
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change, 11 January 2023
Sec. Forests and the Atmosphere
Volume 5 - 2022 | https://doi.org/10.3389/ffgc.2022.1065645
This article is part of the Research TopicForest-Climate Interactions and the Climate Consequences of Forest-Cover ChangeView all 4 articles
 Flavia Machado Durgante1,2*
Flavia Machado Durgante1,2* Niro Higuchi2
Niro Higuchi2 Shinta Ohashi3John Ethan Householder1
Shinta Ohashi3John Ethan Householder1 Adriano José Nogueira Lima2
Adriano José Nogueira Lima2 Moriyoshi Ishizuka3
Moriyoshi Ishizuka3 Florian Wittmann1,2Joaquim dos Santos2Vilany Matilla Colares Carneiro2Xiaomei Xu4Claudete Catanhede do Nascimento2
Florian Wittmann1,2Joaquim dos Santos2Vilany Matilla Colares Carneiro2Xiaomei Xu4Claudete Catanhede do Nascimento2 Jochen Schöngart2
Jochen Schöngart2 Maria Teresa Fernandez Piedade2Adalberto Rodrigo Kossmann Schmitt2
Maria Teresa Fernandez Piedade2Adalberto Rodrigo Kossmann Schmitt2 Yanka Laryssa Almeida Alves2
Yanka Laryssa Almeida Alves2 Jennifer Lehman5
Jennifer Lehman5 Bruno Oliva Gimenez2,6Priscilla Maia Baggio2Lucas Kosvoski de Ourique2
Bruno Oliva Gimenez2,6Priscilla Maia Baggio2Lucas Kosvoski de Ourique2 Susan Trumbore4,7
Susan Trumbore4,7Introduction: The productivity of the Amazon Rainforest is related to climate and soil fertility. However, the degrees to which these interactions influence multiannual to decadal variations in tree diameter growth are still poorly explored.
Methods: To fill this gap, we used radiocarbon measurements to evaluate the variation in tree growth rates over the past decades in an important hyperdominant species, Eschweilera coriacea (Lecythidaceae), from six sites in the Brazilian Amazon that span a range of soil properties and climate.
Results: Using linear mixed-effects models, we show that temporal variations in mean annual diameter increment evaluated over a specific time period reflect interactions between soil fertility and the drought index (SPEI-Standardized Precipitation and Evapotranspiration Index).
Discussion: Our results indicate that the growth response of trees to drought is strongly dependent on soil conditions, a facet of forest productivity that is still underexplored, and which has great potential for improving predictions of future tropical tree growth in the face of projected climate change.
Growth rates and turnover of woody biomass are important components of carbon (C) cycling in the Amazon basin (Zhao and Running, 2010). At the broad scale of the basin, growth rates have been shown to vary according to water availability and soil fertility (Toledo et al., 2011; Quesada et al., 2012; Sousa et al., 2022). High water availability increases forest productivity in the tropics (Pereira da Silva et al., 2002; Zuidema et al., 2022), thus the extreme drought events suppress this productivity (Phillips et al., 2009; Doughty et al., 2015). Additionally, the soil’s fertility gradient is another factor that influences forest productivity in the Amazon basin, as trees in the nutrient-rich soils closer to the Andes have on average lower wood density, faster growth and turnover, while those growing in poorer soils in the Central and Eastern Amazon have generally higher wood density and slower growth and turnover (Quesada et al., 2012).
The environmental resources have diverse effects on forest productivity due to the heterogeneity of habitats in the tropics, where nutrients, water availability and radiance (light) vary over space and time (Muller-Landau et al., 2021; Sousa et al., 2022). However, the effect of these environmental factors on tree growth does not follow a certain pattern. For example, the literature indicates that the increase in water availability increases production (Zuidema et al., 2022), but an inverse behavior is observed in forests with shallow groundwater (Sousa et al., 2022). Another example is photosynthetically active radiation (PAR), which would not be a limiting factor in the tropics, but the cloud cover limits PAR in tropical wet forests, and this limitation affects forest productivity (Clark and Clark, 1994; Schuur, 2003). In the Amazon basin, while soil fertility drives forest productivity from west to east (Malhi et al., 2004; Quesada et al., 2012), Toledo et al. (2011) state that in Bolivia the climate is the strongest driver of spatial variation in tree growth. However, the interactions between climate variation and soil fertility are a potentially important facet of forest productivity.
Many of the studies that link the allocation of tree resources to stem growth and changing environmental conditions rely on permanent plot data and, thus, integrate the response of many tree species. For example, the negative relationship between tree growth and wood density reported across the Amazon basin (Quesada et al., 2012) is driven in part by changes in species composition with different life strategies or traits among the different locations (Baker et al., 2004). Wood density are trait with high heritability (Cornelius, 1994), were 74% of species level wood density variation was explained at the genus level (Chave et al., 2006). However, some authors showed a significant intraspecific variation in that trait varying with an environmental conditions, as flooded levels (Wittmann et al., 2006) or different types of flooded forests (Schöngart et al., 2005). Few studies in terra-firme forest have evaluated whether the gradients in soil fertility can cause similar kinds of variation in wood density and tree growth for a single species.
To fill this gap, we explored the decadal-average tree stem growth rates for a single hyperdominant species, which was sampled in terra firme forest sites in the state of Amazonas, Brazil, that varied in climate and soil fertility. Average tree growth was estimated over different time intervals by using radiocarbon measurements to determine the calendar year of wood growth at several points in radial sections for each tree. Radiocarbon measurements were used to determine whether growth rings were recognizable in the wood (Andreu-Hayles et al., 2015; Schöngart et al., 2017); however, when ring counts were compared with radiocarbon, many missing rings were found in most trees (Ohashi et al., 2016; Herrera-Ramirez et al., 2017; Santos et al., 2021). As we could not rely on ring-counting for some species, instead we used the difference in the 14C-determined calendar year to estimate the time interval during which a specific radial growth increment took place, especially in species without annual rings.
Using the hyperdominant tropical tree species Eschweilera coriacea (Ter Steege et al., 2020) growing in a terra firme forest as a model, we hypothesized that stem growth rates would be lowest in nutrient-poor soils and would increase with soil nutrient content. In soils with higher fertility, rainfall is likely to become a limiting factor, with growth rates increasing with rainfall from dry to wet conditions. In addition, we expect a negative relationship between wood density and tree growth rate within a single species as observed in another tropical forest ecosystem (Schöngart et al., 2005) and a narrow intraspecific variation comparing to interspecific variation in the tree community (Chave et al., 2006). Our study can therefore help determine the role of intraspecific variation in traits that have been observed varying with edaphic conditions across the Amazon (Quesada et al., 2011). However, the main advantage is that it contributes to knowledge on how climatic and edaphic conditions interact to govern spatial and temporal variations in tree growth. These are fundamental for understanding growth dynamics, climate, and carbon cycling of forests across this globally significant region.
For this study, we selected the hyperdominant species Eschweilera coriacea (DC.) S.A Mori, which has a mean estimated population of 5 × 109 trees in the Amazon (ter Steege et al., 2013, 2020) and is among the most widespread and abundant canopy tree species in the terra firme forest (Mori and Lepsch-Cunha, 1995). Eschweilera coriacea belongs to the Lecythidaceae family, whose members are common in mature Neotropical forests (Mori et al., 2007), As there are several species that can be confused with Eschweilera coriacea, all individuals had their identification confirmed by specialists and were compared with reference materials in the herbarium collections at the National Institute for Amazonian Research (INPA) in Manaus, Brazil.
We sampled individuals of E. coriacea from terra firme upland forests in plateau areas at six sites within the state of Amazonas (Figure 1). The vegetation in all sites can be classified as old-growth, tropical evergreen forest with closed canopy (Chambers et al., 2001). Across the sites, mean annual rainfall varied from 2,427 to 3,621 mm (Figure 2). Five sites have a Köppen’s climate classification of Af–without a dry season, while Manicoré (the southern-most site) is Am–tropical monsoon (Alvares et al., 2013).
