 Jeanette S. Blumroeder
Jeanette S. Blumroeder Frederic Schmidt
Frederic Schmidt Pierre L. Ibisch
Pierre L. Ibisch- Faculty of Forest and Environment, Centre for Econics and Ecosystem Management, Eberswalde University for Sustainable Development, Eberswalde, Germany
Introduction: The effects of climate change are exacerbating the fire risk in forests worldwide. Conifer plantations in particular are especially vulnerable to fire outbreaks. At the end of the extraordinarily hot and dry summer of 2018, a forest pine plantation burned in Brandenburg, NE Germany. Different forestry interventions were carried out after the fire, while one area of the damaged plantation remained untouched.
Methods: We investigated the resilience of the forest ecosystem and the effectiveness of different active and passive forest restoration measures during the subsequent relatively warm and dry years 2019–2021.
Results: One year after the fire, Populus tremula showed strong spontaneous colonization at all sites. In contrast, the majority of planted Pinus sylvestris plantlets died on the plots that had been salvage-logged after the fire. Three years after the fire, Populus tremula successfully established itself as the dominant tree species on all plots, with the highest abundance on the plot where the overstorey of the dead pines was left. Betula pendula, Salix caprea, and Pinus sylvestris showed lower abundance, with their proportion increasing with decreasing cover by dead trees. The distribution of regrowing trees is very heterogeneous across the different treatments and plots. In the clear-cut plots, the extreme microclimatic conditions expose the young trees to additional heat and drought, while the retention of deadwood measurably buffers the temperature and water stress.
Discussion: The resilience and adaptability of naturally regenerating forests that develop into ecosystems that are more diverse seem more promising than restoration through intervention. Apart from hampering restoration under extreme weather conditions, post-fire salvage logging contributes to soil degradation and loss of organic carbon.
Introduction
Landscape fires are a substantial and inherent component of many ecosystems around the World (Pausas and Keeley, 2019). Between 2001 and 2010, a global average of approximately 464 Mha burned every year, with a relatively low share in forest biomes (Randerson et al., 2012). In many regions of the world, fires have become larger, more frequent, and more widespread (Iglesias et al., 2022). For instance, in southern boreal forests of Western Siberia, the current fire regime is unprecedented for the last five millennia (Feurdean et al., 2020). In general, Earth seems to enter an age of fire, the Pyrocene (Pyne, 2020). Fires are increasingly perceived as a challenge, and fire suppression management has been implemented (Doerr and Santín, 2016). In Europe, between 1986 and 2016, an average of approximately 140,000 ha of fire-related disturbances was recorded per year, accounting for 0.06% of the total forested area (Senf and Seidl, 2021). The frequency and impact of forest fires depend on the region, climate, vegetation composition, and human land use (Lavorel et al., 2007). By emitting greenhouse gases and aerosols as well as modifying surface properties, fires can affect not only vegetation but also climate (Lavorel et al., 2007). As an effect of ongoing climate change, the increasing frequency of fire weather, the expanding length of the fire season, the area burned and the occurrence of fires will become all the more important in the future (Bowman et al., 2020). Apart from climate change, the forests’ structural, physiological and mechanical characteristics are other decisive factors affecting forest vulnerability and these can be controlled by forest management (Forzieri et al., 2021).
In Central Europe, only 0.5% of fires are of natural origin, while ca. 39% are negligently and 56% deliberately ignited due to arson (Ganteaume et al., 2013). In Germany, the average area burned since 1991–2018 was around 447 ha per year and only 283 ha between 2009 and 2018 with around 0.5 ha burned per fire, while the state of Brandenburg represents the area that burned the most (Fernandez-Anez et al., 2021). Most fires in Germany (65%) were recorded in coniferous forests (Gnilke and Sanders, 2021). The dominant forest type in the region of Brandenburg are Scots pine (Pinus sylvestris) monocultures on sandy soils that have replaced many of the previous and less-flammable broadleaved forest (Dietze et al., 2019).
As summarized by Dimitrakopoulos et al. (2011), the ignitability or flammability of plants is related to the time and temperature of ignition (Anderson, 1970; Martin et al., 1994) as well as the chemical fuel properties (Rundel, 1981). This relates to the heat and ash contents, as high heat content increases fuel combustibility and fire intensity (Shafizadeh et al., 1977) and high ash content reduces fuel flammability (Philpot, 1970). Pinus sylvestris takes 28.063 s to ignite at 248.25°C ignition temperature, and has a relatively high heat and low ash content compared to other coniferous species (Dimitrakopoulos et al., 2011). The flammability of cones and needle litter is another important component that contributes to the fire properties of pine forests (Fonda, 2001; Fonda and Varner, 2004). Especially the moisture content was reported to be a relevant factor due to increasing ignition times and lower ignition frequency (Popović et al., 2021). A moisture content of the surface material of less than 30–35% increases the fire ignition probability (Tanskanen et al., 2005). Besides that, organic metabolites such as cellulose, tannins, terpenes, and alkane wax contents are crucial chemical components (Ormeño et al., 2020). Canopy depth and leaf area index are forest stand characteristics negatively correlated with ignitability, while clearcuts of former Scots pine stands showed the highest ignition probability; also the frequency of ignitions was lower for sites with higher canopy cover and leaf area index (Tanskanen, 2007). The flammability of pine stands was shown to be higher in stands that have burned previously considering flame height and residence, shoot flammability as well as time to ignition or higher combustibility (Romero and Ganteaume, 2021).
Although fire prevention, early detection, and documentation have improved, there is less agreement on how forest owners should manage their forests after fire disturbance, especially in areas where there have been far fewer fires in the past.
As the frequency of fire events is expected to increase in Central European forests (San-Miguel-Ayanz et al., 2019), an appropriate post-fire management approach is needed to support effective forest restoration while safeguarding the provision of ecosystem services and reducing the risk of future fire events (Ascoli et al., 2013). Many strategies focus on fire prevention and increased suppression through the development of detection technologies, and less effort is expended in forest management measures such as improving forest structures and species composition and converting timber plantations into more natural and climate-resilient forests that are less prone to fire (FAO, 2007). Regardless of international scientific evidence, the conventional response to calamities is mostly the immediate clearing of sites and artificial reforestation (Moreira et al., 2012a). This is often also promoted legally and financially by policy makers in the European Union (Vallejo et al., 2012b). However, this is not always the most cost-effective approach and ignores the natural properties of the ecosystem that can facilitate the restoration processes (Moreira et al., 2012b). Soil protection and water regulation, management of trees, habitats, and biodiversity are to be considered as restoration objectives (Robichaud, 2009; Vallejo et al., 2012a). In the worst case, fires and forest management can increase the risk of regeneration failure. In Siberia, it was found that dry pine stands and repeatedly disturbed forests were the most vulnerable to experience a biome shift during regeneration toward steppe vegetation (Kukavskaya et al., 2016).
In this study, we investigate the post-fire succession of a forest fire area that burned in summer 2018 and the effectiveness of different silvicultural treatments on forest regeneration and microclimate regulation. In particular, sites that had been cleared, thinned, or not managed were investigated in both one and two vegetation periods after a forest fire (e.g., after one growing season in December 2019 and after two growing seasons in May 2021) in a Scots pine plantation in north-eastern Germany. Measurements of temperature and relative humidity accompanied the recording of succession.
Materials and methods
Study sites
The study sites are located in a forest fire area that burned in late August 2018 in the south of Brandenburg, Germany, approximately 60 km south of the Berlin city border. The fire affected about 400 ha of Scots pine monocultures that were established for timber production in different years after World War II. Most of the pine trees died immediately or in the following weeks after the forest fire. After the fire, different forest owners chose a variety of restoration interventions, while some of the areas remained unmanaged. After the first investigations started already in 2019, an ecosystemic research project was initiated in May 2020 with comprehensive ecological monitoring.1 Some plots sampled in the present study are part of the PYROPHOB research project. The project is based on the collaboration of eight institutions that investigate the effects of forest fire and different post-fire management approaches on ecosystemic development to formulate strategies for developing more pyrophobic and climate-resilient forests in burned areas.
The study area is characterized by relatively low annual precipitation of 500–550 mm on average and a negative ecoclimatic water balance of –220 to –170 mm per year and therefore represents one of the driest regions in Germany (Schulze et al., 2013). It is also characterized by relatively high temperatures in summer and cold winters, so that the annual fluctuations are relatively large. The average annual temperature is 8.5°C with an annual variation of 18.5 and 19.0°C (Gauer and Aldinger, 2005). The postglacial landscape has an average altitude between approximately 95 m and 110 m above sea level. The soil was formed from deposits of meltwater sands and periglacial and fluviatile sands and gravels.
The years 2018–2021 were exceptionally warm (DWD, 2022) and dry with severe impacts on soil water availability and drought stress (Marx et al., 2016) across Germany.
Tree rejuvenation and microclimate were sampled in the years 2019 and 2021. However, the location of both sampled indicators does not always coincide with each other (Figure 1). Some sites were surveyed before and after forestry interventions had been carried out and therefore had to be attributed to different treatments for the two sampling periods (Table 1).
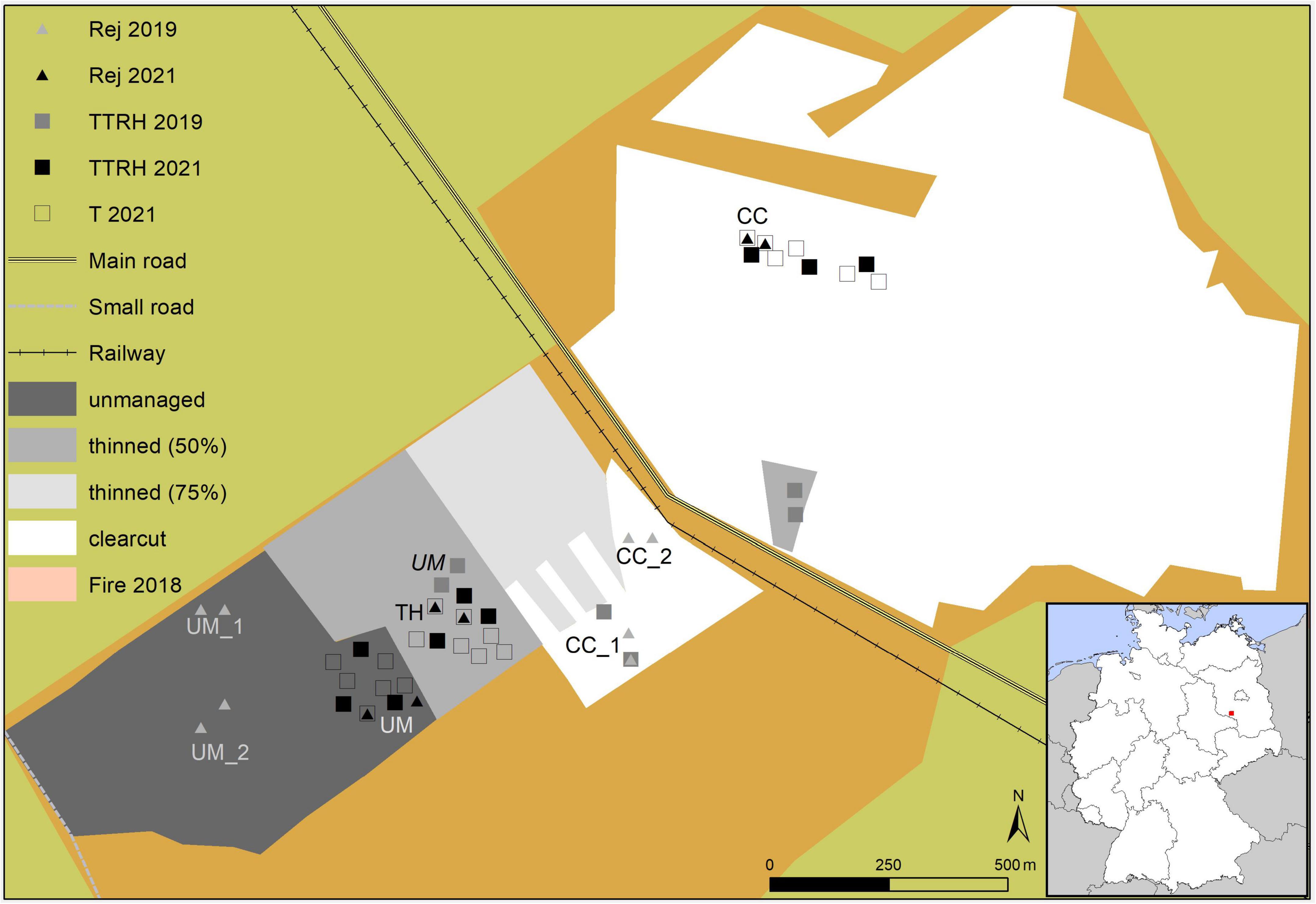
Figure 1. Overview of study sites in the south of Brandenburg, Germany, and respective treatments and sample plots (Rej, location of the rejuvenation assessment and the corresponding year; TTRH, location of temperature data-loggers at 1.3 m above ground together with data-loggers measuring temperature and relative humidity at 0.1 m above ground in the corresponding year; T 2021, temperature data-logger at 1.3 m above ground in the year 2021).
