 Muhammad Fahad Sardar
Muhammad Fahad Sardar Fazila Younas3
Fazila Younas3 Zia Ur Rahman Farooqi
Zia Ur Rahman Farooqi Yanli Li
Yanli Li
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. For. Glob. Change , 12 May 2023
Sec. Forest Soils
Volume 6 - 2023 | https://doi.org/10.3389/ffgc.2023.1144930
This article is part of the Research Topic The Role of Soil Organic Nitrogen in Forest Plant Nutrition View all 6 articles
Nitrogen (N) is an important component of the forest soils and plays a pivotal role in the ecosystem’s health, also in broadleaf and Moso bamboo (Phyllostachys edulis) forests. Nitrogen plays numerous functions in the production and transformation of soil organic matter (SOM) and alterations in the soil’s physicochemical and biological properties. Despite the extensive research on soil properties and microorganism diversity in broadleaf and Moso bamboo forests, we still know very little about N dynamics and its significance in broadleaf and Moso bamboo forests, or how various physical and environmental variables influence soil physicochemical, N dynamics, and biological attributes. This article provides a thorough overview of the role of N dynamics in broadleaf and Moso bamboo forests and changes in soil physicochemical properties processes by summarizing recent advances in our knowledge of forest soil microbial diversity, and carbon (C) and N sink in broadleaf and Moso bamboo forests. As broadleaf and Moso bamboo forests are very sensitive to little change, even a small change in these ecosystems can alter the overall N dynamics. Here, we dissect the soil ecology of broadleaf and Moso bamboo forests to provide insights into the possibilities and consequences of future studies of N dynamics in these ecosystems.
Due to the function in the production and transformation of soil organic matter and transformation of forest soil physicochemical and biological attributes, Nitrogen (N) plays a pivotal role and important component of the forest soil ecosystem’s health, especially in broadleaf and Moso bamboo (Phyllostachys edulis). Soils contain less than 1% of the world’s total N, while other is stored in the atmosphere and sedimentary rocks that are thus unavailable to plants (Follett and Hatfield, 2001), which ultimately affect the forest tree development. Litterfall, biological N fixation (BNF), recycling of tree leftovers, and microbial transformations and bodies are all many of the N sources in forests (Sponseller et al., 2016; Tang et al., 2018), while the N cycle is also governed by these factors in addition to soil organic matter (SOM) breakdown via climatic conditions and/or forest fires, etc. The mineralization-immobilization turnover is one process in the N cycle that is directly influenced by the N, which in turn modifies a variety of other ecosystem processes and functions, such as the soil nutrient cycles, i.e., N cycle and forests ecosystem functionality of the forests (Cheng et al., 2019; Kurniawan et al., 2019). The amount of N released during SOM decomposition directly influences the amount of N consumed by soil microorganisms, and then the mineralization rate. In the N cycle of the forest soils, nitrification/denitrification and ammonification are important key steps. Ammonia (NH3) is converted to nitrite (NO2–) by ammonia-oxidizing archaea and ammonia-oxidizing bacteria, while nitrite (NO2–) is converted to nitrate (NO3–) by nitrite-oxidizing bacteria (Isobe et al., 2018a).
Many crucial ecological and physiological processes in forest ecosystems rely on the activities of soil microbial communities, including N and SOM turnover, the control of mineral nutrient availability, the fixation of atmospheric N, and the development of mycorrhiza (Zhou et al., 2020; Matos et al., 2021; Wang et al., 2022a). Soil microbial populations produce hydrolases to break down plant litter and other organic materials (Tan et al., 2020), recycling nutrients and promoting plant growth as part of the nutrient cycle process in forest environments (Prasad et al., 2021). Soil microbial communities, through controlling the rate of microbial decomposition of SOM, may influence soil attributes including nutrient concentration (Kooch and Noghre, 2020), and hydrolase activity (Wu et al., 2021). Multi-dimensional soil characteristics such as nutrient content, moisture, and pH also affect soil microbial populations. In a similar vein, immature Moso bamboo and broadleaf forest species, a huge woody clonal plant, relies on regular nutrition delivery through clonal integration to develop rapidly. Moreover, the release of BNI substances in the rhizosphere by some forest tree species could also affect N dynamics (Wang et al., 2021). So, an explanatory document about N fixation and inhibition in rhizospheric and bulk soils of Moso bamboo and broadleaf forests is missing in spite of a few short reports in the last several years on how clonal integration affects nutrient transport and allocation in Moso bamboo by Shi et al. (2022). But the N dynamics under different factors and environmental stress conditions remain unclear. In this review, we have summarized the N dynamics in rhizospheres of moso bamboo and broadleaf forests in the presence and absence of BNI substances and environmental factors with the significance of N dynamics in Moso bamboo and broadleaf forests.
Nitrogen is an essential element for plant growth and is a crucial component of forest ecosystems. It is one of the primary nutrients that plants need to synthesize proteins, enzymes, and other essential molecules. It is also a critical limiting factor in many forest ecosystems, meaning that its availability can significantly affect the productivity and health of the ecosystem (Lobell et al., 2004; Kumar et al., 2020; Yan et al., 2020). Nitrogen also plays a vital role in the cycling of nutrients within forest ecosystems. It is taken up by plants from the soil and incorporated into their tissues (Tian et al., 2019). When plants die and decompose, N is released back into the soil, where it can be taken up by other plants or processed by microorganisms. It is also essential for the growth of soil microorganisms, which are critical for nutrient cycling in forest ecosystems (Perakis and Pett-Ridge, 2019). The N in forest trees is found in biological substances (up to 60%) and only a small percentage is found in mineral forms like ammonium (NH4+) and nitrate (NO3–) (Evans and Poorter, 2001; Geisseler et al., 2010). It benefits the trees in many ways, i.e., it boosts the soil water-use efficiency (WUE), and soil serves as an anchor for the expansion and development of tree roots (Shi et al., 2020; Li Y. et al., 2021). When N is up-taken by roots, it regulates and enhances tree growth (Guan et al., 2019; Muhammad et al., 2022). Moreover, N presence in the forest soils accelerates the SOM storage and concentration due to thorough microbial communities manipulation. The N in plant and animal dead bodies stimulates soil microbial activity (Allison et al., 2008). Its concentration also affects residue decomposition and affects nutrient concentration in soils (Drost et al., 2020), and increases phosphorus (P) absorption by the plant by providing acidic compounds in soil and creating an acidic environment which is favorable for nutrient bioavailability to plants (Vitousek and Denslow, 1986). The N in broadleaf and bamboo forests enters via atmospheric deposition, soil minerals, rock weathering, and BNF (Lilleskov et al., 2019). In both types of forests, the cycling of N is largely driven by microorganisms, which play a key role in the decomposition of SOM and the conversion of N between different forms (Mushinski et al., 2021). The N can be immobilized by microorganisms and incorporated into their biomass or can be released back into the soil as inorganic forms (Dong et al., 2019a). However, there are some notable differences between nitrogen dynamics in broadleaf forests and bamboo forests. Moso bamboo is a fast-growing, woody grass species that has unique physiological characteristics, including a high demand for N and a unique root system that can rapidly take up and transport nutrients (Zheng et al., 2021). As a result, bamboo forests typically have higher rates of N uptake and cycling compared to broadleaf forest species.
Additionally, N dynamics in bamboo forests can be influenced by the cultivation practices used for bamboo production. For example, N fertilizers may be applied to promote bamboo growth, which can alter N cycling and have negative impacts on soil and water quality if not managed properly. The detailed importance and fate of N in the broadleaf and bamboo forest tree life cycle and soil biogeochemical cycles are summarized in Figure 1.

Figure 1. Importance of nitrogen in the soil and its role in soil and plant health.
There are over 60,000 tree species in different types of forests worldwide (United Nation’s Environment Program, 2022). So, one of the most significant areas of the study is N dynamics and its enrichment in forest ecosystems owing to its great influence on the forest ecosystem (Reis et al., 2017). Different biotic and abiotic components of the forest ecosystems influence N dynamics, i.e., mycorrhizae. Mycorrhizae works in soils for the decomposition of SOM found in tree litter and dead bodies in the form of different organic complexes, e.g., lignin, and cellulose (Martin et al., 2022; Ward et al., 2022). Along with fungi, bacteria and forest restoration also play crucial roles alone and in combination with fungi in the N cycle as indicated by Wang et al. (2022e). Forests dominated by Oligotrophic Acidobacteria and Actinobacteria shifted to be predominated by Copiotrophic Proteobacteria and Firmicutes after forest restoration (Selmants et al., 2016). Higher C and N stoichiometry in the litter-soil-microbe continuum is mostly because of soil bacterial diversity, whereas increased plant variety is due to tree litter decomposition. But the composition of soil microbes was inversely linked to litter biomass, in addition to N concentrations. The N dynamics and soil microbial community structure along with their functions are also influenced by the tree diversity. Lower plants play a significant role in soil N availability, N mineralization rate, soil bacterial community build-up, and biomass in the top 10 cm of soil compared to deeper horizons. As reported by Xiao et al. (2022), who noted a stronger relationship between soil microbial biomass and N mineralization than there was between soil N content and N mineralization. This indicates that soil N mineralization is affected by forest type and soil microbial biomass. Moreover, soil bacterial composition was also found to have a significant effect on soil microbial biomass, i.e., Actinobacteria, Patescibacteria, and Chloroflexi (Xiao et al., 2022).
Long-term N addition (10 years) has been also reported to be having negative effects on N concentrations in bamboo and other forest ecosystems as indicated by Wang et al. (2022b). In another study, Neves et al. (2022) reported that season and the predominance of various functional groups influence the dynamics of litter and the intake of nutrients in a secondary seasonally dry tropical forest. There may be phenological synchronization across various functional groups since litterfall output was much greater during the two driest months of study. Moreover, trees of the legume family dominated the greatest diameter class, reaching double the basal area of non-legumes. In comparison to non-legume trees, legume trees doubled the quantity of N storage which was dropped throughout the winter. Moreover, altered N addition or mineralization can cause a significant change in soil microbial communities. Soil microbial communities, dissolved organic N, and litterfall all were affected by increased soil water content (Yu et al., 2022). Forest fires also affect the N dynamics in different forest ecosystems during the recovery period and can cause a ninefold increase in soil NH4+-N and a threefold increase in soil NO3–-N concentrations in top soils after 2 years (Xu et al., 2022). Table 1 summarizes the N dynamics in different broadleaf forest and bamboo forest ecosystems around the world.
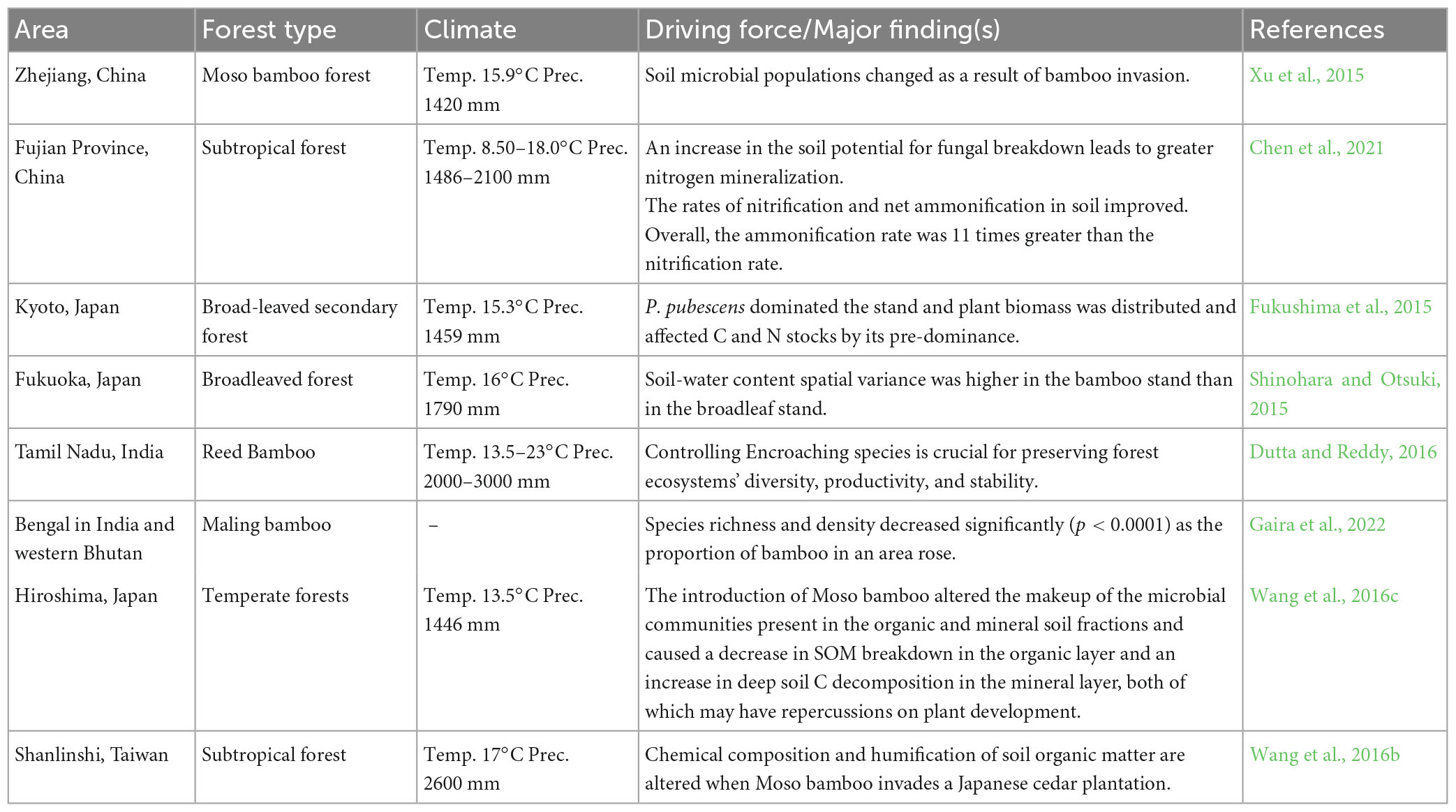
Table 1. Global extent of bamboo invasion into the broadleaf forest ecosystem.
