 Juan Liu
Juan Liu Rui Li1
Rui Li1 Songze Wan
Songze Wan
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 02 March 2023
Sec. Planted Forests
Volume 6 - 2023 | https://doi.org/10.3389/ffgc.2023.1136474
This article is part of the Research Topic Carbon Sequestration in Forest Plantation Ecosystems View all 6 articles
Objective: Lime application and understory plant clearance are two common forest management methods employed to increase forest productivity in subtropical regions. However, the effect of these two management methods, or their combined application, on soil respiration in forest plantation settings is still not fully understood.
Methods: Here, a field experiment was conducted to determine how lime application, understory clearance, and their combined application, would impact total soil respiration (Rs), autotrophic respiration (Ra), and heterotrophic respiration (Rh) in a Chinese subtropical Eucalyptus plantation. Changes in soil microclimate and microbial community (PLFAs), as well as Eucalyptus fine root biomass were also assessed.
Results: Lime application stimulated Rh, but decreased Ra of understory vegetation (primarily Dicranopteris dichotoma), thus consequently inhibiting Rs. Understory clearance also reduced Rs, primarily due to reductions in soil PLFAs and associated Rh, and Ra of understory vegetation. Since the increase in Rh induced by lime application was greater than the decrease in Rh caused by understory clearance, the combined application of lime application and understory clearance decreased Rs primarily by reducing understory root respiration.
Conclusion: Our observations suggest that understory plants can modulate the effect of lime application on Rs, highlighting the important role of understory vegetation in regulating soil carbon cycling in subtropical Eucalyptus plantations. We concluded that the potential interactive effects should be considered in developing management practices that optimize the yield and sustainability of subtropical plantations.
Soil respiration (Rs) is the second largest carbon flux between land and the atmosphere (Yang et al., 2022). In terrestrial ecosystems, forests are important regulators of the global carbon balance, with trees storing up to 56% of both above- and below-ground terrestrial carbon, and forest soil respiration accounting for at least 30% of total terrestrial respiration (Wan et al., 2015; Yang et al., 2022). Therefore, even minor alternations in forest soil respiration can significantly change atmospheric carbon dioxide concentration and the global carbon cycle (Valente et al., 2021). Forest soil respiration is affected by forest management practices such as thinning (Sherman and Coleman, 2020), fertilization (Wang et al., 2017), understory removal (Yao et al., 2019), and litter manipulation (Dai et al., 2021). However, since the two main components of Rs: autotrophic respiration (Ra: from live roots and rhizosphere), and heterotrophic respiration (Rh: from microbial decomposition of soil organic matter) are site-specifically related to forest type, soil characteristics, local climate, and other variables, the impact of forest management practices on actual soil respiration are often variable (Mo et al., 2008; Wan et al., 2015). For example, Zhang et al. (2022) explored that thinning increased Rs in subtropical Cunninghamia lanceolata plantations. Nonetheless, Pang et al. (2013) found that thinning inhibited Rs in a pine plantation in the eastern Tibetan Plateau. Furthermore, multiple silvicultural practices may be applied simultaneously, potentially producing interactive effects on soil respiration (Sherman and Coleman, 2020). Therefore, a comprehensive assessment of the impact of forest management measures on soil respiration will not only help to mitigate climate warming, but also provide a scientific basis for formulating more rational forest management strategies.
Lime application is a common forestry practice to improve acrisols in humid tropical and subtropical regions (Zhao et al., 2015). In addition to preventing or reversing soil acidification, the lime application can profoundly alter soil microbial communities (Sridhar et al., 2022), although the reported results concerning the effect of lime application on soil respiration have so far been mixed. Shah et al. (1990) and Goupil (2014) reported that lime application increased Rh, likely because of enhanced acid intolerant bacterial abundance and fungal abundance, and consequently increased soil respiration. On the contrary, Melvin et al. (2013) reported a reduction in soil respiration by inhibiting soil basal respiration in a broadleaved forest after the application of lime, thereby increasing carbon storage in the forest soil. Meanwhile, Lochon et al. (2019) found that lime application produced a negligible effect on soil respiration because the increase in Rh was counteracted by the decrease in Ra. Taken together, it appears that responses of soil respiration to liming are highly dependent on the environment and the balance between Ra and Rh. High soil pH tends to favor the growth of acid-intolerant bacteria at the expense of both acid-tolerant bacteria and fungi (Wan et al., 2015), thus alternations of microbial communities in limed soils may at least partially explain changes in soil respiration. Meanwhile, the lime application can increase the availability of some nutrients (i.e., Ca and Mn), while decreasing the availability of some micro nutrients, such as Cu and Zn (Florentino et al., 2021). Therefore, alternations in plant growth under lime application may also explain changes in soil respiration. To date, the effect of lime application has been tested in grassland and agricultural soils (Lochon et al., 2019; Xu et al., 2022). However, there is a lack of information on the response of soil respiration to lime application in forest ecosystems, particularly in Eucalyptus plantations grown in strong acidic soil in South China.
Eucalyptus urophylla S.T.Blake is a fast-growing, high-yielding tree species cultivated in subtropical and tropical terrestrial regions around the world. In southern China, cultivation has been expanding rapidly in response to increasing demand for fiber and wood (Wan et al., 2019; Gao et al., 2021). In southern Chinese E. urophylla monoculture plantations, the native fern Dicranopteris dichotoma, tends to create a thick understory blanket (Yang et al., 2021). In practice, understory plants are traditionally removed as they are believed to compete with cultivated trees for both nutrients and water. However, studies have shown that D. dichotoma can have profound impacts on soil organisms (Gao et al., 2021), litter decay (Chen et al., 2019), soil microclimate (Wan et al., 2019), and canopy productivity (Wan et al., 2014). Although understory plants may be a substantial driver of both above and below-ground ecosystem function, few studies have quantitatively explored the effect of understory clearance on soil respiration (Wan et al., 2015; Zhao et al., 2022). Even less is understood regarding the potential interactive effects of combined plantation management practices, such as understory clearance and lime application, on soil respiration.
In this study, a field experiment was conducted in a southern Chinese subtropical E. urophylla plantation to investigate the individual and combined effect of lime application and understory clearance on soil respiration and its two main components (autotrophic respiration and heterotrophic respiration). We hypothesized that: (1) lime application would decrease soil respiration by inhibiting soil total microbial PLFAs and associated Rh (Melvin et al., 2013; Wan et al., 2019); (2) understory clearance would reduce root respiration of understory vegetation and Rh by reducing labile carbon and nutrient substrates for soil microorganisms, thus consequently decreasing soil respiration (Wan et al., 2021; Xiao et al., 2022); (3) lime application combined with understory clearance would reduce soil respiration by inhibiting both Rh and Ra.
The field site is located at the Heshan National Field Research Station of Forest Ecosystem (22°34′N,112°50′E), Guangdong Province, Southern China. The overall climate of the area is subtropical, with an annual mean precipitation of 1,700 mm, and an annual mean temperature of 21.7°C. The soil at the field site is an Ultisol developed from sandstone (FAO, 2006). The Eucalyptus plantation at the site was re-established after the previously-planted trees (Pinus elliottii Engelm.) were clear-cut, and Eucalyptus (Eucalyptus urophylla S.T.Blake) saplings (1,650 per hectare) were planted with a spacing of 3 × 2 m in 2005. From August to September 2011, three independent Eucalyptus plantations, with about 1 ha of area for each, were selected to conduct the experiment. The understory vegetation in these plantations was nearly dominated by Dicranopteris dichotoma (a natural perennial fern species widely distributed in southern China), accompanied by a small number of other species, such as Miscanthus sinensis Andersson and Rhodomyrtus tomentosa (Aiton) Hassk.
