachapoi (a keystone species) had led to its listing in the category of “vulnerable” species by IUCN. However, its current status after decades of conservation efforts remains unknown.
Methods: Here, we evaluate the current status of Chinese V. mangachapoi.
Results and Discussion: We found that its population is now dispersed in 14 protected areas, the largest being a coastal forest that contains 96.84% of all the Chinese V. mangachapoi. Compared to their historic records, the age of this forest was estimated at ≤ 70 years. The mono-culturing of V. mangachapoi in this forest, since 1960, has replaced all the older trees, resulting in its extremely high (91%) relative abundance, and an extensively low (only 44) tree species richness. Further, these V. mangachapoi trees now suffer from vine strangulations and severe Amauroderma perplexum infections: 18.5% of V. mangachapoi have died and 75% are at a high risk, thereby creating a threat of its extinction. Although, the other 13 protected areas have a higher tree species richness (152–451), a lower (6.1–25%) relative abundance of V. mangachapoi, and they neither suffer from vine strangulation or disease infections, they contribute to only 3.16% of total Chinese population of this species. Therefore, an immediate revision of threat status of this species in IUCN, from vulnerable to endangered, is warranted. Further, a change in planting patterns, from monocultures to mix-plantations of native species, is needed to promote biodiversity and restrict other biotic challenges so that this species is not extinct.
Introduction
Tropical rainforests are distributed in Asia, New Guinea, Africa, Madagascar, and the Neotropics. Asian tropical rainforests are uniquely dominated by trees from the family Dipterocarpaceae (Corlett, 2007), where Vatica mangachapoi is a keystone species for tropical rainforest in Hainan Island, China (Ding et al., 2006). This species accounts for 20–50% of forest basal area and > 50% of the tree canopy cover (Corlett and Primack, 2006; Corlett, 2011; Ghazoul, 2016). It is native to Thailand, Brunei, Malaysia, Philippines, Indonesia, and the Hainan Island of China (Supplementary Figure 1). These trees can grow up to 20 m tall, and mostly grow on hill- and mountain-slopes below an altitude of 700 meters (De Guzman et al., 1986). It has two recognized subspecies, V. mangachapoi subsp. obtusifolia (found only in Philippines and Malaysia) and V. mangachapoi subsp. mangachapoi (found in rest of the world, except in China), which grows in deep soils, mixed peat swamp forests, hills and coastal areas (Soepadmo et al., 2004).
Vatica mangachapoi had been commonly found in its natural habitats, but in the last three decades (or any specific year), its population has been declining substantially (Pooma et al., 2017). For instance, in Hainan Island, its population has had a reduction of 35% between 1991 and 2008, something which is of major concern for the conservation of this tree species (Zhang et al., 2010). Due to such large decline in Chinese V. mangachapoi has led to its listing as “vulnerable” in the international union for conservation of nature (IUCN) red list of threatened species (Pooma et al., 2017). After such an IUCN assessment, constant monitoring of V. mangachapoi’s population decline was warranted (Pooma et al., 2017). Yet, an accurate and up-to-date scientific report on the conditions (e.g., fitness and health) in which it is found is lacking. We have therefore addressed this challenge in the current investigation by surveying all places where V. mangachapoi population exists in the Hainan Island, southeast China (Supplementary Figure 1).
Vatica mangachapoi-mixed forests have dominated the old-growth tropical evergreen monsoon rainforests on Hainan Island of China (Ding et al., 2006; Dai et al., 2018). However, the conversion of these natural mixed forests into rubber plantations is the main driver of large decline in V. mangachapoi population (Zhang et al., 2010; Pooma et al., 2017; Guo et al., 2018), which are now dispersed across the whole islands. Therefore, we first surveyed its population density (%) in all the V. mangachapoi pockets to determine its relative abundance and spatial distribution patterns. Moreover, the conversion of tropical lowland rainforest into rubber plantation has resulted in a large decrease in tree diversity (Ahrends et al., 2015; Warren-Thomas et al., 2020). Replanting of mono-cultured V. mangachapoi in these locations raises the threat of diseases and other biotic challenges. Validation of these possibilities require evaluation of tree diversity (tree species richness) and assessment of V. mangachapoi health. Additionally, When asking and hearing from local people, indicates that several protected areas have been planted with monocultures of seedlings of V. mangachapoi to increase its density in Hainan Island. This can be confirmed by tracing the land use history and the age of the oldest V. mangachapoi at various locations. For example, remote sensing (RS) and geographic information systems (GIS) are suitable tools for detecting and evaluating long-term changes in habitat cover and quality of threatened species management (Zou et al., 2022). Moreover, δ14C estimates can also help estimate tree age (Ichie et al., 2022).
In this investigation, we used the currently available remote sensing images of the highest resolution to trace the changes in historical land use. We also used δ14C estimation methods to measure the age of the oldest V. mangachapoi trees to determine whether they have been artificially planted or naturally regenerated.
