 Renzo Motta1*
Renzo Motta1* Matteo Garbarino1Roberta Berretti1Alessia Bono1
Matteo Garbarino1Roberta Berretti1Alessia Bono1 Milic Curovic2
Milic Curovic2 Vojislav Dukić3Paola Nola4
Vojislav Dukić3Paola Nola4- 1Department of Agricultural, Forest and Food Sciences, University of Torino, Grugliasco, Italy
- 2Biotechnical Faculty, University of Montenegro, Podgorica, Montenegro
- 3Faculty of Forestry, University of Banja Luka, Banja Luka, Bosnia and Herzegovina
- 4Department of Earth and Environmental Sciences, University of Pavia, Pavia, Italy
Species-rich mixed silver fir (Abies alba Mill.) forests dominated in the northern Apennines, but climate and, mainly, anthropogenic land use provoked a sharp silver fir decline approximately 5000 years bp. The conservation of the silver fir in this region was mainly due to the establishment of monastic orders that preserved and even planted silver fir for its spiritual and economic value. In 1993, the best silver fir stands were included in the Parco Nazionale delle Foreste Casentinesi (FCNP), Monte Falterona e Campigna, and have been submitted to low-intensive management or strict protection regardless of past land use and cultural history. In this study, we have (1) analyzed the current structure of three silver fir forests that have had different ownership histories and (2) compared the structure of the three Italian forests among them and with two old-growth forests from the Dinaric Alps as a reference of naturalness. The results show that the current structures of the three Italian forests are very different among them and are strictly related to past land use and, mainly, to monastic legacies. Even if the Italian forests have experienced decades of low-intensity management or strict protection, they are currently structurally very different from Dinaric old-growth forests. Developing an old-growth structure in these forests can be very slow and, in some ways, unpredictable. The results also highlight the importance of recognizing protected areas as cultural landscapes that host an important biocultural diversity. The current risk is that by applying almost exclusive biodiversity-centered management and setting difficult or impossible-to-achieve biodiversity goals, total diversity will decrease, and biocultural diversity, the greatest richness of most European parks, will be lost.
1 Introduction
The Apennines are characterized by high forest cover and high regional tree species diversity and provide essential ecosystem services to millions of people. However, past anthropogenic land use has profoundly affected forests of the Apennine region (Vacchiano et al., 2017) and outweighed by far the effects of macroecological constraints in the Holocene (Henne et al., 2012; Brown et al., 2013).
Species-rich mixed silver fir (Abies alba Mill.) forests dominated in the northern Apennines approximately 10,500–5,500 years bp under warmer-than-today conditions. Cooler and moister summers and increased fire activity caused a decline in fire-sensitive species (e.g., silver fir). They favored the establishment of beech (Fagus sylvatica L.) forests approximately 5,500 years ago (Tinner et al., 2013). For 5,000–2000 years bp, beech continued its expansion in this region at the expense of fir under increasing anthropogenic land use, which likely involved an intense use of fire (Morales-Molino et al., 2021). Later, the intensity of land-use changes increased, such as converting forests into chestnut (Castanea sativa Mill.) orchards (Conedera et al., 2004). Fire was no longer used systematically to clear openings. Turkey oaks (Quercus cerris L.) and chestnuts were necessary to produce fodder and nuts for livestock raising. The production of chestnut flour was fundamental for the feeding of farmers, while chestnut coppice was cultivated to make poles for construction and vine growing and beech was used for firewood (Agnoletti and Paci, 2001; Agnoletti, 2018). Beech also took advantage of its lower palatability and sensitivity to fire and browsing compared to other co-existing trees to increase its dominance (Wick and Möhl, 2006).
The conservation of silver fir in this region was mainly related to “monastic silviculture” (Agnoletti and Paci, 2001; Gabbrielli, 2006). Different religious orders, such as Camaldolese, Vallombrosians, and Franciscans, settled around the XI–XIII centuries in the northern Apennines and managed the forests for centuries (Frascaroli, 2013). The monks had great estates where they could differentiate products (crops, firewood, and timber). Besides, they preserved and even planted silver fir for its spiritual and economic value (Romano and Urbinati, 2012). Inspired by spiritual principles and applying traditional ecological knowledge, monastic communities developed distinctive natural resource management models, resulting in peculiar landscapes rich in both biological and cultural diversity (Agnoletti and Rotherham, 2015; Zannini et al., 2021).
Intensive forest utilization occurred from medieval times until the first half of the 20th century. In the last decades, socioeconomic changes caused the abandonment of traditional farming or grazing activities in mountain settlements and the re-establishment and increment of forest cover (Frascaroli, 2013; Marini Govigli et al., 2021; Agnoletti et al., 2022). Since 1993, most of the pure or mixed silver fir forests in the northern Apennines have been included in the Parco Nazionale delle Foreste Casentinesi (FCNP), Monte Falterona e Campigna, thereby creating both positive synergies and new challenges for the conservation of their naturalistic value and the historical, cultural, and spiritual legacies. However, following the FCNP establishment, most pure and mixed silver fir stands have been submitted to low-intensive management or strict protection regardless of past land use and cultural history.
According to the Italian Forest Inventory, in the central and northern Apennines, more than 8,000 ha are covered by monospecific and monolayered silver fir forests, and more than 23,000 ha of forests are covered by mixed forests with conifers and broadleaves.
This study is focused on three sites, the forests of Camaldoli, La Verna, and Sasso Fratino, which have had different land-use and land-ownership histories. We selected two forests of the Dinaric Alps, the forests of Perućica (BiH) and Biogradska Gora (MNE), as old-growth reference forests belonging to the same forest type (Sabatini et al., 2018).
The specific objectives of this study are as follows:
- to analyze the role of monastic rules and the spiritual attitude of religious orders in shaping the forest structure and composition of the current silver fir forests of the northern Italian Apennines.
- to compare the current structure of these forests against old-growth forests of the Dinaric Alps.
- to discuss the current forest dynamics and the effectiveness of present management policies on conserving the cultural heritage and restoring biodiversity.
2 Materials and methods
2.1 Study areas
We conducted this study in three forests of the FCNP established in 1993, containing some of the best remnants of silver fir in the northern Apennines. The forests of Camaldoli (CAM), La Verna (VER), and Sasso Fratino (SFR) are closely located and have similar site characteristics (Table 1; Figure 1). SFR has been strictly protected since 1959 (the core area where our study area is located). In recent decades, CAM and VER have had no harvesting and locally low-intensity forest tending. Study areas that were relatively structurally uniform were selected in each forest based on their representativity and conservation status.

Table 1. Description of the study sites.
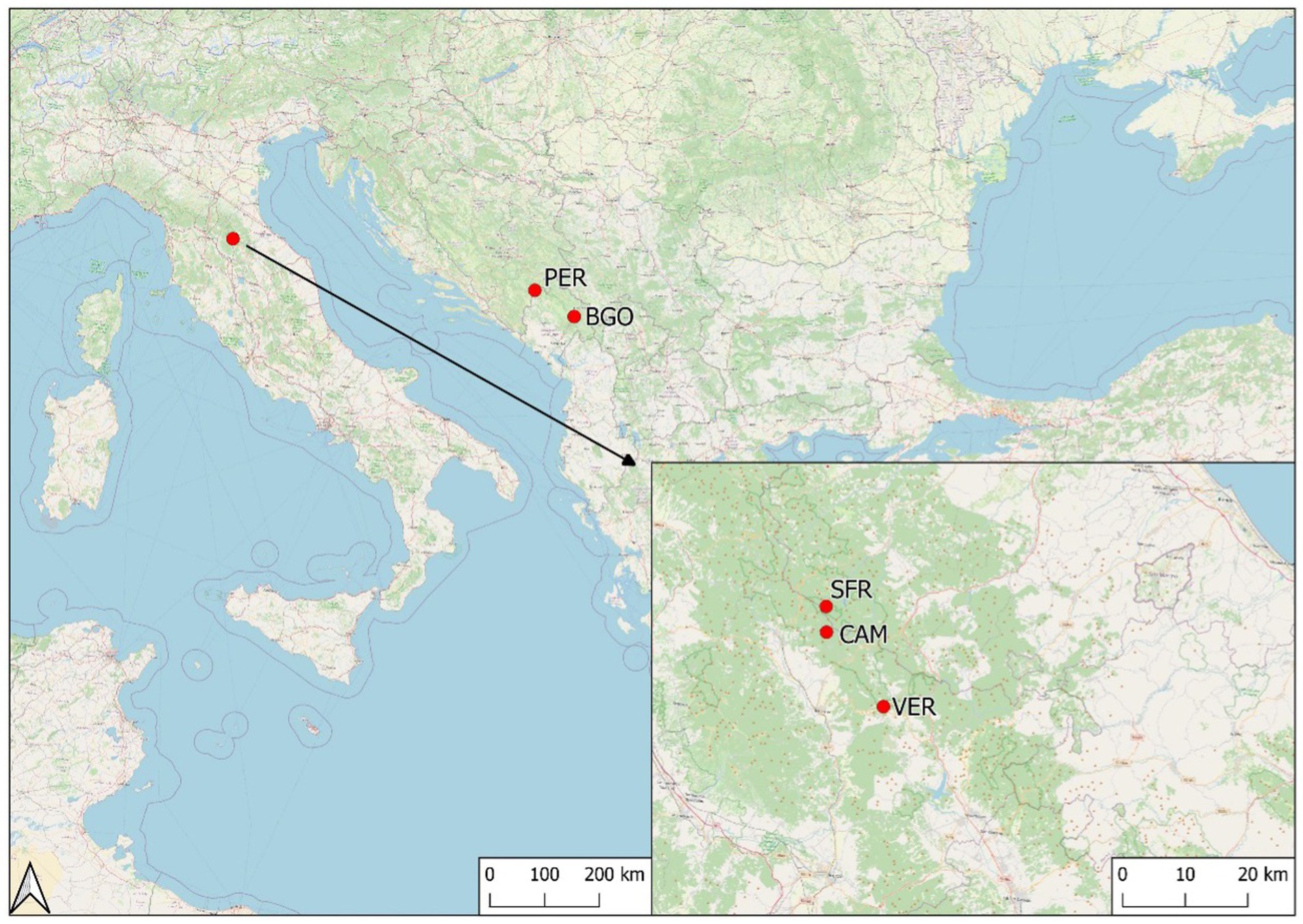
Figure 1. Location of the study sites.