In forests, the soil is continuously enriched with plant litter, dead animal and prey bodies, and plant dead wood (Tang et al., 2018). Therefore, the N cycle is different from agricultural soils along with the factors affecting it as it is majorly regulated by the tree community (Nie et al., 2018; Qin et al., 2019). Tree species determine the nature of N cycling and dynamics (Mushinski et al., 2021). Moreover, soil microbial community and N dynamics change due to several factors like forest fires and N addition from synthetic sources, i.e., microbial community structure, precipitation, soil properties, and litterfall (Yokobe et al., 2018; Tahmasbian et al., 2019; Purwanto and Alam, 2020; Xiao et al., 2020) (Figure 2). Below are the factors which majorly affect the N fixation and cycling in forest ecosystems.
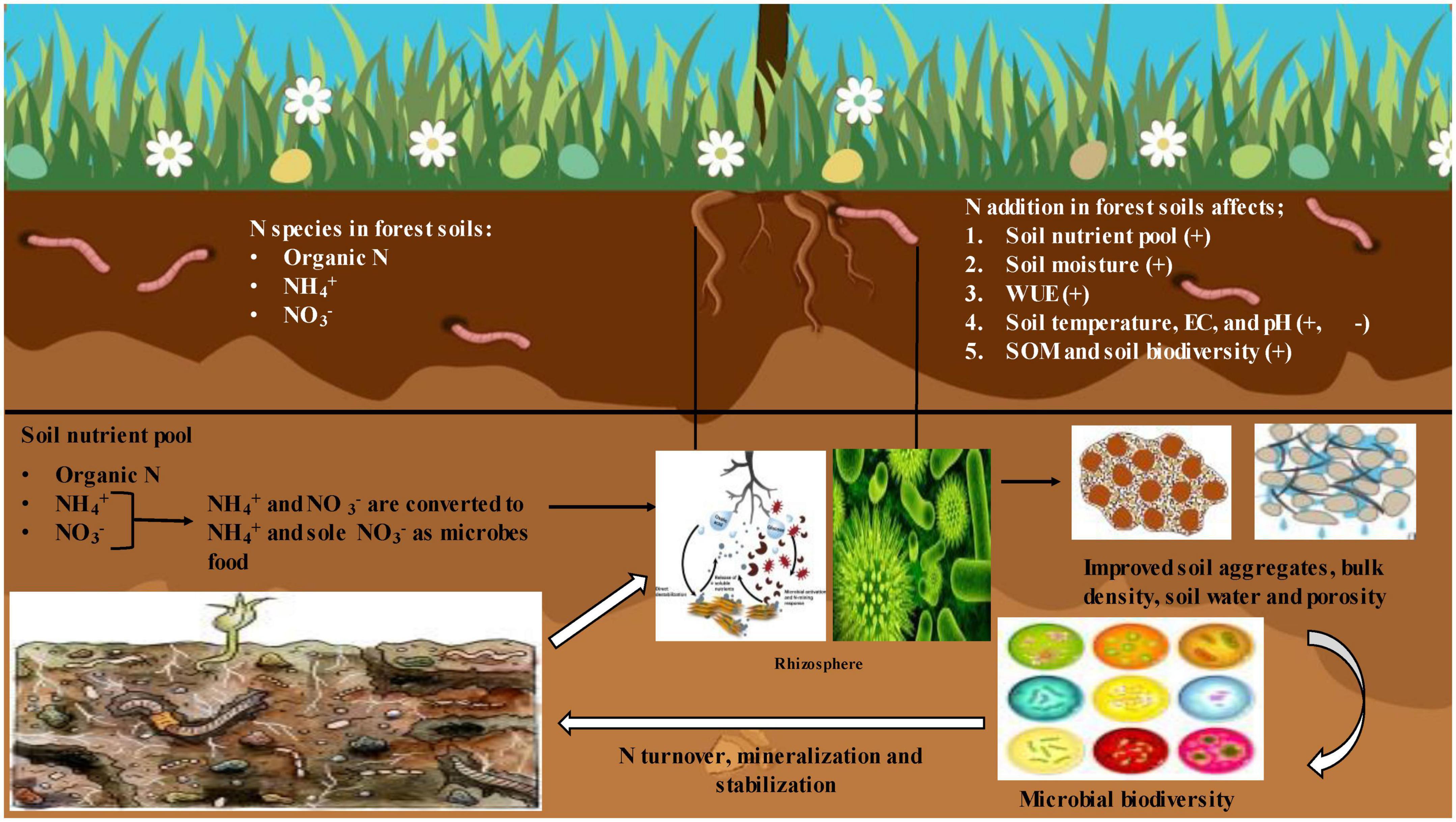
Figure 2. Mechanisms behind nitrogen dynamics laid change in soil properties in forest ecosystem.
As soil under different forest ecosystems inhabitants large amounts of flora and fauna, they also have associated nutrients in the form of SOM and minerals (Dhaliwal et al., 2019; Sun Y. et al., 2021), only a minor alteration in the forest ecosystem can results in a greater response in forest tree species as different factors favor different tree species (Xing et al., 2022). In a study, Isobe et al. (2018b) reported altered N dynamics with the change in tree species as N transformations were driven by the type of tree species and their litterfall, as well as its decomposers’ population diversity. Moreover, some tree species have BNI or BNF characteristics and they greatly influence N dynamics (Ramm et al., 2022). In the same way, different forest ecosystems carry the same responses, i.e., if tree species are replaced with other species, N dynamics changes accordingly. Su et al. (2021) reported the same phenomena in the subtropical forests of China where a change in forest tree ecology resulted in altered nutrient cycles. Table 2 summarizes the N dynamics and their drivers in the numerous forest ecosystem. It can be seen that tree species can influence the N dynamics in a forest ecosystem.
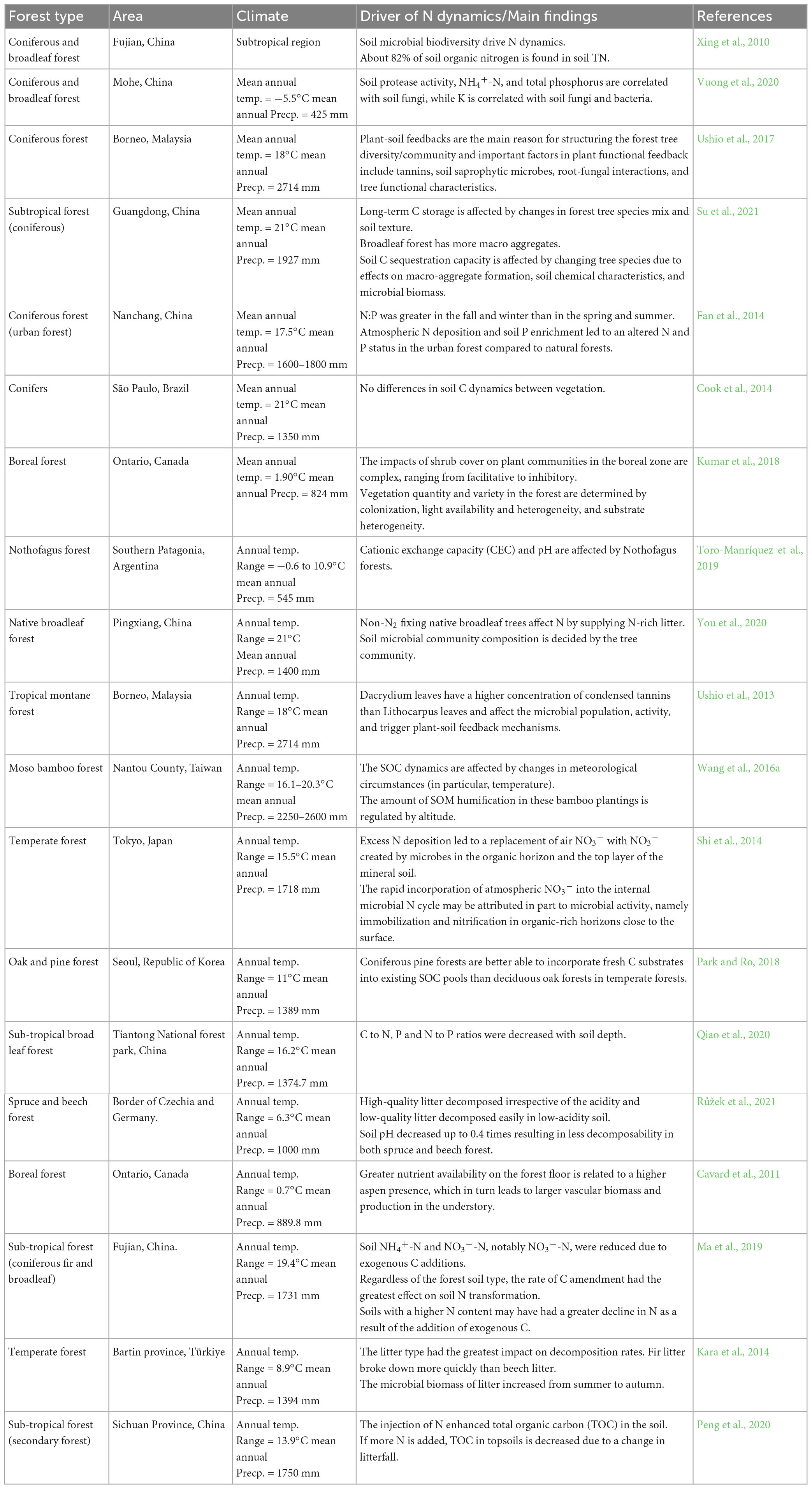
Table 2. Global scenario of forest types and nitrogen dynamics.
Soil microorganisms, forest fires, litterfall, nutrient addition, temperature, and soil characteristics are only a few of the primary elements influencing N dynamics in a forest ecosystem. The availability of N is a significant factor limiting the forest’s primary productivity.
Forests are rich in tree diversity (Schiefer et al., 2020; Babur et al., 2021), and these different kinds of trees have different ecosystem services, e.g., some act as N fixers (Batterman et al., 2018), and some act as a source of N when become dead or both (Thorn et al., 2020), or litter is fallen from them (Lyu et al., 2018; Smith and Wan, 2019). The N-fixing black locust (Robinia pseudoacacia) trees work as N fixation. Black pine (Pinus thunbergii) trees and a mixed stand with black locust trees had higher N-fixing ability when assessed in a study from 2000 to 2019 time period. Cárdenas et al. (2022) reported that using an N-fixing legume and one non-legume tree species in a tropical dry forest results in similar performances across N-fixing and non-fixing legumes in the study area. Similarly oak and non-oak tree species also have different plant litter decomposition rates (Babl-Plauche et al., 2022).
Soil organic matter and humus are produced by the action of microbes. Its mineralization is also governed by soil fauna and flora (Komarov et al., 2017). Wang et al. (2022c) reported more earthworm growth in older forest soil compared to younger forest soil in response to more SOM concentration in older forest soil. The abundance of fungi also has similar effects as indicated by Osono (2015) that fungi pose a prominent role in N dynamics across a variety of forest ecosystems. The bacteria and earthworms in the soil also have reactions to the various functional kinds of plants. Francini et al. (2018) reported that soil microbial and nematode populations vary in natural and urban forests depending upon management, with mostly higher biota in a natural ecosystem, where N is governed by enzymes like arylsulfatase and phosphodiesterase. Earthworms also regulate N dynamics through their diversity and number. Groffman et al. (2015) concluded that N pools in earthworm-colonized plots were more stabilized than in forests dominated by L. rubellus, indicating earthworms help move N from litter into stable SOM and boost the soil microbial biomass. Fugère et al. (2017) reported that N transport in soil was higher with multi-species earthworm communities.