Materials and methods
Study sites
The survey of Chinese V. mangachapoi population was performed across the entire Hainan Island where this tree species is known to be present. The survey of current outcome status of an unique coastal V. mangachapoi forest is conducted at the Provincial National Reserve of Liji costal V. mangachapoi forest (LJCV; 110°15′29.98″E, 18°39′43.14″N) in Xinglong town, Wanning City, Hainan Island, China. In China, Hainan Island has unique and high-diversity tropical lowland rainforest whose keystone species is V. mangachapoi (Guo et al., 2018). Its altitude ranges from 0 to 1,867 m. The sites has mean temperature of 22–27°C, with the lowest temperature (17°C) and the highest temperature (40°C) distributing in January and July. Average annual precipitation is ∼1,600 mm, with most of the precipitation occurring between May and October.
Survey of Chinese V. mangachapoi population density
Chinese V. mangachapoi population is now merely distributed in Hainan Island, China (Supplementary Figure 1). Thus, in 2020, we surveyed the respective V. mangachapoi individuals of all places where present V. mangachapoi individuals, so that we collected calculate their individual V. mangachapoi population density (%).
Remote sensing data collection and processing
The exact boundaries of the coastal forests were first obtained by performing field surveys with a handheld GPS. Then, we collected archived remote sensing images of the forests to clearly trace historical changes in its land use. The earliest remote sensing image (Landsat) could only be found for 1988. The tropical forest species composition is usually surveyed at an interval of every 5 years (Harms et al., 2001), we collected remote-sensing images for coastal V. mangachopoi forest from 1988 to 2020 at a regular interval of 5 years. For 1988, and 1993, only Landsat images were available; spot images were utilized for the other years. Details of all the remote sensing images, their preprocessing and supervised classification are shown in Supplementary Table 1.
Classification of remote-sensing images
Based on ArcGIS 10.2 platform, the maximum likelihood method of supervised classification was used to analyze land cover in the images (Phiri and Morgenroth, 2017). The study area of the coastal forest was classified as tomb, artificial construction and forest.
Survey of current status of the coastal forest
In the season of maximum growth of 2021 (August), in the costal forest, all freestanding trees with diameter of ≥ 1 cm at breast height (DBH) were measured and identified to species. We also recorded how many V. mangachapoi trees had vine damages, had any diseases, and were dead or healthy.
Measurements of age of the oldest V. mangachapoi
As DBH (and plant height/growth rates) could serve as an indicator of plant age, we hypothesized that the V. mangachapoi trees with largest DBH and plant height could be amongst the oldest trees in the costal forest. So, three xylem samples, which were closest to the roots, were collected from such oldest V. mangachapoi in the coastal forest. Specifically, xylem samples were collected by using an a standard 4.3-mm increment borer (Häglof Company Group, Långsele, Sweden) to extract the piece of wood from the stem near the root. These were used to estimate the age with the help of an accelerator mass spectrometry (AMS) technique (Hajdas et al., 2021).
Assessment of fungal disease of V. mangachapoi
In each of the 20 sampling plots (described above), we determined how many V. mangachapoi individuals (including both, dead and alive) had symptomatic leaves, stems or roots. Then, all the symptomatic plant parts were collected from 10 independent V. mangachapoi trees and were brought back to the laboratory on ice packs. Their DBH ranged between 1.1 and 42 cm that corresponded to trees of all sizes groups in the forest. Then, the symptomatic plant parts were washed in distilled water to remove dusts, sterilized by ethanol and NaOCl for three times (Zimmerman and Vitousek, 2012), and dried with sterile absorbent paper.
Symptomatic plant parts were first cut into small pieces and placed on potato dextrose agar (PDA; streptomycin: 40 μg ml–1, 20 g of potato starch, 4 g of dextrose, and 15 g of agar in 1 L), and incubated at 23°C under an 8 h light/16 h dark regime for 5 days in an artificial climate box (Jiangnan, Zhejiang, China). Fungal colonies were purified to individual isolates in order to separate the potential pathogenic strains (Zheng et al., 2021).
We extracted total genomic DNA for each of the fungal isolates with the help of Qiagen Plant DNeasy kits (Qiagen, Hilden, Germany). The extracted DNA was evaluated using gel electrophoresis (1% agarose gel). Then, the internal transcribed spacer 1 (ITS1) region was amplified by PCR using ITS1-F (CTT GGT CAT TTA GAG GAA GTAA) and ITS2 (GCT GCG TTC TTC ATC GAT GC) forward and reverse primers, respectively. A 30 μL PCR reaction mixture included the following: 15 μL of Phusion Master Mix (2×), 10 μL of DNA template, 3 μL of primer, and 2 μL of dd H2O. The PCR conditions were as following: a hot start at 98°C for 1 min; then 30 cycles of 98°C for 30 s, 55°C for 30 s and 72°C for 30 s; and a final extension at 72°C for 5 min (Zheng et al., 2021)
Results
We found that V. mangachapoi populations in China are dispersed across 23 protected areas (Figure 1A). Among them, a costal V. mangachapoi forest accounts for 96.84% of all of its existing population, whereas the remaining 22 protected areas account for merely 0.01–1.25% at each site (Figure 1A). This coastal forest has very low tree species richness (only 44), and an extremely high relative abundance (91%) of V. mangachapoi (Figure 1B). In contrast, the other 13 protected areas have much higher (152–451) tree species richness, and consequently, a much lower (6.1–25%) relative abundance of V. mangachapoi (Figure 1B).
