
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change, 23 March 2023
Sec. Forest Management
Volume 6 - 2023 | https://doi.org/10.3389/ffgc.2023.1138317
This article is part of the Research TopicMountainous Forest Ecosystems: Challenges and Management ImplicationsView all 16 articles
 Jiahua Han1
Jiahua Han1 Huiyan Yin1
Huiyan Yin1 Jie Xue2,3,4*
Jie Xue2,3,4* Zhiwei Zhang1,5*Zhen Xing1Shunke Wang2,3,4Jingjing Chang6Xiaoqiang Chen1,7Bo Yu1
Zhiwei Zhang1,5*Zhen Xing1Shunke Wang2,3,4Jingjing Chang6Xiaoqiang Chen1,7Bo Yu1The vertical distribution understanding of undergrowth herbaceous plants in high altitude mountain areas is of great significance to reveal the environmental gradient change law of plant diversity. However, the species composition of herbaceous plants at different altitude gradients and the relationship between diversity and environmental factors is still vague. This study takes the Sejila Mountain in southeast Tibet at an altitude of 2,000–3,300 m as the study area to analyze the vertical distribution differences of the understory herbs and their driving factors on shady and sunny slopes. The results showed that: (1) The number of families, genera and species of herbaceous plants on sunny slope is greater than that on shady slope. Asteraceae on shady and sunny slopes has the largest number of species; (2) The α diversity index of the herbs on shady slope shows a U-shaped trend. The Cody heterogeneity index (βC) is negatively correlated with the β diversity index. The herb richness index (R) on sunny slope is unimodal. The β diversity index displays the same as that of shady slope; (3) Different environmental factors have different effects on herb diversity. The correlation between environmental factors and diversity indicators indicates that there is a certain degree of clustering among samples. The environmental factors of shady slope and sunny slope are separated; and (4) The difference of water and heat conditions caused by altitude and slope aspect is an important driving factor for the diversity of herbage species, followed by soil clay content, silt content and slope gradient. This study will help to understand the species diversity and ecosystem characteristics of the region, and provide a reference for the study of plant community structure and function and rational utilization of understory plant resources in high-altitude mountain areas.
Species diversity is the basis for understanding the composition, structure and functional state of community species, and also the basis for studying biodiversity (Butchart et al., 2010; Cardinale et al., 2012; Chitale et al., 2014). As the largest terrestrial ecosystem, the forests are an important carrier of species diversity. In addition to trees and shrubs, the herbs are also an important part of the forests. Although the plants of herbaceous plants are small and their biomass proportion is low, they have the highest species diversity. The herbs play an important role in maintaining the material cycle and energy flow of the forest ecosystem, and play an important role in ensuring the function and stability of the ecosystem (Gilliam, 2007; Brockerhoff et al., 2017).
The alpine ecosystems are the most sensitive regions to environmental change, suitable for studying the response of ecosystem to climate change and predicting the response to future climate change (Ecology and Kessler, 2012). Altitude gradient in alpine ecosystems integrates changes in water, temperature, light, soil and other factors, and is considered to be one of the decisive factors affecting species diversity (Andra, 2016; Yılmaz et al., 2018). Many scholars have done some research on species diversity at different altitudes, but the differences are large. Species diversity has a variety of altitude gradient distribution patterns. There was significant positive, negative, unimodal, or u-type correlation between species diversity index and altitude (Sklenář and Jørgensen, 1999; Sklenár and Ramsay, 2001; Austrheim, 2002; Doležal and Šrůtek, 2002; da Silva et al., 2014; Ent et al., 2016; Gómez-Díaz et al., 2017; Dip et al., 2020; Walter et al., 2021).
The Qinghai-Tibet Plateau, known as the “third pole of the world,” is one of the largest original forest regions in China and one of the provinces (regions) with the richest forest resources in China. It is of great significance for the study of undergrowth herbs in the study area. However, the research on herbaceous plants in the Qinghai-Tibet Plateau mainly focuses on the grassland degradation, plant diversity and biomass change of alpine meadow and alpine grassland in the source area of the Three Rivers, Gobi area in the north of the Qinghai-Tibet Plateau (Mu et al., 2018; Xue et al., 2018; Fayiah et al., 2019; Han et al., 2022; She et al., 2022). The research on plant diversity in southeast Tibet mainly focuses on woody plants, ferns and mosses (Li et al., 2013; Zu et al., 2019; Shi et al., 2021; Xian et al., 2021). The research on the diversity of undergrowth herbaceous plants mainly focuses on the composition of plant communities and species diversity in forest fire slash (Zhang et al., 2021, 2022; Yin et al., 2022; Zhou et al., 2022). There are relatively few studies on the diversity of undergrowth herbaceous plants.
Southeast Tibet is located in the southeast of the Qinghai-Tibet Plateau, the western line of Hengduan Mountains. As the Indian Ocean warm moisture flows northward, it brings a lot of rainfall every year, forming many forest vegetation types. The region has a variety of tropical, subtropical, temperate and cold climate zones. Influenced by the Indian Ocean warm and wet monsoon, it is cold and dry in winter, cold and rainy in summer with distinct dry and wet seasons. There are many types of primitive forests in this area, mainly dark coniferous forests (Wang et al., 2019; Xiangyang et al., 2022), and there are rich herbaceous plants under the forests. Studying the distribution characteristics of herbaceous plants at different altitudes and slope directions in Tongmai section of Sejila Mountain is helpful to understand the diversity and ecosystem characteristics of undergrowth herbaceous plants in southeast Tibet.
In this study, it is assumed that the distribution differences of the understory herbs is mainly affected by the alpine topography factors and soil conditions. Based on field investigation and relevant data, the diversity index of herbaceous plants was calculated and the difference analysis and principal component analysis were carried out. We studied the change pattern of herbage species composition and diversity index with altitude gradient in Tongmai section of Sejila Mountain, and the correlation between herbage diversity and environmental factors. The purpose is to provide reference for the study of plant community structure and function, rational utilization of understory plant resources, and ecological environment protection in the southeast of the high altitude Qinghai Tibet Plateau.
Sejila Mountain is located in Bayi District, Linzhi City, at the junction of Nianqing Tanggula Mountain and Himalayas in the southeast of Tibet. It is mainly from Tongmai Town to Lulang Town. The geographical location is 29.79°–30.13°N, 94.75°–95.06 °E (Figure 1). It is located in the transition zone between semi humid and humid areas in southeast Tibet, with unique plateau ecological characteristics. The area is warm in winter and cool in summer, with distinct dry and wet seasons. The average annual temperature is 6–12°C, the average temperature of the warmest month is 10–18°C, the number of days ≥ 0°C is 210–350 days. The annual precipitation is 1,000–1,200 mm. The soil is mainly mountainous brown soil. The arbor layer is mainly composed of moist coniferous and broad-leaved mixed forest zone in the warm temperate zone of mountains and cool moist dark coniferous forest zone in the temperate zone of mountains. The main dominant species are Alnus nepalensis, Populus szechuanica var.tibetica and Pinus densata. The dominant species in shrub layer are Viburnum kansuense, Rosa sericea and Elsholtzia fruticosa. The herb layer is very rich in species in this area, mainly including Pilea indolens, Duchesnea indica, Fragaria moupinensis, Eragrostis pilosa, Synotis saluenensis, Dryopteris redactopinnata, and Oplismus compositus.
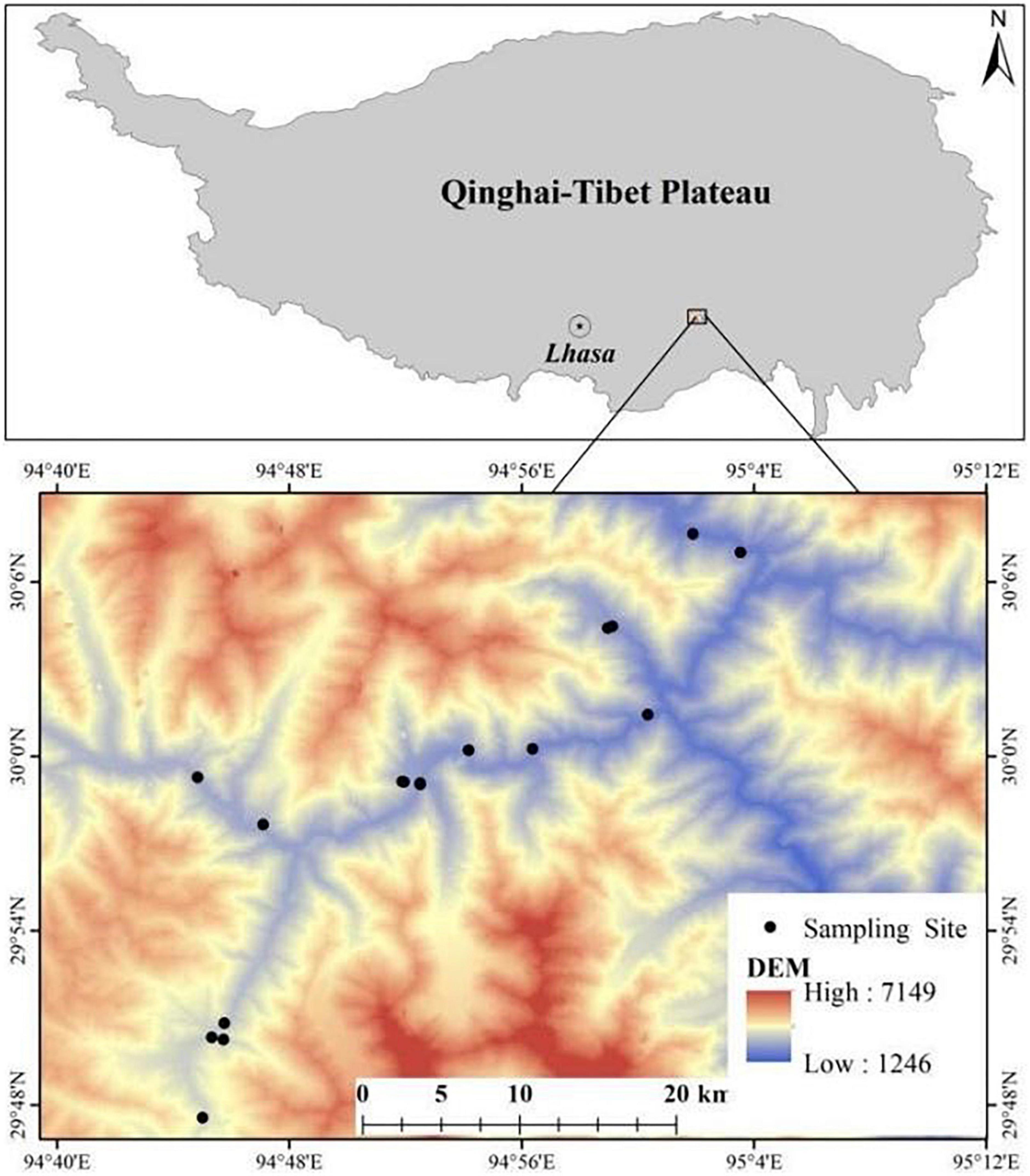
Figure 1. Location of the study area and distribution of soil sampling points.
