 Veronica De Micco
Veronica De Micco Hans Beeckman
Hans Beeckman
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
EDITORIAL article
Front. For. Glob. Change , 04 January 2023
Sec. Forest Growth
Volume 5 - 2022 | https://doi.org/10.3389/ffgc.2022.1090078
This article is part of the Research Topic Vital Rates of Forest Dynamics Driven by Traits and Performance of Trees View all 6 articles
Editorial on the Research Topic
Vital rates of forest dynamics driven by traits and performance of trees
Global change is influencing plant growth and ecosystem productivity. This is a main challenge for forest ecology, aiming at understanding the dynamics of tree communities (Canadell and Raupach, 2008; Gottfried et al., 2012; Anderegg et al., 2015; Gazol et al., 2018). Modeling forest dynamics for different climate changes scenarios is substantially gaining interest, given the key role of forest ecosystem services such as carbon offsetting (Dixon et al., 1994; Favorskaya and Jain, 2017). Climate driven changes of structure and floristic composition of a vegetation are resulting from alterations in vital rates of mutually interfering populations. Vital rates depend on performance of individual trees (Westoby et al., 2002; Beeckman, 2016; De Micco et al., 2019). A wide spectrum of functional traits, including foliar, wood, tree architectural and phenological characteristics, shapes tree performance. Traits are impacted by environmental shifts, including rising temperatures, higher frequency of heat waves and forest fires, more severe and prolonged drought events, changing periodicity of rainfall, rising CO2 concentrations and insect outbreaks (Kautz et al., 2017; Masson-Delmotte et al., 2018; Ogaya et al., 2019).
Traits are input variables for some of the models that support the international policy on climate change. Relationships between traits, tree performance and forest structure are typically complex. A good understanding of trade-offs between traits, vital rates and community assemblies depends on solid data, collected at multiscale levels, involving multidisciplinary methodology (De Micco et al., 2019). The principal vital rates (i.e., regeneration/recruitment, individual growth, and survival/mortality) can be studied in isolation, but interactions can have distinctive effects on populations.
Studying global trends typically simplifies the underlying drivers. This requires selecting plant traits, that are representative and show robust correlations with relevant eco-physiological processes (Suding et al., 2008). Global models also use mean values of species traits, assuming a larger interspecific than intraspecific variation (Shipley et al., 2016), which might not be the case locally.
Hecking et al. addressed the issue of robustness of trait covariation, which can be much weaker on a smaller spatio-temporal range than at global scale. They studied trait variation in different life stages of individuals along an environmental gradient, covering temperate and montane boreal forest in northeast United States. They checked for species crossover, where the ranking of trait values changes along an environmental gradient. The authors analyzed the degree of covariation and the intraspecific trait variation in nine functional traits in overstory and understory. They highlighted that the relative importance of intraspecific trait variation is trait-specific, but globally relevant trends were still significant at local scale.
Especially for long-standing species, an often neglected source of variability is due to plant size and ontogeny. Tree size is the result of both primary and secondary growth that substantially influence the trait spectrum resulting in distinct age- or size-trends. The challenge of disentangling ontogeny and environment has been faced by Corrêa Scalon et al. who modeled growth trajectories of 45 woody species from a regenerating subtropical forest in the Brazilian Atlantic biome. Discrete grouping of growth patterns or plant traits was not detected. The authors suggested a gradual distribution of the analyzed species between two extremes defined by architecture, leaf dry matter content and growth rate dynamics. They recognized the complex nature of growth trajectories that contribute to the intrinsic diversity of the tropical forest system.
Another challenge is disentangling environmental and ontogenetic drivers from genetic effects (Housset et al., 2021). Tree populations with high genetic diversity are assumed to be more resilient, showing a stronger regrowth after disturbance (Reed and Frankham, 2003). However, unequivocal relations between tree growth and genetic diversity have not been found yet, likely because of interferences with age- and size trends. Venegas-González et al. conducted a study on five isolated populations of Nothofagus macrocarpa to evaluate the contribution of genetic variability, biogeography, and tree size on the growth resilience after an extreme drought in the only Mediterranean forest in central Chile. Tree-ring data and nuclear microsatellite markers confirmed that the resilience of N. macrocarpa could be better explained by site and tree size than genetic diversity. Nevertheless, they underlined that a whole-genome analysis might identify genomic regions correlated to the phenotype, including certain traits influencing adaptive patterns.
