
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change, 04 November 2021
Sec. Forests and the Atmosphere
Volume 4 - 2021 | https://doi.org/10.3389/ffgc.2021.776905
This article is part of the Research TopicForest Monitoring to Assess Forest Functioning Under Air Pollution and Climate ChangeView all 8 articles
 Roman Zweifel1*
Roman Zweifel1* Sophia Etzold1
Sophia Etzold1 David Basler1,2
David Basler1,2 Reinhard Bischoff3
Reinhard Bischoff3 Sabine Braun4
Sabine Braun4 Nina Buchmann5
Nina Buchmann5 Marco Conedera1
Marco Conedera1 Patrick Fonti1
Patrick Fonti1 Arthur Gessler1,6Matthias Haeni1
Arthur Gessler1,6Matthias Haeni1 Günter Hoch2Ansgar Kahmen2Roger Köchli1Marcus Maeder7,8Daniel Nievergelt1
Günter Hoch2Ansgar Kahmen2Roger Köchli1Marcus Maeder7,8Daniel Nievergelt1 Martina Peter1
Martina Peter1 Richard L. Peters1,9,10
Richard L. Peters1,9,10 Marcus Schaub1
Marcus Schaub1 Volodymyr Trotsiuk1
Volodymyr Trotsiuk1 Lorenz Walthert1Micah Wilhelm1
Lorenz Walthert1Micah Wilhelm1 Werner Eugster5
Werner Eugster5The TreeNet research and monitoring network has been continuously collecting data from point dendrometers and air and soil microclimate using an automated system since 2011. The goal of TreeNet is to generate high temporal resolution datasets of tree growth and tree water dynamics for research and to provide near real-time indicators of forest growth performance and drought stress to a wide audience. This paper explains the key working steps from the installation of sensors in the field to data acquisition, data transmission, data processing, and online visualization. Moreover, we discuss the underlying premises to convert dynamic stem size changes into relevant biological information. Every 10 min, the stem radii of about 420 trees from 13 species at 61 sites in Switzerland are measured electronically with micrometer precision, in parallel with the environmental conditions above and below ground. The data are automatically transmitted, processed and stored on a central server. Automated data processing (R-based functions) includes screening of outliers, interpolation of data gaps, and extraction of radial stem growth and water deficit for each tree. These long-term data are used for scientific investigations as well as to calculate and display daily indicators of growth trends and drought levels in Switzerland based on historical and current data. The current collection of over 100 million data points forms the basis for identifying dynamics of tree-, site- and species-specific processes along environmental gradients. TreeNet is one of the few forest networks capable of tracking the diurnal and seasonal cycles of tree physiology in near real-time, covering a wide range of temperate forest species and their respective environmental conditions.
Although monitoring in forest ecology has a long history (Schimel and Keller, 2015; Richardson et al., 2018; Etzold et al., 2019, 2020; Schulze et al., 2019), the lack of more automated approaches to deal with in situ ecophysiological measurements becomes more obvious due to the increasing data requirements from Earth system models and stakeholder requests. Many forest ecology networks are based on traditional growth measurements to assess forest functioning and rely either on manually recorded long-term data series or automatically measured data stored in decentral loggers and data bases. In some cases, there are sporadic collection initiatives among such networks with the goal of combining and homogenizing existing data (e.g., SAPFLUXNET, Poyatos et al., 2021). However, such datasets typically remain unupdated and are not capable of providing nowcast, a combination of measurements and modeling to assess current conditions.
New data transmission technologies (e.g., the Internet of Things IoT or the Low Power Network LoRaWAN), which have become well established over much of the world in recent years, enable an integrated approach to data collection also in forests and other ecosystems, consisting not only of automated sensors but also allowing near real-time data transmission (e.g., Franz et al., 2018; Sabbatini et al., 2018; Heiskanen et al., 2021).
All field measurements may contain both systematic and non-systematic errors. Hence, the continuous stream of raw data needs to be processed to remove artifacts and data errors before meaningful and biologically relevant information can be extracted (Yenni et al., 2019). In contrast to meteorological measurement methods (e.g., Heylen, 1992), applicable methods for recording physiological responses of a tree to environmental conditions under field conditions are still limited. Point dendrometers are one of the few forest monitoring methods that can be automated because they are weather resistant, require very little energy and provide an integrated signal representing the condition of an entire tree over several years (Zweifel et al., 2006; Drew and Downes, 2009; Steppe et al., 2015).
TreeNet is a forest monitoring and research network that collects and processes dendrometer measurements along broad temperature and precipitation gradients, currently mainly in Switzerland. The overall goal of TreeNet is to generate scientifically sound data of radial stem growth and tree water deficits, but also to enable information on underlying mechanisms of forest trees in near real-time (nowcasting). The increasing intensity and frequency of extreme weather events, i.e., heat and drought (National Academies of Sciences Engineering and Medicine, 2016; Bastos et al., 2020; Buras et al., 2020; Hari et al., 2020; Trotsiuk et al., 2020), but also cold snaps (Mayr et al., 2012; Geng et al., 2020; Fang et al., 2021) and heavy rain events (IPCC, 2021), require an immediate signal from forest ecosystems to document, understand and predict relevant ecological consequences.
Here we provide a detailed description of the extensive TreeNet network. We present the history and locations of the monitoring sites, the field infrastructure, and the detailed data workflow, including data acquisition, data collection, data processing, the theoretical background of the data processing steps, and examples of data outputs. Finally, opportunities, but also measurement uncertainties and error propagations within the collected data will be discussed, with the goal of making our collected knowledge available to the forest science community.
TreeNet (1Figure 1) was established in 2010 and started collecting data in 2011, but also includes dendrometer time series already ranging back to 1998 (site Davos). As a collaboration between the Swiss Federal Office for the Environment (FOEN), the Swiss Federal Institute for Forest, Snow and Landscape Research (WSL), the Institute of Applied Plant Biology (IAP), ETH Zurich, the University of Basel, and the Zurich University of the Arts (ZHdK) the network was continuously expanded. As of today, TreeNet covers 61 sites in Switzerland with about 420 trees and shares sites with Swiss FluxNet2, ICOS-CH3, Long-Term Forest Ecosystem Research (LWF;4), Mycorrhiza Monitoring Network, Soil and Biochemistry field sites (WSL,5), the Lötschental Tree Growth Monitoring Transect (WSL,6), the Swiss Canopy Crane Project II (Uni Basel,7), and the WSL Insubric Ecosystems in Cadenazzo. Furthermore, TreeNet is linked to the international networks ICOS8, UNECE/ICP Forests9, and eLTER10 considered as emerging sites in the 2018 European Strategy Forum on Research Infrastructures (ESFRI) report11 and considered for the Swiss Roadmap on Research Infrastructures 2025–2028 (Eugster et al., 2021). Moreover, TreeNet also contributes data to related networks e.g., the SapFluxNet (Poyatos et al., 2021), and DendroGlobal12.
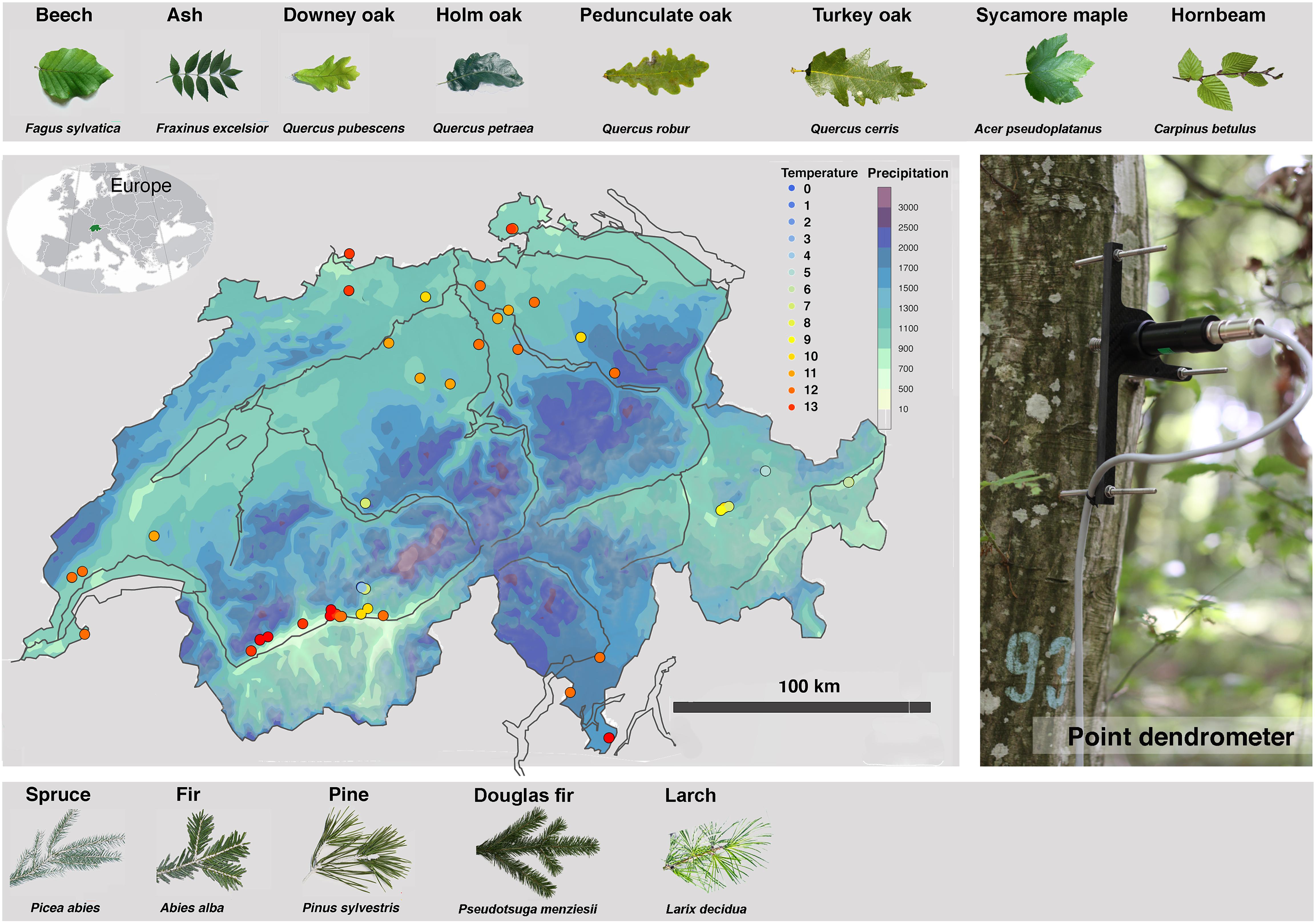
Figure 1. Locations of the 61 TreeNet sites in Switzerland (www.treenet.info) including thirteen tree species of about 420 individually measured trees. Sites are defined as forest patches with a specific species composition and specific environmental conditions. The background color of the map indicates the average annual precipitation. The site dots are color-coded according to the mean annual temperature. Sites cover gradients of mean annual temperature between 0 and 13°C and annual precipitation sum from 600 to 1,800 mm, respectively. The photo shows a point dendrometer mounted on a beech tree. Find further site details in the Supplementary Material (Supplementary Table 1). The background color of the map indicates the average annual precipitation (MeteoSwiss).
