 Branko Kanjevac1*
Branko Kanjevac1* Violeta Babić1Snežana Stajić2Nikola Martać3Branka Pavlović3Danilo Furtula4Vlado Čokeša3
Violeta Babić1Snežana Stajić2Nikola Martać3Branka Pavlović3Danilo Furtula4Vlado Čokeša3- 1Faculty of Forestry, Chair of Silviculture, University of Belgrade, Belgrade, Serbia
- 2Department of Forest Establishment, Silviculture and Ecology, Institute of Forestry, Belgrade, Serbia
- 3Department of Forest Management Planning, Organizing and Economics, Institute of Forestry, Belgrade, Serbia
- 4Department of Forest Protection, Institute of Forestry, Belgrade, Serbia
Understanding the processes occurring in old-growth forests and identifying their key aspects can significantly enrich modern forestry practices with innovative ideas and concepts. The natural regeneration process in beech old-growth forests exhibits distinct spatial heterogeneity and temporal variability. To define the key drivers that influence the spatial heterogeneity of regeneration processes and their effects, research was conducted in three beech old-growth forests situated in Serbia, Southeastern Europe: Felješana, Vinatovača, and Kukavica. In each old-growth forest, a network of circular sample plots with an area of 0.1 ha (totaling 45 plots) was established to gather data on structural characteristics and ecological conditions. Within each circular sample plot, data on the regeneration layer were collected on four square sample plots of 1 m2 (180 in total). Using linear mixed models, the key drivers of spatial heterogeneity of regeneration processes in beech old-growth forests were analyzed. Based on the results, several key factors contribute to the highly heterogeneous distribution of seedlings, including the canopy, the presence of a middle layer comprising young trees, ground vegetation, and soil stoniness, while a significant influence of the combined effect of the canopy and the presence of a middle layer of young trees is also defined. The spatial heterogeneity of the regeneration process is also represented through the assessment of the ratio between the abundance of one-year-old and older seedlings. The dominance of one-year-old seedlings intensifies with increased canopy density (in instances of very dense canopy (1.0), the ratio of one-year-old and older seedlings is 70:30%). Seedling growth characteristics are shaped by multiple factors, including the influence of the canopy, the presence of the middle layer of young trees, slope, and soil stoniness, with a substantial combined influence of the canopy and the middle layer of young trees. This indicates that the spatial variability of the regeneration process in beech old-growth forests is primarily driven by factors with a substantial individual influence, which may also act combined. It is of paramount importance to understand these factors and determine their influence on the regeneration process in managed beech forests.
1 Introduction
Old-growth forests in contemporary conditions constitute distinct components of the global forest ecosystem, characterized by pronounced specificity of ecological conditions, structural characteristics and frequently pronounced spatial heterogeneity (Szewczyk and Szwagrzyk, 2010; Čurović et al., 2011; Keren et al., 2017; Feldmann et al., 2020). The processes occurring within these ecosystems are markedly slow-paced and integrated with the natural laws that dictate their dynamics (Glatthorn et al., 2018). In order to preserve the complex dynamics of forest ecosystems, modern forest management concepts rely on designing treatments that mimic natural processes and disturbances and thus promote and preserve the biodiversity and functionality of forest ecosystems (Wirth et al., 2009; Schröter et al., 2012; Govedar et al., 2018; Bagnato et al., 2021). Unlike old-growth forests that are characterized by a passive regime, managed forests are in a more favorable position since they can provide support to potentially endangered species through silvicultural treatments such as planting, protection from browsing, competition regulation, etc. (Diaci et al., 2022).
Despite beech forests being among the most widespread forest types in Europe, there are only small areas of old-growth beech forests scattered across remote and inaccessible locations in the Carpathians, Balkans and Alps (Commarmot and Brang, 2011; Hobi et al., 2015a). Due to their uniqueness and lack of anthropogenic influence, these forests hold particular importance for the ecology, conservation and management of forests in Europe (Govedar and Stanivuković, 2008; Wirth et al., 2009; Ammer et al., 2018; Čurović et al., 2020; Feldmann et al., 2020).
A very important issue in old-growth forests pertains to the process of regeneration, considering that its characteristics and dynamics can provide insights valuable for the harmonization and improvement of the existing regeneration systems applied in forests that are subject to regular management. Natural regeneration is considered a very effective nature-based solution aimed at restoring and increasing biodiversity, both woody and non-woody species in forest ecosystems (Rozendaal et al., 2019; Jaloviar et al., 2020; Siminski et al., 2021). Consequently, it is well-known that increased diversity has a stabilizing influence on productivity and increases forest resistance in dynamic environmental conditions (Jucker et al., 2014; Ford and Keeton, 2017).
Beech exhibits high regeneration potential due to its key bioecological characteristics, which include extensive and regular seed production, shade tolerance, wide ecological amplitude and the ability to conquer space rapidly under favorable developmental conditions (Wagner et al., 2010; Barna and Bosela, 2015). Although regeneration in old-growth beech forests follows well-established patterns, this process has so far been insufficiently studied, particularly concerning its fundamental characteristics, such as balance and continuity (Szwagrzyk and Szewczyk, 2008; Rugani et al., 2013). In the absence of serious disturbances (stormy winds, etc.), natural regeneration in these forests begins spontaneously on a small scale, initiated by the gradual death of individual trees or under the canopy of mature trees that provide a substantial quantity of seeds (Bottero et al., 2011; Glatthorn et al., 2018). Beech has the capacity to establish dense seedling formations in the initial phase of development under a very dense canopy of mature trees (Collet et al., 2001; Modry et al., 2004). However, it does not guarantee a successful regeneration; on the contrary, it frequently results in very high or complete mortality of one-year-old seedlings (Hobi et al., 2015b; Petrovska et al., 2022). This finding underscores the high dependence of beech seedling development on the available amount of light or canopy openness (Petritan et al., 2009; Vacek et al., 2015). The dynamics of canopy opening creation in these forests represent a key factor affecting the structural heterogeneity and diversity of species (Rozenbergar et al., 2007; Massad et al., 2019). When small to large canopy openings emerge, suppressed seedlings gain space for vigorous growth, possessing a significant advantage over seedlings that have just germinated due to their developed root system (Feldmann et al., 2020). In this way, dynamic structural and microclimatic conditions contribute to the creation of fine uneven-aged and multi-layered structures, which are one of the main characteristics of these forests (Motta et al., 2011; Zenner et al., 2015).
The key features of the natural regeneration process in old-growth beech forests are spatial heterogeneity and temporal variability (Szewczyk and Szwagrzyk, 2010; Hobi et al., 2015a). Despite being very slow, this process is continuous, balanced and marked by pronounced complexity, which results from the direct or indirect influence of multiple factors (Diaci et al., 2012). The emergence of seedling banks, the dynamics of their development, their disappearance due to the canopy closing, and the interspecific and intraspecific competitive interactions that occur within the seedling banks are segments that reflect the overall dynamics of old-growth beech forest regeneration. These segments continuously intertwine, thus sustaining the process and maintaining its balance (Dröβler and von Lüpke, 2005; Madsen and Hahn, 2008). In addition, beech proves to be an extremely dominant species in its habitats, suppressing the presence of other broadleaved trees and thus preserving its composition (Petrovska et al., 2023).
Based on the above, the study aimed to achieve the following objectives: (i) study the impact of spatial heterogeneity in structural characteristics and ecological conditions on natural regeneration in old-growth beech forests in Southeastern Europe, (ii) define the ratio of one-year-old to older seedlings concerning spatial heterogeneity, and (iii) define the characteristics of seedling growth in relation to spatial heterogeneity and identify the dominant factors influencing their development in old-growth beech forests.
2 Materials and methods
2.1 Study site
The research was conducted in three old-growth beech forests in Serbia, Southeastern Europe. These forests included Felješana (44°20′ N; 21°53′ E), Vinatovača (44°04′ N; 21°46′ E) and Kukavica (42°47′ N; 22°00′ E). All three old-growth forests have been designated strict nature reserves by the Government of the Republic of Serbia (Felješana 1950; Vinatovača 1957; Kukavica 1980). In addition, even prior to their protected status, there were no intensive management treatments in these forests, given that they are located in isolated and inaccessible locations.
The examined stands are located at altitudes ranging from 520 to 1,175 m a.s.l., on steep slopes and mostly northern aspects, with a semi-humid climate at lower altitudes and a humid climate at higher altitudes (Lang, 1915; Thornthwaite, 1948) (Table 1). Dystric cambisol soil type prevails in all three old-growth forests, with the bedrock consisting of schist and limestone in Felješana and Vinatovača and gneiss in Kukavica (Table 1).