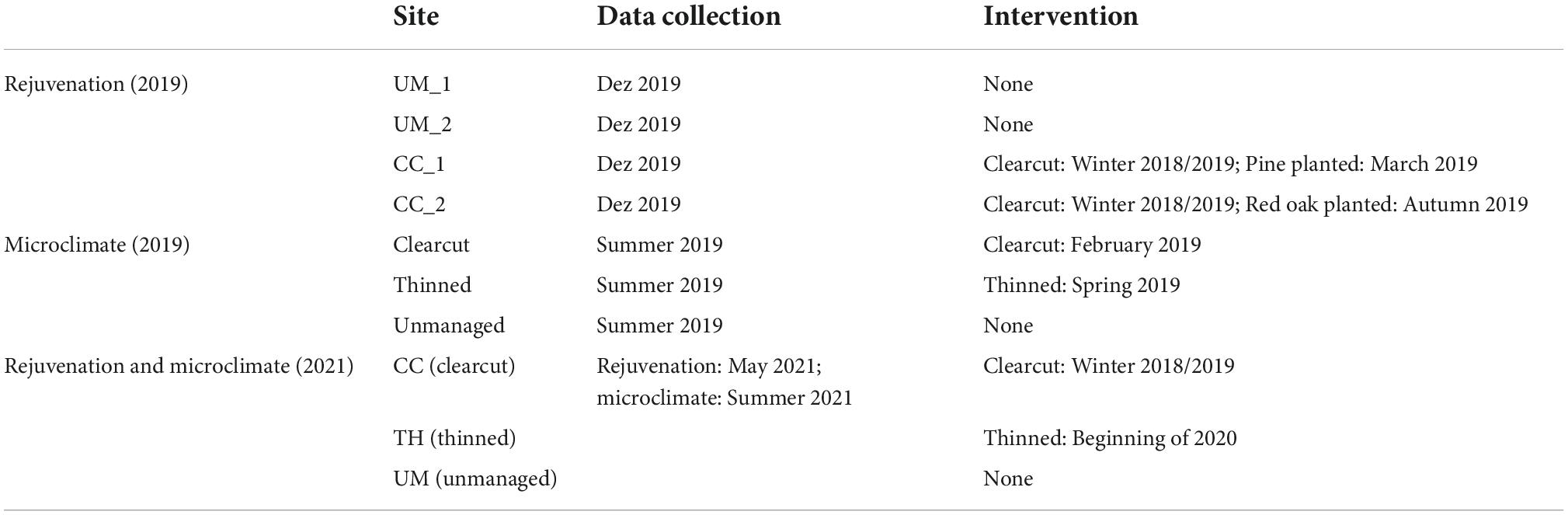
Table 1. Overview of time and kind of intervention on the study sites in relation to the collected data.
The study sites are located in the flat lowlands of Brandenburg on poor sandy soils. Due to the proximity of sampling plots and the absence of significant topographic structures, other factors such as elevation, slope, exposition, and soil were not considered in this analysis.
Rejuvenation
Tree rejuvenation was surveyed in December 2019, the first growing season after the fire and in May 2021, when the third growing season started. In 2019, two clearcut sites (CC_1, CC_2) and two unmanaged sites (UM_1, UM_2) were investigated on two sample plots each. In 2021, one clearcut (CC), one thinned (TH), and one unmanaged (UM) site were surveyed on two sample plots each.
Before the surveys started in 2019, a north-oriented 50 m to 50 m grid with a centered point in each grid was created within the forest fire polygon using ArcMap (Version 10.7.1). Within homogeneous forest sections, centric points were selected as study plots but leaving out forest roads and skidding trails. The points were located with a GPS device with an accuracy of 5–10 m. There, sample plots of 10 m radius were established, every rejuvenating tree individual was recorded and taxonomically determined. For each tree also the vitality status was documented, differentiating apex, side shoot, and leaf damages. In addition, dead individuals were counted in 2019. In 2021, combined damages were registered, which were not encountered in 2019. Dead trees were not recorded in 2021 due to bad identifiability. The number of trees counted on the sampling plots was interpolated to one hectare.
Microclimate
Microclimate data were collected during summer 2019 and 2021. In 2019, two sample plots per site with two data-loggers each were mounted on a wooden pole into a white box facing north in order to protect them from direct sunlight (Blumröder et al., 2021). One data-logger measuring temperature was located at 1.3 m on the pole, another one measuring temperature and relative humidity was at 0.1 m above ground on the same pole. Data-loggers were installed on May 10, 2019, and records were taken starting from May 11, 2019 in 30-min intervals. Before demounting the data-loggers on September 13, data were downloaded twice and all records of installation and demounting days were excluded as samples. From the remaining 123 days in the sampling period, the hottest days were selected, being defined as days with a daily mean temperature over all samples above 23.22°C, which represents the upper 25% quantile. From the resulting 31 days, the five maximum values per day were extracted and averaged as the daily maximum temperature (Tmax) for each data-logger. The driest days were filtered as days with a daily mean vapor pressure deficit (VPD) over all samples above 1.22 kPa, which represents the upper 25% quantile. From the resulting 31 days, the five maximum values were extracted and averaged as daily maximum VPD (VPDmax) for each data-logger.
Data-loggers were installed again on May 18, 2021, but in a slightly different setting. In the frame of the PYROPHOB project, ten sampling plots per treatment were established. On each sampling location, a wooden pole with a white box facing north containing one data-logger measuring temperature at 1.3 m above ground was installed. On three plots per treatment, a second data-logger measuring temperature and relative humidity was installed at 0.1 m above ground. Records were taken starting from May 19, 2021 in 10-min intervals until August 25, 2021. After excluding days with incomplete samples, out of the remaining 86 days the hottest days were selected as days with a daily mean temperature over all samples above 22.52°C, which represents the upper 25% quantile. From these resulting 22 days, the five maximum values were extracted and averaged as daily maximum temperature (Tmax) for each data-logger. The driest days were filtered as days with a daily mean VPD over all samples above 1.19 kPa, which represents the upper 25% quantile. From the resulting 22 days, the five maximum values were extracted and averaged as daily maximum VPD (VPDmax) for each data-logger.
Non-parametric tests (Kruskal–Wallis rank-sum test) were conducted to test for differences among the treatments as normality distribution could not be confirmed according to Shapiro–Wilk normality test with Pairwise Wilcoxon Rank Sum Tests (p-values adjusted using Bonferroni corrections) for pairwise comparisons among treatments. The treatments comprise clearcuts (e.g., post-fire salvage-logged where all trees have been removed using harvesters and skidders), thinned (e.g., about half of the trees have been removed using harvesters and skidders), and unmanaged (none of the trees have been removed after the fire). All data processing, analyses, and figures were computed using R (R Studio Version 1.3.1093).
Results
In 2019, one growing season after the forest fire, the passively restored sites, where no intervention was carried out after the fire, showed the highest number of rejuvenating trees (mean = 39,248 trees per hectare; standard deviation = 15,181; n = 4). On actively restored sites, where trees were planted after clearcutting, fewer trees were recorded (mean = 33,709 trees per hectare; standard deviation = 12,392; n = 4). In 2021, two growing seasons after the fire, fewer trees were recorded. Sites where all timber remained on site after the forest fire showed the highest number of living young trees (mean = 21,740 trees per hectare; standard deviation = 22,328; n = 2) followed by the clearcut sites (mean = 19,752 trees per hectare; standard deviation = 2,814; n = 2) and sites where burnt trees were partially removed (mean = 13,226 trees per hectare; standard deviation = 1,598; n = 2). Populus tremula was the dominant rejuvenating tree species across all sites and years (Figures 2, 3).
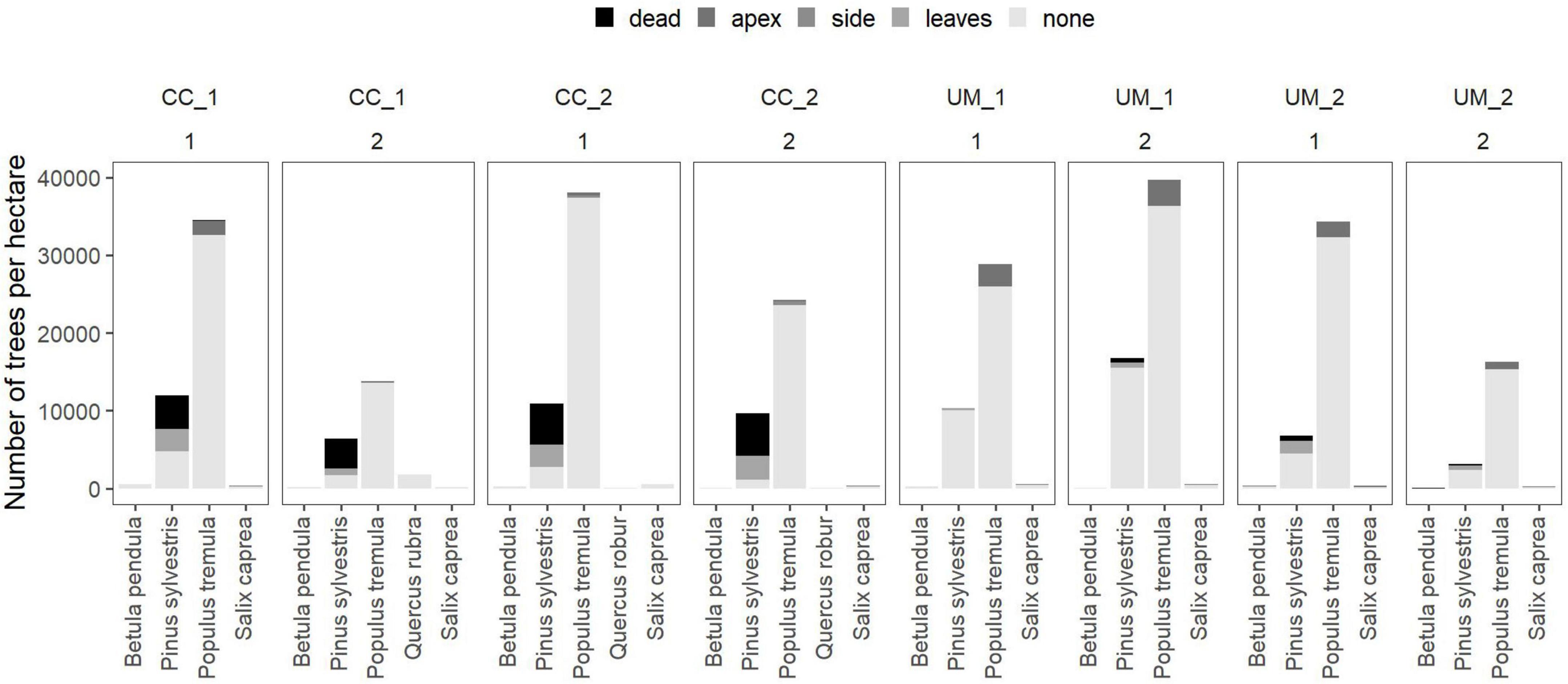
Figure 2. Distribution of vitality and damages for tree species occuring per treatment (CC, clearcut; UM, unmanaged) and plot (two plots per treatment represented by the number below the treatment name) recorded in the year 2019. Colors of the stacked bars indicate the damage recorded on each tree individual.