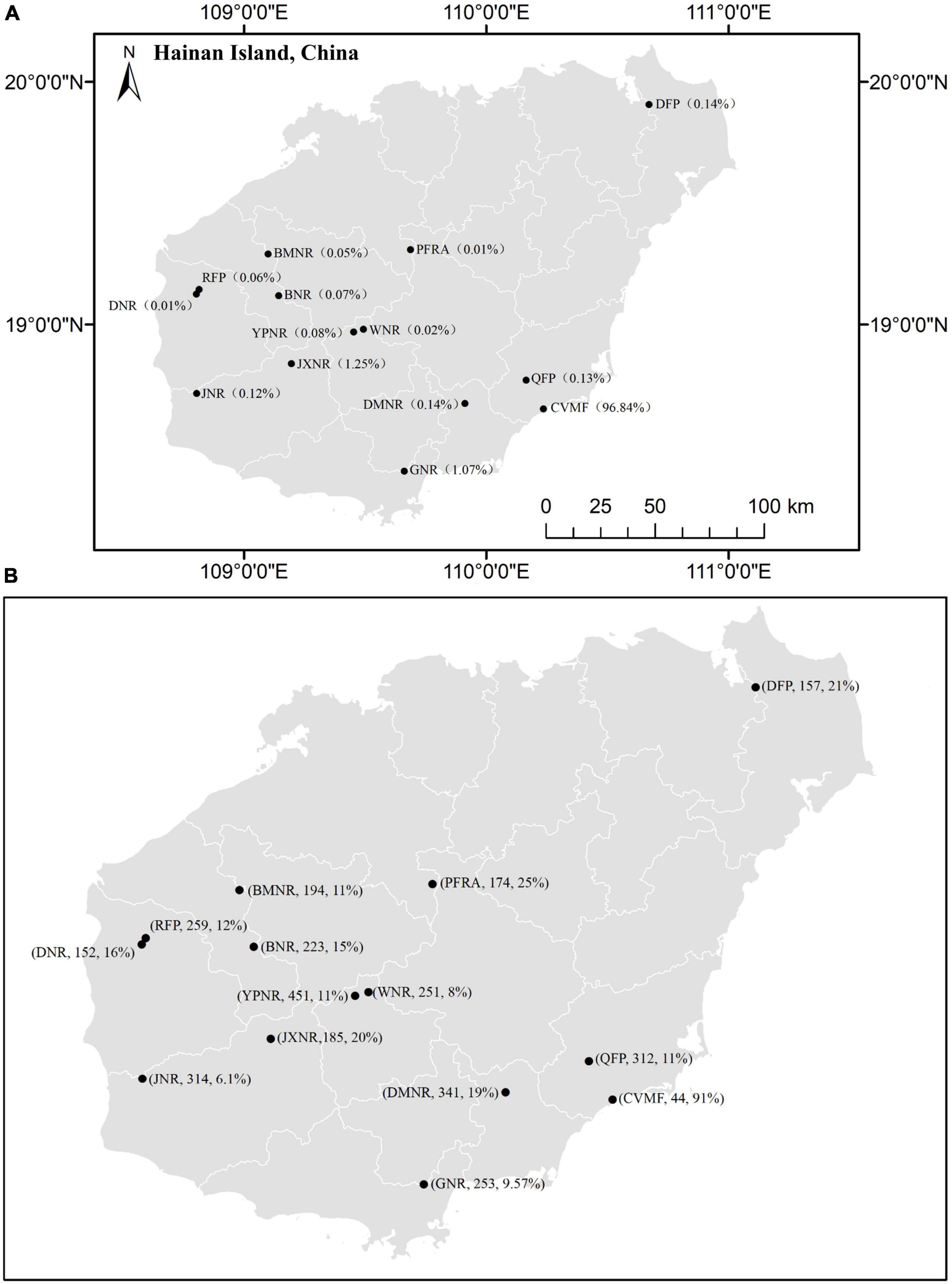
Figure 1. The locations of presently existed Chinese V. mangachapoi population and their respective V. mangachapoi population density (%) (A) and total tree species richness (numbers of species) and relative abundance (%) for V. mangachapoi population in each location (B). These locations are distributed in 14 protected areas in Hainan Island, China. They include the coastal V. mangachapoi forest (CVMF), Qiaoxiang Forest Park (QFP), Ganshiling Nature Reserve (GNR), Gouling Forest Park (GFP), Balong National Forest Park (BNFP), Linchun Ridge Forest Park (LRFP), Institute of Sanya Forestry Research (ISFR), Tropical Paradise Forest Park (TPFP), Bawangling Natural Reserve (BNR), Baomeiling Nature Reserve (BMNR), Hongling Nature Reserve (HNR), Jiaxi Natural Reserve (JXNR), Jianfengling Natural Reserve (JNR), Kafaling Forest Park (KFP), Jiangfengling Fork Estuary (JFE), Datian Nature Reserve (DNR), Rhesus Forest Park (RFP), Dacheng Forest Park (DFP), Diaolou Mountain Natioanl Natural Reserve (DMNR), Nanwan Natural Reserve (NNR), Panga Forest Reserve Area (PFRA), Yinggeling Provincial Natural Reserve (YPNR), and Pojianling Forest Park (PFP).
