 Lesli C. Hernández-Mendoza
Lesli C. Hernández-Mendoza Luis Escalera-Vázquez
Luis Escalera-Vázquez Daniel Arceo-Carranza
Daniel Arceo-Carranza- 1Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Sisal, Mexico
- 2Laboratorio de Biología Acuática, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
- 3Laboratorio de Ecología, Unidad Multidisciplinaria de Docencia e Investigación Sisal, Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico City, Mexico
The loss of mangrove areas due to anthropogenic activities has triggered efforts to recover or restore these ecosystems, their functions, and associated diversity. Such functions include nursery areas and energy flow through trophic relationships for a large variety of inhabitant species (e.g., fish). The food webs and trophic dynamics of estuarine fish provide important information on the food resources in mangrove ecosystems and their response to restoration processes. Nonetheless, few studies focus on using fish feeding characteristics as an approximation to assess the conservation or recovery status of these aquatic ecosystems. Thus, the aim of the present study was to compare the trophic dynamics of an ichthyic community in a mangrove ecosystem related to karstic wetlands in the Mexican–Caribbean using freshwater, estuarine, and marine fish species as bioindicators of the restoration process in mangroves. Stomach contents were analyzed for eight species of fish inhabiting specific mangrove zones (1—conserved zone, and 2—restored zone; a zone exposed to ecological restoration processes due to impacts of anthropic activities) related to karstic wetlands in the Mexican–Caribbean. Four feeding characteristics were considered: trophic guild, trophic level, feeding strategy, and prey abundance. Results showed differences and changes in the use of food sources at the trophic level mainly for Floridichthys polyommus, changing from a secondary consumer in the conserved zones to a primary consumer in the restored zones. This suggests that the feeding characteristics of the inhabiting fish are related to the mangrove’s conservation/restoration status and the trophic dynamics in the community. The results of this study are relevant as a tool for mangrove restoration plans regarding the analysis of fish and their food prey, in order to perform an easy and rapid assessment to determine the conservation/restoration status of these aquatic ecosystems from a functional perspective.
Introduction
Worldwide, mangroves are coastal ecosystems that are distributed in tropical and subtropical zones; they provide many ecosystem services such as water filtration, carbon storage, and coastline maintenance as a buffer to the impact of hurricanes and tides (Nagelkerken et al., 2008; Twilley and Day, 2013). Also, a high primary productivity and complex arrangement of submerged and aerial roots (neumatophores) form a refuge and feeding habitat for many species of fish at different life cycle stages (Blaber, 2000; Laegdsgaar and Johnson, 2001). Even when the ecological relevance of mangrove areas is acknowledged, these ecosystems are the most threatened by anthropic activities (Twilley and Day, 2013; Hamilton and Casey, 2016).
The accelerated reduction in mangrove areas has led to the implementation of different conservation and restoration efforts, with the objective of maintaining ecosystem functions at the local, regional, and global scale (FAO, 2007). Mainly, mangrove restoration activities are based on recovering hydrologic processes and substrate retention through reforestation. Therefore, the ecosystem restoration indicators have focused on hydrology and vegetation structure (McAlpine et al., 2016). However, the biotic interactions that include vertebrates (e.g., fish and birds), invertebrates (e.g., crustaceans and insects), primary producers (e.g., algae and plants), and food webs reflect the restoration progress of ecosystem functions in less time (Bosire et al., 2008).
Fishes serve as bioindicators of the mangrove restoration process, as its narrow relationship with the roots in this habitat (used as refuge, feeding, and reproduction areas) and high recovery rate of species after the restoration process (Laegdsgaar and Johnson, 2001; Nagelkerken et al., 2008; Vaslet et al., 2015). It is estimated that the mangrove fish community in restored mangroves is similar to that of conservation zones in nearly 5 years (Bosire et al., 2008). In addition, mangrove ecosystems are an important source of structure food webs due to the connections with adjacent aquatic systems, such as coral reefs (Nagelkerken et al., 2000; Mumby, 2006) and terrestrial ecosystems, which increase the availability of prey for larger vertebrates such as birds and reptiles (Trexler and Goss, 2009; De Dios Arcos et al., 2019).