Four experimental plots (10 m × 10 m) were established in each plantation, and each plot was applied randomly to one of the four treatments: LA (application of lime), UR (removal of understory), L+UR (application of lime and removal of understory), and CK (control/no treatment). For lime application treatments (LA and L+UR), in order to significantly increase soil pH in the context of heavy rainfall in subtropical regions, the soil surface was treated with lime (CaO) at a rate of 60 kg per 100 m2 per year, with 30 kg applied on 15 December 2011 and the other 30 kg applied on 15 August 2012. For understory removal treatments (UR and L+UR), harvesting hooks were used to manually removal both above and below-ground understory biomass before measurements were conducted (15 December 2011), and any regrowth of the understory plants was removed monthly there after with aid of machete.
LI-8100 Automated Soil CO2 Flux System (LI-COR, Lincoln, NE, USA) was used to measure soil respiration. Five PVC soil collars (5 cm in height, 20 cm in diameter) were randomly placed and inserted at 3 cm depth in each plot, and the measured soil respiration was used to indicate the total soil respiration (Rs). In each plot, a trenched subplot (1 m in length, 1 m in width, and 0.5 m in depth) was outfitted and surrounded by PVC sheets to prevent lateral root entry and to minimize nutrient and water exchange. Trenching was conducted in August 2011, about 3 months before treatments, allowing the decomposition of the roots inside the trenched plots after trenching. All vegetation was removed from the trenched subplot during the experiment. A soil collar inserted to 3 cm depth was placed in each subplot and the measured soil respiration was used to indicate soil heterotrophic respiration (Rh) due to the absence of plant biomass. Autotrophic respiration (Ra) was calculated as the difference between Rs and Rh (Dacal et al., 2020). All soil collars were inserted before any treatments were applied, and remained in place throughout the study. Soil respiration was measured twice a month from December 2011 to November 2012, and each measurement took place between 9 a.m. and 12 p.m. on a sunny day. Proportions of lime application induced soil respiration (RLI) to total soil respiration were calculated as either RLI = (RLA−RCK)/RCK × 100% or RLI = (RL+UR−RUR)/RUR × 100%, where RCK, RLA, RL+UR, RUR is soil respiration in CK, LA, L+ UR, and UR treatment, respectively. Proportions of understory respiration (RU) to total soil respiration was calculated as either RU = (RUR−RCK)/RCK × 100% or RU = (RL+UR−RLA)/RLA × 100%, where RCK, RLA, RL+UR, RUR is soil respiration in CK, LA, L+UR, and UR treatment, respectively. Annual soil average respiration rate was calculated by the sum of the soil respiration rate from each month divided by sampling time (12 months). Both soil moisture (volumetric moisture content) and temperature near the soil collar were measured at a depth of 0–5 cm using a probe attached to the Li-8100.
Soil samples were collected in December 2011 and Feb, May, Sep, December 2012, respectively. The surface litter was carefully removed prior to samplings. In each plot within each plantation, five sites were randomly selected and soil cores were collected from 0 to 10 cm depth. The soil cores from each plot were pooled. All soil samples were clear of visible stones and roots, then sieved (2 mm), and portioned into two sub-samples. Of these, one sub-sample destined for physio-chemical analysis was stored at 4°C and the other one destined for phospholipid fatty acid analysis (PLFAs) was stored at −20°C. Soil microbial community composition and abundance were estimated using PLFAs analysis (Bossio and Scow, 1998). When applying PLFAs analysis, soil bacterial abundance was estimated using PLFA markers cy19:0, cy17:0, a17:0, 17:0, i17:0, i16:0, 16:1ω7, i15:0, a15:0, and 14:0, while soil fungal abundance was determined using 18:2ω6,9c. The soil microbial community composition was estimated using the ratio of total fungal to bacterial abundance (F:B ratio) (Xiao et al., 2022). Soil pH was measured using a 1:2.5 soil: water mixture. Soil dissolved organic carbon (DOC) in fresh soil sample was extracted with 0.5 M K2SO4 and then measured with a TOC analyzer (TOC-VCPH Shimadzu Corp., Shinagawa, Japan). Soil NH4+-N and NO3–-N in fresh soil samples were extracted with 2 M KCL and then measured with a flow injection auto-analyzer (FIA, Lachat Instruments, USA).
Fine root biomass of Eucalyptus was determined in August 2012, when the trees grew most vigorously. Nine samplings were randomly sampled within each plot at a depth of 40 cm with an 8 cm diameter soil corer. Each soil core was divided into four sections based on horizon (0–10 cm, 10–20 cm, 20–30 cm, and 30–40 cm), and each section of soil was soaked and carefully sieved through a 0.5 mm mesh sieve to isolate roots, which were sorted by size (less than 5 mm or 5 mm in diameter) and vitality (living or dead according to color, and elasticity) In plantations, the understory vegetation is dominated by the fern Dicranopteris dichotoma, whose roots are rhizome and adventitious, and the fine roots can be hardly distinguished. In addition, the roots of Dicranopteris dichotoma were harder than those of Eucalyptus due to their higher lignin content (Yang et al., 2021). The roots were then dried in an oven (65°C) until they reached a constant mass and weighed.
Statistical analyses were performed in SPSS 18.0 (SPSS, Chicago, IL, USA). All data met assumptions of normality and homogeneity of variance in the present study. A two-way repeated-measure analysis of variance (ANOVAs) was conducted to determine the effect of LA, UR, and their interactions on soil temperature, moisture, pH, DOC, NH4+-N, NO3–-N, soil respiration rate (Rs, Rh, and Ra), and soil microbial communities. A one-way ANOVA and least significant difference tests (LSD) were then employed to test the significant difference among treatments on soil annual average respiration rate and Eucalyptus fine root biomass. In all cases, the statistical significance was set at P ≤ 0.05. The relationships between soil respiration (Rs, Rh, and Ra) and soil temperature or soil moisture were tested by exponential regression and linear models, respectively. The temperature sensitivity (Q10) of soil respiration (Rs, Rh, and Ra) was established using the following equations (Dacal et al., 2020):
R = aebT
Q10 = e10b
where R is Rs, Rh, or Ra, T is the temperature of the soil at a depth of 0–5 cm, and a and b are the regression coefficients.
Soil respiration (Rs, Rh, and Ra) exhibited clear temporal patterns, with the lowest respiration rate occurring in December and the highest in September (Figures 1A, B). Overall, the soil respiration rate ranged from 0.67 to 4.21 μmol m–2 s–1 (Figure 1). Lime application reduced Rs and Ra, as well as their annual average rates (Table 1 and Figures 1A, C), while it increased Rh and the annual average rate irrespective of understory removal (Table 1 and Figure 1B). Understory clearance reduced both Rs and Rh and their annual average rates (Table 1 and Figures 1A, B). It also inhibited Ra and the annual average rate (Table 1 and Figure 1C). Moreover, the LA treatment decreased Rs by 16.48% compared to the CK treatment, and the L+UR treatment decreased Rs by 4.49% relative to the UR treatment (Figure 2A). The UR treatment decreased Rs by 21.43% compared to the CK treatment, while the L+UR treatment decreased Rs by 10.24% relative to the LA treatment (Figure 2B). Both understory clearance and lime application increased the proportion of heterotrophic respiration to total soil respiration (Supplementary Figure 1A), while decreased the proportion of autotrophic respiration to total soil respiration (Supplementary Figure 1B).