Our analysis of remote sensing data clearly revealed the artificially plantations of V. mangachapoi in this unique costal forest. From 1988 to 1993, the total area of this forest had decreased from 900,900 to 622,800 m2, but it quickly recovered to 1,097,500 m2 between 1993 and 1998 (Figure 2), whereas the total forest area in 2021 was estimated to be 1,016,039 m2 (Figure 2). Moreover, δ14C measurements of the age of the oldest V. mangachapoi trees, in the protected coastal forest, indicate that they could have been planted or regenerated only after 1950 (age of the oldest trees is around 70 years; Supplementary Table 1). Whereas, the age of the oldest V. mangachapoi trees in the other 22 protected area range from 160 to 460 years (Table 1).
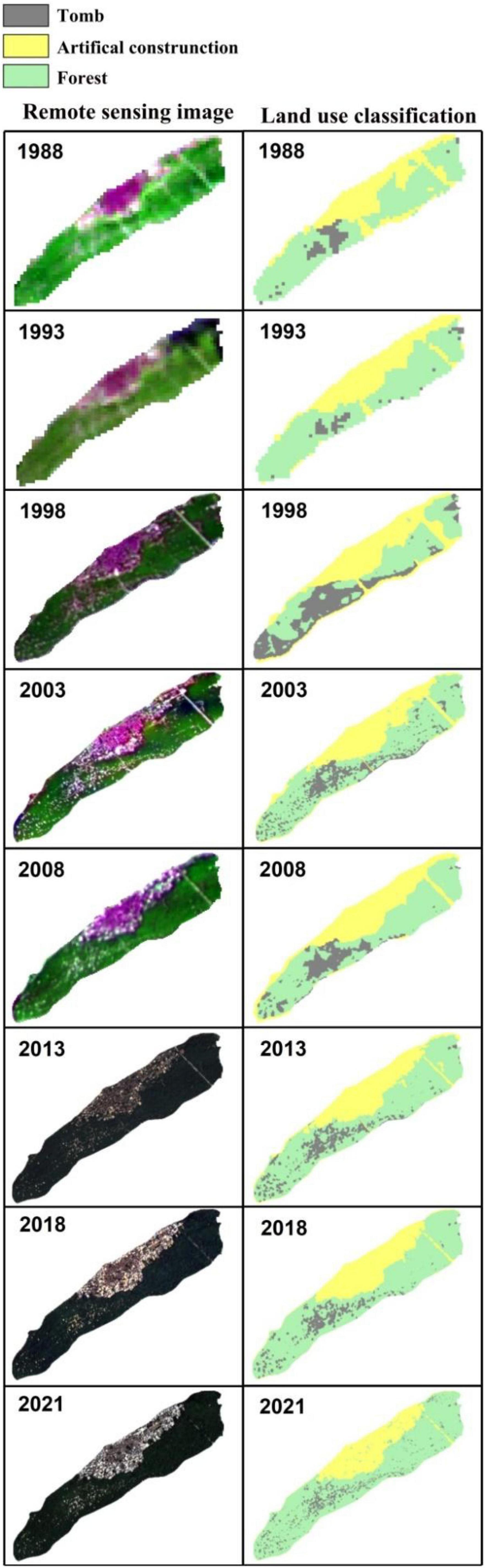
Figure 2. Remote sensing images for a detailed land cover analysis for the coastal V. mangachapoi forest during 1988–2020. The left panel shows the remote sensing images from 1988 to 2020. The right panel demonstrates the detailed land cover classification (tomb, artificial construction or forest) of the corresponding remote sensing images.