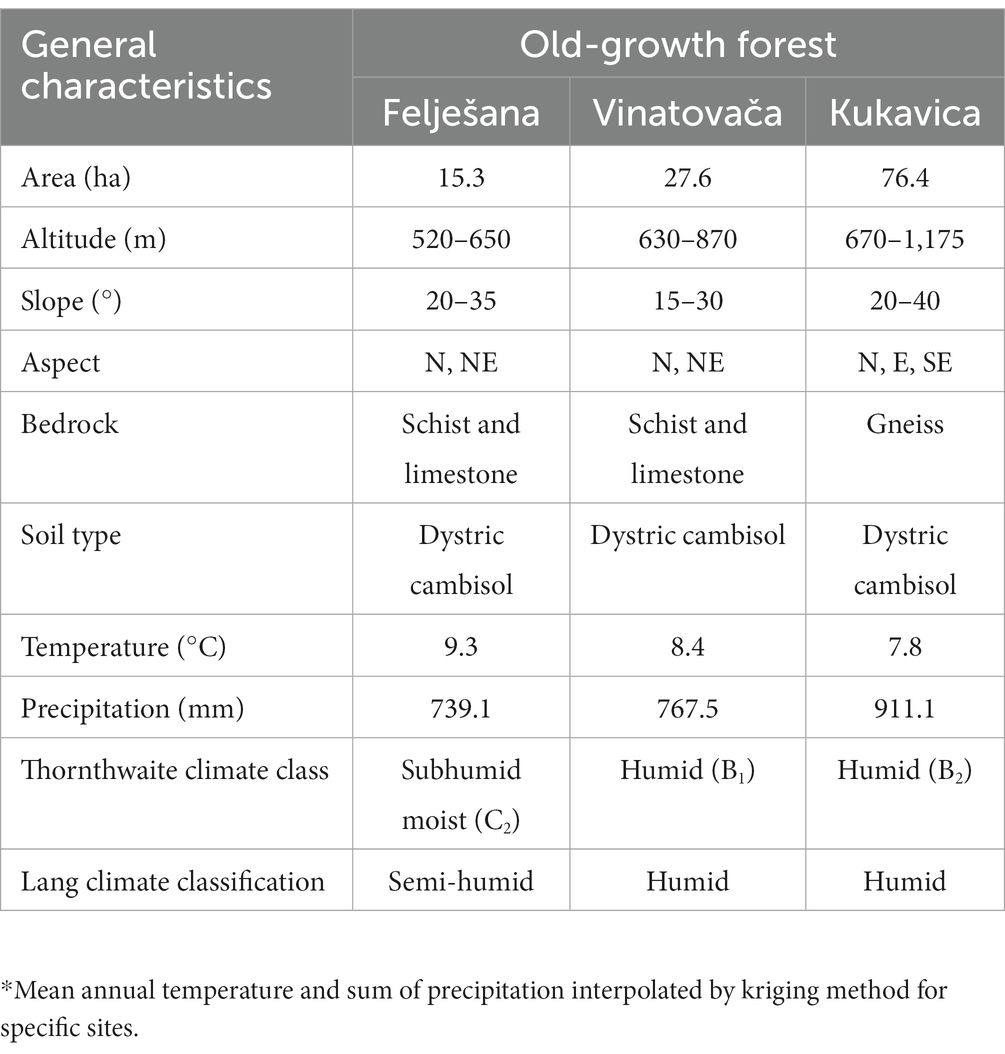
Table 1. General characteristics of the studied old-growth beech forests.
2.2 Study design
In each of the three stands, a network of 15 circular sample plots with an area of 0.1 ha (45 plots in total) was established at appropriate distances to collect data related to ecological conditions and structural characteristics of the stands (Figure 1). To define ecological conditions, data were collected on edaphic (bedrock, soil type, stoniness recorded to the nearest 10% within the range from 10 to 100%) and orographic factors (altitude, slope, aspect). To define the structural characteristics of the stands, the diameter at breast height and the total height of all trees above the estimation threshold of 5 cm were measured.
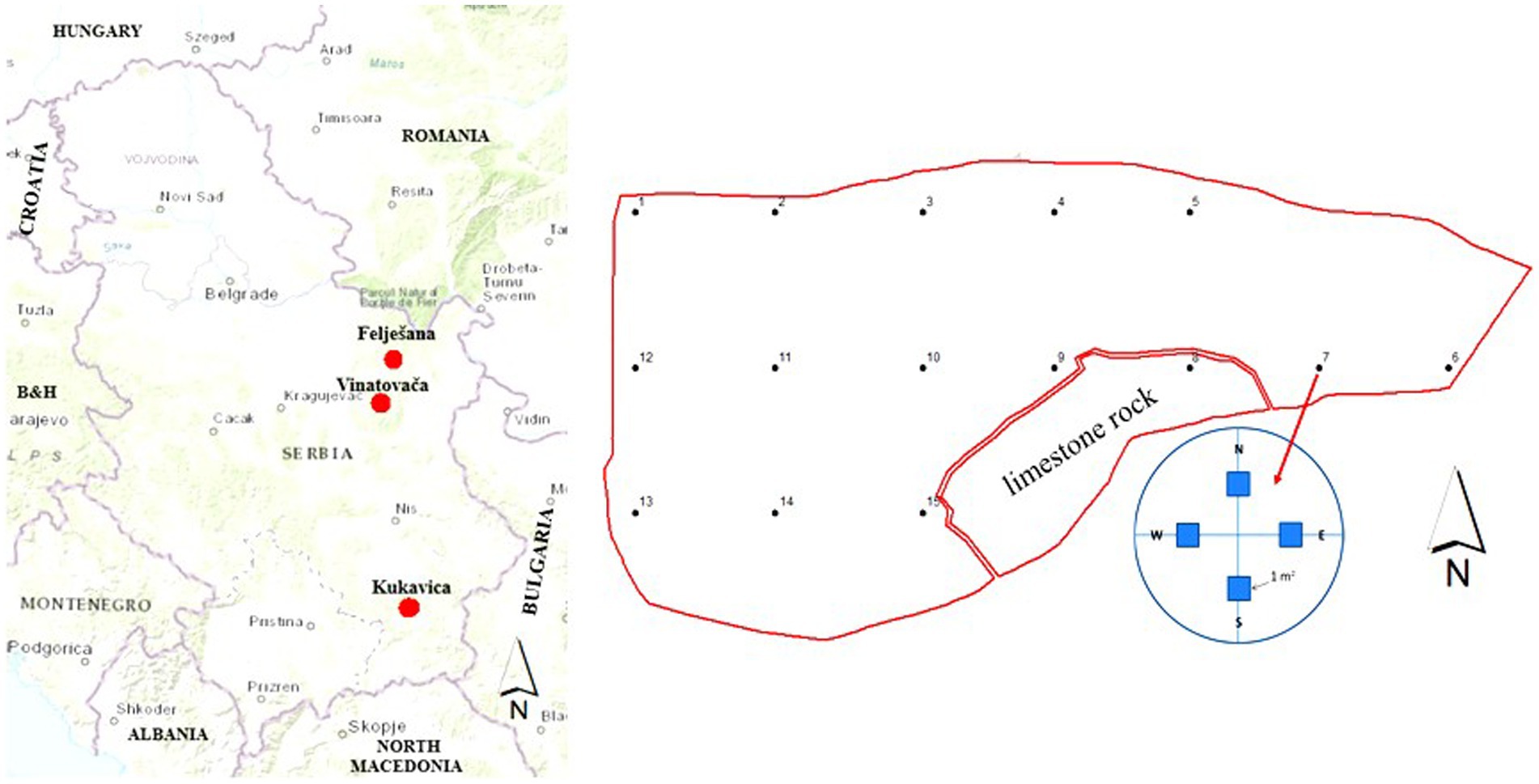
Figure 1. Locations of the studied old-growth forests and the arrangement of circular and square sample plots in the old-growth forest Felješana.
Within each circular sample plot, four square sample subplots of 1 m2 (180 in total), oriented according to the four cardinal directions of the world, were established to collect data on the seedling development conditions, seedling growth characteristics and the presence of ground vegetation (Figure 1). To begin, a visual assessment of the stand canopy recorded to the nearest 10% within the range from 10 to 100% was performed, and the presence of a middle layer of young trees was noted. To assess the presence and growth characteristics of woody seedlings with dbh < 5 cm, the following data were collected: the number of one-year-old and older seedlings, their total height, height increment, and diameter at root collar. The height and height increment measurements were taken using a measuring tape with an accuracy of 1.0 cm, and the diameter at root collar was measured using a digital caliper with an accuracy of 1.0 mm. The percentage cover of ground vegetation was visually estimated from above and recorded to the nearest 10% within the range from 10 to 100% and to the nearest 1% within the range from 1 to 10%.
2.3 Data analysis
Data analysis was conducted using Microsoft Excel Version 2019 and R Version 4.3.1 (R Core Team, 2023).
A comparative analysis of the fundamental structural parameters and regeneration characteristics of the studied stands was performed using descriptive statistical analysis. Given that beech is the dominant species in the regeneration layer, the subsequent analysis, focused on the definition of key drivers that influence the spatial heterogeneity of the regeneration process, is exclusively related to this species.