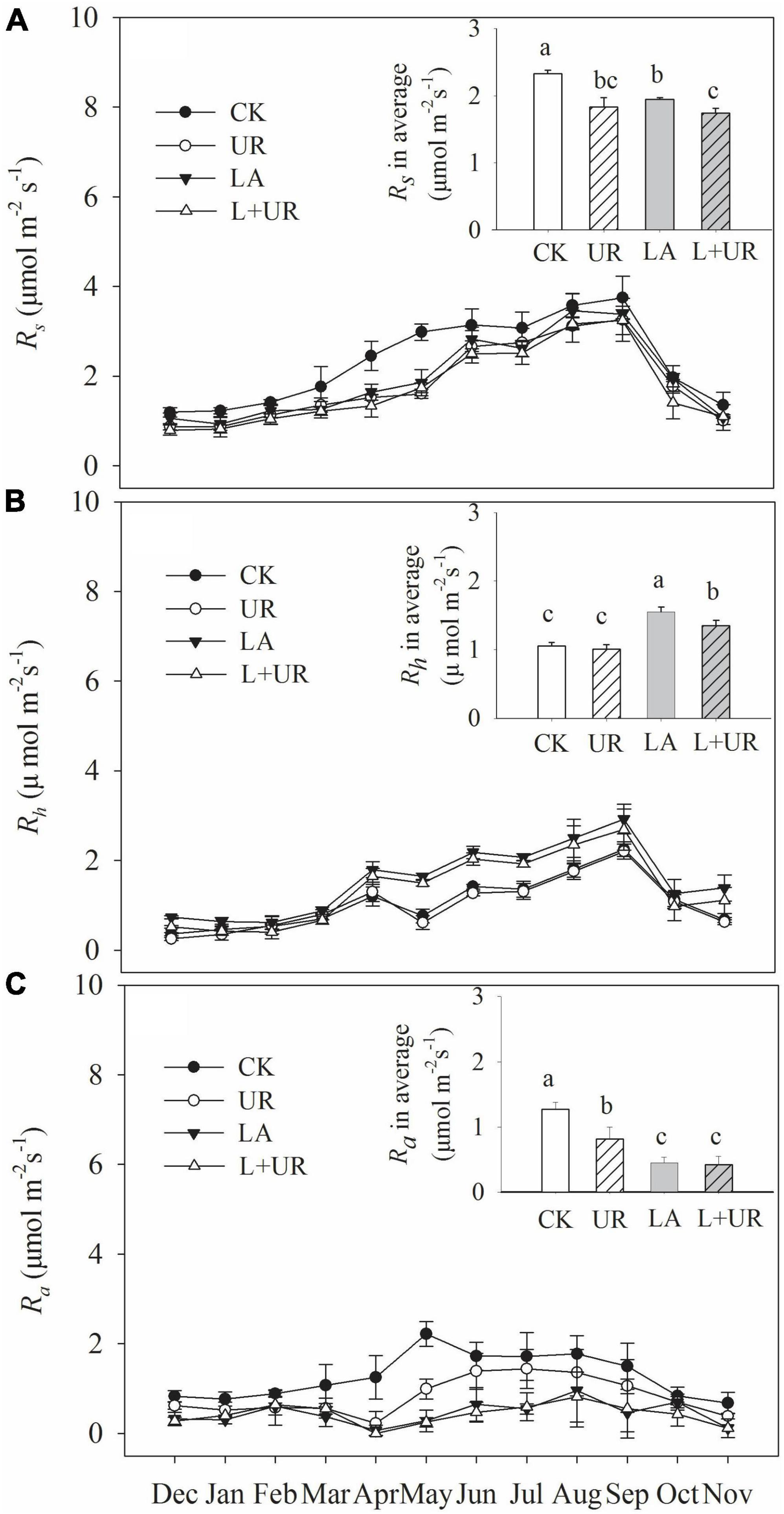
Figure 1. Variations of Rs (A), Rh (B), and Ra (C) under different treatments from December 2011 to November 2012. The inserted figure indicate the annual average of soil respiration (Rs, Rh, and Ra) under different treatments. Different lowercase letters among treatments indicate significant difference (P ≤ 0.05). Values are means with standard errors, n = 3. Control (CK), understory removal (UR), lime application (LA), and lime application with understory removal (L+UR).
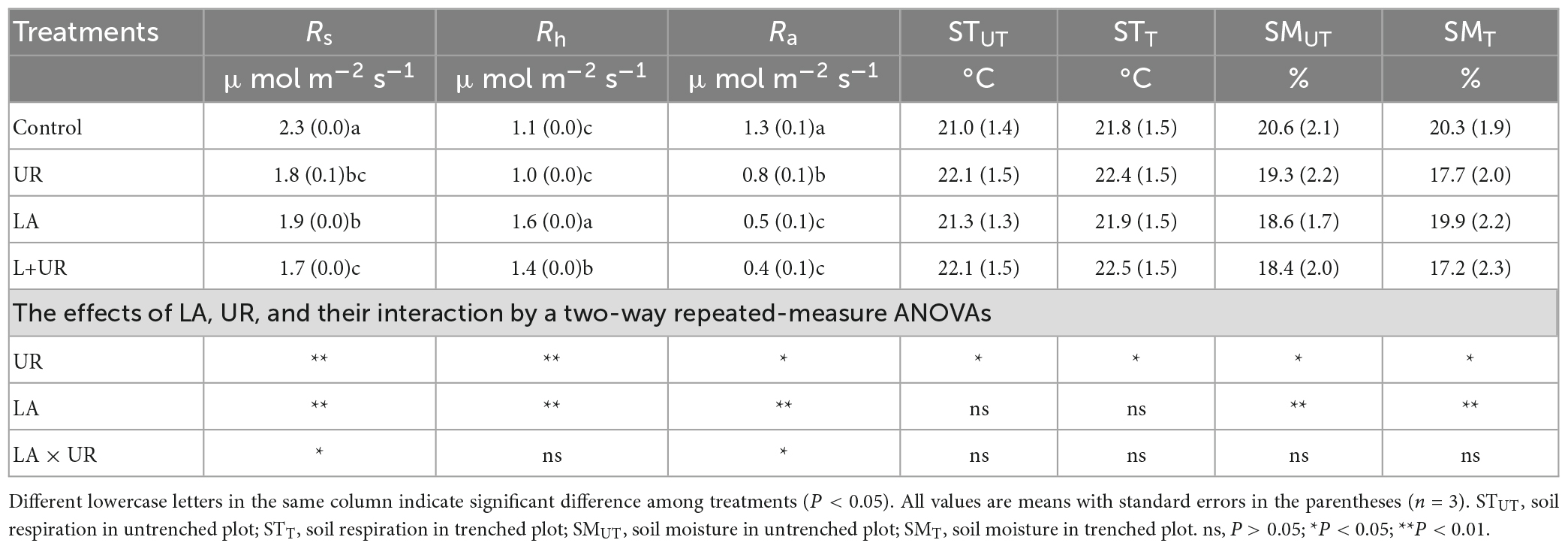
Table 1. Effects of lime application (LA), understory removal (UR), and their interactions (LA × UR) on soil respiration (Rs, Rh, and Ra), soil temperature (ST), and soil moisture (SM) in a southern Chinese Eucalyptus plantation.