Different aspects of the ichthyic community have been assessed in relation to restoration processes, primarily those considering the structure and composition of species (e.g., Trexler and Goss, 2009; Arceo-Carranza et al., 2016; Enchelmaier et al., 2020; Soria-Barreto et al., 2021). Nonetheless, these results only reflect the importance of mangrove ecosystems as refuge (Kovalenko et al., 2019; Le Guen et al., 2019). Recently, the importance in including the trophic interactions among fish and its preys has been proposed as an indirect tool useful to assess the conservation and restoration status of mangrove ecosystems, because of its high connectivity to terrestrial and marine environments and process related to the maintenance of biodiversity and functions at different scales (Palmer et al., 1997; Vander Zanden et al., 2016; Hale et al., 2019; Loch et al., 2020).
Classifying species by functional groups or guilds provides a more detailed perspective of the biotic and abiotic responses related to species ecological relationship, organism–environment responses, incorporating characteristics driving interactions and exploitation of available resources in the ecosystem, and how these influence processes and functions (Elliott et al., 2007; Violle et al., 2007). Also, it can reveal other mechanisms that promote diversity such as the response of communities to environmental changes (i.e., predator–prey relationships), prey preferences, and prey abundance (Kovalenko et al., 2019).
Habitat modification promotes changes in the trophic structure of aquatic ecosystem and modifies the prey diversity availability for the ichthyofauna (Bernardino et al., 2018). Therefore, the feeding habits are modified in order to exploit the available resources in the environment (Elliott et al., 2007; Vander Zanden et al., 2016). In relation to the seasonal dynamics, the ecotone that represents the mangrove ecosystems harbors high functional diversity such as opportunistic species related to a large trophic plasticity, others related to changes in environmental conditions, and in most cases high richness of species with specialized diets, which in most cases migrate to higher, richer, prey-conserved zones. Regarding the aforementioned, the study and comparison of changes in fish diets between recovering zones through restoration processes are relevant in order to assess the status and success of restoration plans and techniques. Therefore, the aim of the present study was to compare the trophic dynamics of eight fish species that inhabit a mangrove ecosystem related to karstic wetlands in the Mexican–Caribbean using freshwater, estuarine, and marine fish species as bioindicators of the restoration process in aquatic environments.
Materials and Methods
Study Area
The Sian Ka’an Biosphere Reserve is located in the State of Quintana Roo, Mexico, at the western region of the Caribbean Sea. Even when this area is decreed as Natural Reserve, it has been significantly modified by anthropic activities (e.g., mangrove deforestation, cattle establishment, water flow modification, etc.). The zone known as “El Playón” in the eastern zone of the reserve (19°49′12.16″ N, 87°29′29.22″ W) remains with a vegetation composed mainly of red mangrove (Rhizophora mangle); the zone presents fragmentation due to a road construction, which altered the vegetation cover, hydrologic flow, and connectivity, particularly for the southern portion of this zone.
Since 2009, the southern zone has been under restoration process (now referred as “restored” zone) in its hydrology and forestry through the construction of culverts, desilting channels, and reforestation of R. mangle. In contrast, the northern zone presents a high density of R. mangle trees, and its hydrology does not present any alteration related to its structure (Herrera-Silveira et al., 2014). This zone was considered as a reference zone (now referred as “conserved” zone) (Figure 1).