About 360 to 380 million hectares of forest area are lost each year due to forest fires, affecting forest soil biogeochemical cycles (Butler et al., 2020; Rabin et al., 2022), by altering soil pH, electrical conductivity (EC), microbial populations, and associated biochemical processes. Forest fires cause the melting of permafrost and remove the insulating layer of SOM, and raise soil temperatures (Aaltonen et al., 2019). The decomposition of plant organic matter is an essential part of the biogeochemical cycle on land and has a significant impact on both the fire fuel load and the C balance of forests. Butler et al. (2020) tested whether three alternative long-term fire regimes (no burning, 4-yearly burning, and 2-yearly burning) were linked to altered N dynamics during litter decomposition by conducting a 277-day experiment in an Australian eucalypt forest. The greatest rates of decomposition and C loss were seen in the no-burning treatment. Hu et al. (2020) concluded after a study in mixed coniferous (Pinus massoniana Lamb.) and broadleaf (Quercus acutissima Carruth.) forests in China that after 3 months of forest fire, the soil pH, NO3–-N concentration, and microbial biomass carbon (MBC) were increased. Four weeks following a fire, the total aboveground biomass in the burnt plots was 39% less than in the unburned plots. In another study, James et al. (2018) reported that over 45 years after a forest fire, changes were still visible in the organic horizon and mineral soil characteristics. Albert-Belda et al. (2022) also reported similar results that burned soils had less microbial biomass compared to unburned forest sites, and burned soils were dominated by gram-positive bacteria.
Nitrogen addition plays an important role in soil biogeochemical cycles in forest ecosystems depending upon the forest tree species, amount of N added, and environmental conditions. Chen et al. (2020) reported that N addition in two different forests with different tree species did not affect SOM and soil organic carbon (SOC). Chen et al. (2016) and Lu et al. (2011) reported similar results that inorganic N, nitrous oxide (N2O) emissions, and NO3– leaching all increased noticeably as a consequence of excessive N addition during 6 years. In another study, the decomposition constant increased by 46% as a result of the N addition. N addition reduced the strong association between litter and decomposers that accelerated the breakdown of home litter and caused stimulated N release from decomposing litter, with plant species effects moderating this process (Cui et al., 2017).
Litterfall is one of the most important factors in forest ecosystems which supply C and N after their decomposition through soil microbes. Bohara et al. (2020) reported that the release of P, potassium (K), C, and N rates followed the order of P > K > C > N from forest trees, suggesting that litter is a major sink of N. In another study, Kamruzzaman et al. (2019) reported C and N content of litterfall decomposition up to 1005.9 g m–2 year–1.
Soil properties were altered with altered N in terms of pH, EC, N, SOM, and P (Lucas-Borja et al., 2022). Siwach et al. (2021) reported that mosses alter soil properties by altering the soil’s capacity to hold nutrients. Xiong et al. (2022) reported that N availability and fluctuations in the microbial community cause significant seasonal changes in the composition of the soil microbiome over an elevation gradient. Molla et al. (2022) concluded that natural forests had the highest concentrations of clay, pH, SOM, N, P, exchangeable bases (Ca+, Mg+, K+, Na+), and cation exchange capacity (CEC). The soil pH, clay, bulk density (BD), and exchangeable bases all improved/increased with depth across all land use change categories in forests.
Mutualistic relationships between tree roots and the soil are how trees get access to the water, nutrients, and minerals in the soil (Authier et al., 2022). Forest ecosystems rely heavily on fine roots for N cycling. Dead roots have more N compared to other tree parts (Hu et al., 2022). Fine roots play a crucial role in the N dynamics of forests, including Moso bamboo forests by uptake absorbing more N from the soil, which is then used for plant growth and metabolism (Drake et al., 2011; Bai et al., 2016). Some tree species have symbiotic relationships with N-fixing bacteria in their fine roots, which converts atmospheric N into a form that plants can used (Wang H. et al., 2019; Staccone et al., 2020). Moreover, decaying fine roots contribute to the forest floor litter and provide N to the soil and can modify soil microbial activity, which in turn can affect N availability and transformations in the soil (Leppälammi-Kujansuu et al., 2014; Solly et al., 2014).
Furthermore, it is well-known fact that roots also play a significant role in the nutrient cycles in the ecosystem through interactions between living and dead roots, the amount and quality of the soil structure and nutrients around the roots, and the impact of the roots themselves may vary depending on the soil profile and the quality of the surrounding soil. Ehrenfeld et al. (1997) examined the various impacts of living and dead roots in forest soils and reported that live roots only promoted ammonification rates in mineral soil when an organic surface horizon was present, while dead roots increased the amount of extractable N, but not in the organic matter. It suggests that N dynamics in forest soils are strongly influenced by the presence or absence of an organic horizon in addition to the root activity.
Humidity affects N dynamics in forests by influencing plant transpiration and microbial activity, both of which play a role in N cycling through the ecosystem (Dong et al., 2019b). Increased humidity can lead to increased transpiration and microbial activity, leading to a faster rate of N uptake and processing by plants and soil microbes (Luce et al., 2011). However, prolonged higher humidity can also result in decreased plant growth and N uptake due to water stress (Deng et al., 2021). Thus, the effect of humidity on N dynamics in forests is complex and dependent on the specific environmental conditions and time frame considered. It is reported that variation in humidity alters the forest’s nutrient and biogeochemical cycles by altering soil temperature. This rise in temperature triggers soil N mineralization rates up to 55% in comparison to the natural forest, with the increase being greatest in the grassland and lowest in the home garden. The concentration of SOC, MBC, and NO3– also decreases in these types of soils in the forests compared to the natural forest (Pandey et al., 2010).
Soil texture also has significant effects on N dynamics in forests. It affects water and nutrient uptake, root growth and distribution, and microbial activity which in turn influences N uptake and cycling (Ma et al., 2016). Fine-textured soils (e.g., clay) have higher water and nutrient-holding capacity but also have lower oxygen levels, which can limit microbial activity and plant root growth (Chivenge et al., 2011). Coarse-textured soils (e.g., sand) have better aeration but lower water and nutrient retention, leading to less efficient N cycling. Soil texture also affects the availability of different forms of N (e.g., NO3–, NH4+) to plants and microbes, which can influence the rate and efficiency of N cycling as reported by Chivenge et al. (2011) that more N and C were stored in soil aggregates in high clay soils.
Due to higher plant inputs and soil flora and fauna in the forest ecosystem, nutrient cycles, and dynamics change during forest growth and regrowth of the forests and largely depend upon the adequate N supply. In younger forests, mineralization and nitrification are decoupled, i.e., more mineralization and low nitrification, suggesting a closed N cycle that simultaneously preserves N availability for forest renewal (Figueiredo et al., 2019). Moreover, the dynamics of litter decomposition, N cycling, SOM, and plant-soil interactions may all be altered when trees associate with various mycorrhizas (Eagar et al., 2022).
Sarai et al. (2022) reported that decomposed litter could alter the soil N transformation which may be further impacted by the changes in the structure and function of the soil microbial community. However, this impact is not for all forest types as N-limited boreal forests due to the substantial contribution of understory plants to soil N cycling (Xiao et al., 2022). In addition, soil and plant organic matter breakdowns are crucial to the biogeochemical cycles even in forest settings where understory solar irradiation stays relatively low. Wang et al. (2022d) reported that the litter decomposition in the forest was 64% which increased up to 75% in adjacent areas with less or no tree cover.
The storage of C in forest soils is an important factor in reducing the effects of climate change and global warming through C sequestration. The N enrichment considerably improves C dynamics in forests by an average of 24 and 10%, respectively (Schulte-Uebbing and de Vries, 2018). In subtropical and tropical forests, both the C pool and inputs increased by 10% in response to N addition, the effect was more apparent in boreal forests (+ 17%) than in subtropical forests (−19%). Soil C sequestration was positively affected by N enrichment in boreal (6%) and temperate (7%), but negatively impacted in subtropical (−30%) and tropical (−10%) forests (Ngaba et al., 2022). Similarly, arbuscular and/or ectomycorrhizal trees can modify soil C and N dynamics. Fungi can easily decompose SOM and also form complexes with it. So, the build-up and decomposition of organic materials can occur (Ward et al., 2022). The essential rhizosphere processes of fine root formation, turnover, and breakdown also help in sequestering C in forest soils during succession. Sarai et al. (2022) reported that the fine root turnover contributes more to C turnover than the standing fine root biomass.
The N and C fertilization decreases SOC and N mineralization which is linked to microbial growth and enzyme activity as fertilization was associated with bacterial growth in the high-C larch forest, concluding that N fertilization contributes to soil C sequestration (Na et al., 2022). Soil C sequestration is significantly impacted by plant roots and the mycorrhizal fungi that live in and around them but contributes to SOC build-up. In an ectomycorrhizae-dominated forest with high N addition (25 kg N ha–1 year–1), SOC increased from 18.02 to 20.55 mg C g–1 (Zhu et al., 2022).
As an indicator of healthy soil, SOC levels are often used for this purpose. To better understand soil functioning as a major element for C sequestration along agricultural production and food and water security. Substantial SOC accumulation in forest soils was agreed upon, providing not just a C sink but also its supply, despite limited data availability and large observed fluctuation in SOC content (Alavi-Murillo et al., 2022). The high rates of SOM/SOC formation play a major role in ecosystem stability (Siwach et al., 2021).
After latitude and altitude, the physicochemical and biological properties of soil may also have a role in forest and tree species distribution. Alternatively, soil formation and associated edaphic qualities may also be affected by variations in tree species composition. The pace at which chemical weathering of parental material occurs, which affects pH and the composition of complex organic acids in soil solutions, might vary with tree species. However, trees mostly influence soil minerals weathering in the top few inches of soil or close to the roots (Joshi and Garkoti, 2021). Some tree-soil interactions, including the interception of airborne contaminants or the podzolization of soil, may have evolved with just a weak or no correlation to plant fitness or closeness. In addition, local factors like litter input, stand species composition, and microclimate variables can have an impact on the chemical properties of forest soils, while long-term effects (like permanent vegetation over a long period) or drastic alterations (like fires, intensive harvesting) primarily influence soil structure and its physical properties (Toro-Manríquez et al., 2019; Joshi and Garkoti, 2021).
One of the most important aspects of the biological health of the soil is bacterial community (Ao et al., 2022). Changes in the microbial biomass of soil are a good indication of the ongoing transformations in the soil nutrients and properties. Knowing how the microbial biomass in plantation soil fluctuates over time is crucial for effective plantation management and cultivation.
It is reported that nutrients serve as useful markers of how forest management methods affect the dynamics of SOC. The largest C:N levels were seen in the autumn and the smallest values were observed in the winter for the three different forest types, although these seasonal patterns were not consistent (Babur et al., 2021). Sun Y. et al. (2021) reported that the rhizosphere’s microbial biomass nitrogen (MBN) was more responsive to mulching and other soil environmental changes. Soil P was shown to correlate with TN and dissolved N, although other N fractions were significantly influenced by soil physical qualities (temperature, water content, bulk density) (Yin et al., 2021). The N content of both fine roots and organic mulch was reduced by mulching, although the N content of the soil was more closely connected to the N content of the leaves (except MBN) (Wang H. et al., 2019; Table 3).
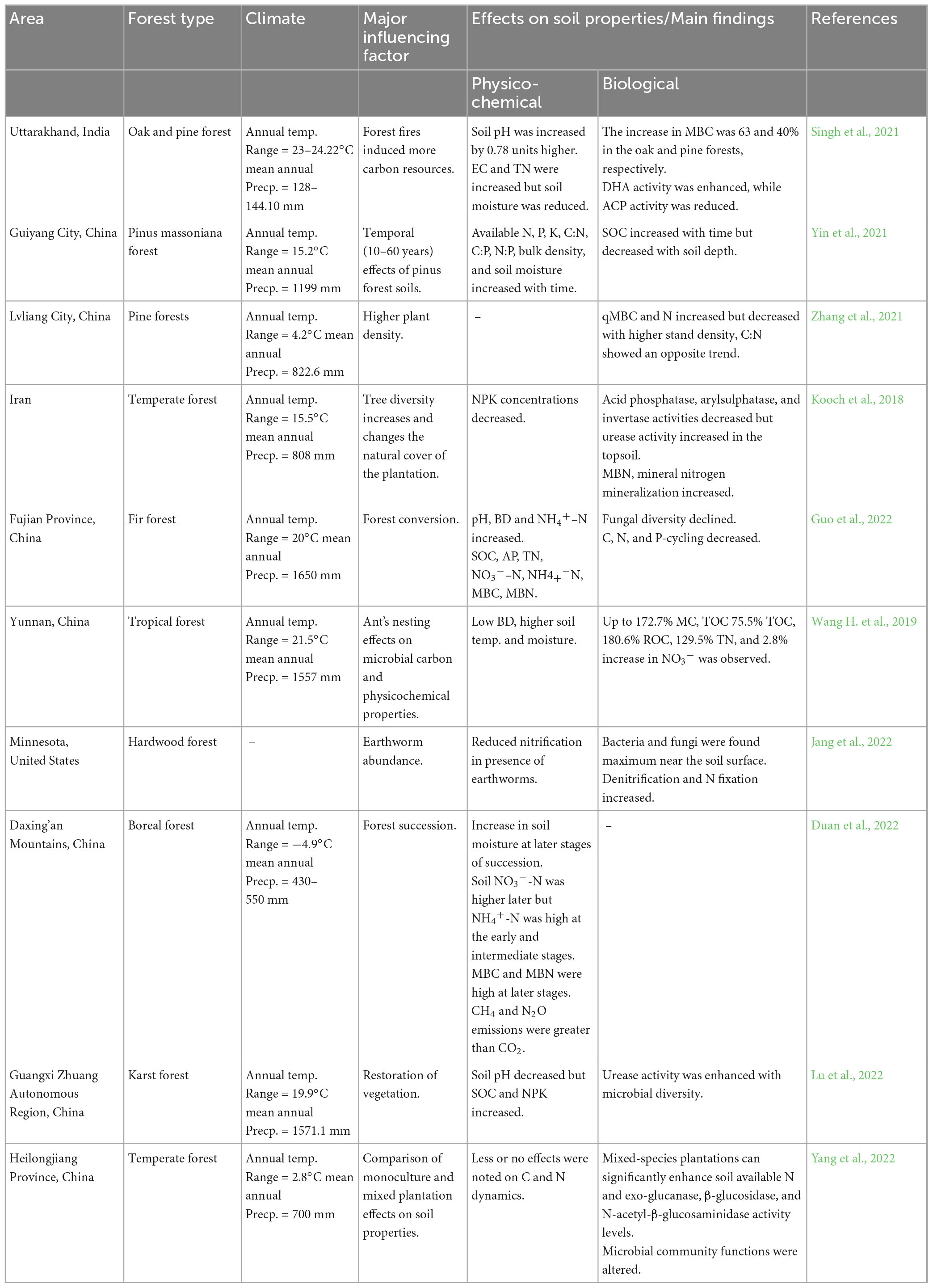
Table 3. Influence of N dynamics on soil properties of different forest ecosystems.