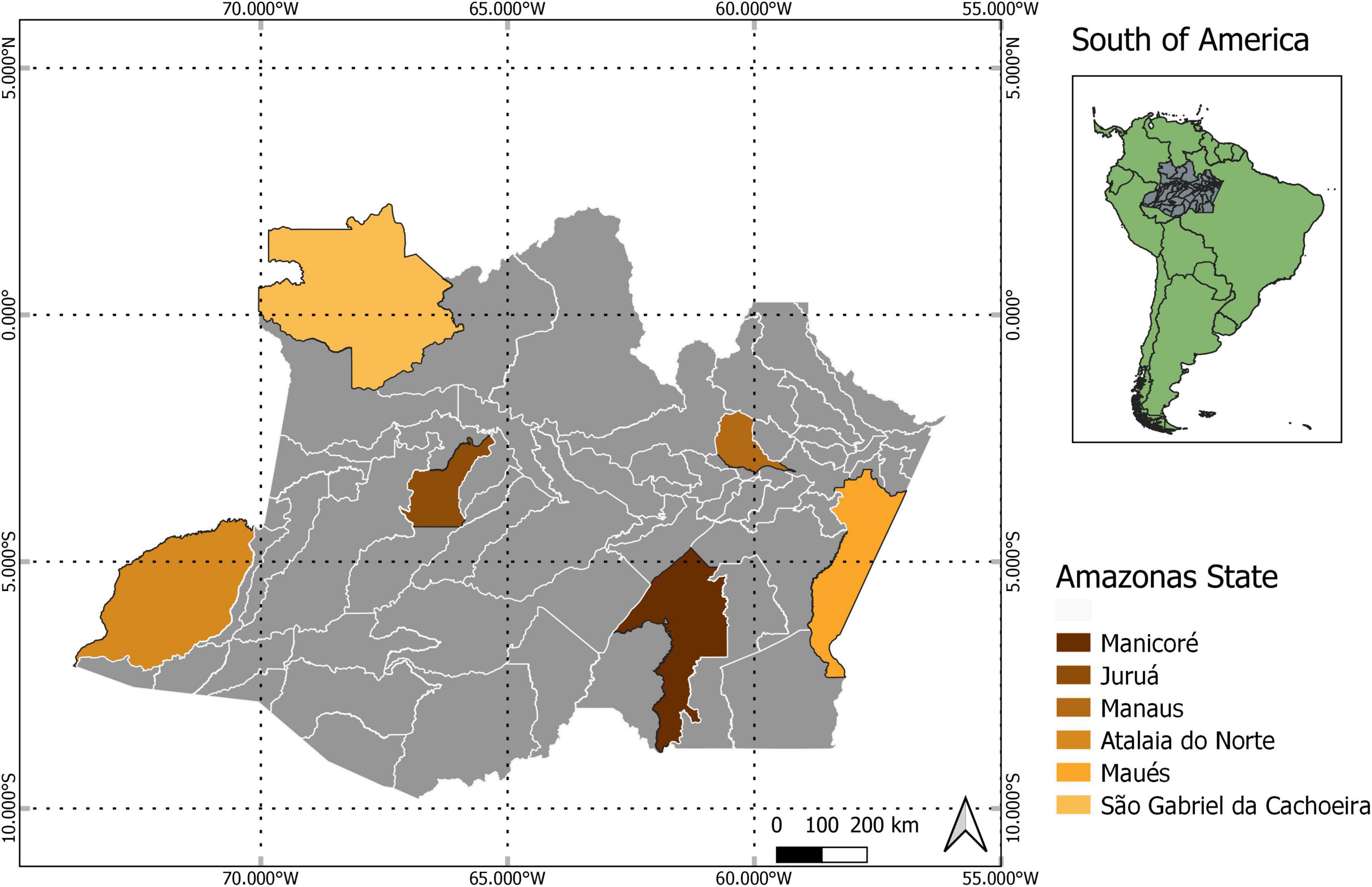
Figure 1. Map showing the locations of the study sites.
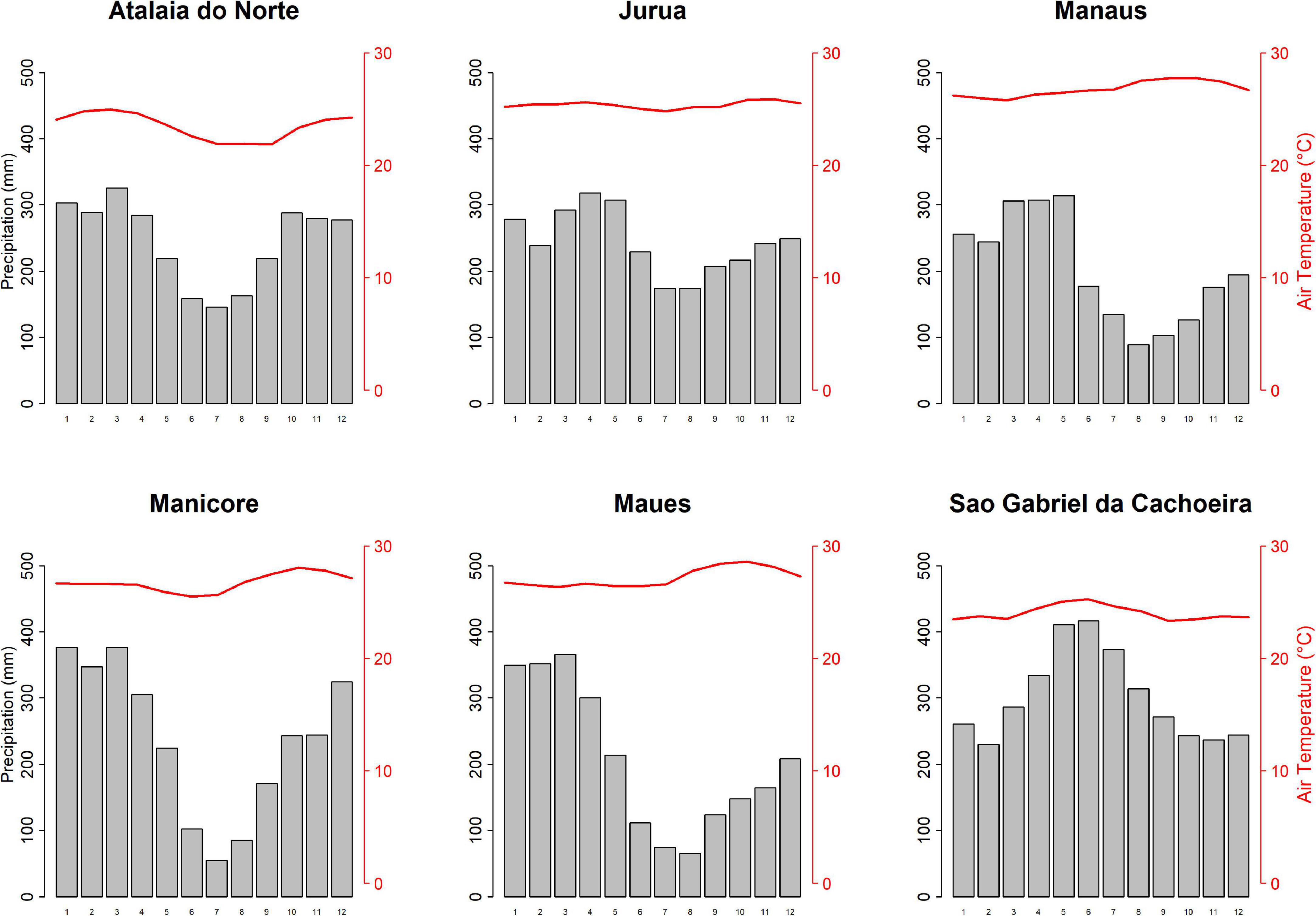
Figure 2. Seasonal distribution of monthly mean precipitation (gray histogram) and temperature (red line) at each study site [adapted from Alvares et al. (2013)].