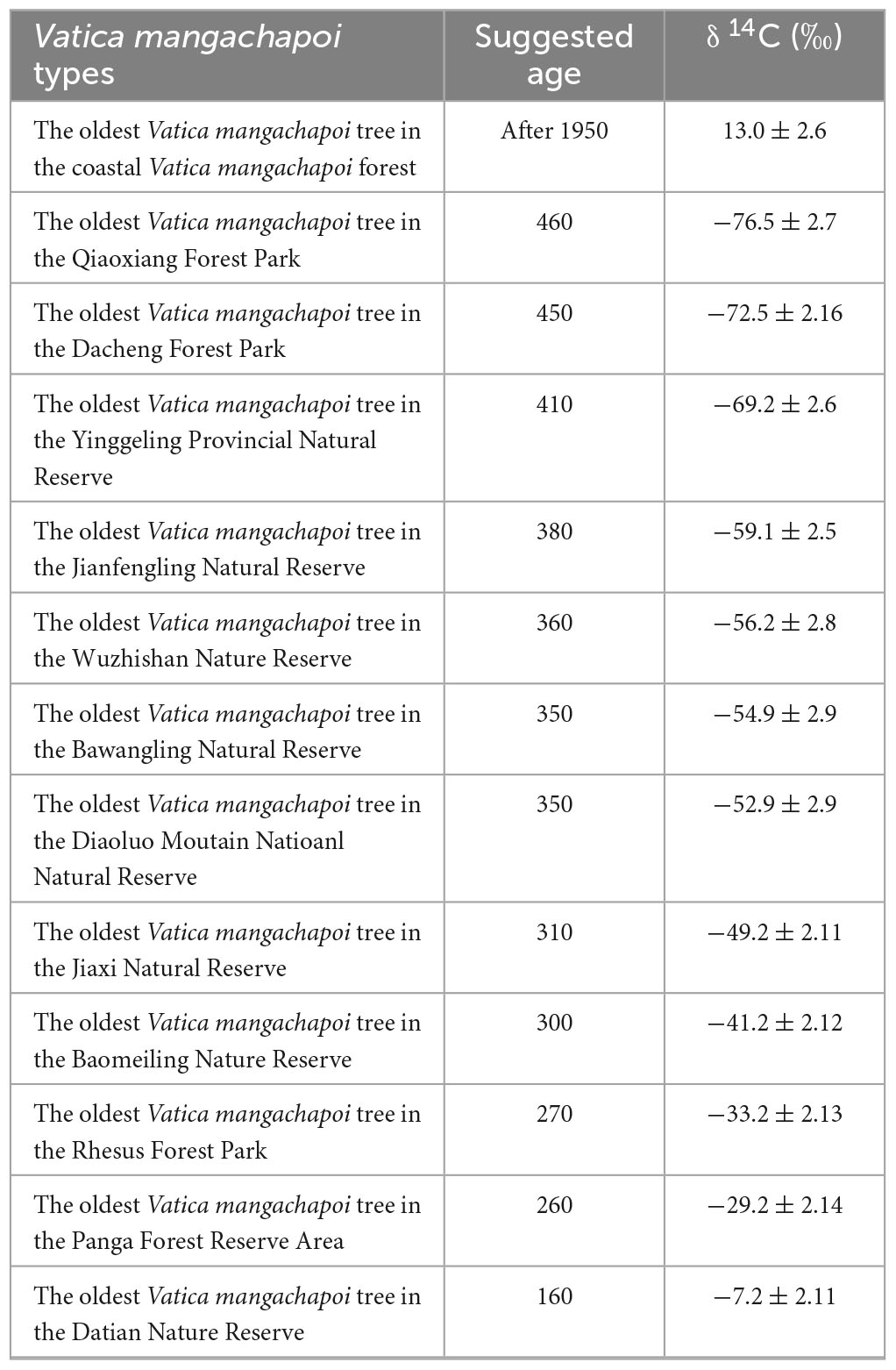
Table 1. δ14C values and suggested age for the oldest Vatica mangachapoi trees in all places where present Vatica mangachapoi in Hainan Island.
When investigating the occurrence of diseases, we found that 89.9% of V. mangachapoi in the coastal forests of Hainan have been infected by a fungal stem rot disease (Figures 3, 4), which is caused by the fungus Amauroderma perplexum (ITS sequencing; Supplementary Figures 2, 3). It is alarming that the 14.9% of V. mangachapoi trees have already died due to A. perplexum-induced stem rot (Figures 3, 4). An additional 75% of the infected V. mangachapoi trees are at an extreme risk of dying soon. Furthermore, 3.6% of V. mangachapoi have died due to vine strangulations (Figures 3, 4). On the other hand, V. mangachapoi in the other 13 protected areas did not suffer from Amauroderma infections and vine strangulation (Figure 3).