Principal component analysis (PCA) was used to analyze relationships between stand parameters (number of trees on a circular sample plot (N_trees), basal area of the 20% of largest trees (Gmax20%_trees), total height of the 20% largest trees (hmax20%_trees) and canopy), regeneration layer (the number of one-year-old seedlings (N_1_year_old), number of older seedlings (N_seedlings), total number of seedlings (N_total), maximum diameter at root collar of seedlings (dmax), maximum height of seedlings (hmax), maximum height increment of seedlings (lmax) and the presence of ground vegetation (Ground_vegetation)) and research plot attributes (Slope, Stoniness). The results of the PCA were visually presented in the form of an ordination diagram. PCA analysis was performed using MASS (Venables and Ripley, 2002), factoextra (Kassambara and Mundt, 2020), and ggplot2 (Wickham, 2016) packages.
The effects of structural characteristics of the studied stands, ecological conditions, and the presence of ground vegetation on the number of one-year-old, older, and total number of seedlings, as well as on the growth characteristics of seedlings, were analyzed using linear mixed-effects models (LMMs). A “top-down” strategy was applied (West et al., 2007). Dependent variables included the number of one-year-old, older, and total number of seedlings, maximum diameter at root collar, maximum height, and maximum height increment of seedlings. Fixed factors included the canopy, the presence of a middle layer of young trees, the presence of ground vegetation, stoniness and inclination, while the locality (Felješana, Vinatovača, Kukavica) was treated as the random factor. Depending on the percentage cover of ground vegetation, the degree of competition in the sample plots was defined as follows: low – the percentage cover of ground vegetation <30%; medium – the percentage cover of ground vegetation 30–60%; high – the percentage cover of ground vegetation >60%. The models were constructed using the “lme4 “package (Bates et al., 2015).
The normality of all dependent variables was checked using Kolmogorov-Smirnoff and Shapiro–Wilk tests. When necessary, variables were transformed using the common logarithm function to meet the assumptions of normality.
3 Results
3.1 Forest structure and regeneration
In Serbia, old-growth forests cover an area of about 1,200 ha, mostly consisting of old-growth beech forests. The old-growth beech forests investigated in this study are probably the best representatives of these forests, considering their condition, the habitats they occupy and the period of their status as protected natural assets.
The structural parameters demonstrate the high productivity of these stands and stress the high-quality of their sites. The volume of the examined stands significantly surpasses the average volume of beech high forests in Serbia, which stands at 269 m3/ha (Table 2). The small number of trees in the studied stands indicates their advanced age and the dominant presence of large trees whose diameters often exceed 80 cm, and heights reach beyond 40 m (Table 2). Additionally, the diameter distributions in all three old-growth forests have a decreasing trend as one moves from smaller to larger diameter classes, which is a typical structural pattern of such forests.
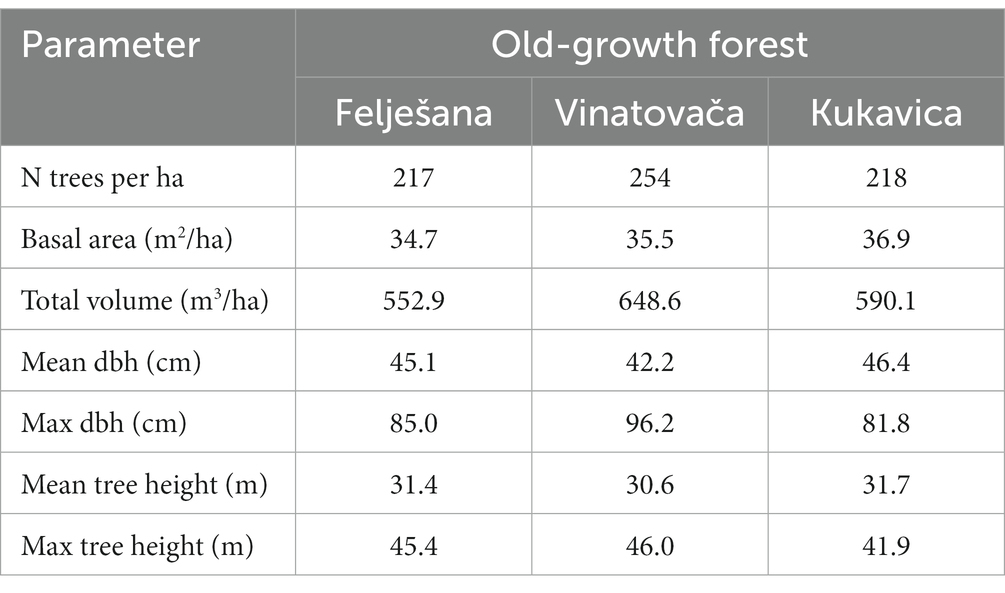
Table 2. Structural indicators of old-growth beech forests.
The analysis of the regeneration process focused on beech seedlings, given the negligible presence of other species (sycamore maple, wild cherry, sessile oak, hornbeam, etc.), which made their separate analyses unfeasible (Table 3). The regeneration process within these stands was characterized by high stability, as evident from the large number of seedlings. The Felješana old-growth forest had the highest number of seedlings, while the Vinatovača and Kukavica old-growth forests had 33.6 and 51.0% fewer seedlings, respectively (Table 3). These findings can be logically explained by the observation that a significant number of standing and fallen dead trees were recorded in the Felješana old-growth forest. These trees spontaneously created conditions favorable for the formation of seedling banks. All three studied stands had a large number of one-year-old seedlings, indicating the significant regenerative potential of parent trees in these stands and the limited conditions for the development of older seedlings. Older seedlings were mostly found in stand openings, where they had enough resources for their development. It was in these openings that beech seedling banks were formed, whereby the seedling height varied with the time of opening formation or the seedling bank formation.
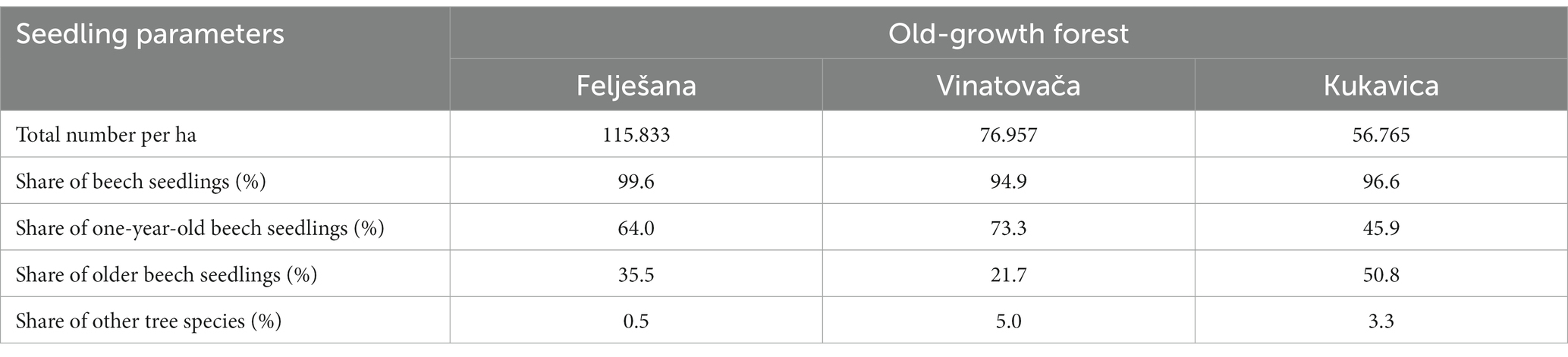
Table 3. Characteristics of old-growth beech forests regeneration.
The height distribution of seedlings in all three studied stands exhibited a decreasing trend, with the maximum presence of seedlings with a height < 30 cm (Figure 2). Due to the small amount of light available in the regeneration layer, seedlings did not have optimal conditions for development, which often initiated the horizontal growth of seedlings in search of diffuse light, small growth in height, and disappearance. The presence of seedlings with a height < 120 cm in the Felješana old-growth forest was 90.1%, in the Vinatovača old-growth forest 86.3%, and in the Kukavica old-growth forest 92.7% (Figure 2). When the conditions became more favorable, i.e., with an additional influx of light in the regeneration layer due to the crown decline or the tree fall, seedlings suddenly intensified their height growth. In the studied stands, the height increment of seedlings increased with the increased seedling height (Figure 3). This relationship is logical because seedlings of greater height were represented in seedling banks within stand openings, where increased competition and light availability favored the conditions for intensive height growth. Depending on the specific stand here studied, seedlings with a height < 120 cm had 62.8–70.8% lower average height increment compared to seedlings with a height > 120 cm (Figure 3).