Soil quality and nutrient storage in forests are interlinked. Although there are minute differences between the broadleaf and bamboo forests according to their respective ecosystem functioning, there are some significant differences that exist and are highlighted in Figure 3. Feng et al. (2020) reported that litter decomposition has a profound effect on N dynamics, and this effect varies greatly with the variety of tree species present in a given forest. Wang et al. (2018) studied the effects of litter mixing and altering the stand environment on the litter decomposition rate and the composition-specific litter organic C and N loss rate of a mixed-species plantation. Mixed-species plantations with lower-quality litter reduced decomposition and resisted N and C loss, whereas higher-quality litter speeded up litter decomposition, which was caused by the higher SOM and SON availability in the mixed-species plantation.
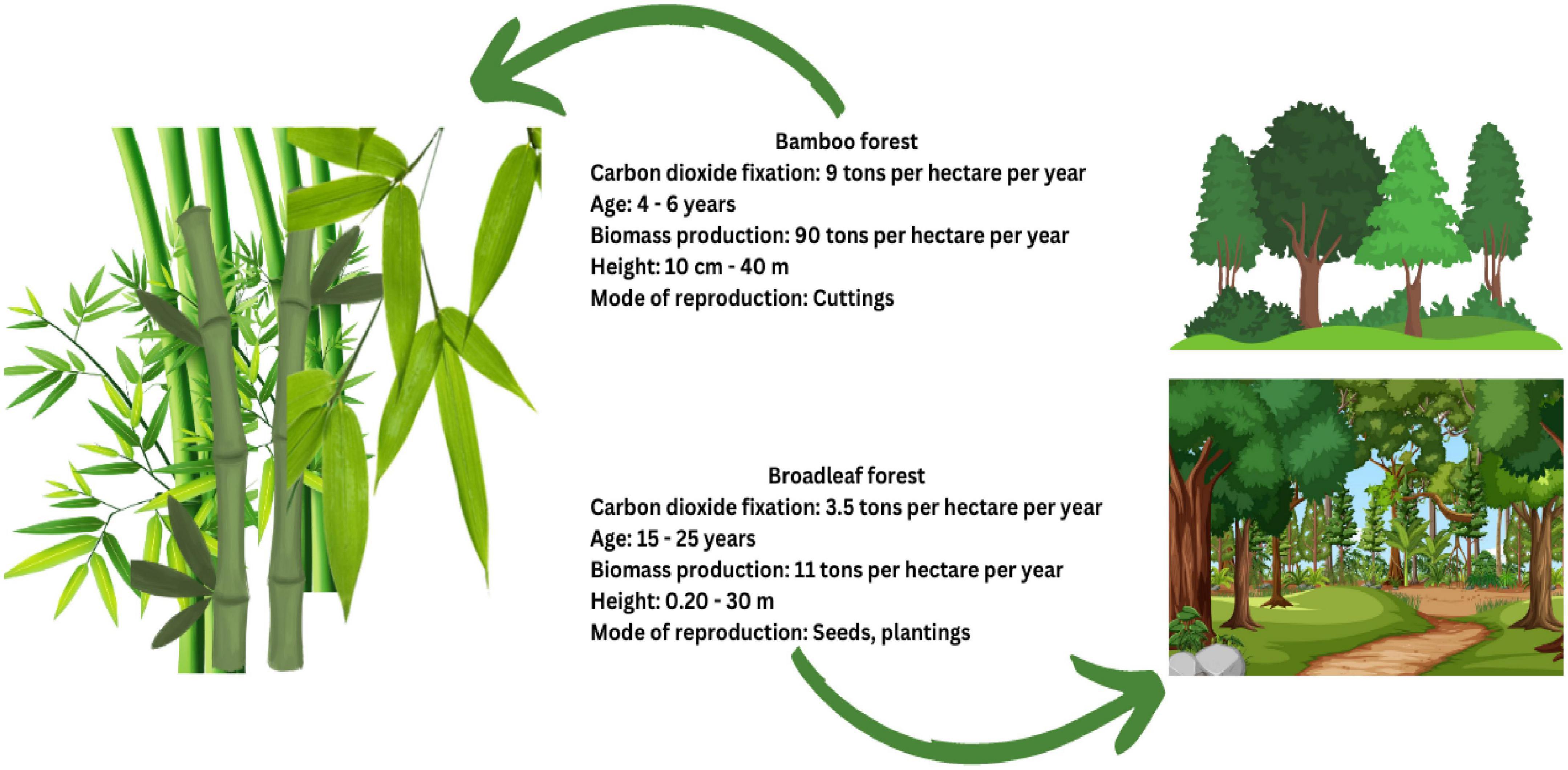
Figure 3. Differences in the bamboo and broadleaf forest species.
It has been reported that tree C, N, and P stocks all fell when moso bamboo and broadleaf forests are converted to other types of forests, with declines of 43.8, 47.9, and 63.1%, respectively, while C and N stocks throughout the whole soil profile fell by 19.1 and 13.0%, respectively as conversion resulted in a decrease in soil P stock from 0 to 20 cm. It indicates that tree biomass can drive the N dynamics and retention of nutrients. In forest soils, the metabolism of C and N is also influenced by the microbial communities and enzyme activities present there. Soil pH, MBC, MBN, C, N, P, NH4+-N, and NO3–N levels were all shown to be strongly impacted by forest type (Ji et al., 2021). Moreover, trees with arbuscular mycorrhizae vary from ecto- and endomycorrhizal tree species in their litter quality and N cycle patterns, which may influence mycorrhizal colonization and the community’s composition and variety (Zhang et al., 2022). Figure 4 explains the factors and processes taking place in broadleaf and bamboo forest ecosystems during N cycling/dynamics. Soil P, calcium (Ca+), K, and magnesium (Mg+), as well as elemental stoichiometric ratios also respond differently in broadleaf and bamboo forests (Feng et al., 2017).
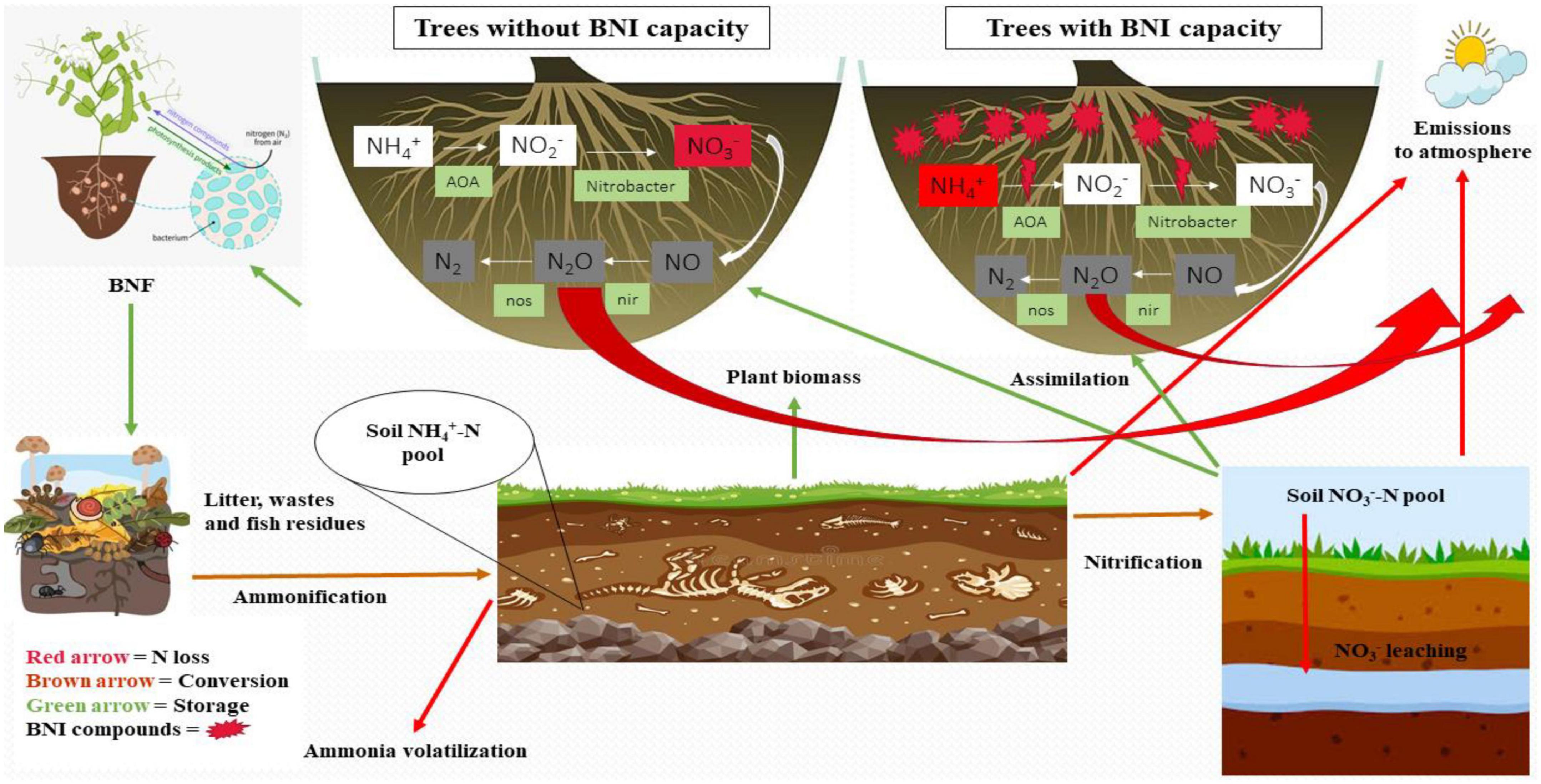
Figure 4. Nitrogen dynamics under the direct influence of trees with and without BNI capacity.
The ecology, as well as the N dynamics in forest soil, is distinct from that of the other soil ecosystems. Nutrient and biogeochemical cycles are necessary for the proper functioning of the forest ecosystem like moso bamboo forests. Moreover, for proper functioning nutrient cycles must work and govern properly. These cycles are run by input factors, i.e., natural, and human processes that affect litter formation and decomposition. The growing influences of different natural and human variables on the nutrient and biogeochemical cycle of tropical forest ecosystems highlight the significance of studying C, N dynamics, litter formation, and its breakdown processes. As the soil microbiome plays a vital role in the feedback between the nutrient cycle and the climate, models used to anticipate changes in ecosystem functioning under changing climatic scenarios must account for this variation among plant species (Xu et al., 2018). As a result of the increase in plant resources and the subsequent rise in soil enzyme activities, soil C, and accessible N, P, and K levels are all enhanced.
Moso bamboo and broadleaf forests have a high SOC storage capacity but also great geographic variation. The SOC in both forest types was shown to be significantly linked with BD (R2 = 0.373), pH (R2 = 0.429), GC (R2 = 0.163), and CEC (R2 = 0.263) (Zhang et al., 2015). Another study revealed that SOC, its fractions, C mineralization rate, soil pH, and total N in 0–20 and 20–40 cm soil layers rose dramatically with increasing duration of intensive management, along with improvements in aggregate stability (Yang et al., 2021). A study in bamboo plantations revealed that SOC, labile soil organic C pools, TN, water-soluble organic N, NH4+-N, NO3–-N, total P, organic and inorganic P content in the rhizosphere are all significantly increased (Ni et al., 2021). In another study, soil samples from the moso bamboo forest were collected at depths of 0, 20, and 60 cm at 138 different spots around the research region. The N, P, and K concentrations varied from 1.01 to 4.11 g kg –1, 0.025 to 0.131 g kg–1, and 0.42 to 5.40 g kg–1, respectively (Guan et al., 2017; Xiao et al., 2021).
The effects of N deposition and management practices on leaf litterfall and N and P return in the moso bamboo (P. edulis) forest are not yet known. However, Zhang et al. (2017) studied the variations between no N, low N (N30), medium N (N60), and high N (N90) levels of simulated N deposition, as well as the control with no N. It was concluded that between 78% and 82% of annual leaf litterfall, 86–94% of annual N return, and 85.6–96% of annual P return occurred during the study period.