The initial collaborations and sites were chosen to cover the widest possible gradient of microclimatic conditions, nitrogen deposition, and elevation of the major tree species in Switzerland. The network is likely one of the largest continuously monitored networks of high-resolution point dendrometers and is continually expanding with partners who have an interest in TreeNet collaboration and progress. The sites are therefore not evenly distributed and show hotspots of forest research in Switzerland (Figure 1), e.g., in the dry, inner alpine Valais, where many research sites pursue questions on climate change and forest dynamics (Rigling et al., 2013; Schönbeck et al., 2018; Walthert et al., 2021). Particularly important are also the super-sites at Hölstein (see text footnote 7), Lägeren13, Pfynwald14 and Davos15 where most extensively a wide variety of additional variables have been assessed and where TreeNet enables partners to extrapolate their site results to a larger area with the help of dendrometer data.
Each field site (Figure 1) is equipped with at least three types of devices (Figure 2): automated point dendrometers (ZN11-T-IP and ZN12-T-WP, Natkon, Switzerland), air temperature and relative humidity sensors (RHT, Sensirion, Switzerland), and soil temperature and soil water potential sensors (TEROS-21, Meter group, United States). Raw data are recorded at 30–60 s execution intervals, averaged and logged at 10-min intervals and transmitted to the respective raw data servers. Further, meteorological variables, i.e., solar radiation, precipitation and wind are not consistently measured at all sites and are additionally derived from gridded data provided by MeteoSwiss and Meteotest. Further, TreeNet analyses benefit from a multitude of additional, site-specific data collected by the site partners, e.g., phenology from automated cameras at the sites Hölstein16, Davos17, and Lägeren18.

Figure 2. Key measurement devices used in TreeNet. Point dendrometer of the type (a) ZN11-T-IP (Natkon) and (b) ZN12-T-WP (Natkon) are mounted to the stem with three stainless steel rods anchored 5–10 cm in the stem wood at about 1.5 m height, (c) air temperature and relative humidity (RHT, Sensirion) are recorded in a weather shield at 2 m height in the forest stand or in a nearby located meteorological station. (d) The combined soil temperature and soil water potential sensors (TEROS21, Metergroup) are located in 10–20 cm soil depth at each site. The TreeNet infrastructure is kept largely harmonized in terms of measurement device types to minimize artifacts caused by different measurement methods.
Data are collected automatically in the field (Figure 2) in a 10-min resolution and continuously transferred to the end user through several steps in near real-time, i.e., stored in databases, cleaned and processed with the R software package treenetproc (R Core Team, 2019; Haeni et al., 2020; Knüsel et al., 2021), filtered, aggregated and visualized in an internet interface with the R software treenetvis19. This fully automated set-up is complemented by user-initiated actions, e.g., the additional manual data cleaning steps to reach the high-quality scientific data sets or the manual download of aggregated data. Figure 3 shows an overview of the different processing steps and color-codes whether they are automated (blue) or user-initiated (red). In the following section, critical processing steps are explained in more detail.
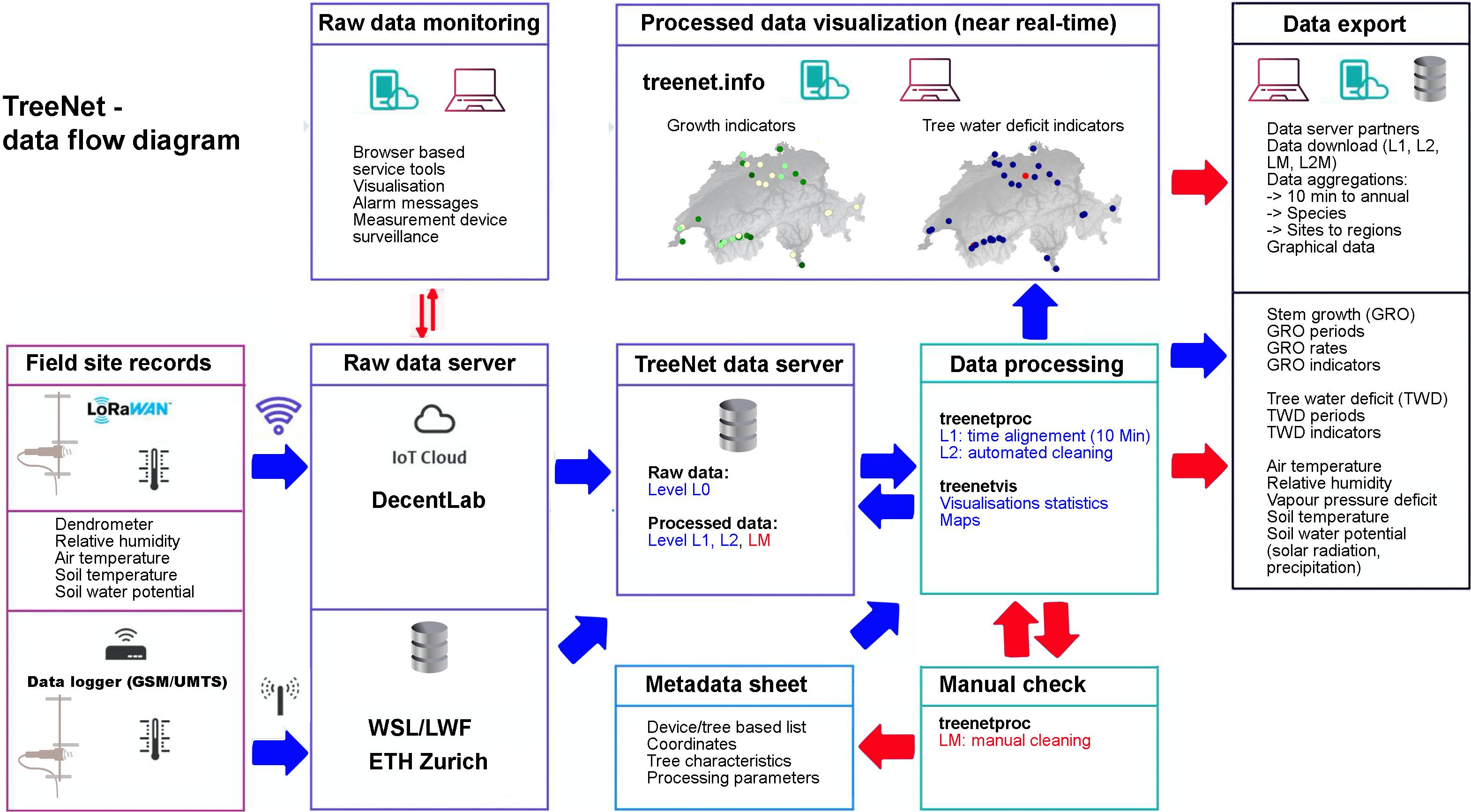
Figure 3. TreeNet data flow diagram from measurement devices in the field to the processed data products, i.e., automated data visualization in the online interface (www.treenet.info) and data output. Blue arrows indicate automated data transfer, red arrows indicate user-initiated data transfer.
Most measuring devices are operated by a LoRaWAN-based logger (Decentlab GmbH, Switzerland) which send the data individually and in near real-time via a LoRaWAN network (Swisscom) or with a local WLAN network and a base station (Decentlab GmbH, Switzerland) via the GSM/UMTS network (Swisscom, Sunrise) to the raw data server. A small number of devices are connected via cables to a central logger (CR1000, Campbell Sci, United States) at the site. The Campbell loggers send the raw data via a GSM/UMTS–connection to the raw data server at WSL and ETH Zurich a couple of times per day. The two ways of data transfer from the field to the raw data server are shown in Figure 3. Although the LoRaWAN-based technology in our case is a pure data transmitter and not a data logger, which means that transmission errors lead to a loss of data, it has the great advantage of being fast in data transmission, connected to a central data server, consuming less power and resources (two 1.5V DC batteries last for >2 years) and making each individual sensor a stand-alone measurement system.
The raw data hosted by Decentlab (about 90% of the data) are accessible via an interactive, browser-based data portal, which allows to setup with user-defined functions, such as an e-mail-based alarm function for low battery status or missing data. The data portal is particularly useful for monitoring the proper functioning of field devices: The raw data can be visualized, aggregated and downloaded in a user-defined resolution (e.g., 10 min, 60 min, 1 day, 1 month, etc.). The download of the raw data as well as its visualization over the entire measurement period is only possible with a user account. A visualization of the raw time series of the last 5 days is openly accessible via https://treenet.info/quickviews-current-raw-data/.
The raw data of the sites with Campbell loggers (Davos, Laegeren, and Lötschental transect) are visually accessible via20 for Davos and21 for Lägeren. A solution for the Lötschental sites is under construction.