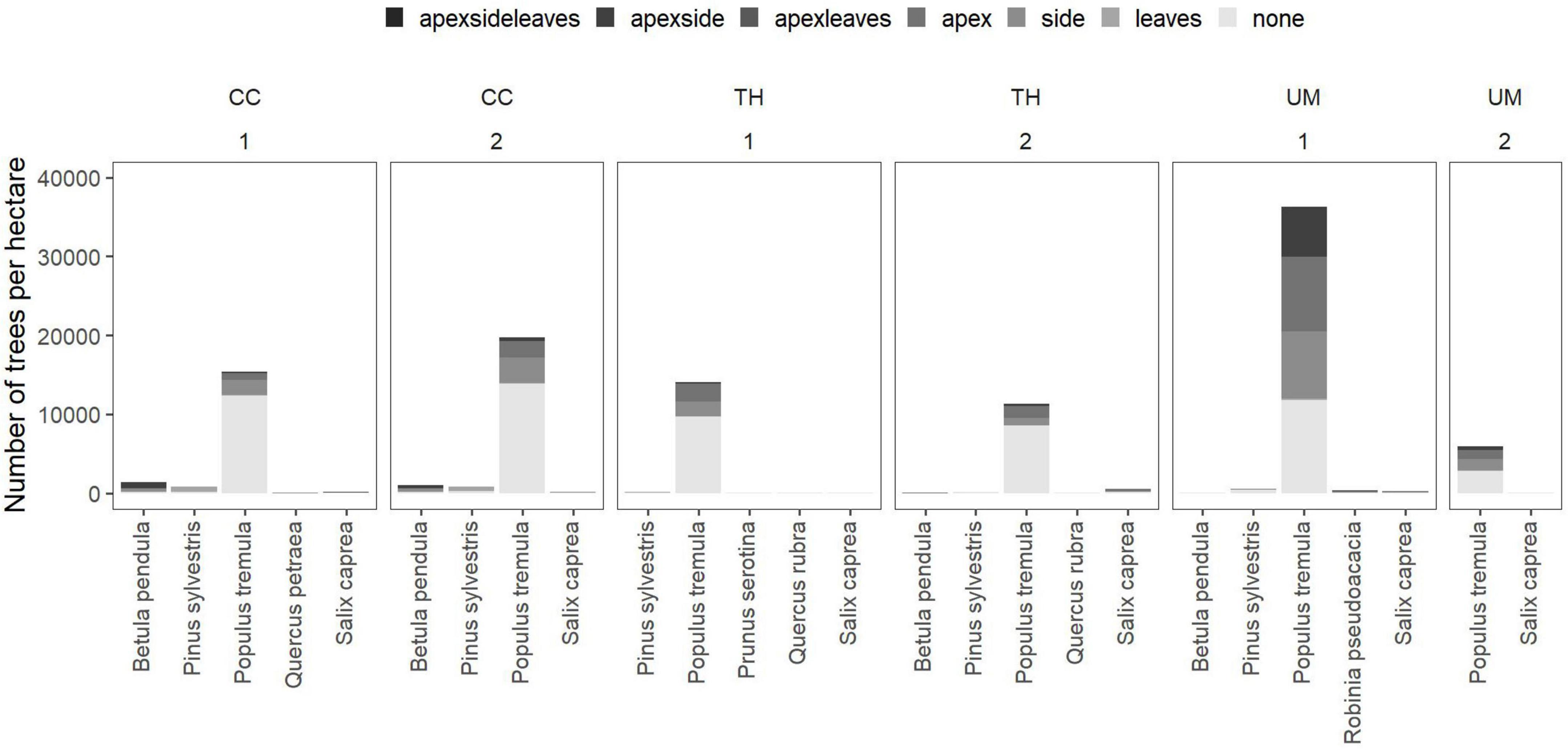
Figure 3. Distribution of vitality and damages for tree species occuring per treatment (CC, clearcut; TH, thinned; UM, unmanaged) and plot (two plots per treatment represented by the number below the treatment name) recorded in the year 2021. Colors indicate the damage recorded on each tree individual.
Except for Pinus sylvestris, after the first growing season, the majority of saplings were alive (Figure 2). Few individuals of Populus tremula showed signs of damages at the apex and side shoots. After two growing seasons, the proportion of damaged trees increased. In addition, multiple damages on the same tree individuals were recorded for some trees (Figure 3). Pine trees showed the highest mortality. In particular, planted seedlings mostly died (23–40% survived) approximately 1 year after planting while naturally rejuvenating pine trees mostly survived (91–100% survived) (Table 2).
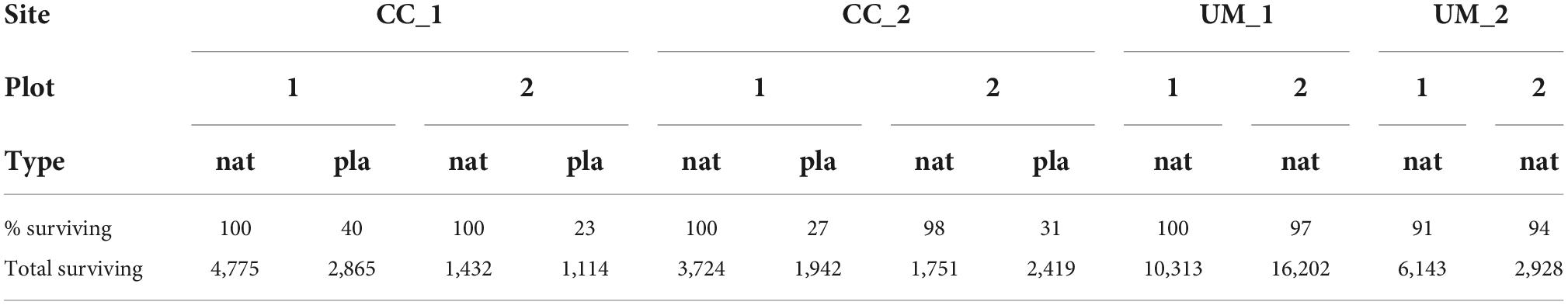
Table 2. Proportion of surviving individuals of Pinus sylvestris in the clearcut (site = CC) and the unmanaged (site = UM) sites comprising naturally established (type = nat) and artificially planted (type = pla) trees, and total number of surviving Pinus sylvestris.
On the actively restored plots, the total number of surviving pine trees was higher for pines that were established naturally compared to the planted ones. On the unmanaged sites, where no pine trees were artificially planted, the number of living pines varied between 2,928 and 16,202 trees per hectare (Figure 4).
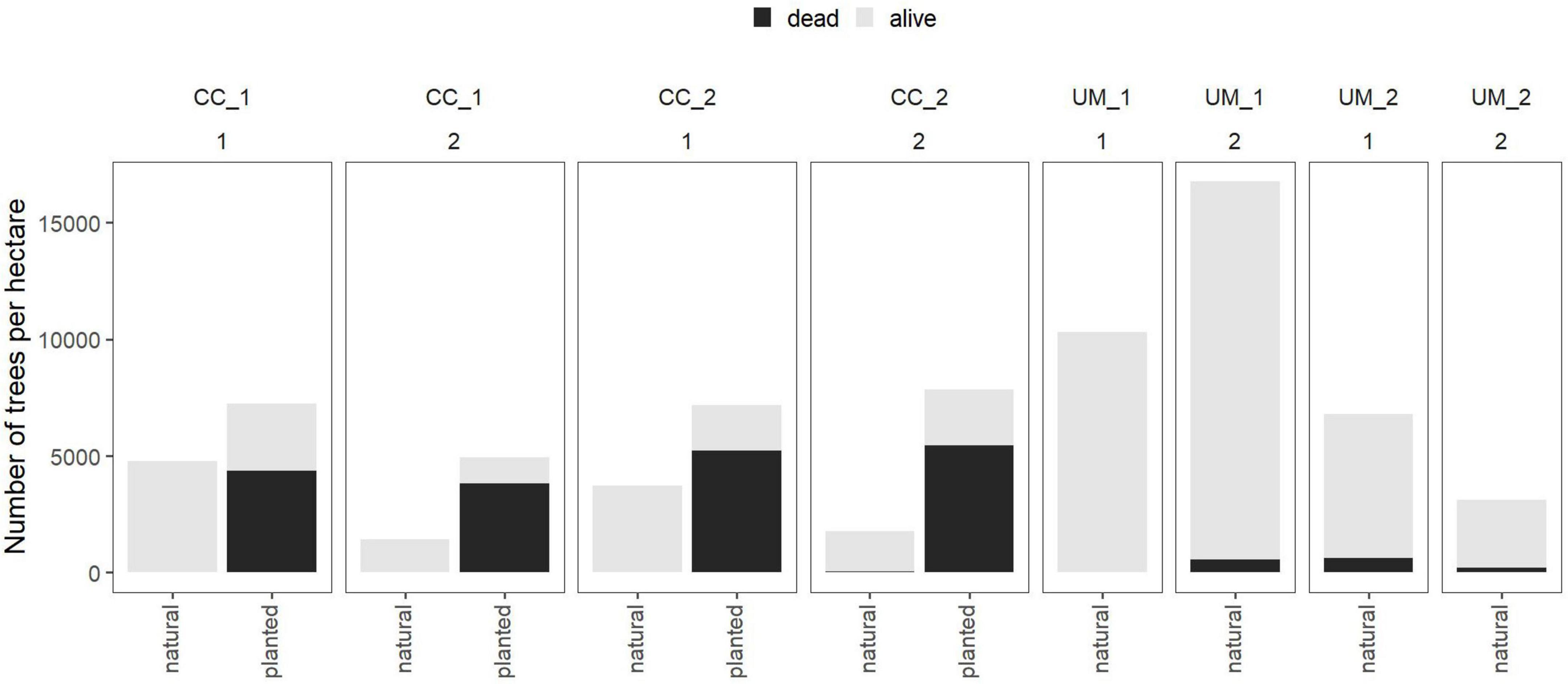
Figure 4. Number of individuals of actively planted and naturally rejuvenating Pinus sylvestris counted per treatment (CC, clearcut; UM, unmanaged) and plot (two plots per treatment represented by the number below the treatment name) recorded in the year 2019 stacked as dead (black) and living (gray) trees.
Species richness varied between four to five species on the clearcuts, including sites where Pinus sylvestris and Quercus rubra were planted. Quercus rubra, representing a non-native species in the region, was the only tree species that did not rejuvenate naturally. On the unmanaged sites, two to five species were recorded, and five on the thinned sites. In the first sampling season, unmanaged sites showed a slightly higher Shannon Diversity Index value compared to the post-fire salvage logged sites (Figure 5A). After the second vegetation season, the increasing dominance of Populus tremula and the lower abundance of other species in the plots resulted in a lower Shannon Diversity Index value (Figure 5B). At this time, the value was highest in the clearcut and lowest in the unmanaged site though this was substantially influenced by planted non-native trees in the clearcuts.
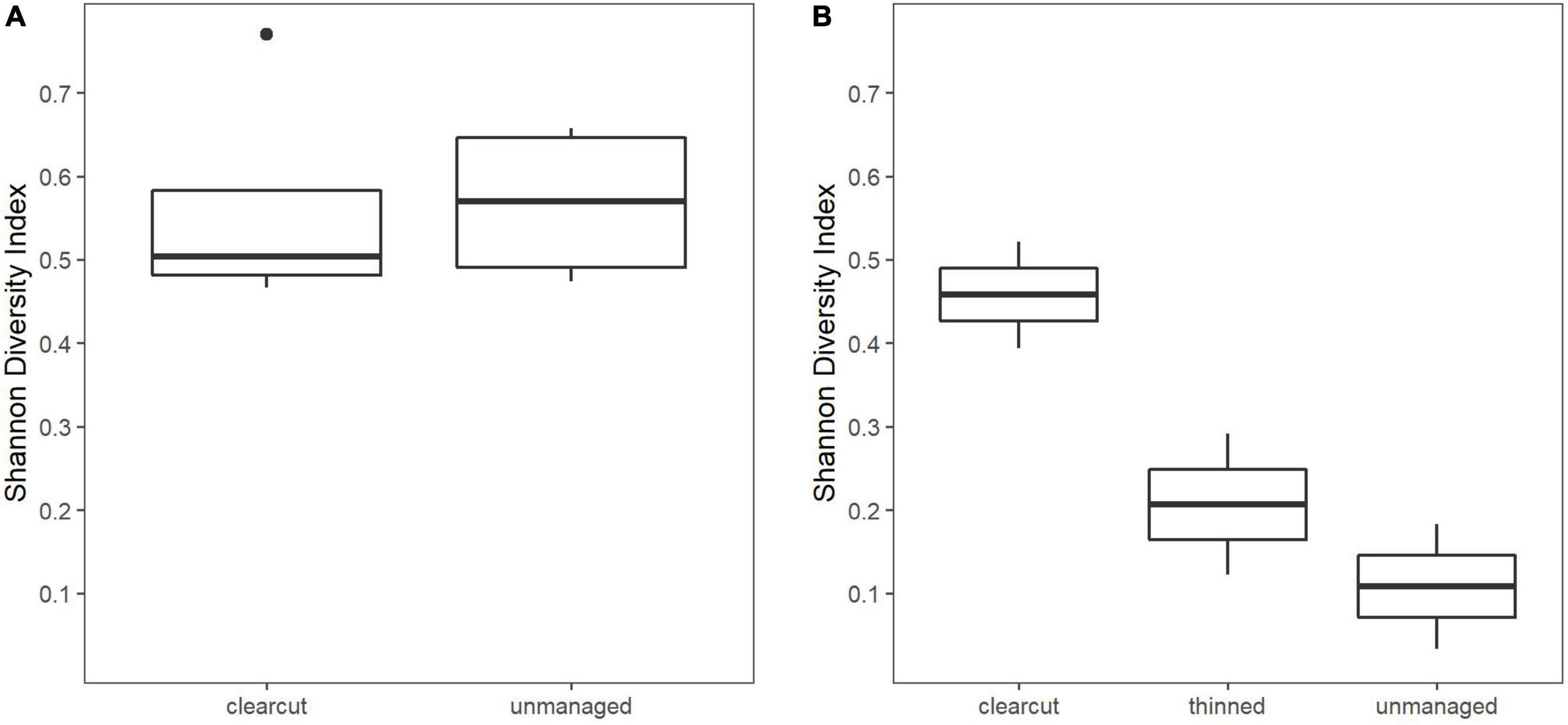
Figure 5. Shannon diversity index for tree rejuvenation: (A) one growing season; (B) two growing seasons after the fire.