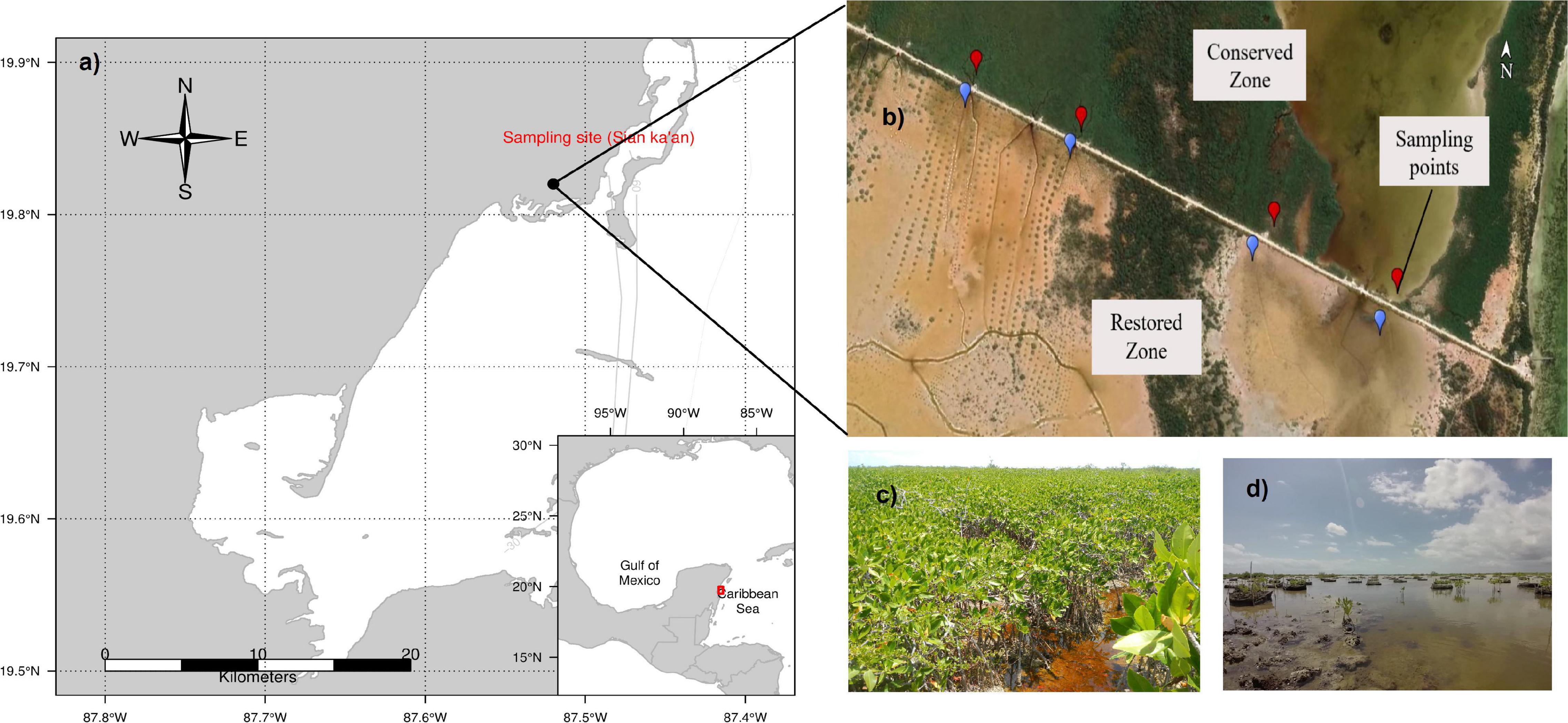
Figure 1. (a) Location of the El Playon mangrove ecosystem, Sian Ka’an, Quintana Roo, Mexico. (b) Sample sites in conserved area of R. mangle and restored area with channels and reforestation of R. mangle. (c) Photography of conserved site of R. mangle and (d) restored zone of mangrove.
This area presents three climatic seasons influenced by regional precipitation: rainy, dry, and cold fronts, regionally known as “nortes”; a saline gradient also is reported in relation to the influence of the freshwater entry due to groundwater upwellings (Herrera-Silveira et al., 2015); the salinity of superficial water presents a variation of 7 to 37 ppm in the southern zone and 3 to 33 ppm in the northern zone; both areas presented a gradient having lower salinity values in the intern zone; the water level varies from 0.30 m in the dry season to 0.75 m in the rainy season in both areas. The temperature of water oscillate was from 24 to 34°C in conserved zone and from 24 to 36°C in restored zone. The higher temperatures are recording in dry season, whereas the lower temperature is related to “nortes” season. Turbidity is higher in the restored area, with many dissolved solids; on the contrary, the conserved area presents crystal clear waters with total transparency (Hernández-Mendoza, 2020).