In October 2021, the herbaceous plants were investigated in the Tongmai section of Sejila Mountain. The survey area has an altitude range of 2,000–3,300 m. The specific altitude range (2,000–3,300 m) spans the warm temperate zone and temperate climate zone. The vegetation zone corresponds to the mountain warm temperate mixed coniferous forest and dark coniferous forest zone, The herbaceous plants are quite different between the two climate zones, so it is of great significance to study the replacement and continuation of herbaceous species in the altitude range of 2,000–3,300 m. Taking 200 m as an altitude gradient zone, there are 6 altitude gradient zones in total. Three 20 m × 30 m standard plots are set on the shady and sunny slopes of each elevation gradient. The 1 m × 1 m standard herbaceous investigation quadrat is set at the four corners and the center of the sample plot. 15 herbaceous investigation quadrats were set on the shady slope and sunny slope respectively for each elevation gradient. The basic contents of the survey include plant name, number of plants/clusters, coverage, height, base diameter, growth and distribution. We measured the longitude and latitude, elevation, slope, aspect and other factors of the quadrat with the real-time dynamic measuring instrument (RTK), and we collected all the herb species in the quadrat. We mark the sample number, collector, longitude and latitude, altitude, slope, aspect and associated plants, and take them back to make specimens for later species identification. Table 1 shows the basic information of the sample plot. Due to the influence of topographic factors, the cliff affected the sampling work in the elevation gradient of 2,800–3,000 m on the sunny slope.
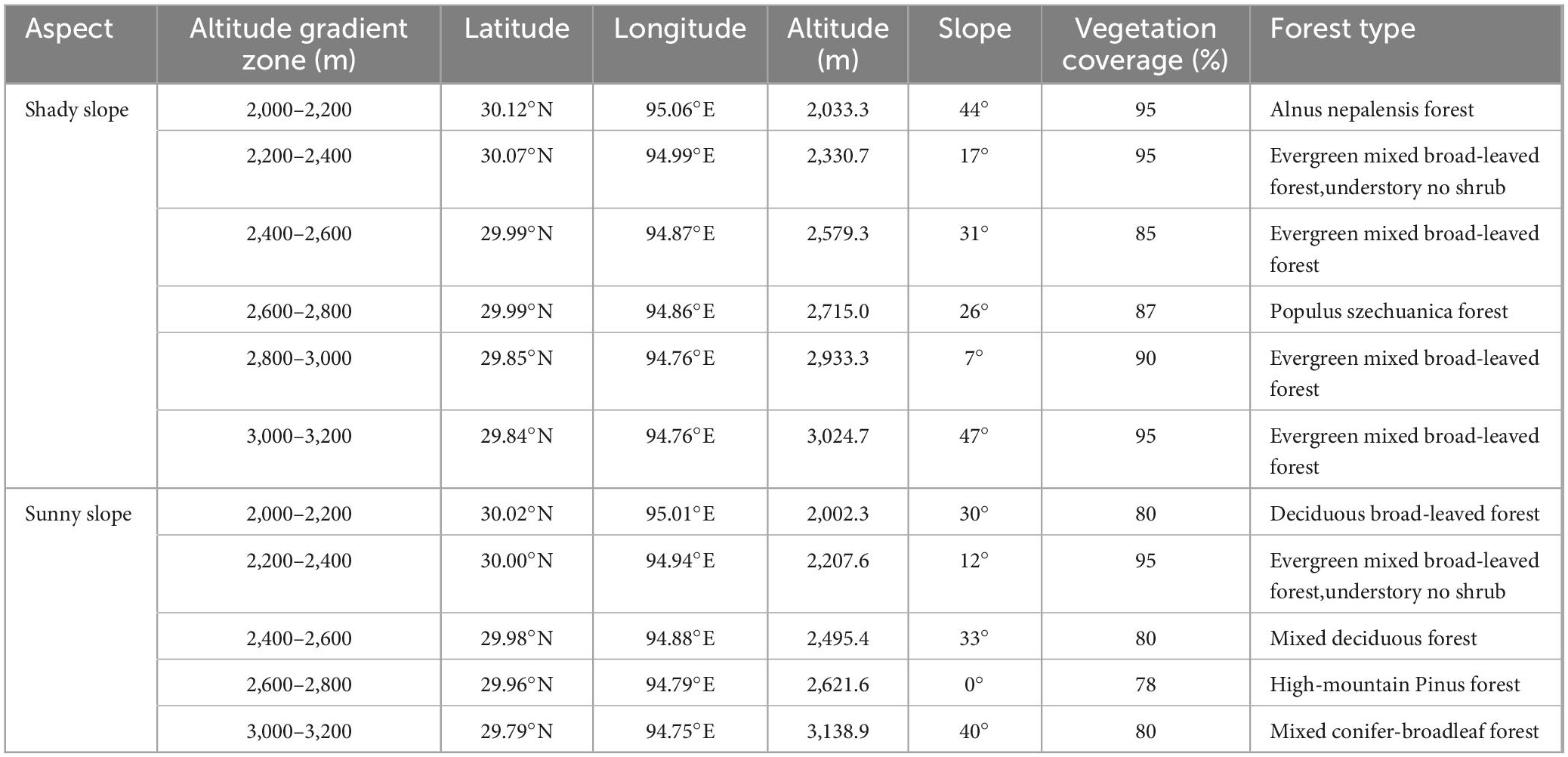
Table 1. Basic information of the research sites.
(1) The important value is a comprehensive quantitative index to study the status and role of a species in the community, which can comprehensively evaluate the status and role of a species in the community (Torresani et al., 2020; Wang et al., 2021), reflecting the structure and functional characteristics of the plant community. The calculation formula of the important value of herbaceous plants is:
where IV is the important value, RA is the relative abundance, RC is the relative coverage, and RH is the relative height.
(2) Species richness is an important indicator of plant diversity, and the calculation formula is:
where S is the number of quadrat species.
(3) α Diversity is used to describe the species diversity of local communities, which is generally affected by microenvironment and interactions between organisms (Ellum et al., 2010; Van Dam, 2019; Wang et al., 2021).
Shannon-Wiener diversity index:
Uniformity index Pielou:
where S is the number of species in the community, and Ni is the important value of species i in the sample plot.
(4) β Diversity refers to the diversity of community species composition between different habitats along the environmental gradient or the rate of species replacement along the environmental gradient (Ellum et al., 2010; Van Dam, 2019; Wang et al., 2021).
Sorenson similarity index:
Cody anisotropy index:
where a and b are the number of species in the 2 communities respectively, c is the number of common species in the 2 communities, g (H) is the number of species increased along the environmental gradient H, and l (H) is the number of species lost along the environmental gradient H.
We summarize and sort out the herbaceous plant specimens we brought back, and identify the collected samples one by one based on relevant materials such as Flora of China, Flora of Tibet, Flora of Botany,1 China Digital Herbarium (CVH)2 and local plant experts in Tibet. Soil particle size was measured by laser particle size distribution instrument, and PH was measured by soil moisture temperature PH velocity meter. Based on the statistical analysis of the composition of families, genera and species of herbaceous plants at different altitudes in Sejila Mountain, the differences were analyzed with SPSS 23 software.ArcGIS 10.5 was used to draw regional overview maps, and Origin 2021 was used to make herb diversity index maps and principal component analysis maps.
It can be seen from Table 2 that 57 species of herbaceous plants belonging to 24 families and 49 genera have been investigated on the shady slope, among which Asteraceae has the most plants. There are 10 species of Asteraceae, accounting for 17.54% of the total herb species investigated on the shady slope of Sejila Mountain. From the perspective of subordination, Asteraceae includes 9 genera, accounting for 18.37% of the total genera of herbs on the shady slope of Sejila Mountain. In the shady slope area, Asteraceae plants are dominant, Dryopteridaceae, Rosaceae and Apiaceae are important families in the shady slope of Sejila Mountain; Poaceae, Rubiaceae and Ranunculaceae enrich the vegetation composition of shady slopes.
