 Christopher M. Gough
Christopher M. Gough1 Introduction
When drafting a research article (Gough et al., 2021), I was at a loss for a term that describes the status of the ecosystem-wide resource base before disturbance that could help buffer against changes in forest functioning during and after disturbance. The concept of “material and information legacies”, summarized by Johnstone et al. (2016), describes the biotic and abiotic resources, including the “information” encoded in genes, that are retained through disturbance and which aid in the recovery of community structure and ecosystem functioning. Such legacies include retained biomass, nutrients, seeds and other propagules, and adaptive traits that pass through a “disturbance filter”, supporting recolonization, regrowth, and the recovery of biological and genetic diversity. But, what term conveys the status and quantity of these functionally-relevant resources prior to disturbance?
The answer: forest ecologists and managers have no agreed-upon term to describe the status of disturbance-buffering materials before disturbance occurs, even though the abundance and diversity of biotic and abiotic resources before disturbance influences how ecosystems respond to droughts and extreme weather, insect pests, pathogens, and fire (Burton et al., 2020). For example, forests depleted in nutrient capital, biological and genetic diversity, seed supply, and plant biomass before disturbance are more prone to significant changes in structure and function when disturbance occurs (Hughes and Stachowicz, 2011; Kranabetter et al., 2016). Moreover, the material and information legacies that persist through disturbance are partly dependent upon the resource base prior to disturbance, with more frequent and severe disturbances progressively eroding stored resources (Latty et al., 2004).
2 Ecosystem capacitance as an indicator of disturbance buffering capacity
Modifying century-old terminology used by physicists and engineers and, more recently, plant physiologists, I propose the use of “ecosystem capacitance” to describe the status of stored biotic and abiotic materials available to sustain or support the recovery of systems-level functioning before disturbance occurs. Broadly, “capacitance” refers to stored energy or mass that, when mobilized, buffers against systems-level change. Originally referencing the storage of electric charge, plant physiologists use capacitance to describe the quantity of stored water within tissues (Bryant et al., 2021) and ecosystems (Konings et al., 2021), a resource that can be remobilized to stabilize water transport thorough stems and into the atmosphere (i.e., transpiration) during periods of drought.
Pivoting from its original definition, “evolutionary capacitance” describes accumulated genetic variation within a population, which, under stressful conditions, may be crucial to adaptation and population persistence (Bergman and Siegal, 2003).
The adoption of such terminology has benefits because, like metrics of ecological stability borrowed from physics, ecosystem capacitance could be quantified and, therefore, serve as a diagnostic measure of an ecosystem's capacity to buffer against or reduce disturbance-driven changes in structure and function. Concepts and terminology borrowed from physics and engineering are not new to ecology. Electrical resistor diagrams have been used by plant physiologists for decades to map and model how water moves from soils through plants and into the atmosphere (Luo et al., 2013). More recently, conceptions and calculations of ecosystem stability, which encompass several component parts, including resistance and resilience, borrow heavily from engineering principles to describe and quantify structural and functional changes in response to disturbance (Hillebrand et al., 2018).
3 Discussion and application to forest management and disturbance mitigation
For forest managers, ecosystem capacitance could serve as an indicator of the potential to sustain or recover functioning, and goods and services after disturbance. Similar to ecosystem health indicators (Shear et al., 2003), a systematic but tailored accounting of ecosystem capacitance could inform conservation and management goals that aim to sustain or recover targeted goods and services following regionally-forecasted disturbance events. In this context, the biotic and abiotic materials assessed to derive ecosystem capacitance would be system- and disturbance-dependent, and consider which resources most limit the functions, goods, or services of interest. For example, in accordance with the “insurance hypothesis”, high biological, functional, and genetic diversity alone may reduce temporal variation in wood production and carbon sequestration during drought and in response to extreme weather (Mori et al., 2013; Morin et al., 2014), but additional accounting of plant biomass and nutrient capital may be critical if the goal is maximizing rates of forest carbon sequestration. The factors underlying ecosystem capacitance also will vary by forest type because principal limiting resources (Running et al., 2004) and prevailing disturbance regimes (Cohen et al., 2018) differ substantially among forest biomes and plant functional types (Amiro et al., 2010). For example, water is a primary limiting resource in evergreen needle leaf forests of western North America, where insects and fire are prominent disturbances (Masek et al., 2013). In contrast, temperate deciduous forests in eastern North America range from nitrogen-limited to nitrogen-polluted (Vitousek et al., 2022), and are more susceptible to insects, pathogens, and wind throw (Fischer et al., 2013; Flower and Gonzalez-Meler, 2015). Extending the diversity-focused insurance hypotheses, indices of ecosystem capacitance would ideally encompass and integrate all quantifiable abiotic and biotic properties that strengthen ecosystem stability in the presence of disturbance (Running et al., 2004; Cohen et al., 2018; Vitousek et al., 2022). While the factors contributing to ecosystem capacitance will differ among forests, the conceptual relationship between capacitance and ecosystem goods and services is the same: across a continuum of disturbance severity, frequency, or duration, forest ecosystems with low capacitance are expected to exhibit a loss of function more rapidly or at lower disturbance thresholds than ecosystems with high capacitance (Figure 1).