In both sampling years, Tmax of the warmest days and VPDmax during the driest days differed between the treatments (p < 0.01). In 2019, Tmax at 1.3 m was significantly higher on the clearcuts compared to thinned (p = 0.04) or unmanaged (p = 0.013) sites, while thinned and unmanaged sites did not significantly differ from each other (p = 1) (Figure 6A, top). At 0.1 m, compared to unmanaged sites Tmax was significantly higher on the clearcuts (p < 0.001) and thinned sites (p < 0.001), while Tmax did not differ between the clearcut and the thinned sites (p = 0.65) (Figure 6A, bottom). Maximum VPD was highest on the clearcut and significantly lower on the thinned sites (p = 0.024). The unmanaged stand showed the lowest VPDmax. It was significantly lower compared to the thinned (p = 0.011) and the clearcut sites (p < 0.001) (Figure 6B).
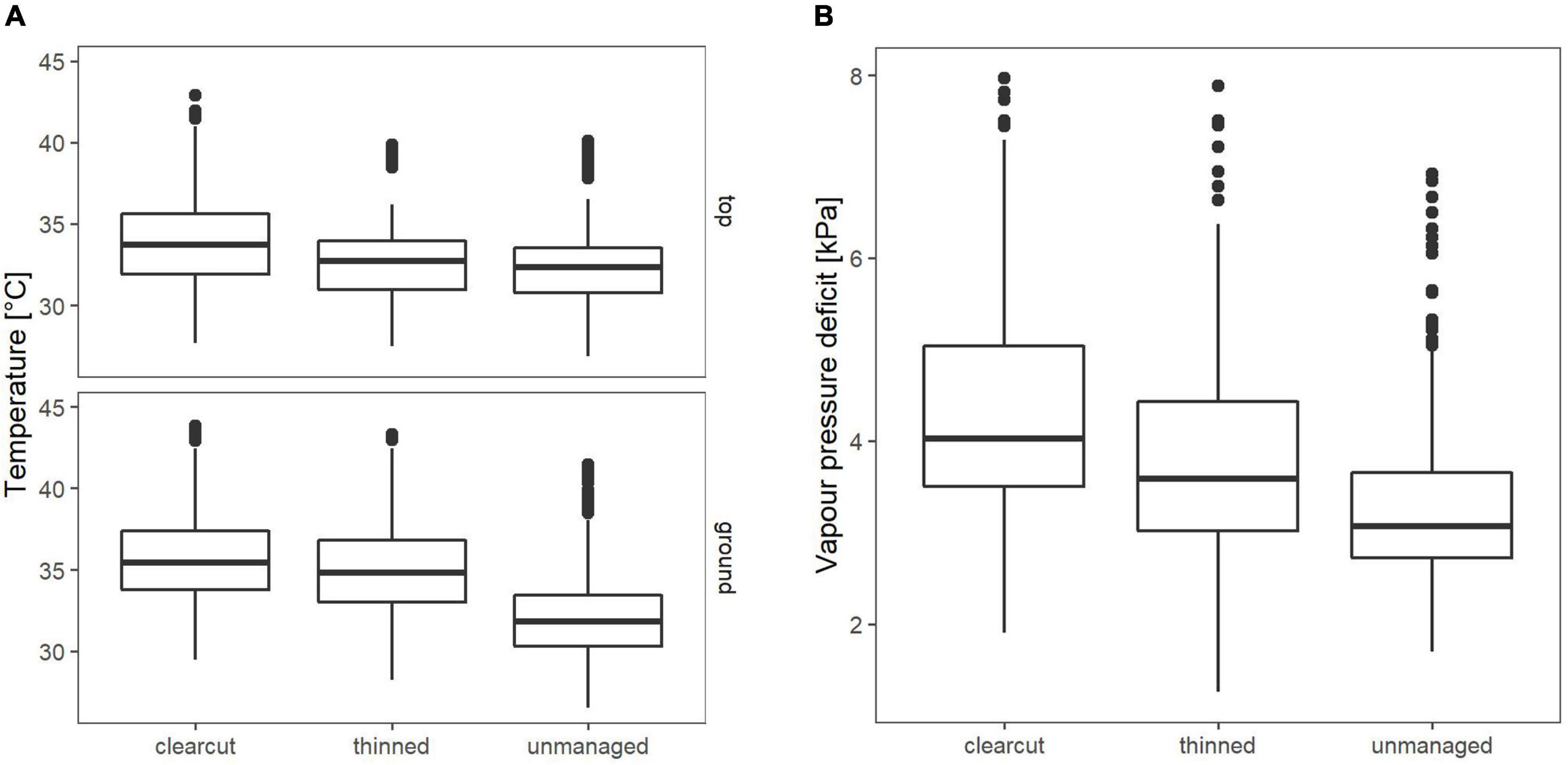
Figure 6. (A) Mean maximum temperature on hottest days (n = 31 days) in 2019 (top = 1.3 m; ground = 0.1 m). (B) Mean VPD on driest days (n = 31 days) in 2019, measured 0.1 m above ground.
In 2021, Tmax at 1.3 m was highest on the clearcut and significantly lower on unmanaged sites (p = 0.0025). There was no significant difference between the clearcut and the thinned sites (Figure 7A, top). At 0.1 m, Tmax was highest on the clearcuts and thinned sites, which did not significantly differ from each other. Unmanaged sites were significantly cooler than thinned (p = 0.00028) and clearcut (p = 0.0045) sites (Figure 7A, bottom). Maximum VPD was lowest on unmanaged sites and significantly higher on thinned and clearcut sites (p < 0.001), which did not differ from each other (Figure 7B).
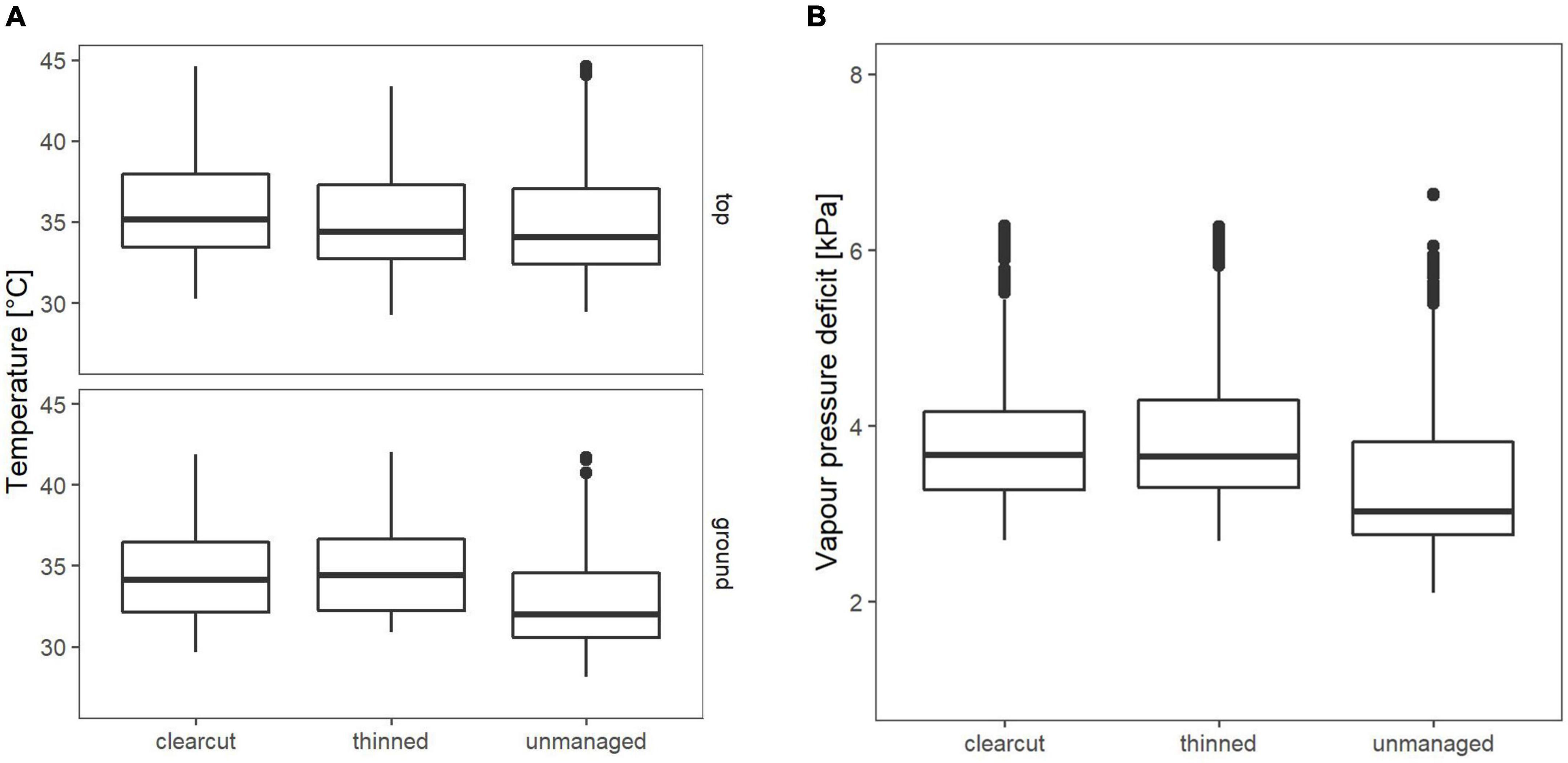
Figure 7. (A) Mean maximum temperature on hottest days (n = 22 days) in 2021 (top = 1.3 m; ground = 0.1 m). (B) Mean VPD on driest days (n = 22 days) in 2021, measured 0.1 m above ground.
Discussion
Natural succession
The results of this study show that 1 year after a forest fire, the pioneer tree species Populus tremula showed strong spontaneous colonization at all sites. In contrast, the majority of planted Pinus sylvestris seedlings had died on the actively restored sites. In the clear-cut plots, the extreme microclimatic conditions exposed the young trees to additional heat and drought stress, while the retention of deadwood buffered both temperature and vapor pressure deficit. Three years after the fire, Populus tremula was the dominant tree species on all plots, with the highest abundance where the dead or dying overstorey was not removed. Betula pendula, Salix caprea, and Pinus sylvestris were also present, but with lower abundance. Strong regeneration of Populus tremula, Salix caprea, Betula pendula, and Pinus sylvestris, and less frequently also Larix decidua in the first 6 years after the fire was also found in burned pine forests in the Czech Republic (Adámek et al., 2018). There, the number of individuals of Populus tremula and Salix caprea decreased in later stages, probably due to unfavorable habitat conditions. The establishment of pioneer tree species is not only relevant for the reforestation of the forest after a fire, but especially the ability of aspen to resprout after disturbance can contribute to a faster regeneration after future disturbances such as subsequent forest fires (Porter et al., 2022).
Although additional trees have been planted on the clearcut sites studied in 2019, the total abundance of naturally established trees clearly exceeded the trees counted on the post-fire salvage logged and actively restored sites (a total of 11,682 individuals that established naturally were counted compared to 8,340 planted trees).
Fire causes the combustion of organic layers covering the soil and can change the soil structure and chemical properties (Ne’eman et al., 1997; Úbeda and Outeiro, 2009). As fire temperatures rise, soil microorganisms, invertebrates, and seeds existing in the upper soil layer are more severely affected (Ne’eman et al., 1997). This might possibly result in a reduction in soil productivity and the survival chances of rejuvenating plants as well as an increase in carbon emissions from soil. The loss of organic matter and the destruction of soil structures can therefore be the cause of a decreased soil water retention capacity after fire (Úbeda and Outeiro, 2009; Pereira et al., 2018). Hence, whether for economic or for ecological reasons, a main goal in the management of the post-fire rejuvenation is a rapid reestablishment of soil cover (Mauri and Pons, 2019).