Since, in the Apennines, there are no more primeval old-growth forests (OGF), we selected the forests of Perućica (PER) and Biogradska Gora, which belong to the Abieti-Fagetum forest type, as a reference for mixed, uneven-aged montane forests. Both PER and BGO among the largest and best-preserved silver fir–beech mixed old-growth forests of the Dinaric Alps (Motta et al., 2015a; Sabatini et al., 2018; Curovic et al., 2020). These forests have been used as a reference for managed stands since the second half of the 20th century (Susmel, 1956; Leibundgut, 1960), but their ecological value has been recognized since the end of the 19th century. The first official “protection” measures concerning these forests date back to 1893 for Perućica and 1878 for Biogradska Gora (Motta et al., 2015a; Stupar and Milanović, 2017). In both cases, the primary protection goal was hunting and game protection. The forest of Perućica was proclaimed a national park in 1962, while Biogradska Gora was declared a national park in 1952. However, even if Perućica and Biogradska Gora are among the most significant and best-preserved old-growth forest relicts in the whole of south-eastern Europe, both forests are surrounded by grasslands and meadows, as well as pure beech stands and forests that have been and are intensively managed. According to paleo-ecological data, during the middle ages, the land around the Biogradska forest was used for cereal crop cultivation and cattle herding, and later local, higher severity catchment fires occurred (Cagliero et al., 2022). This evidence supports the notion that historical land-use pressure reduced the extent of old-growth forests, and some sporadic anthropogenic disturbances could have affected all the European forest stands (Finsinger et al., 2022). On the other hand, our reference areas have developed naturally and have experienced only small-scale disturbances typical of old-growth and late seral stages of this forest type at least for the last three to four centuries (Nagel and Svoboda, 2008; Bottero et al., 2011; Motta et al., 2011).
2.2 Field methods
2.2.1 Forest structure
After extensive reconnaissance, we selected 29–50 ha study areas in each forest that were relatively structurally uniform and representative of the current forest structure. Inside each area, a regular grid (100 m for CAM, VER, SFR, and PER, and 120 m for BGO) was superimposed on the 1:10.000 raster map, resulting in a variable number of sampling plots ranging from 29 to 37.
At each plot, four types of measurements were applied (Castagneri et al., 2010): (a1) in circular plots of 14 m (CAM, VER, and SFR) or 12 m of radius (PER and BGO), we recorded species type, diameter at breast height (dbh, to the nearest 0.01 m) and height (to the closest 0.5 m) for all living trees; (a2) in a 113.1-m2 round plot (radius = 6 m), we recorded species type and height of each regeneration individual (h > 10 cm and dbh < 7.5 cm); (a3) on two 50-m lines that intersect and were oriented northward and eastward from the center of the sampling point, we measured the logs crossing the line (Van Wagner, 1968); and (a4) in a 50 × 4 m rectangular plot centered on the previous line, we measured stumps (diameter at the ground and the top) and snags (dbh). The size of the plots was the result of reliability and agreement with already established local systems (Lombardi et al., 2015). For each element of coarse woody debris (CWD), the species (when possible) and decay class were recorded (Motta et al., 2006). The volume for living trees and snags was calculated according to local volume tables. The volume for snags, logs, and stumps was calculated according to methods described by Motta et al. (2006). Aboveground and CWD carbon stocks were calculated using direct measurements, for stems and CWD, and using IPCC allometric algorithms (IPCC, 2006).
2.2.2 Tree cores
We collected at dbh two cores from 10 to 20 trees of each species in each study area to determine the tree age and growth patterns of the dominant trees. The cores were collected in three species (beech, silver fir, and Norway spruce) in the Dinaric sites, two species (beech and silver fir) in SFR and VER, and only one (silver fir) in CAM where beech is currently related in the intermediate-suppressed layer. All cores were fixed to wooden supports in the laboratory and prepared with a razor until an optimal surface resolution was obtained. Tree ring widths were measured to the nearest 0.01 mm using the LINTAB device and TSAP-Win software package (Rinntech, Heidelberg, Germany). All the tree-ring series were cross-dated, both visually and statistically, by CooRecorder software (Larsson and Larsson, 2018). For age estimation, in the cores, including the pith, tree age was taken as the number of rings between the pith and the cambium (age at the coring height). The pith location was estimated graphically for cores that fell short of the pith (Motta and Nola, 2001). Once the pith location had been estimated, the missing radius was estimated starting from the innermost ring. Rings taken from the innermost part of the core were then counted until a segment equal in length to the estimated missing radius was obtained; this number was added to the number of rings in the core to obtain the estimated tree age at the coring height. When it was impossible to estimate the age, we indicated the number of rings counted (e.g.,> 480 years). Given the uncertainty from estimation, age was represented for 10-year classes (Payette et al., 1990).
To describe past disturbance events, we analyzed the growth pattern to identify releases from suppression using a running mean radial-averaging method (Rubino and McCarthy, 2004; Fraver and White, 2005). The detection of growth releases was obtained using Jolt software (version 6.01P by R.L. Holmes, University of Arizona, Tucson, Arizona), taking into account abrupt growth increment over 166% of the previous years in a window of 10 years. Only the growth changes during at least 4 years were classified as releases from suppression (Schweingruber et al., 1990). A chronology of all releases found in the dominant trees of all species analyzed for each site was constructed. Release data were summarized as the percentage of trees in a given decade showing a growth release (Motta et al., 1999).
2.2.3 Multivariate ordination analyses
Classical forest structure attributes for living trees (basal area, density, volume, mean diameter, the standard deviation of diameter, and stature), dead wood (volume of stumps, logs, snags, overall CWD, and density of snags having dbh > 47.5 cm), and regeneration (density of saplings divided by species) were used to assess the between-forest variability and the between-plot variability. Other between-forest parameters were the species composition calculated as a proportion of a particular species on a broad basal area and the diameter distribution.
Between-plot variability was explored by using a multivariate approach. Two principal components analyses (PCAs) were performed to investigate the correlation structure among living trees, regeneration, and CWD attributes. The first PCA was performed on the Italian study areas only, and the second one was performed to compare the forest structure of Italian vs. Dinaric study areas. PCAs were performed using PC-ORD 7.10 (McCune & Mefford 1999) statistical packages, and their statistical significance was tested by the Monte Carlo permutation method based on 10,000 runs with randomized data.
3 Results
3.1 Forest structure
The three Italian forests have very distinct forest structures (Table 2). The CAM volume of living trees is the highest observed in the Italian study areas, reaching 929 m3 ha−1. The high current volume originated from dense artificial regeneration, the cessation of thinning after the FCNP establishment (1993), and the consequent overstocking typical of the late pole stage (Oliver and Larson, 1996). The diameter distribution of the study area simulates a negative exponential typical of multilayered forests (Figure 2). However, it results from the coexistence of monolayered dominant silver fir stands, with a normal distribution and a dense intermediate-suppressed beech layer that has progressively been artificially established in recent decades. Beech accounts for 55% of the trees but only 10% of the basal area. Dominant trees are smaller (average of the 10 largest trees ha−1: 77 cm) than in VER and SFR because of past intensive management. The volume of CWD (57 m3 ha−1) is mainly a consequence of the endogenous competition mortality observed in the last 50 years (only two snags and 1 log ha−1 with diameter > 50 cm) after the cessation of thinning. The regeneration density (287 individuals ha−1) is low.
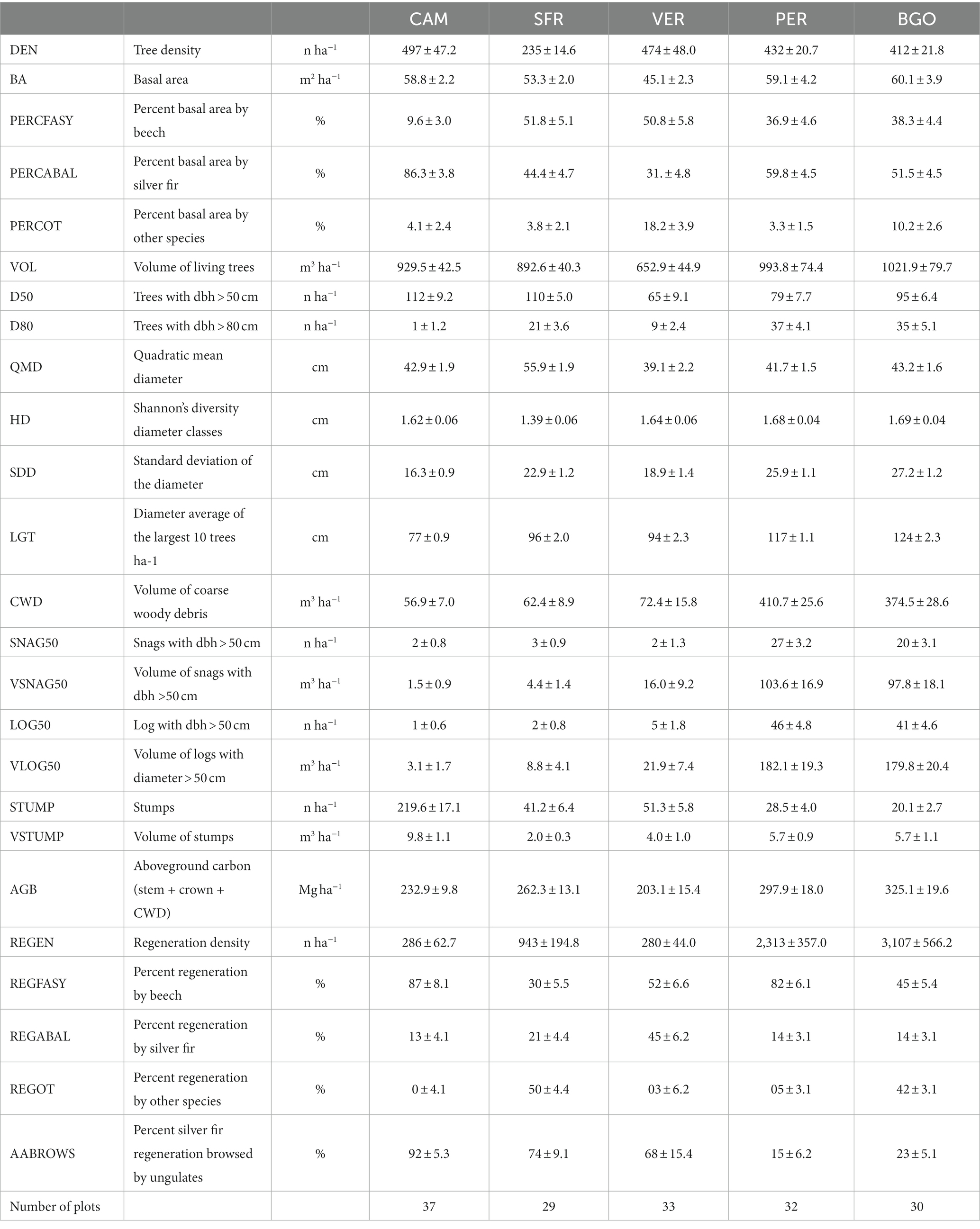
Table 2. Structural characteristics of the study areas.