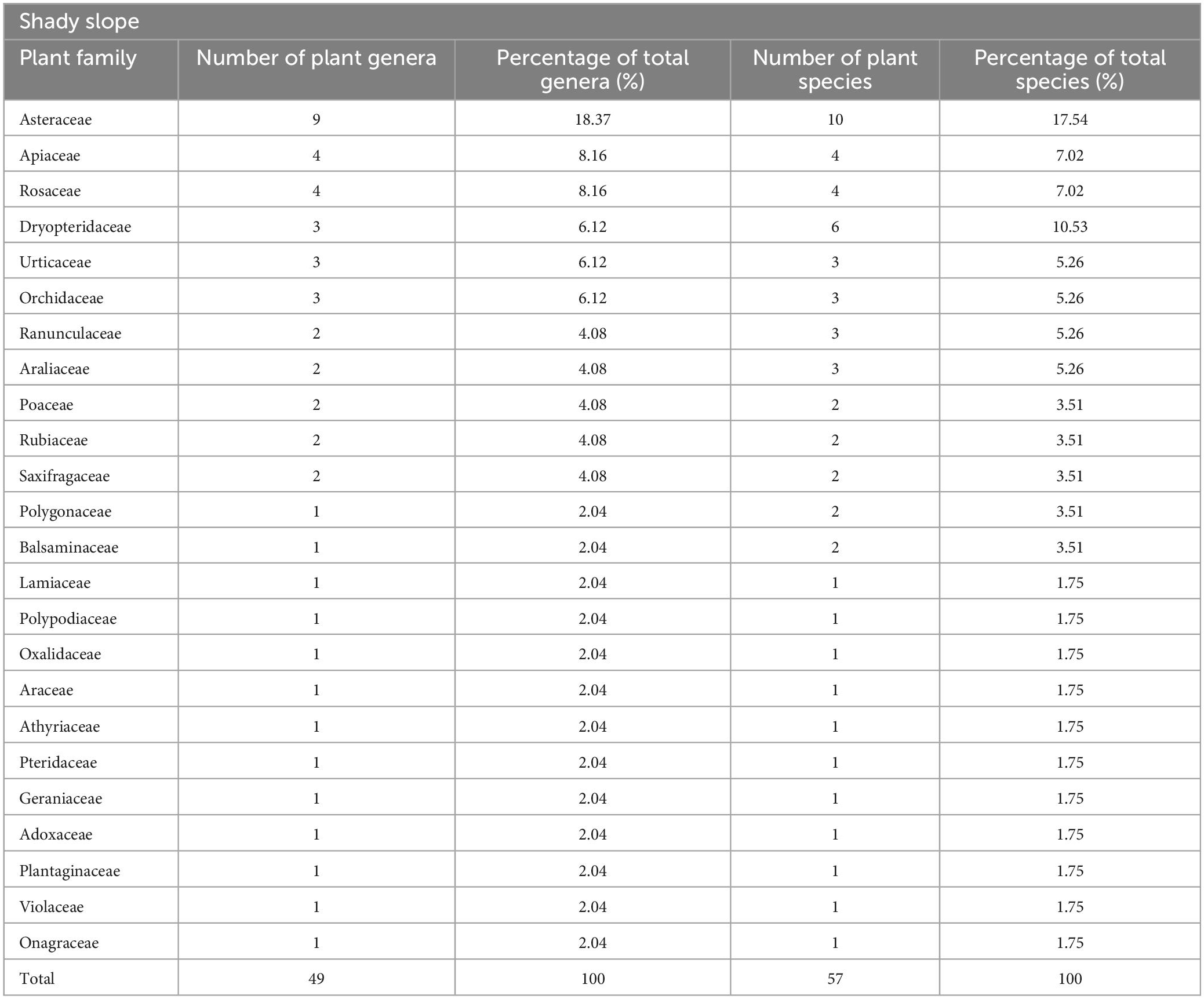
Table 2. Composition of herbaceous plants on shady slope.
In the sample plot on the sunny slope, according to Table 3, 73 species of herbaceous plants belonging to 35 families and 62 genera are found on the sunny slope, with the largest number of Asteraceae plants. There are 9 species of Asteraceae, accounting for 12.33% of the total herbaceous species investigated on the sunny slope of Sejila Mountain. From the perspective of subordinate species, Asteraceae includes 7 genera, accounting for 11.29% of the total genera of herbaceous plants on the sunny slope of Sejila Mountain. It can be seen that Asteraceae plants are dominant in the sunny slope of study area, and Lamiaceae and Rosaceae are important constituent families in the sunny slope of Sejila Mountain; Zingiberaceae, Geraniaceae and Rubiaceae enrich the vegetation composition of the sunny slope.
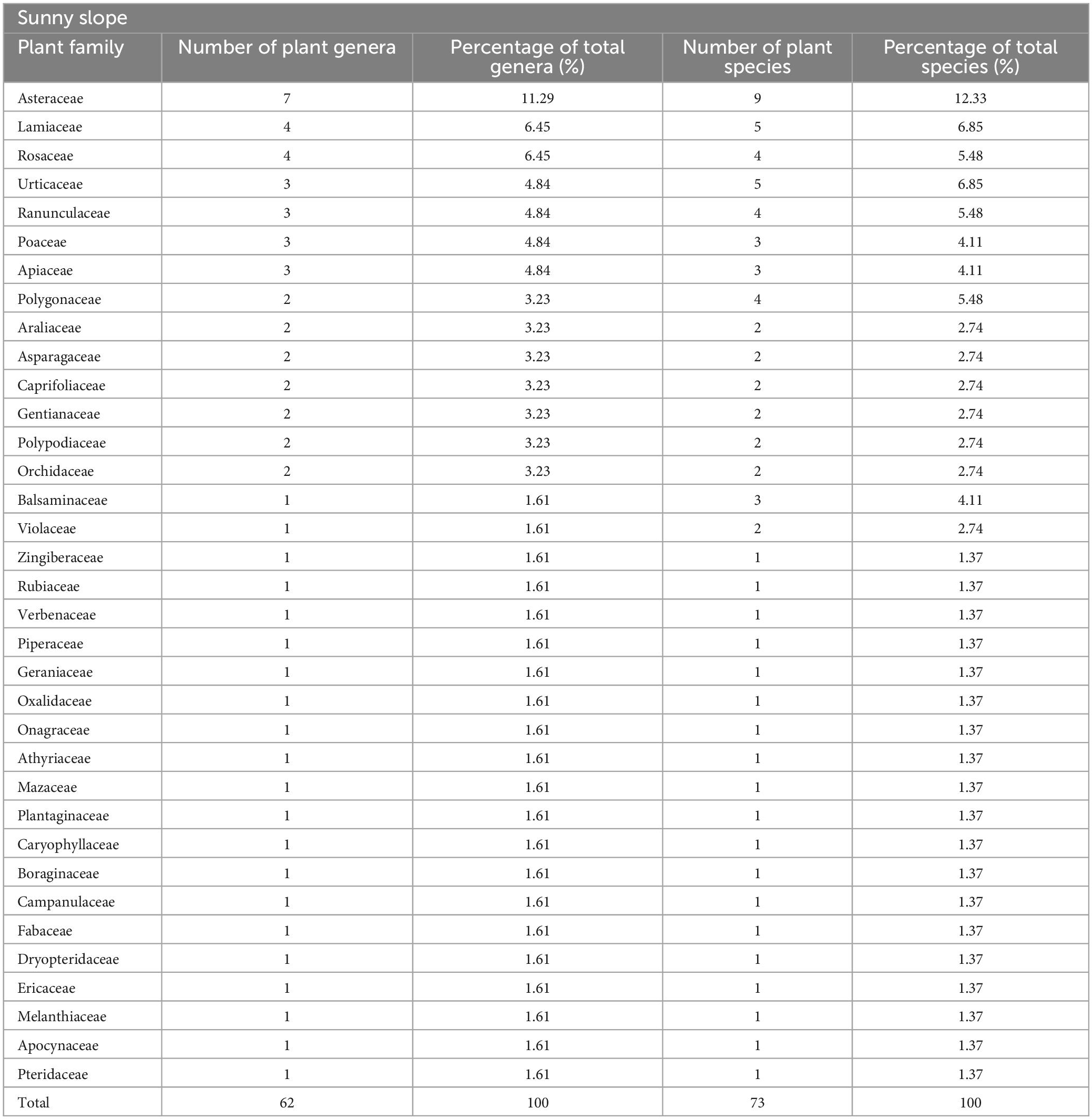
Table 3. Composition of herbaceous plants in sunny slope.
It can be seen from Figures 2, 3 that the species composition of herbaceous plants on shady and sunny slopes varies greatly at different altitudes. In the shady slope of the study area, the number of herbaceous species decreased first and then increased with the increase of altitude. Herbal species with higher important values in each altitude gradient zone include Pilea insolens, Duchesnea indica and Fragaria moupinensis; The most dominant is that the important value of Pilea insolens decreases with the increase of altitude. The important value of Duchesnea indica and Fragaria moupinensis increased with the elevation. In the sunny slope of the study area, the number of herbaceous species first increased and then decreased with the increase of altitude. In each elevation gradient zone, the importance value of herbaceous plants decreases with the increase of herbaceous species; On the whole, the higher herbaceous plant species on the sunny slope are Fragaria moupinensis, Peperomia tetraphylla and Duchesnea indica.