Fish Sampling
Considering an annual cycle, six bimonthly collections were done from August 2017 to May 2018 at four sampling points near the culverts (2–5 m from the shore) within each zone (conserved and restored; Figure 1). Fish were collected in each sampling point using a cast net (two throws; 0.70-m radius and a mesh size of 1 cm). The collected fish were placed in thermal boxes at 4°C and further fixed using formaldehyde at 4%.
Laboratory and Statistical Analysis
To ecologically characterize the species of fish that feed in the mangrove, fish were identified to species level and classified according to their salinity tolerance (marine, freshwater, and estuarine fish) and ontogenic phases such as juveniles and adults, depending on its length at first sexual maturity (Table 1) regarding the specialized literature (Schmitter-Soto, 1998; Castro-Aguirre et al., 1999; FAO, 2002; Miller, 2009; Froese and Pauly, 2019). Based on the presence of fish in the conserved and restored mangrove zones, a diet analysis was done through stomach content analysis (Table 2) for marine (Atherinomorus stipes, Eucinostomus gula, Gerres cinereus, Lutjanus griseus, and Sphoeroides testudineus), estuarine (Gambusia yucatana and Floridichthys polyommus), and freshwater species (Mayaheros urophthalmus).
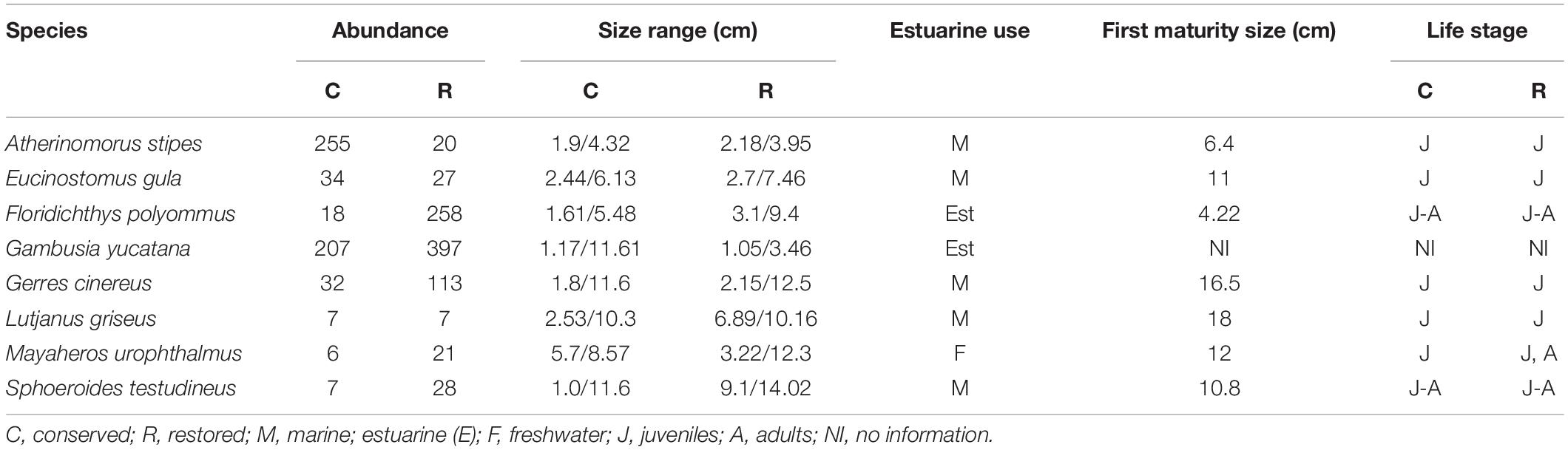
Table 1. Abundance, size range (min–max), and juvenile use of mangrove site (first maturity size) of fish species in mangrove areas.