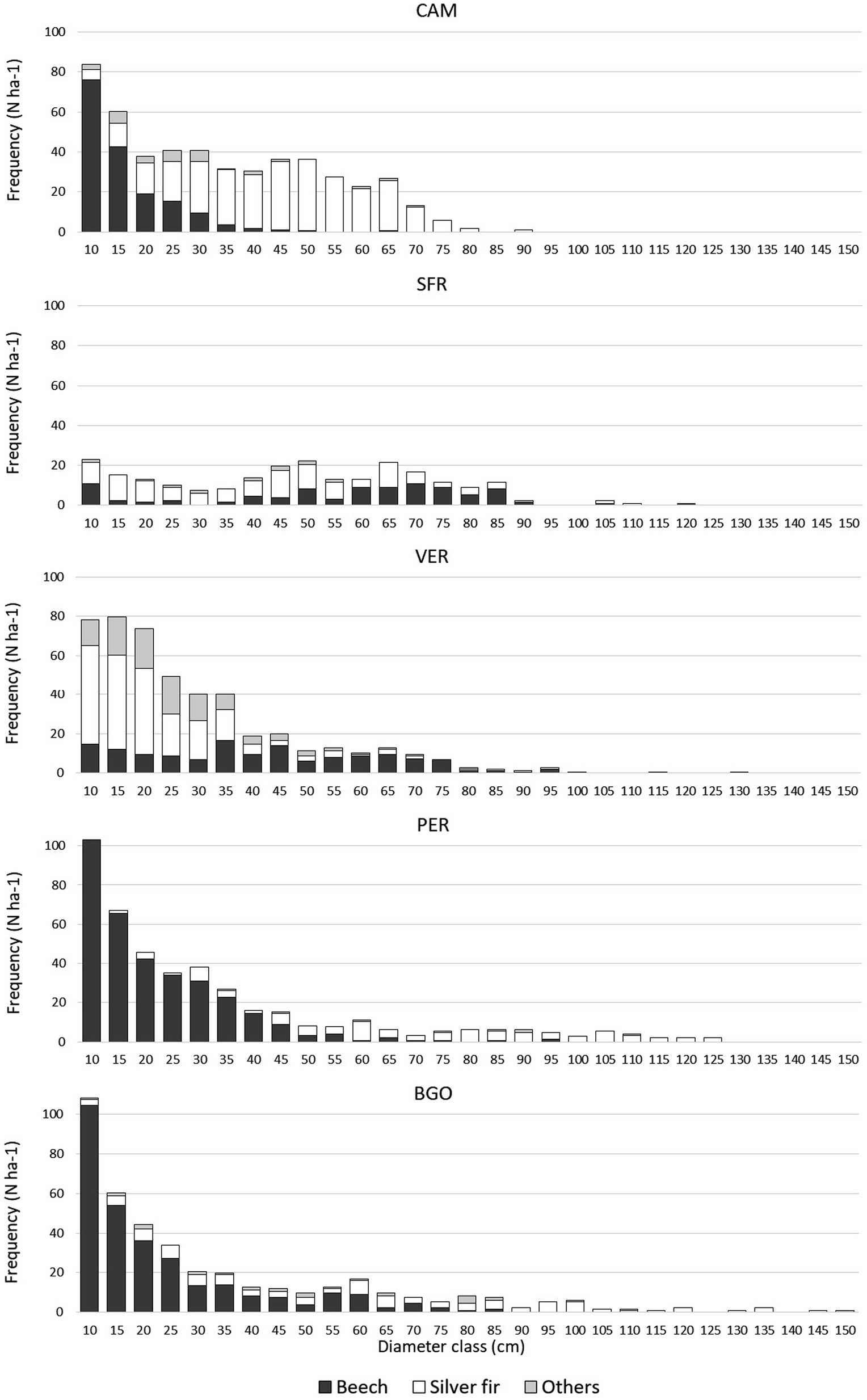
Figure 2. Diameter distribution in the five study areas.
The SFR current volume of living trees is 893 m3 ha−1. The irregular diameter distribution shows a relatively flat shape due to the overlap between past sporadic but intense cutting and recent strict protection. Beech accounts for 52% of the basal area. Dominant trees are among the largest that we can observe in Apennine silver fir–beech forests, reaching 96 cm (average of the 10 largest tree ha−1). The volume of CWD (62 m3 ha−1), despite the relatively long period of strict protection, is lower than the volume in VER. This is because the study area is in the transition stage (Oliver and Larson, 1996); most of the CWD input is still due to endogenous competition and mortality has not yet affected dominant trees (3 snags and 2 logs per ha−1 > 50 cm diameter). Besides, the slope (closer to 100% and much higher than VER and CAM) reduces CWD accumulation in situ. The regeneration density (943 individuals ha−1) in SFR is higher than that in CAM and VER.
The VER current volume of living trees is 653 m3 ha−1. The diameter distribution shows a negative exponential shape typical of selection-managed multilayered forests. Beech accounts for 51% of the basal area. Dominant trees are larger than typical managed mixed silver fir–beech forests, reaching 94 cm (average of the 10 largest tree ha−1). The volume of CWD (72 m3 ha−1) is the highest observed in the Apennines. This record is because the mortality has affected dominant trees (2 snags and 5 logs per ha−1 > 50 cm diameter), which is the result of both endogenous and exogenous sporadic windthrow mortality (Miozzo and Borchi, 2009). The regeneration density (280 individuals ha−1) in VER is low.
The regeneration of the three Italian study areas is relatively low both because they are characterized by density/crown cover characteristics of the late pole stage (CAM) or the beginning of the transition stage (SFR and VER) where mortality of dominant trees and creation of regeneration gaps is low, and because all the study areas are strongly affected by ungulate browsing. The incidence of browsing of the silver fir, which is the most browse-sensitive species, ranges from 68 to 92%, compromising the future processes of regeneration establishment. The regeneration is higher in SFR because the study area is steeper on the northern side of the Apennines, has a higher incidence of safe sites (Harper, 1977), and is less favorable for the ungulate wintering.
The two Dinaric study areas have similar forest structures and show relevant differences from the Italian ones.
The volume of living trees of Dinaric OGF is 994 m3 ha−1 in PER and 1,021 in BGO. The diameter distribution is a rotated sigmoid typical of the old-growth stage (Motta et al., 2015b). Beech accounts for 37% (PER) and 38% (BGO) of the basal area. Dominant trees reach a relevant size and an average size (10 largest tree ha−1) of 117 cm in PER and 124 cm in BGO. The volume of CWD in the Dinaric study areas is much higher than that in the Italian study areas, with the values being 411 m3 ha−1 in PER and 375 in BGO. The difference in the CWD quantity can be explained mainly by the critical incidence of large snags (respectively, 103 ha−1 and 97 with a diameter of >50 cm in PER and BGO) and logs (respectively, 46 ha−1 and 41 with a diameter of >50 cm in PER and BGO). The Shannon diversity index and diameter standard deviation are higher in the Dinaric study areas than in the Italian ones.
The regeneration density is also much higher in the Dinaric study areas than in the Italian study areas (2,313 individuals ha−1 in PER and 3,107 individuals in BGO), showing the continuity of the recruitment in the forest dynamics coherent from the old-growth stage characterized by continuous minor scale disturbances. Besides, the regeneration is less affected by ungulate browsing (15% browsed silver firs in PER and 23% in BGO). The carbon stock in the Dinaric forests (aboveground biomass and CWD) is 25% higher in the Dinaric study areas than in the Italian ones.
3.2 Tree cores
The maximum ages reached by the dominant trees range from 164 (CAM) to 303 (SFR) years in the Italian sites (Table 3). The age in the Dinaric sites is much older, reaching more than 480 years in both sites. In these forests, which represent two of the best-preserved old-growth forests in southern Europe, we can hypothesize that the oldest trees are near their maximum biological age.
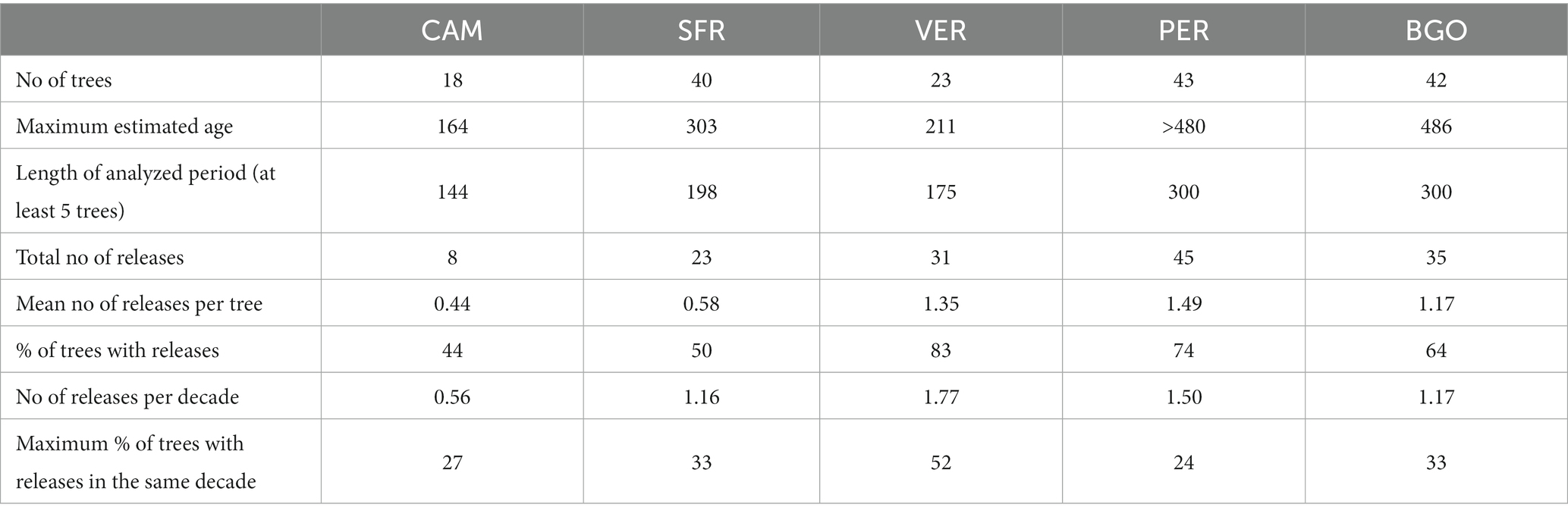
Table 3. Maximum age and disturbance (releases from suppression) regime in the five study areas according to dendrochronological analysis.
