 Zorica Popović
Zorica Popović Vera Vidaković
Vera Vidaković Jovana Janković
Jovana Janković- Department of Ecology, Institute for Biological Research “Siniša Stanković”—National Institute of Republic of Serbia, University of Belgrade, Belgrade, Serbia
Introduction: A variety of plant traits, from architectural to the cellular level, have been connected to flammability, but intraspecific variability of plant traits (ITV) and components of flammability is poorly studied. The lack of knowledge about ITV of plant traits related to flammability appears to be a major shortcoming in further interpreting species flammability and fire behavior and incorporating the data into models.
Methods: Morpho-ecophysiological traits (width, length, thickness, weight, area, volume, moisture content, flatness, specific leaf area, density of leaf tissue, ratio of area to volume) and time-to-ignition of fresh foliage were measured in seven populations of Picea omorika.
Results: All leaf traits are presented along with their correlations to the flammability trait. The seven populations differed in terms of fresh leaves’ time-to-ignition. Differences among populations in morpho-ecophysiological traits were also significant but not consistent among populations. PCA classified 49 elements into three different groups, where three populations were clustered by higher leaf area-related traits, other three populations were clustered by higher leaf length, volume, thickness, time-to-ignition, density index, moisture content, width, weight, and one population was classified between the two main groups. The first two principal components accounted for 87% of the total variance: variability in leaf area- and leaf weight-derived parameters (specific leaf area and density index) and time-to-ignition primarily defined the formation of the first axis, while variability in leaf flatness (based on leaf weight and thickness) primarily contributed to the formation of the second axis.
Discussion: Results suggest high ITV in natural populations of P. omorika regardless of site fire history.
Introduction
Serbian spruce [Picea omorika (Pančić) Purk.] is an endangered tertiary relict and endemic species with a restricted and fragmented natural range in western Serbia and eastern Bosnia and Herzegovina. Its current geographic distribution is between 43°21′ and 44°08′ North and 18°37′ and 19°45′ East, in the middle and upper basin of the Drina River. The list of threatened plants classifies the Serbian spruce as endangered due to its patchy population structure, consisting of about 30 groups of several tens to a few hundred trees (IUCN, 2023).1 Despite legal protection, there is growing concern for these populations as they have declined dramatically due to a variety of environmental causes, including climate change, land use change, and fire (Dell’Oro et al., 2020). Wildfires are considered one of the greatest threats to Serbian spruce, and populations of this species have been destroyed by fire several times throughout its long history (Wardle, 1956; Horvat et al., 1974). However, regeneration and expansion of populations occurred after these events, and the remarkable ability of P. omorika seedlings to regenerate after fires has been confirmed by several studies on the species’ ecology and reintroduction after fires (Čolić, 1987; Dinić and Mišić, 1991). A paleoecological study suggests that moderate fire disturbance may have been beneficial to P. omorika, allowing rapid regeneration in the absence of more competent, shade-tolerant species (Finsinger et al., 2017).
Fire events appear to have played an important role in the evolution and ecology of Serbian spruce and likely led to habitat fragmentation and population differentiation (Kuittinen et al., 1991; Gajić et al., 1994; Ballian et al., 2006; Aleksić and Geburek, 2014), which contributed to the development of population-specific traits that may be crucial for the response to fire disturbance. Evidence suggests that a variety of diverse traits enhance fitness after fires (either improving fire survival or promoting recruitment in the post-fire environment), and these adaptive traits vary depending on the fire regime (Keeley and Pausas, 2022). However, many plant traits that are beneficial for fire survival are also useful for adaptation to other selective pressures such as competition, stress and herbivory (Mason et al., 2016; Pausas et al., 2017). A lot of attention has been directed to characterizing tree species based on their flammability and evaluating the relevance of morphological, chemical and ecophysiological traits to flammability (reviewed in Popović et al., 2021). However, there are very few studies of intraspecific variation in plant flammability that either focus on the influence of the fire regime (Pausas et al., 2012; Battersby et al., 2017; Kane et al., 2022) or provide evidence that plant flammability in local populations is an incidental property not selected primarily by fire, at least in environments that have not experienced frequent fire historically (Mason et al., 2016; Cui et al., 2022).
The trait-based approach has been extensively studied in both genetic and environmental contexts, contributing to the development of knowledge in evolutionary and population biology, as well as community and comparative ecology. Intraspecific variability of plant traits (ITV) has become a hot topic in ecology simultaneously with the development of ecological modeling, as the shortcomings of many modeling approaches have become apparent due to a lack of data on ITV (Moran et al., 2016). Studies suggest that selecting relevant traits, describing intraspecific trait variation, and incorporating this variation into models should provide for scaling trait data to community and ecosystem level processes (Violle et al., 2014; Aubin et al., 2016; Funk et al., 2017; Bricca et al., 2022). Morphological, physiological, and phenological traits and their variations have been widely used to explain plant strategies (Reich et al., 2003) and the mechanisms underlying ecosystem responses to environmental change and disturbance (Eviner, 2004). It has been suggested that a deeper understanding of ITV, which accounts for more than 25% of the total variation in leaf functional traits (Albert et al., 2010), could improve understanding of both species’ responses to environmental gradients and community functioning (Martin et al., 2017).
There is a large body of knowledge related to the linkage between plant traits and flammability, and the most commonly examined plant trait has been leaf moisture content, followed by a range of morphological traits (reviewed in Popović et al., 2021). In these studies, a number of species have been tested to establish relationships between traits and flammability, with a tendency to increase the basic knowledge of species fire ecology, but also to provide valuable information for restoration and conservation ecology (Mola et al., 2014; Kane et al., 2019; Krix et al., 2019; Molina et al., 2019). However, more data, including trait intraspecific variability, are needed for our understanding of the relevance of plant traits to flammability, consistency of functional trait covariation from the individual to the ecosystem level and species responses under current and future climate scenarios (Pausas et al., 2017; Alam et al., 2020).
Intraspecies variability of flammability-related traits exists in populations from fire-prone environments with different fire histories, implying that such traits play an adaptive role (Pausas et al., 2012; Battersby et al., 2017; Kane et al., 2022). However, research from forests where no fires were observed over a longer period suggest that intraspecies variability in traits related to flammability may be due to adaptive responses to other environmental disturbances (Mason et al., 2016; Cui et al., 2022). Given that natural populations of P. omorika can be viewed from both perspectives (fire events had an impact on current population differentiation and geographic location is not pronouncedly fire-prone), we hypothesized that ITV in natural populations may have an impact on intraspecies variability of flammability-related traits. The natural populations of P. omorika were selected based on their fire history and overall biogeographic conditions. The goal of our study was to (i) determine intraspecific variability of plant traits related to flammability and (ii) establish relationship between morpho-ecophysiological traits and ignitability.
Materials and methods
Study site
The Tara National Park is a globally significant area of natural values that has largely preserved the character of an autochthonous natural environment. It covers most of the Tara Mountain in the west of Serbia (43° 57′ N, 19° 28′ E). The average altitude is 1,000 to 1,200 m. The climate of Tara Mt. is characterized by fresh to cool summers and quite cold winters. The average annual temperature is 7.9°C and the annual temperature fluctuations are small: January −4.5°C, August 16.7°C. The average annual precipitation is 977 mm, and the average annual humidity is very high (83.4%). Almost all species of boreal dendroflora of Europe and their communities can be found in the extensive area of Tara Mt. between 900 and 1,200 m. Depending on the aspect and slope, all communities that include mixed (deciduous-coniferous) forests and coniferous forests are spread out like a mosaic, which is the basic ecological and biogeographical feature of Tara Mt. Locations inhabited by P. omorika are declared as strict nature reserves or nature monuments (smaller groups or individual trees). P. omorika inhabits steep limestone cliffs or fire pits at elevations ranging from 700 to 1,600 m. The species is found in a variety of communities, usually with spruce, fir, beech, white and black pine and other deciduous species. Seven populations assessed in this study differed by fire history (Kanjon Brusnice was completely destroyed by fire and naturally reestablished to the climax forest community), the presence of water courses (Zmajevački potok and Vranjak are near water courses; Studenac, Bilo, and Trenice do not have surface or underground watercourses), and overall vitality of individuals (drying of trees is observed in Crvene stene). The geographic locations, site and vegetation characteristics of these populations are listed in Table 1. Plant material was collected following the protocol given by Cornelissen et al. (2003).