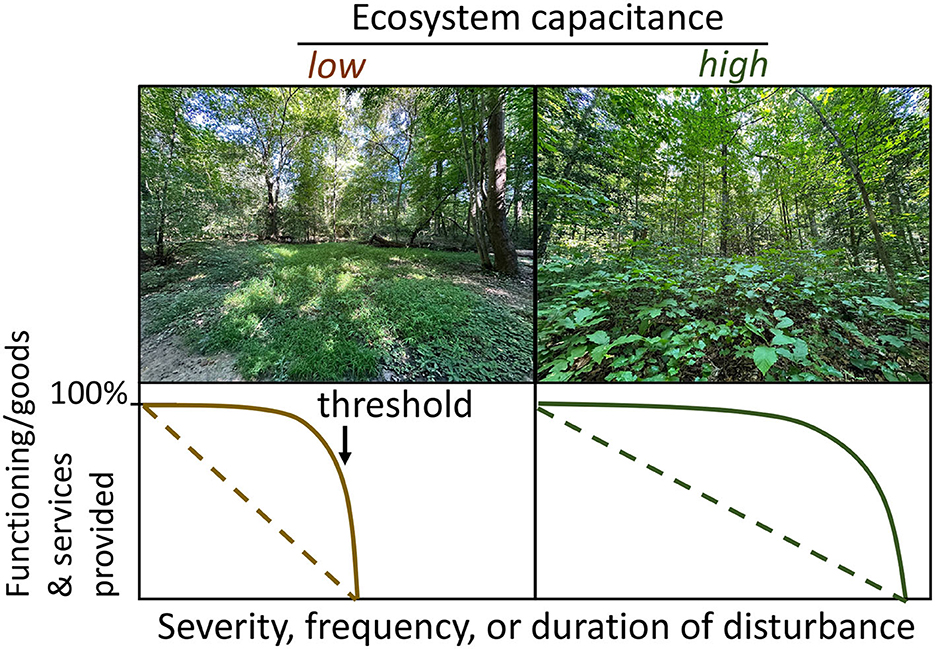
Figure 1. Temperate deciduous broadleaf forests with contrasting low and high ecosystem capacitance, respectively. The ecosystem on the left is depauperate of indigenous plant biodiversity, mid-canopy plant biomass, and soil organic matter, resources that are crucial to sustaining a number of functions—from wood production to nutrient retention—during and after insect pest and pathogen outbreaks or drought, disturbances that are increasingly common in temperate deciduous broadleaf forests. The high capacitance ecosystem on the right, contrastingly, has abundant stored resources and is expected to exhibit less or delayed functional change as disturbance severity, frequency, or duration increases. In both cases, ecological theory and observations suggest declines in ecosystem goods and services with increasing disturbance severity, frequency, or duration could proceed linearly (dashed line) or non-linearly.