Same as the other forests ecosystem, N addition in the bamboo forests also results in more stable N and C concentrations and efficient biogeochemical cycling. Li Y. et al. (2021) studied the changes in soil microbial abundance, variety, and community structure in response to N addition (0, 30, 60, or 90 kg N ha–1 year–1) in a moso bamboo (P. edulis) forest in subtropical China. Results showed an increase in microbial biodiversity (Lv et al., 2022). A 7-year field experiment was conducted in a subtropical moso bamboo (P. edulis) plantation in Zhejiang Province, China to examine the effects of varying N addition rates (0, 30, 60, and 90 kg N ha–1 year–1) on soil respiration rates with stumps of three ages (1, 3, and 5 years after cutting). Results indicated that N addition reduced soil respiration by 4.69 to 27.37% compared to the control treatment (Cai et al., 2021; Li et al., 2022).
The C and N cycle of the forest ecosystem, i.e., moso bamboo forests could be greatly impacted by increases in environmental stresses, such as drought frequency and severity. In a simulated study involving drought as a stress factor, three different litterfall treatments were tested for responses about soil respiration, temperature sensitivity, and the lagging effect of the soil. The treatments were ambient litterfall, litter addition, and litter removal. It was also noted that the annual soil respiration rate was lower in the unchanged treatment (2.34 mmol m–2s–1) compared to the control (3.15 mmol m–2s–1) (Ge et al., 2018). Soil respiration’s temperature sensitivity was lowered by 8.4% due to drought, 15.4% due to litter addition, and 7.6% due to litter removal (Ge et al., 2018). It was also reported that drought reduces litter decomposition rates by up to 30%, thereby affecting N dynamics (Jin et al., 2020; Tong et al., 2020; Ge et al., 2022).
Forest ecosystems play a crucial role in global C and N cycles, as well as temperature and precipitation patterns. Hence, they are the focus of study by scientists from all over the world. Because of the worldwide goals of carbon neutrality and carbon sequestration. They constitute an essential component of the global carbon sink, i.e., moso bamboo forests. Moso bamboo and broadleaf forests are crucial elements of the carbon and N cycles since they are so prevalent in the subtropics. Because it is essential to have clear quantification of changes in bamboo forest soil to understand the N dynamics of bamboo forests, soil C sequestration, N dynamics, and the biomass of the entire broadleaf and moso bamboo forest ecosystem. Moreover, C and N dynamics are greatly influenced by biotic and abiotic factors. Even small changes in these factors can cause severe negative impacts on soil biogeochemical cycles and disturbance in soil ecology, ultimately destroying balance in an ecosystem. More and improved research on soil microbes and microbial residues is needed to understand the mechanisms of soil microbial responses to different settings and the carbon and nitrogen cycle and sequestration process for C and N accumulation by microbial residues in broadleaf and moso bamboo forests.
MS: conceptualization and original draft writing. YL: funding acquisition. FY, ZF, and YL: visualization and revision. MS, FY, and YL: original draft review and funding acquisition. All authors have read and agreed to the published version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aaltonen, H., Köster, K., Köster, E., Berninger, F., Zhou, X., Karhu, K., et al. (2019). Forest fires in Canadian permafrost region: The combined effects of fire and permafrost dynamics on soil organic matter quality. Biogeochemistry 143, 257–274. doi: 10.1007/s10533-019-00560-x
Alavi-Murillo, G., Diels, J., Gilles, J., and Willems, P. (2022). Soil organic carbon in Andean high-mountain ecosystems: Importance, challenges, and opportunities for carbon sequestration. Reg. Environ. Change 22:128. doi: 10.1007/s10113-022-01980-6
Albert-Belda, E., Hinojosa, M. B., Laudicina, V. A., García-Ruiz, R., Pérez, B., and Moreno, J. M. (2022). Previous fire occurrence, but not fire recurrence, modulates the effect of charcoal and ash on soil C and N dynamics in Pinus pinaster Aiton forests. Sci. Total Environ. 802:149924. doi: 10.1016/j.scitotenv.2021.149924
Allison, S. D., Czimczik, C. I., and Treseder, K. K. (2008). Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest. Glob. Change Biol. 14, 1156–1168. doi: 10.1111/j.1365-2486.2008.01549.x
Ao, L., Zhao, M., Li, X., and Sun, G. (2022). Different urban forest tree species affect the assembly of the soil bacterial and fungal community. Microb. Ecol. 83, 447–458. doi: 10.1007/s00248-021-01754-3
Authier, L., Violle, C., and Richard, F. (2022). Ectomycorrhizal networks in the anthropocene: From natural ecosystems to urban planning. Front. Plant Sci. 13:900231. doi: 10.3389/fpls.2022.900231
Babl-Plauche, E. K., Alexander, H. D., Siegert, C. M., Willis, J. L., and Berry, A. I. (2022). Mesophication of upland oak forests: Implications of species-specific differences in leaf litter decomposition rates and fuelbed composition. For. Ecol. Manag. 512:120141. doi: 10.1016/j.foreco.2022.120141
Babur, E., Dindaroğlu, T., Solaiman, Z. M., and Battaglia, M. L. (2021). Microbial respiration, microbial biomass and activity are highly sensitive to forest tree species and seasonal patterns in the eastern mediterranean karst ecosystems. Sci. Total Environ. 775:145868. doi: 10.1016/j.scitotenv.2021.145868
Bai, S., Conant, R. T., Zhou, G., Wang, Y., Wang, N., Li, Y., et al. (2016). Effects of moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pools. Sci. Rep. 6:31480. doi: 10.1038/srep31480
Batterman, S. A., Hall, J. S., Turner, B. L., Hedin, L. O., Lahaela Walter, J. K., Sheldon, P., et al. (2018). Phosphatase activity and nitrogen fixation reflect species differences, not nutrient trading or nutrient balance, across tropical rainforest trees. Ecol. Lett. 21, 1486–1495. doi: 10.1111/ele.13129
Bohara, M., Acharya, K., Perveen, S., Manevski, K., Hu, C., Yadav, R. K. P., et al. (2020). In situ litter decomposition and nutrient release from forest trees along an elevation gradient in central Himalaya. Catena 194:104698. doi: 10.1016/j.catena.2020.104698
Butler, O. M., Lewis, T., Rezaei Rashti, M., and Chen, C. (2020). Long-term fire regime modifies carbon and nutrient dynamics in decomposing Eucalyptus pilularis leaf litter. Front. For. Glob. Change 3:22. doi: 10.3389/ffgc.2020.00022
Cai, C., Yang, Z., Liu, L., Lai, Y., Lei, J., Fan, S., et al. (2021). Consistent effects of canopy vs. Understory nitrogen addition on soil respiration and net ecosystem production in moso bamboo forests. Forests 12:1427. doi: 10.3390/f12101427
Cárdenas, C. D., Varón-García, D., Suárez-Rodríguez, F., and Pizano, C. (2022). Overcoming the regeneration barriers of tropical dry forest: Effects of water stress and herbivory on seedling performance and allocation of key tree species for restoration. J. Trop. Ecol. 38, 210–218. doi: 10.1017/S0266467422000074
Cavard, X., Bergeron, Y., Chen, H. Y. H., and Paré, D. (2011). Effect of forest canopy composition on soil nutrients and dynamics of the understorey: Mixed canopies serve neither vascular nor bryophyte strata. J. Veg. Sci. 22, 1105–1119. doi: 10.1111/j.1654-1103.2011.01311.x
Chen, H., Gurmesa, G. A., Zhang, W., Zhu, X., Zheng, M., Mao, Q., et al. (2016). Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing. Funct. Ecol. 30, 305–313. doi: 10.1111/1365-2435.12475
Chen, J., Ji, C., Fang, J., He, H., and Zhu, B. (2020). Dynamics of microbial residues control the responses of mineral-associated soil organic carbon to N addition in two temperate forests. Sci. Total Environ. 748:141318. doi: 10.1016/j.scitotenv.2020.141318
Chen, Z., Li, Y., Chang, S. X., Xu, Q., Li, Y., Ma, Z., et al. (2021). Linking enhanced soil nitrogen mineralization to increased fungal decomposition capacity with Moso bamboo invasion of broadleaf forests. Sci. Total Environ. 771:144779. doi: 10.1016/j.scitotenv.2020.144779
Cheng, Y., Wang, J., Chang, S. X., Cai, Z., Müller, C., and Zhang, J. (2019). Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review. Environ. Pollut. 244, 608–616. doi: 10.1016/j.envpol.2018.10.054
Chivenge, P., Vanlauwe, B., Gentile, R., and Six, J. (2011). Comparison of organic versus mineral resource effects on short-term aggregate carbon and nitrogen dynamics in a sandy soil versus a fine textured soil. Agric. Ecosyst. Environ. 140, 361–371. doi: 10.1016/j.agee.2010.12.004
Cook, R. L., Binkley, D., Mendes, J. C. T., and Stape, J. L. (2014). Soil carbon stocks and forest biomass following conversion of pasture to broadleaf and conifer plantations in Southeastern Brazil. For. Ecol. Manag. 324, 37–45. doi: 10.1016/j.foreco.2014.03.019
Cui, X., Yue, P., Gong, Y., Li, K., Tan, D., Goulding, K., et al. (2017). Impacts of water and nitrogen addition on nitrogen recovery in Haloxylon ammodendron dominated desert ecosystems. Sci. Total Environ. 601-602, 1280–1288. doi: 10.1016/j.scitotenv.2017.05.202
Deng, L., Peng, C., Kim, D.-G., Li, J., Liu, Y., Hai, X., et al. (2021). Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems. Earth Sci. Rev. 214:103501. doi: 10.1016/j.earscirev.2020.103501
Dhaliwal, S., Naresh, R., Mandal, A., Singh, R., and Dhaliwal, M. (2019). Dynamics and transformations of micronutrients in agricultural soils as influenced by organic matter build-up: A review. Environ. Sustain. Indic. 1:100007. doi: 10.1016/j.indic.2019.100007
Dong, Z., Driscoll, C. T., Campbell, J. L., Pourmokhtarian, A., Stoner, A. M., and Hayhoe, K. (2019a). Projections of water, carbon, and nitrogen dynamics under future climate change in an alpine tundra ecosystem in the southern Rocky Mountains using a biogeochemical model. Sci. Total Environ. 650, 1451–1464. doi: 10.1016/j.scitotenv.2018.09.151
Dong, Z., Driscoll, C. T., Johnson, S. L., Campbell, J. L., Pourmokhtarian, A., Stoner, A. M., et al. (2019b). Projections of water, carbon, and nitrogen dynamics under future climate change in an old-growth Douglas-fir forest in the western Cascade Range using a biogeochemical model. Sci. Total Environ. 656, 608–624. doi: 10.1016/j.scitotenv.2018.11.377
Drake, J. E., Gallet-Budynek, A., Hofmockel, K. S., Bernhardt, E. S., Billings, S. A., Jackson, R. B., et al. (2011). Increases in the flux of carbon belowground stimulate nitrogen uptake and sustain the long-term enhancement of forest productivity under elevated CO2. Ecol. Lett. 14, 349–357. doi: 10.1111/j.1461-0248.2011.01593.x
Drost, S. M., Rutgers, M., Wouterse, M., De Boer, W., and Bodelier, P. L. (2020). Decomposition of mixtures of cover crop residues increases microbial functional diversity. Geoderma 361:114060. doi: 10.1016/j.geoderma.2019.114060
Duan, B., Cai, T., Man, X., Xiao, R., Gao, M., Ge, Z., et al. (2022). Different variations in soil CO2, CH4, and N2O fluxes and their responses to edaphic factors along a boreal secondary forest successional trajectory. Sci. Total Environ. 838:155983. doi: 10.1016/j.scitotenv.2022.155983
Dutta, K., and Reddy, C. S. (2016). Geospatial analysis of reed bamboo (Ochlandra travancorica) invasion in Western Ghats, India. J. Indian Soc. Remote Sens. 44, 699–711. doi: 10.1007/s12524-015-0534-5
Eagar, A. C., Mushinski, R. M., Horning, A. L., Smemo, K. A., Phillips, R. P., and Blackwood, C. B. (2022). Arbuscular mycorrhizal tree communities have greater soil fungal diversity and relative abundances of saprotrophs and pathogens than ectomycorrhizal tree communities. Appl. Environ. Microbiol. 88:e0178221. doi: 10.1128/AEM.01782-21
Ehrenfeld, J. G., Parsons, W. F. J., Han, X., Parmelee, R. W., and Zhu, W. (1997). Live and dead roots in forest soil horizons: Contrasting effects on nitrogen dynamics. Ecology 78, 348–362. doi: 10.2307/2266012
Evans, J. R., and Poorter, H. (2001). Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 24, 755–767. doi: 10.1046/j.1365-3040.2001.00724.x
Fan, J., Wang, J.-Y., Hu, X.-F., and Chen, F.-S. (2014). Seasonal dynamics of soil nitrogen availability and phosphorus fractions under urban forest remnants of different vegetation communities in Southern China. Urban For. Urban Green. 13, 576–585. doi: 10.1016/j.ufug.2014.03.002
Feng, C., Ma, Y., Fu, S., and Chen, H. Y. H. (2017). Soil carbon and nutrient dynamics following cessation of anthropogenic disturbances in degraded subtropical forests. Land Degrad. Dev. 28, 2457–2467. doi: 10.1002/ldr.2781
Feng, Y., Han, S., Wei, Y., Xiao, J., Geng, S., Gu, Y., et al. (2020). Comparative study of lignin stabilizing mechanisms in soil aggregates at virgin mixed broadleaf-pine forest and secondary broadleaf forest at Changbai mountain nature reserve, Northeast China. Ecol. Indic. 117:106665. doi: 10.1016/j.ecolind.2020.106665
Figueiredo, V., Enrich-Prast, A., and Rütting, T. (2019). Evolution of nitrogen cycling in regrowing Amazonian rainforest. Sci. Rep. 9:8538. doi: 10.1038/s41598-019-43963-4
Follett, R. F., and Hatfield, J. L. (2001). Nitrogen in the environment: Sources, problems, and management. ScientificWorldJournal 1, (Suppl. 2), 920–926. doi: 10.1100/tsw.2001.269
Francini, G., Hui, N., Jumpponen, A., Kotze, D. J., Romantschuk, M., Allen, J. A., et al. (2018). Soil biota in boreal urban greenspace: Responses to plant type and age. Soil Biol. Biochem. 118, 145–155. doi: 10.1016/j.soilbio.2017.11.019
Fugère, M., Bradley, R. L., and Vellend, M. (2017). Exotic earthworms dispersion through protected forest areas and their potential impacts on nitrous oxide production. Biol. Invasions 19, 773–783. doi: 10.1007/s10530-016-1331-y
Fukushima, K., Usui, N., Ogawa, R., and Tokuchi, N. (2015). Impacts of moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in Kyoto, western Japan. Plant Species Biol. 30, 81–95. doi: 10.1111/1442-1984.12066
Gaira, K. S., Pandey, A., Sinha, S., Badola, H. K., Lepcha, J., Dhyani, P. P., et al. (2022). Maling bamboo (Yushania maling) overdominance alters forest structure and composition in Khangchendzonga landscape, Eastern Himalaya. Sci. Rep. 12:4468. doi: 10.1038/s41598-022-08483-8
Ge, X., Wang, C., Wang, L., Zhou, B., Cao, Y., Xiao, W., et al. (2022). Drought changes litter quantity and quality, and soil microbial activities to affect soil nutrients in moso bamboo forest. Sci. Total Environ. 838:156351. doi: 10.1016/j.scitotenv.2022.156351
Ge, X. G., Tong, R., Cao, Y. H., Zhou, B. Z., Xiao, W. F., Wang, X. M., et al. (2018). [Effect of litterfall input on soil respiration and its temperature sensitivity in moso bamboo forest under simulated drought]. Ying Yong Sheng Tai Xue Bao 29, 2233–2242.