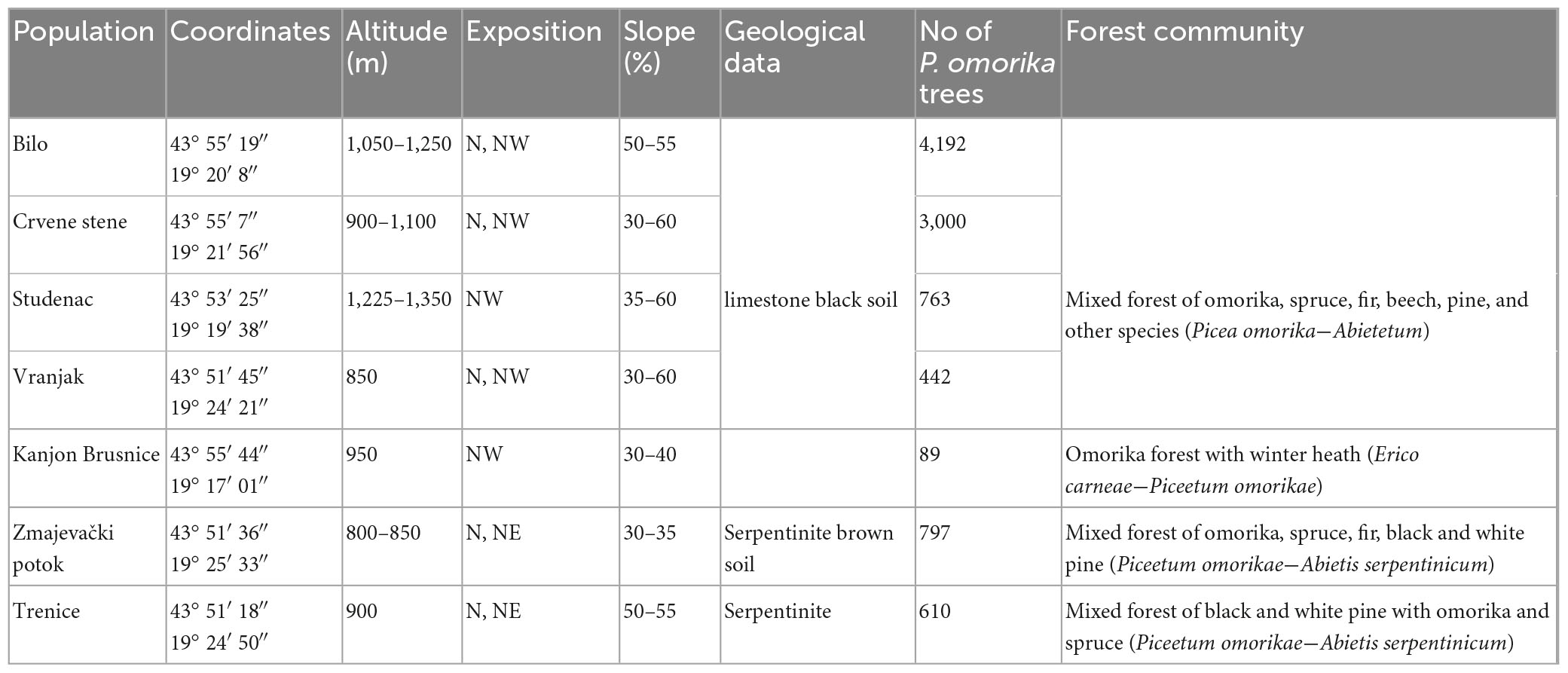
Table 1. Geographical locations, site and vegetation characteristics of seven Picea omorika populations.
Leaf traits measurements
From each population, seven well-grown trees free of pathogen or herbivore damage were selected. Five branches from the outer part of the crown from each tree were sampled (approx. height 4 m) with a telescopic forestry scissors and three twigs with 2-year needles from the same branch were used as a sampling material for (i) leaf trait measurements, (ii) moisture content measurements, and (iii) flammability testing. Leaf trait measurements were conducted on the fresh material (in accordance with testing the fresh-leaf ignitability) and five measurements (from five sampled branches) per tree for needle traits were averaged and used in further analyses. To avoid water loss during transportation from the field to the laboratory, the material from each plant was enclosed in a hermetic plastic bag and stored in an ice box.
The cross-sectional shape of the central part of the needle of P. omorika is generally rhomboidal and can be determined by two diameters: major (needle width) and minor (needle thickness) (Nikolić et al., 2015). These two diameters are noticeable even if the cross-sectional shape is more elliptical (Radovanović et al., 2014). Leaf length (LL, mm) was measured to the nearest millimeter with a standard ruler. Leaf width (LWi, μm) and thickness (LT, μm) were measured in the central part of the needle with a digital micrometer (Käfer Messuhrenfabrik GmbH & Co., Germany). Leaf flatness (LF) was calculated as the ratio between width and thickness (Sellin, 2001). Leaf weight (LWe, g) was measured using a laboratory analytical balance (0.1 mg precision) (Kern, Germany). The needles were scanned and area of an image (scanned area, SA, cm2) was determined in ImageJ program. Specific leaf area (SLA, cm2 g–1) was calculated by dividing the scanned leaf area by its dry weight. The density of leaf tissue was characterized by the density index, DI (g cm–3), which is the ratio between leaf weight and scanned area multiplied by thickness (Sprugel et al., 1996).
The needle volume was measured according to the water displacement principle (Brand, 1987; Chen et al., 1997). The twig containing 30–40 needles was submerged in water and the increase in volume was recorded using a balance (V1). The twig was removed from the water and allowed to drain. The needles were carefully removed and the water displacement procedure was repeated with the bare twig (V2). The difference between V1 and V2 is considered the volume of needles (n = 30–40), and the volume of one needle (V, cm3) was obtained as a V/n.
Leaf moisture content (MC, %) was determined based on the weight of fresh needles (FW) and dry needles (48 h oven-dried at 70°C) (DW), according to calculation (Pellizzaro et al., 2007): MC (%) = (FW−DW)/DW × 100.
Flammability testing
The time-to-ignition (TTI, s) of fresh foliage was measured by 500 W epiradiator with 10 cm radiant disk and nominal surface temperature of 420°C (model 534Rc2, Saint-Gobain Corporation, France). The epiradiator was positioned horizontally, with a pilot flame mounted 4 cm above the center of the disk, which allowed more regular ignition but did not contribute to sample decay (Bianchi and Defossé, 2015). The samples were placed directly on the radiant disk after the device reached the steady-state and the material formed a uniform layer on the epiradiator surface. A total of 245 samples were analyzed consecutively, and each sample contained 3.0 ± 0.1 grams of plant material (Kauf et al., 2015). A digital timer was used to record the time elapsed between placing a sample on the epiradiator surface and the appearance of a flame (TTI). All experiments were conducted in a closed environment.
Statistical analyses
Seven populations of P. omorika were compared on the basis of twelve measured variables: leaf traits (SA/V, SLA, leaf scanned area, weight, thickness, length, width, volume, flatness, density index), moisture content (MC) and time-to-ignition (TTI). The assumption of normality was assessed using the Shapiro–Wilk test; p > 0.05 indicated normal distribution. The differences between populations were tested with ANOVA when the variables appeared to follow normal distribution, or with the non-parametric Kruskal–Wallis test when the assumption of normality was violated. Benjamini and Yekutieli (BY) p-adjustment method was used to control the type I error rate in hypothesis testing. Non-parametric Spearman’s rho correlations were used for the analyses related to plant trait-flammability relationships. Principal component analysis (PCA) was carried out to visualize the data structure. Statistical analyses were performed using R statistical software, version 4.2.1 (R Core Team, 2022).