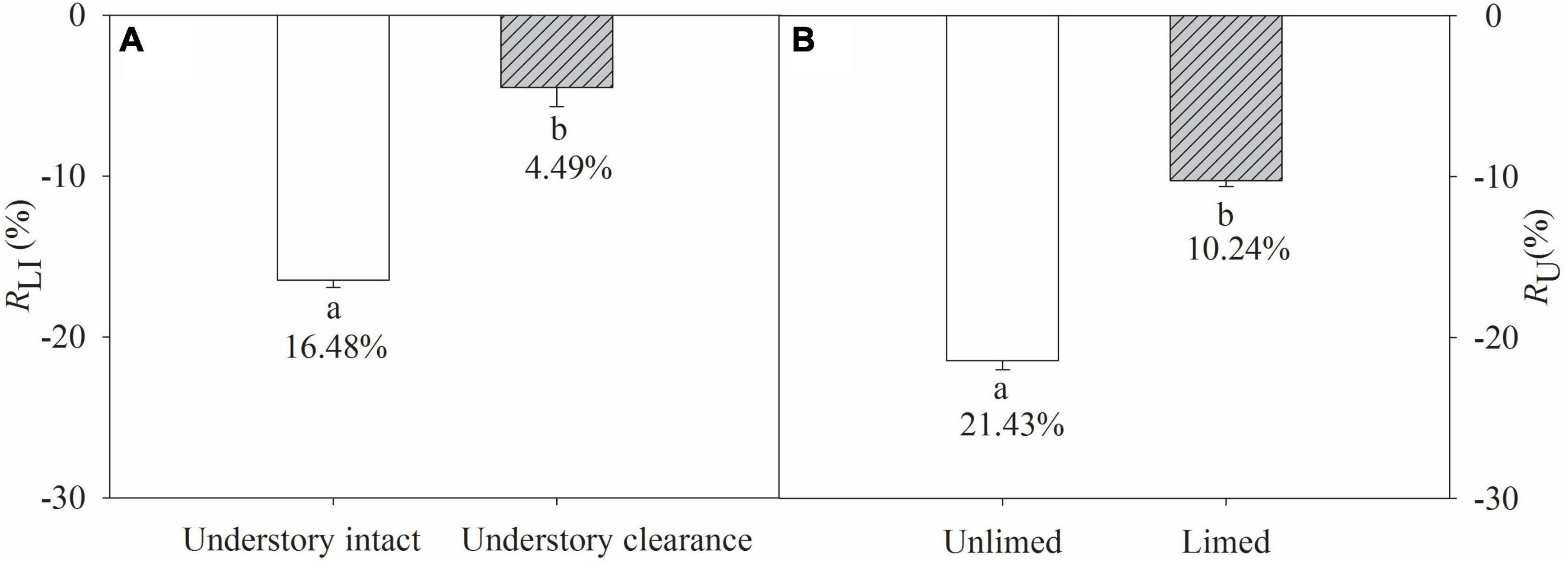
Figure 2. (A) Proportions of lime application induced soil respiration to total soil respiration (RLI) in the presence and absence of understory vegetation; (B) proportions of understory removal caused changes in soil respiration to total soil respiration (RU) in unlimed and limed soil. Values are means with standard errors, n = 3. Different lowercase letters indicate significant difference (P ≤ 0.05).
Lime application significantly increased soil pH, and soil NO3–-N content, but decreased soil moisture, with no significant impact on soil temperature and soil NH4+-N content (Table 1, Figure 3, and Supplementary Table 1). Understory clearance elevated soil temperature by 0.74°C, but decreased soil moisture content and soil NO3–-N content (Table 1, Figure 3, and Supplementary Table 1). In addition, understory clearance caused a trend of decline in DOC (P = 0.06). No significant interactions of lime application and understory clearance were observed on soil moisture, soil temperature, and soil physical-chemical properties during the experimental period (Table 1 and Supplementary Table 1).
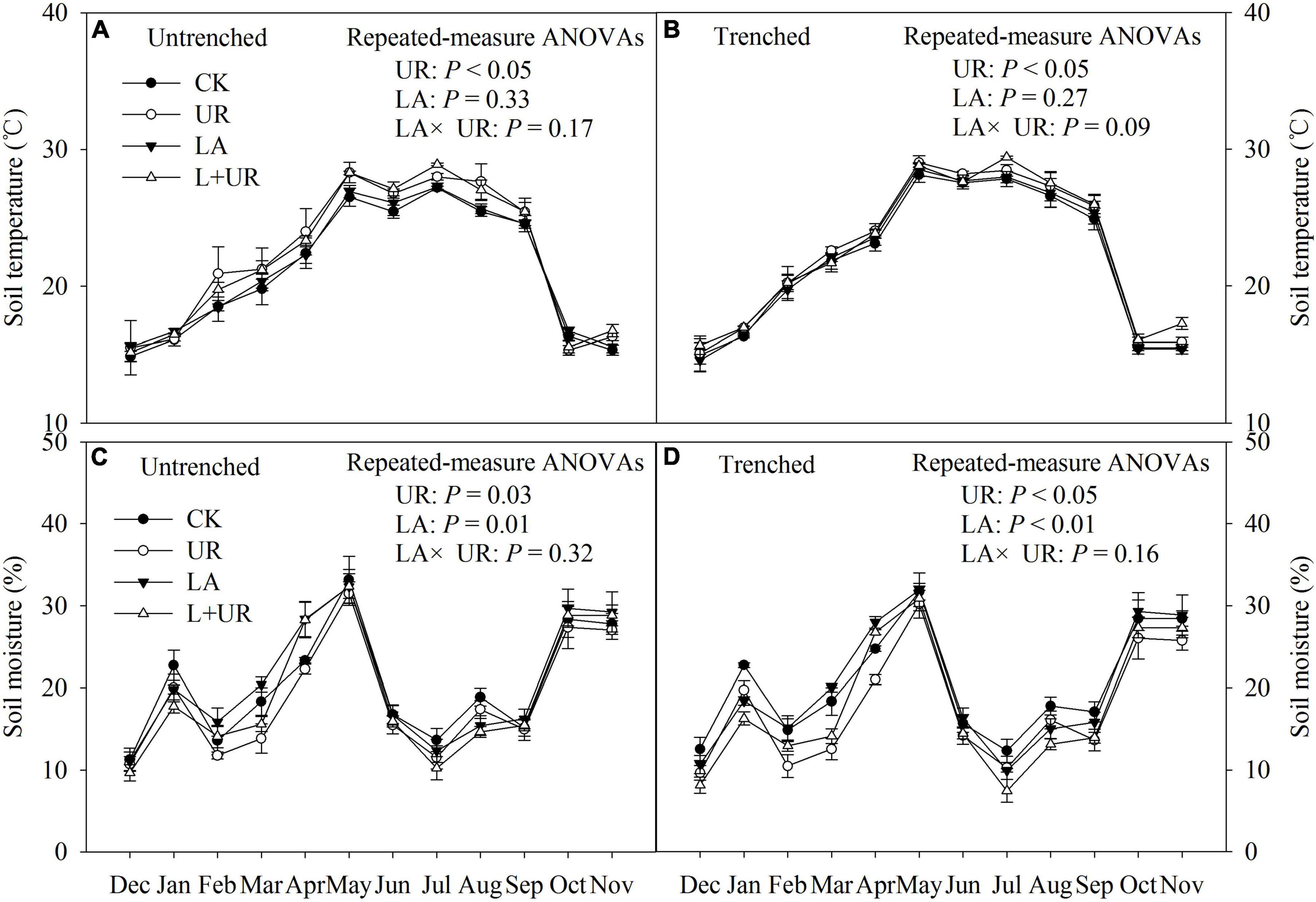
Figure 3. (A,B) Are variations of soil temperature under different treatments in untrenched and trenched plot, respectively; (C,D) are variations of soil moisture under different treatments in untrenched and trenched plot, respectively, from December 2011 to November 2012. The inserted statistical results are from a two-way repeated-measure ANOVAs. Values are means with standard errors, n = 3.
The soil microbial biomass ranged from 453 to 2,701 ng g–1 for each treatment during the experiment. Sampling time caused a significant effect on soil microbial biomass and communities. Lime application significantly decreased both soil fungal and bacterial PLFAs, and the soil total PLFAs, despite that the F:B ratio remained unchanged (Table 2). Understory clearance significantly decreased the fungal and total PLFAs without affecting bacterial PLFAs, overall leading to a reduced F:B ratio. No interactive effects of lime application and understory clearance were found on the composition of soil microbial communities.