Developing tractable approaches for quantifying meaningful measures of ecosystem capacitance will require dialogue and collaboration among forest scientists and managers, akin to the iterative process that led to practitioner- and policy-relevant wetland assessment approaches and measures (Herlihy et al., 2019). Assessments of wetlands, similarly motivated by ecological restoration and sustainability goals, were developed through long-term partnerships among researchers, practitioners, and policy makers. With standardized wetland assessment protocols in wide use, these tools provide intercomparable data-driven and evidence-based diagnostics with which to guide preemptive and adaptive management, while employing methods and providing metrics that are accessible and meaningful to managers (Maltby, 2022). Indeed, national forest inventories, such as the United States Department of Agriculture's Forest Inventory and Analysis, report a suite of longitudinal data that could be used to develop quantitative indicators of ecosystem capacitance, including tree crown conditions, stand biomass, tree mortality, and forest composition (Woodall et al., 2011). Moreover, such ground-based inventories provide crucial assessments of ecosystem functioning and goods and services, including carbon sequestration and timber production. When paired with increasingly available remotely-sensed data describing the physical arrangement and taxonomic diversity of vegetation within forest canopies, inventory data provide a rich complementary dataset with which to develop and test the efficacy of ecosystem capacitance as an indicator of preparedness against future disturbance.
Do we need another term in the already extensive glossary of forest ecology? Possibly not, but the conceptual and quantitative frameworks used to characterize, predict, and interpret the effects of disturbance on forest biodiversity, structure, function, goods and services remain unsettled (Müller et al., 2023). With global change-related disturbances increasing, the standardization and maturation of language and concepts that forest ecologists and managers use to describe, predict, mitigate, and measure disturbance effects on ecosystems is, arguably, more important than ever.
Author contributions
CG: Conceptualization, Writing – original draft.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by National Science Foundation, Department of Environmental Biology, Award 2219695.
Acknowledgments
The author thanks Gough Lab members and a peer reviewer for their comments and contributions.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Amiro, B. D., Barr, A. G., Barr, J. G., Black, T. A., Bracho, R., Brown, M., et al. (2010). Ecosystem carbon dioxide fluxes after disturbe in forests of North America. J. Geophysi. Res.-Biogeosci. 115. doi: 10.1029/2010JG001390
Bergman, A., and Siegal, M. (2003). Evolutionary capacitance as a general feature of complex gene networks. Nature 424, 549–552. doi: 10.1038/nature01765
Bryant, C., Fuenzalida, T. I., Brothers, N., Mencuccini, M., Sack, L., Binks, O., et al. (2021). Shifting access to pools of shoot water sustains gas exchange and increases stem hydraulic safety during seasonal atmospheric drough. Plant Cell Environm. 44, 2898–2911. doi: 10.1111/pce.14080
Burton, P. J., Jentsch, A., and Walker, L. R. (2020). The ecology of disturbance interions. Bioscience 70, 854–870. doi: 10.1093/biosci/biaa088
Cohen, W. B., Yang, Z. Q., Heale, S. P., Kennedy, R. E., and Gorelic, N. (2018). A LandTrendr multispectral ensemble for forest disturbance detection. Remote Sens. Environm. 205, 131–140. doi: 10.1016/j.rse.2017.11.015
Fischer, A., Marshall, P., and Camp, A. (2013). Disturbances in deciduous temperate forest ecosystems of the Northern Hemisphere: their effects on both recent anuture forest development. Biodivers. Conserv. 22, 1863–1893. doi: 10.1007/s10531-013-0525-1
Flower, C. E., and Gonzalez-Meler, M. A. (2015). Responses of temperate forest productivity to insect and pathogen disturbances. Annu Rev Plant Biol. 66, 547–569. doi: 10.1146/annurev-arplant-043014-115540
Gough, C. M., Atkins, J., Ben, B.-L., Agee, E. A., Dorheim, K. R., Fahey, R. T., et al. (2021). Forest structural complexity and biomass predict firsar carbon cycling responses to disturbance. Ecosystems 24, 699–712. doi: 10.1007/s10021-020-00544-1