Results
The seven populations differed in terms of fresh-leaves time-to-ignition (Figure 1). Population IV had the longest TTI, followed by populations I and VII, after which populations II and III, and finally populations V and VI had the shortest TTI. Inter-population differences in morpho-ecophysiological parameters were also significant, but not consistent among populations (Figure 1). The most distinct inter-population differences were found for leaf density index (where all seven populations significantly differed from each other), SLA, SA/V and leaf width (where only two of seven populations did not show a significant difference), and scanned area (with three populations sharing similar values). The smaller inter-population differences were found for MC, leaf thickness, length, weight and volume (four significantly different values were found among seven populations), and leaf flatness (three significantly distinct values were found among seven populations).
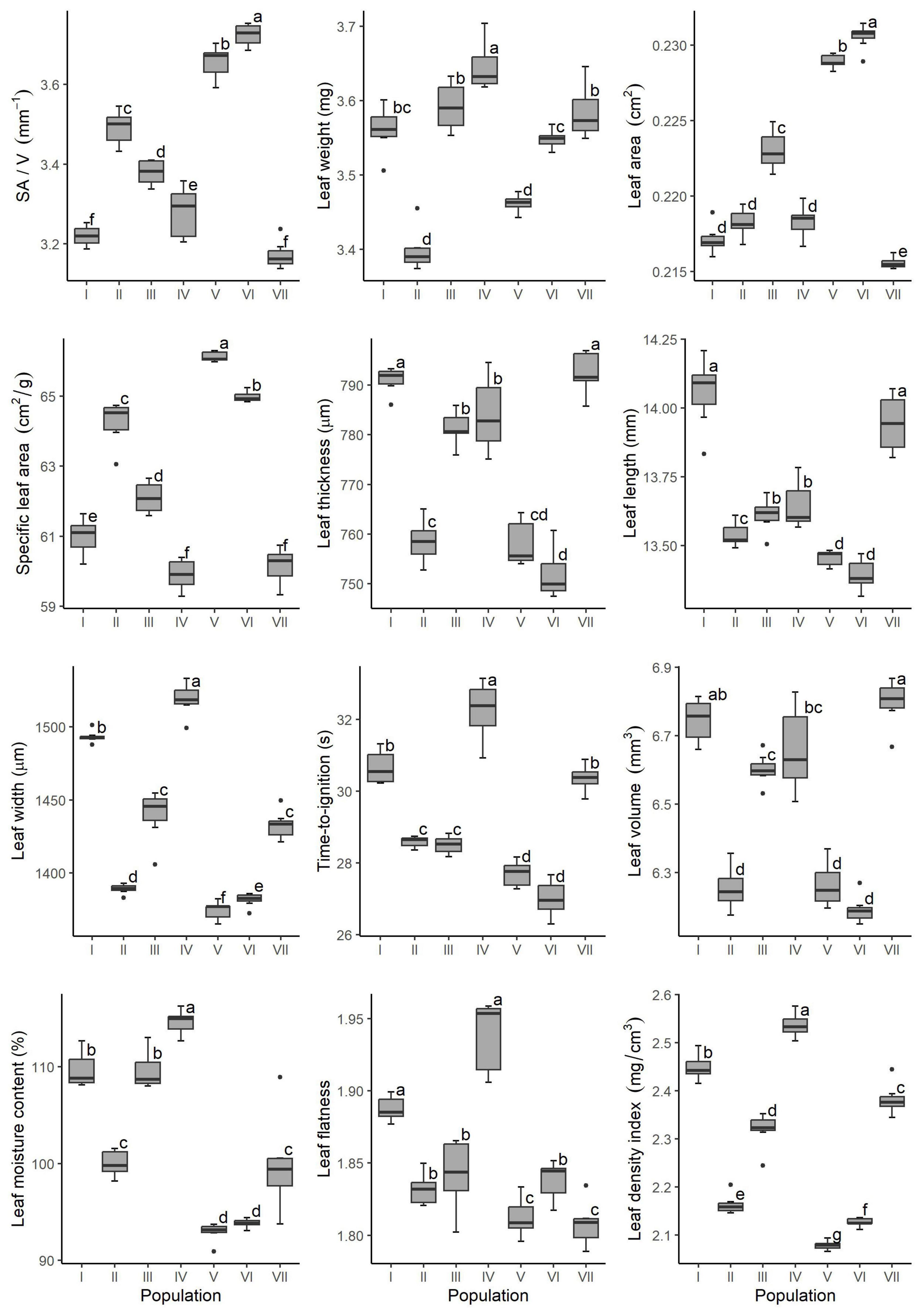
Figure 1. Inter-population differences in morpho-ecophysiological and flammability properties of seven Picea omorika populations based on ANOVA and Kruskal–Wallis test (I, Studenac; II, Vranjak; III, Trenice; IV, Zmajevački potok; V, Crvene stene; VI, Bilo; VII, Kanjon Brusnice).
Time-to-ignition (TTI) was negatively influenced by leaf area-related measures SA/V (ρ = −0.81, p < 0.0001), scanned area (ρ = −0.75, p < 0.0001), and SLA (ρ = −0.87, p < 0.0001), with larger leaves having a shorter time-to-ignition (Figure 2). TTI was positively influenced by the following measures, in order of strength of correlation: density index (ρ = +0.92, p < 0.0001), leaf width (ρ = +0.88, p < 0.0001), MC (ρ = +0.79, p < 0.0001), leaf length (ρ = +0.78, p < 0.0001), thickness (ρ = +0.74, p < 0.0001), volume (ρ = +0.74, p < 0.0001), weight (ρ = +0.59, p < 0.0001), flatness (ρ = +0.52, p < 0.001).
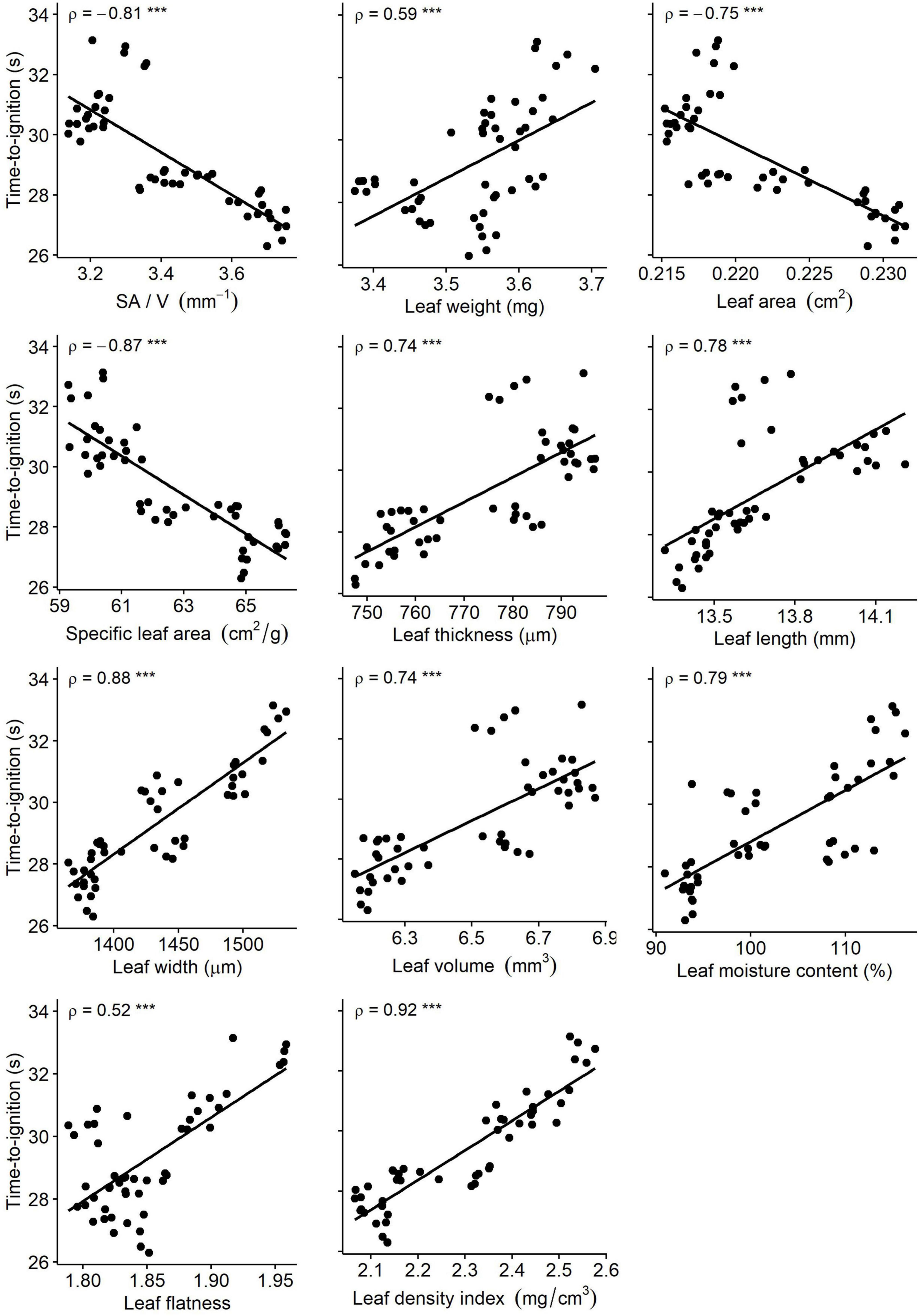
Figure 2. Bivariate relationships between the morpho-ecophysiological traits and time-to-ignition of fresh foliage of Picea omorika. *** Denotes statistically significant correlation, p < 0.001.