Active restoration
Reforestation by seeding or planting is commonly applied in post-fire forest restoration projects (Mazza, 2007; Moreira et al., 2009; Beghin et al., 2010; Morat, 2019), especially in forests where timber production is a main management objective (Franklin and Agee, 2003; Sessions et al., 2004; Ürker et al., 2018). In many countries, forest owners are obliged by law to reforest burned forests by replanting which can be subsidized by institutional funding programs (Mavsar et al., 2012; Ryu et al., 2017). Artificial reforestation is generally recommended in stands where the existing species are not fire-adapted, where no reproductive material has survived the fire and natural regeneration is not expected, or when forest species conversion is planned (Ne’eman et al., 1997; Beschta et al., 2004; Moreira et al., 2012b; Vallejo et al., 2012b; Ryu et al., 2017; Morat, 2019). The common perception is that planting on degraded areas is essential in order to ensure sufficient post-fire regeneration (Moreira et al., 2009; Vallejo et al., 2012a; Tinya et al., 2020). It is claimed that natural regeneration is unsuccessful, where post-fire salvage logging was applied and that artificial regeneration on these stands might be indispensable but as shown in the Italien Alps natural restoration is an effective alternative (Beghin et al., 2010). It is also argued that in ecosystems that have been modified by human interventions over a long time, natural succession is not sufficient to ensure site regeneration and might therefore not comply with the restoration objectives (Ne’eman et al., 1997; Pausas et al., 2004). However, on sites where sufficient natural regeneration is expected, artificial reforestation measures are generally not recommended (Ne’eman et al., 1997; Beschta et al., 2004; Donato et al., 2006; Mavsar et al., 2012; Ibisch, 2019).
Nevertheless, reforestation is often applied before the ecosystem was given the chance to regenerate by itself, even when natural regeneration would suffice (Moreira et al., 2009). This also happened at the clearcut and replanted sites investigated in the frame of this study and the PYROPHOB project. Human intervention, such as post-fire salvage logging, aiming to speed up reforestation might impair natural regeneration processes, negatively impact biodiversity of usually slowly regenerating forests, promote exotic alien species or simply recreate the fire-prone stand condition prevailing before the forest fire (Franklin and Agee, 2003; Lindenmayer and Noss, 2006). Our findings show, that in the studied area, sufficient natural regeneration is taking place, even on clearcuts despite the lower abundance compared to the unmanaged sites. There is no need for active restoration in the study area, and the secondary site degradation due to post-fire salvage logging on the cleared and thinned sites could have been prevented by allowing natural regeneration.
Planting is usually conducted using saplings that were grown in tree nurseries, giving them a developmental advantage over seedlings germinating on the site. Due to this advantage, planted trees are claimed to have higher survival chances than natural regeneration, especially given the harsh post-disturbance conditions (Pausas et al., 2004; Newton et al., 2006). However, Alanís-Rodríguez et al. (2015) have observed a survival rate of 35% of the planted individuals even after costly replacement of dead seedlings over the first years after planting. In our case, the survival rate of pine was even lower. Nevertheless, pine saplings were planted repeatedly after the fire and subsequent clearcutting, from 2019 to 2021. While Scots pine plantations have always been established and favored as economically viable tree species by many forest owners in Brandenburg, the growing conditions seem to have become unsuitable in the climatically extreme years 2019–2021, which might represent a new average within a few years. Based on our findings, we cannot recommend establishing or replanting Scots pine stands. This type of forest plantation has already been described as “neither environmental-friendly nor climate smart, given their enhancement of climate-warming, low climate change mitigation potential, and negative effect on groundwater recharge” (Leuschner et al., 2022).
In a study conducted by Moreira et al. (2009) in the first regeneration phase after a forest fire in central Portugal, survival and growth rates of naturally regenerating oak (Quercus faginea) and ash (Fraxinus angustifolia) resprouters were higher than when the same species were planted. Tinya et al. (2020) obtained similar results with naturally regenerating seedlings (Quercus petraea, Carpinus betulus, and Fraxinus ornus) in a mountain forest in Hungary. In a study conducted by Donato et al. (2006) in regenerating Douglas-fir (Pseudotsuga menziesii) stands in Oregon, USA, natural conifer regeneration on sites was generally abundant and sufficient for the reestablishment of the stand according to regional standards, implying that active reforestation may not be necessary while post-fire salvage logging killed 71% of the natural regeneration and increased fine fuels on the site that added to greater subsequent fire risks. Ne’eman et al. (1997) made similar observations on regenerating Pinus halepensis stands in the first year after a forest fire on Mount Carmel in Israel. In a post-fire remote sensing-based study conducted by Vlassova and Pérez-Cabello (2016) in Northeast Spain, various regeneration mechanisms of the naturally regenerating species were sufficient to ensure the recovery of the future forest stand regardless of the applied treatment method. Additionally, it was argued that planting might have undesired effects on the stands’ native biodiversity and genetic composition (Ne’eman et al., 1997; Mavsar et al., 2012). In a study by Schmidt and Wichmann (2000), both the mean number of species and diversity in the tree and herb layers were lower in the planted plots than on the naturally rejuvenated plots. Although Ürker et al. (2018) measured a more successful establishment of planted saplings over natural regeneration, they also acknowledged the negative effects of pine plantations on various ecosystem services like habitat provision and biodiversity maintenance and concluded that seeding or natural regeneration should be preferred over plantations in post-fire restoration.
In many countries, reforestation by planting after site preparation is considered as the most successful reforestation technique and is often preferred over seeding (Espelta et al., 2003; Pausas et al., 2004; Ginzburg and Steinberger, 2012; Alanís-Rodríguez et al., 2015). This could not be confirmed by our study. However, it is important to ensure that sufficient seed trees are available, especially pioneer tree species that can recolonize degraded forests rapidly. It is therefore advisable to conserve and promote these species in regions with large monocultures that are, or can become, calamity prone areas. In addition, they play an important role in conversion of monocultures to far less flammable forests.
Direct seeding is another possible reforestation strategy used in active restoration. It is cheaper and considered to have a lower environmental impact than planting. Broadcast seeding can also be conducted from the air, representing an option for the restoration of remote areas (Lamb and Gilmour, 2003; Pausas et al., 2004; Mansourian et al., 2005; Vallejo et al., 2009). Pausas et al. (2004) reported research performed after a wildfire in Catalonia where a relatively successful germination rate (circa 5%) was obtained from an aerial seeding. It is also argued that seedlings which directly regenerated from seeds and were exposed to the stands’ conditions in their early development, will be better acclimatized and less susceptible to stand conditions than nursery-grown and transplanted seedlings (Vallejo et al., 2009). However, establishment rates of seeds are often lower comparing to planted seedlings, whether due to unsuitable site conditions for germination, extreme weather events, competition through fast growing ground vegetation or due to seed predation by birds and rodents (Espelta et al., 2003; Lamb and Gilmour, 2003; Pausas et al., 2004; Mansourian et al., 2005). Espelta et al. (2003) reported higher seedling establishment rates on planted plots in comparison to plots where seeding was conducted. In a research project by Pausas et al. (2004) after a wildfire in eastern Spain, predation rates of pine (Pinus halepensis) seeds applied by aerial seeding were high with more than 80% predated during the first 6 months after seeding. Restoration by seeding has eventually failed, as germination was not observed in any of the examined plots. In a burned pine (Pinus halepensis) forest in Greece, Spanos et al. (2010) observed the same woody species composition but a lower seedling establishment on the seeded plots compared to the non-seeded, naturally regenerating control. Thus, natural regeneration following fires, without intervention, may be preferable.
Post-fire salvage logging
Post-fire salvage logging is the most common post-fire activity, and often the first measure applied after fire in various forest types around the world (Ne’eman et al., 1997; McIver and Starr, 2001; Beschta et al., 2004; Lindenmayer and Noss, 2006; Peterson et al., 2009; Vallejo et al., 2012b; Ascoli et al., 2013; Leverkus et al., 2014). This strategy seems to be especially attractive for private forest owners dependent on the revenues from charred wood and on the subsidies often granted by the state for post-fire salvage logging operations (Vallauri, 2005; Vallejo et al., 2012b). However, the harvested charred logs have often only very little economic value, sometimes not even covering the operation costs (Ibisch, 2019). This was the case in post-fire studies conducted in Mediterranean forests in Italy and Spain, where the high costs for post-fire logging resulted in no economic benefits (Beghin et al., 2010; Leverkus et al., 2012). Nevertheless, economic output will depend on the level of damage caused to the trees, harvesting technologies, and market conditions (Pereira et al., 2018). Further arguments in favor of post-fire salvage logging are fuel reduction on the forest floor reducing the risk of future fires, the prevention of pest outbreaks, the safety of forest visitors, and the facilitation of further restoration measures (like planting) planned for the stand (Ne’eman et al., 1997; McIver and Starr, 2001; Lindenmayer and Noss, 2006; Beghin et al., 2010; Castro et al., 2011; Leverkus et al., 2012; Ascoli et al., 2013). Additionally, for certain tree species (e.g., pines, oaks, beeches), the exposure of the mineral soil, soil scarification through the logging operations and the improvement in light conditions through tree removal can support a more successful germination and seedling establishment (McIver and Starr, 2001; Sessions et al., 2004; Beghin et al., 2010; Ascoli et al., 2013).
On the other hand, salvage logging might cause more disturbance to the ecosystem than the fire itself (Lindenmayer and Noss, 2006). Arguments against salvage logging include the risk of soil compaction and erosion through logging operations and additional road building, an increase in runoff and sediment loads, the damage to ground vegetation and especially natural rejuvenation, the modification of hydrological cycles, the removal of biological legacies responsible for the improvement of microclimatic conditions and habitat provision for regenerating organisms and hence, the reduction in structural and species diversity (McIver and Starr, 2001; Beschta et al., 2004; DellaSala et al., 2006; Lindenmayer and Noss, 2006; Lindenmayer et al., 2008; Robichaud et al., 2011; Ibisch, 2019). In studies conducted in coniferous mountain forests in the USA and Canada, logging operations have generated soil compaction, reduced infiltration, and increased runoff rates and sedimentation in recently burned areas. Logged areas also experienced a slower recovery rate of regenerating vegetation compared to the unlogged control (Donato et al., 2006; Robichaud et al., 2011; Wagenbrenner et al., 2016). Beghin et al. (2010) showed that even when logging was conducted 4 years after the fire, it contributed to a prolonged soil disturbance. Other post-fire research projects have obtained similar results of increased soil compaction and erosion and a decrease in seedling recruitment and soil cover following post-fire logging operations (Inbar et al., 1997; Martínez-Sánchez et al., 1999; Spanos et al., 2010; de las Heras et al., 2012; Marañón-Jiménez et al., 2013; Moya et al., 2015; Malvar et al., 2017; Urretavizcaya and Defossé, 2019). Further studies have observed an increased fire risk due to fine woody debris left on the site after logging (Ne’eman et al., 1997; Donato et al., 2006), a reduction in soil nutrients (Merino et al., 2005), a decrease in seedling height growth (Martínez-Sánchez et al., 1999), and a decrease in species richness and diversity (Leverkus et al., 2014). In a study comparing different treatment methods in a mountain forest in Hungary, large cleared areas also experienced a limited seed dispersal, apparently due to the lack of seed sources on the stand as well as the lack of habitat for seed-dispersing animals (Tinya et al., 2020). The microclimatic conditions induced by salvage logging, such as decreased soil moisture and increased surface and soil temperature, have been commonly observed as unfavorable for woody species regeneration (Castro et al., 2011; Ginzburg and Steinberger, 2012; Marcolin et al., 2019; Tinya et al., 2020) but favorable for competing ground vegetation (Sessions et al., 2004; Beghin et al., 2010). In combination, this can result in seedling dehydration and mortality.