Between July 2013 and April 2014, five individuals of E. coriacea were cut down at each site to analyze growth rates and wood specific gravity. The diameter at breast height (dbh) of these trees ranged from 18 to 47 cm. Discs cut at 1.3 m were taken from each of the 30 trees (Sisbio License: 27391311). The surfaces of the discs were coarsely polished with a disc sander (grit 60) and then with a hand sander (grit 80 to 200). Two radii per tree were selected at an angle that varied from 120 to 180 degrees in order to avoid holes, insect damage, or rot. Radial slices with tangential widths of 12 mm were removed with a band saw. These were trimmed to a thickness of ∼1 mm with a circular table saw or a band saw. Cellulose (holocellulose) was extracted from these thin wood sections using the method of Kagawa et al. (2015). The chemical process for holocellulose extraction was described by Ohashi et al. (2016) and included freeze drying and correction for any shrinkage of the wood from its original dimensions.
Radiocarbon dating of the resulting holocellulose was performed by removing 1 mm-thick slices of a length needed to achieve 2–3 mg of material. These were centered at distances of 5, 10, and 15 mm away from the bark on each of the two radii from each individual.
The 2–3 mg of sampled holocellulose was combusted with CuO in evacuated and sealed quartz tubes at 900°C, and the resulting CO2 was purified and converted to graphite using sealed-tube zinc reduction (Xu et al., 2007). Radiocarbon analyses were performed at the W. M. Keck Carbon Cycle Accelerator Mass Spectrometry Facility at the University of California, Irvine, USA. Radiocarbon data are reported as the fraction modern (FM), the ratio of 14C/12C in the sample divided by that of a pre-industrial wood standard. Using this standard annotation, a correction is applied to remove the effects of mass-dependent isotopic fractionation using measured δ13C values (Stuiver and Polach, 1977). The accuracy for 14C analyses is 2–3‰ for modern samples based on the long-term reproducibility of secondary standards.
To convert fraction modern (FM) data to calendar years of wood formation, we used OxCal [version 4.2] (Ramsey, 2009) with the calibration curve: Bomb 13 SH 1 2 (Hogg et al., 2013; Andreu-Hayles et al., 2015). Only 14C data in the bomb period after 1950 AD were considered (errors of at most 1–2 years). Dates older than 1950 (0.96 to 1.0 FM) indicate wood formed between ∼1650 and 1950 AD and were not used in further analyses. If one or more of the samples from our standard depth increments of 5, 10, and 15 mm dated as being older than 1950, we took additional samples at depths closer to the bark in order to have three samples dating post-1960 per radial transect in order to calculate mean annual increments. Calendar year results were reported using 95.4% probability determined from combined uncertainties in the 14C analyses and calibration curves (Ramsey, 2009).
The periodic annual increment (PAI) was determined by dividing the growth distance between two radiocarbon-dated sampling points by the number of calendar years elapsed. We assumed the outermost xylem represents the year the tree was cut down (Figure 3). The distance between two dated points or from a dated point to the outermost xylem indicates how many millimeters the radius grew in the time that had elapsed, and was determined by the difference between the two 14C calibrated years or between the calibrated date and the year that the tree was cut down. The increment divided by time elapsed was defined as the periodic annual increment (PAI; mm/year). Figure 3 shows the example of tree EC-Ju-1, in which we can observe a variation in the growth rate of over time.
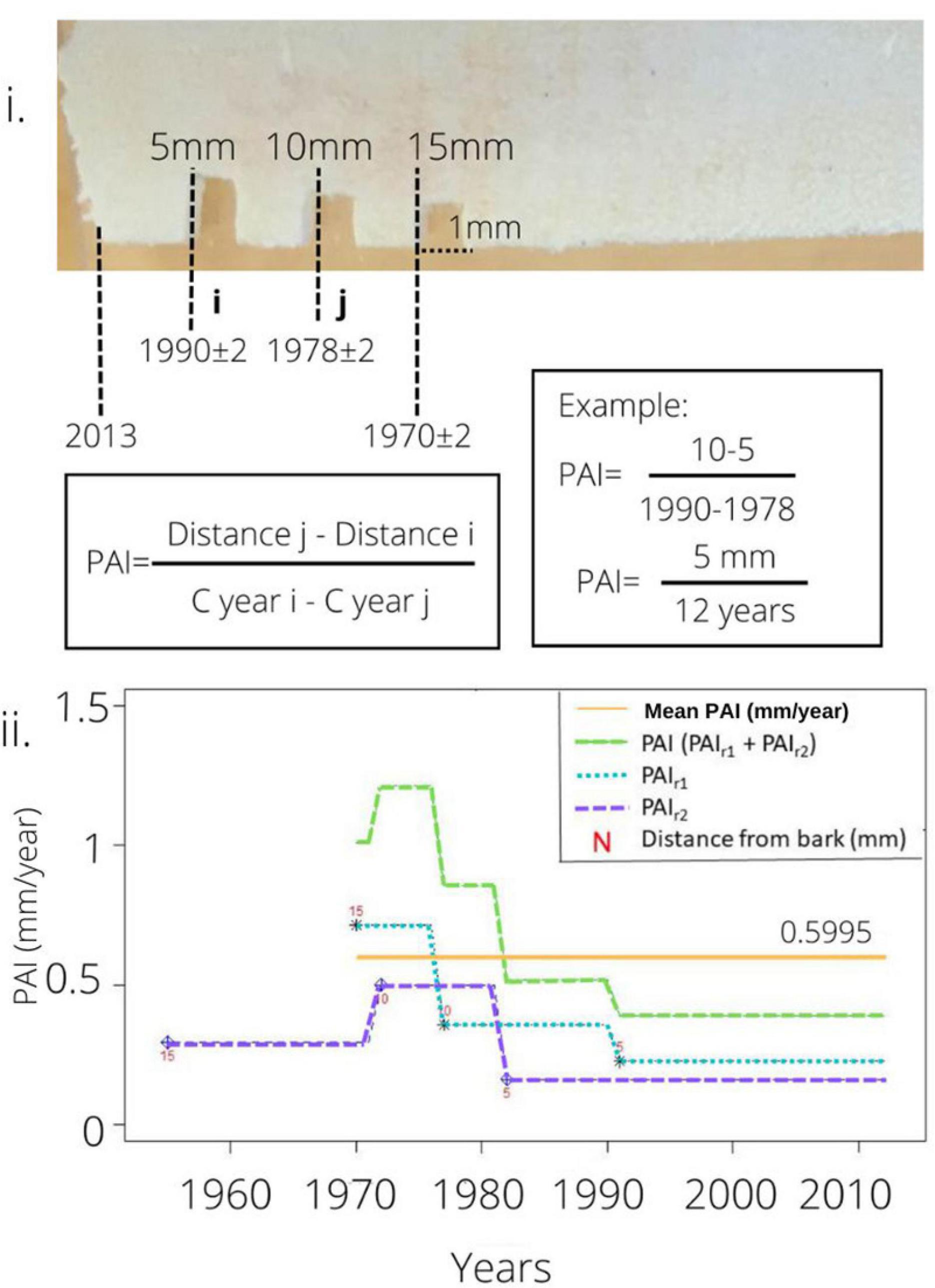
Figure 3. Explanatory figure for how we used 14C dating to estimate growth rates, expressed as periodic annual increment (PAI). (i) Photo showing the holocellulose (white) slab that is ∼1 mm thick; the portions removed for 14C measurements at 5, 10, and 15 cm radial positions can be seen. The calendar age determined for each sample is shown, as well as the assumed year of tree cut at the bark, and the calculation of PAI is shown as an example. (ii) Graph showing how PAI varies with time for each interval dated with 14C. The red number (N) indicates the distance (mm) from the dated point to the bark; PAIr1- periodic annual increment of radius 1; PAIr2- periodic annual increment of radius 2; PAI = The sum of the two radii over the entire period (assuming the sum of the two radii provides a measurement of the diameter increment; For this individual, PAI declined over the decades since the 1970s. Mean PAI is the periodic annual increment averaged out over the entire period represented in the outer 15 mm (usually representing multiple decades of growth).