Principal component analysis (PCA) classified 49 elements into three distinct groups (Figure 3). The results show that the first two principal components explain 87% of the data variability (74% and 13%, respectively). Based on the sum of squared correlations between the variables and the factors (0.63 < sum of r2 < 0.99), all variables were well represented in the plane of PC1 and PC2. The variability of density index, SLA, leaf width, SA/V, leaf thickness, volume and time-to-ignition (TTI) primarily defined the formation of the first axis, whereas the variability of leaf flatness primarily contributed to the formation of the second axis. The clustering of individuals into several groups is visible in Figure 3. Populations II, V, and VI (higher SA/V and SLA) formed the right-side cluster, while populations I, IV, and VII (higher leaf length, volume, thickness, TTI, density index, width, weight) formed the left-side cluster. Population III was placed in the center, forming a distinct cluster that was closer to populations I, IV, and VII by the majority of parameters. The trait coordination revealed a close relationship between leaf thickness and volume, with both exhibiting similar contribution to the PC1-2 plane. Strong correlations were found between TTI and density index (with DI contributing more to the formation of PC 1-2 plane) and between leaf width and MC (with LWi contributing more to the formation of PC 1-2 plane) (Figure 3). Leaf volume was strongly positively correlated with leaf thickness, and both measures were strongly negatively correlated with SA/V (Figure 3).
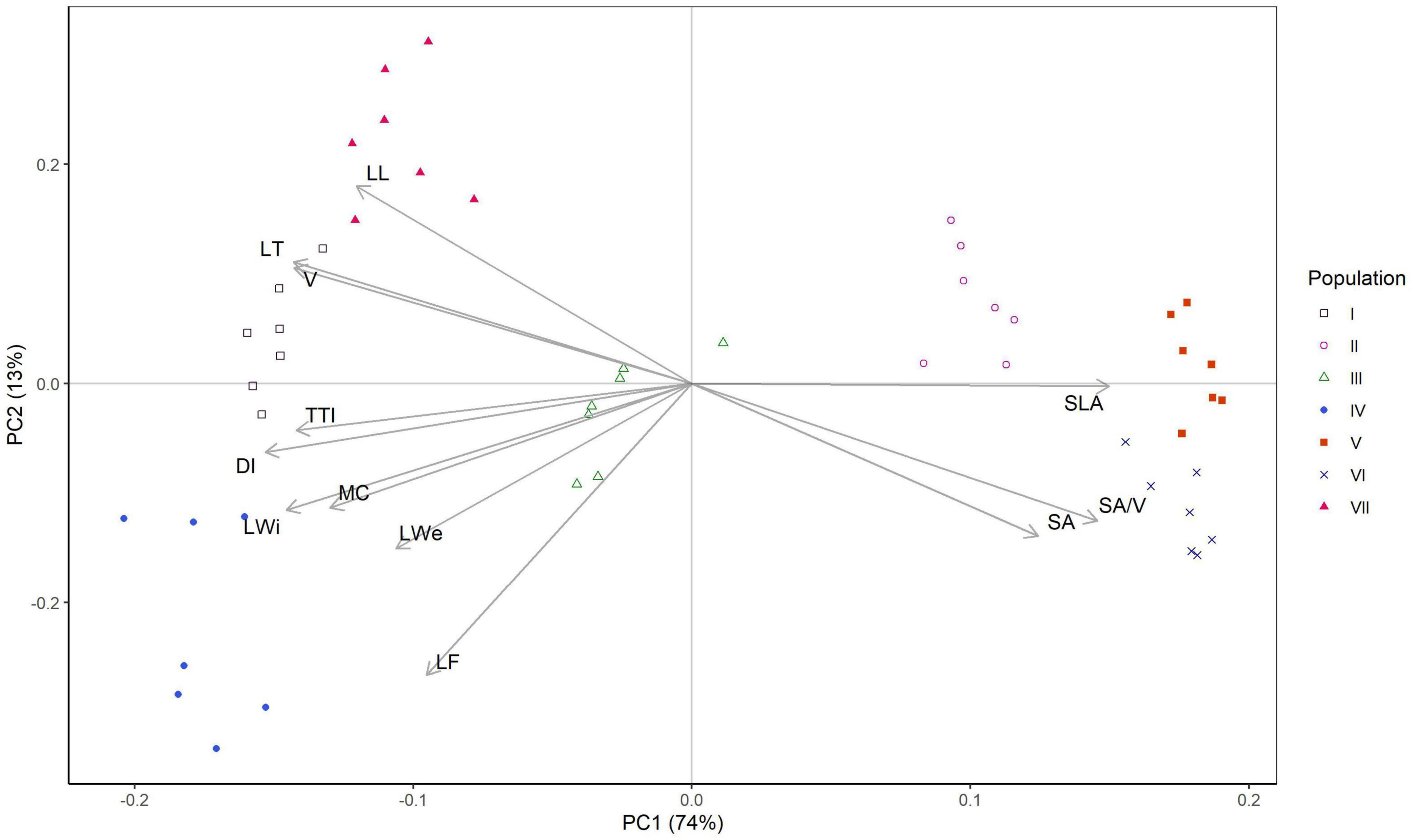
Figure 3. Principal component analysis (PCA) of twelve variables measured from 49 trees of seven Picea omorika populations. Population labels are given in Figure 1.
Discussion
The results presented in this study correspond to previous findings on plant trait-flammability relationships. All measured parameters were significantly correlated with TTI. Leaf density index and TTI had the strongest positive correlation (+0.92). Studies suggest that leaf density index is related to flammability (Burton et al., 2021), and it is also a component of litterbed packing density (a function of tissue density and packing ratio) that strongly influences differences among species and flammability of mixtures (Zhao et al., 2016). TTI and leaf width showed a strong positive correlation (+0.88). Flammability of fresh leaves was found to be significantly correlated with leaf width, with wider and broader leaves having significantly faster ignition times (Murray et al., 2013; Krix and Murray, 2018). Strong negative correlations were found between TTI and both SA/V and SLA (−0.81 and −0.87, respectively). The ratio of leaf area to volume (SA/V), which describes the particle geometry and relative dimensions of the elements of the fuel complex and has a significant influence on the components of flammability (Anderson, 1970), was found to be the most important factor in delaying ignition. Higher values correspond to higher energy and mass transfer rates, resulting in shorter ignition delays and faster fire spread (Anderson, 1970; Dimitrakopoulos and Panov, 2001; Weise et al., 2005; Saura-Mas et al., 2010; Engber and Varner, 2012; Simeoni et al., 2012). Leaf area and SLA were found to be significant predictors of flammability, with larger and wider leaves having a lower time-to-ignition (Murray et al., 2013; Ganteaume, 2018). Leaf thickness, on the other hand, had a negative effect on flammability (increased time-to-ignition) (Grootemaat et al., 2017; Ganteaume, 2018; Romero et al., 2019). Leaf moisture content, the most studied parameter related to flammability and shown to be negatively correlated with TTI (Chuvieco et al., 2004), also had a negative effect on TTI of fresh P. omorika leaves, but with a lower correlation coefficient compared with some morphological measures (e.g., SLA, density index, leaf width).