For the disturbance history detected from abrupt growth release (Figure 3), we have fixed a period of three centuries for the Dinaric sites, while in the Apennines, the period analyzed depends on the availability of old dominant trees and was 160 years for CAM, 180 years for VER, and 200 years for SFR. Most of the release from suppression detected in the Italian sites were legacies of the past management (Müllerová et al., 2016; Muigg et al., 2020). The only exception was the release peak observed in VER, which was due to a large windthrow in 1943 (Miozzo and Borchi, 2009). In the Italian sites, there was a peak in the releases’ incidence in 1930 in CAM, 1950 in VER, and 1830 in SFR. CAM is relatively young and has a low incidence of release. At the same time, VER has been managed until recently with a selection system and, therefore, shows high release incidence in the decades of intensive management. In CAM and VER, the release distribution is irregular. At the same time, in SFR, after the major event detected in 1830, the frequency of disturbances is relatively low and regular, reflecting the recent shift from management disturbances to natural processes. In the Dinaric sites, the growth releases occurred quite regularly throughout the history of the stand. There was only one decade in in PER (1880) and BGO (1750) when the incidence of the releases was higher than 20%. The disturbance history is coherent with a small-scale continuous disturbance regime typical of the old-growth stage (Motta et al., 2011; Fraver et al., 2020).
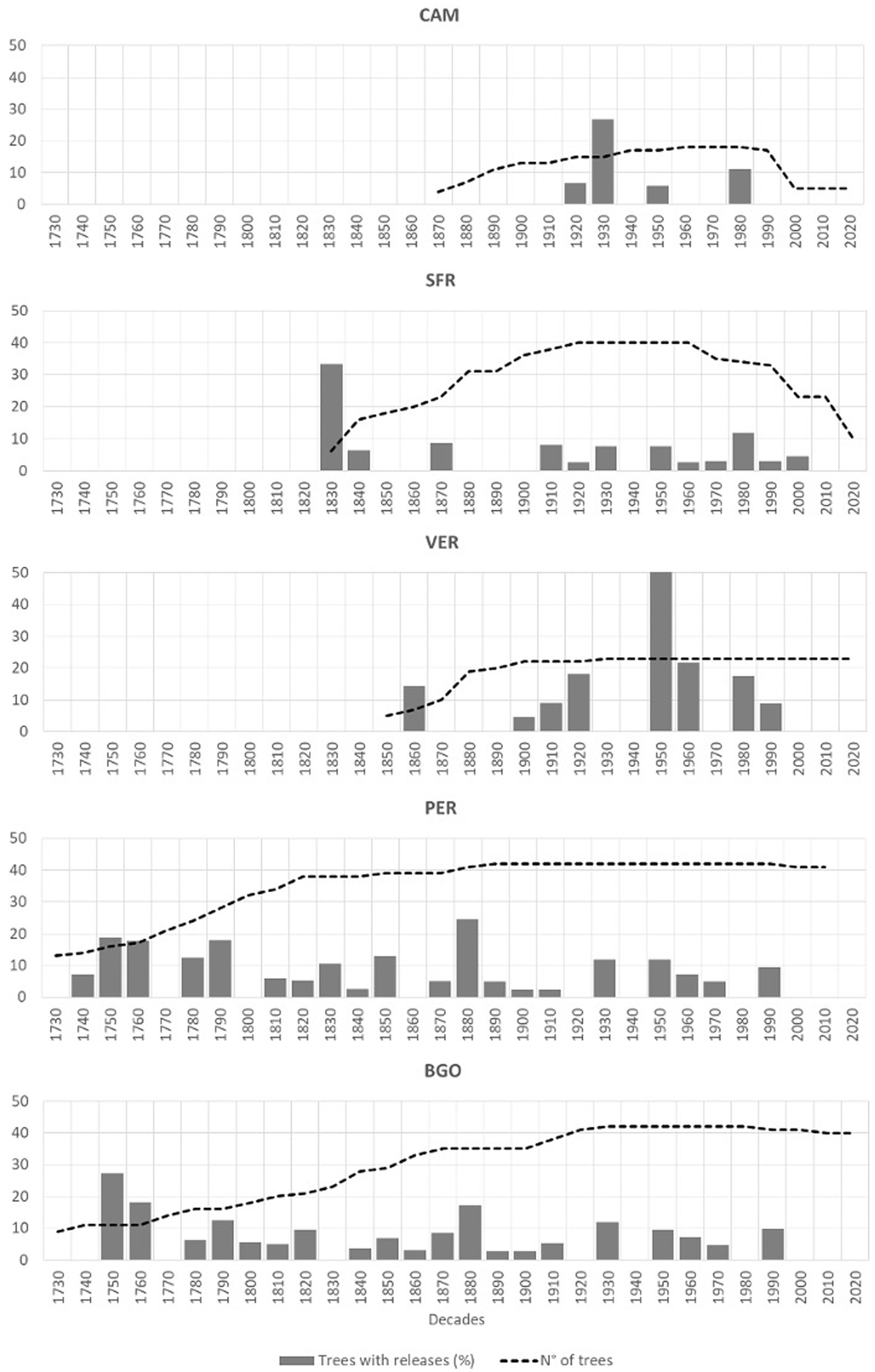
Figure 3. Release from suppression for the five study sites (gray bars: percent of trees showing abrupt growth release; dotted line: sample depth).
3.3 Multivariate ordination analyses
The forest structures of the Apennine study areas are different, and this diversity is expressed by the distance between their centroids in the ordination environment (Figure 4). VER is characterized by lower basal area (BA) and high biodiversity related to both species (SH_sp) and structure (SH_dbh). CAM and SFR are characterized by a higher basal area (BA), but silver fir is dominant at CAM and beech at SFR. Another considerable difference between the latter areas is the amount of regeneration (higher at SFR) and CWD (higher at CAM).
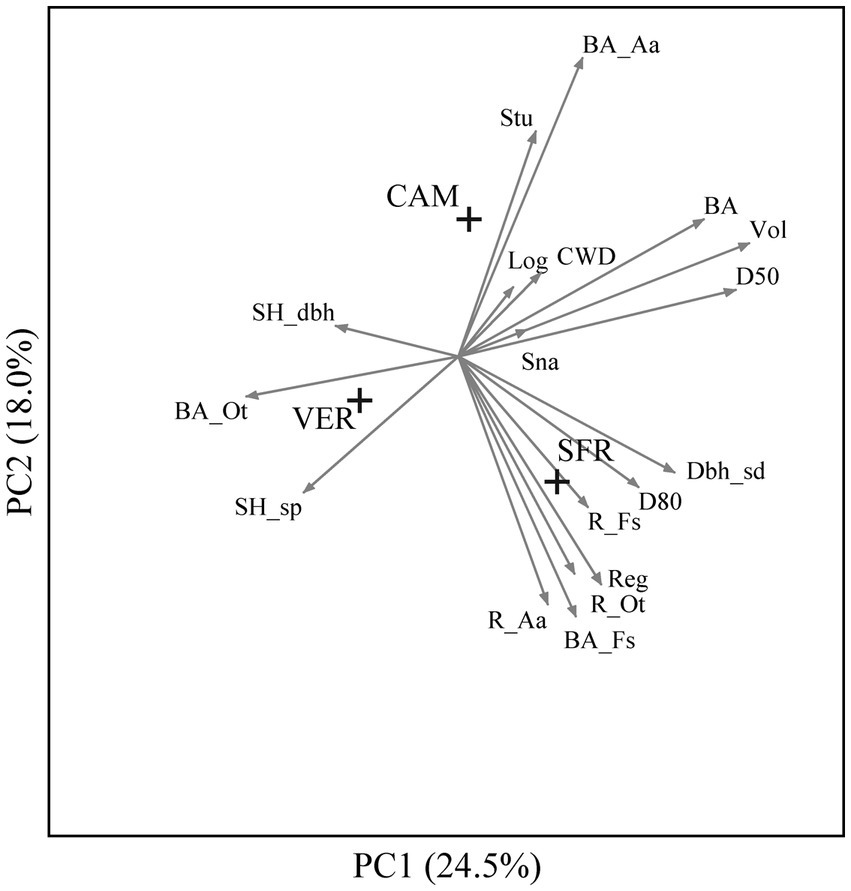
Figure 4. Principal component analysis of 99 plots of 3 sites of the Italian Apennines (CAM, Camaldoli; SFR, Sasso Fratino; VER, La Verna) indicated by crosses.
By adding the two reference study areas (PER and BGO) to the ordination analysis, we observed a marked difference between the Apennine study areas and the two OGFs of the Dinaric Alps (Figure 5). The latter is characterized by a higher amount of CWD, regeneration density (R), large living trees (D80), and large dead trees (Log>50, Snag>50). This multivariate comparison highlights the similarities between the two OGFs of the Dinaric Alps and their marked structural distance from the Apennine study areas.