Herlihy, A. T., Kentula, M. E., Magee, T. K., Lomnicky, G. A., Nahlik, A. M., and Serenbetz, G. (2019). Striving for consistency in the National Wetland Condition Assessment: developing a reference condition approach for assessing wetlands at a continental scale. Environ. Monit. Assess. 191, 327. doi: 10.1007/s10661-019-7325-3
Hillebrand, H., Langenheder, S., Lebret, K., Lindstrom, E., Ostman, O., and Stri, M. (2018). Decomposing multiple dimensions of stability in global change experiments. Ecol. Lett. 21, 21–30. doi: 10.1111/ele.12867
Hughes, A. R., and Stachowicz, J. J. (2011). Seagrass genotypic diversity increases disturbance response via complementarand dominance. J. Ecol. 99, 445–453. doi: 10.1111/j.1365-2745.2010.01767.x
Johnstone, J. F., Allen, C. D., Franklin, J. F., Frelich, L. E., Harvey, B. J., Higuera, P. E., et al. (2016). Changing disturbancgimes, ecological memory, and forest resilience. Front. Ecol. Environ. 14, 369–378. doi: 10.1002/fee.1311
Konings, A. G., Saatchi, S. S., Frankenberg, C., Keller, M., Leshyk, V., Anderegg, W. R. L., et al. (2021). Detecting forest response to droughts with global observations of vegetation water ent. Glob. Chang. Biol. 27, 6005–6024. doi: 10.1111/gcb.15872
Kranabetter, J. M., McLauchlan, K. K., Enders, S. K., Fraterrigo, J. M., Higuera, P. E., Morris, J. L., et al. (2016). A framework assess biogeochemical response to ecosystem disturbance using nutrient partitioning ratios. Ecosystems 19, 387–395. doi: 10.1007/s10021-015-9934-1
Latty, E. F., Canham, C. D., and Marks, P. L. (2004). The effects of land-use history on soil properties and nutr dynamics in northern hardwood forests of the Adirondack Mountains. Ecosystems 7, 193–207. doi: 10.1007/s10021-003-0157-5
Luo, X. Y., Liang, X., and McCarthy, H. R. (2013). VIC plus for water-lim conditions: a study of biological and hydrological processes and their interactions in soil-plant-atmosphere continuum. Water Resourc. Res. 49, 7711–7732. doi: 10.1002/2012WR012851
Maltby, E. (2022). The wetlands paradigm shift in response to changing societal priorities: a reflective review. Land. 11, 1526. doi: 10.3390/land11091526
Masek, J. G., Goward, S. N., Kennedy, R. E., Cohen, W. B., Moisen, G. G., Schleeweis, K., et al. (2013). United States forest disturbance trends observed using landsat time series. Ecosyst. 16, 1087–1104. doi: 10.1007/s10021-013-9669-9
Mori, A. S., Furukawa, T., and Sasaki, T. (2013). Response diversity determines the resilience of ecosystems to environmental change. Biol. Rev. 88, 349–364. doi: 10.1111/brv.12004
Morin, X., Fahse, L., de Mazancourt, C., Scherer-Lorenzen, M., and Bugmann, H. (2014). Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 17, 1526–1535. doi: 10.1111/ele.12357
Müller, J., Mitesser Cadotte, M. W., van der Plas, F., Mori, A. S., Ammer, C., et al. (2023). Enhancing the structural diversity between forest patches-a concept and real-world experiment to study biersity, multifunctionality and forest resilience across spatial scales. Glob. Chang. Biol. 29, 1437–1450. doi: 10.1111/gcb.16564
Running, S. W., Nemani, R. R., Heinsch, F. A., Zhao, M. S., and Reeves Hashimoto, H. (2004). A continuous satellite-derived measure of global terrestrial primary production. Bioscience 54, 547–560. doi: 10.1641/0006-3568(2004)054(0547:ACSMOG)2.0.CO;2
Shear, H., Stadler-Salt, N., and Bertram Horvatin, P. (2003). The development and implementation of indicators of ecosystem health in the Great Lakes basin. Environm. Moning Assessm. 88, 119–152. doi: 10.1023/A:1025504704879
Vitousek, P. M., Treseder, K. K., Howarth, R. W., and Menge, D. N. L. (2022). A “toy model” analysis of causes of nitrogen limitation in terrestrial ecosystems. Biogeochemistry 160, 381–394. doi: 10.1007/s10533-022-00959-z
Keywords: disturbance, stability, resistance, diversity, insurance hypothesis
Citation: Gough CM (2024) Ecosystem capacitance: an integrative buffer against disturbance. Front. For. Glob. Change 6:1326569. doi: 10.3389/ffgc.2023.1326569
Received: 23 October 2023; Accepted: 21 December 2023;
Published: 12 January 2024.
Edited by:
Sigrid Netherer, University of Natural Resources and Life Sciences Vienna, AustriaReviewed by:
Alexander Krichels, Rocky Mountain Research Station, Forest Service (USDA), United StatesCopyright © 2024 Gough. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christopher M. Gough, Y21nb3VnaEB2Y3UuZWR1