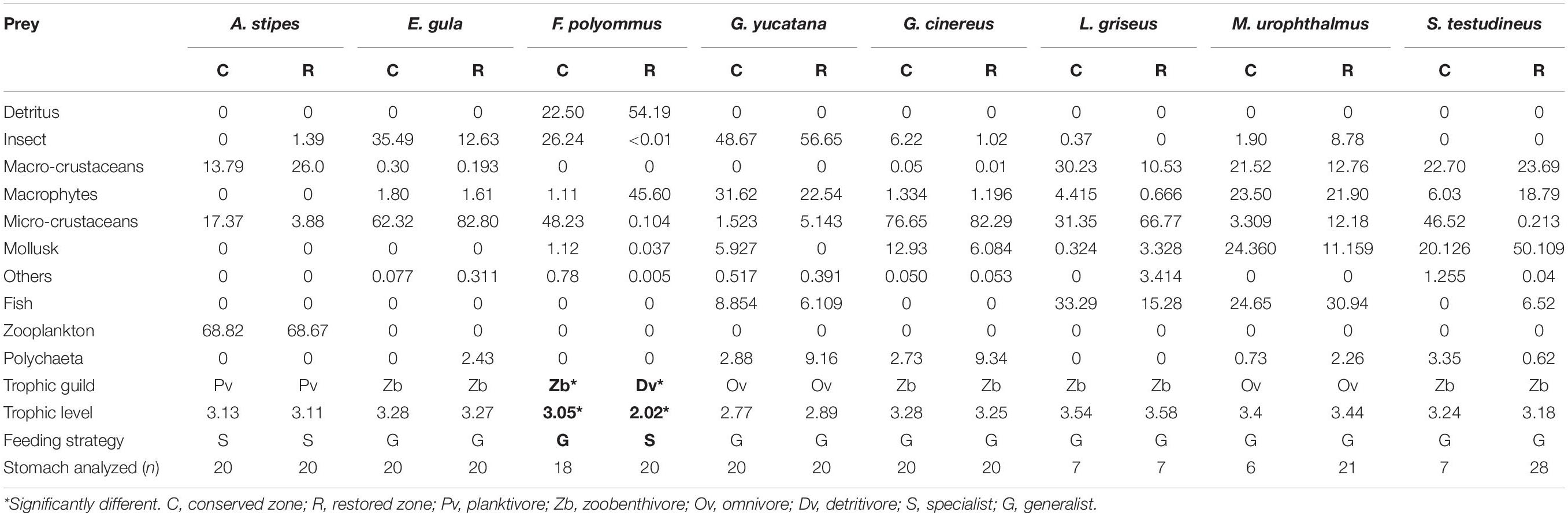
Table 2. Relative importance of the preys, trophic guild, trophic level, feeding strategy, and number of stomach analyzed of the fish species in restored and conserved mangroves.
A total of 274 stomachs of the fishes collected in the mangrove zone were dissected using a stereo-microscope zoom 20 × (Nikon C-LEDS SMZ445). The entire digestive tract from the esophagus to the anus for each individual was examined. The prey species were separated and identified up to the lowest possible taxa. All identified prey items were counted and weighed (in grams). Prey items were grouped in 11 prey categories: macrocrustaceans, macrophytes (remains of terrestrial vegetation) macroalgae, insects, detritus, polychaetes, fish, mollusks, zooplankton (zoeae, decapods, and copepods), and others (foraminifera). In order to obtain the main preys included in the diet, the percentage of the prey-specific index of relative importance (PSIRI%) was used (Brown et al., 2012).
where % FO is frequency of occurrence i; % PN, numeric proportion i; % PW, weight proportion i.
To identify differences between fish species diet, an analysis of similarities (ANOSIM) was performed using “site” as a factor. In addition, an analysis of similarity percentage (SIMPER) was used to identify the preys that mostly contributed to differences in the diet. Analyses were done using the statistical software PRIMER 7.0 for Windows (PRIMER Ltd., Plymouth, United Kingdom).