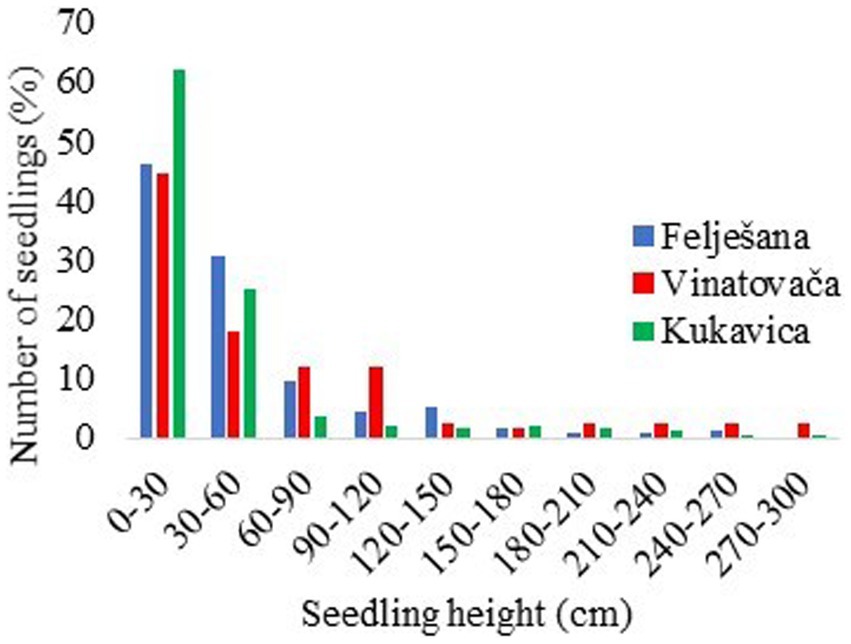
Figure 2. Height distribution of seedlings.
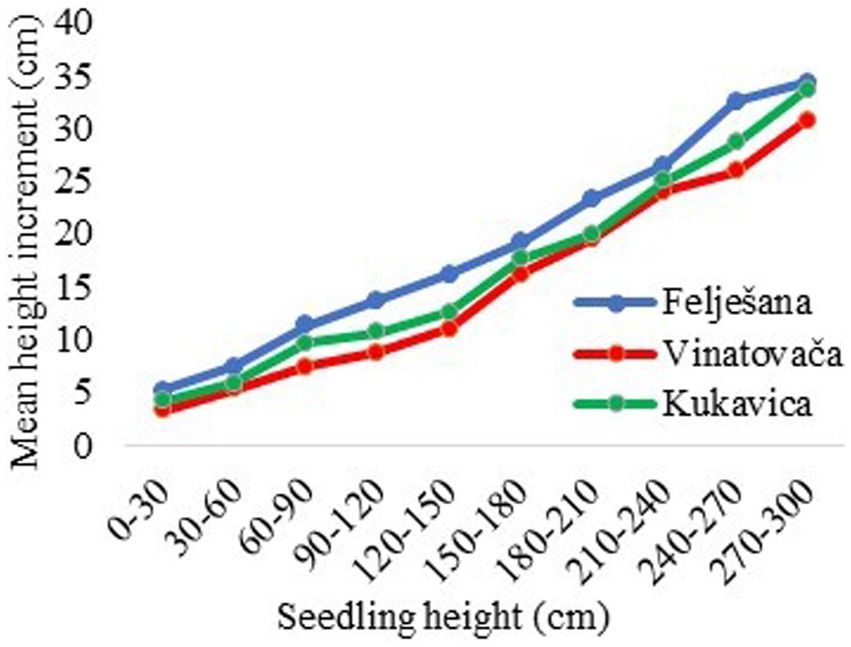
Figure 3. The ratio between height and height increment of seedlings.
3.2 Seedling occurrence and abundance
Regeneration of old-growth beech forests is characterized by marked spatial heterogeneity, resulting in an irregular distribution of seedlings through an area. Our analysis, conducted using Principal Component Analysis (PCA) and represented as an ordination diagram, identified the factors that significantly influenced the occurrence and development of seedlings in the studied stands (Figure 4). The first and second ordination axes accounted for 30.6 and 16.9% of the variance, respectively. The variables with statistical significance greater than 95% are presented. As part of the PCA analysis, it was observed that the number of one-year-old, older, and the total number of seedlings were negatively correlated with the presence of ground vegetation and soil stoniness (Figure 4). It was further observed that the stand canopy was negatively correlated with the number of older seedlings and the total number of seedlings (Figure 4).
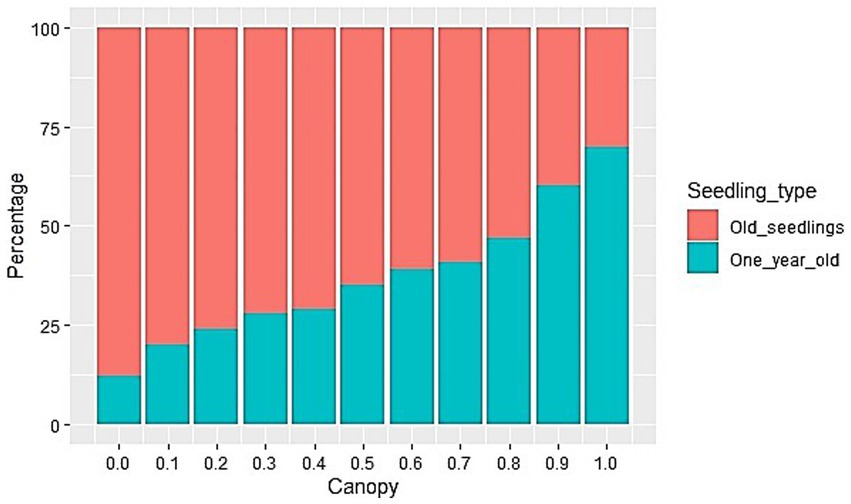
Figure 4. The ratio of the number of one-year-old and older beech seedlings under different canopy.
The results of linear mixed models indicate that the presence of a middle layer of young trees (p < 0.001), ground vegetation (p < 0.05) and stoniness (p < 0.05) significantly influenced the independent number of one-year-old and older seedlings and the total number of seedlings (Table 4). Furthermore, the analysis revealed that the canopy (p < 0.05), as well as the combined effect of the canopy and the presence of the middle layer of young trees (p < 0.05), had a significant influence on the number of older seedlings and the total number of seedlings, although this influence was not observed for one-year-old seedlings (Table 4). The obtained values of the marginal R2fix. and conditional R2tot. coefficients indicated that a significant share of variance was accounted for by fixed and random factors (Table 4).
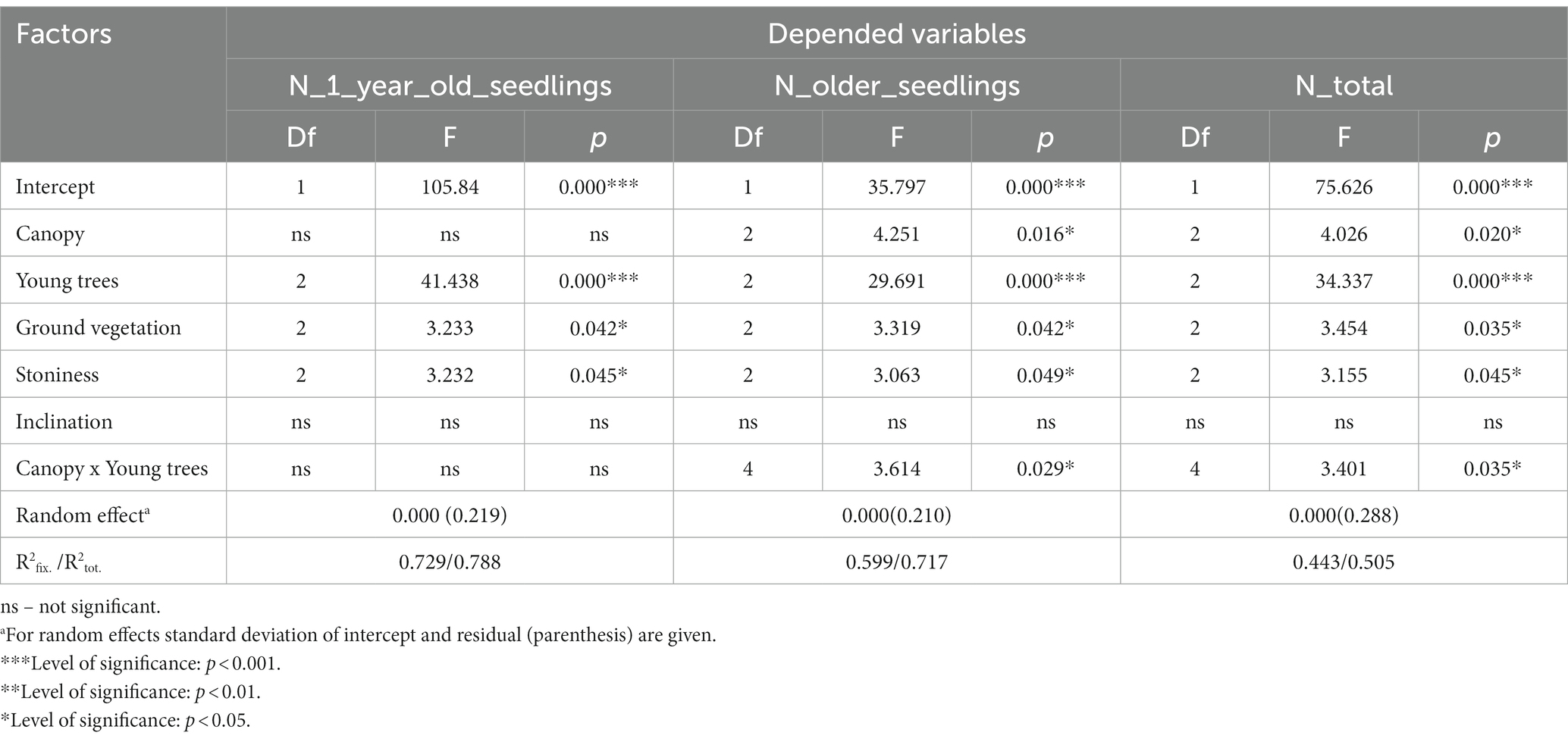
Table 4. Results of the linear mixed models for estimating the number of one-year-old, older and all seedlings in relation to the stand canopy, the presence of young trees, ground vegetation, stoninnes and inclination (fixed factors) at different study sites (random factor).