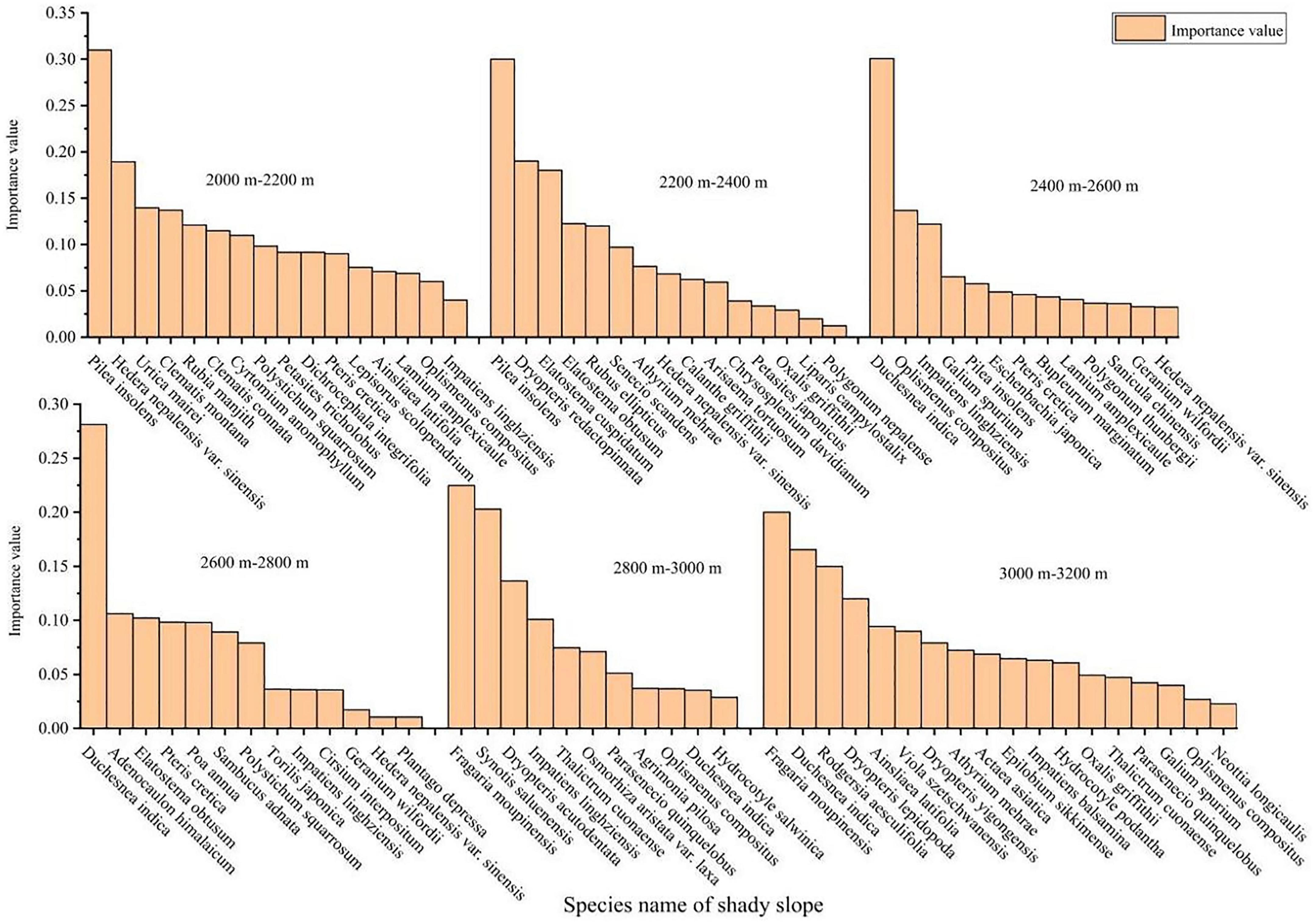
Figure 2. Important values of herbaceous plants in different altitude gradients of shady slope.
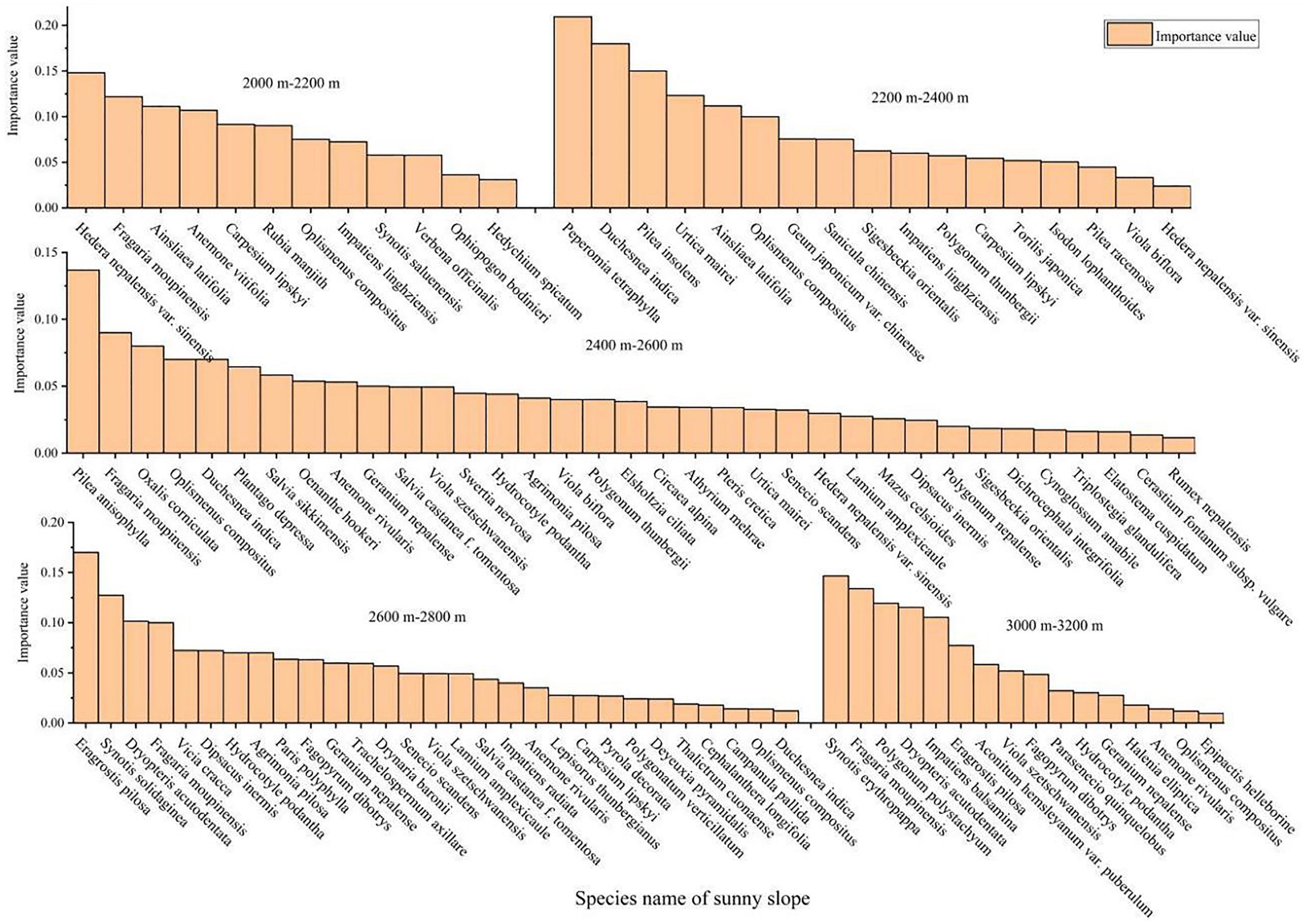
Figure 3. Important values of herbaceous plants in different altitude gradients of sunny slope.
It can be seen from Figures 4A, B that the species richness index (R), Shannon-Wiener diversity index (H) and Pielou evenness index (J) of herbaceous plants on shady slopes show a trend of first decreasing and then increasing with the increase of altitude. Shannon-Wiener diversity index (H) and Pielou evenness index (J) showed no significant difference among different altitudes. It can be seen from Figures 5A, B that the species richness index (R) of herbaceous plants on the sunny slope increases first and then decreases with the increasing altitude. Shannon-Wiener diversity index (H) showed a trend of first decreasing and then increasing. Shannon-Wiener diversity index (H) was significantly higher than the altitude gradient of 2,400–2,600 m, 2,600–2,800 m and 3,000–3,200 m at 2,000–2,200 m and 2,200–2,400 m (P < 0.05). The Pielou evenness index (J) decreases first and then increases with the increase of altitude. The Pielou evenness index (J) at the altitude gradient of 2,000–2,200 m and 2,200–2,400 m was significantly higher than the altitude gradient of 2,400–2,600 m, 2,600–2,800 m and 3,000–3,200 m (P < 0.05), and there was no significant difference between 2,600–2,800 m and 3,000–3,200 m (P > 0.05).