Dendroecology usually doesn't consider the genetic diversity within populations, assuming a stronger environmental effect on tree plasticity than genetics (King et al., 2013). Since long-lasting plants offer opportunities to construct time series based on pith-to-bark profiles of wood traits such as density, vessel features and other anatomical characteristics, a dendrochronological dimension to the topic of traits and vital rates is appealing. An often used trait is leaf phenology which is closely linked to the cambial activity and contributes to the formation of tree-ring series. Asynchronous growth patterns among trees can be interpreted in terms of resilience capacity of a population (Fritts, 2001). A study of Prioria balsamifera in the Mayombe rainforest in western Congo lead Angoboy Ilondea et al. to conclude that, despite a distinct periodicity of leaf phenology, crossdating of tree-ring series was weak. This confirms that populations of tropical rainforest trees developed a bet hedging strategy making them resilient against climatic changes.
Climate change also increased incidence and severity of insects outbreaks resulting into tree damage and eventually tree death. Finding early signals for significant tree stress after defoliation remains a challenge. Effects on plant water relations and tree growth can be positive or negative with unclear underlying mechanisms. Studying Picea mariana and Abies balsamea, defoliated by eastern spruce budworm, Balducci et al. confirmed that monitoring stem radius variation (e.g., duration and amplitude of expansion/contraction) can be useful to detect early signals of stress caused by mild defoliation.
Taken together, the contributions to this Research Topic covered some key aspects of the multiple relations between traits and forest dynamics (Figure 1). While the relationship between tree performance, vital rates of populations and composition of forest communities is unmistakable in general, empirical evidence is still needed for better predictions of population dynamics and forest tree species assemblies. Data-supported clarifications on which traits influence which vital rates are needed at multiple scales given the numerous sources of variability.
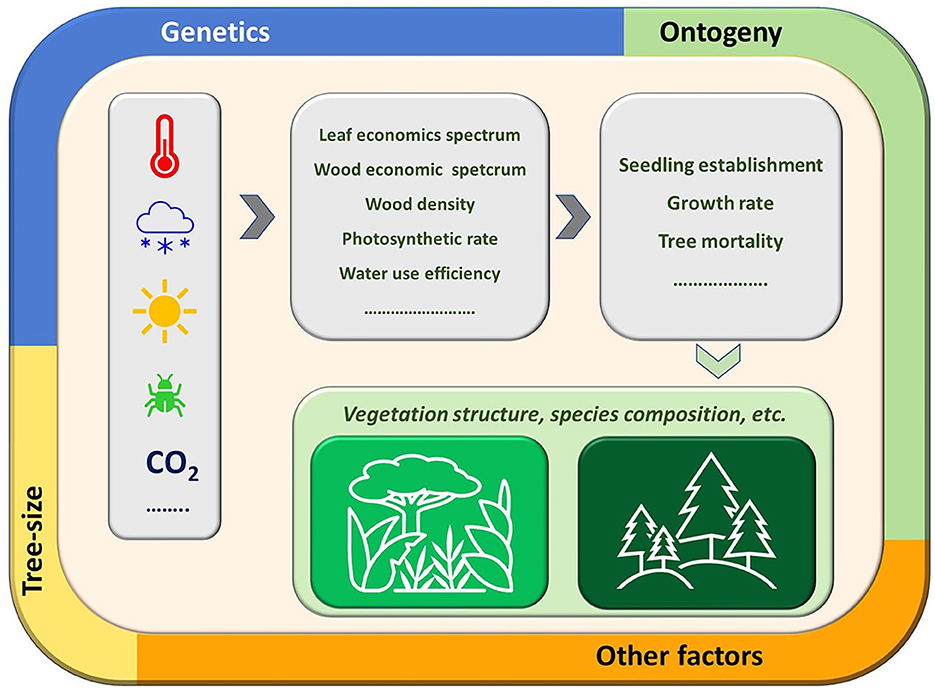
Figure 1. Schematic representation of the impact of climate-driven environmental changes on plant traits and thus on vital rates which in turn determine the composition of forest communities. Such impacts can be different on the various biomes also due to the interaction with other drivers including genetics, tree-size, and ontogeny whose weight is variable and needs further research at multiscale levels.