Three ecological aspects related to fish feeding were evaluated: (1) trophic guild, (2) feeding strategy (specialist or generalist), and (3) trophic level. Following Elliott et al. (2007), the guilds were classified in detritivores, omnivores, zooplanktivorous, and zoobenthivores and analyzed through a cluster analysis using the average linkage method by similarity matrices based on Bray–Curtis distances. Based on the %PSIRI values obtained from the analyses of feeding groups. Similarity profile analysis was performed to identify true groups with an α level of 0.05. The feeding strategy was determined through the graphic method by Costello (1990) modified by Amundsen et al. (1996). The trophic level was calculated starting from the percentage weight of the prey for each species using the software TrophLab (Pauly et al., 2000).
Results
Diet Composition
The diet regarding the marine species in its juvenile stage, A. stipes fed primarily on decapod larvae and copepods, with no significant differences in its diet between zones (ANOSIM R = 0.119, p = 0.08). The feeding of the mojarras E. gula was based on microcrustaceans and insects in both zones (ANOSIM R = 0.13, p = 0.06), whereas G. cinereus fed on microcrustaceans and mollusks (ANOSIM R = −0.05, p = 0.82); none of the two species presented differences between the conserved and restored zones. Snappers (L. griseus) preferred and consumed microcrustaceans and fish in both sites with no significant differences (ANOSIM R = −0.132, p = 0.97). S. testudineus showed differences in its diet between the conserved and restored zone (R = 0.335, p = 0.001). However, the main prey consumed in both zones was mollusks. The SIMPER analysis showed a dissimilarity of 66.53%, highlighting that microcrustaceans and mollusks contributed with more of the 50% in the total diet.