The spatial heterogeneity of the regeneration process can further be explained through the ratio between the number of one-year-old and older seedlings (Figure 5). An increase in the canopy resulted in a higher prevalence of one-year-old seedlings. In a very dense canopy, the ratio of one-year-old to older seedlings was 70:30%, while in the canopy openings, it amounted to 12:88% (Figure 5).
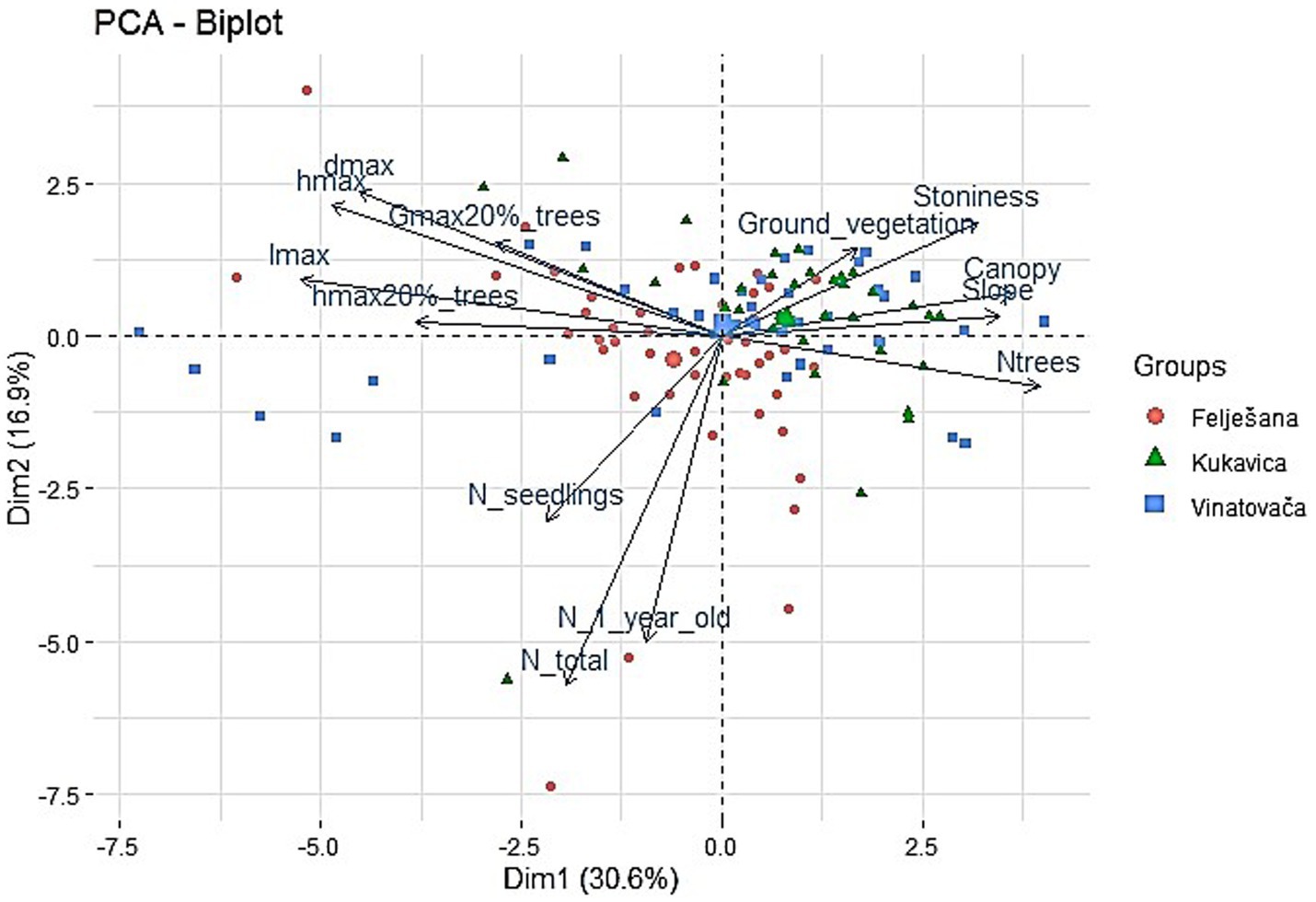
Figure 5. PCA analysis of relationships beetwen stand parameters (N_trees, Gmax20%_trees, hmax20%_trees, Canopy), regeneration layer (N_seedlings, N_1_year_old, N_total, dmax, hmax, lmax, Ground_vegetation) and research plot attributes (Slope, Stoniness).
3.3 Seedling growth
Similarly to presence, the spatial heterogeneity of the growth characteristics of beech seedlings is clearly evident in the studied stands. PCA analysis reveals a positive correlation between the main seedling growth parameters, i.e., maximum diameter at root collar (dmax), maximum height (hmax) and maximum height increment (lmax) of the seedlings and the structural stand parameters as represented by basal area (Gmax20%_trees) and the height of the largest 20% of trees (hmax20%_trees) (Figure 5). In contrast, there is a pronounced negative correlation between the same parameters of seedling growth and ecological factors (slope, stoniness), as well as structural stand parameters (canopy, number of trees) (Figure 5). Furthermore, a positive correlation is evident between the aforementioned growth parameters and the number of seedlings, observed through the total value or independent number of one-year-old and older seedlings (Figure 5).
Regarding seedling growth parameters, the results of linear mixed models revealed that canopy (p < 0.001), the presence of a middle layer of young trees (p < 0.001), stoniness (p < 0.05), slope (p < 0.05), as well as the combined effect of the canopy and the presence of a middle layer of young trees (p < 0.05) significantly affected the maximum values of the diameter at root collar (dmax), height (hmax) and height increment (lmax) of seedlings (Table 5). The obtained values of the marginal R2fix. and conditional R2tot. coefficients further indicated that a significant share of variance was accounted for by fixed and random factors (Table 5).