The radial and longitudinal variation of the wood specific gravity (wood density) was determined for all the tree trunks. Eight subsamples (2 × 2 × 2 cm3) were removed from each part of the trunk, four in the radial direction (bark to pith) from two heights and at four different positions on the trunk (base, 1.3 m, 50 and 100% of commercial height), which totaled 32 subsamples per tree. The wood specific gravity from each subsample was determined as the ratio between water-saturated volume and dry weight (Williamson and Wiemann, 2010). The saturated volume was determined using Archimedes principle of liquid displacement in the saturated samples after 15 days submerged in water. Afterward, the samples were oven-dried for 3 days at 103°C until a reaching constant weight.
The standardized precipitation and evapotranspiration index–SPEI (Vicente-Serrano et al., 2010) was determined at each site and used to assess diameter growth responses to drought events at the different temporal scales represented by our 5 mm radial samples. SPEI is a drought index that captures the main impact of temperature increase on water demand, as it uses both precipitation and potential evapotranspiration data at a spatial resolution of 0.5 degrees (Beguería et al., 2014). SPEI is a relative measurement that is comparable across space and time. In other words, SPEI can correct for the fact that the same amount of rain could represent extreme drought at one site, but could represent greater than average rainfall at another site. Monthly average SPEI values (negative values equal drought) were calculated at each site and then averaged over each dated time interval represented by wood formed between 1950 and 2014.
To assess differences in soil properties, we sampled mineral soils (0–20 cm depth) from two collection points at a distance of 5 meters from the base of each sampled tree. The two samples were combined and used for subsequent analyses at the Soil and Plant Thematic Laboratory of the National Institute for Amazonian Research (INPA) according to method described by Quesada et al. (2010). The parameters evaluated include sand, silt and clay content (%), pH (hydrogen potential), Mg (magnesium), K (potassium), Ca (calcium), Na (sodium), and SB (sum of base cations) in cmol*kg–1. Differences in soils between sites were summarized using principal component analysis (PCA) and, in particular, the axis of first component (PC1) provides a useful measure of the fertility gradient between sites with the highest scores for Mg, K, pH, SB, and sand (Supplementary Figure 1). The macronutrients Mg and K are essential elements for plant growth and are involved in several essential physiological functions (Zekri and Obreza, 2003).
The relationships between tree growth, wood density and environmental variables were verified using linear regression models. To test for interactions, three increasingly complex models were generated (Table 1) and used tree growth as a dependent variable. Initial models explored tree growth as a function of either SPEI or fertility (PC1 axis value for each tree). Tree growth as an additive effect of SPEI and fertility was subsequently modeled. In this model, soil fertility would be expected to have a positive effect on growth and its effect is independent of SPEI. In a final model, interactions between SPEI and fertility were tested, by which soil fertility is expected to moderate growth response to SPEI. Competing models were compared with a drop in deviance test: a model with higher deviance according to AIC (Akaike Information Criterion) and BIC (Schwartz’s Bayesian Information Criterion), which provides a poorer model fit to the data than a model with lower deviance (Bates et al., 2015). Fixed effect variables are the variables that influence the response, and are known as explanatory variables in standard linear regression (Bolker et al., 2009). Random effect variables are categorical variables of the factors that need to be controlled (Bolker et al., 2009). The analyses were performed in R with the main packages MASS (Venables and Ripley, 2002), lme4 (Bates et al., 2015), bbmle (Tools and Team, 2022), ggplot2 (Wickham, 2016), and stargazer (Hlavac, 2022).

Table 1. The three linear mixed-models that were tested to describe the relationship between the variation in the tree growth (periodic annual increment – PAI), and the drought indices (Mean negative SPEI), soil fertility (PC1 axis of the soil data), and the random effect at each site and on each tree.
Across all the sites, the mean periodic annual increment (PAI, henceforth tree growth) of E. coriacea varied from 0.4 to 6 mm/year with an overall average of 2.63 ± 0.30 mm/year. The tree growth was related to the soil fertility index (PC1) (r2 = 0.58 and p < 0.001, Figure 4A) and had a negative relationship with wood specific gravity (r2= 0.37 and p < 0.001). The mean wood specific gravity was 0.77 ± 0.01 g/cm3, which varied by 0.69 to 0.89 g/cm3 and had an inverse relationship with the soil fertility index (PC1) (r2 = 0.59 and p < 0.01, Figure 4B).
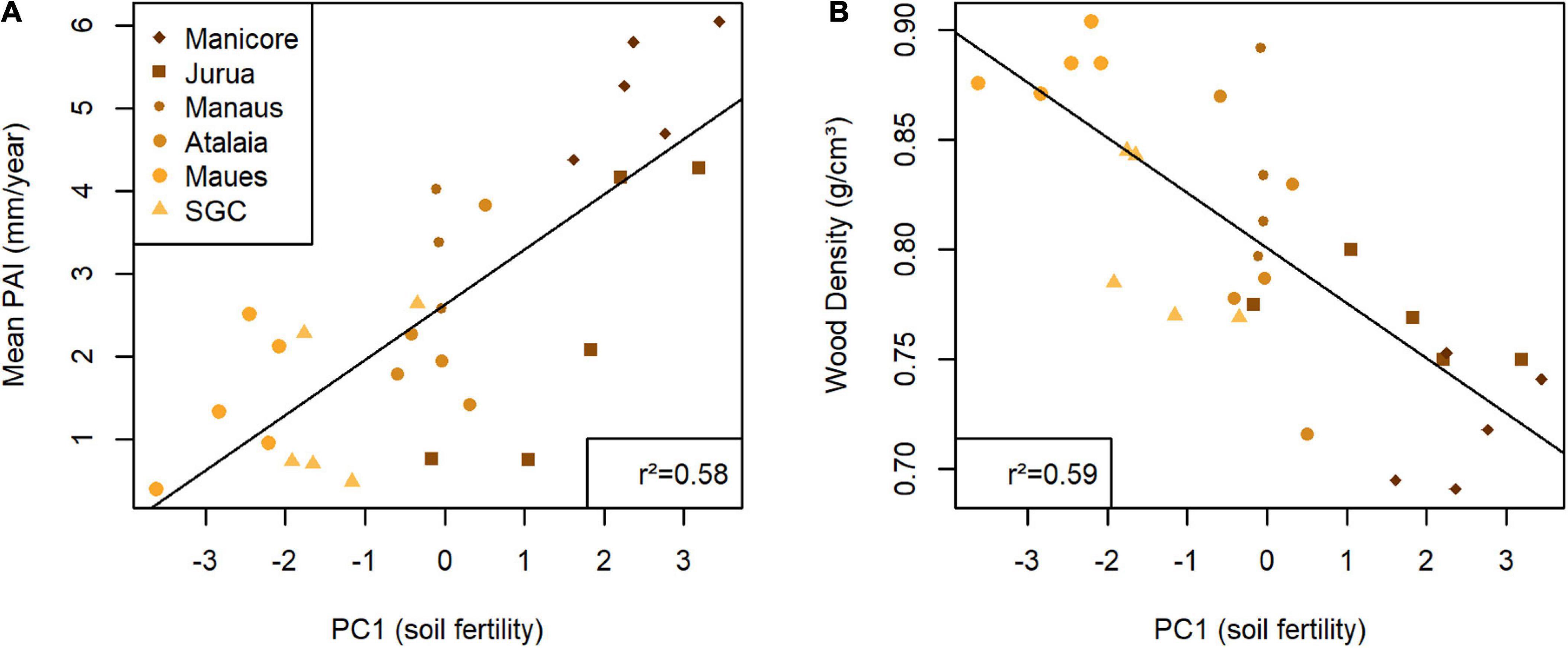
Figure 4. Graphics showing the distribution points between Eschweilera coriacea traits varying by axis 1 (PC1) from the principal component analysis calculated using soil data expressing the soil fertility, were: (A) PAI (mm/year)–Periodic annual increment. (B) Wood density (g/cm3). The black line indicates the tendency line of the regression from each data set. Acronyms of the study sites: Manicore, Manicoré; Jurua, Juruá; Manaus, Manaus; Atalaia, Atalaia do Norte; Maues, Maués; SGC, São Gabriel da Cachoeira.