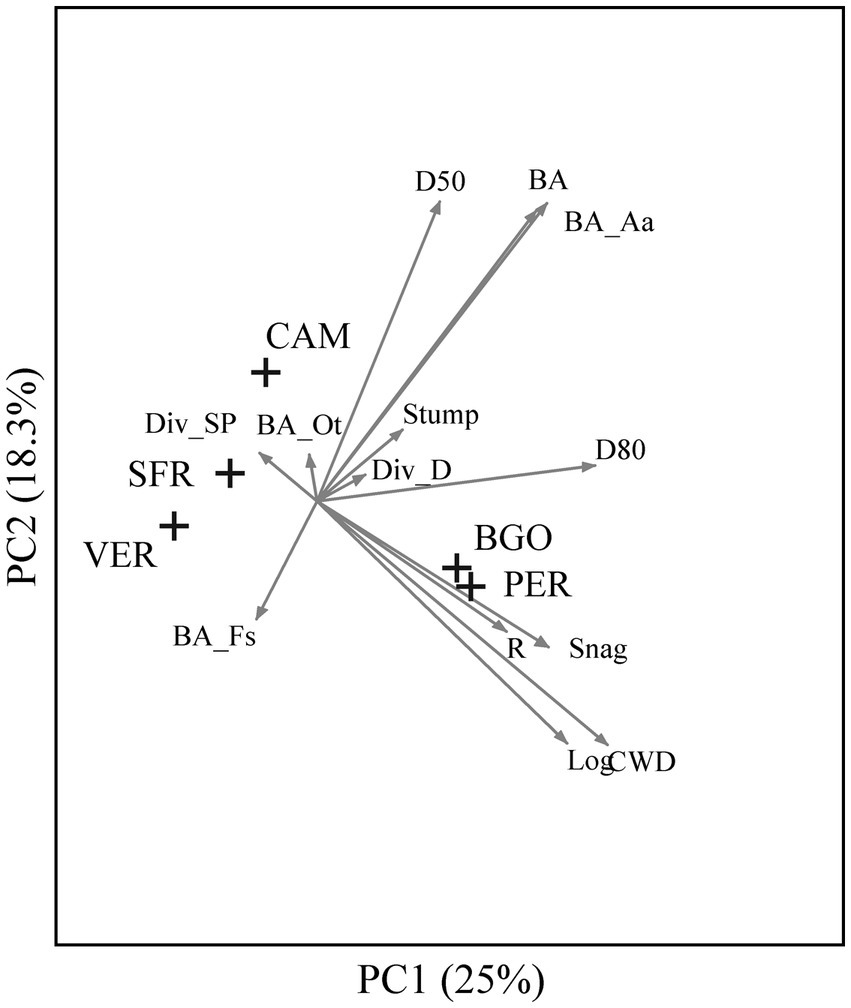
Figure 5. Principal component analysis of 161 plots of 3 sites of the Italian Apennines (CAM, Camaldoli; SFR, Sasso Fratino; VER, La Verna) and 2 sites of the Dinaric Alps (PER, Perućica; BGO, Biogradska Gora) indicated by crosses.
4 Discussion
4.1 Cultural and land-use legacies on the current structure of the Apennine study areas
The Camaldoli hermitage was established in the 11th century (Table 4). The Camaldolesi, since 1,278, appointed a “keeper of the forest” and, in the middle ages, started to carry out an intensive timber trade (Borchi, 1996, 2004). The management principles were originally orally transmitted but later became an official part of the monastic rules described, in 1525, in the “Forestry code,” the first example of written rules aiming at both reaching a sustained production while preserving long-term productivity of the forest (Romano and Urbinati, 2012). They gradually replaced the beech with even-aged and monospecific fir stands. As the code describes, they regulated logging by locals. Every year, they planted thousands of trees to integrate the natural regeneration. The silvicultural system ranged from a single tree and small group selection to large forest gaps, which were replanted and developed the Camaldolese “monastic silviculture” that preserved or even increased the presence of silver fir in the region (Agnoletti, 2013). Order ownership was interrupted in the 19th century when management governance changed at different times. During the 19th century, a prominent role was held by the Bohemian forest engineer Karl Siemon, who was appointed Inspector and Administrator of the Royal Casentine Forests by Grand Duke Leopold II (who was the owner of the forests at that time) in 1837. Karl Siemon and his colleague Anton Seeland carried out a profound reorganization of forest management, improving the forest road network and reforesting extensive areas. The central European silvicultural principles introduced clear-cutting followed by artificial regeneration (Vazzano et al., 2011). The CAM forest has been intensively harvested, mainly with clear-cut, during and after the First and the Second World Wars (Borchi, 2013). Starting from the middle of the 20th century, the State Forestry Agency made an extensive post-war restoration through the entire forest by planting silver fir in the clear-cut areas and of beech under the cover of the dominant silver fir, which originated the current intermediate and suppressed beech layers. The current monolayered structure of most of the Camaldoli study area reflects the central European silvicultural principles based on clear-cutting and dense artificial afforestation of silver fir and thinning of the monolayered stands introduced by Karl Siemon more than the Benedictine monastic approach.
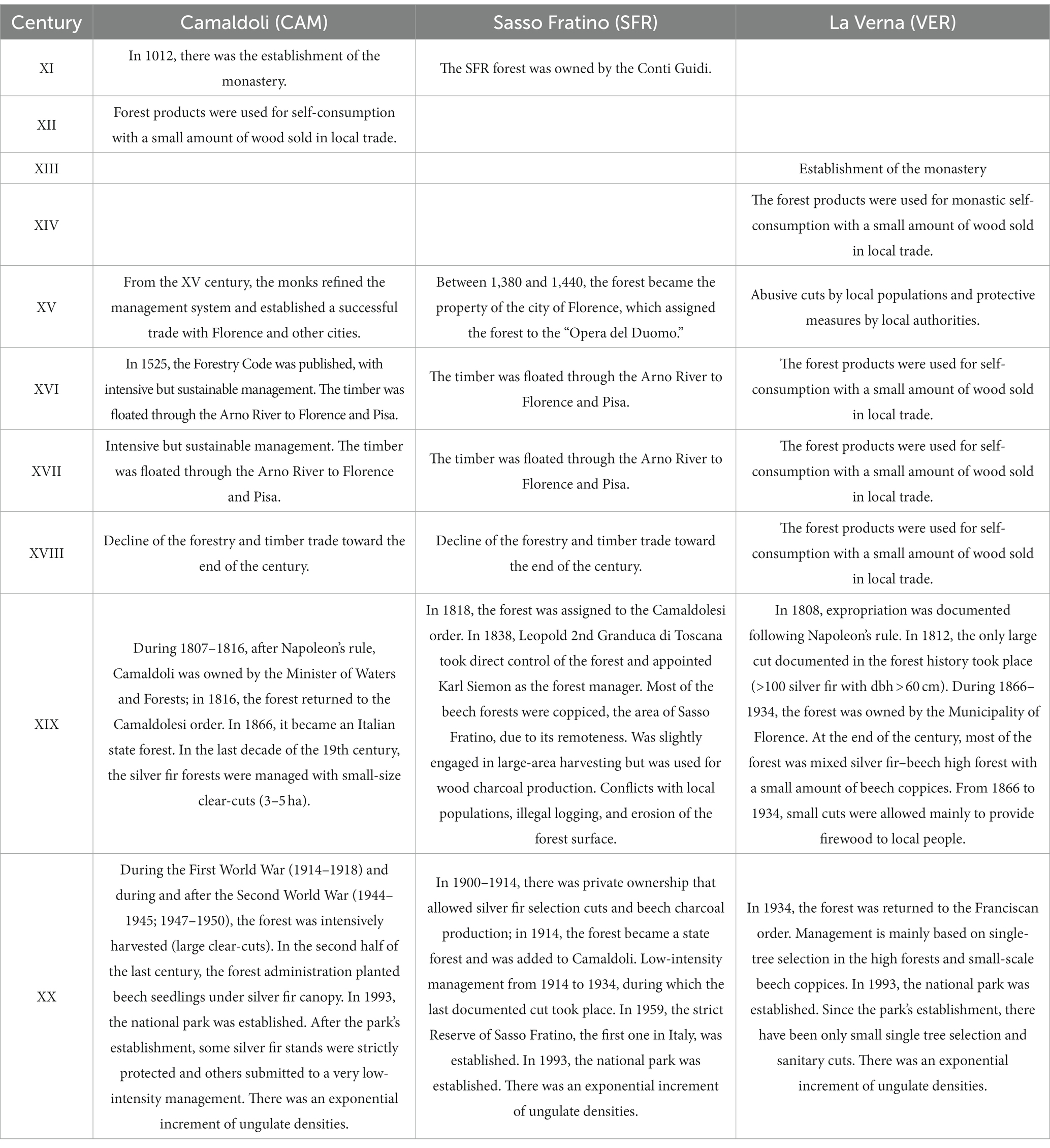
Table 4. Relevant historical events in the three northern Apennine study areas.
SFR was held by Opera del Duomo, an institution created in 1380 by the Florentine Republic to build the church of Santa Maria del Fiore in Florence, which owned a large forest estate extending across the Apennine ridge and was the most important producer of silver fir timber in Tuscany. Since the beginning, the area was disputed with local communities that needed land for agriculture and pasture. The property and the management changed at different times in the 19th century and became the property of the Italian Kingdom (Azienda del Demanio Forestale di Stato) in 1914. Due to the complex site conditions (slope and remoteness), the area of Sasso Fratino has not been the object of continuous and regular forest management but was only sporadically harvested for timber (silver fir) and was often used for charcoal production and the assignment of firewood to local communities (Bottacci, 2009a). The legacies of the charcoal production activity are dozens of abandoned charcoal kiln platforms still visible in the Reserve (Carrari et al., 2017). According to the management plans, the most recent timber harvests (approximately 400 m3) were conducted by the State Forestry Agency in 1936 (Bianchi et al., 2011). In 1959, a first block of 113 ha became the first established Italian strict Reserve. The Reserve was enlarged in the following years and currently covers 764 ha (Bottacci, 2009b). The current irregular forest structure reflects the past sporadic but intense harvesting followed by some decades of strict protection. Probably because of its importance for past charcoal production, the beech dominates all the vertical layers and still shows signs of past coppicing. Sporadic silver fir veteran trees surrounded by beech were probably left when the beech was coppiced as reserves due to the difficulty of transporting the trees out of the forest. The chronology of the releases from suppression in the last decades shows that natural processes (small-scale disturbances) are slowly shaping the forest structure.
The VER forest has been owned and managed since the 13th century by the Franciscan religious order. Unlike the Camaldolese monks, the Franciscan monks were not interested in increasing the abbey’s properties or carrying out any form of trading as poverty was their most important precept. They cultivated the land necessary for their living collecting and using the firewood or timber needed. The timber trade was almost ignored except for selling limited wood to local inhabitants (Borchi, 2004). Furthermore, respect for nature was part of the teachings of Saint Francis; thus, no intensive management rules were developed. Consequently, the silviculture mainly consisted of low-intensity single tree or group selection timber felling in the high forest. Small beech coppiced stands (Borchi and Miozzo, 2015) were harvested only to supply the essential needs of the friary. The only extensive commercial cuts were made at the beginning of the 19th century during the Napoleonic period. Small salvage loggings were conducted after big storms (e.g., in 1888, 1943, and 2015) that broke or uprooted large silver fir or beech, those with dbh > 1 m, that could grow because of the conservative approach of the Franciscan monks (Miozzo and Borchi, 2009). Since the 80s of the last century, the management has been based on salvage low-intensity restoration loggings. Even if it has always been managed with low intensity, at least in the previous century, the VER forest is the only one, among the three Apennine forests studied, that has had a continuous forest cover without large clear-cuts or coppicing.