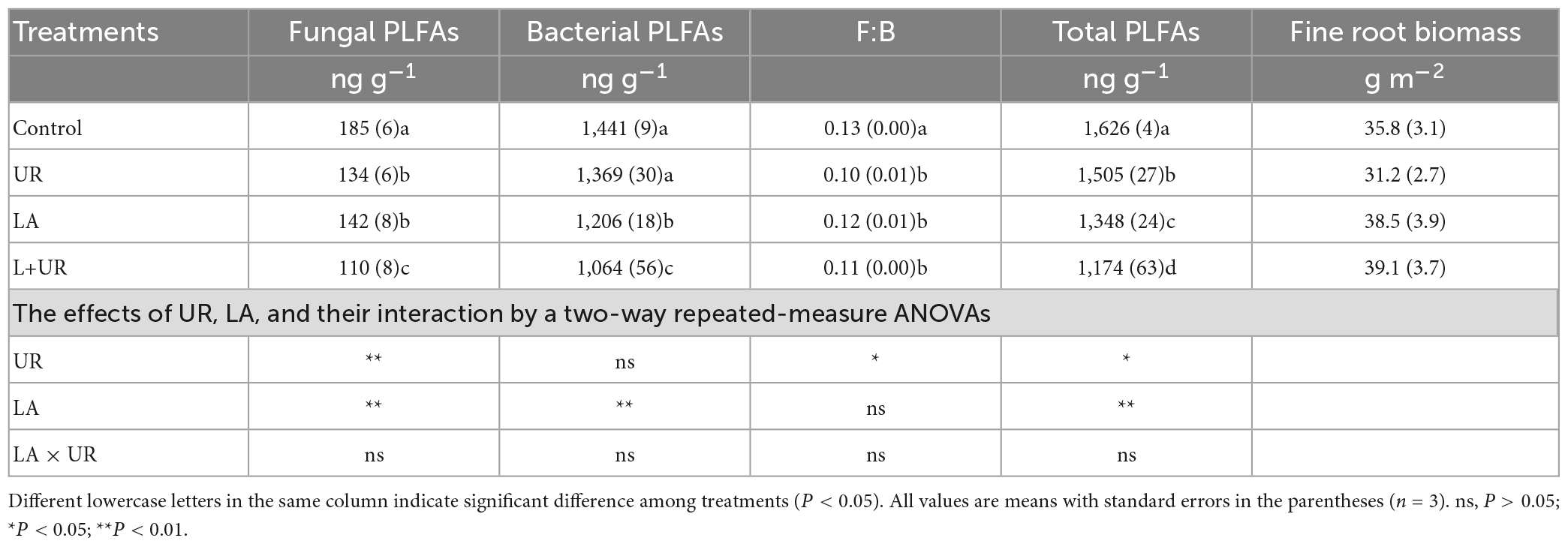
Table 2. Effects of lime application (LA), understory removal (UR), and their interactions (LA × UR) on soil microbial PLFAs, and Eucalyptus fine root biomass in a southern Chinese Eucalyptus plantation.
Eight months after treatment, the fine root biomass, was 35.8 ± 3.0, 31.2 ± 2.7, 38.5 ± 3.9, and 39.1 ± 3.7 g m–2 for CK, UR, LA, and L+UR, respectively. Neither lime application nor understory clearance had a significant impact on Eucalyptus fine root biomass (Table 2 and Figure 4).
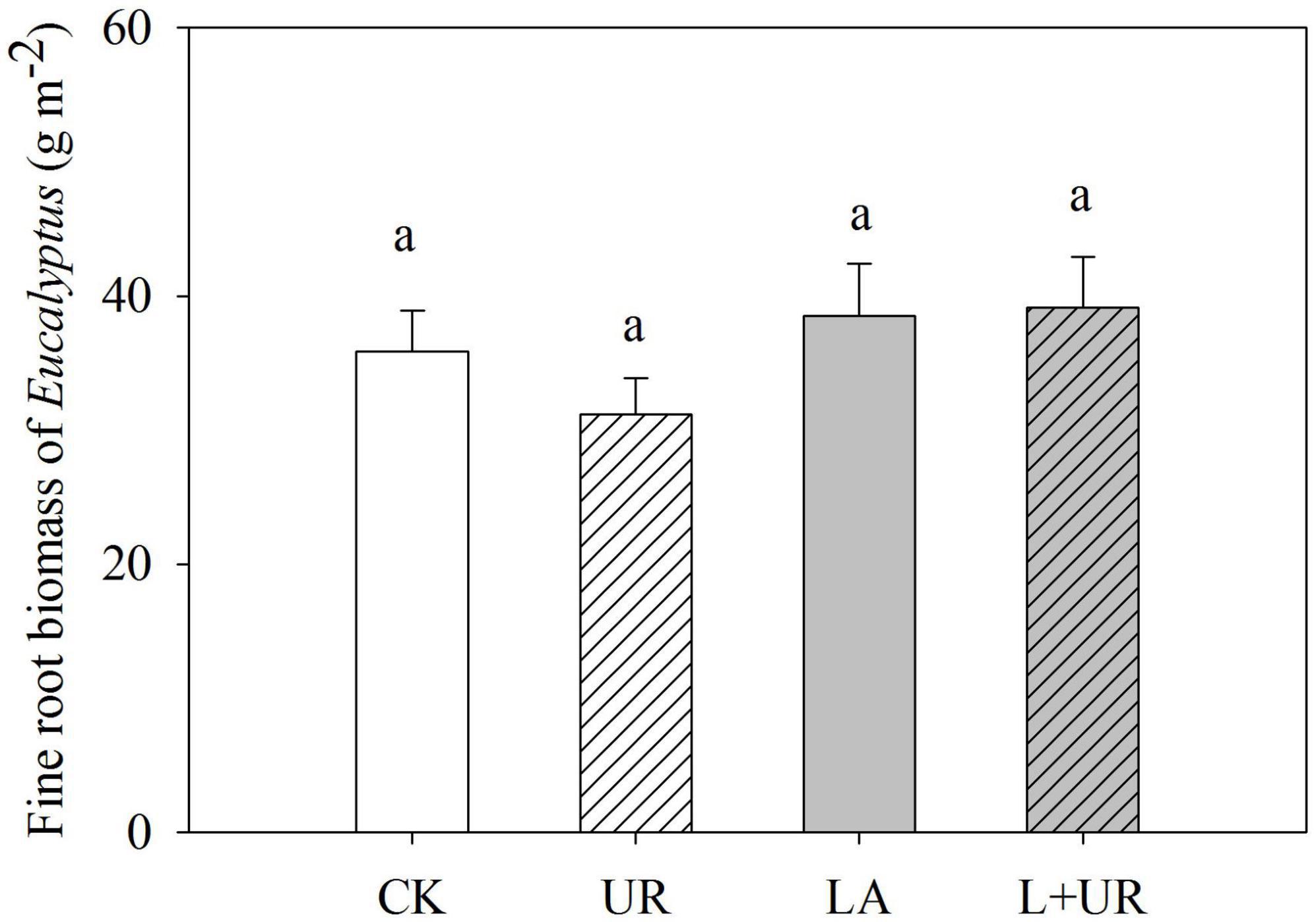
Figure 4. Eucalyptus fine root biomass after treatments were applied 8 month. Values are means with standard errors, n = 3. Different lowercase letters indicate significant difference (P < 0.05).
Soil temperature was found to be significantly positively correlated with Rs, Rh, and Ra, with the exception of Ra under lime application (Figures 5A–C). Based on exponential regression models, regardless of the treatment applied, soil temperature explained 56–82% of the variation in Rs, 33–59% of variation in Rh, and 58–92% of variation in Ra. The temperature sensitivity (Q10) varied from 2.08 to 2.21 in Rs, from 1.83 to 2.37 in Rh, and from 2.3 to 2.35 in Ra, regardless of treatment (Figures 5A–C). Neither lime application nor understory removal had a significant effect on Q10 of soil respiration and its components. Linear regression modeling showed that the effect of soil moisture on Rs, Rh, and Ra was negligible (Figures 6A–C).