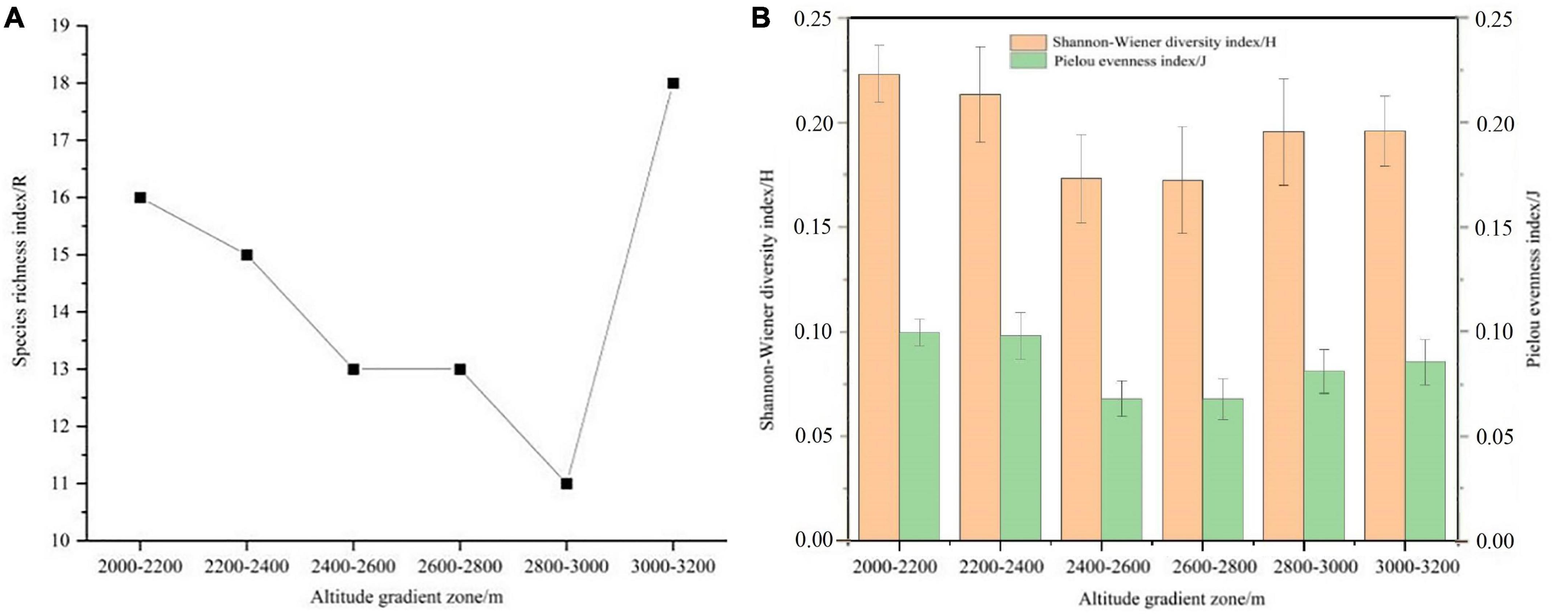
Figure 4. α diversity index of herbaceous plants in different altitude gradients of shady slope. (A) Species richness index, and (B) Shannon-Wiener diversity index and Pielou evenness index.
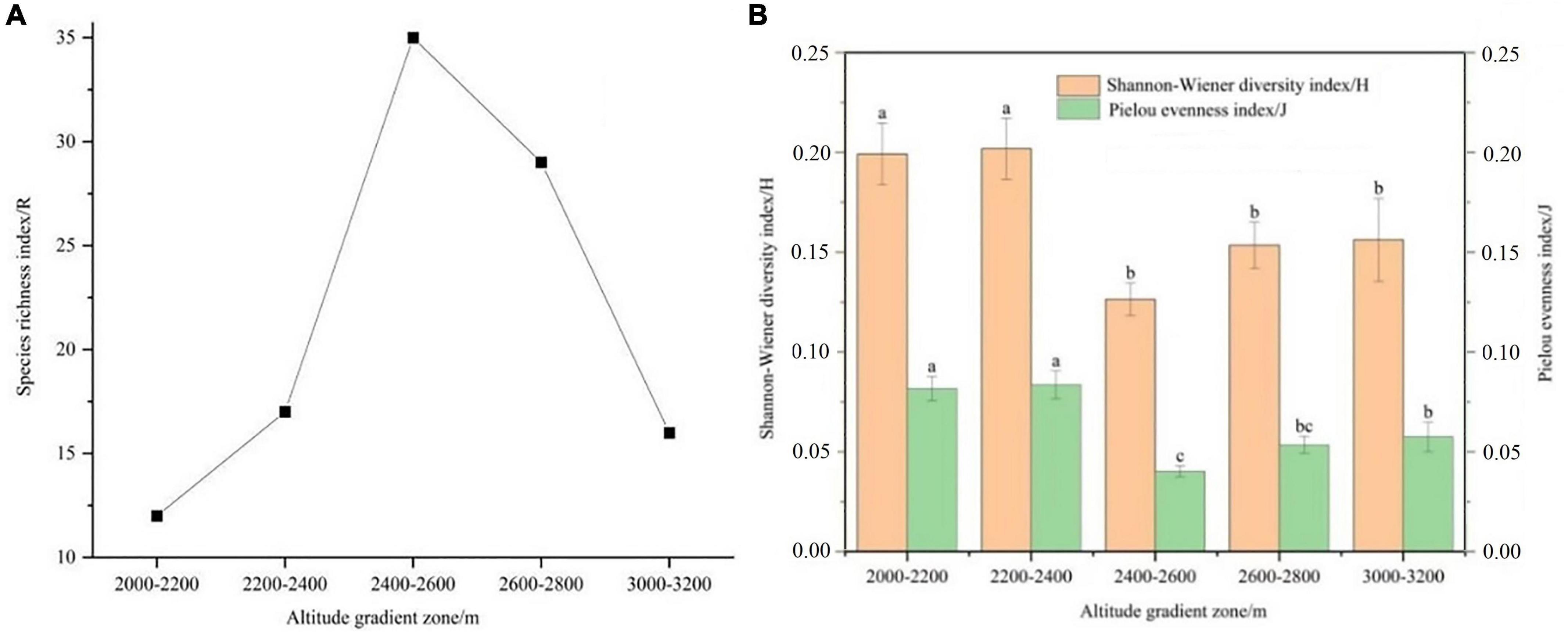
Figure 5. α diversity index of herbaceous plants in different altitude gradients of sunny slope. (A) Species richness index, and (B) Shannon-Wiener diversity index and Pielou evenness index. Different lower case letters in the figure indicate that there are significant differences between different altitude gradients (P < 0.05).
According to Figures 6A, C, Sorenson similarity index (SI) of herbaceous plants in different altitude gradient zones on the shady slope is between 0.129 and 0.385. Cody anisotropy index(βc) is between 8 and 13.5. Sorenson similarity index (SI) is the highest between 2,400–2,600 m and 2,600–2,800 m of the altitude gradient zone, indicating that there are many common species between the two altitude gradient zones. Cody anisotropy index(βc) is the lowest, indicating that the replacement rate of herb species between the two altitude gradient zones is relatively slow. Sorenson Similarity Index (SI) is the lowest between 2,000–2,200 m and 2,200–2,400 m elevation gradient zones, indicating that there are fewer species in common between these two elevation gradient zones. Cody anisotropy index(βc) is the highest, indicating that the area between the two altitude gradient zones is rich in herb species on the shady slope. It can be seen from Figures 6B, D that Sorenson similarity index (SI) of herbaceous plants in different altitude gradient zones on the sunny slope is between 0.269 and 0.690, and Cody dissimilarity index(βc) is between 9.5 and 20. Sorenson similarity index (SI) is the highest between 2,000–2,200 m and 2,200–2,400 m in the altitude gradient zone, indicating that there are many common species between the two altitude gradient zones. Cody anisotropy index(βc) is the lowest, indicating that the replacement rate of herb species between the two altitude gradient zones is relatively slow. Sorenson Similarity Index (SI) between 2,200–2,400 m and 2,400–2,600 m altitude gradient zones is the lowest, and there are few common species between the two altitude gradient zones.
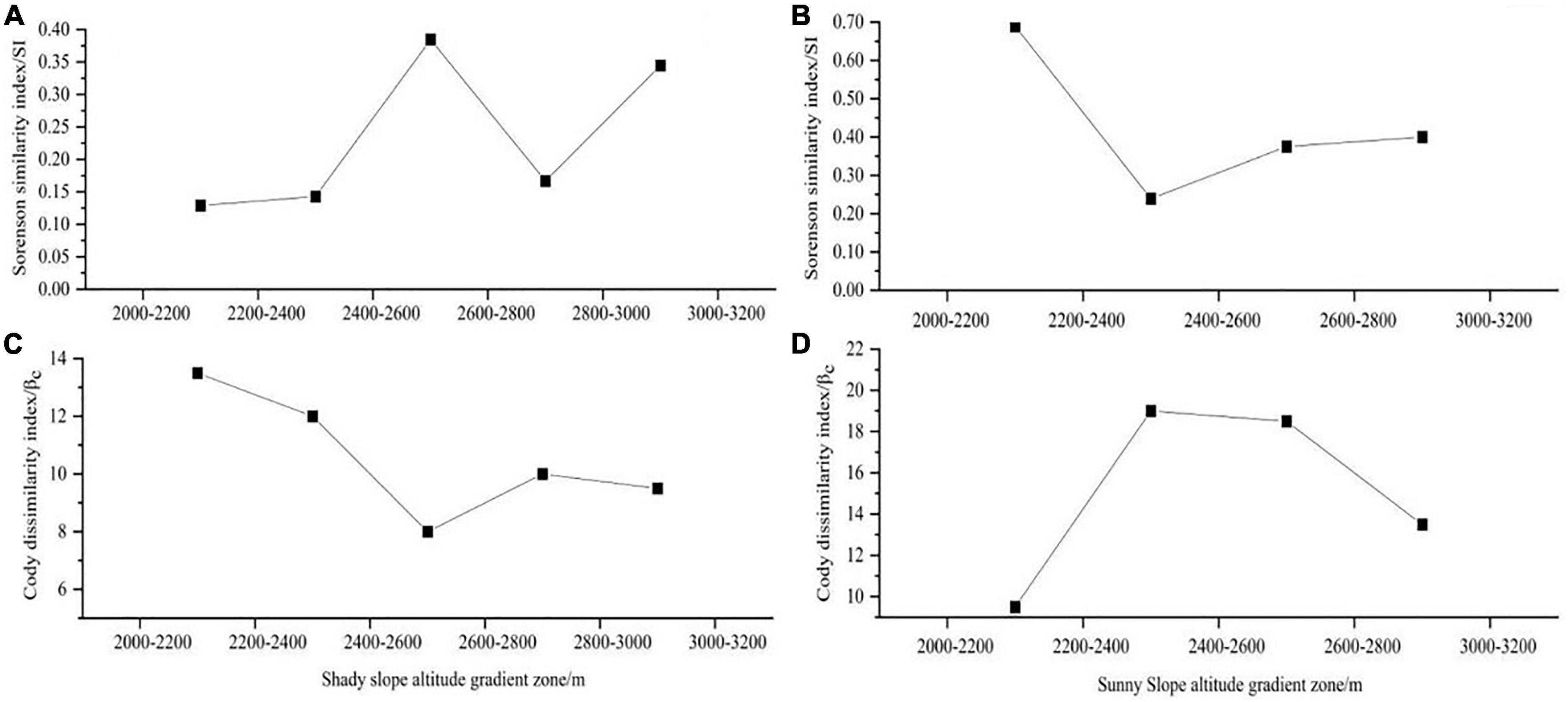
Figure 6. β diversity index of herbaceous plants in different altitude gradients. Panels (A,C) are Sorenson similarity index and Cody dissimilarity index on shady slope, and panels (B,D) are Sorenson similarity index and Cody dissimilarity index on sunny slope.