By comparing the history of land use and the current structures, it is evident that past land use has been the main driver for the current structure, which is very different in each of the three forests (Figure 4). The role of monasteries in the conservation and management of silver fir stands was so crucial that foresters coined the term “monastic silviculture” to define the historical relationship between the monks and the forest (Agnoletti and Paci, 2001). This term was mainly related to the Camaldolesi role because this was the place where a silvicultural system was developed. For the Benedictines, the forest was a tool to increase the monastery’s power and god’s glory. However, the management was based on the principle “only to use that what will regrow” (Freyer et al., 2018) and the “Camaldoli forestry code” can be considered one of the first examples of sustainable forest management that was published and applied 250 years before von Carlowitz’s Sylvicultura oeconomica (Brink, 2022). There was a significant difference between Benedictine and Franciscan rules, and this different “spiritual” and “behavioral” attitude has substantially affected the landscape that is still visible today. The productive but sustainable benedictine management model contrasted with the Franciscans’ more “natural” approach. The history of SFR (located 3 km from Camaldoli and 16 km from La Verna) is slightly different for its different ownership and relative remoteness (located on the northern side of the Apennines, with no road crossing the forest) and its slope steepness. The forest structure has been more influenced by intense but sporadic harvesting and by local farmers more interested in firewood than timber.
Consequently, there was a higher incidence of beech coppiced, and many charcoal kiln platforms are still visible today. At the same time, the sporadic harvesting and the difficulties of wood extraction have allowed the conservation of some old and large trees. The last silvicultural intervention dates back to 1936, and in the last decades, the natural dynamics have shaped and partially overlapped the previous anthropogenic structure (Travaglini et al., 2012).
4.2 Old-growthness and dynamics
All the studied Apennine forests, although they have been strictly protected for more than 60 years as is the case of SFR or have had in recent decades only low-intensity or sanitary interventions (CAM and VER), still have structural characteristics very different from the Dinaric old-growth forests (Figure 5).
The Italian study areas have much higher living tree biomass than low-intensity managed forests (Motta et al., 2015b) and close to the amount of old-growth stands in SFR and CAM (Table 2). This amount is mainly due to the cessation/substantial reduction of harvesting in the last decades.
The main structural differences between the Italian and Dinaric forests are related to CWD’s quantity (total volume) and quality (large snags and logs). According to the CWD diameter distribution, endogenous processes dominate mortality more than exogenous small- or large-scale disturbances (except some small windthrows in VER). All three study areas lack large trees, snags, and logs that characterize late seral stages. Large old trees play an extraordinary range of critical ecological roles, including in hydrological regimes, nutrient cycles, and numerous ecosystem processes (Lutz et al., 2012, 2018; Lee et al., 2022). In addition, large-diameter trees and CWD store disproportionally massive amounts of carbon and are a significant driver of carbon cycle dynamics in forests (Mildrexler et al., 2020). Even if the age of the oldest trees observed in the three Italian forests is relevant for the Apennines (Table 3), still there are significant differences with Dinaric OGFs, where both beech and silver fir reach their biological age limits. As expected, the Shannon diversity index and standard deviation of the diameter are higher in the Dinaric forests, highlighting the presence of a wide range of development stages characterized by different tree sizes.
Considering the recent starting point and the long-term development of the forest dynamics, the Apennine study areas are currently, according to Oliver and Larson (1996), in the late pole stage (CAM) or at the beginning of the transition stage (SRF and VER). After logging cessation, legacy effects affect species composition and structure for decades or even for centuries (Martin-Benito et al., 2022) and the altered forests can return to a more natural level of structural complexity and biodiversity, but conversion can be slow and, in some way, unpredictable (Whitlock et al., 2017; Albrich et al., 2021).
4.3 Biocultural diversity, conservation, and current management
The management of silver fir forests in the last decades has shifted from traditional use to almost strict protection, where human intervention is banished. This management has been applied in some state forests before the FCNP establishment: the Sasso Fratino Reserve was established in 1959, and then other forests owned by the state (Badia Prataglia, Campigna, and La Scodella) became Biogenetic Nature Reserves in 1977. After the FCNP establishment (1993), a substantial restriction for forest management and tending was extended to all the silver fir forests. This policy is understandable for Sasso Fratino, the first Italian strict reserve. Even if the SFR forests have been intensely harvested in the past, even coppiced in most parts, SFR forest is currently mixed, has irregular structure, has never been artificially regenerated, and is probably currently within its natural range of variability (Landres et al., 1999). The same approach is not advisable for La Verna. In this case, the forest is mixed, the structure is multilayered, the regeneration has always been natural, and the forest study area certainly is in its natural range of variability. The difference is that, in La Verna, strict protection means the loss of the historical role of the forest and its relationships with the Franciscan monastery. Finally, the strict protection in Camaldoli is not comprehensible in pure silver fir forests that have been clear-cut and artificially regenerated. Furthermore, when taking into account that the original goal of strict protection was the conservation of the genetic heritage (Riserva Biogenetica) it becomes clear that such a goal is impossible to apply in forests artificially regenerated with seeds of unknown origin. At the same time, this type of management does not take into account the cultural value of forests that have been managed for centuries by Camaldolesi monks, who have shaped a cultural landscape and created a biocultural diversity (Figure 6) that, with the current management approach will be lost in a few decades (Scherzinger, 1996). According to Frascaroli et al. (2016), “Forbidding traditional management to privilege minimal intervention… will result in assimilating sacred natural forests to surrounding areas, with consequent loss of biological distinctiveness and decrease of landscape scale diversity.” According to Lelli et al. (2021), “Forests are getting poorer in species in the understorey, with a higher tree layer and denser canopy cover.” In their Sacred Natural Sites (SNS) review, Zannini et al. (2022) recommend “traditional management to be encouraged at SNS… especially when this is supported by local people and traditional uses of the SNS are still alive” because “it would be the most effective way to conserve both nature and culture at some of Europe’s last hot spots of biocultural diversity.”
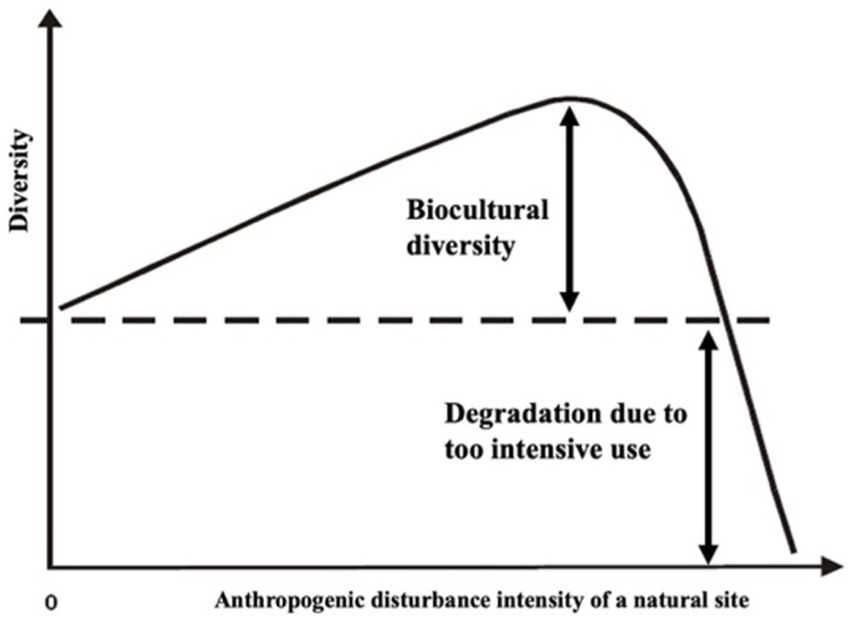
Figure 6. Biocultural diversity. Based on low/intermediate land-use intensity and landscape heterogeneity, traditional land use increases the original diversity, creating biocultural diversity. A degradation and diversity decline exist if the land-use intensity is too high. On the other hand, if the traditional land use is abandoned or if it is withdrawn from management for other purposes, there is a diversity decline due to the loss of disturbances that maintain biocultural diversity (From Scherzinger (1996); modified).
4.4 Conclusion
The studied forests represent the northern Apennines’ most important silver fir remains. The conservation of these forests is mainly due to the “monastic silviculture” that has preserved and even planted and spread the silver fir for economic and spiritual purposes (Agnoletti and Paci, 2001). Even if the three studied forests are relatively close and share similar site conditions, their current structures are significantly different. The differences are mainly due to past ownership and different monastic rules and spiritual attitudes. The monks have created a cultural landscape and a peculiar biocultural diversity over the centuries, even with other historical contributions (the most important one is due to the Bohemian forest engineer Karl Siemon, who managed Camaldoli and most of the silver fir forests in the 19th century). There are currently substantial structural differences between Apennine forests and Dinaric old-growth forests mainly due to the amount and the quality of CWD, the lack of large and old living trees, and the lack of large snags and logs.
The current management privileges strict protection and very low-intensity management (unsustainable from the economic point of view) with the purpose of restoring biodiversity, old-growth conditions, and natural processes. This policy sounds legitimate for the remote forest of SFR and its biological and symbolic role as the first Italian strict reserve. However, it is not shareable for VER and even more for CAM, where the current stands are planted and strict protection will induce the loss of a unique cultural landscape. Simply abandoning forest management in previously intensively managed landscapes has limited conservation value (Bruun and Heilmann-Clausen, 2021) and has a negative impact on biodiversity conservation and on the development of natural processes (Mölder et al., 2014). A return to old-growth conditions can take centuries, and for Mediterranean forests, primarily located in densely inhabited areas with high land-use pressure, restoring the attributes of primary forests remains challenging (Sabatini et al., 2020).
Another serious issue for the future of the silver fir Apennine forests is the high browsing incidence that is almost preventing regeneration establishment. The primary objectives of a national park are the protection of natural processes and species conservation. When these objectives come into conflict, as it usually occurs because of the cascading effects of large mammals on lower trophic levels (Carpio et al., 2021), park managers have to decide upon the appropriate management while considering local peculiarities (van Beeck Calkoen et al., 2020).