A major motivation for forest owners to clear the site early after the fire is the risk of losing timber revenues for rotten timber (Moreira and Vallejo, 2009; Robichaud et al., 2011; Vallejo et al., 2012b; Pereira et al., 2018). Sessions et al. (2004) estimated a volume loss of 22% of the still merchantable logs due to wood deterioration in the first years after a large forest fire in Oregon. Nevertheless, when applied immediately after the fire, logging operations might have a greater negative effect on the exposed vulnerable soil compared to a later implementation (Ne’eman et al., 1997; Mazza, 2007; Peterson et al., 2009; Wagenbrenner et al., 2016). Although soil degradation can be reduced or even avoided by certain harvesting techniques (e.g., cable systems, skyline yarding, helicopter), these are usually more expensive and their cost-effectiveness is dependent on the timber price (McIver and Starr, 2001; Vallejo et al., 2012b; Leverkus et al., 2018; Marcolin et al., 2019). On the other hand, the later post-fire logging takes place, the higher is the potential damage to tree rejuvenation already established on the stand (Martínez-Sánchez et al., 1999; McIver and Starr, 2001; Lindenmayer and Noss, 2006; Castro et al., 2011; Fischer and Fischer, 2012). According to Franklin and Agee (2003), post-fire logging generally does not facilitate ecological ecosystem recovery and is therefore often inappropriate for post-fire forest restoration.
Deadwood retention
Passive restoration favors natural succession and prescribes no artificial intervention in the natural processes of the ecosystem. Apart from natural rejuvenation, this also includes leaving burned residual wood on site, regardless of its state, size, or position. Organic materials remaining from the pre-disturbed stand are also called biological legacies, defined by Franklin et al. (2000) as “…the organisms, organic materials, and organically generated environmental patterns that persist through a disturbance and are incorporated into the recovering ecosystem.” In burned forests, they are represented by surviving living trees, standing charred trees, standing dead snags as well as lying deadwood in different sizes and degrees of combustion (Franklin and Agee, 2003; Noss et al., 2006). Post-fire ecosystems are rich in biological legacies that are considered essential for post-fire recovery processes, especially in the initial regeneration phase (Franklin and Agee, 2003; Beschta et al., 2004; Noss et al., 2006; Leverkus et al., 2014). Many researchers have described the positive effects of biological legacies on regenerating ecosystems (DellaSala and Hanson, 2015). One major benefit provided through the sheltering effect of both standing and lying residual deadwood is the amelioration of the stands’ microclimatic conditions—keeping temperatures stable, protecting the soil and vegetation from direct solar radiation, and retaining soil moisture levels more favorable for germination and for the establishment of natural regeneration (Martínez-Sánchez et al., 1999; Pausas et al., 2004; Moreira and Vallejo, 2009; Peterson et al., 2009; Beghin et al., 2010; Castro et al., 2011; Marañón-Jiménez et al., 2013; Moya et al., 2015; Ibisch, 2019; Marcolin et al., 2019).
Our findings show that the retention of the dead trees provides shade and reduces both surface and ambient temperature. In turn, the vapor pressure deficit is reduced, lowering additional water losses through evapotranspiration and desiccation of plants. As water presents one of the most limiting resources for tree growth in the study region and these conditions are likely to worsen in the future, it is becoming ever more important to prevent additional water loss. Our data confirm the microclimatic benefits provided by deadwood left on site. Apart from contributions to cooling and moisture retention in the vegetation period, it can also be beneficial under winter conditions, as it absorbs heat radiation and promotes snow melting, providing adequate growth conditions for the seedlings in its proximity (Castro et al., 2011). Surviving charred vegetation with undamaged root system supports soil stabilization while coarse and fine woody debris protect the soil from compaction and erosion (Marañón-Jiménez et al., 2013; Leverkus et al., 2014; Vlassova and Pérez-Cabello, 2016; Marcolin et al., 2019). Surviving living trees also act as seed sources providing reproductive material for the regeneration onsite, while both living trees and dead snags can be used as perches for seed-dispersing species and thereby help accelerate natural succession (Franklin and Agee, 2003; Whisenant, 2005; Moreira and Vallejo, 2009; Marzano et al., 2013). Additionally, coarse woody debris and snags provide habitat for many post-disturbance specialized species, like cavity-living birds or wood decomposing insects, thereby increasing species diversity and often forming regional diversity hotspots. Increased species diversity is also promoted by increased structural diversity and heterogeneous light conditions on uncleared stands (Beschta et al., 2004; Mazza, 2007; Beghin et al., 2010; Marzano et al., 2013; Leverkus et al., 2014; Marcolin et al., 2019). Another benefit is the retention of nutrients stored in the wood and their later provision through decomposition onsite (Peterson et al., 2009; Gustafsson et al., 2012; Marañón-Jiménez et al., 2013; Leverkus et al., 2014; Vlassova and Pérez-Cabello, 2016). Uncleared stands also act as carbon pools (Gustafsson et al., 2012; Leverkus et al., 2018). Finally, it is argued that burned trees left on site do not increase future fire risk, as the snags slowly decay and are gradually integrated back into the soil in comparison to logging treatments where large amounts of flammable slash are often left on the ground (Donato et al., 2006).
Various study projects investigating the effects of different post-fire treatments have measured higher rates of vegetation recovery, natural seedlings emergence and abundance (Beghin et al., 2010; Castro et al., 2011; Marañón-Jiménez et al., 2013; Marzano et al., 2013; Chen et al., 2014; Leverkus et al., 2014; Moya et al., 2015; Vlassova and Pérez-Cabello, 2016), growth rates (Martínez-Sánchez et al., 1999; Marañón-Jiménez et al., 2013; Moya et al., 2015), species richness and diversity (Marzano et al., 2013; Leverkus et al., 2014), soil moisture, and soil microbial activity (Ginzburg and Steinberger, 2012), as well as increased structural diversity (Beghin et al., 2010), when deadwood remained on the regenerating stand in comparison to cleared stands. These positive effects were attributed to the ecological functions of deadwood residuals. Besides the protective effects of deadwood on plant desiccation by providing more favorable site conditions for tree growth, deadwood significantly reduces herbivorous browsing through increased surface roughness (Marangon et al., 2022).
Beschta et al. (2004) conducted a literature review investigating the ecological effects of common post-fire treatments and concluded that post-fire forest restoration aiming to maintain the stands’ ecological functions should include the retention of large, living or dead, standing or lying trees. Regarding the concern that deadwood could increase stand flammability, they argue that woody debris is principally not responsible for carrying wildland fire but rather for the finer fuel sources such as grasses and tree foliage. Although Castro et al. (2011) measured no difference in seedling density between logged and unlogged stands, they concluded that the passive method should be preferred since the “no-intervention” plots seemed to enjoy an improvement in microclimatic conditions without suffering the disturbing effects of logging related heavy machinery observed on the logged plots. In addition to the better height growth and seedling recruitment rates, Moya et al. (2015) also observed the emergence of other tree species only in the “no-intervention” plots in a Mediterranean region, generating a higher species diversity in those stands. In a study conducted by Marzano et al. (2013), coniferous species (Pinus sylvestris and Larix decidua) considered as pioneers did not benefit from the exposed conditions on the salvaged areas and performed better in the proximity of deadwood. They concluded that there is a higher probability for seedling survival and establishment in the presence of at least one deadwood element within one meter from the regeneration. Once biological legacies have been removed from the stand, it takes a long time until they are replaced and their functions are recovered (Franklin and Agee, 2003).
Logging operations also increase the risk of further soil degradation as well as potentially damaging surviving or emerging natural regeneration (Lindenmayer and Noss, 2006; Marzano et al., 2013; Moya et al., 2015; Vlassova and Pérez-Cabello, 2016; Marcolin et al., 2019). In a study conducted in a burned pine stand (Pinus halepensis) in Spain, wood removal was shown to generate high seedling mortality while decreasing seedling growth due to mechanical damage and the exposed site conditions created by the logging operations (Martínez-Sánchez et al., 1999). Besides the physical damage, negative effects include the alteration of soil properties, water, carbon, and nutrient cycles, the facilitation of stand colonization by invasive species, the reduction in connectivity between non-burned forest areas, the homogenization of the forest structure, and the loss of structural diversity as well as a change in the stands’ plant and animal species composition (Beschta et al., 2004; Lindenmayer and Noss, 2006; Lindenmayer et al., 2008Peterson et al., 2009; Vlassova and Pérez-Cabello, 2016). Beghin et al. (2010) reported that while the passive strategy applied in their study was successful, the active measures traditionally prescribed by regional authorities were shown to alter the natural forest structure and slow down its regeneration. Others claim that the removal of biological legacies, especially of the surviving, still living trees, could potentially decrease the natural regeneration capacity of the stand and slow down the recovery of ecosystem functions and service provision (Ne’eman et al., 1997; Beschta et al., 2004; Beghin et al., 2010; Fischer and Fischer, 2012; DellaSala, 2020).
Conclusion
Questions about how to manage forests after such an event become more pressing, especially with an increasing fire risk under climate change. Catastrophic events and disturbances in timber plantations such as forest fires in artificial Scots pine monocultures can also help forests to develop into more structurally diverse, broad-leaved or mixed stands compared to actively restored sites where dead trees are removed and economically targeted trees are planted simultaneously. Even in planted Scots pine monocultures that were affected by a forest fire, there is a potential for natural succession, which supports the development into more broad-leaved stands. At the same time, the conditions for the reestablishment of coniferous plantations are becoming ever more severe, and effective active restoration becomes more difficult than in the past. Maintaining and developing a sufficient seed tree abundance, especially of pioneer tree species, can contribute to the forest resilience and help reestablish and maintain ecological functions and services after disturbance. In addition, the ability of certain tree species to resprout adds to the resilience of ecosystems and promotes their recovery from stress and disturbances. The development of the forest ecosystem under unfavorable growing conditions during extreme years with exceptionally hot temperatures and ongoing water deficits shows that the temperate forests of Central Europe still hold a certain inherent adaptive capacity that might be helpful in coping with the upcoming impacts of climate change. This does not mean, of course, that drastic changes in vegetation can be ruled out if the climate crisis continues to intensify as unchecked as it does at present.
Data availability statement
The datasets presented in this study can be found in online repositories. The name of the repository: Zenodo; https://zenodo.org/record/7371511#.Y4SRd32ZOUk.
Author contributions
JB did the conceptualization of the study and collected the microclimatic data, computed all data analyses, and wrote the first manuscript draft. FS, AG, and SG collected the rejuvenation data in the field in the frame of their bachelor thesis. PI was responsible for the conceptualization and supervision of the study. All authors contributed to finalizing the manuscript.
Funding
This study was partly conducted in the frame of the PYROPHOB project funded by the forest-climate funds provided by the German Federal Ministry of Food and Agriculture and the Federal Ministry for the Environment, Nature Conservation, Nuclear Safety and Consumer Protection (Grant No. 2219WK50A4).
Acknowledgments
We thank the forest owner for providing the research sites. PI supervised the study in the framework of his long-term research program facilitated by the research professorship “Ecosystem-based sustainable development” (since 2015) granted by the Eberswalde University for Sustainable Development. We thank all students and assistances including Maren Flagmeier for supporting the data collection in the field.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Adámek, M., Jankovská, Z., Hadincová, V., Kula, E., and Wild, J. (2018). Drivers of forest fire occurrence in the cultural landscape of Central Europe. Landscape Ecol. 33, 2031–2045. doi: 10.1007/s10980-018-0712-2
Alanís-Rodríguez, E., Valdecantos, A., Jiménez-Pérez, J., Rubio-Camacho, E. A., Yerena-Yamallel, J. I., and González-Tagle, M. A. (2015). Post-fire ecological restoration of a mixed Pinus-Quercus forest in northeastern Mexico. Rev. Chapingo 21, 157–170. doi: 10.5154/r.rchscfa.2014.07.031
Ascoli, D., Castagneri, D., Valsecchi, C., Conedera, M., and Bovio, G. (2013). Post-fire restoration of beech stands in the Southern Alps by natural regeneration. Ecol. Eng. 54, 210–217. doi: 10.1016/j.ecoleng.2013.01.032
Beghin, R., Lingua, E., Garbarino, M., Lonati, M., Bovio, G., Motta, R., et al. (2010). Pinus sylvestris forest regeneration under different post-fire restoration practices in the northwestern Italian Alps. Ecol. Eng. 36, 1365–1372. doi: 10.1016/j.ecoleng.2010.06.014
Beschta, R. L., Rhodes, J. J., Kauffman, J. B., Gresswell, R. E., Minshall, G. W., Karr, J. R., et al. (2004). Postfire management on forested public lands of the western united states. Conserv. Biol. 18, 957–967. doi: 10.1111/j.1523-1739.2004.00495.x
Blumrö,der, J. S., May, F., Härdtle, W., and Ibisch, P. L. (2021). Forestry contributed to warming of forest ecosystems in northern Germany during the extreme summers of 2018 and 2019. Ecol. Solut. Evid. 2, 1–14. doi: 10.1002/2688-8319.12087
Bowman, D. M. J. S., Kolden, C. A., Abatzoglou, J. T., Johnston, F. H., van der Werf, G. R., and Flannigan, M. (2020). Vegetation fires in the anthropocene. Nat. Rev. Earth Environ. 1, 500–515. doi: 10.1038/s43017-020-0085-3
Castro, J., Allen, C. D., Molina-Morales, M., Marañón-Jiménez, S., Sánchez-Miranda, Á, and Zamora, R. (2011). Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment. Restor. Ecol. 19, 537–544. doi: 10.1111/j.1526-100X.2009.00619.x
Chen, W., Moriya, K., Sakai, T., Koyama, L., and Cao, C. (2014). Monitoring of post-fire forest recovery under different restoration modes based on time series Landsat data. Eur. J. Remote Sens. 47, 153–168. doi: 10.5721/EuJRS20144710
de las Heras, J., Moya, D., Vega, J. A., Daskalakou, E., Vallejo, V. R., Grigoriadis, N., et al. (2012). “Post-fire management of serotinous pine forests,” in Post-fire management and restoration of Southern European Forests, eds F. Moreira, M. Arianoutsou, P. Corona, and J. de las Heras (Dordrecht: Springer Netherlands), 121–150. doi: 10.1016/j.scitotenv.2020.144222
DellaSala, D. A. (2020). “Fire-mediated biological legacies in dry forested ecosystems of the Pacific Northwest,” in Disturbance ecology and biological diversity, eds E. A. Beaver, S. Prange, and D. A. DellaSala (Boca Raton, FL: CRC Press Taylor and Francis Group), 38–85.