The results of principal component analysis (Figure 7A) show that different environmental factors have different loads in different principal components. Clay content, silt content, elevation, slope and other factors are positively correlated with the first principal component. Clay content, silt content, slope, PH value are positively correlated with the second principal component. Only the loadings of clay content, silt content and slope in the two principal components are positive, while the loadings of vegetation coverage and sand content in the two principal components are negative, indicating that different environmental factors have different effects on the herb diversity indicators. The grain size composition of soil determines its texture. Soil texture directly affects the maintenance and movement of soil moisture, soil fertility, soil aeration, soil temperature, and is closely related to crop growth and development. Vegetation coverage can reflect the quality of local habitats and is the result of the comprehensive effect of various environmental factors and plants. The correlation between environmental factors and diversity indicators (Figure 7B) shows that there is a certain degree of clustering among samples. The environmental factors of shady slope and sunny slope are separated. The sample plot on the shady slope has a higher altitude, with a higher content of clay and silt in the soil. The sunny slope sample plot has higher vegetation coverage and slope. The Shannon-Wiener diversity index (H) and evenness index (J) of herbaceous plant diversity showed significant positive correlation. Shannon-Wiener diversity index (H) and evenness index (J) were negatively correlated with species richness (R).
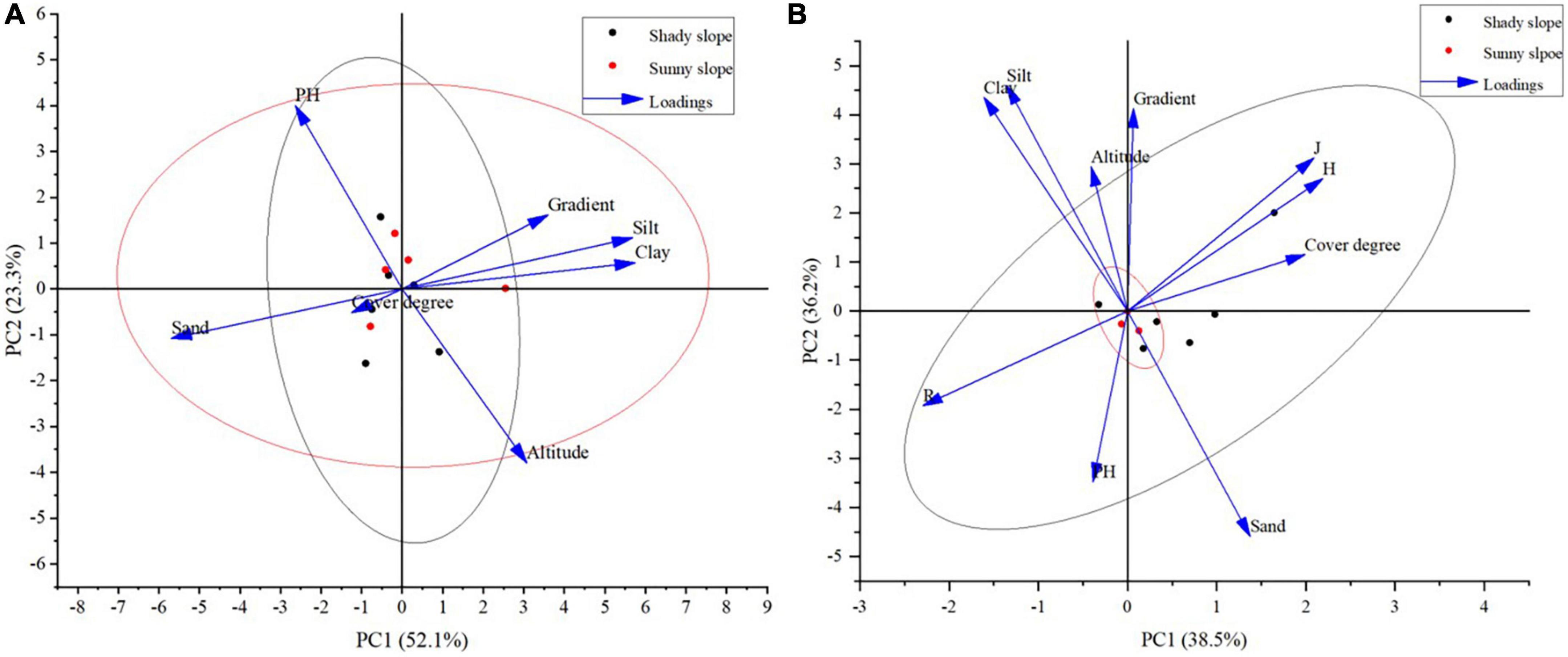
Figure 7. Principal component analysis of different environmental factors and herb diversity index. (A) Principal component analysis of environmental factors. (B) Correlation between environmental factors and diversity indices.
Herbs are an important part of the biodiversity of the forest ecosystem. Their species composition and structure are also important indicators of the natural recovery of forests and the healthy structure of the forest ecosystem (Chitale et al., 2014; Wani et al., 2022). Altitude has always been considered as the dominant factor affecting regional habitat differences. Different altitudinal gradients lead to different spatial distributions of hydrothermal conditions, thus forming different herbage community structures in different regions (Cuesta et al., 2016; Zhao et al., 2016; Borisade, 2020; Pérez-Postigo et al., 2021).
According to the survey results in the Sejila Mountain research area, there are relatively many single family (genus) plant species in the undergrowth herb diversity in southeast Tibet, and most families only include one genus and one species (Wang et al., 2021). The number of families, genera and species of herbaceous plants on sunny slope is greater than that on shady slope, which is caused by the difference of surface soil temperature between shady slope and sunny slope. Slope aspect plays a leading role in the distribution of herbaceous plants and the change of soil temperature, indicating that the distribution of mountain plants is greatly affected by the change of soil temperature. The soil temperature on the sunny slope is higher, which is more suitable for the growth of a variety of herbs (Zhao et al., 2016; Xue et al., 2018; Kwaku, 2021). Asteraceae species on shady slope and sunny slope are the largest, and the number of Asteraceae species on shady slope accounts for 17.54% of the total herbaceous species investigated on shady slope of Sejila Mountain. The number of Asteraceae species on the sunny slope accounts for 12.33% of the total herbaceous species investigated on the sunny slope of Sejila Mountain, which is related to the habitat heterogeneity under different altitude gradients in the study area. Habitat heterogeneity makes different herbs occupy different niches (Xu et al., 2011).
In the α diversity index, Shannon-Wiener diversity index (H) and Pielou evenness index (J) of shady slope and sunny slope decreased first and then increased with the increasing altitude (Zhang et al., 2016). The main reason is that due to the uplift of mountains, the precipitation will increase with the elevation and then decrease to a certain extent (Brien et al., 2000; Ent et al., 2016; Tesfaye et al., 2016). The soil water content is high between 2,400–2,600 m above sea level, and the good habitat conditions meet the growth of trees and shrubs. This leads to the occupation of habitat conditions by trees and shrubs in this altitude gradient zone, squeezing the growth of herbaceous plants. Therefore, the Shannon Wiener diversity index (H) and Pielou evenness index (J) of herbaceous plants in this altitude gradient zone are the lowest. The number of herbaceous species is the largest in the area of 2,600–2,800 m above sea level on the sunny slope. This area has the lowest vegetation coverage, which is caused by more solar radiation reaching the ground through alpine pine forests (Kleinn et al., 2020). Therefore, in the same forest coverage area, the smaller the vegetation coverage, the more species of herbaceous plants. However, relevant scholars pointed out that the family and genus composition of undergrowth herbaceous vegetation species is complex, the community species diversity level is high, the evenness is large, and the habitat conditions are stable. The species diversity shows a different trend of increasing first and then decreasing with the increase of altitude gradient (Austrheim, 2002; Gómez-Díaz et al., 2017; Dip et al., 2020).