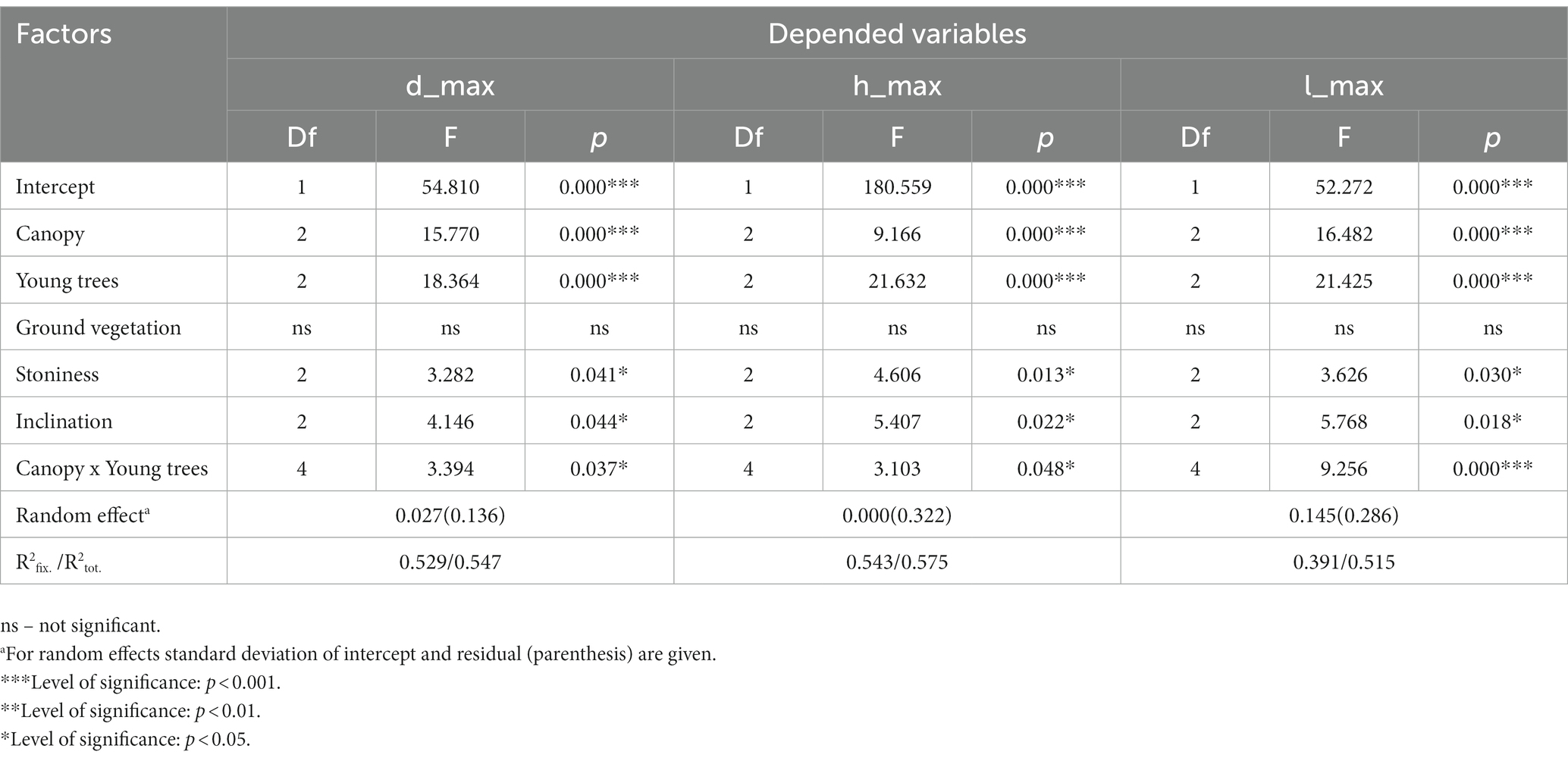
Table 5. Results of the linear mixed models for estimating the maximum root collar diameter (dmax), maximum height (hmax) and maximum height increment (lmax) in relation to the stand canopy, the presence of young trees, ground vegetation, stoninnes and inclination (fixed factors) at different study sites (random factor).
4 Discussion
4.1 Regeneration potential of old-growth beech forests
The balance and continuity of the regeneration process are some of the key features of old-growth beech forests. Compared to managed forests, this process is notably slower, yet it offers a higher level of confidence that the regeneration will be successful. Regeneration dynamics is a very important factor affecting the quality of the habitat, the provision of forest ecosystem services, as well as the capacity of forests to adapt to environmental changes (Duveneck and Scheller, 2015; Aquilué et al., 2021). Gradual changes resulting from minor or major natural disturbances or the mortality of individual or groups of trees lead to the establishment of beech seedling banks. These canopy openings provide light necessary for regeneration, although at the expense of reducing the microclimatic buffering effect of forests (Thom et al., 2023). Besides intensifying the beech regeneration, these openings also foster the emergence of herbaceous plants and shrubs that compete with tree seedlings for resources (Holeksa, 2003; Diaci et al., 2012). All these factors make the regeneration process complex and stress the remarkable capacity of beech to dominate in competitive relationships with other herbaceous and tree species. In the stands under our investigation, the strong competitive power of beech limited the participation of other tree species in the regeneration layer, which ranged from 0.5 to 5.0%.
The specificity of the old-growth beech forests lies in the prolonged capacity of beech trees to produce seeds and the frequent occurrence of mast years that sustain the continuity of the regeneration process (Evans, 1988). To eliminate variables related to the quality of beech seeds, the potential for regeneration of old-growth beech forests is best assessed through the presence of one-year-old seedlings in the stand. The results presented in this paper reveal a large number of seedlings in old-growth beech forests, as well as a very significant, often dominant, participation of one-year-old beech seedlings in the regeneration layer, ranging from 45.9 to 73.3%. Our analysis further determined that the share of one-year-old seedlings in the total number of seedlings increased with an increase in the canopy cover. Considering that beech is a shade-tolerant species, the occurrence of seedling banks under a closed canopy is a very common phenomenon (Szwagrzyk et al., 2001; Caquet et al., 2010; Orman et al., 2018). This finding is also supported by the positive correlation between small seedlings and the stand basal area (Diaci et al., 2022), as well as their negative correlation with direct light (Rozenbergar et al., 2007).
Another very important issue that has to be studied in order to gain a complete understanding of the regeneration process is seedling mortality. However, it was not within the scope of our research. The mortality rate of beech seedlings is lowest in seedling banks, where individuals have a better-developed root system and necessary resources for growth (Topoliantz and Ponge, 2000). Furthermore, the largest presence of tall beech seedlings is within seedling banks in canopy openings (Wagner et al., 2010; Orman et al., 2018; Diaci et al., 2022). Accordingly, in beech old-growth forests, a balance is established between seedling mortality and the occurrence of new seedlings, which ensures the continuity of the regeneration process, allowing beech to use any additional influx of light and suppress other herbaceous and tree species.
4.2 Key drivers affecting the spatial heterogeneity of seedling occurrence
Successful regeneration results from the joint action of multiple factors such as flowering, seed production, seed dispersal, storage, germination, seedling development, competition with other species and successful establishment of juvenile trees (Fischer et al., 2016; Axer et al., 2021). It is imperative to recognize the key drivers that influence the spatial heterogeneity of the occurrence and presence of seedlings in old-growth beech forests.
The occurrence and presence of beech seedlings are influenced by both habitat and stand characteristics (Barna and Bosela, 2015; Čurović et al., 2020; Axer et al., 2021). Our study identified and singled out a number of factors affecting the spatial heterogeneity of beech seedlings. Structural indicators such as the canopy and the presence of a middle layer of young trees strongly influenced the distribution and density of older seedlings. On the other hand, the occurrence of seedlings was strongly influenced by the presence of a middle layer of young trees, while the stand canopy was not recognized as a key factor. This finding is related to the fact that seedling occurrence can take place under varying canopy conditions (Žemaitis et al., 2019; Axer et al., 2021), while seedlings survival is highly dependent on the available amount of light (Barna and Bosela, 2015). In addition, numerous studies have pointed out a weak connection between the occurrence of seedlings and light availability in beech forests (Szwagrzyk et al., 2001; Ammer et al., 2008). Conversely, the presence of a middle layer of young trees does not provide the minimum resources necessary for the establishment of a new regeneration layer. Besides these factors, the adverse effect of the presence of ground vegetation and stoniness on the occurrence and presence of one-year-old and older seedlings was identified. Similar factors have been identified in numerous other studies (Ammer et al., 2008; Diaci et al., 2012; Vacek et al., 2017; Žemaitis et al., 2019). The most influential factors determining the total number of beech seedlings in the studied stands, regardless of their age, are the canopy and the presence of a middle layer of young trees, as well as their combined influence. In fact, this combined influence significantly reduces available light, which results in high seedling mortality (Collet et al., 2001; Szwagrzyk et al., 2001; Klopcic et al., 2012; Barna and Bosela, 2015), indirectly affecting the total number of seedlings.
4.3 Key drivers affecting the spatial heterogeneity of seedling growth
The growth characteristics of seedlings in old-growth beech forests are directly dependent on the changes in the stand canopy. These changes affect the regime of light and other microclimatic factors that govern seedling development (Feldmann et al., 2020). Our study documented the dominant presence of small seedlings, which aligns with numerous studies of old-growth beech forests in Central and Eastern Europe (Commarmot et al., 2005; Dröβler and von Lupke., 2005; Diaci et al., 2012).
Accordingly, the growth dynamics of seedlings were very slow in the investigated old-growth beech forests. Only seedlings with a height > 120 cm, whose presence ranged from 7.3 to 13.7% in the studied stands, were characterized by the height increment necessary for the establishment of stable seedling banks (Szwagrzyk et al., 2001; Feldmann et al., 2020). As with the occurrence of seedlings, their growth dynamics were influenced by habitat conditions and structural characteristics. The growth dynamics of seedlings were strongly influenced by the structural elements of stands, such as the canopy and the presence of a middle layer of young trees, also manifested through their combined influence. Similar observations are reported in numerous other studies of the regeneration dynamics of old-growth beech forests (Petritan et al., 2009; Vacek et al., 2015; Orman et al., 2018; Čurović et al., 2020). In areas where a middle layer of young trees was formed, only the presence of small, usually one-year-old individual seedlings was recorded. On the other hand, the highest presence of tall beech seedlings was noted within seedling banks formed in canopy openings, which is a typical trait of the regeneration in these forests (Wagner et al., 2010; Čurović et al., 2011; Orman et al., 2018; Diaci et al., 2022). Competitive interactions in the regeneration layer seemed to be quite limited in the old-growth beech forests included in these studies. Ground vegetation was not identified as a factor with adverse effects on the seedling growth dynamics. The insufficient amount of light in the regeneration layer often leads to the absence or mitigation of the negative impact of ground vegetation (Diaci et al., 2022). Under these conditions, beech is usually more dominant than other tree and herbaceous species. In our study, habitat factors such as slope and stoniness proved to have a negative effect on the growth dynamics of seedlings. This negative effect could be attributed to the reduced potential of the soil to retain moisture on higher slopes and greater stoniness, which also affected the growth of seedlings (Vacek et al., 2015; Liang et al., 2023).