The TreeNet processes are operated under two independent Unix-like servers hosted under the WSL IT infrastructure. The data storage server hosts the PostgreSQL database22, where all the raw and processed data from the various sources are stored. The data processing server hosts the automated processing (treenetproc) and visualization (treenetvis). Those processes are run on the regular intraday interval, and are scheduled and executed using crontab (Reznick, 1993; Figure 3). Data are stored and labeled according to their data processing levels: L0-data are raw data as obtained from the data logger in the field, L1-data are time-aligned to 10 min resolution, L2-data are automatically cleaned and further processed into the derivatives (growth GRO, tree water deficit TWD, see also Supplementary Table 2), and LM-data are manually checked. A versioning hierarchy identifies the processed LM data according to the end date of the data set (e.g., “12.2020” contains data until the end of 2020). In addition, each processing step is documented with a version of treenetproc (e.g., V0.1.5). While L1 and L2 datasets are regularly recalculated and updated, LM data are usually produced once a year and remain archived in order to enable the reconstruction of (published) results retrospectively. Finally, there is an additional data level L2M, which combines the existing LM data with the most recent L2 data.
Central to the data processing are the functions collected in the R package treenetproc. treenetproc is currently run twice a day to automatically obtain cleaned L1 and L2 data from the raw data.
In summary, treenetproc collects the raw data (data level: L0) from the TreeNet data server and converts them into time-aligned time series (data level: L1, 10-min resolution), automatically cleans them for outliers and data shifts (Knüsel et al., 2021), and linearly interpolates data gaps < 30 min, and writes the data back to the data server (data level: L2). Linear interpolation makes sense for such short periods of time, as the stem radius changes steadily and relatively slowly. Each change in the data is documented with specific flags assigned to the respective timestamp (Knüsel et al., 2021). Furthermore, the cleaned dendrometer L2 data are separated into their two components, the reversible, water-induced shrinkage and expansion of the radial stem circumference (tree water deficit, TWD) and the irreversible radial expansion due to newly formed cells in wood and bark (growth, GRO) with the zero-growth concept (Zweifel et al., 2016; Figure 4; see also sections “Measurement Devices in the Field” and “Potential Inadequacies Associated With Dendrometer Data”). The detailed functionalities of treenetproc are described in Knüsel et al. (2021) and, in addition to the automatically executed functions, also include features for user-initiated checking, cleaning and processing of time series, e.g., to raise the data quality from L2 to LM data (Figure 3) or to obtain seasonal characteristics as beginning and ending of the stem growth period.
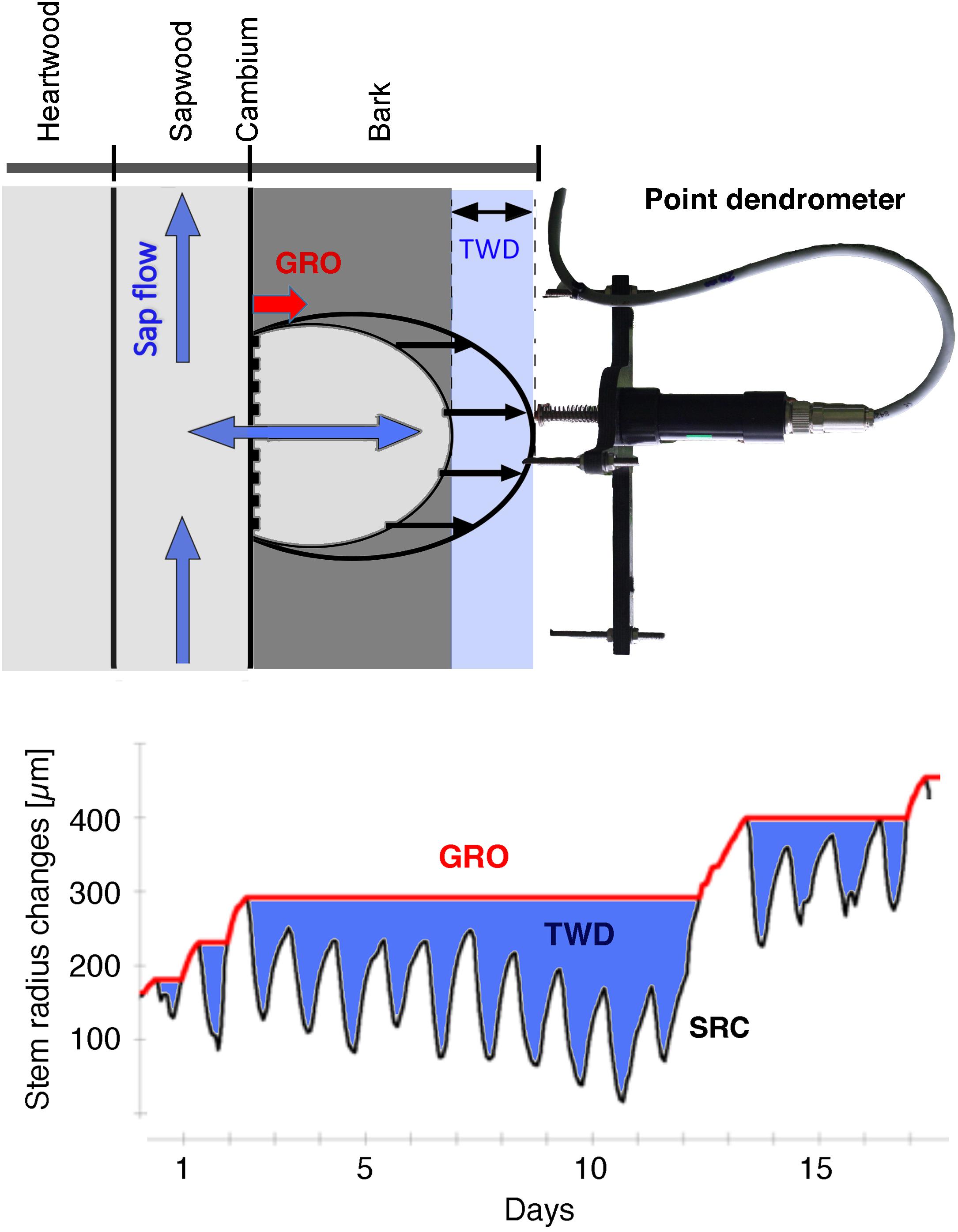
Figure 4. Stem radius changes (SRC; including the bark) measured by a point dendrometer are separated into the fractions tree water deficit (TWD) and growth (GRO) according to the zero-growth concept. TWD is reversible and is induced by changes in plant water potentials. GRO is irreversible and consists of new built cells in the wood (xylem) and in the bark (phloem).
The R-package treenetproc offers a variety of functions to manually check and clean data. See (Knüsel et al., 2021) for further details. Any changes to the data are documented with flags in the produced data set (Supplementary Table 2), ensuring a reproducible data treatment. Quality-controlled (LM) data sets are written back to the TreeNet data server on a user-initiated basis. Further, the manual procedure may lead to improved settings of processing parameters that are manually written to the metadata sheet and become immediately part of the automated data processing with treenetproc.
The metadata sheet contains a device-based list with all available metadata information on the applied measurement methods (e.g., type of sensor, series name, device exposition), the tree it is attached to (e.g., species, tree height, diameter at breast height, bark thickness), and the site the tree is located in (e.g., coordinates, elevation, average environmental conditions, Supplementary Table 1). The unique identifier for each device is the series name, which in the case of Decentlab-based hardware is given by the hardware supplier, comparable to a Media-Access-Control- (MAC-) address of a computer device. The metadata sheet contains all information to locate this MAC address in the field and to relate it to a specific tree.
Additionally, this sheet contains all the necessary parameters used for cleaning and processing the data with treenetproc (e.g., filter settings, threshold temperature for frost conditions, length of period to be gap-filled). The data processing from L0 to LM relies on these parameters and treenetproc queries them every time the processing is executed. In practice, it was found to be useful to make this metadata sheet accessible and editable (as GoogleDoc) for all network partners in order to keep it up to date.
Further, the information about time series that were recorded with different hardware components (e.g., date when the logger was exchanged) and thus had different series names over time is listed in the historical order of appearance in the metadata sheet. This information allows user queries with treenetproc that contain only one of the series names but leads to the return of correctly merged time series over the entire requested time period (download function of treenetproc).
TreeNet supplies its network partners with processed data. L2 and LM data are automatically exported to the partner’s data server or can be downloaded manually. The download function of treenetproc (download_treenet) enables an automatic merging of LM and L2 data to obtain the best available data quality (L2M) supplemented by the latest, automatically processed L2 data. Also in this case, the essential information for the correct merging of fragmented time series is automatically taken from the metadata sheet. A list of the available variables of the data of the different levels (L0 to LM) can be found in Supplementary Table 2 and is partially visualized in Figure 3.
Point dendrometers measure stem radius changes in high spatial resolution of a few micrometers. We here apply the zero-growth concept (Zweifel et al., 2016) to turn cleaned stem radius change L2 data into physiologically meaningful measures for growth (irreversible stem increment, GRO) and tree water deficit (reversible, water related swelling and shrinking of the stem, TWD) (Figure 4). The theoretical background is based on the functional linkage of the tree water relations with the conditions that are needed for cell division and cell expansion (Zweifel et al., 2016). The core of the proposed mechanism is the soil-plant-atmosphere continuum (SPAC) including the cohesion-tension theory (Dixon and Joly, 1894), which links soil water dynamics with tree hydraulics and atmospheric water demand. The movement of water upward along the tree but also in and out of elastic water storing tissues depend on the water potential gradients within the SPAC. Water potentials also affect the turgor pressure in the lateral/secondary meristem, and the turgor pressure in the cambium ultimately determines whether new wood and bark cells can be formed (Lockhart, 1965; Woodruff and Meinzer, 2011; Peters et al., 2021b). Solar radiation, vapor pressure deficit (VPD) of the air and soil water potential (SWP) are the most important factors for determining transpiration (Penman, 1948; Monteith, 1965; Monteith and Unsworth, 1990) and thus directly alter the water potentials in the tree tissues. The changing water potentials themselves affect the turgor pressure in a tree’s meristems, making, VPD and SWP key limiting factors for the turgor pressure and thus growth (Zweifel et al., 2021).
Accordingly, the zero-growth concept assumes GRO to only occur under water potential conditions in the stem that do not initiate stem shrinkage (Zweifel et al., 2016). Technically, GRO is equivalent to an irreversible, incremental increase in stem radius when the measured radius gets larger than it was at any time in the monitored past. Any shrinkage or expansion of the stem below the previously reached maximum radius is handled as a reversible change in TWD (Figure 4).