In the β diversity index, the Cody dissimilarity index(βc) of shady slope and sunny slope generally shows a downward trend with the increase of altitude, while Sorenson similarity index (SI) is opposite. Cody anisotropy index(βc) of shady slope and sunny slope generally shows a downward trend with the increase of altitude, and the substitution of herbaceous plants in different habitats is generally slow (Sánchez-gonzález and López-mata, 2005). This is related to the competition among species, resulting in the heterogeneity of habitats. Sorenson similarity index (SI) of herbaceous plants in different altitude gradient zones is generally higher on sunny slopes than on shady slopes, indicating that there are more common species on sunny slopes than on shady slopes among different altitude gradient zones. Cody anisotropy index(βc) is generally higher on sunny slope than on shady slope, and the replacement rate of sunny slope is slower than that of shady slope.
The human disturbance, grazing pressure, and land use change is important in shaping and driving herbaceous plant communities. The Sejila Mountain on the Qinghai-Tibet Plateau is a region with dense original vegetation and steep mountain terrain. The human disturbance intensity and grazing pressure have little influence on the growth of herbaceous plants, and the change of land use under the original forest area is small (Xue et al., 2018; Zu et al., 2019; Han et al., 2022). Topography and local habitats are obviously different in the alpine environment. Among the driving factors affecting herb diversity, the topography factors including the slope, slope direction and altitude have a great impact on vegetation growth and spatial distribution (Davies et al., 2007). In this study, the elevation gradient and slope aspect have significant effects on the diversity index, which is consistent with the research findings of Korner (2001), Liao et al. (2008), and Sherman et al. (2008). Moreover, the physical and chemical properties of soil also have an important impact on the distribution of plant communities. The previous studies have shown that the soil factors strongly affect the species composition of alpine vegetation (Yan, 2007). This study also found that the soil clay content and silt content are key factors affecting the diversity of herbaceous plants. Under the influence of micro-climate and micro-topography in different regions, the soil texture also changes correspondingly. The influence of soil particle size on herb species diversity in Sejira Mountain shows a remarkable change.
In the future research on the vertical distribution pattern of herbaceous plants in high altitude mountain areas, we should expand to multiple biological groups, and deeply study the interaction between various groups (such as the influence of some microorganisms on herbaceous plant roots, the influence of insects on herbaceous plant pollination, etc.). This will greatly improve our understanding of the vertical distribution pattern of biodiversity and its driving factors. In addition, the study relies on a limited number of samples, which may not fully reflect the herbaceous plant communities in the alpine regions. Due to the influence of alpine geographical environment, it is difficult to reach the sufficient number of samples. This partly affects the reliability of conclusions. Further studies will provide a more accurate picture of the species composition and diversity of herbaceous plants using a larger number of samples and a more systematic sampling design in high-altitude mountain areas.
The types of herb communities in the study area are diverse, and the species diversity is high. A total of 57 species of herbs belonging to 49 genera and 24 families were found on the shady slope. A total of 73 species belonging to 62 genera and 35 families were found on the sunny slope. Asteraceae contains the most species; α Among the diversity indices, the richness index (R) of shady slope, Shannon-Wiener diversity index (H) and Pielou evenness index (J) all decreased first and then increased with the increasing altitude. The richness index (R) of the sunny slope increased first and then decreased with the elevation. Shannon-Wiener diversity index (H) and Pielou evenness index (J) both decreased first and then increased with the increasing altitude; In the β diversity index, Cody dissimilarity index (βc) of shady slope and sunny slope generally showed a downward trend, while Sorenson similarity index (SI) was opposite; Altitude gradient and aspect have significant effects on diversity index. The diversity of herbaceous plants is affected by many factors. The difference of water and heat conditions caused by altitude and slope aspect is an important driving factor for the diversity of herbaceous plants. Moreover, the soil clay content, silt content and slope gradient are also important driving factors. The correlation between environmental factors and diversity index shows that there is a certain degree of clustering among samples. The driving environmental factors of shady and sunny slope are separated due to the different degrees of influence on alpine herbaceous plant diversity index. Therefore, the differential measure is necessary in the rational utilization of understory plant resources and ecological environment protection of the shady and sunny slopes in high-altitude mountain regions.
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
JH and HY: conceptualization. BY and SW: data curation. JX: formal analysis, supervision, and writing—review and editing. ZZ: methodology. XC: resources. JH and JC: validation. JH and ZZ: writing—original draft. All authors read and agreed to the published version of the manuscript.
This work was financially supported by Investigation on resource environment and biodiversity of typical mountainous areas in different climatic zones (2019FY101601-2), the original innovation project of the basic frontier scientific research program, Chinese Academy of Sciences (ZDBS-LY-DQC031), Ecological cultivation demonstration of Gentian at the Himalayan headwater of Chcedo, the National Natural Science Foundation of China (42071259), the Third Batch of Tianshan Talents Program of Xinjiang Uygur Autonomous Region (2021–2023), the Youth Innovation Promotion Association of the Chinese Academy of Sciences (2019430), the study on the support system of modern forestry technology in Tibet (2018xz503118002), and the study on the model of planting and breeding circular agriculture in Tibet.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Andra, T. (2016). Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in hungary. Plant Ecol. 217, 549–556.
Austrheim, G. (2002). Plant diversity patterns in semi-natural grasslands along an elevational gradient in southern Norway. Plant Ecol. 161, 193–206. doi: 10.1023/A:1020315718720
Borisade, T. V. (2020). Nutrient status in herbaceous layer of riparian forests in Southwestern. Trop. Ecol. 61, 589–593.
Brien, E. M. O., Sciences, F., Wales, U., Road, D., Uw, L. L., Field, R., et al. (2000). Climatic gradients in woody plant (tree and shrub) diversity?: water-energy dynamics, residual variation, and topography. Oikos 89, 588–600.
Brockerhoff, E. G., Barbaro, L., Castagneyrol, B., Forrester, D. I., Gardiner, B., González-Olabarria, J. R., et al. (2017). Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodiversity Conserv. 26, 3005–3035. doi: 10.1007/s10531-017-1453-2
Butchart, S. H., Walpole, M., Collen, B., van Strien, A., Scharlermann, J. P., and Almond, R. E. (2010). Global biodiversity: indicators of recent declines. Science 328, 1164–1168. doi: 10.1126/science.1187512
Cardinale, B. J., Duffy, J. E., Gonzalez, A., Hooper, D. U., Perrings, C., and Venail, P. (2012). Biodiversity loss and its impact on humanity. Nature 486, 59–67. doi: 10.1038/nature11148
Chitale, V. S., Behera, M. D., and Roy, P. S. (2014). Future of endemic flora of biodiversity hotspots in India. PLoS One 9:e115264. doi: 10.1371/journal.pone.0115264
Cuesta, F., Muriel, P., Llambí, L. D., Halloy, S., Aguirre, N., Beck, S., et al. (2016). Accepted article. Ecography 40, 1381–1394. doi: 10.1111/ecog.02567
da Silva, F. K. G., de Faria Lopes, S., Lopez, L. C. S., de Melo, J. I. M., and de Brito Melo Trovão, D. M. (2014). Patterns of species richness and conservation in the Caatinga along elevational gradients in a semiarid ecosystem. J. Arid Environ. 110, 47–52. doi: 10.1016/j.jaridenv.2014.05.011
Davies, K. W., Bates, J. D., and Miller, R. F. (2007). Environmental and vegetation relationships of the Artemisia tridentata spp. wyomingensis alliance. J. Arid Environ. 70, 478–494. doi: 10.1016/j.jaridenv.2007.01.010
Dip, A. B., Sampietro-Vattuone, M. M., Garey, M. V., and Eleuterio, A. A. (2020). Altitudinal gradient of plant diversity patterns in the sierra de quilmes (monte desert – Argentina). J. Arid Environmen. 182:104274. doi: 10.1016/j.jaridenv.2020.104274
Doležal, J., and Šrůtek, M. (2002). Altitudinal changes in composition and structure of mountain-temperate vegetation: a case study from the Western Carpathians. Plant Ecol. 158, 201–221. doi: 10.1023/A:1015564303206
Ecology, S. P., and Kessler, M. (2012). Elevational gradients in species richness and endemism of selected plant groups in the central bolivian andes author (s): Michael Kessler reviewed work (s): elevational gradients in species richness and endemism groups in the central Bolivian Andes of. Plant Ecol. 149, 181–193.
Ellum, D. S., Ashton, M. S., and Siccama, T. G. (2010). Spatial pattern in herb diversity and abundance of second growth mixed deciduous-evergreen forest of southern New England. U S A. For. Ecol. Manag. 259, 1416–1426. doi: 10.1016/j.foreco.2010.01.011
Ent, A., Van Der Erskine, P., Mulligan, D., Repin, R., and Karim, R. (2016). Vegetation on ultramafic edaphic ‘ islands ’ in Kinabalu park (Sabah, Malaysia) in relation to soil chemistry and elevation. Plant Soil 403, 77–101.
Fayiah, M., Dong, S., Li, Y., Xu, Y., Gao, X., Li, S., et al. (2019). The relationships between plant diversity, plant cover, plant biomass and soil fertility vary with grassland type on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 286:106659. doi: 10.1007/s11104-016-2831-3
Gilliam, F. S. (2007). The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience 57, 845–858. doi: 10.1890/12-1582.1
Gómez-Díaz, J. A., Krömer, T., Kreft, H., Gerold, G., Carvajal-Hernández, C. I., and Heitkamp, F. (2017). Diversity and composition of herbaceous angiosperms along gradients of elevation and forest-use intensity. PLoS One 12:e0182893. doi: 10.1371/journal.pone.0182893
Han, W., Chen, L., Su, X., Liu, D., Jin, T., Shi, S., et al. (2022). Effects of soil physico-chemical properties on plant species diversity along an elevation gradient over alpine grassland on the Qinghai-Tibetan Plateau, China. Front. Plant Sci. 13:822268. doi: 10.3389/fpls.2022.822268
Kleinn, C., Magnussen, S., Nölke, N., Magdon, P., and Álvarez-gonzález, J. G. (2020). Improving precision of field inventory estimation of aboveground biomass through an alternative view on plot biomass. For. Ecosyst. 7:57. doi: 10.1186/s40663-020-00268-7
Korner, C. (2001). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin: Springer.