Land-use activities of different types, durations, and intensities have modified the Italian forests. Most forest landscapes support both cultural and natural elements (Figure 6). These intermediate conditions are shaped by complex interactions of changing climate and land use that operate over different temporal and spatial scales and pose the conservation challenge of supporting both natural structure and diversity, on one hand, and cultural features on the other (Lindenmayer and Hunter, 2010). Recognizing protected areas as cultural landscapes that host an important biocultural diversity requires site and forest-specific management while considering both naturalness and culturalness issues (Sarmiento-Mateos et al., 2019). The risk is that by applying almost exclusive biodiversity-centered management, applying strict protection measures, and setting difficult or even impossible-to-achieve biodiversity goals, the total diversity will decrease, and biocultural diversity, which is the greatest richness of most European parks, will be lost.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
MR developed the conception and the method of the study. MR, GM, BR, CM, DV, and BA made the field data collection. GM made the multivariate ordination analyses. NP made the dendrochronological analyses. BR, BA, and MR made the structural analysis. All authors contributed to the article and approved the submitted version.
Acknowledgments
Thanks to Reparto Carabinieri per la Biodiversità di Pratovecchio (Pratovecchio, AR, Italy), Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (Pratovecchio, AR, Italy), Biogradska Gora National Park (Kolasin, MNE), and Sutjeska National Park (Tjentiste, BiH) for providing information, data, and logistic support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Agnoletti, M. (2013). “Tuscany” in Italian historical rural landscapes. ed. M. Agnoletti (Dordrecht: Springer)
Agnoletti, M., and Paci, M. (2001). “Monks, foresters and ecology. Silver fir in Tuscany from XIV to XX century” in Le Sapin - Enjeux anciens, Enjeux actuels. ed. A. Corvol (Paris: L'Harmattan), 173–194.
Agnoletti, M., Piras, F., Venturi, M., and Santoro, A. (2022). Cultural values and forest dynamics: the Italian forests in the last 150 years. For. Ecol. Manag. 503:119655. doi: 10.1016/j.foreco.2021.119655
Agnoletti, M., and Rotherham, I. D. (2015). Landscape and biocultural diversity. Biodivers. Conserv. 24, 3155–3165. doi: 10.1007/s10531-015-1003-8
Albrich, K., Thom, D., Rammer, W., and Seidl, R. (2021). The long way back: development of central European mountain forests towards old-growth conditions after cessation of management. J. Veg. Sci. 32:e13052. doi: 10.1111/jvs.13052
Bianchi, L., Bottacci, A., Calamini, G., Maltoni, A., Mariotti, B., Quilghini, G., et al. (2011). Structure and dynamics of a beech forest in a fully protected area in the northern Apennines (Sasso Fratino, Italy). IForest 4, 136–144. doi: 10.3832/ifor0564-004
Borchi, S. (1996). Ordini religiosi e modalità di rapporto col bosco: le fonti di Camaldoli e La Verna. Sherwood - Foreste ed Alberi Oggi 14, 17–21.
Borchi, S. (2004). “La gestion des forêts par les ordres religieux: camaldoli, vallombreuse et La Verne” in Abbayes et monastères aux racines de l'Europe. eds. P. Poupard and B. Ardura (Paris: Cerf)
Borchi, S. (2013). Tagli bellici nella foresta di Camaldoli. Sherwood - Foreste ed alberi oggi 195, 10–15.
Borchi, S., and Miozzo, M. (2015, 2014). “Monitoraggio compositivo e strutturale della foresta de La Verna (Italia - AR) attraverso la gestione selvicolturale dal 1890 a oggi” in Second international congress of Silviculture. Designing the future of the forestry sector. ed. O. Ciancio (Firenze: Accademia Italiana di Scienze Forestali), 249–256.
Bottacci, A. (2009a). “Cenni storici” in La Riserva Naturale Integrale di Sasso Fratino. 1959–2009, 50 anni di conservazione della biodiversità. ed. A. Bottacci (Pratovecchio: CFS/UTB Pratovecchio), 29–38.
Bottacci, A. (ed.). (2009b). La Riserva Naturale Integrale di Sasso Fratino. 1959–2009, 50 anni di conservazione della biodiversità. Pratovecchio: CFS/UTB.
Bottero, A., Garbarino, M., Dukic, V., Govedar, Z., Lingua, E., Nagel, T. A., et al. (2011). Gap-phase dynamics in the old-growth Forest of Lom, Bosnia and Herzegovina. Silva Fenn. 45, 875–887. doi: 10.14214/sf.76
Brink, S. C. (2022). Sustainability: a 21st century concept? Trends Plant Sci. 27, 619–620. doi: 10.1016/j.tplants.2022.05.001
Brown, A. G., Hatton, J., Selby, K. A., Leng, M. J., and Christie, N. (2013). Multi-proxy study of Holocene environmental change and human activity in the central Apennine Mountains, Italy. J. Quat. Sci. 28, 71–82. doi: 10.1002/jqs.2591
Bruun, H. H., and Heilmann-Clausen, J. (2021). What is unmanaged forest and how does it sustain biodiversity in landscapes with a long history of intensive forestry? J. Appl. Ecol. 58, 1813–1816. doi: 10.1111/1365-2664.13754
Cagliero, E., Morresi, D., Paradis, L., Curovic, M., Spalevic, V., Marchi, N., et al. (2022). Legacies of past human activities on one of the largest old-growth forests in the south-east European mountains. Veg. Hist. Archaeobotany 31, 415–430. doi: 10.1007/s00334-021-00862-x
Carpio, A. J., Apollonio, M., and Acevedo, P. (2021). Wild ungulate overabundance in Europe: contexts, causes, monitoring and management recommendations. Mammal Rev. 51, 95–108. doi: 10.1111/mam.12221
Carrari, E., Ampoorter, E., Bottalico, F., Chirici, G., Coppi, A., Travaglini, D., et al. (2017). The old charcoal kiln sites in central Italian forest landscapes. Quat. Int. 458, 214–223. doi: 10.1016/j.quaint.2016.10.027
Castagneri, D., Garbarino, M., Berretti, R., and Motta, R. (2010). Site and stand effects on coarse woody debris in montane mixed forests of eastern Italian Alps. For. Ecol. Manag. 260, 1592–1598. doi: 10.1016/j.foreco.2010.08.008
Conedera, M., Krebs, P., Tinner, W., Pradella, M., and Torriani, D. (2004). The cultivation of Castanea sativa (mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobotany 13, 161–179. doi: 10.1007/s00334-004-0038-7
Curovic, M., Spalevic, V., Sestras, P., Motta, R., Dan, C., Garbarino, M., et al. (2020). Structural and ecological characteristics of mixed broadleaved old-growth forest (Biogradska Gora - Montenegro). Turk. J. Agric. For. 44, 428–438. doi: 10.3906/tar-2003-103
Finsinger, W., Cagliero, E., Morresi, D., Paradis, L., Čurović, M., Garbarino, M., et al. (2022). The value of long-term history of small and fragmented old-growth forests for restoration ecology. Past Glob. Chang. Mag. 30, 8–9. doi: 10.22498/pages.30.1.8
Frascaroli, F. (2013). Catholicism and conservation: the potential of sacred natural sites for biodiversity management in Central Italy. Hum. Ecol. 41, 587–601. doi: 10.1007/s10745-013-9598-4
Frascaroli, F., Bhagwat, S., Guarino, R., Chiarucci, A., and Schmid, B. (2016). Shrines in Central Italy conserve plant diversity and large trees. Ambio 45, 468–479. doi: 10.1007/s13280-015-0738-5
Fraver, S., Kenefic, L. S., Cutko, A. R., and White, A. S. (2020). Natural disturbance and stand structure of old-growth northern white-cedar (Thuja occidentalis) forests, northern Maine, USA. For. Ecol. Manag. 456:117680. doi: 10.1016/j.foreco.2019.117680
Fraver, S., and White, A. S. (2005). Identifying growth releases in dendrochronological studies of forest disturbance. Can. J. For. Res. 35, 1648–1656. doi: 10.1139/x05-092
Freyer, B., Aversano-Dearborn, V., Winkler, G., Leipold, S., Haidl, H., Brand, K. W., et al. (2018). Is there a relation between ecological practices and spirituality? The case of benedictine monasteries. J. Agric. Environ. Ethics 31, 559–582. doi: 10.1007/s10806-018-9745-4
Gabbrielli, A. (2006). Le vicende storiche e demografiche italiane come causa dei cambiamenti del paesaggio forestale. Annali Accademia Italiana di Scienze Forestali 55, 133–166.
Henne, P. D., Elkin, C., Colombaroli, D., Samartin, S., Bugmann, H., Heiri, O., et al. (2012). Impacts of changing climate and land use on vegetation dynamics in a Mediterranean ecosystem: insights from paleoecology and dynamic modeling. Landsc. Ecol. 28, 819–833. doi: 10.1007/s10980-012-9782-8
IPCC (2006). “2006 IPCC guidelines for national greenhouse gas inventories” in A primer, Prepared by the National Greenhouse Gas Inventories Programme. eds. H. S. Eggleston, K. Miwa, N. Srivastava, and K. Tanabe (Japan: IGES)
Landres, P. B., Morgan, P., and Swanson, F. J. (1999). Overview of the use of natural variability concepts in managing ecological systems. Ecol. Appl. 9, 1179–1188.
Larsson, L.A., and Larsson, P.O. (2018). CDendro and CooRecorder (v. 9.3. 1). Saltsjöbaden, Sweden: Cybis Elektronik.
Lee, C. B., Ali, A., Yuan, Z., Lutz, J. A., Svenning, J. C., and Lee, M. K. (2022). Large-diameter trees control forest structure and function in successional temperate forests. Biogeosci. Discuss. 2022, 1–20. doi: 10.5194/bg-2022-60
Leibundgut, H. (1960). Risultati delle ricerche in foreste vergini europee. Annali Accademia Italiana di Scienze Forestali 9, 277–287.