Importantly, we use the term “growth” (GRO) to refer to the irreversible radial expansion of the tree stem (i.e., cell division and cell enlargement), neglecting all accompanying stem growth processes (e.g., cell wall thickening) (Cuny et al., 2015; Rathgeber et al., 2016; Hilty et al., 2021). In addition, it is important to note that GRO contains wood and bark cells and is therefore different from tree ring data that focus exclusively on stem wood. The proportions of bark and wood cells in GRO cannot be distinguished without further effort and varies largely between species and growth conditions. The smaller the total annual increment is, the potentially larger is the fraction of bark cells (Gricar and Cufar, 2008).
Reliable visualization of data in near real-time requires additional data filtering to avoid misleading visualization results of automatically cleaned but still error-prone L2 data. The R package treenetproc was developed to correct data errors, but is not able to solve all cases in every time series. One of the reasons for this is the large variety of correctly measured biological responses, which are difficult to separate from artifacts that occur in the same order of magnitude of measurements. For example, the fluctuations in stem radius in winter can be ten times the fluctuations in the growing season. This is due to frost processes that cause enormous changes in stem tissues (Zweifel and Häsler, 2000; Sevanto et al., 2012; Charrier et al., 2017). Especially the bark experiences a great shrinkage when the trunk starts to freeze. The shrinkage process is thought to protect the living stem tissue from frost damage by drawing water out of the sensitive cells, which lowers the water content of the tissue while increasing osmolality. Both in combination lower the temperature to initialize ice formation and additionally result in smaller ice crystals with less potential for damage to living tissue (Single, 1964; Kitaura, 1967; Sevanto et al., 2012).
Some artifacts in L2 data remain, despite the fact that treenetproc relies on temperature-dependent parameters (Knüsel et al., 2021). Since these artifacts, even in a small number only, would strongly affect the automated data visualization of GRO and TWD indicators, they have to be filtered out by a plausibility check. The R script treenetvis was developed for this task and to aggregate and visualize TreeNet data (nowcasting). It uses the best available data quality, i.e., each time series is a combination of historical LM (where available) and the most recent L2 data (L2M). The data aggregation level for this visualization is daily. Time series are filtered out if the current GRO or TWD value exceeds the smoothed 100% percentile (stat_smooth of the R package ggplot2) of the LM data by a factor of 2. Only after this filtering step are the remaining time series used for visualization.
The visualization of the GRO and TWD data of TreeNet with treenetvis follows the idea of presenting relative performances rather than absolute values. This has the great advantage that generally slow-growing tree individuals or tree species (e.g., at generally dry sites) can be compared directly with fast-growing ones. Or, in other words, the current GRO and TWD performances are not assessed on the basis of their absolute changes, but on the basis of their current value in relation to the historical performance of a tree individual at a given point in time. Each tree can perform “poor/low” (<25% percentile), “below average” (25–50% percentile), “above average” (50–75% percentile), and “very good/high” (>75% percentile) regardless of its absolute overall (growth) performance (Figure 5). This type of data processing also allows for the detection of unusual seasonal deviations from the norm in the current GRO and TWD data. Additionally, periods outside of the potential growth period are labeled with “off season.”
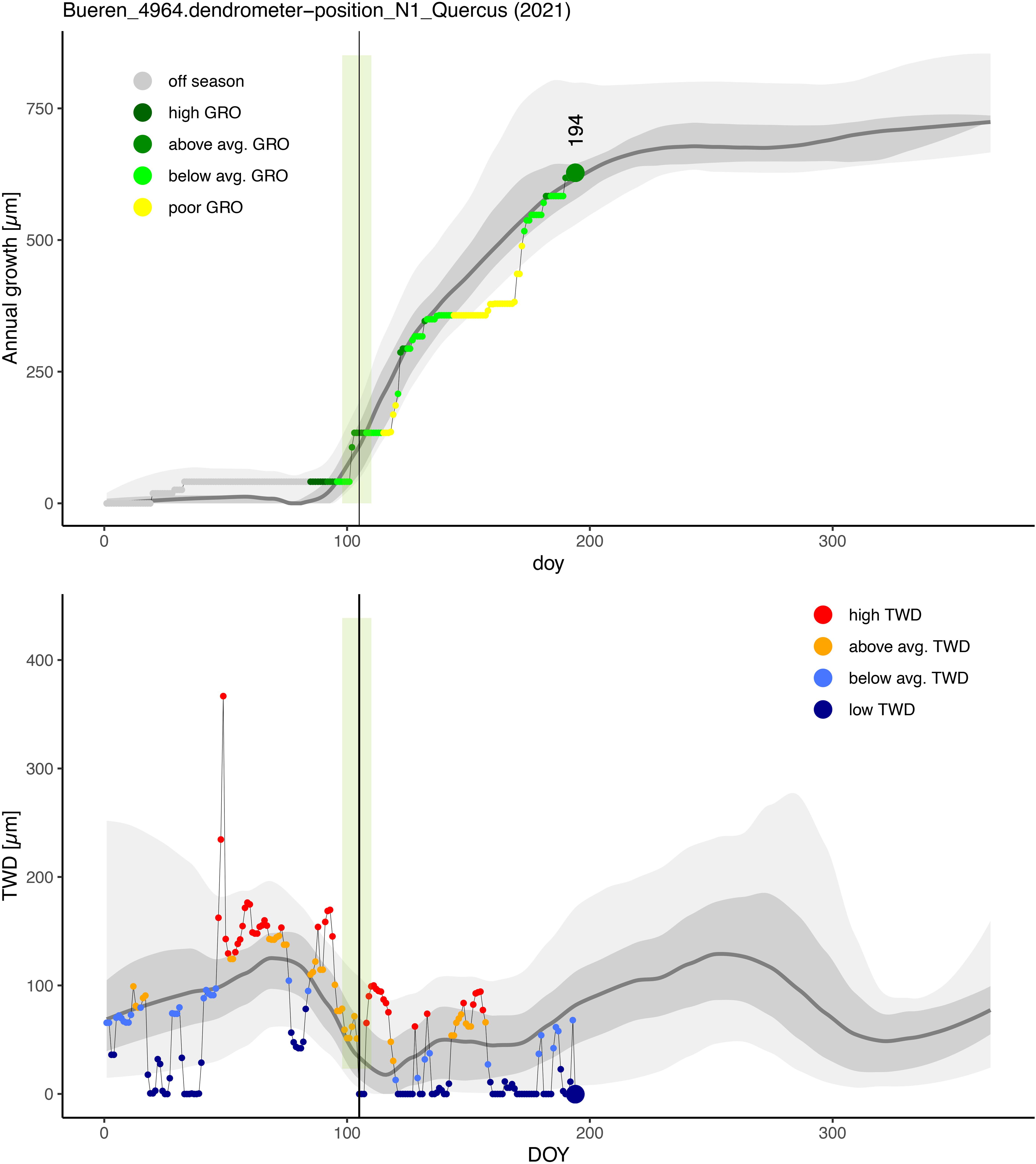
Figure 5. Performance of an oak tree at the site Bueren until the day of the year (DOY) 191 in the year 2021, in terms of stem growth (GRO, upper panel) and tree water deficit (TWD, lower panel). The grey colored areas indicate the 5–95% percentile range (light grey) and the 25–75% percentile range (dark grey) of all available years (L2M data). The grey line indicates the median of the respective data. The colored symbols (L2 data) indicate whether a tree is currently growing poorly (yellow, <25% percentile), below average (light green, 25–50% percentile), above average (green, 50–75%) or highly (dark green, >75%) relative to the years before. The period outside the potential stem growth time is indicated with grey circles and the label ‘off season’. Analogue for TWD, low TWD (black blue, <25% percentile), below average (light blue, 25–50% percentile), above average (orange, 50–75% percentile) or high TWD (red, >75% percentile). The vertical black line with the green area indicates the median DOY when stem growth of a season crosses the 5% threshold of annual growth (based on historic LM data).
The daily data of each individual tree, as shown in Figure 5, were aggregated to GRO and TWD performances of sites (Figure 6) or sites and species (Figure 7) according to the four performance levels described above. As an example, the exceptionally wet summer of 2021 resulted in predominantly water saturated trees with TWD closer to zero or well below historical TWD values at this time of the year (mid of July). Radial stem growth status, on the other hand, appeared to be rather below average overall. Increased growth was only observed in the driest parts of Switzerland (Figure 1) at this time of the season (Figure 6), e.g., in the inner alpine valleys in the southwest of the country (Valais). Considering the generally delayed growth response of trees to changing environmental conditions (Ogle et al., 2015; Zweifel and Sterck, 2018; Kannenberg et al., 2019; Bastos et al., 2020; Bose et al., 2020; Pappas et al., 2020; Zweifel et al., 2020; Fang et al., 2021), a uniform rapid growth response to wet conditions was not expected. However, trees growing normally under most drought-stressed conditions appeared to benefit more immediately in terms of an above-average growth performance.