Geisseler, D., Horwath, W. R., Joergensen, R. G., and Ludwig, B. (2010). Pathways of nitrogen utilization by soil microorganisms – A review. Soil Biol. Biochem. 42, 2058–2067. doi: 10.1016/j.soilbio.2010.08.021
Groffman, P. M., Fahey, T. J., Fisk, M. C., Yavitt, J. B., Sherman, R. E., Bohlen, P. J., et al. (2015). Earthworms increase soil microbial biomass carrying capacity and nitrogen retention in northern hardwood forests. Soil Biol. Biochem. 87, 51–58. doi: 10.1016/j.soilbio.2015.03.025
Guan, B., Xie, B., Yang, S., Hou, A., Chen, M., and Han, G. (2019). Effects of five years’ nitrogen deposition on soil properties and plant growth in a salinized reed wetland of the Yellow River Delta. Ecol. Eng. 136, 160–166. doi: 10.1016/j.ecoleng.2019.06.016
Guan, F., Xia, M., Tang, X., and Fan, S. (2017). Spatial variability of soil nitrogen, phosphorus and potassium contents in Moso bamboo forests in Yong’an City, China. CATENA 150, 161–172. doi: 10.1016/j.catena.2016.11.017
Guo, J., Feng, H., Roberge, G., Feng, L., Pan, C., Mcnie, P., et al. (2022). The negative effect of Chinese fir (Cunninghamia lanceolata) monoculture plantations on soil physicochemical properties, microbial biomass, fungal communities, and enzymatic activities. For. Ecol. Manag. 519:120297. doi: 10.1016/j.foreco.2022.120297
Hu, M., Liu, Y., Wang, T., Hao, Y., Li, Z., and Wan, S. (2020). Fire alters soil properties and vegetation in a coniferous–broadleaf mixed forest in central china. Forests 11:164. doi: 10.3390/f11020164
Hu, M., Ma, Z., and Chen, H. Y. H. (2022). Intensive plantations decouple fine root C:N:P in subtropical forests. For. Ecol. Manag. 505:119901. doi: 10.1016/j.foreco.2021.119901
Isobe, K., Ikutani, J., Fang, Y., Yoh, M., Mo, J., Suwa, Y., et al. (2018a). Highly abundant acidophilic ammonia-oxidizing archaea causes high rates of nitrification and nitrate leaching in nitrogen-saturated forest soils. Soil Biol. Biochem. 122, 220–227. doi: 10.1016/j.soilbio.2018.04.021
Isobe, K., Oka, H., Watanabe, T., Tateno, R., Urakawa, R., Liang, C., et al. (2018b). High soil microbial activity in the winter season enhances nitrogen cycling in a cool-temperate deciduous forest. Soil Biol. Biochem. 124, 90–100. doi: 10.1016/j.soilbio.2018.05.028
James, J. A., Kern, C. C., and Miesel, J. R. (2018). Legacy effects of prescribed fire season and frequency on soil properties in a Pinus resinosa forest in northern Minnesota. For. Ecol. Manag. 415-416, 47–57. doi: 10.1016/j.foreco.2018.01.021
Jang, J., Xiong, X., Liu, C., Yoo, K., and Ishii, S. (2022). Invasive earthworms alter forest soil microbiomes and nitrogen cycling. Soil Biol. Biochem. 171:108724. doi: 10.1016/j.soilbio.2022.108724
Ji, L., Yang, Y., Yang, N., Khan, A., and Yang, L. (2021). Seasonal variation of diversity and co-occurrence patterns of arbuscular mycorrhizal fungal communities in mixed broadleaf-conifer forests. Appl. Soil Ecol. 158:103782. doi: 10.1016/j.apsoil.2020.103782
Jin, K.-M., Zhuo, R.-Y., Xu, D., Wang, Y.-J., Fan, H.-J., Huang, B.-Y., et al. (2020). Genome-wide identification of the expansin gene family and its potential association with drought stress in Moso Bamboo. Int. J. Mol. Sci. 21:9491. doi: 10.3390/ijms21249491
Joshi, R. K., and Garkoti, S. C. (2021). Influence of nepalese alder on soil physico-chemical properties and fine root dynamics in white oak forests in the central Himalaya, India. CATENA 200:105140. doi: 10.1016/j.catena.2020.105140
Kamruzzaman, M., Basak, K., Paul, S. K., Ahmed, S., and Osawa, A. (2019). Litterfall production, decomposition and nutrient accumulation in Sundarbans mangrove forests, Bangladesh. For. Sci. Technol. 15, 24–32. doi: 10.1080/21580103.2018.1557566
Kara, O., Bolat, I., Cakıroglu, K., and Senturk, M. (2014). Litter decomposition and microbial biomass in temperate forests in Northwestern Turkey. J. Soil Sci. Plant Nutr. 14, 31–41.
Komarov, A., Chertov, O., Bykhovets, S., Shaw, C., Nadporozhskaya, M., Frolov, P., et al. (2017). Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing. Ecol. Modell. 345, 113–124. doi: 10.1016/j.ecolmodel.2016.08.007
Kooch, Y., and Noghre, N. (2020). Nutrient cycling and soil-related processes under different land covers of semi-arid rangeland ecosystems in northern Iran. Catena 193:104621. doi: 10.1016/j.catena.2020.104621
Kooch, Y., Sanji, R., and Tabari, M. (2018). Increasing tree diversity enhances microbial and enzyme activities in temperate Iranian forests. Trees 32, 809–822. doi: 10.1007/s00468-018-1674-3
Kumar, P., Chen, H. Y. H., Thomas, S. C., and Shahi, C. (2018). Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 106, 1266–1276. doi: 10.1111/1365-2745.12861
Kumar, S., Meena, R. S., Datta, R., Verma, S. K., Yadav, G. S., Pradhan, G., et al. (2020). “Legumes for carbon and nitrogen cycling: An organic approach,” in Carbon and nitrogen cycling in soil, eds R. Datta, R. Meena, S. Pathan, and M. Ceccherini (Singapore: Springer), 337–375. doi: 10.1007/978-981-13-7264-3_10
Kurniawan, S., Utami, S. R., Mukharomah, M., Navarette, I. A., and Prasetya, B. (2019). Land use systems, soil texture, control carbon and nitrogen storages in the forest soil of UB forest, Indonesia. AGRIVITA 41, 416–427. doi: 10.17503/agrivita.v41i3.2236
Leppälammi-Kujansuu, J., Aro, L., Salemaa, M., Hansson, K., Kleja, D. B., and Helmisaari, H.-S. (2014). Fine root longevity and carbon input into soil from below-and aboveground litter in climatically contrasting forests. For. Ecol. Manag. 326, 79–90. doi: 10.1016/j.foreco.2014.03.039
Li, Q., Ma, Q., Gao, J., Zhang, J., Li, Y., Shi, M., et al. (2022). Stumps increased soil respiration in a subtropical Moso bamboo (Phyllostachys edulis) plantation under nitrogen addition. Agric. For. Meteorol. 323:109047. doi: 10.1016/j.agrformet.2022.109047
Li, Q., Peng, C., Zhang, J., Li, Y., and Song, X. (2021). Nitrogen addition decreases methane uptake caused by methanotroph and methanogen imbalances in a Moso bamboo forest. Sci. Rep. 11:5578. doi: 10.1038/s41598-021-84422-3
Li, Y., Xu, X., Lei, B., Zhuang, J., Zhang, X., Hu, C., et al. (2021). Magnesium-nitrogen co-doped carbon dots enhance plant growth through multifunctional regulation in photosynthesis. Chem. Eng. J. 422:130114. doi: 10.1016/j.cej.2021.130114
Lilleskov, E. A., Kuyper, T. W., Bidartondo, M. I., and Hobbie, E. A. (2019). Atmospheric nitrogen deposition impacts on the structure and function of forest mycorrhizal communities: A review. Environ. Pollut. 246, 148–162. doi: 10.1016/j.envpol.2018.11.074
Lobell, D. B., Ortiz-Monasterio, J. I., and Asner, G. P. (2004). Relative importance of soil and climate variability for nitrogen management in irrigated wheat. Field Crops Res. 87, 155–165. doi: 10.1016/j.fcr.2003.10.004
Lu, X., Mo, J., Gilliam, F. S., Yu, G., Zhang, W., Fang, Y., et al. (2011). Effects of experimental nitrogen additions on plant diversity in tropical forests of contrasting disturbance regimes in southern China. Environ. Pollut. 159, 2228–2235. doi: 10.1016/j.envpol.2010.10.037
Lu, Z. X., Wang, P., Ou, H. B., Wei, S. X., Wu, L. C., Jiang, Y., et al. (2022). Effects of different vegetation restoration on soil nutrients, enzyme activities, and microbial communities in degraded karst landscapes in southwest China. For. Ecol. Manag. 508:120002. doi: 10.1016/j.foreco.2021.120002
Lucas-Borja, M. E., De Las Heras, J., Moya Navarro, D., González-Romero, J., Peña-Molina, E., Navidi, M., et al. (2022). Short-term effects of prescribed fires with different severity on rainsplash erosion and physico-chemical properties of surface soil in Mediterranean forests. J. Environ. Manag. 322:116143. doi: 10.1016/j.jenvman.2022.116143
Luce, M. S., Whalen, J. K., Ziadi, N., and Zebarth, B. J. (2011). Nitrogen dynamics and indices to predict soil nitrogen supply in humid temperate soils. Adv. Agron. 112, 55–102. doi: 10.1016/B978-0-12-385538-1.00002-0
Lv, J., Li, Q., Zhang, J., Li, Y., Peng, C., and Song, X. (2022). Nitrogen addition increases CO2, CH4, and N2O concentrations of topsoils and subsoils in a subtropical Moso bamboo forest. CATENA 216:106397. doi: 10.1016/j.catena.2022.106397
Lyu, M., Xie, J., Vadeboncoeur, M. A., Wang, M., Qiu, X., Ren, Y., et al. (2018). Simulated leaf litter addition causes opposite priming effects on natural forest and plantation soils. Biol. Fertil. Soils 54, 925–934. doi: 10.1007/s00374-018-1314-5
Ma, H., Yin, Y., Gao, R., Taqi, R., and He, X. (2019). Response of nitrogen transformation to glucose additions in soils at two subtropical forest types subjected to simulated nitrogen deposition. J. Soils Sediments 19, 2166–2175. doi: 10.1007/s11368-018-02237-8
Ma, W., Li, Z., Ding, K., Huang, B., Nie, X., Lu, Y., et al. (2016). Soil erosion, organic carbon and nitrogen dynamics in planted forests: A case study in a hilly catchment of Hunan Province, China. Soil Tillage Res. 155, 69–77. doi: 10.1016/j.still.2015.07.007
Martin, G. D., Morrissey, E. M., Carson, W. P., and Freedman, Z. B. (2022). A legacy of fire emerges from multiple disturbances to most shape microbial and nitrogen dynamics in a deciduous forest. Soil Biol. Biochem. 169:108672. doi: 10.1016/j.soilbio.2022.108672
Matos, P. S., Silva, C. F. D., Damian, J. M., Cerri, C. E. P., Pereira, M. G., and Zonta, E. (2021). Beneficial services of Glomalin and Arbuscular Mycorrhizal fungi in degraded soils in Brazil. Sci. Agric. 79, 1–13. doi: 10.1590/1678-992x-2021-0064
Molla, E., Getnet, K., and Mekonnen, M. (2022). Land use change and its effect on selected soil properties in the northwest highlands of Ethiopia. Heliyon 8:e10157. doi: 10.1016/j.heliyon.2022.e10157
Muhammad, I., Yang, L., Ahmad, S., Farooq, S., Al-Ghamdi, A. A., Khan, A., et al. (2022). Nitrogen fertilizer modulates plant growth, chlorophyll pigments and enzymatic activities under different irrigation regimes. Agronomy 12:845. doi: 10.3390/agronomy12040845
Mushinski, R. M., Payne, Z. C., Raff, J. D., Craig, M. E., Pusede, S. E., Rusch, D. B., et al. (2021). Nitrogen cycling microbiomes are structured by plant mycorrhizal associations with consequences for nitrogen oxide fluxes in forests. Glob. Change Biol. 27, 1068–1082. doi: 10.1111/gcb.15439
Na, M., Hicks, L. C., Zhang, Y., Shahbaz, M., Sun, H., and Rousk, J. (2022). Semi-continuous C supply reveals that priming due to N-mining is driven by microbial growth demands in temperate forest plantations. Soil Biol. Biochem. 173:108802. doi: 10.1016/j.soilbio.2022.108802