5 Conclusion
Old-growth beech forests in Southeastern Europe are characterized by a very slow, yet balanced and continuous regeneration process, which should be viewed from three different aspects: occurrence, survival and development of seedlings. This process exhibits spatial heterogeneity governed by factors with profound effects that can sometimes be combined. Our study identified structural and habitat factors that strongly influence and dictate the regeneration dynamics of beech. In contrast, beech, known as a species of pronounced biological strength, dominates in these situations, and having a high regenerative potential leaves very small room for the establishment of other tree and herbaceous species. The growth dynamics of seedlings are limited by slow changes in the stand canopy, as they are highly dependent on the available amount of light.
These findings underscore the importance of identifying and understanding the factors that influence the spatial heterogeneity of regeneration processes in old-growth beech forests. Consequently, it is of utmost significance to understand these factors and determine their influence on the regeneration process in managed beech forests.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
BK: Conceptualization, Data curation, Investigation, Methodology, Supervision, Writing – original draft. VB: Resources, Writing – review & editing. SS: Formal analysis, Investigation, Writing – review & editing. NM: Conceptualization, Data curation, Formal analysis, Investigation, Visualization, Writing – review & editing. BP: Formal analysis, Investigation, Writing – review & editing. DF: Formal analysis, Investigation, Writing – review & editing. VČ: Investigation, Methodology, Resources, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
The authors express gratitude to the State Enterprise “Srbijašume,” Belgrade, Serbia, the Institute of Forestry, Belgrade, and the Faculty of Forestry University of Belgrade for the assistance provided during the fieldwork. This study was funded by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia, Contract No. 451-03-47/2023-01/ 200027.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ammer, C., Fichtner, A., Fischer, A., Gossner, M. M., Meyer, P., Seidl, R., et al. (2018). Key ecological research questions for central European forests. Basic Appl. Ecology 32, 3–25. doi: 10.1016/j.baae.2018.07.006
Ammer, C., Stimm, B., and Mosandl, R. (2008). Ontogenetic variation in the relative infuence of light and belowground resources on European beech seedling growth. Tree Physiol. 28, 721–728. doi: 10.1093/treephys/28.5.721
Aquilué, N., Messier, C., Martins, K. T., Dumais-Lalonde, V., and Mina, M. (2021). A simple-to-use management approach to boost adaptive capacity of forests to global uncertainty. For. Ecol. Manag. 481:118692. doi: 10.1016/j.foreco.2020.118692
Axer, M., Martens, S., Schlicht, R., and Wagner, S. (2021). Modelling natural regeneration of European beech in Saxony, Germany: identifying factors infuencing the occurrence and density of regeneration. Eur. J. For. Res. 140, 947–968. doi: 10.1007/s10342-021-01377-w
Bagnato, S., Marziliano, P. A., Sidari, M., Mallamaci, C., Marra, F., and Muscolo, A. (2021). Effects of gap size and cardinal directions on natural regeneration, growth dynamics of trees outside the gaps and soil properties in European beech forests of southern Italy. Forests 12:1563. doi: 10.3390/f12111563
Barna, M., and Bosela, M. (2015). Tree species diversity change in natural regeneration of a beech forest under different management. For. Ecol. Manag. 342, 93–102. doi: 10.1016/j.foreco.2015.01.017
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Bottero, A., Garbarino, M., Dukic, V., Govedar, Z., Lingua, E., Nagel, T. A., et al. (2011). Gap-phase dynamics in the old-growth forest of Lom. Bosnia and Herzegovina. Silva Fennica 45:76. doi: 10.14214/sf.76
Caquet, B., Montpied, P., Dreyer, E., Epron, D., and Collet, C. (2010). Response to canopy opening does not act as a filter to Fagus sylvatica and Acer sp. advance regeneration in a mixed temperate forest. Ann. For. Sci. 67:105. doi: 10.1051/forest/2009086
Collet, C., Lanter, O., and Pardos, M. (2001). Effects of canopy opening on height and diameter growth in naturally regenerated beech seedlings. Ann. For. Sci. 58, 127–134. doi: 10.1051/forest:2001112
Commarmot, B., Bachofen, H., Bundziak, Y., Burgi, A., Ramp, B., Shparyk, Y., et al. (2005). Structures of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): a comparative study. For. Snow. Landsc. Res. 79, 45–56.
Commarmot, B., and Brang, P. (2011). “Was sind Naturwälder, was Urwälder?” in Waldreservate. 50 Jahre natürliche Waldentwicklung in der Schweiz. eds. P. Brang, C. Heiri, and H. Bugmann (Bern, Switzerland: Eidg. Forschungsanstalt WSL, Birmensdorf; ETH Zürich, Zürich. Haupt, Bern, CH), 12–25.
Čurović, M., Medarević, M., Cvjetićanin, R., and Knežević, M. (2011). Major characteristics of the mixed fir and beech virgin forests in National Park Biogradska Gora in Montenegro. Bulletin of the Faculty of Forestry 103, 157–172. doi: 10.2298/GSF1103157C
Čurović, M., Spalević, V., Sestras, P., Motta, R., Dan, C., Garbarino, M., et al. (2020). Structural and ecological characteristics of mixed broadleaved old-growth forest (Biogradska Gora - Montenegro). Turk. J. Agric. For. 44, 428–438. doi: 10.3906/tar-2003-103
Diaci, J., Adamic, T., Fidej, G., and Rozenbergar, D. (2022). Toward a beech-dominated alternative stable state in Dinaric mixed montane forests: a long-term study of the Pecka old-growth Forest. Front. For. Glob. Change 5:937404. doi: 10.3389/ffgc.2022.937404
Diaci, J., Adamic, T., and Rozman, A. (2012). Gap recruitment and partitioning in an old-growth beech forest of the Dinaric Mountains: influences of light regime, herb competition and browsing. For. Ecol. Manag. 285, 20–28. doi: 10.1016/j.foreco.2012.08.010
Dröβler, L., and von Lüpke, B. (2005). Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 51, 446–457. doi: 10.17221/4578-JFS
Duveneck, M. J., and Scheller, R. M. (2015). Climate-suitable planting as a strategy for maintaining forest productivity and functional diversity. Ecol. Appl. 25, 1653–1668. doi: 10.1890/14-0738.1
Evans, J. (1988). Natural regeneration of broadleaves. Forestry commission bulletin 78 HMSO ; Her Majesty’s stationery office: London, UK
Feldmann, E., Glatthorn, J., Ammer, C., and Leuschner, C. (2020). Regeneration dynamics following the formation of understory gaps in a Slovakian beech virgin Forest. Forests 11:585. doi: 10.3390/f11050585
Fischer, H., Huth, F., Hagemann, U., and Wagner, S. (2016). “Developing restoration strategies for temperate forests using natural regeneration processes” in Restoration of boreal and temperate forests. ed. J. Stanturf (Boca Raton: CRC Press), 103–164.
Ford, S. E., and Keeton, W. S. (2017). Enhanced carbon storage through management for old-growth characteristics in northern hardwood-conifer forests. Ecosphere 8:e01721. doi: 10.1002/ecs2.1721
Glatthorn, J., Feldmann, E., Tabaku, V., Leuschner, C., and Meyer, P. (2018). Classifying development stages of primeval European beech forests: is clustering a useful tool? BMC Ecol. 18:47. doi: 10.1186/s12898-018-0203-y
Govedar, Z., Krstić, M., Keren, S., Babić, V., Zlokapa, B., and Kanjevac, B. (2018). Actual and balanced stand structure: examples from beech-fir-spruce old-growth forests in the area of the Dinarides in Bosnia and Herzegovina. Sustainability 10:540. doi: 10.3390/su10020540
Govedar, Z., and Stanivuković, Z. (2008). Natural regeneration of beech in mixed stands of beech and fir with spruce (Piceo-Abieti-Fagetum) in the virgin forest “Perućica”. Bulletin of the Faculty of Forestry 97, 107–126. doi: 10.2298/GSF0897107G
Hobi, M. L., Commarmot, B., and Bugmann, H. (2015a). Pattern and process in the largest primeval beech forest of Europe (Ukrainian Carpathians). J. Veg. Sci. 26, 323–336. doi: 10.1111/jvs.12234
Hobi, M. L., Ginzler, C., Commarmot, B., and Bugmann, H. (2015b). Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing. Ecosphere 6, 1–15. doi: 10.1890/ES14-00390.1
Holeksa, J. (2003). Relationship between field-layer vegetation and canopy openings in a Carpathian subalpine spruce forests. Plant Ecol. 168, 57–67. doi: 10.1023/A:1024457303815
Jaloviar, P., Sedmáková, D., Pittner, J., Jarčušková Danková, L., Kucbel, S., Sedmák, R., et al. (2020). Gap structure and regeneration in the mixed old-growth forests of National Nature Reserve Sitno. Slovakia. Forests 11:81. doi: 10.3390/f11010081
Jucker, T., Bouriaud, O., Avacaritei, D., and Coomes, D. A. (2014). Stabilizing effects of diversity on aboveground wood production in forest ecosystems: linking patterns and processes. Ecol. Lett. 17, 1560–1569. doi: 10.1111/ele.12382
Kassambara, A., and Mundt, F. (2020). Factoextra: extract and visualize the results of multivariate data analyses. R package version 1.0.7. Available at: https://CRAN.R-project.org/package=factoextra
Keren, S., Diaci, J., Motta, R., and Govedar, Z. (2017). Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina. For. Ecol. Manag. 400, 531–541. doi: 10.1016/j.foreco.2017.06.009
Klopcic, M., Poljanec, A., and Boncina, A. (2012). Modelling natural recruitment of European beech (Fagus sylvatica L.). For. Ecol. Manag. 284, 142–151. doi: 10.1016/j.foreco.2012.07.049
Lang, R. (1915). Versuch einer Exakten Klassifikation der Böden in Klimatischer und Geologischer Hinsicht. Internationale Mitteilungen für Bodenkunde 5, 312–346.