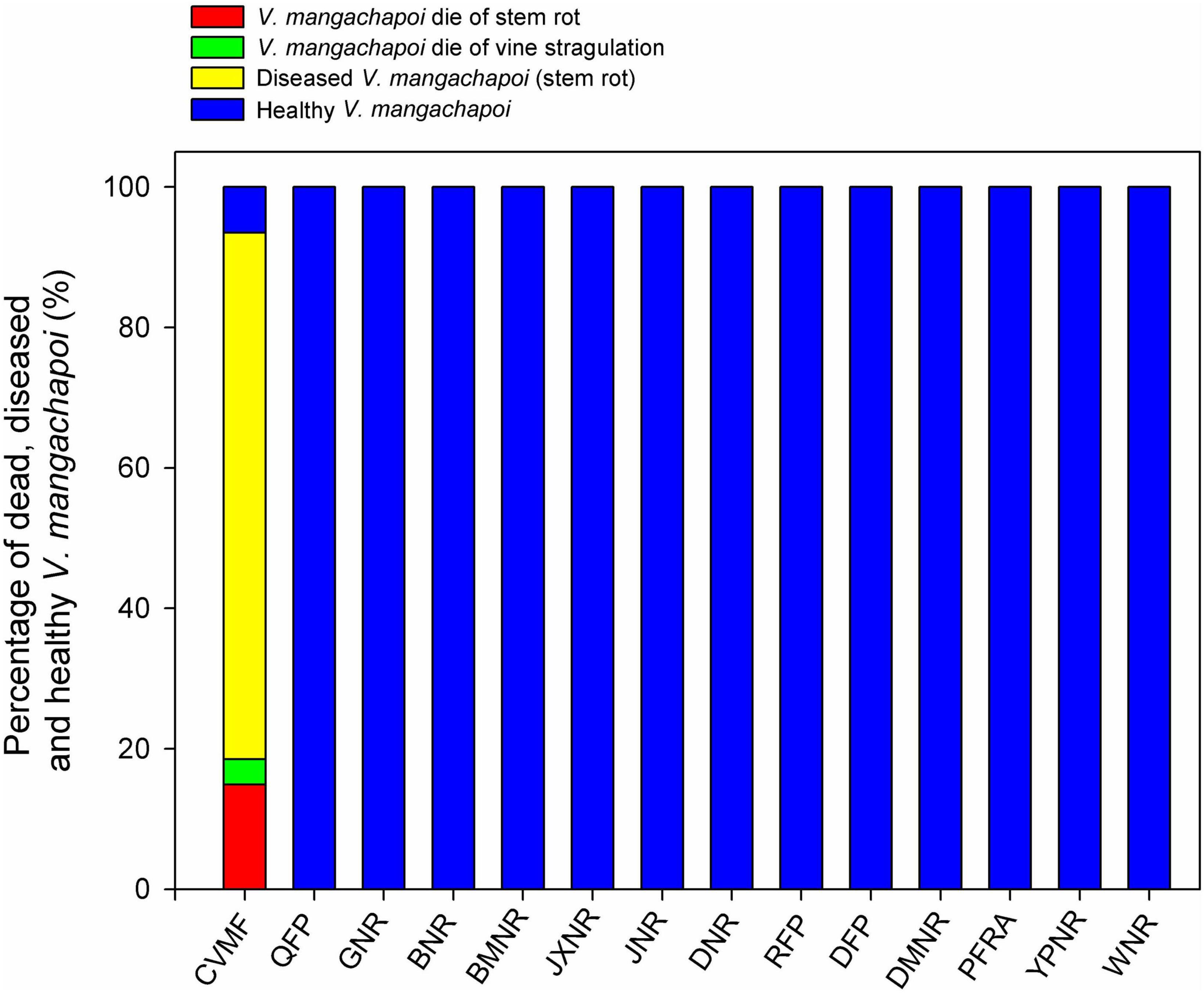
Figure 3. Composition of V. mangachapoi trees that are either infected (with stem rot), dead (due to stem rot or vine strangulation), or are healthy for Chinese V. mangachapi.

Figure 4. A fast-spreading stem rot disease of V. mangachapoi and vine strangulation in the coastal V. mangachapoi forest. (A) Few examples of dead and severely infected V. mangachapoi and stem rot symptoms (pathogen’s fruiting body in a live stem of V. mangachapoi). (B) Empirical proof of vine strangulation-induced death of V. mangachapoi.
Discussion
After the listing of Chinese V. mangachapoi in the IUCN red list of threatened species (Pooma et al., 2017), it is absolutely necessary to monitor the accurate status of its population. Our results provide the first evidence that V. mangachapoi is now sparingly distributed in 14 protected areas, of which a coastal forest in Hainan contains 96.84% of all of its Chinese populations. Therefore, this coastal forest becomes a unique site for conservation of V. mangachapoi as it would directly determine the threat that this species faces.
Unfortunately, the current situation of V. mangachapoi in this coastal forest is extremely bad. Since 1988, nearly half of this coastal forest has been destroyed by construction work. Further, the relative abundance of V. mangachapoi in this forest is now extremely high (90.8%), whereas plant diversity is very low. In contrast, the other 13 protected areas have much higher tree species richness (152–451), and lower (6.1–25%) relative abundance for V. mangachapoi (Figure 2). Tree communities with low diversity are at a high risk of epidemic diseases (Chen and Zhou, 2015; Liu et al., 2016, 2020), and of vine invasions (Livingstone et al., 2020; Carboni et al., 2021). Indeed, a total of 18.5% of V. mangachapoi trees in the coastal forests of Hainan have died due to vine strangulations or an infection of a fungal stem rot disease, which is caused by Amauroderma perplexum (Figures 3, 4 and Supplementary Figures 2, 3). Additional 75% of the V. mangachapoi trees are already infected by stem rot and are at an extreme risk of dying soon. On the other hand, V. mangachapoi the other 13 protected areas neither suffer from any vine strangulation nor any fungal diseases. It is evident that a low species diversity and an extremely high relative abundance of V. mangachapoi in this coastal forest are subjecting it to a threat of extinction.