DellaSala, D. A., and Hanson, C. T. (2015). The ecological importance of mixed-severity fires: Nature’s phoenix. Amsterdam: Elsevier.
DellaSala, D. A., Karr, J. R., Schoennagel, T., Perry, D., Noss, R. F., Lindenmayer, D., et al. (2006). Post-fire logging debate ignores many issues. Science 314, 51–52. doi: 10.1126/science.314.5796.51b
Dietze, E., Brykała, D., Schreuder, L. T., Jażdżewski, K., Blarquez, O., Brauer, A., et al. (2019). Human-induced fire regime shifts during 19th century industrialization: A robust fire regime reconstruction using northern Polish lake sediments. PLoS One 14:e0222011. doi: 10.1371/journal.pone.0222011
Dimitrakopoulos, A. P., Mitsopoulos, I. D., and Kaliva, A. (2011). Short communication. Comparing flammability traits among fire-stricken (low elevation) and non fire- stricken (high elevation) conifer forest species of Europe: A test of the mutch hypothesis. For. Syst. 22, 134–137. doi: 10.5424/fs/2013221-02475
Doerr, S. H., and Santín, C. (2016). Global trends in wildfire and its impacts: Perceptions versus realities in a changing world. Philos. Trans. R. Soc. Lond B Biol. Sci. 371:20150345. doi: 10.1098/rstb.2015.0345
Donato, D. C., Fontaine, J. B., Campbell, J. L., Robinson, W. D., Kauffman, J. B., and Law, B. E. (2006). Post-wildfire logging hinders regeneration and increases fire risk. Science 311:352. doi: 10.1126/science.1122855
Espelta, J. M., Retana, J., and Habrouk, A. (2003). An economic and ecological multi-criteria evaluation of reforestation methods to recover burned Pinus nigra forests in NE Spain. For. Ecol. Manag. 180, 185–198. doi: 10.1016/S0378-1127(02)00599-6
FAO (2007). Fire management: Global assessment 2006 : A thematic study prepared in the framework of the global forest resources assessment 2005. Rome: Food and Agriculture Organization of the United Nations.
Fernandez-Anez, N., Krasovskiy, A., Müller, M., Vacik, H., Baetens, J., Hukiæ, E., et al. (2021). Current Wildland fire patterns and challenges in Europe: A synthesis of national perspectives. Air Soil Water Res. 14, 1–19. doi: 10.1177/11786221211028185
Feurdean, A., Florescu, G., Tanţău, I., Vannière, B., Diaconu, A.-C., Pfeiffer, M., et al. (2020). Recent fire regime in the southern boreal forests of western Siberia is unprecedented in the last five millennia. Quat. Sci. Rev. 244:106495. doi: 10.1016/j.quascirev.2020.106495
Fischer, A., and Fischer, H. (2012). “Restoration of temperate Forests: An European approach,” in Restoration ecology: The new frontier, 2nd Edn, eds J. van Andel and J. Aronson (Hoboken, NJ: Wiley-Blackwell), 145–160. doi: 10.1016/j.ppees.2018.01.002
Fonda, R. W. (2001). Burning characteristics of needles from eight pine species. For. Sci. 47, 390–396. doi: 10.1093/forestscience/47.3.390
Fonda, R. W., and Varner, J. M. (2004). Burning characteristics of cones from eight pine species. Northwest Sci. 78, 322–333.
Forzieri, G., Girardello, M., Ceccherini, G., Spinoni, J., Feyen, L., Hartmann, H., et al. (2021). Emergent vulnerability to climate-driven disturbances in European forests. Nat. Commun. 12:1081. doi: 10.1038/s41467-021-21399-7
Franklin, J. F., and Agee, J. K. (2003). Forging a science-based national forest fire policy. Issues Sci. Technol. 20, 59–66. doi: 10.4324/9780203495261.bmatt
Franklin, J. F., Lindenmayer, D., Macmahon, J. A., McKee, A., Magnuson, J., Perry, D. A., et al. (2000). Threads of continuity: Ecosystem disturbance, recovery, and the theory of biological legacies. Conserv. Spring 1, 8–17.
Ganteaume, A., Camia, A., Jappiot, M., San-Miguel-Ayanz, J., Long-Fournel, M., and Lampin, C. (2013). A review of the main driving factors of forest fire ignition over Europe. Environ. Manag. 51, 651–662. doi: 10.1007/s00267-012-9961-z
Gauer, J., and Aldinger, E., eds (2005). Waldökologische naturräume deutschlands: Forstliche wuchsgebiete und wuchsbezirke; mit Karte 1:1.000.000. Freiburg: Verein für Forstliche Standortskunde und Forstpflanzenzüchtung.
Ginzburg, O., and Steinberger, Y. (2012). Salvage logging versus natural regeneration post-fire practices in a forest: Soil chemical and microbial aspects. OJE 02, 29–37. doi: 10.4236/oje.2012.21004
Gnilke, A., and Sanders, T. (2021). Forest fire history in Germany (2001-2020). Eberswalde: Thünen Institute of Forest Ecosystems.
Gustafsson, L., Baker, S. C., Bauhus, J., Beese, W. J., Brodie, A., Kouki, J., et al. (2012). Retention forestry to maintain multifunctional forests: A world perspective. Bioscience 62, 633–645. doi: 10.1525/bio.2012.62.7.6
Ibisch, P. L. (2019). Umgang mit der aktuellen extremwetterbedingten Waldkrise (Deutscher Bundestag Ausschussdrucksache 19(10)280-A) für die 41. Sitzung des Ausschusses für Ernährung und Landwirtschaft. Eberswalde: Hochschule für nachhaltige Entwicklung Eberswalde.
Iglesias, V., Balch, J. K., and Travis, W. R. (2022). U.S. fires became larger, more frequent, and more widespread in the 2000s. Sci. Adv. 8:eabc0020. doi: 10.1126/sciadv.abc0020
Inbar, M., Wittenberg, L., and Tamir, M. (1997). Soil erosion and forestry management after wildfire in a mediterranean woodland, Mt. Carmel, Israel. Int. J. Wildland Fire 7, 285–294. doi: 10.1071/WF9970285
Kukavskaya, E. A., Buryak, L. V., Shvetsov, E. G., Conard, S. G., and Kalenskaya, O. P. (2016). The impact of increasing fire frequency on forest transformations in southern Siberia. For. Ecol. Manag. 382, 225–235. doi: 10.1016/j.foreco.2016.10.015
Lamb, D., and Gilmour, D. (2003). Rehabilitation and restoration of degraded forests. Gland: IUCN Gland.
Lavorel, S., Flannigan, M. D., Lambin, E. F., and Scholes, M. C. (2007). Vulnerability of land systems to fire: Interactions among humans, climate, the atmosphere, and ecosystems. Mitig. Adap. Strateg. Glob. Change 12, 33–53. doi: 10.1007/s11027-006-9046-5
Leuschner, C., Förster, A., Diers, M., and Culmsee, H. (2022). Are northern German Scots pine plantations climate smart? The impact of large-scale conifer planting on climate, soil and the water cycle. For. Ecol. Manag. 507:120013. doi: 10.1016/j.foreco.2022.120013
Leverkus, A. B., Lorite, J., Navarro, F. B., Sánchez-Cañete, E. P., and Castro, J. (2014). Post-fire salvage logging alters species composition and reduces cover, richness, and diversity in Mediterranean plant communities. J. Environ. Manag. 133, 323–331. doi: 10.1016/j.jenvman.2013.12.014
Leverkus, A. B., Puerta-Piñero, C., Guzmán-Álvarez, J. R., Navarro, J., and Castro, J. (2012). Post-fire salvage logging increases restoration costs in a Mediterranean mountain ecosystem. New For. 43, 601–613. doi: 10.1007/s11056-012-9327-7
Leverkus, A. B., Rey Benayas, J. M., Castro, J., Boucher, D., Brewer, S., Collins, B. M., et al. (2018). Salvage logging effects on regulating and supporting ecosystem services — a systematic map. Can. J. For. Res. 48, 983–1000. doi: 10.1139/cjfr-2018-0114
Lindenmayer, D. B., and Noss, R. F. (2006). Salvage logging, ecosystem processes, and biodiversity conservation. Conserv. Biol. 20, 949–958. doi: 10.1111/j.1523-1739.2006.00497.x
Lindenmayer, D., Burton, P., and Franklin, J. (2008). Salvage logging and its ecological consequences. Washington DC: Island Press.
Malvar, M. C., Silva, F. C., Prats, S. A., Vieira, D. C. S., Coelho, C. O. A., and Keizer, J. J. (2017). Short-term effects of post-fire salvage logging on runoff and soil erosion. For. Ecol. Manag. 400, 555–567. doi: 10.1016/j.foreco.2017.06.031
Mansourian, S., Lamb, D., and Gilmour, D. (2005). “Overview of technical approaches to restoring tree cover at the site level,” in Forest restoration in landscapes: Beyond planting trees, eds S. Mansourian, D. Vallauri, and N. Dudley (New York, NY: Springer), 241–249.
Marangon, D., Marchi, N., and Lingua, E. (2022). Windthrown elements: A key point improving microsite amelioration and browsing protection to transplanted seedlings. For. Ecol. Manag. 508:120050. doi: 10.1016/j.foreco.2022.120050
Marañón-Jiménez, S., Castro, J., Querejeta, J. I., Fernández-Ondoño, E., and Allen, C. D. (2013). Post-fire wood management alters water stress, growth, and performance of pine regeneration in a Mediterranean ecosystem. For. Ecol. Manag. 308, 231–239. doi: 10.1016/j.foreco.2013.07.009
Marcolin, E., Marzano, R., Vitali, A., Garbarino, M., and Lingua, E. (2019). Post-fire management impact on natural forest regeneration through altered microsite conditions. Forests 10:1014. doi: 10.3390/f10111014
Martin, R. E., Gordon, D. A., Gutierrez, M. E., Lee, D. S., Molina, D. M., and Schroeder, R. A. (1994). Assessing the flammability of domestic and wildland vegetation. Proc. Int. Confer. Fire For. Meteorol. 12, 130–137.
Martínez-Sánchez, J. J., Ferrandis, P., de las Heras, J., and Herranz, J. M. (1999). Effect of burnt wood removal on the natural regeneration of Pinus halepensis after fire in a pine forest in Tus valley (SE Spain). For. Ecol. Manag. 123, 1–10. doi: 10.1016/S0378-1127(99)00012-2
Marx, A., Samaniego, L., Kumar, R., Thober, S., Mai, J., and Zink, M. (2016). “Der dürremonitor – Aktuelle information zur bodenfeuchte in deutschland,” in Wasserressourcen – Wissen in flussgebieten vernetzen: Beiträge zum tag der hydrologie am 17./18. März 2016 in Koblenz, ausgerichtet von der hochschule koblenz und der bundesanstalt für gewässerkunde, eds G. Wernecke, A.-D. Ebner von Eschenbach, Y. Strunck, L. Kirschbauer, and A. Müller (Leipzig: Forum für Hydrologie und Wasserbewirtschaftung), 131–142.