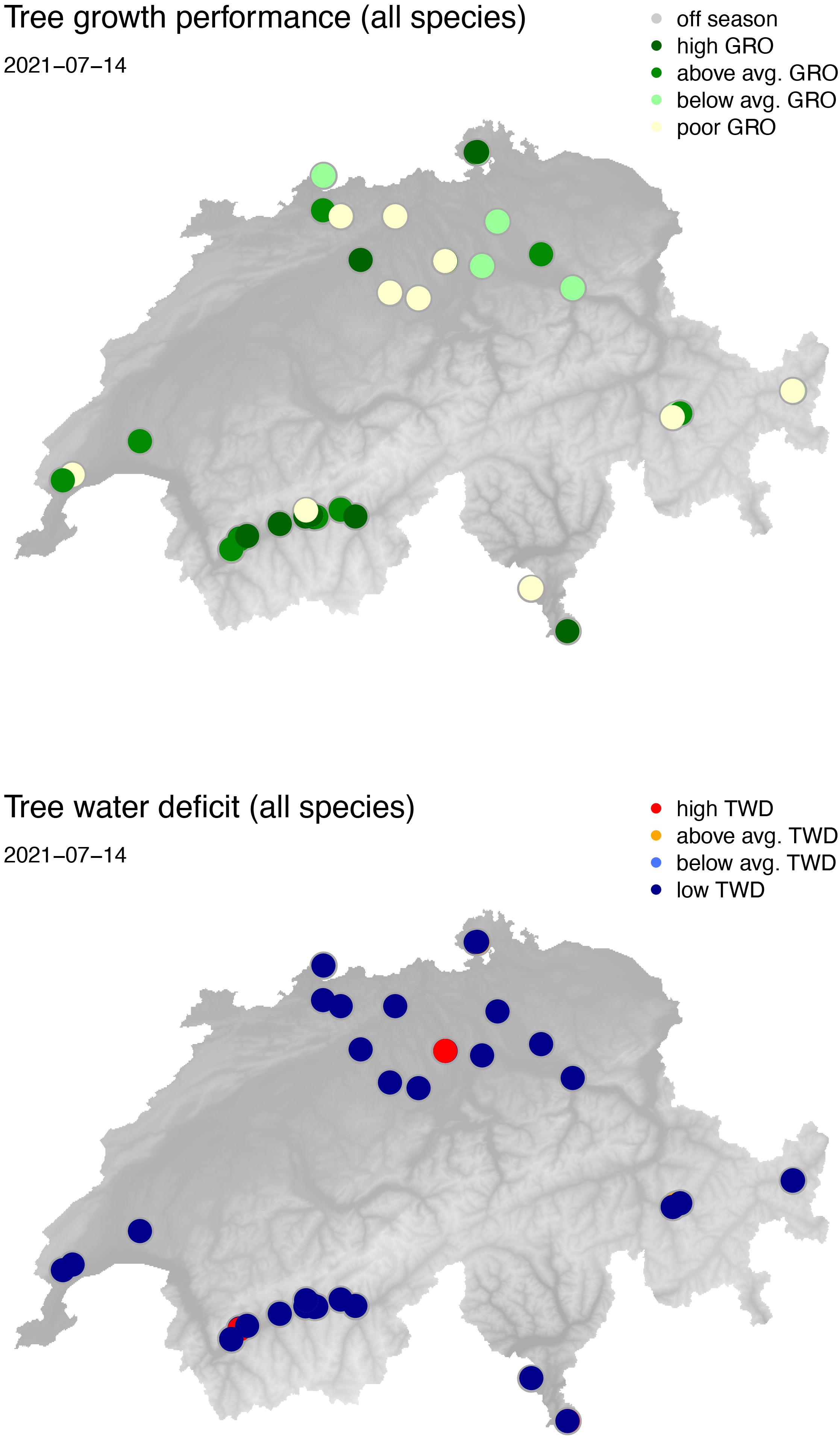
Figure 6. Stem growth (GRO) and tree water deficit (TWD) status of all available TreeNet sites on 14 July 2021. Upper map shows the color-coded growth performance, the lower map shows the same for TWD. Actual maps can be found under https://treenet.info. Color codes correspond to the ones shown in Figure 5.
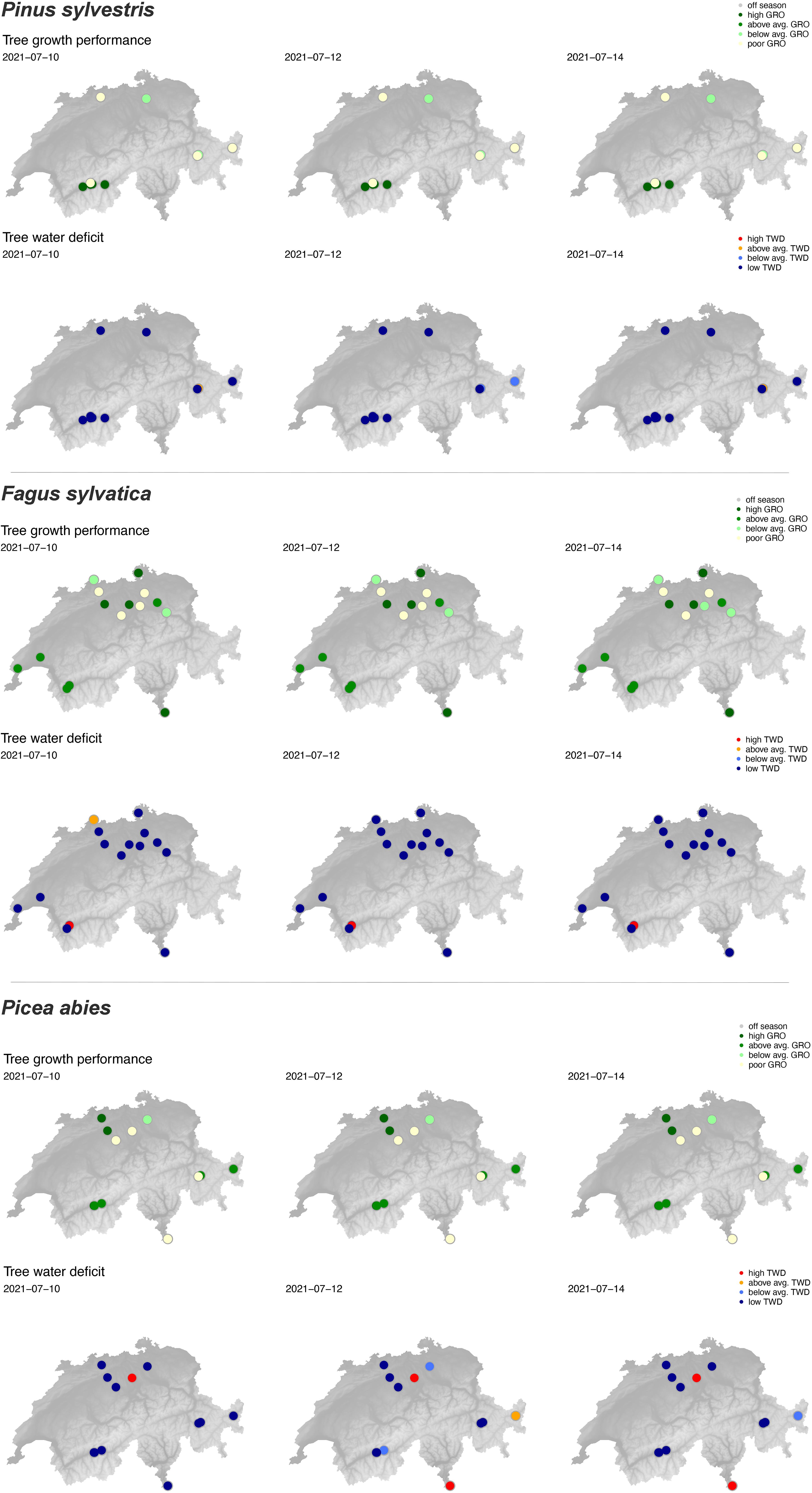
Figure 7. Examples of species-specific stem growth (GRO) and tree water deficit (TWD) status at the available TreeNet sites on 10, 12, and 14 July 2021. Upper maps show the color-coded growth performance, the lower maps the same for TWD.
The general TWD and GRO response patterns (Figure 6) were confirmed by the species-specific patterns (Figure 7). Spruces showing strongly increased TWD (red dots) and poor growth performance (yellowish dots) at some sites (Figure 7) may indicate already damaged tree individuals that are physiologically unable to recover and grow, although water is available in abundance.
Although all processing steps presented ensure the high quality of TreeNet data and analyses, potential uncertainties and discrepancies in this and other dendrometer studies must be considered to improve our understanding of tree physiological processes at high temporal resolution. Uncertainties in high temporal resolution dendrometer data can occur due to technical deficiencies or differences in data processing methods.
Technical shortcomings include, for example, the electronic and mechanical temperature behavior of dendrometers, the way the sensor is anchored in the tree, or an unstable construction of the dendrometer. Important for an accurate dendrometer is the combination of all components including the cable, the power source and the data logger used. The temperature sensitivity of the electronic part of a dendrometer alone, as given in many dendrometer studies, is not a sufficient characteristic to judge its quality. Furthermore, we have found that performing temperature corrections is only useful if the temperature measurement accurately reflects the sensor (and not environment in a nearby meteorological station) and the temperature-related artifact is linear. The point dendrometers used in TreeNet have a temperature sensitivity of −0.2 to −0.3 μm K–1 (including power source, cable and recording device) which was not further corrected.
Temperature-related changes in the size of the wood with temperature are another potential source of uncertainty. The reported coefficient of expansion of 10−30 * 10−6 K–1 for wood (Goli et al., 2019) would theoretically affect a 20 cm diameter tree stem with a day-night temperature change of 10°C by about 20 μm. This is quite big in relation to a normal daily fluctuation of 100−200 μm. However, note that the temperature expansion coefficient is usually determined for dry wood (5−15% moisture content) with no such coefficients existing for moist, live tree stems, including bark. As such, a 10°C change in air temperature results in a smaller temperature change in the stem because the sap transported in the wood cools the stem during the day (Majdak et al., 2021). Furthermore, any size changes due to temperature affecting the wood deeper in the stem than the anchoring of the dendrometer takes place (about 5−10 cm in the case of the dendrometers used in TreeNet) is likely irrelevant as the dendrometer reading is not affected. The negative temperature response of the dendrometers used also means that the positive temperature-related size effect of the stem is partially compensated. Therefore, we assume that the temperature effect is less relevant for the quality of the dendrometer data (during non-freezing periods) than the expansion coefficient applied to the total stem size would suggest. We assume that radial stem changes in the order of 1−5 μm represent the maximum reliable resolution within TreeNet, while the measurement system provides resolutions in the order of one hundredth of a micrometer.
Another problem can be tree species with thick bark and a crown that does not keep water away from the stem (e.g., pine). Here, hygroscopic effects of the bark can occur when the stem surface is moistened by rain (Oberhuber et al., 2020). This can be remedied by removing dead bark at the measuring point of the dendrometer on the stem, as is done in the TreeNet infrastructure. This also helps against artifacts caused by drying and bending pieces of the outermost dead layer of bark. Hygroscopic and bark bending effects are generally more of a problem with band dendrometers that span the entire stem circumference and are therefore more difficult to avoid, which is less the case with the point dendrometers used here.