Using AIC, BIC, and Pr values, the temporal variation in the tree growth was tested via three mixed-effects models to determine the dependent variables affecting this factor (Table 2). By itself, the dependent variable SPEI (drought index) did not explain the tree growth. However, the interaction between soil fertility and drought index is the most explicative model for the temporal growth variation (model 3 and Figure 5). The vast majority of growth responses to drought were negative, decreasing growth when there was a strong drought. However, the effect of the drought on the growth decreased with soil fertility. It is possible to determine these interaction effects via the slope variation between sites (Figure 5). The decreasing growth on fertile soils is abrupt and in infertile soils the tree growth remains slow, and with little variation. However, two trees at the Maués site showed a slightly positive growth response to drought.
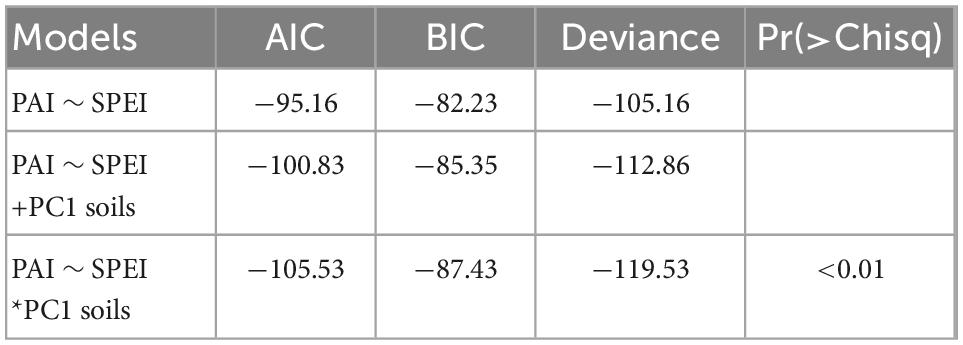
Table 2. Statistical parameters comparing the three linear mixed-effects models tested to describe the relationship between the variations in the PAI (tree growth), SPEI (drought index), and PC1 Soils (soil fertility index).
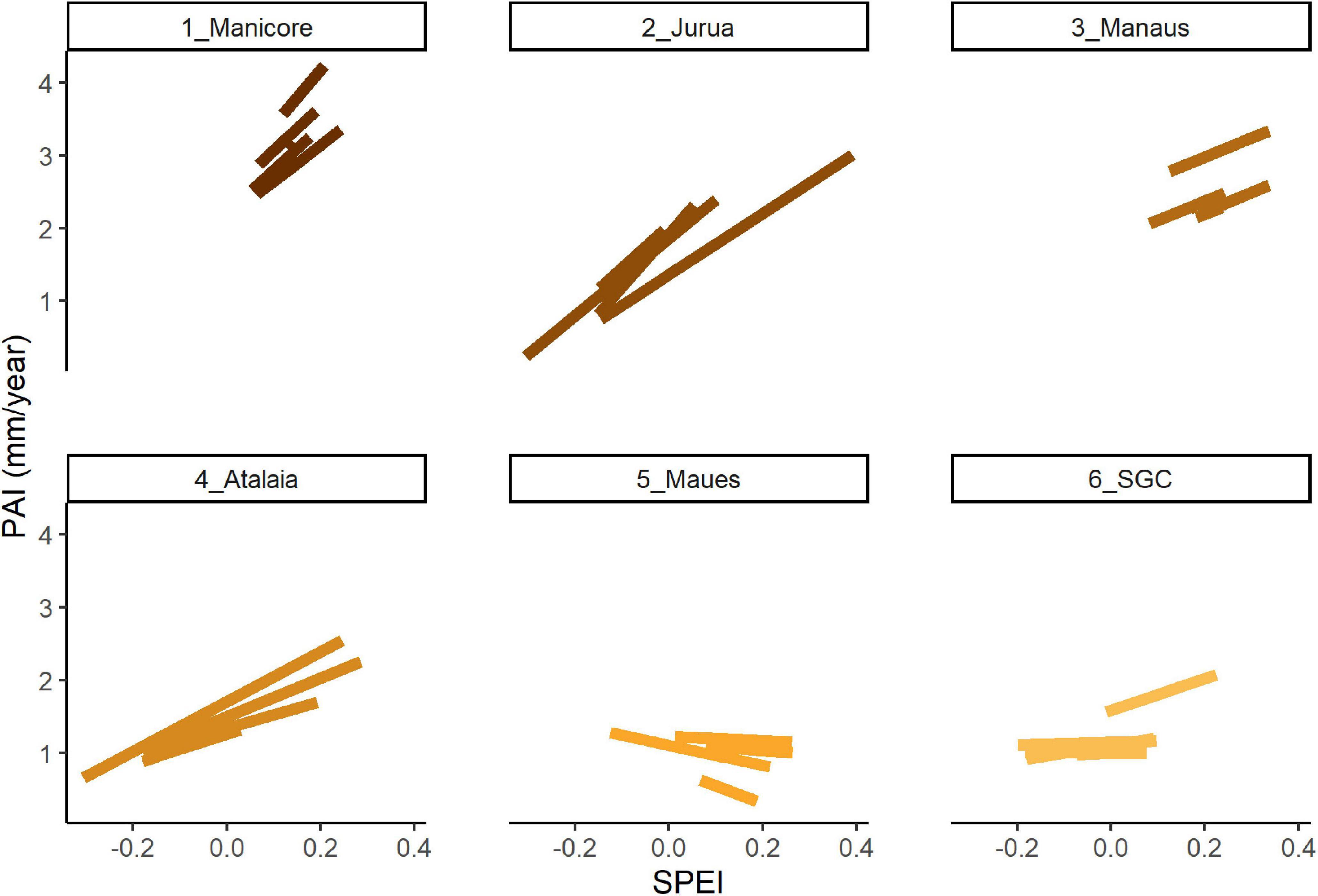
Figure 5. The linear mixed-model predictions show the periodic annual increment (PAI) of Eschweilera coriacea in mm/year responding the interaction between soil fertility and drought index. Sites are distributed in descending order according to the soil fertility gradient. SPEI corresponds to the Standardized Precipitation-Evapotranspiration index (drought index). The trend lines are the values predicted by the linear mixed model for each tree. Acronyms of the study sites: 1_Manicore: Manicoré; 2_Jurua: Juruá; 3_Manaus: Manaus; 4_Atalaia: Atalaia do Norte; 5_Maues: Maués; 6_SGC: São Gabriel da Cachoeira.
Our results prove the strong effect of the interaction between soil fertility and climate on the temporal variation in tree growth of the hyperdominant species Eschweilera coriacea. Trees at sites with fertile soils grew the fastest and those with the least fertile soils grew the slowest. This result agrees with the overall pattern found for the tree community as a whole at the basin scale (Malhi et al., 2004; Quesada et al., 2012). The tendency for trees to decrease wood productivity with drought intensification has already been described in the literature (Brando et al., 2008; Phillips et al., 2009; Doughty et al., 2015; Feldpausch et al., 2016); however, we were able to show that the magnitude of the drought effect differed among sites. Individuals growing in fertile soils decreased their growth rates dramatically with drought intensification, while, in less fertile sites, growth rates remained slow and showed less decline with the increase in the drought index. The drought index on its own does not significantly influence tree growth without the fertile soils effect in our models. Thus, adding the soil fertility index as a dependent variable in the mixed-effects models, we determined that, overall, the nutrient limitation is the strongest factor for limiting the growth of E. coriacea over this large portion of the Amazon. If this is true for other trees as well, the sensitivity of wood production to drought would be expected to be greatest in faster-growing forests.