Neves, N., Paula, R., Araujo, E., Gorsani, R., Abreu, K., and Kunz, S. (2022). Contribution of legume and non-legume trees to litter dynamics and C-N-P inputs in a secondary seasonally dry tropical forest. iForest 15, 8–15. doi: 10.3832/ifor3442-014
Ngaba, M. J. Y., Uwiragiye, Y., Bol, R., De Vries, W., and Zhou, J. (2022). Low-level nitrogen and short-term addition increase soil carbon sequestration in Chinese forest ecosystems. CATENA 215:106333. doi: 10.1016/j.catena.2022.106333
Ni, H., Su, W., Fan, S., and Chu, H. (2021). Effects of intensive management practices on rhizosphere soil properties, root growth, and nutrient uptake in Moso bamboo plantations in subtropical China. For. Ecol. Manag. 493:119083. doi: 10.1016/j.foreco.2021.119083
Nie, Y., Wang, M., Zhang, W., Ni, Z., Hashidoko, Y., and Shen, W. (2018). Ammonium nitrogen content is a dominant predictor of bacterial community composition in an acidic forest soil with exogenous nitrogen enrichment. Sci. Total Environ. 624, 407–415. doi: 10.1016/j.scitotenv.2017.12.142
Osono, T. (2015). Hyphal length in the forest floor and soil of subtropical, temperate, and subalpine forests. J. For. Res. 20, 69–76. doi: 10.1007/s10310-014-0461-2
Pandey, C. B., Singh, G. B., Singh, S. K., and Singh, R. K. (2010). Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of India. Plant Soil 333, 453–467. doi: 10.1007/s11104-010-0362-x
Park, J. S., and Ro, H. M. (2018). Temporal variations in soil profile carbon and nitrogen during three consecutive years of 15N deposition in temperate oak and pine forest stands. Forests 9:338. doi: 10.3390/f9060338
Peng, Y., Song, S. Y., Li, Z. Y., Li, S., Chen, G. T., Hu, H. L., et al. (2020). Influences of nitrogen addition and aboveground litter-input manipulations on soil respiration and biochemical properties in a subtropical forest. Soil Biol. Biochem. 142:107694. doi: 10.1016/j.soilbio.2019.107694
Perakis, S. S., and Pett-Ridge, J. C. (2019). Nitrogen-fixing red alder trees tap rock-derived nutrients. Proc. Natl. Acad. Sci. U.S.A. 116, 5009–5014. doi: 10.1073/pnas.1814782116
Prasad, S., Malav, L. C., Choudhary, J., Kannojiya, S., Kundu, M., Kumar, S., et al. (2021). “Soil microbiomes for healthy nutrient recycling,” in Current trends in microbial biotechnology for sustainable agriculture, eds A. N. Yadav, J. Singh, C. Singh, and N. Yadav (Singapore: Springer), 1–21. doi: 10.1007/978-981-15-6949-4_1
Purwanto, B. H., and Alam, S. (2020). Impact of intensive agricultural management on carbon and nitrogen dynamics in the humid tropics. Soil Sci. Plant Nutr. 66, 50–59. doi: 10.1080/00380768.2019.1705182
Qiao, Y., Wang, J., Liu, H., Huang, K., Yang, Q., Lu, R., et al. (2020). Depth-dependent soil C-N-P stoichiometry in a mature subtropical broadleaf forest. Geoderma 370:114357. doi: 10.1016/j.geoderma.2020.114357
Qin, H., Xing, X., Tang, Y., Hou, H., Yang, J., Shen, R., et al. (2019). Linking soil N2O emissions with soil microbial community abundance and structure related to nitrogen cycle in two acid forest soils. Plant Soil 435, 95–109. doi: 10.1007/s11104-018-3863-7
Rabin, S., Gérard, F., and Arneth, A. (2022). The influence of thinning and prescribed burning on future forest fires in fire-prone regions of Europe. Environ. Res. Lett. 17:055010. doi: 10.1088/1748-9326/ac6312
Ramm, E., Liu, C., Mueller, C. W., Gschwendtner, S., Yue, H., Wang, X., et al. (2022). Alder-induced stimulation of soil gross nitrogen turnover in a permafrost-affected peatland of Northeast China. Soil Biol. Biochem. 172:108757. doi: 10.1016/j.soilbio.2022.108757
Reis, C. R. G., Nardoto, G. B., and Oliveira, R. S. (2017). Global overview on nitrogen dynamics in mangroves and consequences of increasing nitrogen availability for these systems. Plant Soil 410, 1–19. doi: 10.1007/s11104-016-3123-7
Růžek, M., Tahovská, K., Guggenberger, G., and Oulehle, F. (2021). Litter decomposition in European coniferous and broadleaf forests under experimentally elevated acidity and nitrogen addition. Plant Soil 463, 471–485. doi: 10.1007/s11104-021-04926-9
Sarai, S. S., De Jong, B. H. J., Esperanza, H. L., Jorge, M. V., Danilo, M. R., and Aryal, D. R. (2022). Fine root biomass stocks but not the production and turnover rates vary with the age of tropical successional forests in Southern Mexico. Rhizosphere 21:100474. doi: 10.1016/j.rhisph.2022.100474
Schiefer, F., Kattenborn, T., Frick, A., Frey, J., Schall, P., Koch, B., et al. (2020). Mapping forest tree species in high resolution UAV-based RGB-imagery by means of convolutional neural networks. ISPRS J. Photogramm. Remote Sens. 170, 205–215. doi: 10.1016/j.isprsjprs.2020.10.015
Schulte-Uebbing, L., and de Vries, W. (2018). Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: A meta-analysis. Glob. Change Biol. 24, 416–431. doi: 10.1111/gcb.13862
Selmants, P. C., Adair, K. L., Litton, C. M., Giardina, C. P., and Schwartz, E. (2016). Increases in mean annual temperature do not alter soil bacterial community structure in tropical montane wet forests. Ecosphere 7:e01296. doi: 10.1002/ecs2.1296
Shi, J., Ohte, N., Tokuchi, N., Imamura, N., Nagayama, M., Oda, T., et al. (2014). Nitrate isotopic composition reveals nitrogen deposition and transformation dynamics along the canopy–soil continuum of a suburban forest in Japan. Rapid Commun. Mass Spectrom. 28, 2539–2549. doi: 10.1002/rcm.7050
Shi, M., Zhang, J., Sun, J., Li, Q., Lin, X., and Song, X. (2022). Unequal nitrogen translocation pattern caused by clonal integration between connected ramets ensures necessary nitrogen supply for young Moso bamboo growth. Environ. Exp. Bot. 200:104900. doi: 10.1016/j.envexpbot.2022.104900
Shi, W., Ju, Y., Bian, R., Li, L., Joseph, S., Mitchell, D. R., et al. (2020). Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci. Total Environ. 701:134424. doi: 10.1016/j.scitotenv.2019.134424
Shinohara, Y., and Otsuki, K. (2015). Comparisons of soil-water content between a Moso bamboo (Phyllostachys pubescens) forest and an evergreen broadleaved forest in western Japan. Plant Species Biol. 30, 96–103. doi: 10.1111/1442-1984.12076
Singh, D., Sharma, P., Kumar, U., Daverey, A., and Arunachalam, K. (2021). Effect of forest fire on soil microbial biomass and enzymatic activity in oak and pine forests of Uttarakhand Himalaya, India. Ecol. Process. 10:29. doi: 10.1186/s13717-021-00293-6
Siwach, A., Kaushal, S., and Baishya, R. (2021). Effect of Mosses on physical and chemical properties of soil in temperate forests of Garhwal Himalayas. J. Trop. Ecol. 37, 126–135. doi: 10.1017/S0266467421000249
Smith, G. R., and Wan, J. (2019). Resource-ratio theory predicts mycorrhizal control of litter decomposition. New Phytol. 223, 1595–1606. doi: 10.1111/nph.15884
Solly, E. F., Schöning, I., Boch, S., Kandeler, E., Marhan, S., Michalzik, B., et al. (2014). Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant Soil 382, 203–218. doi: 10.1007/s11104-014-2151-4
Sponseller, R. A., Gundale, M. J., Futter, M., Ring, E., Nordin, A., Näsholm, T., et al. (2016). Nitrogen dynamics in managed boreal forests: Recent advances and future research directions. Ambio 45, 175–187. doi: 10.1007/s13280-015-0755-4
Staccone, A., Liao, W., Perakis, S., Compton, J., Clark, C. M., and Menge, D. (2020). A spatially explicit, empirical estimate of tree-based biological nitrogen fixation in forests of the United States. Glob. Biogeochem. Cycles 34:e2019GB006241. doi: 10.1029/2019GB006241
Su, F., Xu, S., Sayer, E. J., Chen, W., Du, Y., and Lu, X. (2021). Distinct storage mechanisms of soil organic carbon in coniferous forest and evergreen broadleaf forest in tropical China. J. Environ. Manag. 295:113142. doi: 10.1016/j.jenvman.2021.113142
Sun, X., Wang, G., Ye, Y., Ma, Q., Guan, Q., and Jones, D. L. (2021). Response of nitrogen fractions in the rhizosphere and bulk soil to organic mulching in an urban forest plantation. J. For. Res. 32, 2577–2588. doi: 10.1007/s11676-021-01310-2
Sun, Y., Xiong, X., He, M., Xu, Z., Hou, D., Zhang, W., et al. (2021). Roles of biochar-derived dissolved organic matter in soil amendment and environmental remediation: A critical review. Chem. Eng. J. 424:130387. doi: 10.1016/j.cej.2021.130387
Tahmasbian, I., Xu, Z., Nguyen, T. T. N., Che, R., Omidvar, N., Lambert, G., et al. (2019). Short-term carbon and nitrogen dynamics in soil, litterfall and canopy of a suburban native forest subjected to prescribed burning in subtropical Australia. J. Soils Sediments 19, 3969–3981. doi: 10.1007/s11368-019-02430-3
Tan, X., Machmuller, M. B., Huang, F., He, J., Chen, J., Cotrufo, M. F., et al. (2020). Temperature sensitivity of ecoenzyme kinetics driving litter decomposition: The effects of nitrogen enrichment, litter chemistry, and decomposer community. Soil Biol. Biochem. 148:107878. doi: 10.1016/j.soilbio.2020.107878
Tang, Y., Yu, G., Zhang, X., Wang, Q., Ge, J., and Liu, S. (2018). Changes in nitrogen-cycling microbial communities with depth in temperate and subtropical forest soils. Appl. Soil Ecol. 124, 218–228. doi: 10.1016/j.apsoil.2017.10.029
Thorn, S., Seibold, S., Leverkus, A. B., Michler, T., Müller, J., Noss, R. F., et al. (2020). The living dead: Acknowledging life after tree death to stop forest degradation. Front. Ecol. Environ. 18, 505–512. doi: 10.1002/fee.2252
Tian, J., Dungait, J. A., Lu, X., Yang, Y., Hartley, I. P., Zhang, W., et al. (2019). Long-term nitrogen addition modifies microbial composition and functions for slow carbon cycling and increased sequestration in tropical forest soil. Glob. Change Biol. 25, 3267–3281. doi: 10.1111/gcb.14750
Tong, R., Zhou, B., Cao, Y., Ge, X., and Jiang, L. (2020). Metabolic profiles of moso bamboo in response to drought stress in a field investigation. Sci. Total Environ. 720:137722. doi: 10.1016/j.scitotenv.2020.137722
Toro-Manríquez, M., Soler, R., Lencinas, M. V., and Promis, Á. (2019). Canopy composition and site are indicative of mineral soil conditions in Patagonian mixed Nothofagus forests. Ann. For. Sci. 76:117. doi: 10.1007/s13595-019-0886-z
United Nation’s Environment Program. (2022). Earth’ biodiversity depends on the world’s forests. Available at: https://www.unep-wcmc.org/en/news/earths-biodiversity-depends-on-the-worlds-forests (accessed March 21, 2023).