VD and HB conceived and designed the Speciality Section, wrote, revised, and approved the submitted version of the manuscript. Both authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Anderegg, W. R. L., Schwalm, C., Biondi, F., Camarero, J. J., Koch, G., Litvak, M., et al. (2015). Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. For. Ecol. 349, 528–532. doi: 10.1126/science.aab1833
Beeckman, H. (2016). Wood anatomy and trait-based ecology. IAWA J. 37, 127–151. doi: 10.1163/22941932-20160127
Canadell, J. G., and Raupach, M. R. (2008). Managing forests for climate change mitigation. Science 320, 1456–1457. doi: 10.1126/science.1155458
De Micco, V., Carrer, M., Rathgeber, C. B. K., Camarero, J. J., Voltas, J., Cherubini, P., et al. (2019). From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 40, 155–182. doi: 10.1163/22941932-40190246
Dixon, R. K., Brown, S., Houghton, R. A., Solomon, A. M., Trexler, M. C., and Wisniewski, J. (1994). Carbon pool and flux of global forest ecosystems. Science 263, 185–190. doi: 10.1126/science.263.5144.185
Favorskaya, M. N., and Jain, L. C. (2017). “Modelling of forest ecosystems,” in Handbook on Advances in Remote Sensing and Geographic Information Systems. Intelligent Systems Reference Library (Cham: Springer), 122. doi: 10.1007/978-3-319-52308-8
Gazol, A., Camarero, J. J., Vicente-Serrano, S. M., Sánchez-Salguero, R., Gutiérrez, E., de Luis, M., et al. (2018). Forest resilience to drought varies across biomes. Glob. Chang. Biol. 24, 2143–2158. doi: 10.1111/gcb.14082
Gottfried, M., Pauli, H., Futschik, A., Akhalkatsi, M., Barancok, P., Alonso, J. L. B., et al. (2012). Continent-wide response of mountain vegetation to climate change. Nat. Clim. Change 2, 111–115. doi: 10.1038/nclimate1329
Housset, J. M., Tóth, E. G., Girardin, M. P., Tremblay, F., Motta, R., Bergeron, Y., et al. (2021). Tree-rings, genetics and the environment: Complex interactions at the rear edge of species distribution range. Dendrochronologia 69, 125863. doi: 10.1016/j.dendro.2021.125863
Kautz, M., Meddens, A. J. H., Hall, R. J., and Arneth, A. (2017). Biotic disturbances in Northern Hemisphere forests—A synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr. 26, 533–552. doi: 10.1111/geb.12558
King, G. M., Gugerli, F., Fonti, P., and Frank, D. C. (2013). Tree growth response along an elevational gradient: Climate or genetics? Oecologia 173, 1587–1600. doi: 10.1007/s00442-013-2696-6
Masson-Delmotte, V., Zhai, P., Pörtner, H. O., Roberts, D., Skea, J., Shukla, P. R., et al. (2018). Global warming of 1.5°C. IPCC Spec. Rep. Impacts Glob. Warm. 1, 1–9.
Ogaya, R., Liu, D., Barbeta, A., and Peñuelas, J. (2019). Stem mortality and forest dieback in a 20-year experimental drought in a Mediterranean holm oak forest. Front. For. Glob. Chang. 2, 89. doi: 10.3389/ffgc.2019.00089
Reed, D. H., and Frankham, R. (2003). Correlation between fitness and genetic diversity. Conserv. Biol. 17, 230–237. doi: 10.1046/j.1523-1739.2003.01236.x
Shipley, B., De Bello, F., Cornelissen, J. H. C., Laliberté, E., Laughlin, D. C., and Reich, P. B. (2016). Reinforcing loose foundation stones in trait-based plant ecology. Oecologia 180, 923–931. doi: 10.1007/s00442-016-3549-x
Suding, K. N., Lavorel, S., Chapin, F. S., Cornelissen, J. H., Díaz, S., Garnier, E., et al. (2008). Scaling environmental change through the community-level: A trait-based response-and-effect framework for plants. Glob. Change Biol. 14, 1125–1140. doi: 10.1111/j.1365-2486.2008.01557.x
Keywords: shifting tree flora, tree performance, forest regeneration, tree growth, tree mortality, plant traits
Citation: De Micco V and Beeckman H (2023) Editorial: Vital rates of forest dynamics driven by traits and performance of trees. Front. For. Glob. Change 5:1090078. doi: 10.3389/ffgc.2022.1090078
Received: 04 November 2022; Accepted: 19 December 2022;
Published: 04 January 2023.
Edited by:
Giorgio Vacchiano, University of Milan, ItalyReviewed by:
Mart-Jan Schelhaas, Wageningen University and Research, NetherlandsCopyright © 2023 De Micco and Beeckman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Veronica De Micco,  ZGVtaWNjb0B1bmluYS5pdA==
ZGVtaWNjb0B1bmluYS5pdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.