Lelli, C., Nascimbene, J., Alberti, D., Agostini, N., Zoccola, A., Piovesan, G., et al. (2021). Long-term changes in Italian mountain forests detected by resurvey of historical vegetationdata. J. Veg. Sci. 32:e12939. doi: 10.1111/jvs.12939
Lindenmayer, D., and Hunter, M. (2010). Some guiding concepts for conservation biology. Conserv. Biol. 24, 1459–1468. doi: 10.1111/j.1523-1739.2010.01544.x
Lombardi, F., Marchetti, M., Corona, P., Merlini, P., Chirici, G., Tognetti, R., et al. (2015). Quantifying the effect of sampling plot size on the estimation of structural indicators in old-growth forest stands. For. Ecol. Manag. 346, 89–97. doi: 10.1016/j.foreco.2015.02.011
Lutz, J. A., Furniss, T. J., Johnson, D. J., Davies, S. J., Allen, D., Alonso, A., et al. (2018). Global importance of large-diameter trees. Glob. Ecol. Biogeogr. 27, 849–864. doi: 10.1111/geb.12747
Lutz, J. A., Larson, A. J., Swanson, M. E., and Freund, J. A. (2012). Ecological importance of large-diameter trees in a temperate mixed-conifer Forest. PLoS One 7:e36131. doi: 10.1371/journal.pone.0036131
Marini Govigli, V., Efthymiou, A., and Stara, K. (2021). From religion to conservation: unfolding 300 years of collective action in a Greek sacred forest. Forest Policy Econ. 131:102575. doi: 10.1016/j.forpol.2021.102575
Martin-Benito, D., Molina-Valero, J. A., Pérez-Cruzado, C., Bigler, C., and Bugmann, H. (2022). Development and long-term dynamics of old-growth beech-fir forests in the Pyrenees: evidence from dendroecology and dynamic vegetation modelling. For. Ecol. Manag. 524:120541. doi: 10.1016/j.foreco.2022.120541
Mildrexler, D. J., Berner, L. T., Law, B. E., Birdsey, R. A., and Moomaw, W. R. (2020). Large trees dominate carbon storage in forests east of the Cascade crest in the United States Pacific northwest. Front. for. glob. change 3:594274. doi: 10.3389/ffgc.2020.594274
Miozzo, M., and Borchi, S. (2009). "La foresta della Verna in Casentino (AR): influenza della gestione selvicolturale degli ultimi secoli sulla struttura della foresta", In: Congresso Nazionale di Selvicoltura: accademia Italiana di Scienze Forestali. Ed. O. Ciancio. Taormina, 869–876.
Mölder, A., Streit, M., and Schmidt, W. (2014). When beech strikes back: how strict nature conservation reduces herb-layer diversity and productivity in central European deciduous forests. For. Ecol. Manag. 319, 51–61. doi: 10.1016/j.foreco.2014.01.049
Morales-Molino, C., Steffen, M., Samartin, S., van Leeuwen, J. F. N., Hurlimann, D., Vescovi, E., et al. (2021). Long-term responses of Mediterranean Mountain forests to climate change, fire and human activities in the northern Apennines (Italy). Ecosystems 24, 1361–1377. doi: 10.1007/s10021-020-00587-4
Motta, R., Berretti, R., Castagneri, D., Dukic, V., Garbarino, M., Govedar, Z., et al. (2011). Toward a definition of the range of variability of central European mixed Fagus-Abies-Picea forests: the nearly steady-state forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 41, 1871–1884. doi: 10.1139/X11-098
Motta, R., Berretti, R., Lingua, E., and Piussi, P. (2006). Coarse woody debris, forest structure and regeneration in the Valbona Forest reserve, Paneveggio, Italian Alps. For. Ecol. Manag. 235, 155–163. doi: 10.1016/j.foreco.2006.08.007
Motta, R., Garbarino, M., Berretti, R., Bjelanovic, I., Borgogno Mondino, E., Čurović, M., et al. (2015a). Structure, spatio-temporal dynamics and disturbance regime of the mixed beech–silver fir–Norway spruce old-growth forest of Biogradska Gora (Montenegro). Plant Biosyst. 149, 966–975. doi: 10.1080/11263504.2014.945978
Motta, R., Garbarino, M., Berretti, R., Meloni, F., Nosenzo, A., and Vacchiano, G. (2015b). Development of old-growth characteristics in uneven-aged forests of the Italian Alps. Eur. J. For. Res. 134, 19–31. doi: 10.1007/s10342-014-0830-6
Motta, R., and Nola, P. (2001). Growth trends and dynamics in sub-alpine forest stands in the Varaita Valley (Piedmont, Italy) and their relationships with human activities and global change. J. Veg. Sci. 12, 219–230. doi: 10.2307/3236606
Motta, R., Nola, P., and Piussi, P. (1999). Structure and stand development in three subalpine Norway spruce (Picea abies (L.) karst.) stands in Paneveggio (Trento, Italy). Glob. Ecol. Biogeogr. 8, 455–471. doi: 10.1046/j.1365-2699.1999.00165.x
Muigg, B., Skiadaresis, G., Tegel, W., Herzig, F., Krusic, P. J., Schmidt, U. E., et al. (2020). Tree rings reveal signs of Europe’s sustainable forest management long before the first historical evidence. Sci. Rep. 10:21832. doi: 10.1038/s41598-020-78933-8
Müllerová, J., Pejcha, V., Altman, J., Plener, T., Dörner, P., and Doležal, J. (2016). Detecting coppice legacies from tree growth. PLoS One 11:e0147205. doi: 10.1371/journal.pone.0147205
Nagel, T. A., and Svoboda, M. (2008). Gap disturbance regime in an old-growth Fagus-Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 38, 2728–2737. doi: 10.1139/X08-110
Payette, S., Filion, L., and Delwaide, A. (1990). Disturbance regime of a cold temperate forest as deduced from tree-ring patterns: the Tantaré ecological reserve, Québec. Can. J. For. Res. 20, 1228–1241. doi: 10.1139/x90-162
Romano, R., and Urbinati, C. (eds.). (2012). Foresta e monaci di Camaldoli: un rapporto millenario tra gestione e conservazione. Roma: INEA.
Rubino, D. L., and McCarthy, B. C. (2004). Comparative analysis of dendroecological methods used to assess disturbance events. Dendrochronologia 21, 97–115. doi: 10.1078/1125.7865.00047
Sabatini, F. M., Burrascano, S., Keeton, W. S., Levers, C., Lindner, M., Potzschner, F., et al. (2018). Where are Europe's last primary forests? Divers. Distrib. 24, 1426–1439. doi: 10.1111/ddi.12778
Sabatini, F. M., Keeton, W. S., Lindner, M., Svoboda, M., Verkerk, P. J., Bauhus, J., et al. (2020). Protection gaps and restoration opportunities for primary forests in Europe. Divers. Distrib. 26, 1646–1662. doi: 10.1111/ddi.13158
Sarmiento-Mateos, P., Arnaiz-Schmitz, C., Herrero-Jáuregui, C., Pineda, F. D., and Schmitz, M. F. (2019). Designing protected areas for social–ecological sustainability: effectiveness of management guidelines for preserving cultural landscapes. Sustainability 11:2871. doi: 10.3390/su11102871
Scherzinger, W. (1996). Naturschutz im Wald. Qualitäts- ziele einer dynamischen Waldentwicklung. Stuttgart: Ulmer.
Schweingruber, F. H., Eckstein, D., Serre-Bachet, F., and Braker, O. U. (1990). Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 8, 9–38.
Stupar, V., and Milanović, D. (2017). History of nature protection in the Sutjeska National Park. Bullet. Facul.For.Univer. Banja Luka 1, 113–128. doi: 10.7251/GSF1726113S
Susmel, L. (1956). Caratteri comparati delle abetine primarie delle Alpi dinariche e delle abetine secondarie delle Alpi orientali italiane. Ann. Acc. Ital. Sci. For. 5, 115–146.
Tinner, W., Colombaroli, D., Heiri, O., Henne, P. D., Steinacher, M., Untenecker, J., et al. (2013). The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 83, 419–439. doi: 10.1890/12-2231.1
Travaglini, D., Paffetti, D., Bianchi, L., Bottacci, A., Bottalico, F., Giovannini, G., et al. (2012). Characterization, structure and genetic dating of an old-growth beech-fir forest in the northern Apennines (Italy). Plant Biosyst. 146, 175–188. doi: 10.1080/11263504.2011.650731
Vacchiano, G., Garbarino, M., Lingua, E., and Motta, R. (2017). Forest dynamics and disturbance regimes in the Italian Apennines. For. Ecol. Manag. 388, 57–66. doi: 10.1016/j.foreco.2016.10.033
van Beeck Calkoen, S. T. S., Mühlbauer, L., Andrén, H., Apollonio, M., Balčiauskas, L., Belotti, E., et al. (2020). Ungulate management in European national parks: why a more integrated European policy is needed. J. Environ. Manag. 260:110068. doi: 10.1016/j.jenvman.2020.110068
Vazzano, E., Quilghini, G., Travaglini, D., and Nocentini, S. (2011). Changes in forest cover in the Foresta della Lama (Casentino forests National Park) from Karl Siemon and Anton Seeland 1837 forest management plan. For. J. Silvic. For. Ecol. 8, 78–87. doi: 10.3832/efor0655-008
Whitlock, C., Colombaroli, D., Conedera, M., and Tinner, W. (2017). Land-use history as a guide for forest conservation and management. Conserv. Biol. 32, 84–97. doi: 10.1111/cobi.12960
Wick, L., and Möhl, A. (2006). The mid-Holocene extinction of silver fir (Abies alba) in the southern Alps: a consequence of forest fires? Palaeobotanical records and forest simulations. Veg. Hist. Archaeobotany 15, 435–444. doi: 10.1007/s00334-006-0051-0
Zannini, P., Frascaroli, F., Nascimbene, J., Halley, J. M., Stara, K., Cervellini, M., et al. (2022). Investigating sacred natural sites and protected areas for forest area changes in Italy. Conserv. sci. pract. 4:e12695. doi: 10.1111/csp2.12695
Keywords: Abies alba, Fagus sylvatica, biocultural diversity, protected areas, forest structure, forest dynamics, old-growth forests
Citation: Motta R, Garbarino M, Berretti R, Bono A, Curovic M, Dukić V and Nola P (2023) Monastic silviculture legacies and current old-growthness of silver fir (Abies alba) forests in the northern Apennines (Italy). Front. For. Glob. Change. 6:1252462. doi: 10.3389/ffgc.2023.1252462
Edited by:
Annemarie Bastrup-Birk, European Environment Agency, DenmarkReviewed by:
Fabio Lombardi, Mediterranea University of Reggio Calabria, ItalyLuigi Portoghesi, University of Tuscia, Italy
Copyright © 2023 Motta, Garbarino, Berretti, Bono, Curovic, Dukić and Nola. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renzo Motta, cmVuem8ubW90dGFAdW5pdG8uaXQ=