Liang, W., Feng, Y., Li, F., Zhao, Y., Zhu, J., Yao, J., et al. (2023). Drivers of vegetation and soil determine natural regeneration of a single plantation at different slope positions. Front. For. Glob. Change 6:1167284. doi: 10.3389/ffgc.2023.1167284
Madsen, P., and Hahn, K. (2008). Natural regeneration in a beech-dominated forest managed by close-to-nature principles – a gap cutting based experiment. Can. J. For. Res. 38, 1716–1729. doi: 10.1139/X08-026
Massad, T. J., Williams, G. L., Wilson, M., Hulsey, C. E., Deery, E., and Bridges, L. E. (2019). Regeneration dynamics in old-growth urban forest gaps. Urban For. Urban Green. 43:126364. doi: 10.1016/j.ufug.2019.06.007
Modry, M., Hubeny, D., and Rejsek, K. (2004). Differential response of naturally regenerated European shade tolerant tree species to soil type and light availability. For. Ecol. Manag. 188, 185–195. doi: 10.1016/j.foreco.2003.07.029
Motta, R., Berretti, R., Castagneri, D., Dukić, V., Garbarino, M., Govedar, Z., et al. (2011). Toward a defnition of the range of variability of central European mixed Fagus–Abies–Picea forests: the nearly steady-state forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 41, 1871–1884. doi: 10.1139/X11-098
Orman, O., Dobrowolska, D., and Szwagrzyk, J. (2018). Gap regeneration patterns in Carpathian old-growth mixed beech forests – interactive effects of spruce bark beetle canopy disturbance and deer herbivory. For. Ecol. Manag. 430, 451–459. doi: 10.1016/j.foreco.2018.08.031
Petritan, A. M., von Lüpke, B., and Petritan, I. C. (2009). Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 128, 61–74. doi: 10.1007/s10342-008-0239-1
Petrovska, R., Bugmann, H., Hobi, M. L., and Brang, P. (2023). Replace me if you can: abundance of advance regeneration under canopy trees in a primeval beech forest. For. Ecol. Manag. 537:120939. doi: 10.1016/j.foreco.2023.120939
Petrovska, R., Bugmann, H., Hobi, M. L., Ghosh, S., and Brang, P. (2022). Survival time and mortality rate of regeneration in the deep shade of a primeval beech forest. Eur. J. For. Res. 141, 43–58. doi: 10.1007/s10342-021-01427-3
R Core Team (2023). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rozenbergar, D., Mikac, S., Anić, I., and Diaci, J. (2007). Gap regeneration patterns in relationship to light heterogeneity in two old-growth beech – fir forest reserves in south East Europe. Forestry 80, 431–443. doi: 10.1093/forestry/cpm037
Rozendaal, D. M. A., Bongers, F., Aide, T. M., Alvarez-Dávila, E., Ascarrunz, N., Balvanera, P., et al. (2019). Biodiversity recovery of Neotropical secondary forests. Sci. Adv. 5:eaau3114. doi: 10.1126/sciadv.aau3114
Rugani, T., Diaci, J., and Hladnik, D. (2013). Gap dynamics and structure of two old-growth beech Forest remnants in Slovenia. PLoS One 8:e52641. doi: 10.1371/journal.pone.0052641
Schröter, M., Härdtle, W., and von Oheimb, G. (2012). Crown plasticity and neighborhood interactions of European beech (Fagus sylvatica L.) in an old-growth forest. Eur. J. For. Res. 131, 787–798. doi: 10.1007/s10342-011-0552-y
Siminski, A., Zambiazi, D. C., dos Santos, K. L., and Fantini, A. C. (2021). Dynamics of natural regeneration: implications for landscape restoration in the Atlantic Forest. Brazil. Front. For. Glob. Change 4:576908. doi: 10.3389/ffgc.2021.576908
Szewczyk, J., and Szwagrzyk, J. (2010). Spatial and temporal variability of natural regeneration in a temperate old-growth forest. Ann. For. Sci. 67:202. doi: 10.1051/forest/2009095
Szwagrzyk, J., and Szewczyk, J. (2008). Is natural regeneration of forest stands a continuous process? A case study of an old-growth forest of the western Carpathians. Pol. J. Ecol. 56, 623–633.
Szwagrzyk, J., Szewczyk, J., and Bodziarczyk, J. (2001). Dynamics of seedling banks in beech forest: results of a 10-year study on germination, growth and survival. For. Ecol. Manag. 141, 237–250. doi: 10.1016/S0378-1127(00)00332-7
Thom, D., Ammer, C., Annighöfer, P., Aszalós, R., Dittrich, S., Hagge, J., et al. (2023). Regeneration in European beech forests after drought: the efects of microclimate, deadwood and browsing. Eur. J. For. Res. 142, 259–273. doi: 10.1007/s10342-022-01520-1
Thornthwaite, C. W. (1948). An approach toward a rational classification of climate. Geogr. Rev. 38, 55–94. doi: 10.2307/210739
Topoliantz, S., and Ponge, J.-F. (2000). Influence of site conditions on the survival of Fagus sylvatica seedlings in an old-growth beech forest. J. Veg. Sci. 11, 369–374. doi: 10.2307/3236629
Vacek, Z., Bulušek, D., Vacek, S., Hejcmanová, P., Remeš, J., Bílek, L., et al. (2017). Effect of microrelief and vegetation cover on natural regeneration in European beech forests in Krkonoše national parks (Czech Republic, Poland). Austrian J. For. Sci. 134, 75–96.
Vacek, Z., Vacek, S., Podrázský, V., Bílek, L., Štefančík, I., Moser, W. K., et al. (2015). Effect of tree layer and microsite on the variability of natural regeneration in autochthonous beech forests. Pol. J. Ecol. 63, 233–246. doi: 10.3161/15052249PJE2015.63.2.007
Venables, W. N., and Ripley, B.D. (2002). Modern applied statistics with S, 4th. Springer, New York.
Wagner, S., Collet, C., Madsen, P., Nakashizuka, T., Nyland, R. D., and Sagheb-Talebi, K. (2010). Beech regeneration research: from ecological to silvicultural aspects. For. Ecol. Manag. 259, 2172–2182. doi: 10.1016/j.foreco.2010.02.029
West, B.T., Welch, K.B., and Galecki, A.T. (2007). Linear mixed models: A practical guide using statistical software. Chapman Hall/CRC Press, Boca Raton.
Wirth, C., Gleixner, G., and Heimann, M. (2009). Old-growth forests: Functions, fate and value. Ecological Studies 207. Springer, New York
Žemaitis, P., Gil, W., and Borowski, Z. (2019). Importance of stand structure and neighborhood in European beech regeneration. For. Ecol. Manag. 448, 57–66. doi: 10.1016/j.foreco.2019.05.066
Keywords: old-growth, beech, regeneration process, seedlings, spatial heterogeneity, Serbia, southeastern Europe
Citation: Kanjevac B, Babić V, Stajić S, Martać N, Pavlović B, Furtula D and Čokeša V (2023) Key drivers affecting the spatial heterogeneity of the regeneration process in old-growth beech forests in southeastern Europe. Front. For. Glob. Change. 6:1304037. doi: 10.3389/ffgc.2023.1304037
Edited by:
Zoran Govedar, University of Banja Luka, Bosnia and HerzegovinaReviewed by:
Admir Avdagić, University of Sarajevo, Bosnia and HerzegovinaMilic Curovic, University of Montenegro, Montenegro
Copyright © 2023 Kanjevac, Babić, Stajić, Martać, Pavlović, Furtula and Čokeša. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Branko Kanjevac, YnJhbmtvLmthbmpldmFjQHNmYi5iZy5hYy5ycw==