In addition, there is uncertainty about the largely unknown processes of bark degradation or phloem collapse (Gricar et al., 2015; Güney et al., 2020), especially after frost in winter, which induces large changes in stem size (Zweifel and Häsler, 2000; Charrier et al., 2017). Bark degradation may falsely delay the determined growth initiation in spring obtained with the zero-growth concept, as the missing (degraded) tissue is interpreted as a reversible water deficit of the tree and not as an irreversible structural volume loss.
Uncertainty also remains with the assumption of the zero-growth concept that no growth takes place in the case of stem shrinkage, which means that the turgor necessary for growth is not achieved (Zweifel et al., 2016) and no cell expansion is possible under such conditions. Even though the underlying concept of Lockhart (1965) is theoretically sound and many recent papers have provided evidence for its usefulness (Dietrich and Kahmen, 2019; Schafer et al., 2019; Aryal et al., 2020; Eitel et al., 2020; Güney et al., 2020; Lamacque et al., 2020; Pappas et al., 2020; Sellier and Segura, 2020; Dukat et al., 2021; Meng et al., 2021; Nehemy et al., 2021; Pierrat et al., 2021; Zweifel et al., 2021), the separation of dendrometer data into GRO and TWD has not yet been tested with an independent method. The main reason for this lack of rigorous testing is that there is no alternative method capable of cross-checking hourly resolved dendrometer data in mature forest trees over a reasonable period of time (i.e., without excessive destructive sampling). Zweifel et al. (2016) estimated that > 95% of growth can be attributed to the correct hour using the zero-growth approach, a very robust but not perfect separation approach. Furthermore, they concluded that even with a lower temporal accuracy of the zero-growth concept, the timing of growth is not significantly affected because the pressure drop in the tree (lowered water potentials) and the shrinkage of the stem occur so rapidly in the morning hours that a slightly different turgor threshold is of little consequence (Zweifel et al., 2021).
There are alternative approaches to obtaining physiologically interpretable data extracts from dendrometers, but these are also subject to a number of uncertainties and assumptions. The approach of Mencuccini et al. (2017), which aims to separate GRO and TWD from dendrometer and sap flow data, does not provide a real alternative to validate the assumptions associated with the zero-growth concept, as the approach cannot be tested against independently measured growth data either. The remaining uncertainties of Mencuccini’s approach are the same as for the zero-growth concept, and the calculations additionally require sap flow data (using the thermal dissipation method), which are not consistently available in TreeNet (and also in other measurement networks) and may also contain methodological inconsistencies (Peters et al., 2018, 2021a). Similarly, the approach of Deslauriers et al. (2007, 2011), which treats each individual day independently of the historical trend of stem radius changes, is not better suited to separate dendrometer data into GRO and TWD, as in this approach any absolute increase in stem size over 24 h is recorded as growth. This means that even longer periods of rehydration are counted as growth if a given day starts with a remaining TWD from the previous day. All daily increments determined with the zero-growth approach add up to an annual increment that matches the total annual change in stem size measured by the dendrometer, whereas in the case of Deslauriers’ approach the daily increments add up to more than the increment measured by the dendrometer over 1 year.
Also poorly suited to the analysis of high-resolution dendrometer data is the use of the Gompertz growth function (Gricar et al., 2008), which is based on a general growth trajectory. The Gompertz function assumes constant and uniform growth throughout the growth phase, which is generally not the case, as shown for wood formation by Cuny et al. (2015), but is particularly incorrect for the growth of trees with water limitation (Zweifel et al., 2021). Furthermore, the Gompertz function is not a reliable method for determining the onset of growth, as it neglects the fact that the stem is first rehydrated before growth starts in spring.
Biologically based data are generally subject to a variety of potential artifacts, but the process of cleaning the data is also a potential source of error and can increase the uncertainty of the derived results. First, by failing to detect the artifacts present, and second, by failing to adequately correct for a known artifact (García Criado et al., 2020). Manual data cleaning is still a time-consuming necessity and involves arbitrary expert decisions, such as the placement of the first data point after a data gap. The TreeNet method for converting raw data into cleaned and processed data is described above (Section “From Stem Size Changes to Physiologically Meaningful Measures”) and follows the functionality of the R package treenetproc (Haeni et al., 2020; Knüsel et al., 2021). Manual data cleaning is still necessary to achieve clean scientific data sets, as the functions of treenetproc cannot detect all errors and artifacts. However, the time required to manually check the data is less and, most importantly, all data manipulations are systematically documented, ensuring a high level of data quality, transparency and reproducibility. As a side note on the practical handling of point dendrometers: We recommend never touching a dendrometer during a data interruption until the data flow is restored. In this way, the data gap remains, but the absolute change of the stem radius is measured correctly.
TreeNet is a research and monitoring network that aims to automatically collect and analyze dendrometer data with high temporal resolution over large spatial scales. TreeNet’s setup is particularly well suited for relative comparisons of the temporal and spatial dynamics of tree water relations and tree growth, which is also reflected in the official title of the network: TreeNet–the biological drought and growth indicator network. TreeNet processes data for science-based research on the one hand, and for online visualization of drought and growth indicators in near real-time for a wider audience on the other (nowcasting). This requires not only reliable sensor technology and data transmission that can be operated anywhere with low energy consumption, but also a data workflow that detects errors in the data and provides for automated cleaning and evaluation steps.
The keystones to establish this in TreeNet were (i) the use of high-precision dendrometer and LoRaWAN-based loggers to make each sensor an individual data provider with minimal power consumption; (ii) the development of reliable, automated processing, filtering and visualization functions (R-scripts treenetproc and treenetvis) and (iii) the establishment of a data processing system that uses individual processing parameters for each dendrometer, which led to a significant improvement of the data cleaning functions.
Growth (GRO) as well as tree water deficit (TWD) have proven to be the most promising physiological variables derived from dendrometer data that allow for the assessment of the current state of forests. However, our ∼10-year data set also showed that both GRO and TWD have large seasonal and tree-individual differences that make it challenging to directly compare absolute values of trees growing over wide gradients of site conditions. For our online visualization, we therefore calculated growth and drought indicators that represent the current tree status relative to historical conditions at the same time in the season. Of course, this approach can only be applied meaningfully if one has multi-year time series at one’s disposal.
There is still room for improvement in the automated cleaning of the measurement data. For the daily visualization of GRO and TWD indicators, the automatically cleaned data need an additional filtering to avoid misleading output. For the scientific analyses and interpretations of the data, manual post-processing is still required to clean up data errors that have not been recorded. We foresee to get a better handle on this shortcoming in the future with novel machine-learning methods.
From an economic point of view, such an integrated monitoring and analysis system not only provides the basis for so-called nowcasting, by which is meant the combination of near real-time measurements and a modeling approach to assess the current situation. It also significantly reduces the manpower required to maintain the infrastructure while increasing data quality. Since any measurement error is immediately detected online, no time is lost until it can be repaired and it allows ad hoc diagnosis of the potential damage (e.g., power failure indicates low battery power; faulty data indicates a defective cable or sensor; no signal indicates a defective LoRaWAN device). Together, such an integrated system more than makes up for the higher installation costs compared to conventional logging systems that store data in a decentralized manner and where it has to be collected and combined manually on a regular basis. The same applies to the maintenance costs, which are continuously incurred per measurement channel but are more than offset by the greatly reduced manpower costs. The technical maintenance of the TreeNet infrastructure (a total of approx. 750 measuring channels, over a large and partly impassable area of 300 km × 150 km) could be managed in the last 10 years with less than a 100% position including the help of a data scientist. Not counted is the scientific work to develop, implement and improve the system.
More than 10 years of TreeNet dendrometer and climate data are now available, and their analysis has not yet been exploited. This huge dataset along a wide gradient of environmental conditions provides an excellent opportunity to gain novel insights into tree-climate relationships for all major forest tree species in Europe or to test models simulating the climate-growth dynamics. In addition, TreeNet provides a unique “ground-truthing” platform for testing drone- and satellite-based indicators. Last but not least, the presented TreeNet concepts can help to establish other networks with similar requirements with the aim to better link and add value to monitoring data.
Selected datasets from TreeNet can be found in the repository Pangea: doi.pangaea.de/10.1594/PANGAEA.935394 (Zweifel et al., 2021) doi.org/10.1594/PANGAEA.918631 (Zweifel et al., 2020) doi.org/10.1594/PANGAEA.859055 (Zweifel et al., 2016) doi.org/10.1594/PANGAEA.859051 (Zweifel et al., 2016) doi.org/10.1594/PANGAEA.859048 (Zweifel et al., 2016) doi.org/10.1594/PANGAEA.859037 (Zweifel et al., 2016).
WE and RZ initiated and established the network. RZ, MH, DB, SE, RK, LW, PF, DN, MP, MC, MM, RP, and RB set up the infrastructure in the field. MH, MW, VT, RB, RP, and RZ designed and realized the software and data base processes. SE, LW, and RZ observed and improved the data quality. SE and LW supplied the environmental data. NB, WE, AG, AK, GH, MS, MM, PF, SB, MC, LW, MP, RP, and RZ acquired the funding and established the scientific exchange between the different networks connected by TreeNet. RZ wrote the first draft of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Federal Office for the Environment FOEN, project TreeNet infrastructure and data processing, grants 18.0031.PJ/E2C9DDCC1 (MW and RZ), 00.25992A/E3D4E3424 (SE and RZ), 00.25992A/08B6EC6C3 (SE and LW), and 18.0031.PJ/E2C9DDCC1 (RZ). SwissForestLab: project: SCC II – REACT, grant: WSL, Grant receiver: GH and RZ. The Intercantonal Monitoring Network of IAP is funded by the Swiss cantons AG, BL, BS, GR, LU, NW, OW, SO, SZ, TG, UR, and ZH). Swiss National Science Foundation SNF: project ICOS-CH Phase 1 (20FI21_148992), Phase 2 (20FI20_173691), Phase 3 (20FI20_198227), Grant receiver: NB and RZ Swiss National Science Foundation (SNF), grant: P2BSP3_184475, Grant receiver: RP. WSL, major investment grant 2020, and Grant receiver: PF, AG, and MS.