Ushio, M., Aiba, S.-I., Takeuchi, Y., Iida, Y., Matsuoka, S., Repin, R., et al. (2017). Plant–soil feedbacks and the dominance of conifers in a tropical montane forest in Borneo. Ecol. Monogr. 87, 105–129. doi: 10.1002/ecm.1236
Ushio, M., Balser, T. C., and Kitayama, K. (2013). Effects of condensed tannins in conifer leaves on the composition and activity of the soil microbial community in a tropical montane forest. Plant Soil 365, 157–170. doi: 10.1007/s11104-012-1365-6
Vitousek, P. M., and Denslow, J. S. (1986). Nitrogen and phosphorus availability in treefall gaps of a lowland tropical rainforest. J. Ecol. 74, 1167–1178. doi: 10.2307/2260241
Vuong, T.-M.-D., Zeng, J.-Y., and Man, X.-L. (2020). Soil fungal and bacterial communities in southern boreal forests of the Greater Khingan Mountains and their relationship with soil properties. Sci. Rep. 10:22025. doi: 10.1038/s41598-020-79206-0
Wang, G., Wang, L., and Ma, F. (2022a). Effects of earthworms and arbuscular mycorrhizal fungi on improvement of fertility and microbial communities of soils heavily polluted by cadmium. Chemosphere 286:131567. doi: 10.1016/j.chemosphere.2021.131567
Wang, J., Han, S., Wang, C., and Li, M.-H. (2022b). Long-term nitrogen-addition-induced shifts in the ectomycorrhizal fungal community are associated with changes in fine root traits and soil properties in a mixed Pinus koraiensis forest. Eur. J. Soil Biol. 112:103431. doi: 10.1016/j.ejsobi.2022.103431
Wang, J., Zheng, Y., Shi, X., Lam, S. K., Lucas-Borja, M. E., and Huang, Z. (2022c). Nature restoration shifts the abundance and structure of soil nematode communities in subtropical forests. Plant Soil 471, 315–327. doi: 10.1007/s11104-021-05229-9
Wang, Q.-W., Robson, T. M., Pieristè, M., Kenta, T., Zhou, W., and Kurokawa, H. (2022d). Canopy structure and phenology modulate the impacts of solar radiation on C and N dynamics during litter decomposition in a temperate forest. Sci. Total Environ. 820:153185. doi: 10.1016/j.scitotenv.2022.153185
Wang, S., Zhao, S., Yang, B., Zhang, K., Fan, Y., Zhang, L., et al. (2022e). The carbon and nitrogen stoichiometry in litter-soil-microbe continuum rather than plant diversity primarily shapes the changes in bacterial communities along a tropical forest restoration chronosequence. CATENA 213:106202. doi: 10.1016/j.catena.2022.106202
Wang, H., Liu, S., Song, Z., Yang, Y., Wang, J., You, Y., et al. (2019). Introducing nitrogen-fixing tree species and mixing with Pinus massoniana alters and evenly distributes various chemical compositions of soil organic carbon in a planted forest in southern China. For. Ecol. Manag. 449:117477. doi: 10.1016/j.foreco.2019.117477
Wang, H., Liu, S., Wang, J., You, Y., Yang, Y., Shi, Z., et al. (2018). Mixed-species plantation with Pinus massoniana and Castanopsis hystrix accelerates C loss in recalcitrant coniferous litter but slows C loss in labile broadleaf litter in southern China. For. Ecol. Manag. 422, 207–213.
Wang, S., Li, J., Zhang, Z., Chen, M., Li, S., and Cao, R. (2019). Feeding-strategy effect of Pheidole ants on microbial carbon and physicochemical properties in tropical forest soils. Appl. Soil Ecol. 133, 177–185. doi: 10.1016/j.apsoil.2018.10.006
Wang, H.-C., Chou, C.-Y., Chiou, C.-R., Tian, G., and Chiu, C.-Y. (2016a). Humic acid composition and characteristics of soil organic matter in relation to the elevation gradient of moso bamboo plantations. PLoS One 11:e0162193. doi: 10.1371/journal.pone.0162193
Wang, H.-C., Tian, G., and Chiu, C.-Y. (2016b). Invasion of moso bamboo into a Japanese cedar plantation affects the chemical composition and humification of soil organic matter. Sci. Rep. 6:32211. doi: 10.1038/srep32211
Wang, X., Sasaki, A., Toda, M., and Nakatsubo, T. (2016c). Changes in soil microbial community and activity in warm temperate forests invaded by moso bamboo (Phyllostachys pubescens). J. For. Res. 21, 235–243. doi: 10.1007/s10310-016-0533-6
Wang, Q., Xiao, J., Ding, J., Zou, T., Zhang, Z., Liu, Q., et al. (2021). Differences in root exudate inputs and rhizosphere effects on soil N transformation between deciduous and evergreen trees. Plant Soil 458, 277–289. doi: 10.1007/s11104-019-04156-0
Ward, E. B., Duguid, M. C., Kuebbing, S. E., Lendemer, J. C., and Bradford, M. A. (2022). The functional role of ericoid mycorrhizal plants and fungi on carbon and nitrogen dynamics in forests. New Phytol. 235, 1701–1718. doi: 10.1111/nph.18307
Wu, G., Chen, Z., Jiang, D., Jiang, N., Jiang, H., and Chen, L. (2021). Oxidases and hydrolases mediate soil organic matter accumulation in chernozem of Northeastern China. Geoderma 403:115206. doi: 10.1016/j.geoderma.2021.115206
Xiao, L., Li, C., Cai, Y., Zhou, T., Zhou, M., Gao, X., et al. (2021). Interactions between soil properties and the rhizome-root distribution in a 12-year Moso bamboo reforested region: Combining ground-penetrating radar and soil coring in the field. Sci. Total Environ. 800:149467. doi: 10.1016/j.scitotenv.2021.149467
Xiao, R., Man, X., Duan, B., and Cai, T. (2020). Short-term litter manipulations have strong impact on soil nitrogen dynamics in Larix gmelinii forest of Northeast China. Forests 11:1205. doi: 10.3390/f11111205
Xiao, R., Man, X., Duan, B., Cai, T., Ge, Z., Li, X., et al. (2022). Changes in soil bacterial communities and nitrogen mineralization with understory vegetation in boreal larch forests. Soil Biol. Biochem. 166:108572. doi: 10.1016/j.soilbio.2022.108572
Xing, A., Du, E., Shen, H., Xu, L., Zhao, M., Liu, X., et al. (2022). High-level nitrogen additions accelerate soil respiration reduction over time in a boreal forest. Ecol. Lett. 25, 1869–1878. doi: 10.1111/ele.14065
Xing, S., Chen, C., Zhou, B., Zhang, H., Nang, Z., and Xu, Z. (2010). Soil soluble organic nitrogen and active microbial characteristics under adjacent coniferous and broadleaf plantation forests. J. Soils Sediments 10, 748–757. doi: 10.1007/s11368-009-0159-9
Xiong, X., Lyu, M., Deng, C., Li, X., Lu, Y., Lin, W., et al. (2022). Carbon and nitrogen availability drives seasonal variation in soil microbial communities along an elevation gradient. Forests 13:1657. doi: 10.3390/f13101657
Xu, Q.-F., Jiang, P.-K., Wu, J.-S., Zhou, G.-M., Shen, R.-F., and Fuhrmann, J. J. (2015). Bamboo invasion of native broadleaf forest modified soil microbial communities and diversity. Biol. Invasions 17, 433–444. doi: 10.1007/s10530-014-0741-y
Xu, W., Elberling, B., and Ambus, P. L. (2022). Pyrogenic organic matter as a nitrogen source to microbes and plants following fire in an Arctic heath tundra. Soil Biol. Biochem. 170:108699. doi: 10.1016/j.soilbio.2022.108699
Xu, Z., Yu, G., Zhang, X., He, N., Wang, Q., Wang, S., et al. (2018). Biogeographical patterns of soil microbial community as influenced by soil characteristics and climate across Chinese forest biomes. Appl. Soil Ecol. 124, 298–305. doi: 10.1016/j.apsoil.2017.11.019
Yan, M., Pan, G., Lavallee, J. M., and Conant, R. T. (2020). Rethinking sources of nitrogen to cereal crops. Glob. Change Biol. 26, 191–199. doi: 10.1111/gcb.14908
Yang, C., Wang, A., Zhu, Z., Lin, S., Bi, Y., and Du, X. (2021). Impact of extensive management system on soil properties and carbon sequestration under an age chronosequence of Moso bamboo plantations in subtropical China. For. Ecol. Manag. 497:119535. doi: 10.1016/j.foreco.2021.119535
Yang, K., Zhu, J., Zhang, W., Gu, J., Wang, Z., and Xu, S. (2022). Comparison of soil chemical and microbial properties in monoculture larch and mixed plantations in a temperate forest ecosystem in Northeast China. Ecol. Process. 11:12. doi: 10.1186/s13717-022-00358-0
Yin, X., Zhao, L., Fang, Q., and Ding, G. (2021). Differences in soil physicochemical properties in different-aged Pinus massoniana plantations in Southwest China. Forests 12:987. doi: 10.3390/f12080987
Yokobe, T., Hyodo, F., and Tokuchi, N. (2018). Seasonal effects on microbial community structure and nitrogen dynamics in temperate forest soil. Forests 9:153. doi: 10.3390/f9030153
You, Y., Xu, H., Wu, X., Zhou, X., Tan, X., Li, M., et al. (2020). Native broadleaf tree species stimulate topsoil nutrient transformation by changing microbial community composition and physiological function, but not biomass in subtropical plantations with low P status. For. Ecol. Manag. 477:118491. doi: 10.1016/j.foreco.2020.118491
Yu, S., Sayer, E. J., Li, Z., Mo, Q., Wang, M., Li, Y., et al. (2022). Delayed wet season increases soil net N mineralization in a seasonally dry tropical forest. Sci. Total Environ. 823:153314. doi: 10.1016/j.scitotenv.2022.153314
Zhang, H., Zhuang, S., Qian, H., Wang, F., and Ji, H. (2015). Spatial variability of the topsoil organic carbon in the Moso bamboo forests of southern China in association with soil properties. PLoS One 10:e0119175. doi: 10.1371/journal.pone.0119175
Zhang, J., Lv, J., Li, Q., Ying, Y., Peng, C., and Song, X. (2017). Effects of nitrogen deposition and management practices on leaf litterfall and N and P return in a Moso bamboo forest. Biogeochemistry 134, 115–124. doi: 10.1007/s10533-017-0349-2
Zhang, Y., Liu, T., Guo, J., Tan, Z., Dong, W., and Wang, H. (2021). Changes in the understory diversity of secondary Pinus tabulaeformis forests are the result of stand density and soil properties. Glob. Ecol. Conserv. 28:e01628. doi: 10.1016/j.gecco.2021.e01628
Zhang, Y., Wang, Q., Xu, L., Ma, S., Cui, D., Zhu, K., et al. (2022). Mixed conifer-broadleaf trees on arbuscular mycorrhizal and ectomycorrhizal communities in rhizosphere soil of different plantation stands in the temperate zone, Northeast China. Front. Microbiol. 13:986515. doi: 10.3389/fmicb.2022.986515
Zheng, Y., Guan, F., Fan, S., Zhou, Y., and Jing, X. (2021). Functional trait responses to strip clearcutting in a Moso bamboo forest. Forests 12:793. doi: 10.3390/f12060793
Zhou, J., Zang, H., Loeppmann, S., Gube, M., Kuzyakov, Y., and Pausch, J. (2020). Arbuscular mycorrhiza enhances rhizodeposition and reduces the rhizosphere priming effect on the decomposition of soil organic matter. Soil Biol. Biochem. 140:107641. doi: 10.1016/j.soilbio.2019.107641
Keywords: forest ecosystem, bamboo invasion, nitrogen cycling, broadleaf forests, soil properties
Citation: Sardar MF, Younas F, Farooqi ZUR and Li YL (2023) Soil nitrogen dynamics in natural forest ecosystem: a review. Front. For. Glob. Change 6:1144930. doi: 10.3389/ffgc.2023.1144930
Received: 15 January 2023; Accepted: 24 April 2023;
Published: 12 May 2023.
Edited by:
Ziliang Zhang, University of Illinois Urbana-Champaign, United StatesReviewed by:
Mengya Song, Swedish University of Agricultural Sciences, SwedenCopyright © 2023 Sardar, Younas, Farooqi and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yanli Li, MTA3MTIzMjUzNUBxcS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.