The estuarine species present in the restored zone and the conserved zones were as follows: G. yucatana, which fed mostly on insects and microphytas (ANOSIM R = −0.018, p = 0.62); F. polyommus, showed changes in its diet between zones (Figure 2), since it predominantly consumed detritus and macrophytes in the restored zone. By the other hand, in the conserved zone this species fed mainly on microcrustaceans (ANOSIM R = 0.65, p = 0.001) in comparison to the restored zone. The SIMPER analysis showed a dissimilarity of 88.14% in the diet between zones, detritus, and macrophytes contributed with 66.87% of the total diet. Finally, the only freshwater species recorded was M. urophthalmus and showed no statistical differences in the diet in both zones (ANOSIM R = −0.014, p = 0.5), which was dominated by smaller fish, macrophytes, and mollusks in both zones.
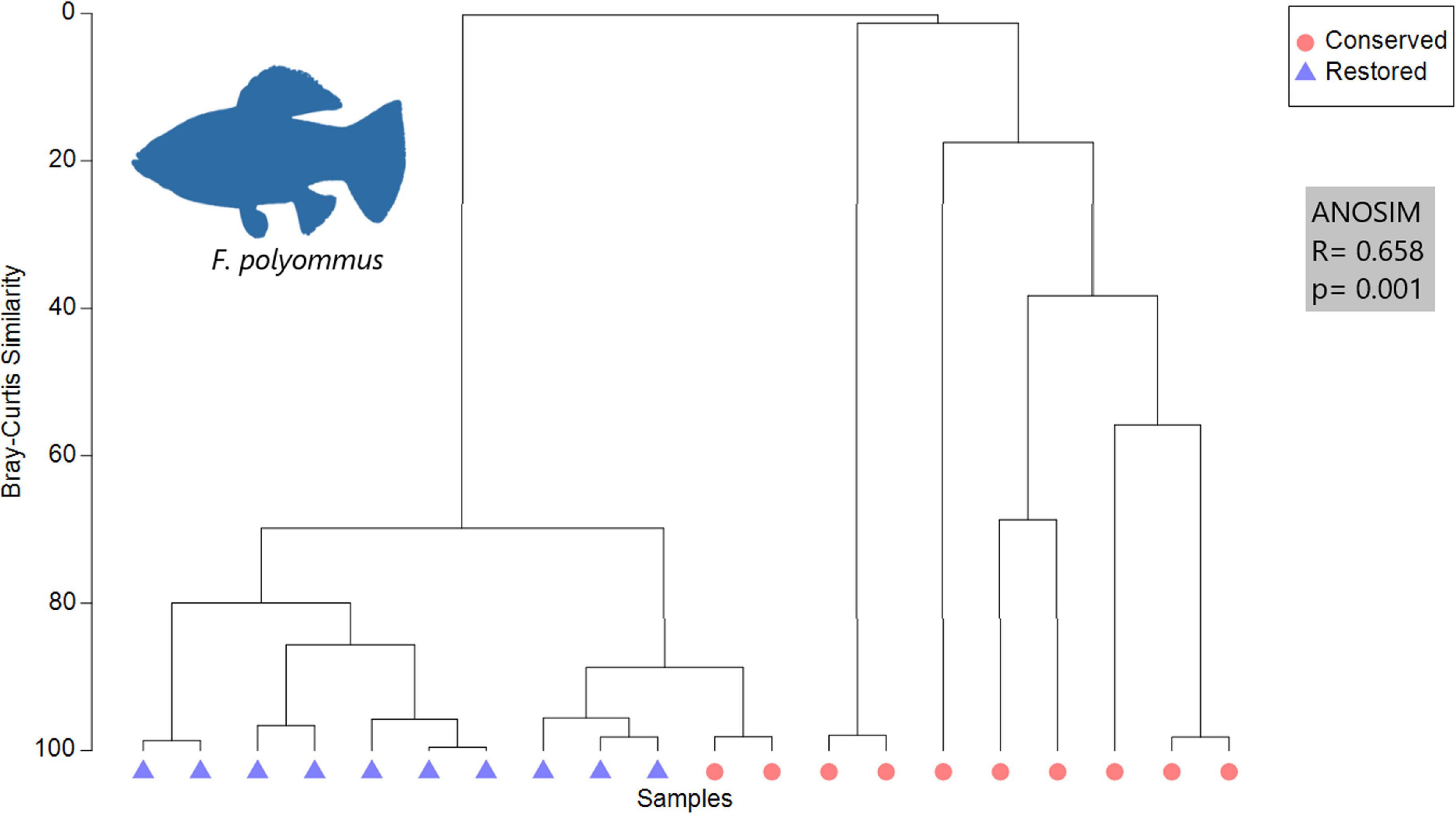
Figure 2. Cluster analysis of F. polyommus showing the similarity of the preys in conserved and restored mangrove zones, and the results of the ANOSIM analysis.
Guilds and Trophic Levels
In accordance with the diet analysis, four trophic guilds were identified: planktivores (PV), detritivores (Dv), omnivores (Ov), and zoobenthivores (Zb) (Figure 3 and Table 2). The trophic level values for the fish species in the restored zone ranged from 2.02 for F. polyommus to 3.58 for L. griseus, whereas for the conserved zone, the minimum value obtained was 2.77 in G. yucatana, and the maximum was 3.54 for L. griseus. The species with the higher trophic level were characterized according to their consumption of fish and macrocrustaceans (Figure 4). No changes were registered either in the guilds or trophic levels in the majority of the species recorded between the conserved and restored mangrove zones. Only F. polyommus showed a difference in guild and trophic level, changing from detritivore and a level of 2.02 in the restored zone to a zoobenthivore and level 3.24 in the conserved zone (Table 2 and Figure 4). According to the feeding strategies, only two species were characterized as specialists, as a result of the low variation in the preys consumed. Atherinomorus stipes consumed from only three different food groups in both zones, and F. polyommus in the restored zone only consumed detritus and macrophytes (Table 2).