Many tropical soils are highly weathered, reflecting ancient landscapes exposed to hot and humid climate conditions. Nutrient availability is thus highly dependent on soil age (Quesada et al., 2011). Chemically, the poorest soils are acidic, highly leached, with low cation exchange capacity and high aluminum saturation (Dalling et al., 2016). In these soils, phosphorus and nitrogen are the main limiting nutrients in vegetation biomass production (Quesada et al., 2012; Dalling et al., 2016; Lambers and Oliveira, 2019). In the present study, the macronutrients Mg and K were the most explanatory elements of PCA axis 1, which defined our fertility gradient. These two elements are indispensable for plant growth and are involved in several essential physiological functions. Mg is the central element of chlorophyll and is thus directly involved in photosynthesis. It also has an important function to activate several enzymes and is involved in carbohydrate metabolism and nucleic acid synthesis (Zekri and Obreza, 2003). K is a basic need for growth functions and is required for the formation of sugars and starches, protein synthesis, cell division, and neutralization of organic acids (Zekri and Obreza, 2003). Nutritional deficiency affects several metabolic factors, including the hydraulic conductance of the root system (Minshall, 1975), osmotic adjustment and turgor maintenance (Mengel and Arneke, 1982). The interaction between soils and water deficit was expected because both water and nutrients are important for metabolic processes to occur effectively (Sands and Mulligan, 1990).
Extreme drought events limit water availability and are associated with high canopy temperatures and increased vapor pressure deficit. To survive these conditions, trees use physiological mechanisms such as stomatal control of the leaves which limit carbon uptake and prevent further water loss through transpiration, thus protecting the plant from possible hydraulic failure (McDowell et al., 2008). Decreased growth rate is an immediate response to decreased carbon uptake, as newly fixed C is redirected to survival functions, and reduces secondary growth (Chapin et al., 1990). In addition, trees adjust allocation to anatomical tissues according to environment conditions (Olson et al., 2014). Acquisitive trees with higher growth rates tend to develop larger vessels with lower hydraulic safety (Oliveira et al., 2021), which is represented by low wood density. The negative relationship between growth rate and wood density observed in the present study (r = −0.58 and p < 0.001) indicates intraspecific adjustment in resource allocation in the same species across different environments (Hacke et al., 2001; Chave et al., 2009). The intraspecific variation in wood density reported in this study (0.20 g/cm3) was narrow comparing to tree community level (Chave et al., 2006), however varied with environmental conditions as was showed for another tropical trees species (Schöngart et al., 2005; Wittmann et al., 2006). Future studies regarding the variation in hydraulic traits in this species across the environmental gradients would be important in order to understand how this hyperdominant species, which is present across much of the Amazon basin, adapts to its local environment.
Maués was the second lowest fertile site in our study (see PCA soils in Supplementary Figure 1), where two trees increased their growth during periods with a higher drought index (negative SPEI values). This pattern of increasing productivity in dry years was also observed in forests that have a shallow water table and excess water limits growth (Sousa et al., 2022). We do not have information about the depth of the water table at the Maués site, but our results suggest that more studies monitoring the influence of a shallow water table on tree growth across the Amazon basin are necessary in order to understand this tree growth pattern.
This paper is the first to use radiocarbon data to estimate tree growth rates over periods of multiple decades and examines the underlying reasons for temporal variation in growth. Vieira et al. (2005) used 14C dating to determine growth rates, and found that the results corroborate those obtained using dendrometer bands over the same time periods in permanent plots. This method is applicable for studying tree growth in species and does not rely on visibility or annual tree rings. While we used radii of sections cut for allometry measurements, the small sample size needed for radiocarbon means that it can also be measured using wood cores such as those used for dendrochronological analysis. In the years since about 1960, radiocarbon dating can determine the age of samples to within ±2 years of resolution. Thus, 14C dating is useful in the field of forestry for confirming the annual nature of rings, for independently estimating tree age, and for quantifying temporal variation in growth rates over time.
The tree flora of the Amazon is estimated to contain around 15,000 species (Ter Steege et al., 2020) with some 227 species being hyperdominant species. The evolutionary history for many species in the Amazon has been poorly investigated, especially for hyperdominant species (Heuertz et al., 2020; Damasco et al., 2021). Given the wide distribution of Eschweilera spp. across the Amazon basin, more phylogenetic studies of this genus are necessary because it remains a non-monophyletic group (Huang et al., 2015). For another hyperdominant species (Protium heptaphylum), eight evolving lineages that are adapted to distinct soil and climate conditions were identified (Damasco et al., 2021). We restricted our sites to terra firme plateau areas, which are well-drained forests dominated by clay-rich soils, in order to avoid additional speciation that could be associated with environmental filtering.
The Amazon basin has 227 hyperdominant species that comprise 50% of the trees in the basin (ter Steege et al., 2013). Only 1% of these species account for 50% of the basin’s carbon stock and productivity (Fauset et al., 2015). The species Eschweilera coriacea is a hyperdominant species in the Amazon (Ter Steege et al., 2020) and is ranked second highest in biomass production in the basin (Fauset et al., 2015). As a highly representative species, the patterns of diameter growth in response to soil-climate interactions described in this paper may reflect more general responses for forest trees in terms of their carbon balance, especially in years of extreme drought.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
FD, NH, and ST designed the study. SO, AL, MI, JdS, FW, JH, JS, MP, and BG made scientific contributions to the research. FD, SO, AL, LO, and VC collected data. VC and FD identified the species. FD, ST, XX, SO, CN, AS, YA, and PB performed the laboratory analysis. FD, JH, FW, NH, and ST performed modeling work and analyzed output data. FD wrote the first draft of the manuscript, and all authors contributed substantially to revisions.
We thank for Mr. Francisco Quintiliano Reis (in memoriam) for his more than 30 years of dedication to forest science: studying, teaching and identifying species in the Amazon Forest. His expertise has taught many colleagues, including F. R. de Araujo who also collaborated in the fieldwork and species identification in this study. We thank the CADAF Project and the Science and Technology Research Partnership for Sustainable Development program of the Japan Science and Technology Agency and the Japan International Cooperation Agency for financial support. We also thank INCT Madeiras da Amazonia, who were financially supported by CNPq and FAPEAM, the Max Planck Society for support of the Tacape Project, and Grupo MAUA. As part of the ATTO project, FD, ST, JS, MP, and FW acknowledge support from the German Federal Ministry of Education and Research (BMBF contracts 01LB1001A, 01LK1602A, 01LK1602F, and 01LK2101D) and the Brazilian Ministério da Ciência, Tecnologia e Inovação (MCTI/FINEP contract 01.11.01248.00) as well as the Amazonas State University (UEA), FAPEAM, LBA/INPA, and SDS/CEUC/RDS-Uatumã. We also acknowledge FAPEAM for funding via the Universal Amazonas 030/2013 and Universal Amazonas 006/2019 calls, as well as support from the KIT-Publication Fund of the Karlsruhe Institute of Technology. BG acknowledges the support of the Next Generation Ecosystem Experiments-Tropics (NGEE-Tropics) funded by the U.S. Department of Energy, Office of Science, Office of Biological and Environmental Research through Contract No. DE-AC02-05CH11231 to LBNL, as part of DOE’s Terrestrial Ecosystem Science Program.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2022.1065645/full#supplementary-material
Alvares, C. A., Stape, J. L., Sentelhas, P. C., De Moraes Gonçalves, J. L., and Sparovek, G. (2013). Köppen’s climate classification map for Brazil. Meteorol. Z. 22, 711–728. doi: 10.1127/0941-2948/2013/0507
Andreu-Hayles, L., Santos, G. M., Herrera-Ramírez, D. A., Martin-Fernández, J., Ruiz-Carrascal, D., Boza-Espinoza, T. E., et al. (2015). Matching dendrochronological dates with the Southern Hemisphere 14 C bomb curve to confirm annual tree rings in Pseudolmedia rigida from Bolivia. Radiocarbon 57, 1–13. doi: 10.2458/azu_rc.57.18192
Baker, T. R., Phillips, O. L., Malhi, Y., Almeida, S., Arroyo, L., Di Fiore, A., et al. (2004). Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob. Change Biol. 10, 545–562. doi: 10.1111/j.1365-2486.2004.00751.x
Bates, D., Mächler, M., Bolker, B. M., and Walker, S. C. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Beguería, S., Vicente-Serrano, S. M., Reig, F., and Latorre, B. (2014). Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 34, 3001–3023. doi: 10.1002/joc.3887
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., et al. (2009). Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Brando, P. M., Nepstad, D. C., Davidson, E. A., Trumbore, S. E., Ray, D., and Camargo, P. (2008). Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: Results of a throughfall reduction experiment. Philos. Trans. R. Soc. B Biol. Sci. 363, 1839–1848. doi: 10.1098/rstb.2007.0031
Chambers, J. Q., Dos Santos, J., Ribeiro, R. J., and Higuchi, N. (2001). Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. For. Ecol. Manag. 152, 73–84. doi: 10.1016/S0378-1127(00)00591-0
Chapin, F. S., Schulze, E., and Mooney, H. A. (1990). The Ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 21, 423–447. doi: 10.1146/annurev.es.21.110190.002231
Chave, J., Coomes, D., Jansen, S., Lewis, S. L., Swenson, N. G., and Zanne, A. E. (2009). Towards a worldwide wood economics spectrum. Ecol. Lett. 12, 351–366. doi: 10.1111/j.1461-0248.2009.01285.x
Chave, J., Muller-Landau, H. C., Baker, T. R., Easdale, T. A., Hans Steege, T. E. R., and Webb, C. O. (2006). Regional and phylogenetic variation of wood density across 2456 neotropical tree species. Ecol. Appl. 16, 2356–2367. doi: 10.1890/1051-07612006016[2356:RAPVOW]2.0.CO;2
Clark, D. A., and Clark, D. B. (1994). Climate-induced annual variation in canopy tree growth in a Costa Rican tropical rain. J. Ecol. 82, 865–872.