RB is a co-owner of the company Decentlab Ltd., which supplied most of the logging infrastructure of TreeNet. RZ is owner of the private company developing and supplying the Natkon point dendrometers.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank the TreeNet project partners Swiss Long-term Forest Ecosystem Research Programme LWF, The Intercantonal Monitoring Network of IAP, the National Air Pollution Monitoring Network NABEL, ICOS-CH, and MeteoSwiss for their assistance with site infrastructure and providing data. We would also like to thank Nicolas De Girardi, Käthi Liechti, Philip Meier, Thomas Baur, Stephanie Pfister, and Moise Groelli for their fieldwork.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2021.776905/full#supplementary-material
Aryal, S., Hausser, M., Griessinger, J., Fan, Z. X., and Brauning, A. (2020). “dendRoAnalyst”: a tool for processing and analysing dendrometer data. Dendrochronologia 64:125772. doi: 10.1016/j.dendro.2020.125772
Bastos, A., Ciais, P., Friedlingstein, P., Sitch, S., Pongratz, J., Fan, L., et al. (2020). Direct and seasonal legacy effects of the 2018 heat wave and drought on European ecosystem productivity. Sci. Adv. 6:eaba2724. doi: 10.1126/sciadv.aba2724
Bose, A. K., Moser, B., Rigling, A., Lehmann, M. M., Milcu, A., Peter, M., et al. (2020). Memory of environmental conditions across generations affects the acclimation potential of scots pine. Plant Cell Environ. 43, 1288–1299. doi: 10.1111/pce.13729
Buras, A., Rammig, A., and Zang, C. S. (2020). Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences 17, 1655–1672.
Charrier, G., Nolf, M., Leitinger, G., Charra-Vaskou, K., Losso, A., Tappeiner, U., et al. (2017). Monitoring of freezing dynamics in trees: a simple phase shift causes complexity. Plant Physiol. 173, 2196–2207. doi: 10.1104/pp.16.01815
Cuny, H. E., Rathgeber, C. B. K., Frank, D., Fonti, P., Makinen, H., Prislan, P., et al. (2015). Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 1, 1–6.
Deslauriers, A., Rossi, S., and Anfodillo, T. (2007). Dendrometer and intra-annual tree growth: what kind of information can be inferred? Dendrochronologia 25, 113–124. doi: 10.1016/j.dendro.2007.05.003
Deslauriers, A., Rossi, S., Turcotte, A., Morin, H., and Krause, C. (2011). A three-step procedure in SAS to analyze the time series from automatic dendrometers. Dendrochronologia 29, 151–161. doi: 10.1016/j.dendro.2011.01.008
Dietrich, L., and Kahmen, A. (2019). Water relations of drought-stressed temperate trees benefit from short drought-intermitting rainfall events. Agric. For. Meteorol. 265, 70–77. doi: 10.1016/j.agrformet.2018.11.012
Dixon, H. H., and Joly, J. (1894). On the ascent of sap. Philos. Trans. R. Soc. Lond. B Biol. Sci. 186, 563–576.
Drew, D. M., and Downes, G. M. (2009). The use of precision dendrometers in research on daily stem size and wood property variation: a review. Dendrochronologia 27, 159–172.
Dukat, P., Ziemblinska, K., Olejnik, J., Malek, S., Vesala, T., and Urbaniak, M. (2021). Estimation of biomass increase and CUE at a young temperate scots pine stand concerning drought occurrence by combining eddy covariance and biometric methods. Forests 12:867. doi: 10.3390/f12070867
Eitel, J. U. H., Griffin, K. L., Boelman, N. T., Maguire, A. J., Meddens, A. J. H., Jensen, J., et al. (2020). Remote sensing tracks daily radial wood growth of evergreen needleleaf trees. Glob. Change Biol. 26, 4068–4078. doi: 10.1111/gcb.15112
Etzold, S., Ferretti, M., Reinds, G. J., Solberg, S., Gessler, A., Waldner, P., et al. (2020). Nitrogen deposition is the most important environmental driver of growth of pure, even-aged and managed European forests. For. Ecol. Manag. 458:117762. doi: 10.1016/j.foreco.2019.117762
Etzold, S., Zieminska, K., Rohner, B., Bottero, A., Bose, A. K., Ruehr, N. K., et al. (2019). One century of forest monitoring data in Switzerland reveals species- and site-specific trends of climate-induced tree mortality. Front. Plant Sci. 10:307.
Eugster, W., Baumgartner, L. P., Bachmann, O., Baltensperger, U., Dèzes, P., Dubois, N., et al. (2021). “Geosciences Roadmap for Research Infrastructures 2025–2028 by the Swiss Geosciences Community”. Swiss Academies Reports. Bern: RoTaGeo team.
Fang, O. Y., Zhang, Q. B., Vitasse, Y., Zweifel, R., and Cherubini, P. (2021). The frequency and severity of past droughts shape the drought sensitivity of juniper trees on the Tibetan plateau. For. Ecol. Manag. 486:118968. doi: 10.1016/j.foreco.2021.118968
Franz, D., Acosta, M., Altimir, N., Arriga, N., Arrouays, D., Aubinet, M., et al. (2018). Towards long-term standardised carbon and greenhouse gas observations for monitoring Europe’s terrestrial ecosystems: a review. Int. Agrophys. 32, 439–455.
García Criado, M., Myers-Smith, I., Baeten, L., Cunliffe, A., Daskalova, G., Gallois, E., et al. (2020). Sharing is caring: working with other people’s data. Methods Ecol. Evol. Methods Blog Available online at: https://methodsblog.com/2020/09/04/sharing-is-caring-working-with-other-peoples-data/ (accessed September 24, 2020),Google Scholar
Geng, X., Fu, Y. H., Hao, F., Zhou, X., Zhang, X., Yin, G., et al. (2020). Climate warming increases spring phenological differences among temperate trees. Glob. Change Biol. 26, 5979–5987. doi: 10.1111/gcb.15301
Goli, G., Becherini, F., Di Tuccio, M. C., Bernardi, A., and Fioravanti, M. (2019). Thermal expansion of wood at different equilibrium moisture contents. J. Wood Sci. 65, 1–7.
Gricar, J., and Cufar, K. (2008). Seasonal dynamics of phloem and xylem formation in silver fir and Norway spruce as affected by drought. Russ. J. Plant Physiol. 55, 538–543. doi: 10.1134/s102144370804016x
Gricar, J., Jagodic, S., and Prislan, P. (2015). Structure and subsequent seasonal changes in the bark of sessile oak (Quercus petraea). Trees Struct. Funct. 29, 747–757. doi: 10.1007/s00468-015-1153-z
Gricar, J., Levanic, T., and Oven, P. (2008). Parameters of Gompertz function for evaluation of wood formation dynamics expressed as number of cells or measured widths in Norway spruce. Wood Res. 53, 35–44.
Güney, A., Zweifel, R., Turkan, S., Zimmermann, R., Wachendorf, M., and Guney, C. O. (2020). Drought responses and their effects on radial stem growth of two co-occurring conifer species in the Mediterranean mountain range. Ann. For. Sci. 77, 1–16.
Haeni, M., Knüsel, S., Wilhelm, M., Peters, R. L., and Zweifel, R. (2020). “Treenetproc - Clean, Process and Visualise Dendrometer Data. R Package Version 0.1.4.”. Github Repository. Available online at: https://github.com/treenet/treenetproc (accessed June 9, 2021).
Hari, V., Rakovec, O., Markonis, Y., Hanel, M., and Kumar, R. (2020). Increased future occurrences of the exceptional 2018-2019 Central European drought under global warming. Sci. Rep. 10:12207.
Heiskanen, J., Brümmer, C., Buchmann, N., Calfapietra, C., Chen, H., Gielen, B., et al. (2021). Integrated carbon observation system in Europe. Bull. Am. Meteorol. Soc. (in press)
Heylen, R. (1992). “A meteorological information processing system (MIPS),” in International Weather Radar Networking, ed. C. G. Collier (Dordrecht: Springer).
Hilty, J., Muller, B., Florent, P., and Leuzinger, S. (2021). Plant growth: the what, the how, and the why. New Phytol. 232, 25–41.
IPCC (2021). Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Kannenberg, S. A., Novick, K. A., Alexander, M. R., Maxwell, J. T., Moore, D. J. P., Phillips, R. P., et al. (2019). Linking drought legacy effects across scales: from leaves to tree rings to ecosystems. Glob. Change Biol. 25, 2978–2992. doi: 10.1111/gcb.14710
Kitaura, K. (1967). “Supercooling and ice formation in mulberry,” in Cryobiology, ed. E. Asahina (Sapporo: Hokkaido Univ), 143–156.
Knüsel, S., Haeni, M., Wilhelm, M., Peters, R. L., and Zweifel, R. (2021). Processing and extraction of seasonal tree physiological parameters from stem radius time series. Forests 12:765. doi: 10.3390/f12060765
Lamacque, L., Charrier, G., Farnese, F. D., Lemaire, B., Ameglio, T., and Herbette, S. (2020). Drought-induced mortality: branch diameter variation reveals a point of no recovery in Lavender species. Plant Physiol. 183, 1638–1649. doi: 10.1104/pp.20.00165
Lockhart, J. A. (1965). An analysis of irreversible plant cell elongation. J. Theor. Biol. 8, 264–275. doi: 10.1016/0022-5193(65)90077-9
Majdak, A., Jakus, R., and Blazenec, M. (2021). Determination of differences in temperature regimes on healthy and bark-beetle colonised spruce trees using a handheld thermal camera. Iforest Biogeosci. For. 14, 203–211. doi: 10.3832/ifor3531-014
Mayr, S., Schmid, P., and Beikircher, B. (2012). “Plant water relations in alpine winter,” in Plants in Alpine Regions, ed. C. Lütz (Vienna: Springer). doi: 10.1007/978-3-030-59538-8_9
Mencuccini, M., Salmon, Y., Mitchell, P., Holtta, T., Choat, B., Meir, P., et al. (2017). An empirical method that separates irreversible stem radial growth from bark water content changes in trees: theory and case studies. Plant Cell Environ. 40, 290–303. doi: 10.1111/pce.12863
Meng, S. W., Fu, X. L., Zhao, B., Dai, X. Q., Li, Q. K., Yang, F. T., et al. (2021). Intra-annual radial growth and its climate response for Masson pine and Chinese fir in subtropical China. Trees Struct. Funct. 1–14. doi: 10.1007/s00468-021-02152-5
Monteith, J. L., and Unsworth, M. H. (1990). Principles of Environmental Physics. Oxford: Edward Arnold.