Analysis of remote sensing data, δ14C measurements of the age of the oldest trees, and local provided insights into such a low tree diversity and an extremely high relative abundance of V. mangachapoi in the coastal forest. Since 1960, all old and mature V. mangachapoi trees of this forest have been cut down for building fishing boats and supporting the local steel industry. In Circular of the people’s Government of Hainan Province (1988), the government of the People’s Republic of China established a “provincial natural reserve” (PNR) for a high-level protection of this forest. Local surveys revealed that since 1993, PNR has created monocultures of the new V. mangachapoi saplings after cutting down other native tree species that originally coexisted with V. mangachapoi. This has transformed an original mixed-forest into a cultivated monoculture of V. mangachapoi in an agricultural setup. This was evident in our analysis of remote sensing data (1988–2021; Supplementary Figure 3). Further, δ14C measurements showed that the oldest V. mangachapoi tree in this forest, were planted/regenerated only after 1950 (age of the oldest trees is around 70 years; Supplementary Table 1). Whereas, the age of the oldest V. mangachapoi in the other 13 protected area range from 160 to 460 years (Supplementary Table 1).
Conclusion
Taken together, our results show that V. mangachapoi is under a threat of extinction in China. This warrants an immediate revision of the threat status of this species, from vulnerable to endangered or extremely endangered, in IUCN and in China. Moreover, the PNR needs a paradigm shift in their strategy to conserve this unique costal forest. This should involve measures to increase biodiversity by planting native species in the understory of the V. mangachapoi or at the sites where dead V. mangachapoi trees are standing. It would gradually restore the current monoculture plantations into a mix-V. mangachapoi forest, and would also enhance the ecosystem functioning and curtail biotic challenges. In addition, liana removal should also be performed to aid V. mangachapoi to grow better. Similar to China, large areas of tropical rainforest has been destroyed by rubber plantations in Thailand, Brunei, Malaysia, Philippines, and in Indonesia (Ahrends et al., 2015). Thus, an accurate survey of the current status of V. mangachapoi is also needed in these countries to ascertain the status of this species and prevent it from a possible acute threat of extinction.
Data availability statement
The original contributions presented in this study are included in this article/Supplementary materials, further inquiries can be directed to the corresponding author.
Author contributions
HY: Conceptualization, Data curation, Investigation, Methodology, Writing – review & editing. XP: Conceptualization, Investigation, Writing – original draft. HZ: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Resources, Writing – original draft, Writing – review & editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by the Hainan Provincial Natural Science Foundation of China (422CXTD508) and the Specific Research Fund of the Innovation Platform for Academicians of Hainan Province.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1356104/full#supplementary-material
References
Ahrends, A., Hollingsworth, P. M., Ziegler, A. D., Fox, J. M., Chen, H., Su, Y., et al. (2015). Current trends of rubber plantation expansion may threaten biodiversity and livelihoods. Glob. Environ. Chang. 34, 48–58. doi: 10.1016/j.gloenvcha.2015.06.002
Carboni, M., Livingstone, S. W., Isaac, M. E., and Cadotte, M. W. (2021). Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 109, 3587–3601. doi: 10.1111/1365-2745.13739
Chen, L., and Zhou, S. (2015). A combination of species evenness and functional diversity is the best predictor of disease risk in multihost communities. Am. Nat. 186, 755–765. doi: 10.1086/683774
Circular of the people’s Government of Hainan Province (1988). The first batch of key scenic spots and nature reserves in Hainan Province. Available online at: http://hnsthb.hainan.gov.cn/xxgk/0200/ 0202/zwgk/zcfg/ 2019 01/t20190106_ 2123256 (accessed 22 October 2022).
Corlett, R. T. (2011). Impacts of warming on tropical lowland rainforests. Trends Ecol. Evol. 26, 606–613. doi: 10.1016/j.tree.2011.06.015
Corlett, R. T., and Primack, R. B. (2006). Tropical rainforests and the need for cross-continental comparisons. Trends. Ecol. Evol. 21, 104–110. doi: 10.1016/j.tree.2005.12.002
Dai, Z., Si, C., Zhai, D., Huang, P., Qi, S., Lin, Y., et al. (2018). Genetic effects of historical anthropogenic disturbance on a long-lived endangered tropical tree Vatica mangachapoi. J. For. Res. 29, 291–299. doi: 10.1007/s11676-017-0470-z
De Guzman, E. D., Umali, R. M., and Sotalbo, E. D. (1986). Guide to Philippine flora and fauna. Quezon City: Natural Resources Management Center, Ministry of Natural Resources and University of the Philippines, 1–73.