Cornelius, J. (1994). Heritabilities and additive genetic coefficients of variation in forest trees. Can. J. For. Res. 24, 372–379. doi: 10.1139/x94-050
Dalling, J. W., Heineman, K., Lopez, O. R., Wright, S. J., and Turner, B. L. (2016). “Nutrient availability in tropical rain forests: The paradigm of phosphorus limitation,” in Tropical tree physiology, eds G. Goldstein and L. Santiago (Cham: Springer), 261–273. doi: 10.1007/978-3-319-27422-5_12
Damasco, G., Baraloto, C., Vicentini, A., Daly, D. C., Baldwin, B. G., and Fine, P. V. A. (2021). Revisiting the hyperdominance of neotropical tree species under a taxonomic, functional and evolutionary perspective. Sci. Rep. 11:9585. doi: 10.1038/s41598-021-88417-y
Doughty, C. E., Metcalfe, D. B., Girardin, C. A. J., Amézquita, F. F., Cabrera, D. G., Huasco, W. H., et al. (2015). Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 519, 78–82. doi: 10.1038/nature14213
Fauset, S., Johnson, M. O., Gloor, M., Baker, T. R., Monteagudo, M. A., Brienen, R. J. W., et al. (2015). Hyperdominance in Amazonian forest carbon cycling. Nat. Commun. 6:6857. doi: 10.1038/ncomms7857
Feldpausch, T. R., Phillips, O. L., Brienen, R. J. W., Gloor, E., Lloyd, J., Lopez-Gonzalez, G., et al. (2016). Amazon forest response to repeated droughts. Glob. Biogeochem. Cycles 30, 964–982. doi: 10.1002/2015GB005133
Hacke, U. G., Sperry, J. S., Pockman, W. T., Davis, S. D., and McCulloh, K. A. (2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126, 457–461. doi: 10.1007/s004420100628
Herrera-Ramirez, D., Andreu-Hayles, L., del Valle, J. I., Santos, G. M., and Gonzalez, P. L. M. (2017). Nonannual tree rings in a climate-sensitive Prioria copaifera chronology in the Atrato River, Colombia. Ecol. Evol. 7, 6334–6345. doi: 10.1002/ece3.2905
Heuertz, M., Caron, H., Scotti-Saintagne, C., Pétronelli, P., Engel, J., Tysklind, N., et al. (2020). The hyperdominant tropical tree Eschweilera coriacea (Lecythidaceae) shows higher genetic heterogeneity than sympatric Eschweilera species in French Guiana. Plant Ecol. Evol. 153, 67–81. doi: 10.5091/plecevo.2020.1565
Hlavac, M. (2022). stargazer: Well-formatted regression and summary statistics tables. Bratislava: Social Policy Institute.
Hogg, A. G., Hua, Q., Blackwell, P. G., Niu, M., Buck, C. E., Guilderson, T. P., et al. (2013). SHCal13 Southern Hemisphere Calibration, 0–50,000 years cal BP. Radiocarbon 55, 1889–1903. doi: 10.2458/azu_js_rc.55.16783
Huang, Y. Y., Mori, S. A., and Kelly, L. M. (2015). Toward a phylogenetic-based generic classification of neotropical Lecythidaceae–II. Status of Allantoma, Cariniana, Couratari, Couroupita, Grias and Gustavia. Phytotaxa 203, 122–137. doi: 10.11646/phytotaxa.203.2.2
Kagawa, A., Sano, M., Nakatsuka, T., Ikeda, T., and Kubo, S. (2015). An optimized method for stable isotope analysis of tree rings by extracting cellulose directly from cross-sectional laths. Chem. Geol. 393–394, 16–25. doi: 10.1016/j.chemgeo.2014.11.019
Lambers, H., and Oliveira, R. S. (2019). Plant physiological ecology. Cham: Springer International Publishing. doi: 10.1007/978-3-030-29639-1
Malhi, Y., Baker, T. R., Phillips, O. L., Almeida, S., Alvarez, E., Arroyo, L., et al. (2004). The above-ground coarse wood productivity of 104 neotropical forest plots. Glob. Change Biol. 10, 563–591. doi: 10.1111/j.1529-8817.2003.00778.x
McDowell, N., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., et al. (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 178, 719–739. doi: 10.1111/j.1469-8137.2008.02436.x
Mengel, K., and Arneke, W.-W. (1982). Effect of potassium on the water potential, the pressure potential, the osmotic potential and cell elongation in leaves of Phaseolus vulgaris. Physiol. Plant. 54, 402–408. doi: 10.1111/j.1399-3054.1982.tb00699.x
Minshall, W. H. (1975). Stimulation of transpiration by nitrogenous materials. Can. J. Bot. 53, 1259–1265. doi: 10.1139/b75-151
Mori, S. A., and Lepsch-Cunha, N. (1995). The Lecythidaceae of a central Amazonian moist forest. Mem. N. Y. Bot. Gard. 75, 1–55.
Mori, S. A., Tsou, C. H., Wu, C. C., Cronholm, B., and Anderberg, A. A. (2007). Evolution of Lecythidaceae with an emphasis on the circumscription of neotropical genera: Information from combined ndhF and trnL-F sequence data. Am. J. Bot. 94, 289–301. doi: 10.3732/ajb.94.3.289
Muller-Landau, H. C., Cushman, K. C., Arroyo, E. E., Martinez Cano, I., Anderson-Teixeira, K. J., and Backiel, B. (2021). Patterns and mechanisms of spatial variation in tropical forest productivity, woody residence time, and biomass. New Phytol. 229, 3065–3087. doi: 10.1111/nph.17084
Ohashi, S., Durgante, F. M., Kagawa, A., Kajimoto, T., Trumbore, S. E., Xu, X., et al. (2016). Seasonal variations in the stable oxygen isotope ratio of wood cellulose reveal annual rings of trees in a central Amazon terra firme forest. Oecologia 180, 685–696. doi: 10.1007/s00442-015-3509-x
Oliveira, R. S., Eller, C. B., Barros, F., de, V., Hirota, M., Brum, M., et al. (2021). Linking plant hydraulics and the fast–slow continuum to understand resilience to drought in tropical ecosystems. New Phytol. 230, 904–923. doi: 10.1111/nph.17266
Olson, M. E., Anfodillo, T., Rosell, J. A., Petit, G., Crivellaro, A., Isnard, S., et al. (2014). Universal hydraulics of the flowering plants: Vessel diameter scales with stem length across angiosperm lineages, habits and climates. Ecol. Lett. 17, 988–997. doi: 10.1111/ele.12302
Pereira da Silva, R., dos Santos, J., Tribuzy, S., Chambers, J. Q., Nakamura, S., and Higuchi, N. (2002). Diameter increment and growth patterns for individual tree growing in Central Amazon. Brazil. For. Ecol. Manag. 166, 295–301.