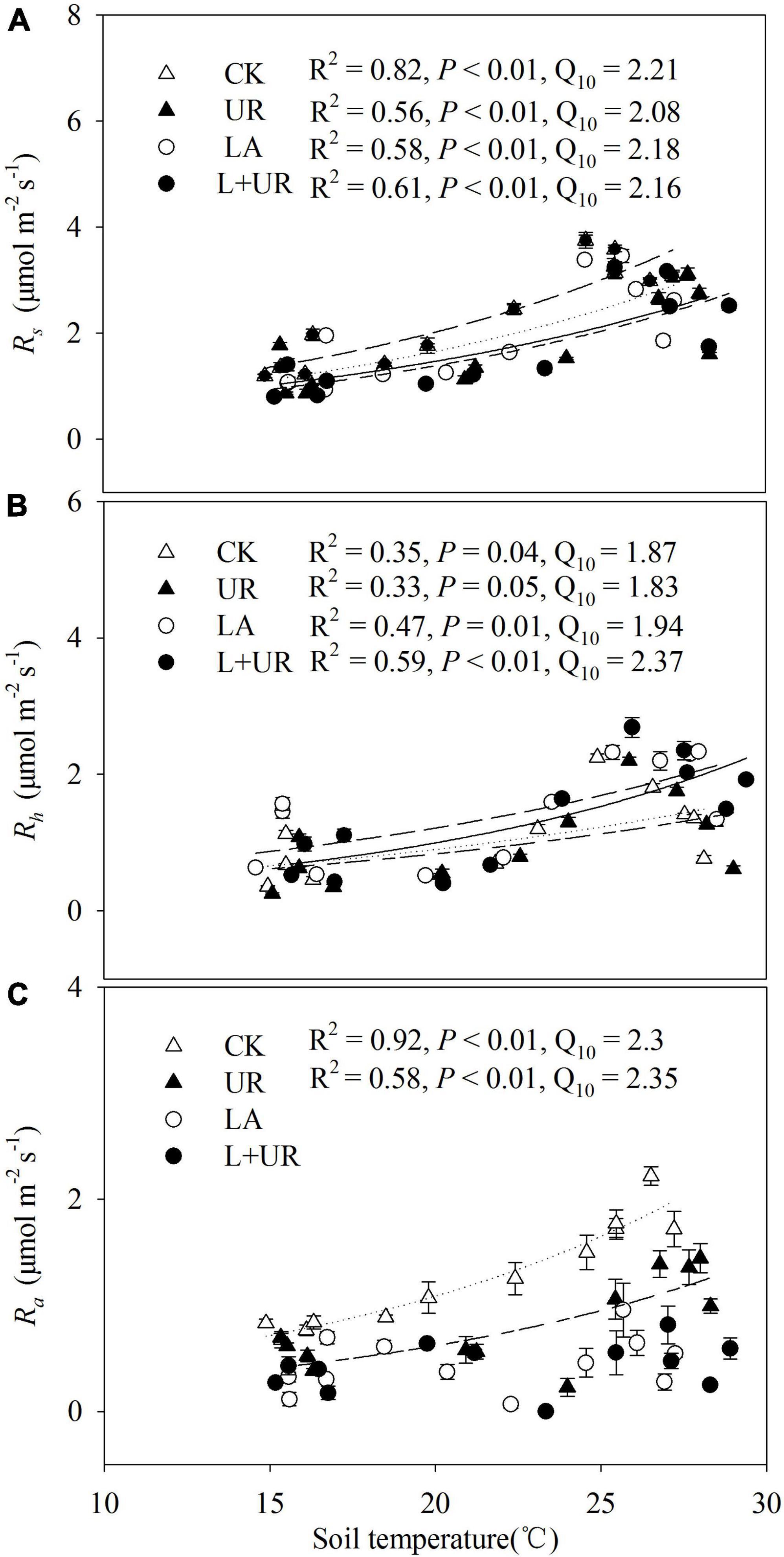
Figure 5. (A–C) Are Rs, Rh, and Ra in relation to soil temperature at 5 cm depth across the four treatments, respectively. CK, dotted line; UR, short-long line; LA, long-dash line; L+UR, solid line.

Figure 6. (A–C) Are Soil respiration (Rs, Rh, and Ra) in relation to soil moisture at 5 cm depth across the four treatments, respectively.
Studies have shown that lime application can increase Rs by increasing plant carbon input, improving soil properties and structure, and activating the activities of soil microorganisms (Ingvar Nilsson et al., 2001; Dada and Ewulo, 2011; Jiang et al., 2018; Abalos et al., 2020). However, in the present study, we did not find such a positive relationship between lime application and Rs, and lime application significantly reduced soil respiration rate irrespective of understory removal, which is consistent with the first hypothesis. Generally, soil respiration mainly includes soil heterotrophic respiration and autotrophic respiration, therefore, the effect of lime application on soil respiration depends on the balance of these two components. Although we found that lime application tended to increase Rh, this treatment also led to the consistent inhibition of Rs, primarily due to the inhibition of Ra (Figures 1A–C), with the extent of inhibition varying according to the occurrence of understory vegetation. Fine root respiration is the primary contributor to total root respiration (Chen et al., 2011). Nevertheless, lime application had no effect on Eucalyptus fine root biomass (Figure 4), thus the reduction in understory plant root respiration is likely responsible for the decreased Ra under lime application. This conclusion is further supported by the finding that lime application caused a greater reduction in Rs when the understory vegetation was intact (Figure 2A). Indeed, in the Eucalyptus plantation we studied, the understory vegetation was almost dominated by Dicranopteris dichotoma, which is considered as an indicator plant for acidic soils in tropical and subtropical regions. Its roots are densely distributed close to the soil surface, therefore, the root growth and associated root respiration are thus negatively affected by higher soil pH under lime application (Yang et al., 2021; Zhang et al., 2021). Børja and Nilsen (2009) and Lin et al. (2015) also explored that lime application can inhibit plant root growth and decomposition. Notably, lime application increased Rh (Table 1 and Figure 1B), but decreased both fungal and bacterial PLFAs (Table 2), implying that some microbial populations were inhibited at higher soil pH but others with low carbon use efficiency were promoted in growth and activity (Xiao et al., 2022). Consequently, lime application stimulated soil microbial activities and associated Rh following the findings of other researchers (Ingvar Nilsson et al., 2001; Dada and Ewulo, 2011). Given that the inhibitory effect of lime application on soil respiration was significantly greater when the understory was intact compared with when the understory was removed (Figure 2A), our results suggest that understory vegetation of Eucalyptus plantation can play a crucial role in regulating the effects of lime application on soil respiration. It should be noted that the effect of lime application on Rs has been found to vary with forest, lime formulation, treatment time, and lime application rate (Klaus et al., 2001; Børja and Nilsen, 2009).
Consistent with the second hypothesis, understory clearance caused a significant reduction in Rs (Figure 1), which is in accordance with the majority of previous work in subtropical plantations (Wan et al., 2015; Zhao et al., 2022). Studies on natural tropical forests, subtropical plantations, and boreal forests show that understory vegetation can affect Rs by modulating Rh and Ra (Fukuzawa et al., 2015; Subedi et al., 2019; Dacal et al., 2020). In the present study, understory clearance allowed a greater amount of solar radiation to reach the soil surface, and increased soil temperature by 0.74°C, which may stimulate Rh, as soil temperature was significantly positively correlated with Rh (Figure 5). Overall, however, we found that understory clearance had a negative effect on Rh. This result may be due to the reduction of DOC and soil NO3–-N content caused by the clearance of understory vegetation (Supplementary Table 1), because the growth of soil microbiota is often limited by the accessibility of soil nutrients and carbon substrates (Subedi et al., 2019). The decreased soil total PLFAs after understory clearance can further support this finding (Table 2). Furthermore, despite the effects of understory clearance on Eucalyptus root respiration and Eucalyptus fine root biomass were negligible (Figure 4), it decreased Ra, suggesting that the inhibition of Ra was mainly due to the reduction in understory root respiration. Overall, understory clearance reduced Rs by decreasing both Rh and Ra. These observations highlight that understory vegetation can play a crucial role in modulating soil microorganisms and carbon cycling in subtropical plantations. In addition, our previous studies have found that understory vegetation had a positive effect on Eucalyptus fine root biomass and productivity in 4-year-old plantations (Wan et al., 2014, 2015), while this was not the case in the present study with plantations in 6-year-old, implying that the positive effect of understory vegetation on the growth of Eucalyptus is age-dependent.