Kwaku, E. A. (2021). Biomass and species diversity of different alpine plant. Plants 10, 1–11. doi: 10.3390/plants10122719
Li, X., Liang, E., Gričar, J., Prislan, P., Rossi, S., and Čufar, K. (2013). Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol. 33, 48–56. doi: 10.1093/treephys/tps113
Liao, B. H., Nan, H. U., and Ding, S. Y. (2008). Dynamics of plant functional groups along altitudinal gradient on the northern slope of Funiu mountain. J. Henan Univ. 38, 618–632.
Mu, C., Li, L., Zhang, F., Li, Y., Xiao, X., Zhao, Q., et al. (2018). Impacts of permafrost on above- and belowground biomass on the northern Qinghai-Tibetan Plateau. Arctic Antarctic Alpine Res. 50:e1447192. doi: 10.1080/15230430.2018.1447192
Pérez-Postigo, I., Bendix, J., Vibrans, H., and Cuevas-Guzmán, R. (2021). Diversity of alien roadside herbs along an elevational gradient in western Mexico. NeoBiota 65, 71–91. doi: 10.3897/neobiota.65.67192
Sánchez-gonzález, A., and López-mata, L. (2005). Plant species richness and diversity along an altitudinal gradient in the Sierra. Divers. Distrib. 11, 567–575. doi: 10.1111/j.1366-9516.2005.00186.x
She, Y., Zhang, Z., Ma, L., Xu, W., Huang, X., and Zhou, H. (2022). Vegetation attributes and soil properties of alpine grassland in different degradation stages on the Qinghai-Tibet Plateau, China: a meta-analysis. Arabian J. Geosci. 15, 15–193. doi: 10.1007/s12517-021-09400-5
Sherman, R., Mullen, R., and Haomin, L. (2008). Spatial patterns of plant diversity and communities in alpine ecosystems of the Hengduan mountains, Northwest Yunnan, China. J. Plant Ecol. 1, 117–136. doi: 10.1016/j.pld.2021.09.001
Shi, S., Liu, G., Li, Z., and Ye, X. (2021). Elevation-dependent growth trends of forests as affected by climate warming in the southeastern Tibetan Plateau. For. Ecol. Manag. 498:119551. doi: 10.1016/j.foreco.2021.119551
Sklenář, P., and Jørgensen, P. M. (1999). Distribution patterns of paramo plants in ecuador. J. Biogeogr. 26, 681–691. doi: 10.3390/insects11010064
Sklenár, P., and Ramsay, P. M. (2001). Diversity of zonal páramo plant communities in ecuador. Divers. Distrib. 7, 113–124. doi: 10.1046/j.1472-4642.2001.00101.x
Tesfaye, M. A., Bravo, F., Ruiz-peinado, R., Pando, V., and Bravo-oviedo, A. (2016). Geoderma impact of changes in land use, species and elevation on soil organic carbon and total nitrogen in Ethiopian central highlands. Geoderma 261, 70–79. doi: 10.1016/j.geoderma.2015.06.022
Torresani, M., Rocchini, D., Sonnenschein, R., Zebisch, M., Hauffe, H. C., Heym, M., et al. (2020). Height variation hypothesis: a new approach for estimating forest species diversity with CHM LiDAR data. Ecol. Indic. 117:106520. doi: 10.1016/j.ecolind.2020.106520
Van Dam, A. (2019). Diversity and its decomposition into variety, balance and disparity. R. Soc. Open Sci. 6:190452. doi: 10.1098/rsos.190452
Walter, J. A., Stovall, A. E. L., and Atkins, J. W. (2021). Vegetation structural complexity and biodiversity in the Great Smoky mountains. Ecosphere 12:e03390. doi: 10.1002/ecs2.3390
Wang, G., Sun, Y., Zhou, M., Guan, N., Wang, Y., Jiang, R., et al. (2021). Effect of thinning intensity on understory herbaceous diversity and biomass in mixed coniferous and broad-leaved forests of Changbai mountain. For. Ecosyst. 8:53. doi: 10.1186/s40663-021-00331-x
Wang, J., Wang, G., Fu, Y., Chen, X., and Song, X. (2019). Short-term effects of nitrogen deposition on soil respiration components in two alpine coniferous forests, southeastern Tibetan Plateau. J. For. Res. 30, 1029–1041. doi: 10.1007/s11676-018-0678-6
Wani, S. A., Ahmad, R., Gulzar, R., Rashid, I., Malik, A. H., and Khuroo, A. A. (2022). Diversity, distribution and drivers of alien flora in the Indian Himalayan region. Global Ecol. Conserv. 38:e02246. doi: 10.1016/j.gecco.2022.e02246
Xian, H., Hao, Y., Lv, J., Wang, C., Zuo, P., Pei, Z., et al. (2021). Novel brominated flame retardants (NBFRs) in soil and moss in Mt. Shergyla, southeast Tibetan Plateau: occurrence, distribution and influencing factors. Environ. Pollut. 291:118252. doi: 10.1016/j.envpol.2021.118252
Xiangyang, S., Genxu, W., Juying, S., Shouqin, S., Zhaoyong, H., Chunlin, S., et al. (2022). Contrasting water sources used by a coniferous forest in the high-altitude, southeastern Tibetan Plateau. Sci. Total Environ. 849:157913. doi: 10.1016/j.scitotenv.2022.157913
Xu, X., Ouyang, H., Cao, G., Richter, A., Wanek, W., and Kuzyakov, Y. (2011). Dominant plant species shift their nitrogen uptake patterns in response to nutrient enrichment caused by a fungal fairy in an alpine meadow. Plant Soil 341, 495–504. doi: 10.1007/s11104-010-0662-1
Xue, R., Yang, Q., Miao, F., Wang, X., and Shen, Y. (2018). Slope aspect influences plant biomass, soil properties and microbial composition in alpine meadow on the Qinghai-Tibetan Plateau. J. Soil Sci. Plant Nutr. 18, 1–12. doi: 10.4067/S0718-95162018005000101
Yan, W. (2007). The impact of snow-cover on alpine vegetation type of different aspects in the west of Sichuan province. Acta Ecol. Sin. 27, 5120–5129.
Yılmaz, O. Y., Yılmaz, H., and Akyüz, Y. F. (2018). Effects of the overstory on the diversity of the herb and shrub layers of Anatolian black pine forests. Eur. J. For. Res. 137, 433–445. doi: 10.1007/s10342-018-1114-3
Yin, H., Yu, B., Zhang, Z., Jia, L., Xue, J., Chen, X., et al. (2022). Evaluating the impact of different afforestation time scales on the soil quality in a typical watershed valley using the criteria importance through intercriteria correlation method and gray target model. Front. For. Glob. Change 5:997949. doi: 10.3389/ffgc.2022.997949
Zhang, W., Huang, D., Wang, R., Liu, J., and Du, N. (2016). Altitudinal patterns of species diversity and phylogenetic diversity across temperate Mountain forests of Northern China. PLoS One 11:e159995. doi: 10.1371/journal.pone.0159995
Zhang, Z. W., Yin, H. Y., Chang, J. J., and Jie, X. (2022). Spatial variability of surface soil water content and its influencing factors on shady and sunny slopes of an alpine meadow on the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 34:e02035. doi: 10.1016/j.gecco.2022.e02035
Zhang, Z. W., Yin, H. Y., Zhao, Y., Wang, S. P., Han, J. H., Yu, B., et al. (2021). Spatial heterogeneity and driving factors of soil moisture in alpine desert using the geographical detector method. Water 13:2652. doi: 10.3390/w13192652
Zhao, J., Li, X., Li, R., Tian, L., and Zhang, T. (2016). Effect of grazing exclusion on ecosystem respiration among three different alpine grasslands on the central Tibetan Plateau. Ecol. Eng. 94, 599–607. doi: 10.1016/j.ecoleng.2016.06.112
Zhou, X. L., Yan, Y. E., Zhang, J., Zhou, X. J., Yan, Y. Q., Yang, F. Q., et al. (2022). Vegetation community structure and diversity in a burned area of Picea asperata-Abies fabri forest on different aspects on the northeastern margin of the Qinghai-Tibetan Plateau. Acta Prataculturae Sinica 31, 144–155. doi: 10.11686/cyxb2021372
Keywords: herbs, species diversity, altitude gradient, driving factors, Qinghai-Tibet Plateau
Citation: Han J, Yin H, Xue J, Zhang Z, Xing Z, Wang S, Chang J, Chen X and Yu B (2023) Vertical distribution differences of the understory herbs and their driving factors on shady and sunny slopes in high altitude mountainous areas. Front. For. Glob. Change 6:1138317. doi: 10.3389/ffgc.2023.1138317
Received: 05 January 2023; Accepted: 06 March 2023;
Published: 23 March 2023.
Edited by:
Zishan Ahmad Wani, Baba Ghulam Shah Badshah University, IndiaReviewed by:
Zeeshan Ahmad, Quaid-i-Azam University, PakistanCopyright © 2023 Han, Yin, Xue, Zhang, Xing, Wang, Chang, Chen and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jie Xue, eHVlamllMTFAbXMueGpiLmFjLmNu; Zhiwei Zhang, YWl3b3dlaWdlQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.