The leaf moisture content is a primary factor in the canopy ignition process, but being a physiologically based plant character, it can show variations on diurnal and seasonal basis, with leaf age and stand structure (Keyes, 2006). Leaf morphology is the result of a species’ resource-use strategy (Wright et al., 2004) which also influences leaf flammability (Dimitrakopoulos and Papaioannou, 2001; Weise et al., 2005; Engber and Varner, 2012). Leaf thickness, leaf surface area-to-volume ratio, and tissue density, which are all closely related to ignitibility (Grootemaat et al., 2017; Ganteaume, 2018; Romero et al., 2019), also provide information on the strategy of resource utilization in the spectrum of leaf economy (Wright et al., 2004). Thicker and denser leaves, which are less flammable, may be less able to recover from tissue loss during fire, which also occurs following herbivory damage (Michelaki et al., 2020).
Picea omorika populations were generally well separated in the plane of the first two axes (except for the Crvene stene and Bilo populations, where there was a slight overlap). The Studenac and Kanjon Brusnice populations differed from the others by higher values for leaf length, leaf volume, and leaf thickness and lower values for SA/V compared to the other populations. Zmajevački potok population differed from the others by higher values for TTI, density index, leaf weight, MC and leaf width. The populations Crvene stene and Bilo differ from the others by higher values for SLA, SA/V and scanned area, and lower values for leaf length, thickness, volume and density index. The most frequently reported plant trait in plant flammability studies is leaf moisture content, which correlates positively with a longer ignition time and negatively with ignition frequency (reviewed in Popović et al., 2021). Our findings confirmed its impact on TTI, trait coordination between MC and leaf width, and its significant contribution to differentiation of Zmajevački potok population (also distinguished from other populations by higher values of TTI). The high intraspecific variability in almost all examined plant traits supports the usefulness of using the plant trait approach to characterize plant response to disturbance, and more empirical data should be provided for advanced statistical methods with an expanded set of spatial and temporal data (Sturtevant and Fortin, 2021).
The ignitability of plant material is commonly used to compare ignition probability between species and to evaluate the relevance of various traits to ignition (Gill and Moore, 1996; Dimitrakopoulos and Papaioannou, 2001; Pellizzaro et al., 2007; Murray et al., 2013; Dahanayake and Chow, 2018). It is suggested that high variability in ignitability among individuals should lead to variability in the probability of burning and in the spread of fire within and between plants (Pausas et al., 2016). Some general relationships between plant traits and ignition delay have been established by investigating a variety of deciduous and coniferous species (e.g., negative correlation between MC, leaf thickness, and TTI; positive correlation between SA/V, SLA, density index, leaf length, leaf width, leaf area, and TTI) (Shan et al., 2008; Grootemaat et al., 2015; Bianchi et al., 2019; Michelaki et al., 2020). Previous studies on flammability of Picea species found that P. engelmannii stands affected by the spruce beetle had a decrease in MC, higher levels of lignin and cellulose, and lower levels of carbohydrate-based compounds, resulting in increased flammability (Page et al., 2014).
On the territory of Serbia, all sites of P. omorika are preserved in situ. For the sites within the Tara National Park, rigid protection measures (without active intervention) are proposed, resulting in the development of untouched forests. Considering fire events in the near and distant past, burned populations can be expected to recover through passive restoration, i.e., natural rejuvenation and succession. In addition, all post-fire biological legacies (structures, organisms, patterns, and processes that constitute continuity between pre- and post-disturbance ecosystems) (Franklin et al., 2000) have been integrated into the recovering ecosystem. Evidence suggests that naturally regenerating forest ecosystems exhibit greater resilience and adaptive capacity compared to those that have been restored through intervention (Blumroeder et al., 2022). The Kanjon Brusnice population, which was established after the great fire in 1946, is the only one that has been restored with autochthonous vegetation in the burned area within Tara National Park. Heather indicates a progressive succession (common in areas with frequent fires) from a polydominant forest type to the final stage of a Piceetum omorikae mixtum forest type. There was no evidence of a fire history in the other six populations in our data set. Some sites are near watercourses (Zmajevački potok, Vranjak), while others do not have surface or underground watercourses (Studenac, Bilo, Trenice), and the drying of trees is observed in Crvene stene location. However, regardless of the fire history of the stand, significant differences in morpho-ecophysiological traits and ignitibility were observed in the total dataset.
In studies of vegetation flammability, a trait-based approach is considered useful for upscaling attributes of the fire regime, such as spatial patterns of fire severity (Stevens et al., 2020). However, the predictive potential of data based on species mean values is limited at the community and ecosystem level unless interactions among species, habitat resource availability, niche and trait overlap among species, and intraspecific trait variation are accounted for Violle et al. (2012), Fréjaville et al. (2018), and Michelaki et al. (2020). According to studies from fire-prone communities, ITV is greater than interspecific trait variability (Dantas et al., 2013), it is the primary response to altered disturbance regimes (Mitchell et al., 2021), and it should be taken into account to improve mechanistic understanding of plant community structure rules (Cianciaruso et al., 2012). ITV has also been shown to account for a significant proportion of the variance in fire resistance traits in open and closed forests in the sub-Mediterranean region, as well as in open subalpine forests and moist montane forests (Fréjaville et al., 2018).
Intraspecific variability of plant traits (ITV) is thought to be caused by genotypic variation (drift, developmental noise, or random mutations) and phenotypic plasticity (variation in trait expression within genotypes) (Scheiner, 1993). The majority of studies within natural populations reveal ITV based on either phenotypic plasticity or local genetic adaptation. However, the main shortcoming in ITV studies of natural populations is that only one approach is used, and both the extent of gene flow and the degree of environmental heterogeneity should be considered (Via and Lande, 1985). The differences in morpho-ecophysiological parameters between populations of P. omorika can be related to the findings on genetic diversity, which suggest that populations only a few kilometers or less apart were poorly connected and highly differentiated over the glacial and post-glacial periods and characterized as independent gene pools at the nuclear DNA level (Aleksić and Geburek, 2010). These findings suggest that fragmentation, admixture, size reductions/expansions and extinction contributed to a continuous increase in the genetic distinctness of populations over the large time periods, decreased gene flow, and almost completely excluded seed flow between populations (Aleksić and Geburek, 2010). The results presented herein suggest that the population differences are a source of ITV, which can be a starting point toward identifying generalities and the underlying mechanisms that shape ITV (Westerband et al., 2021).
Disturbances are a natural and integral part of forest ecosystems and are usually followed by forest succession (Dale et al., 2000). Fires, which are among the key disturbances in forest ecosystems, play an important role in fire-prone ecosystems, where various species- and community-level adaptations, including flammability, to fire regimes can be observed (Foster et al., 1998). The search for similar patterns in forest ecosystems from areas without frequent fire has suggested that other environmental filters have shaped plants adaptive traits, and traits related to flammability may be considered a secondary byproduct of selection for other traits (Mason et al., 2016; Fréjaville et al., 2018; Cui et al., 2022). There is open debate about the adaptive importance of plant traits related to flammability in different areas where fire does or does not exert strong selection pressure (Bradshaw et al., 2011; Keeley et al., 2011; Bowman et al., 2014). However, given that projected climate change may favor fire-promoting environments and alter fire regimes (Moritz et al., 2014), there is strong interest in expanding global knowledge of the relationship between fire and forest ecosystems (FAO and UNEP, 2020).