Following with the third hypothesis, Rs was decreased when lime application was combined with understory clearance (Table 1 and Figure 1). In the present study, the annual average of Rh under the L+UR treatment were significantly higher than those under the CK treatment (Figure 2B), indicating that the increase in Rh due to lime application was greater than that the decrease in Rh after understory clearance. Therefore, when lime application and understory clearance was combined, the increased Rh by lime application can be partially offset by the reduction in Rh caused by understory clearance, and the other by the reduction in Ra caused by understory clearance. Given that both lime application and understory clearance had negligible effects on Eucalyptus root respiration, we believe that the combined lime application and understory clearance can inhibit soil respiration, mainly by reducing root respiration associated with understory vegetation. In the context of the prevalence of lime application and understory clearance in subtropical plantations (Moore and Ouimet, 2021; Wan et al., 2021), our results suggest that the combined effect of lime application and understory clearance should be considered to understand and predict carbon cycling in the plantation ecosystems.
Overall, lime application and understory removal, and their combined application reduced soil respiration, suggesting that both forest management practices have the potential to increase soil carbon stocks of subtropical Eucalyptus plantations. However, whether lime application or understory removal contributes to carbon sequestration in subtropical Eucalyptus plantation requires further study. Soil microbial abundance and activity generally have positive effects on plant growth, however in this study, lime application and understory significantly reduced soil microbial biomass. Therefore, we advise that plant productivity and soil carbon stocks should be considered when taking lime application and understory removal measures in subtropical Eucalyptus plantations.
In conclusion, lime application inhibited soil respiration by decreasing the root respiration of understory vegetation. Understory clearance reduced soil respiration by inhibiting both soil heterotrophic respiration and understory root respiration. The interaction of lime application and understory clearance resulted in a reduction in soil respiration, which was mainly due to the decrease in understory root respiration. These results demonstrate that the effects of lime application on soil respiration are highly modulated by understory vegetation, highlighting the important role of understory vegetation in regulating soil carbon balance in subtropical Eucalyptus plantations. Given the critical role of soil respiration in regulating soil carbon cycling, our findings will contribute to understanding and predicting the effect of forest management practices on soil carbon storage and global carbon cycling in a changing climate.
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
JL performed the study data collection, analyzed the results, wrote the original draft, and reviewed the manuscript. RL performed the formal analysis and visualization. JX analyzed the results and reviewed the manuscript. SF designed the experiment, reviewed the manuscript, and wrote the final manuscript. SW designed the experiment, writing—review and editing, and acquired funding. All authors contributed to the article and approved the submitted version.
This study was financed by the National Natural Science Foundation of China (Nos. 41867007 and 32060267) and the Double Thousand Plan of Jiangxi Province (jxsq2018102058).
We thank the editor and the reviewers for their constructive comments on this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2023.1136474/full#supplementary-material
Abalos, D., Liang, Z., Dörsch, P., and Elsgaard, L. (2020). Trade-offs in greenhouse gas emissions across a liming-induced gradient of soil pH: Role of microbial structure and functioning. Soil Biol. Biochem. 150:108006. doi: 10.1016/j.soilbio.2020.108006
Børja, I., and Nilsen, P. (2009). Long term effect of liming and fertilization on ectomycorrhizal colonization and tree growth in old Scots pine (Pinus sylvestris L.) stands. Plant Soil 314, 109–119. doi: 10.1007/s11104-008-9710-5
Bossio, D. A., and Scow, K. M. (1998). Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty Acid profiles and substrate utilization patterns. Microb. Ecol. 35, 265–278. doi: 10.1007/s002489900082
Chen, D., Zhang, C., Wu, J., Zhou, L., Lin, Y., and Fu, S. (2011). Subtropical plantations are large carbon sinks: Evidence from two monoculture plantations in South China. Agr. Forest Meteorol. 151, 1214–1225. doi: 10.1016/j.agrformet.2011.04.011
Chen, Y., Zhang, Y., Cao, J., Fu, S., Hu, S., Wu, J., et al. (2019). Stand age and species traits alter the effects of understory removal on litter decomposition and nutrient dynamics in subtropical Eucalyptus plantations. Glob. Ecol. Conserv. 20:e00693. doi: 10.1016/j.gecco.2019.e00693
Dacal, M., García-Palacios, P., Asensio, S., Cano-Díaz, C., Gozalo, B., Ochoa, V., et al. (2020). Contrasting mechanisms underlie short- and longer-term soil respiration responses to experimental warming in a dryland ecosystem. Glob. Change Biol. 26, 5254–5266. doi: 10.1111/gcb.15209
Dada, B. F., and Ewulo, B. S. (2011). Manure and lime amendment effects on soil pH, fertility, microbe population and respiration. South Asian J. Exp. Biol. 1, 172–177. doi: 10.38150/sajeb.1(4).p172-177
Dai, W., Peng, B., Liu, J., Wang, C., Wang, X., Jiang, P., et al. (2021). Four years of litter input manipulation changes soil microbial characteristics in a temperate mixed forest. Biogeochemistry 154, 371–383. doi: 10.1007/s10533-021-00792-w
FAO (2006). World reference base for soil resources 2006. World Soil resources report 103. Rome: FAO.
Florentino, A., Masullo, L., Ferrza, A., Mateus, N., Monteleone, R., Pastoriza, L., et al. (2021). Nutritional status of Eucalyptus plantation and chemical attributes of a Ferralsol amended with lime and copper plus zinc. Forest Ecol. Manag. 502:119742. doi: 10.1016/j.foreco.2021.119742
Fukuzawa, K., Shibata, H., Takagi, K., Satoh, F., Koike, T., and Sasa, K. (2015). Roles of dominant understory Sasa bamboo in carbon and nitrogen dynamics following canopy tree removal in a cool-temperate forest in northern Japan. Plant Spec. Biol. 30, 104–115. doi: 10.1111/1442-1984.12086
Gao, D., Wan, S., Fu, S., and Zhao, J. (2021). Effects of understory or overstory removal on the abundances of soil nematode genera in a Eucalyptus plantation. Front. Plant Sci. 12:640299. doi: 10.3389/fpls.2021.640299
Goupil, K. (2014). Effects of liming on soil respiration, fungi diversity and abundance in a metal-contaminated region in Northern Ontario. Ph.D. thesis, Laurentian University of Sudbury, Sudbury.