Marzano, R., Garbarino, M., Marcolin, E., Pividori, M., and Lingua, E. (2013). Deadwood anisotropic facilitation on seedling establishment after a stand-replacing wildfire in Aosta Valley (NW Italy). Ecol. Eng. 51, 117–122. doi: 10.1016/j.ecoleng.2012.12.030
Mauri, E., and Pons, P. (2019). Handbook of good practices in post-wildfire management: Anifog project I+D+i CGL2014-54094-R. Girona: Universitat de Girona.
Mavsar, R., Varela, E., Corona, P., Barbati, A., and Marsh, G. (2012). “Economic, legal and social aspects of post-fire management,” in Post-fire management and restoration of southern european forests, eds F. Moreira, M. Arianoutsou, P. Corona, and J. de las Heras (Dordrecht: Springer Netherlands), 45–78.
McIver, J. D., and Starr, L. (2001). A literature review on the environmental effects of postfire logging. Western J. Appl. For. 16, 159–168. doi: 10.1093/wjaf/16.4.159
Merino, A., Balboa, M. A., Rodríguez Soalleiro, R., and Álvarez González, J. G. (2005). Nutrient exports under different harvesting regimes in fast-growing forest plantations in southern Europe. For. Ecol. Manag. 207, 325–339. doi: 10.1016/j.foreco.2004.10.074
Moreira, F., and Vallejo, V. R. (2009). “What to do after fire? Post-fire restoration,”,” in Living with wildfires: What science can tell us: A contribution to the science-policy dialogue. EFI discussion paper 15, ed. Y. Birot (Joensuu: European Forest Institute), 53–58.
Moreira, F., Arianoutsou, M., Corona, P., and de las Heras, J e (2012a). Post-fire management and restoration of Southern European Forests. Dordrecht: Springer Netherlands.
Moreira, F., Arianoutsou, M., Vallejo, V. R., de las Heras, J., Corona, P., Xanthopoulos, G., et al. (2012b). “Setting the scene for post-fire management,” in Post-fire management and restoration of Southern European Forests, eds F. Moreira, M. Arianoutsou, P. Corona, and J. de las Heras (Dordrecht: Springer Netherlands), 1–19.
Moreira, F., Catry, F. X., Lopes, T., Bugalho, M. N., and Rego, F. (2009). Comparing survival and size of resprouts and planted trees for post-fire forest restoration in central Portugal. Ecol. Eng. 35, 870–873. doi: 10.1016/j.ecoleng.2008.12.017
Moya, D., de las Heras, J., López-Serrano, F. R., and Ferrandis, P. (2015). Post-fire seedling recruitment and morpho-ecophysiological responses to induced drought and salvage logging in Pinus halepensis Mill. stands. Forests 6, 1858–1877. doi: 10.3390/f6061858
Ne’eman, G., Perevolotsky, A., and Schiller, G. (1997). The management implications of the Mt. Carmel research project. Int. J. Wildland Fire 7, 343–350. doi: 10.1071/WF9970343
Newton, M., Fitzgerald, S., Rose, R. R., Adams, P. W., Tesch, S. D., Sessions, J., et al. (2006). Comment on “Post-wildfire logging hinders regeneration and increases fire risk”. Science 313:615;authorrely615. doi: 10.1126/science.1126478
Noss, R. F., Franklin, J. F., Baker, W. L., Schoennagel, T., and Moyle, P. B. (2006). Ecology and management of fire-prone forests of the western united states: Society for conservation biology scientific panel on fire in western U.S. Forests. Arlington, VA: Society for Conservation Biology.
Ormeño, E., Ruffault, J., Gutigny, C., Madrigal, J., Guijarro, M., Hernando, C., et al. (2020). Increasing cuticular wax concentrations in a drier climate promote litter flammability. For. Ecol. Manag. 473:118242. doi: 10.1016/j.foreco.2020.118242
Pausas, J. G., and Keeley, J. E. (2019). Wildfires as an ecosystem service. Front. Ecol. Environ. 17:289–295. doi: 10.1002/fee.2044
Pausas, J. G., Bladé, C., Valdecantos, A., Seva, J. P., Fuentes, D., Alloza, J. A., et al. (2004). Pines and oaks in the restoration of mediterranean landscapes of Spain: New perspectives for an old practice – a review. Plant Ecol. 171, 209–220. doi: 10.1023/B:VEGE.0000029381.63336.20
Pereira, P., Francos, M., Brevik, E. C., Úbeda, X., and Bogunovic, I. (2018). Post-fire soil management. Curr. Opin. Environ. Sci. Health 5, 26–32. doi: 10.1016/j.coesh.2018.04.002
Peterson, D. L., Agee, J. K., Aplet, G. H., Dykstra, D. P., Graham, R. T., Lehmkuhl, J. F., et al. (2009). Effects of timber harvest following wildfire in western North America. Portland, OR: U.S. Dept. of Agriculture.
Philpot, C. W. (1970). Influence of mineral content on the pyrolysis of plant materials. For. Sci. 16, 461–471. doi: 10.1093/forestscience/16.4.461
Popović, Z., Bojović, S., Marković, M., and Cerdà, A. (2021). Tree species flammability based on plant traits: A synthesis. Sci.. Total Environ. 800:149625. doi: 10.1016/j.scitotenv.2021.149625
Porter, A., Assal, T., Hobbs, N. T., and Sibold, J. (2022). Wildfire catalyzed shift from conifer to aspen dominance in montane zone, Colorado. SSRN J 40. doi: 10.2139/ssrn.4153771
Pyne, S. J. (2020). From pleistocene to pyrocene: Fire replaces ice. Earths Future 8, 1–5. doi: 10.1029/2020EF001722
Randerson, J. T., Chen, Y., van der Werf, G. R., Rogers, B. M., and Morton, D. C. (2012). Global burned area and biomass burning emissions from small fires. J. Geophys. Res. Biogeosci. 117:G04012. doi: 10.1029/2012JG002128
Robichaud, P. R. (2009). “Post-fire stabilization and rehabilitation,” in Fire Effects on soils and restoration strategies, eds A. Cerdà and P. R. Robichaud (Enfield, NH: Science Publishers), 299–320.
Robichaud, P. R., Elliot, W. J., MacDonald, L., Coats, R., Wagenbrenner, J. W., Lewis, S. A., et al. (2011). Evaluating post-fire salvage logging effects on erosion. JFSP research project reports. 64. Available online at: http://digitalcommons.unl.edu/jfspresearch/64
Romero, B., and Ganteaume, A. (2021). Effect of fire frequency on the flammability of two mediterranean pines: Link with needle terpene content. Plants (Basel) 10:2164. doi: 10.3390/plants10102164
Rundel, P. W. (1981). “Structural and chemical components of flammability,” in Fire regimes and ecosystem properties: Proceedings of the conference. General technical report WO-GTR-26, eds H. A. Mooney, T. M. Bonnicksen, N. L. Christensen Jr., J. E. Lotan, and W. A. Reiners (Washington, DC: USDA Forest Service), 183–207.
Ryu, S.-R., Choi, H.-T., Lim, J.-H., Lee, I.-K., and Ahn, Y.-S. (2017). Post-Fire restoration plan for sustainable forest management in South Korea. Forests 8:188. doi: 10.3390/f8060188
San-Miguel-Ayanz, J., Durrant, T., Boca, R., Libertà, G., Branco, A., Rigo, D., et al. (2019). Forest fires in Europe, middle East and North Africa 2018: JRC technical report. Brussels: European Commission.
Schmidt, W., and Wichmann, I. (2000). Zur sukzession von waldbrandflächen in der lüneburger heide. Forst und Holz 55, 481–487.
Schulze, G., Kopp, D., and Wirner, M. (2013). Anleitung für die forstliche standortserkundung im nordostdeutschen tiefland (Standortserkundungsanleitung): SEA 95. Malchin: Landesforst Mecklenburg Vorpommern AöR.
Senf, C., and Seidl, R. (2021). Storm and fire disturbances in Europe: Distribution and trends. Glob. Change Biol. 27, 3605–3619. doi: 10.1111/gcb.15679
Sessions, J., Bettinger, P., Buckman, R., Newton, M., and Hamann, J. (2004). Hastening the return of complex forests following fire: The consequences of delay. J. For. 102, 38–45.
Shafizadeh, F., Chin, P. P. S., and DeGroot, W. F. (1977). Effective heat content of green forest fuels. For. Sci. 23, 81–89.
Spanos, I. A., Raftoyannis, Y., Platis, P., and Xanthopoulou, E. (2010). Post-fire management and recovery of a pine forest in Greece. Web Ecol. 10, 27–31.
Tanskanen, H. (2007). Fuel conditions and fire behavior characteristics of managed Picea abies and Pinus sylvestris forests in Finland. Academic dissertation. Helksinki: Department of Forest Ecology, Faculty of Agriculture and Forestry.
Tanskanen, H., Venäläinen, A., Puttonen, P., and Granström, A. (2005). Impact of stand structure on surface fire ignition potential in Picea abies and Pinus sylvestris forests in southern Finland. Canadian J. For. Res. 35, 410–420. doi: 10.1139/x04-188
Tinya, F., Kovács, B., Aszalós, R., Tóth, B., Csépányi, P., Németh, C., et al. (2020). Initial regeneration success of tree species after different forestry treatments in a sessile oak-hornbeam forest. For. Ecol. Manag. 459:117810. doi: 10.1016/j.foreco.2019.117810
Úbeda, X., and Outeiro, L. R. (2009). “Physical and chemical effects of fire on soil,” in Fire effects on soils and restoration strategies, eds A. Cerdà and P. R. Robichaud (Enfield, NH: Science Publishers), 105–132.
Ürker, O., Tavşanoðlu, Ç, and Gürkan, B. (2018). Post-fire recovery of the plant community in Pinus brutia forests: Active vs. indirect restoration techniques after salvage logging. iForest 11, 635–642. doi: 10.3832/ifor2645-011
Urretavizcaya, M. F., and Defossé, G. E. (2019). Restoration of burned and post-fire logged Austrocedrus chilensis stands in Patagonia: Effects of competition and environmental conditions on seedling survival and growth. Int. J. Wildland Fire 28:365. doi: 10.1071/WF18154
Vallauri, D. (2005). “Restoring forests after violent storms,” in Forest restoration in landscapes: Beyond planting trees, eds S. Mansourian, D. Vallauri, and N. Dudley (New York, NY: Springer), 339–344.
Vallejo, V. R., Allen, E. B., Aronson, J., Pausas, J. G., Cortina, J., and Gutiérrez, J. R. (2012a). “Restoration of mediterranean-type woodlands and shrublands,” in Restoration ecology: The new frontier, 2nd Edn, eds J. van Andel and J. Aronson (Hoboken, NJ: Wiley-Blackwell), 130–144.
Vallejo, V. R., Arianoutsou, M., and Moreira, F. (2012b). “Fire ecology and post-fire restoration approaches in southern european forest types,” in Post-fire management and restoration of southern european forests, eds F. Moreira, M. Arianoutsou, P. Corona, and J. de las Heras (Dordrecht: Springer Netherlands), 93–119.
Vallejo, V. R., Serrasolses, I., Alloza, J. A., Baeza, M. J., Bladé, C., Chirino, E., et al. (2009). “Long-term Restoration strategies and techniques,” in Fire effects on soils and restoration strategies, eds A. Cerdà and P. R. Robichaud (Enfield, NH: Science Publishers), 373–398.
Vlassova, L., and Pérez-Cabello, F. (2016). Effects of post-fire wood management strategies on vegetation recovery and land surface temperature (LST) estimated from Landsat images. Int. J. Appl. Earth Observ. Geoinform. 44, 171–183. doi: 10.1016/j.jag.2015.08.011
Wagenbrenner, J. W., Robichaud, P. R., and Brown, R. E. (2016). Rill erosion in burned and salvage logged western montane forests: Effects of logging equipment type, traffic level, and slash treatment. J. Hydrol. 541, 889–901. doi: 10.1016/j.jhydrol.2016.07.049
Keywords: restoration, rejuvenation, microclimate, temperature, VPD, salvage logging, deadwood retention
Citation: Blumroeder JS, Schmidt F, Gordon A, Grosse S and Ibisch PL (2022) Ecosystemic resilience of a temperate post-fire forest under extreme weather conditions. Front. For. Glob. Change 5:1070958. doi: 10.3389/ffgc.2022.1070958
Received: 15 October 2022; Accepted: 23 November 2022;
Published: 08 December 2022.
Edited by:
William R. Moomaw, Tufts University, United StatesReviewed by:
Dominick DellaSala, Earth Island Institute, United StatesChad Hanson, Earth Island Institute, United States
Copyright © 2022 Blumroeder, Schmidt, Gordon, Grosse and Ibisch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeanette S. Blumroeder, ai5ibHVtcm9lZGVyQGhuZWUuZGU=