While strategic forest management focuses on fire prevention and suppression through the development of advanced detection technologies (FAO, 2007), less emphasis is placed on plant trait approaches, which should be widely used and supplemented with more data on ITV (Harrison et al., 2021). This study provides evidence for population-based intraspecific variability of plant traits (morpho-ecophysiological traits and flammability) in P. omorika, as well as relationships between traits that can be useful in future modeling projects.
Conclusion
Although trait analyses have been recognized as useful for evaluating the fire responses of a variety of species, the studies have primarily focused on average species trait values and assumed that intraspecific variation is small or negligible. However, the need for evidence on ITV in plant trait-flammability studies has raised along with the need for more empirical data in modeling. According to this study, population differences are a source of ITV (morpho-ecophysiological traits and ignitability). There is strong evidence that the high genetic diversity of P. omorika natural populations is due to fragmentation, admixture, size reductions/expansions, and extinction, which all contribute to the genetic distinctness of populations. Given that this study focused on ITV of flammability-related traits, it can help to identify underlying mechanisms that affect ITV in ecosystems that are not classified as fire-prone. It may also contribute to the promotion of plant trait approaches that should be widely used in future modeling projects.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
ZP conceived the ideas, designed methodology, and wrote the manuscript. ZP and JJ conducted the fieldwork, material sampling, and lab analyses. VV analyzed data and contributed to the writing of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was financed by the Rufford Fund for Nature Conservation, small project granted to JJ (316924/2021-0) and the Ministry of Science, Technological Development and Innovation, Republic of Serbia (contract number 451-03-47/2023-01/200007).
Acknowledgments
We are thankful to the management and employees of National Park Tara for their assistance in fieldwork. Also, helpful comments by two reviewers referees are gratefully acknowledged.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
DI (g cm–3), density index; ITV, intraspecific variability of plant traits; (SA/V), lear area to volume ratio; (LL, mm), leaf length; (LWi, μ m), leaf width; (LT, μ m), leaf thickness; LF, leaf flatness; (LWe, g), leaf weight; (SA, cm2), leaf scanned area; (MC, %), leaf moisture content; (V, cm3), leaf volume; (SLA, cm2 g–1), specific leaf area; (TTI, s), time-to-ignition.
Footnotes
References
Alam, M. A., Wyse, S. V., Buckley, H. L., Perry, G. L., Sullivan, J. J., Mason, N. W., et al. (2020). Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits. J. Ecol. 108, 641–653. doi: 10.1111/1365-2745.13289
Albert, C. H., Thuiller, W., Yoccoz, N. G., Soudant, A., Boucher, F., Saccone, P., et al. (2010). Intraspecific functional variability: extent, structure and sources of variation. J. Ecol. 98, 604–613. doi: 10.1111/j.1365-2745.2010.01651.x
Aleksić, J. M., and Geburek, T. (2010). Mitochondrial DNA reveals complex genetic structuring in a stenoendemic conifer Picea omorika [(Panč.) Purk.] caused by its long persistence within the refugial Balkan region. Plant Syst. Evol. 285, 1–11. doi: 10.1007/s00606-009-0250-0
Aleksić, J. M., and Geburek, T. (2014). Quaternary population dynamics of an endemic conifer, Picea omorika, and their conservation implications. Conserv. Genet. 15, 87–107. doi: 10.1007/s10592-013-0523-6
Aubin, I., Munson, A. D., Cardou, F., Burton, P. J., Isabel, N., Pedlar, J. H., et al. (2016). Traits to stay, traits to move: a review of functional traits to assess sensitivity and adaptive capacity of temperate and boreal trees to climate change. Environ. Rev. 24, 164–186. doi: 10.1139/er-2015-0072
Ballian, D., Longauer, R., Mikić, T., Paule, L., Kajba, D., and Gömöry, D. (2006). Genetic structure of a rare European conifer, Serbian spruce (Picea omorika (Panč.) Purk.). Plant Syst. Evol. 260, 53–63. doi: 10.1007/s00606-006-0431-z
Battersby, P. F., Wilmshurst, J. M., Curran, T. J., McGlone, M. S., and Perry, G. L. (2017). Exploring fire adaptation in a land with little fire: serotiny in Leptospermum scoparium (Myrtaceae). J. Biogeogr. 44, 1306–1318. doi: 10.1111/jbi.12950
Bianchi, L. O., and Defossé, G. E. (2015). Live fuel moisture content and leaf ignition of forest species in Andean Patagonia, Argentina. Int. J. Wildland Fire 24, 340–348. doi: 10.1071/WF13099
Bianchi, L. O., Oddi, F. J., Muñoz, M., and Defossé, G. E. (2019). Comparison of leaf moisture content and ignition characteristics among native species and exotic conifers in Northwestern Patagonia, Argentina. For. Sci. 65, 375–386. doi: 10.1093/forsci/fxy054
Blumroeder, J. S., Schmidt, F., Gordon, A., Grosse, S., and Ibisch, P. L. (2022). Ecosystemic resilience of a temperate post-fire forest under extreme weather conditions. Front. For. Glob. Change 5:1070958. doi: 10.3389/ffgc.2022.1070958
Bowman, D., French, B., and Prior, L. D. (2014). Have plants evolved to self-immolate? Front. Plant Sci. 5:590. doi: 10.3389/fpls.2014.00590
Bradshaw, S. D., Dixon, K. W., Hopper, S. D., Lambers, H., and Turner, S. R. (2011). Little evidence for fire-adapted plant traits in Mediterranean climate regions. Trends Plant Sci. 16, 69–76. doi: 10.1016/j.tplants.2010.10.007
Brand, D. G. (1987). Estimating the surface area of spruce and pine foliage from displaced volume and length. Can. J. For. Res. 17, 1305–1308. doi: 10.1139/x87-203
Bricca, A., Di Musciano, M., Ferrara, A., Theurillat, J. P., and Cutini, M. (2022). Community assembly along climatic gradient: contrasting pattern between-and within-species. Perspect. Plant Ecol. Evol. Syst. 56:125675. doi: 10.1016/j.ppees.2022.125675
Burton, J. E., Cawson, J. G., Filkov, A. I., and Penman, T. D. (2021). Leaf traits predict global patterns in the structure and flammability of forest litter beds. J. Ecol. 109, 1344–1355. doi: 10.1111/1365-2745.13561
Chen, J. M., Rich, P. M., Gower, S. T., Norman, J. M., and Plummer, S. (1997). Leaf area index of boreal forests: theory, techniques, and measurements. J. Geophys. Res.: Atmos. 102, 29429–29443. doi: 10.1029/97JD01107
Chuvieco, E., Aguado, I., and Dimitrakopoulos, A. P. (2004). Conversion of fuel moisture content values to ignition potential for integrated fire danger assessment. Can. J. For. Res. 34, 2284–2293. doi: 10.1139/x04-101
Cianciaruso, M. V., Silva, I. A., Batalha, M. A., Gaston, K. J., and Petchey, O. L. (2012). The influence of fire on phylogenetic and functional structure of woody savannas: moving from species to individuals. Perspect. Plant Ecol. Evol. Syst. 14, 205–216. doi: 10.1016/j.ppees.2011.11.004
Čolić, D. B. (1987). Spontana obnova Pančićeve omorike (Picea omorika Panč.) posle požara [Spontaneous regeneration of Serbian spruce (Picea omorika Panč.) after the fire]. Zaštita Prirode 40, 37–56.