Phillips, O. L., Aragaþo, L. E. O. C., Lewis, S. L., Fisher, J. B., Lloyd, J., Loìpez-Gonzaìlez, G., et al. (2009). Drought sensitivity of the Amazon rainforest. Science 323, 1344–1347. doi: 10.1126/science.1164033
Quesada, C. A., Lloyd, J., Anderson, L. O., Fyllas, N. M., Schwarz, M., and Czimczik, C. I. (2011). Soils of Amazonia with particular reference to the RAINFOR sites. Biogeosciences 8, 1415–1440. doi: 10.5194/bg-8-1415-2011
Quesada, C. A., Lloyd, J., Schwarz, M., Patiño, S., Baker, T. R., Czimczik, C., et al. (2010). Variations in chemical and physical properties of Amazon forest soils in relation to their genesis. Biogeosciences 7, 1515–1541. doi: 10.5194/bg-7-1515-2010
Quesada, C. A., Phillips, O. L., Schwarz, M., Czimczik, C. I., Baker, T. R., Patiño, S., et al. (2012). Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate. Biogeosciences 9, 2203–2246. doi: 10.5194/bg-9-2203-2012
Ramsey, C. B. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360. doi: 10.1017/s0033822200033865
Sands, R., and Mulligan, D. R. (1990). Water and nutrient dynamics and tree growth. For. Ecol. Manag. 30, 91–111.
Santos, G. M., Rodriguez, D. R. O., Barreto, N., de, O., Assis-Pereira, G., Barbosa, A. C., et al. (2021). Article growth assessment of native tree species from the southwestern Brazilian amazonia by post-AD 195014C analysis: Implications for tropical dendroclimatology studies and atmospheric14C reconstructions. Forests 12:1177. doi: 10.3390/f12091177
Schöngart, J., Bräuning, A., Barbosa, A. C. M. C., Lisi, C. S., and Oliveira, J. M. (2017). “Dendroecological studies in the neotropics: History, status and future challenges,” in Dendroecology: Ecological studies, Vol. 231, eds M. Amoroso, L. Daniels, P. Baker, and J. Camarero (Cham: Springer), 35–73.
Schöngart, J., Piedade, M. T. F., Wittmann, F., Junk, W. J., and Worbes, M. (2005). Wood growth patterns of Macrolobium acaciifolium (Benth.) Benth. (Fabaceae) in Amazonian black-water and white-water floodplain forests. Oecologia 145, 454–461. doi: 10.1007/s00442-005-0147-8
Schuur, E. A. G. (2003). Productivity and global climate revisited: The sensitivity of tropical forest growth to precipitation. Ecology 84, 1165–1170. doi: 10.1890/0012-96582003084[1165:PAGCRT]2.0.CO;2
Sousa, T. R., Schietti, J., Ribeiro, I. O., Emílio, T., Fernández, R. H., ter Steege, H., et al. (2022). Water table depth modulates productivity and biomass across Amazonian forests. Glob. Ecol. Biogeogr. 31, 1571–1588. doi: 10.1111/geb.13531
Stuiver, M., and Polach, H. A. (1977). Discussion reporting of 14 C data. Radiocarbon 19, 355–363. doi: 10.1017/s0033822200003672
ter Steege, H., Pitman, N. C. A., Sabatier, D., Baraloto, C., Salomão, R. P., Guevara, J. E., et al. (2013). Hyperdominance in the Amazonian tree flora. Science 342:1243092. doi: 10.1126/science.1243092
Ter Steege, H., Prado, I., Lima, R. A. F., Pos, E., de Souza Coelho, L., de Andrade Lima Filho, D., et al. (2020). Biased-corrected richness estimates for the Amazonian tree flora. Sci. Rep. 10:10130. doi: 10.1038/s41598-020-66686-3
Toledo, M., Poorter, L., Peña-Claros, M., Alarcón, A., Balcázar, J., Leaño, C., et al. (2011). Climate is a stronger driver of tree and forest growth rates than soil and disturbance. J. Ecol. 99, 254–264. doi: 10.1111/j.1365-2745.2010.01741.x
Tools, B. B., and Team, R. D. C. (2022). bbmle: Version, for general maximum likelihood estimation. R package 1.0.25. Available online at: https://cran.r-project.org/package=bbmle (accessed October 04, 1983)
Venables, W. N., and Ripley, B. D. (2002). Modern applied statistics with S. Fourth. New York, NY: Springer.
Vicente-Serrano, S. M., Beguería, S., and López-Moreno, J. I. (2010). A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 23, 1696–1718. doi: 10.1175/2009JCLI2909.1
Vieira, S., Trumbore, S., Camargo, P. B., Selhorst, D., Chambers, J. Q., Higuchi, N., et al. (2005). Slow growth rates of Amazonian trees: Consequences for carbon cycling. Proc. Natl. Acad. Sci. U.S.A. 102, 18502–18507. doi: 10.1073/pnas.0505966102
Williamson, G. B., and Wiemann, M. C. (2010). Measuring wood specific gravity.correctly. Am. J. Bot. 97, 519–524. doi: 10.3732/ajb.0900243
Wittmann, F., Schöngart, J., Parolin, P., Worbes, M., Piedade, M. T. F., and Junk, W. J. (2006). Wood specific gravity of trees in Amazonian white-water forests in relation to flooding. IAWA J. 27, 255–268.
Xu, X., Trumbore, S. E., Zheng, S., Southon, J. R., McDuffee, K. E., Luttgen, M., et al. (2007). Modifying a sealed tube zinc reduction method for preparation of AMS graphite targets: Reducing background and attaining high precision. Nucl. Instr. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 259, 320–329. doi: 10.1016/j.nimb.2007.01.175
Zekri, M., and Obreza, T. A. (2003). Plant nutrients for citrus trees: SL 200/SS419, 1/2003. EDIS 2003. doi: 10.32473/edis-ss419-2003
Zhao, M., and Running, S. W. (2010). Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 329, 940–943. doi: 10.1126/science.1192666
Keywords: Eschweilera coriacea, radiocarbon, SPEI, tree growth, tropical forest, wood density, intraspecific variation
Citation: Durgante FM, Higuchi N, Ohashi S, Householder JE, Lima AJN, Ishizuka M, Wittmann F, dos Santos J, Carneiro VMC, Xu X, do Nascimento CC, Schöngart J, Piedade MTF, Schmitt ARK, Alves YLA, Lehman J, Gimenez BO, Baggio PM, de Ourique LK and Trumbore S (2023) Soil fertility and drought interact to determine large variations in wood production for a hyperdominant Amazonian tree species. Front. For. Glob. Change 5:1065645. doi: 10.3389/ffgc.2022.1065645
Received: 10 October 2022; Accepted: 22 December 2022;
Published: 11 January 2023.
Edited by:
Dashan Wang, Southern University of Science and Technology, ChinaReviewed by:
Luciana F. Alves, University of California, Los Angeles, United StatesCopyright © 2023 Durgante, Higuchi, Ohashi, Householder, Lima, Ishizuka, Wittmann, dos Santos, Carneiro, Xu, do Nascimento, Schöngart, Piedade, Schmitt, Alves, Lehman, Gimenez, Baggio, de Ourique and Trumbore. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Flávia Machado Durgante,  ZmxhdmlhbWRmbG9yZXN0YWxAZ21haWwuY29t, ZmxhdmlhLmR1cmdhbnRlQGtpdC5lZHU=
ZmxhdmlhbWRmbG9yZXN0YWxAZ21haWwuY29t, ZmxhdmlhLmR1cmdhbnRlQGtpdC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.