Ding, Y., Zang, R.-G., and Jiang, Y.-X. (2006). Effect of Hillslope gradient on vegetation recovery on abandoned land of shifting cultivation in Hainan Island, South China. J. Integr. Plant Biol. 48, 642–653. doi: 10.1111/j.1744-7909.2006.00262.x
Ghazoul, J. (2016). Dipterocarp biology, ecology, and conservation. Oxford: Oxford University Press, doi: 10.1093/acprof:oso/9780199639656.001.0001
Guo, J., Shang, S., and Zeng, J. (2018). Morphometric study confirms the presence of only Vatica mangachapoi on Hainan Island, China. J. For. Res. 29, 639–646. doi: 10.1007/s11676-017-0458-8
Hajdas, I., Ascough, P., Garnett, M. H., Fallon, S. J., Pearson, C. L., Quarta, G., et al. (2021). Radiocarbon dating. Nat. Rev. Methods Prim. 1:62. doi: 10.1038/s43586-021-00058-7
Harms, K. E., Condit, R., Hubbell, S. P., and Foster, R. B. (2001). Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 89, 947–959. doi: 10.1111/j.1365-2745.2001.00615.x
Ichie, T., Igarashi, S., Yoshihara, R., Takayama, K., Kenzo, T., Niiyama, K., et al. (2022). Verification of the accuracy of the recent 50 years of tree growth and long-term change in intrinsic water-use efficiency using xylem Δ14C and δ13C in trees in an aseasonal tropical rainforest. Methods Ecol. Evol. 13, 1135–1147. doi: 10.1111/2041-210X.13823
Liu, X., Chen, L., Liu, M., García-Guzmán, G., Gilbert, G. S., and Zhou, S. (2020). Dilution effect of plant diversity on infectious diseases: Latitudinal trend and biological context dependence. Oikos 129, 457–465. doi: 10.1111/oik.07027
Liu, X., Lyu, S., Zhou, S., and Bradshaw, C. J. (2016). Warming and fertilization alter the dilution effect of host diversity on disease severity. Ecology 97, 1680–1689. doi: 10.1890/15-1784.1
Livingstone, S. W., Isaac, M. E., and Cadotte, M. W. (2020). Invasive dominance and resident diversity: Unpacking the impact of plant invasion on biodiversity and ecosystem function. Ecol. Monogr. 90:e01425. doi: 10.1002/ecm.1425
Phiri, D., and Morgenroth, J. (2017). Developments in landsat land cover classification methods: A review. Remote Sens. 9:967. doi: 10.3390/rs9090967
Pooma, R., Barstow, M., and Newman, M. F. (2017). Vatica mangachapoi. IUCN Red List Threat. Species 2017:e.T32461A2819415. doi: 10.2305/IUCN.UK.2017-3.RLTS.T32461A2819415.en
Soepadmo, E., Saw, L. G., and Chung, R. C. K. (2004). Tree flora of Sabah and Sarawak, Vol. 5. Kuching: Sabah Forestry Department, Forest Research Institute Malaysia (FRIM), Sarawak Forestry Department, 63–388.
Warren-Thomas, E., Nelson, L., Juthong, W., Bumrungsri, S., Brattström, O., Stroesser, L., et al. (2020). Rubber agroforestry in Thailand provides some biodiversity benefits without reducing yields. J. Appl. Ecol. 57, 17–30. doi: 10.1111/1365-2664.13530
Zhang, M., Fellowes, J. R., Jiang, X., Wang, W., Chan, B. P., Ren, G., et al. (2010). Degradation of tropical forest in Hainan, China, 1991–2008: Conservation implications for Hainan gibbon (Nomascus hainanus). Biol. Conserv. 143, 1397–1404. doi: 10.1016/j.biocon.2010.03.014
Zheng, Y., Xie, Y., Xie, Y., and Yu, S. (2021). Asexual reproduction and vegetative growth of Bionectria ochroleuca in response to temperature and photoperiod. Ecol. Evol. 11, 10515–10525. doi: 10.1002/ece3.7856
Zimmerman, N. B., and Vitousek, P. M. (2012). Fungal endophyte communities reflect environmental structuring across a Hawaiian landscape. Proc. Natl. Acad. Sci. U.S.A. 109, 13022–13027. doi: 10.1073/pnas.1209872109
Keywords: Chinese Vatica mangachapoi, fungal disease, plant diversity, stem rot disease, vine strangulation
Citation: Yu H, Pei X and Zhang H (2024) Threat categories of Vatica mangachapoi should be reassessed. Front. For. Glob. Change 7:1356104. doi: 10.3389/ffgc.2024.1356104
Received: 18 December 2023; Accepted: 15 February 2024;
Published: 07 March 2024.
Edited by:
Chengjin Chu, Sun Yat-sen University, ChinaReviewed by:
Diego Rodriguez, University of Bristol, United KingdomXiang Liu, Lanzhou University, China
Copyright © 2024 Yu, Pei and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hui Zhang, OTkzNzgxQGhhaW5hbnUuZWR1LmNu
†These authors have contributed equally to this work