Cornelissen, J. H. C., Lavorel, S., Garnier, E., Díaz, S., Buchmann, N., Gurvich, D. E., et al. (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 51, 335–380. doi: 10.1071/BT02124
Cui, X., Paterson, A. M., Perry, G. L., Wyse, S. V., Alam, M. A., Huang, C., et al. (2022). Intraspecific variation in shoot flammability in Dracophyllum rosmarinifolium is not predicted by habitat environmental conditions. For. Ecosyst. 9:100017. doi: 10.1016/j.fecs.2022.100017
Dahanayake, K. C., and Chow, C. L. (2018). Moisture content, ignitability, and fire risk of vegetation in vertical greenery systems. Fire Ecol. 14, 125–142. doi: 10.4996/fireecology.140112514
Dale, V. H., Joyce, L. A., McNulty, S., and Neilson, R. P. (2000). The interplay between climate change, forests, and disturbances. Sci. Total Environ. 262, 201–204. doi: 10.1016/S0048-9697(00)00522-2
Dantas, V. D. L., Pausas, J. G., Batalha, M. A., Loiola, P. D. P., and Cianciaruso, M. V. (2013). The role of fire in structuring trait variability in Neotropical savannas. Oecologia 171, 487–494. doi: 10.1007/s00442-012-2431-8
Dell’Oro, M., Mataruga, M., Sass-Klaassen, U., and Fonti, P. (2020). Climate change threatens on endangered relict Serbian spruce. Dendrochronologia 59:125651. doi: 10.1016/j.dendro.2019.125651
Dimitrakopoulos, A. P., and Panov, P. I. (2001). Pyric properties of some dominant Mediterranean vegetation species. Int. J. Wildland Fire 10, 23–27. doi: 10.1071/WF01003
Dimitrakopoulos, A. P., and Papaioannou, K. K. (2001). Flammability assessment of mediterranean forest fuels. Fire Technol. 37, 143–152. doi: 10.1023/A:1011641601076
Dinić, A., and Mišić, V. (1991). Značaj rezultata dvadesetogodišnjih eksperimentalinih fitocenoloških istraživanja šumskih ekosistema za šumarsku praksu [Significance of results of the twenty-year experimental phytocoenological investigations of forest ecosystems for the forestry practice]. Bull. Fac. For. Univ. Belgrade 73, 105–112.
Engber, E. A., and Varner, J. M. (2012). Patterns of flammability of the California oaks: the role of leaf traits. Can. J. For. Res. 42, 1965–1975. doi: 10.1139/x2012-138
Eviner, V. T. (2004). Plant traits that influence ecosystem processes vary independently among species. Ecology 85, 2215–2229. doi: 10.1890/03-0405
FAO (2007). Fire Management – Global Assessment 2006: A Thematic Study Prepared in the Framework of the Global Forest Resources Assessment 2005. Rome: Food and Agriculture Organization of the United Nations.
FAO, and UNEP (2020). The State of the World’s Forests 2020: Forests, Biodiversity and People. Rome: Food and Agriculture Organization of the United Nations and United Nations Environment Programme.
Finsinger, W., Morales-Molino, C., Gałka, M., Valsecchi, V., Bojovic, S., and Tinner, W. (2017). Holocene vegetation and fire dynamics at Crveni Potok, a small mire in the Dinaric Alps (Tara National Park, Serbia). Quat. Sci. Rev. 167, 63–77. doi: 10.1016/j.quascirev.2017.04.032
Foster, D. R., Knight, D. H., and Franklin, J. F. (1998). Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1, 497–510. doi: 10.1007/s100219900046
Franklin, J. F., Lindenmayer, D., Macmahon, J. A., Mckee, A., Perry, D. A., Waide, R., et al. (2000). Threads of continuity: ecosystem disturbance, recovery, and the theory of biological legacies. Conserv. Pract. 1, 8–17. doi: 10.1111/j.1526-4629.2000.tb00155.x
Fréjaville, T., Vilà-Cabrera, A., Curt, T., and Carcaillet, C. (2018). Aridity and competition drive fire resistance trait covariation in mountain trees. Ecosphere 9:e02493. doi: 10.1002/ecs2.2493
Funk, J. L., Larson, J. E., Ames, G. M., Butterfield, B. J., Cavender-Bares, J., Firn, J., et al. (2017). Revisiting the Holy Grail: using plant functional traits to understand ecological processes. Biol. Rev. 92, 1156–1173. doi: 10.1111/brv.12275
Gajić, M., Vilotić, D., Karadžić, D., Mihajlović, L., and Isajev, V. (1994). Omorika – Picea omorika (Pančić) Purkyne na Području Nacionalnog Parka Tara (monografska studija). Beograd: Šumarski fakultet.
Ganteaume, A. (2018). Does plant flammability differ between leaf and litter bed scale? Role of fuel characteristics and consequences for flammability assessment. Int. J. Wildland Fire 27, 342–352. doi: 10.1071/WF17001
Gill, A. M., and Moore, P. H. (1996). Ignitibility of Leaves of Australian Plants,” in Contract Report to the Australian Flora Foundation. Canberra, NSW: CSIRO.
Grootemaat, S., Wright, I. J., van Bodegom, P. M., and Cornelissen, J. H. (2017). Scaling up flammability from individual leaves to fuel beds. Oikos 126, 1428–1414. doi: 10.1111/oik.03886
Grootemaat, S., Wright, I. J., van Bodegom, P. M., Cornelissen, J. H., and Cornwell, W. K. (2015). Burn or rot: leaf traits explain why flammability and decomposability are decoupled across species. Funct. Ecol. 29, 1486–1497. doi: 10.1111/1365-2435.12449
Harrison, S. P., Prentice, I. C., Bloomfield, K. J., Dong, N., Forkel, M., Forrest, M., et al. (2021). Understanding and modelling wildfire regimes: an ecological perspective. Environ. Res. Lett. 16:125008. doi: 10.1088/1748-9326/ac39be
Horvat, I., Glavac, V., and Ellenberg, H. (1974). Vegetation of Southeast Europe. Stuttgart: Gustav Fischer Verlag.
Kane, J. M., Gallagher, M. R., Varner, J. M., and Skowronski, N. S. (2022). Evidence of local adaptation in litter flammability of a widespread fire-adaptive pine. J. Ecol. 110, 1138–1148.
Kane, J. M., Varner, J. M., and Saunders, M. R. (2019). Resurrecting the lost flames of American chestnut. Ecosystems 22, 995–1006. doi: 10.1007/s10021-018-0318-1
Kauf, Z., Fangmeier, A., Rosavec, R., and Španjol, Ž (2015). Seasonal and local differences in leaf litter flammability of six Mediterranean tree species. Environ. Manage 55, 687–701. doi: 10.1007/s00267-014-0427-3
Keeley, J. E., and Pausas, J. G. (2022). Evolutionary ecology of fire. Annu. Rev. Ecol. Evol. Syst. 53, 203–225. doi: 10.1146/annurev-ecolsys-102320-095612
Keeley, J. E., Pausas, J. G., Rundel, P. W., Bond, W. J., and Bradstock, R. A. (2011). Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 16, 406–411. doi: 10.1016/j.tplants.2011.04.002
Keyes, C. R. (2006). “Foliar moisture contents of North American conifers,” in Fuels Management-how to Measure Success: Conference Proceedings. 2006 March 28-30; Portland, OR. Proceedings RMRS-P-41, Comps, eds P. L. Andrews and B. W. Butler (Fort Collins, CO: USDA Forest Service, Rocky Mountain Research Station), 395–399. doi: 10.1002/ecy.2824
Krix, D. W., and Murray, B. R. (2018). Landscape variation in plant leaf flammability is driven by leaf traits responding to environmental gradients. Ecosphere 9:e02093. doi: 10.1002/ecs2.2093
Krix, D. W., Phillips, M. L., and Murray, B. R. (2019). Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland–urban interface. Int. J. Wildland Fire 28, 295–307. doi: 10.1071/WF18167
Kuittinen, H., Muona, O., Kärkkäinen, K., and Borzan, Ž (1991). Serbian spruce, a narrow endemic, contains much genetic variation. Can. J. For. Res. 21, 363–367. doi: 10.1139/x91-044
Martin, A. R., Rapidel, B., Roupsard, O., Van den Meersche, K., de Melo Virginio Filho, E., et al. (2017). Intraspecific trait variation across multiple scales: the leaf economics spectrum in coffee. Funct. Ecol. 31, 604–612. doi: 10.1111/1365-2435.12790