National Academies of Sciences Engineering and Medicine (2016). Attribution of Extreme Weather Events in the Context of Climate Change. Washington, DC: The National Academies Press.
Nehemy, M. F., Benettin, P., Asadollahi, M., Pratt, D., Rinaldo, A., and Mcdonnell, J. J. (2021). Tree water deficit and dynamic source water partitioning. Hydrol. Process. 35, e14004. doi: 10.1002/hyp.14004
Oberhuber, W., Sehrt, M., and Kitz, F. (2020). Hygroscopic properties of thin dead outer bark layers strongly influence stem diameter variations on short and long time scales in Scots pine (Pinus sylvestris L.). Agric. For. Meteorol. 290, 1–7. doi: 10.1016/j.agrformet.2020.108026
Ogle, K., Barber, J. J., Barron-Gafford, G. A., Bentley, L. P., Young, J. M., Huxman, T. E., et al. (2015). Quantifying ecological memory in plant and ecosystem processes. Ecol. Lett. 18, 221–235. doi: 10.1111/ele.12399
Pappas, C., Peters, R. L., and Fonti, P. (2020). Linking variability of tree water use and growth with species resilience to environmental changes. Ecography 43, 1386–1399. doi: 10.1111/ecog.04968
Penman, H. L. (1948). Natural evaporation from open water, bare soil and grass. Proc. R. Soc. Lond. 193, 120–146. doi: 10.1098/rspa.1948.0037
Peters, R. L., Fonti, P., Frank, D. C., Poyatos, R., Pappas, C., Kahmen, A., et al. (2018). Quantification of uncertainties in conifer sap flow measured with the thermal dissipation method. New Phytol. 219, 1283–1299. doi: 10.1111/nph.15241
Peters, R. L., Steppe, K., Cuny, H. E., De Pauw, D. J. W., Frank, D. C., Schaub, M., et al. (2021b). Turgor - a limiting factor for radial growth in mature conifers along an elevational gradient. New Phytol. 229, 213–229. doi: 10.1111/nph.16872
Peters, R. L., Pappas, C., Hurley, A. G., Poyatos, R., Flog, V., Zweifel, R., et al. (2021a). Assimilate, process, and analyse thermal dissipation sap flow data using the TREX R package. Methods Ecol. Evol. 12, 342–350. doi: 10.1111/2041-210X.13524
Pierrat, Z., Nehemy, M. F., Roy, A., Magney, T., Parazoo, N. C., Laroque, C., et al. (2021). Tower-based remote sensing reveals mechanisms behind a two-phased spring transition in a mixed-species boreal forest. J. Geophys. Res. Biogeosci. 126:e2020JG006191. doi: 10.1029/2020JG006191
Poyatos, R., Et, A., Zweifel, R., and MartiíNez-Vilalta, J. (2021). Global transpiration data from sap flow measurements: the SAPFLUXNET database. Earth Syst. Sci. Data 13, 2607–2649. doi: 10.5194/essd-2020-227
R Core Team (2019). R: A Language and Environment for Statistical Computing (Version 3.6.2). Vienna: R Foundation for Statistical Computing.
Rathgeber, C. B. K., Cuny, H. E., and Fonti, P. (2016). Biological basis of tree-ring formation: a crash course. Front. Plant Sci. 7:734. doi: 10.3389/fpls.2016.00734
Richardson, A. D., Hufkens, K., Milliman, T., Aubrecht, D. M., Chen, M., Gray, J. M., et al. (2018). Tracking vegetation phenology across diverse North American biomes using PhenoCam imagery. Sci. Data 5:180028. doi: 10.1038/sdata.2018.28
Rigling, A., Bigler, C., Eilmann, B., Feldmeyer-Christe, E., Gimmi, U., Ginzler, C., et al. (2013). Driving factors of a vegetation shift from Scots pine to pubescent oak in dry alpine forests. Glob. Change Biol. 19, 229–240. doi: 10.1111/gcb.12038
Sabbatini, S., Mammarella, I., Arriga, N., Fratini, G., Graf, A., Hortriagl, L., et al. (2018). Eddy covariance raw data processing for CO2 and energy fluxes calculation at ICOS ecosystem stations. Int. Agrophys. 32, 495–515. doi: 10.1515/intag-2017-0043
Schafer, C., Rotzer, T., Thurm, E. A., Biber, P., Kallenbach, C., and Pretzsch, H. (2019). Growth and tree water deficit of mixed Norway spruce and European beech at different heights in a tree and under heavy drought. Forests 10, 1–20. doi: 10.3390/f10070577
Schimel, D., and Keller, M. (2015). Big questions, big science: meeting the challenges of global ecology. Oecologia 177, 925–934. doi: 10.1007/s00442-015-3236-3
Schönbeck, L., Gessler, A., Hoch, G., Mcdowell, N. G., Rigling, A., Schaub, M., et al. (2018). Homeostatic levels of nonstructural carbohydrates after 13 yr of drought and irrigation in Pinus sylvestris. New Phytol. 219, 1314–1324. doi: 10.1111/nph.15224
Schulze, E.-D., Beck, E., Buchmann, N., Clemens, S., Müller-Hohenstein, K., and Scherer-Lorenzen, M. (2019). Plant Ecology. Berlin: Springer-Verlag. doi: 10.1007/978-3-662-56233-8
Sellier, D., and Segura, R. (2020). Radial growth anisotropy and temporality in fast-growing temperate conifers. Ann. For. Sci. 77, 1–12. doi: 10.1007/s13595-020-00991-9
Sevanto, S., Holbrook, N. M., and Ball, M. C. (2012). Freeze/Thaw-induced embolism: probability of critical bubble formation depends on speed of ice formation. Front. Plant Sci. 3:107. doi: 10.3389/fpls.2012.00107
Single, W. V. (1964). Studies on frost injury to wheat. II. Ice formation within the plant. Austr. J. Agric. Res. 15, 869–875. doi: 10.1071/AR9640869
Steppe, K., Sterck, F., and Deslauriers, A. (2015). Diel growth dynamics in tree stems: linking anatomy and ecophysiology. Trends Plant Sci. 20, 335–343.
Trotsiuk, V., Hartig, F., Cailleret, M., Babst, F., Forrester, D. I., Baltensweiler, A., et al. (2020). Assessing the response of forest productivity to climate extremes in Switzerland using model-data fusion. Glob. Change Biol. 26, 2463–2476. doi: 10.1111/gcb.15011
Walthert, L., Ganthaler, A., Mayr, S., Saurer, M., Waldner, P., Walser, M., et al. (2021). From the comfort zone to crown dieback: sequence of physiological stress thresholds in mature European beech trees across progressive drought. Sci. Total Environ. 753:14. doi: 10.1016/j.scitotenv.2020.141792
Woodruff, D. R., and Meinzer, F. C. (2011). “Size-dependent changes in biophysical control of tree growth: the role of turgor,” in Size- and Age-Related Changes in Tree Structure and Function, eds F. C. Meinzer, B. Lachenbruch, and T. E. Dawson (Dordrecht: Springer), 363–384. doi: 10.1007/978-94-007-1242-3_14
Yenni, G. M., Christensen, E. M., Bledsoe, E. K., Supp, S. R., Diaz, R. M., White, E. P., et al. (2019). Developing a modern data workflow for regularly updated data. PLoS Biol. 17:e3000125. doi: 10.1371/journal.pbio.3000125
Zweifel, R., Etzold, S., Sterck, F., Gessler, A., Anfodillo, T., Mencuccini, M., et al. (2020). Determinants of legacy effects in pine trees - implications from an irrigation-stop experiment. New Phytol. 227, 1081–1096. doi: 10.1111/nph.16582
Zweifel, R., Haeni, M., Buchmann, N., and Eugster, W. (2016). Are trees able to grow in periods of stem shrinkage? New Phytol. 211, 839–849. doi: 10.1111/nph.13995
Zweifel, R., and Häsler, R. (2000). Frost-induced reversible shrinkage of bark of mature, subalpine conifers. Agric. For. Meteorol. 102, 213–222. doi: 10.1016/S0168-1923(00)00135-0
Zweifel, R., and Sterck, F. (2018). A conceptual tree model explaining legacy effects on stem growth. Front. Plant Sci. 1:9. doi: 10.3389/ffgc.2018.00009
Zweifel, R., Sterck, F., Braun, S., Buchmann, N., Eugster, W., Gessler, A., et al. (2021). Why trees grow at night. New Phytol. 231, 2174–2185.
Keywords: early-warning, radial wood and bark growth, forest dynamics, Research Infrastructure, tree water relations, observation system, near real-time monitoring, nowcast
Citation: Zweifel R, Etzold S, Basler D, Bischoff R, Braun S, Buchmann N, Conedera M, Fonti P, Gessler A, Haeni M, Hoch G, Kahmen A, Köchli R, Maeder M, Nievergelt D, Peter M, Peters RL, Schaub M, Trotsiuk V, Walthert L, Wilhelm M and Eugster W (2021) TreeNet–The Biological Drought and Growth Indicator Network. Front. For. Glob. Change 4:776905. doi: 10.3389/ffgc.2021.776905
Received: 14 September 2021; Accepted: 05 October 2021;
Published: 04 November 2021.
Edited by:
Alessandra De Marco, Energy and Sustainable Economic Development (ENEA), ItalyReviewed by:
Makoto Watanabe, Tokyo University of Agriculture and Technology, JapanCopyright © 2021 Zweifel, Etzold, Basler, Bischoff, Braun, Buchmann, Conedera, Fonti, Gessler, Haeni, Hoch, Kahmen, Köchli, Maeder, Nievergelt, Peter, Peters, Schaub, Trotsiuk, Walthert, Wilhelm and Eugster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roman Zweifel, cm9tYW4uendlaWZlbEB3c2wuY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.