Ingvar Nilsson, S., Andersson, S., Valeur, I., Persson, T., Bergholm, J., and Wirén, A. (2001). Influence of dolomite lime on leaching and storage of C, N and S in a Spodosol under Norway spruce (Picea abies (L.) Karst.). Forest Ecol. Manage. 146, 55–73. doi: 10.1016/S0378-1127(00)00452-7
Jiang, Y., Liao, P., van Gestel, N., Sun, Y., Zeng, Y., Huang, S., et al. (2018). Lime application lowers the global warming potential of a double rice cropping system. Geoderma 325, 1–8. doi: 10.1016/j.geoderma.2018.03.034
Klaus, L., Karl, H. F., and Ellen, K. (2001). The response of soil microbial biomass and activity of a Norway spruce forest to liming and drought. J. Plant Nutr. Soil Sci. 164, 9–19. doi: 10.1002/1522-2624(200102)164:1<9::AID-JPLN9>3.0.CO;2-Q
Lin, N., Bartsch, N., Heinrichs, S., and Vor, T. (2015). Long-term effects of canopy opening and liming on leaf litter production, and on leaf litter and fine-root decomposition in a European beech (Fagus sylvatica L.) forest. Forest Ecol. Manag. 338, 183–190. doi: 10.1016/j.foreco.2014.11.029
Lochon, I., Carrère, P., Yvin, J.-C., Houdusse-Lemenager, D., and Bloor, J. M. G. (2019). Impacts of low-level liming on soil respiration and forage production in a fertilized upland grassland in Central France. Sci. Total Environ. 697:134098. doi: 10.1016/j.scitotenv.2019.134098
Melvin, A. M., Lichstein, J. W., and Goodale, C. L. (2013). Forest liming increases forest floor carbon and nitrogen stocks in a mixed hardwood forest. Ecol. Appl. 23, 1962–1975. doi: 10.1890/13-0274.1
Mo, J., Zhang, W., Zhu, W., Gundersen, P., Fang, Y., Li, D., et al. (2008). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Change Biol. 14, 403–412. doi: 10.1111/j.1365-2486.2007.01503.x
Moore, J. D., and Ouimet, R. (2021). Liming still positively influences sugar maple nutrition, vigor and growth, 20 years after a single application. Forest Ecol. Manag. 490:119103. doi: 10.1016/j.foreco.2021.119103
Pang, X., Bao, W., Zhu, B., and Cheng, W. (2013). Responses of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agr. Forest Meteorol. 171, 57–64. doi: 10.1016/j.agrformet.2012.12.001
Shah, Z., Adamst, W. A., and Haven, C. D. V. (1990). Composition and activity of the microbial population in an acidic upland soil and effects of liming. Soil Biol. Biochem. 22, 257–263. doi: 10.1016/0038-0717(90)90095-H
Sherman, L., and Coleman, M. D. (2020). Forest soil respiration and exoenzyme activity in western North America following thinning, residue removal for biofuel production, and compensatory soil amendments. GCB Bioenergy 12, 223–236. doi: 10.1111/gcbb.12668
Sridhar, B., Wilhelm, R. C., Debenport, S. J., Fahey, T. J., Buckley, D. H., and Goodale, C. L. (2022). Microbial community shifts correspond with suppression of decomposition 25 years after liming of acidic forest soils. Glob. Change Biol. 28, 5399–5415. doi: 10.1111/gcb.16321
Subedi, P., Jokela, E. J., and Vogel, J. G. (2019). Soil respiration and organic matter decomposition dynamics respond to legacy fertilizer and weed control treatments in loblolly pine stands. Soil Biol. Biochem. 137:107548. doi: 10.1016/j.soilbio.2019.107548
Valente, F. D. A., Gomes, L. C., Castro, M. F., Neves, J. C. L., Silva, I. R., and de Oliveira, T. S. (2021). Influence of different tree species on autotrophic and heterotrophic soil respiration in a mined area under reclamation. Land Degrad. Dev. 32, 4288–4299. doi: 10.1002/ldr.4035
Wan, S., Fu, S. L., Zhang, C. L., Liu, J., Zhang, Y., and Mao, R. (2021). Effects of understory removal and litter addition on leaf and twig decomposition in a subtropical Chinese fir plantation. Land Degrad. Dev. 32, 5004–5011. doi: 10.1002/ldr.4086
Wan, S., Liu, Z., Chen, Y., Zhao, J., Ying, Q., and Liu, J. (2019). Effects of lime application and understory removal on soil microbial communities in subtropical Eucalyptus L’Hér. plantations. Forests 10:338. doi: 10.3390/f10040338
Wan, S., Zhang, C., Chen, Y., Zhao, J., Wang, X., Wu, J., et al. (2014). The understory fern Dicranopteris dichotoma facilitates the overstory Eucalyptus trees in subtropical plantations. Ecosphere 5:51. doi: 10.1890/ES14-00017.1
Wan, S., Zhang, C., Chen, Y., Zhao, J., Zhu, X., Wu, J., et al. (2015). Interactive effects of understory removal and fertilization on soil respiration in subtropical Eucalyptus plantations. J. Plant Ecol. 8, 284–290. doi: 10.1093/jpe/rtu013
Wang, Q., Zhang, W., Sun, T., Chen, L., Pang, X., Wang, Y., et al. (2017). N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agr. Forest Meteorol. 232, 66–73. doi: 10.1016/j.agrformet.2016.08.007
Xiao, Y., Xu, J. H., Zhou, B., Li, K., Liu, J., Zhang, L. P., et al. (2022). Contrasting effect of thinning and understory removal on soil microbial communities in a subtropical moso bamboo plantation. Forests 13:1574. doi: 10.3390/f13101574
Xu, C., Wong, V., Tuoninen, A., and Simojoki, A. (2022). Effects of liming on oxic and anoxic N2O and CO2 production in different horizons of boreal acid sulfate soil and non-acid soil under controlled conditions. Sci. Total Environ. 857:159505. doi: 10.1016/j.scitotenv.2022.159505
Yang, L., Huang, Y., Lima, L., Sun, Z., Liu, M., Wang, J., et al. (2021). Rethinking the ecosystem functions of Dicranopteris, a widespread genus of ferns. Front. Plant Sci. 11:581513. doi: 10.3389/fpls.2020.581513
Yang, L., Qin, J., Geng, Y., Zhang, C., Pan, J., Niu, S., et al. (2022). Long-term effects of forest thinning on soil respiration and its components in a pine plantation. Forest Ecol.Manag. 513:120189. doi: 10.1016/j.foreco.2022.120189
Yao, X., Yu, K., Wang, G., Deng, Y., Lai, Z., Chen, Y., et al. (2019). Effects of soil erosion and reforestation on soil respiration, organic carbon and nitrogen stocks in an eroded area of Southern China. Sci. Total Environ. 683, 98–108. doi: 10.1016/j.scitotenv.2019.05.221
Zhang, H., Ying, B., Hu, Y., Wang, Y., Yu, X., and Tang, C. (2022). Response of soil respiration to thinning is altered by thinning residue treatment in Cunninghamia lanceolata plantations. Agr. Forest Meteorol. 324:109089. doi: 10.1016/j.agrformet.2022.109089
Zhang, Y., Zhang, X., Qiu, Z., and Chen, Y. (2021). Ecological effect of Dicranopteris pedata and its resource utilization: A review. Pratacultural Sci. 38, 1525–1536.
Zhao, B., Ballantyne, A. P., Meng, S., Zhao, G., Zheng, Z., Zhu, J., et al. (2022). Understory plant removal counteracts tree thinning effect on soil respiration in a temperate forest. Glob. Change Biol. 28, 6102–6113. doi: 10.1111/gcb.16337
Keywords: lime application, understory clearance, soil respiration, autotrophic respiration, heterotrophic respiration
Citation: Liu J, Li R, Xu J, Fu S and Wan S (2023) Effect of lime application on soil respiration is modulated by understory vegetation in subtropical Eucalyptus L’Hér. plantations. Front. For. Glob. Change 6:1136474. doi: 10.3389/ffgc.2023.1136474
Received: 03 January 2023; Accepted: 16 February 2023;
Published: 02 March 2023.
Edited by:
Jianping Wu, Yunnan University, ChinaReviewed by:
Lingyan Zhou, East China Normal University, ChinaCopyright © 2023 Liu, Li, Xu, Fu and Wan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Songze Wan, c3dhbjA3MjJAanhhdS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.