Mason, N. W., Frazao, C., Buxton, R. P., and Richardson, S. J. (2016). Fire form and function: evidence for exaptive flammability in the New Zealand flora. Plant Ecol. 217, 645–659. doi: 10.1007/s11258-016-0618-5
Michelaki, C., Fyllas, N. M., Galanidis, A., Aloupi, M., Evangelou, E., Arianoutsou, M., et al. (2020). Adaptive flammability syndromes in thermoMediterranean vegetation, captured by alternative resource-use strategies. Sci. Total Environ. 718:137437. doi: 10.1016/j.scitotenv.2020.137437
Mitchell, R. M., Ames, G. M., and Wright, J. P. (2021). Intraspecific trait variability shapes leaf trait response to altered fire regimes. Ann. Bot. 127, 543–552. doi: 10.1093/aob/mcaa179
Mola, J. M., Varner, J. M., Jules, E. S., and Spector, T. (2014). Altered community flammability in Florida’s apalachicola ravines and implications for the persistence of the endangered conifer Torreya taxifolia. PLoS One 9:e103933. doi: 10.1371/journal.pone.0103933
Molina, J. R., Lora, A., Prades, C., and y Silva, F. R. (2019). Roadside vegetation planning and conservation: new approach to prevent and mitigate wildfires based on fire ignition potential. For. Ecol. Manage 444, 163–117. doi: 10.1016/j.foreco.2019.04.034
Moran, E. V., Hartig, F., and Bell, D. M. (2016). Intraspecific trait variation across scales: implications for understanding global change responses. Global Change Biol. 22, 137–150. doi: 10.1111/gcb.13000
Moritz, M. A., Batllori, E., Bradstock, R. A., Gill, A. M., Handmer, J., Hessburg, P. F., et al. (2014). Learning to coexist with wildfire. Nature 515, 58–66. doi: 10.1038/nature13946
Murray, B. R., Hardstaff, L. K., and Phillips, M. L. (2013). Differences in leaf flammability, leaf traits and flammability-trait relationships between native and exotic plant species of dry sclerophyll forest. PLoS One 8:e79205. doi: 10.1371/journal.pone.0079205
Nikolić, B., Bojović, S., and Marin, P. D. (2015). Variability of morpho-anatomical characteristics of the needles of Picea omorika from natural populations in Serbia. Plant Biosyst. 149, 61–67. doi: 10.1080/11263504.2013.810180
Page, W. G., Jenkins, M. J., and Runyon, J. B. (2014). Spruce beetle-induced changes to Engelmann spruce foliage flammability. For. Sci. 60, 691–702. doi: 10.5849/forsci.13-050
Pausas, J. G., Alessio, G. A., Moreira, B., and Corcobado, G. (2012). Fires enhance flammability in Ulex parviflorus. New Phytol. 193, 18–23.
Pausas, J. G., Alessio, G. A., Moreira, B., and Segarra-Moragues, J. G. (2016). Secondary compounds enhance flammability in a Mediterranean plant. Oecologia 180, 103–110. doi: 10.1007/s00442-015-3454-8
Pausas, J. G., Keeley, J. E., and Schwilk, D. W. (2017). Flammability as an ecological and evolutionary driver. J. Ecol. 105, 289–297. doi: 10.1111/1365-2745.12691
Pellizzaro, G., Duce, P., Ventura, A., and Zara, P. (2007). Seasonal variations of live moisture content and ignitability in shrubs of the Mediterranean Basin. Int. J. Wildland Fire 16, 633–641. doi: 10.1071/WF05088
Popović, Z., Bojović, S., Marković, M., and Cerdà, A. (2021). Tree species flammability based on plant traits: a synthesis. Sci. Total Environ. 800:149625. doi: 10.1016/j.scitotenv.2021.149625
Radovanović, B., Šinžar-Sekulić, J., Rakić, T., Živković, I., and Lakušić, D. (2014). Variation in needle anatomy of Picea omorika (Pinaceae) plants belonging to different gene pools in natural populations on Tara Mt. Serbia. Bot. Serbica 38, 237–246.
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). The evolution of plant functional variation: traits, spectra, and strategies. Int. J. Plant Sci. 164, S143–S164. doi: 10.1086/374368
Romero, B., Fernandez, C., Lecareux, C., Ormeño, E., and Ganteaume, A. (2019). How terpene content affects fuel flammability of wildland–urban interface vegetation. Int. J. Wildland Fire 28, 614–627. doi: 10.1071/WF18210
Saura-Mas, S., Paula, S., Pausas, J. G., and Lloret, F. (2010). Fuel loading and flammability in the Mediterranean Basin woody species with different post-fire regenerative strategies. Int. J. Wildland Fire 19, 783–794. doi: 10.1071/WF09066
Scheiner, S. M. (1993). Genetics and evolution of phenotypic plasticity. Annu. Rev. Ecol. Evol. Syst. 24, 35–68.
Sellin, A. (2001). Morphological and stomatal responses of Norway spruce foliage to irradiance within a canopy depending on shoot age. Environ. Exp. Bot. 45, 115–131. doi: 10.1016/s0098-8472(00)00086-1
Shan, Y., Liu, N., and Shu, L. (2008). Flammability ranking of foliage species by factor analysis of physical and chemical pyric properties. Fire Mater. 32, 371–382. doi: 10.1002/fam.974
Simeoni, A., Thomas, J. C., Bartoli, P., Borowieck, P., Reszka, P., Colella, F., et al. (2012). Flammability studies for wildland and wildland–urban interface fires applied to pine needles and solid polymers. Fire Saf. J. 54, 203–217. doi: 10.1016/j.firesaf.2012.08.005
Sprugel, D. G., Brooks, J. R., and Hinckley, T. M. (1996). Effects of light on shoot geometry and needle morphology in Abies amabilis. Tree Physiol. 16, 91–98. doi: 10.1093/treephys/16.1-2.91
Stevens, J. T., Kling, M. M., Schwilk, D. W., Varner, J. M., and Kane, J. M. (2020). Biogeography of fire regimes in western US conifer forests: a trait-based approach. Global Ecol. Biogeogr. 29, 944–955. doi: 10.1111/geb.13079
Sturtevant, B. R., and Fortin, M. J. (2021). Understanding and modeling forest disturbance interactions at the landscape level. Front. Ecol. Evol. 9:653647. doi: 10.3389/fevo.2021.653647
Via, S., and Lande, R. (1985). Genotype-environment interaction and the evolution of phenotypic plasticity. Evolution 39, 505–522. doi: 10.1111/j.1558-5646.1985.tb00391.x
Violle, C., Enquist, B. J., McGill, B. J., Jiang, L., Albert, C. H., Hulshof, C., et al. (2012). The return of the variance: intraspecific variability in community ecology. Trends Ecol. Evol. 27, 244–252. doi: 10.1016/j.tree.2011.11.014
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., and Kattge, J. (2014). The emergence and promise of functional biogeography. Proc. Natl. Acad. Sci. U S A. 111, 13690–13696. doi: 10.1073/pnas.1415442111
Wardle, P. (1956). Picea omorika in its natural habitat. Forestry 29, 91–117. doi: 10.1093/forestry/29.2.91
Weise, D. R., White, R. H., Beall, F. C., and Etlinger, M. (2005). Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation. Int. J. Wildland Fire 14, 321–338. doi: 10.1071/WF04035
Westerband, A. C., Funk, J. L., and Barton, K. E. (2021). Intraspecific trait variation in plants: a renewed focus on its role in ecological processes. Ann. Bot. 127, 397–410. doi: 10.1093/aob/mcab011
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature 428, 821–827. doi: 10.1038/nature02403
Keywords: leaf morphology, leaf moisture content, ignitability, trait variability, intraspecific differences
Citation: Popović Z, Vidaković V and Janković J (2023) Variability of leaf traits in natural populations of Picea omorika determines ignitability of fresh foliage. Front. For. Glob. Change 6:1196809. doi: 10.3389/ffgc.2023.1196809
Received: 30 March 2023; Accepted: 30 May 2023;
Published: 12 June 2023.
Edited by:
Chad M. Hoffman, Colorado State University, United StatesReviewed by:
Jose Climent, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), SpainAlexandros P. Dimitrakopoulos, Aristotle University of Thessaloniki, Greece
Copyright © 2023 Popović, Vidaković and Janković. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zorica Popović, em9yaWNhakBpYmlzcy5iZy5hYy5ycw==