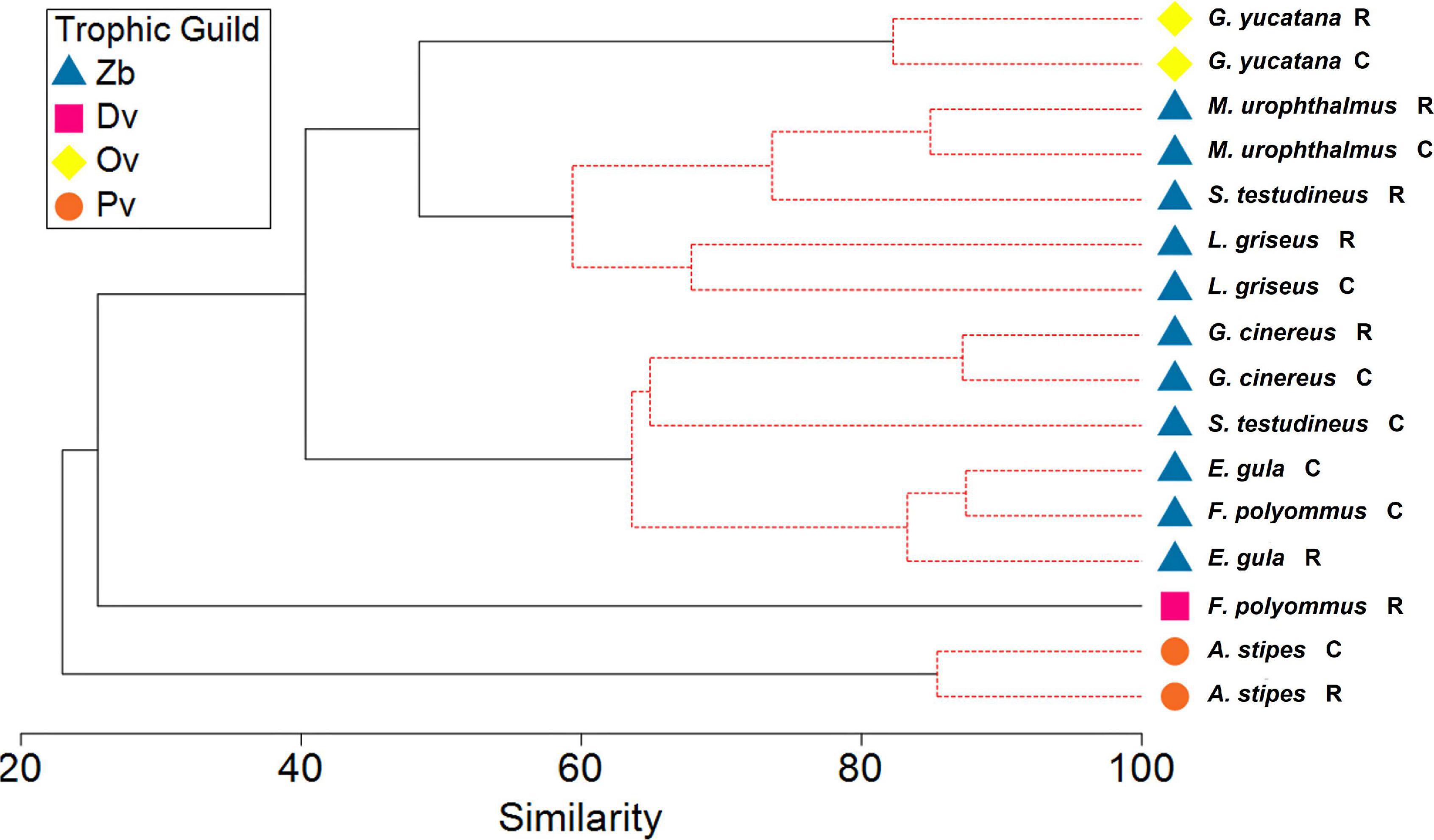
Figure 3. Cluster analysis showing the groups by trophic guilds of the fish species. Pv, planktivores; Dv, detritivores; Ov, omnivores; Zb, zoobenthivores; C, conserved; R, restored.
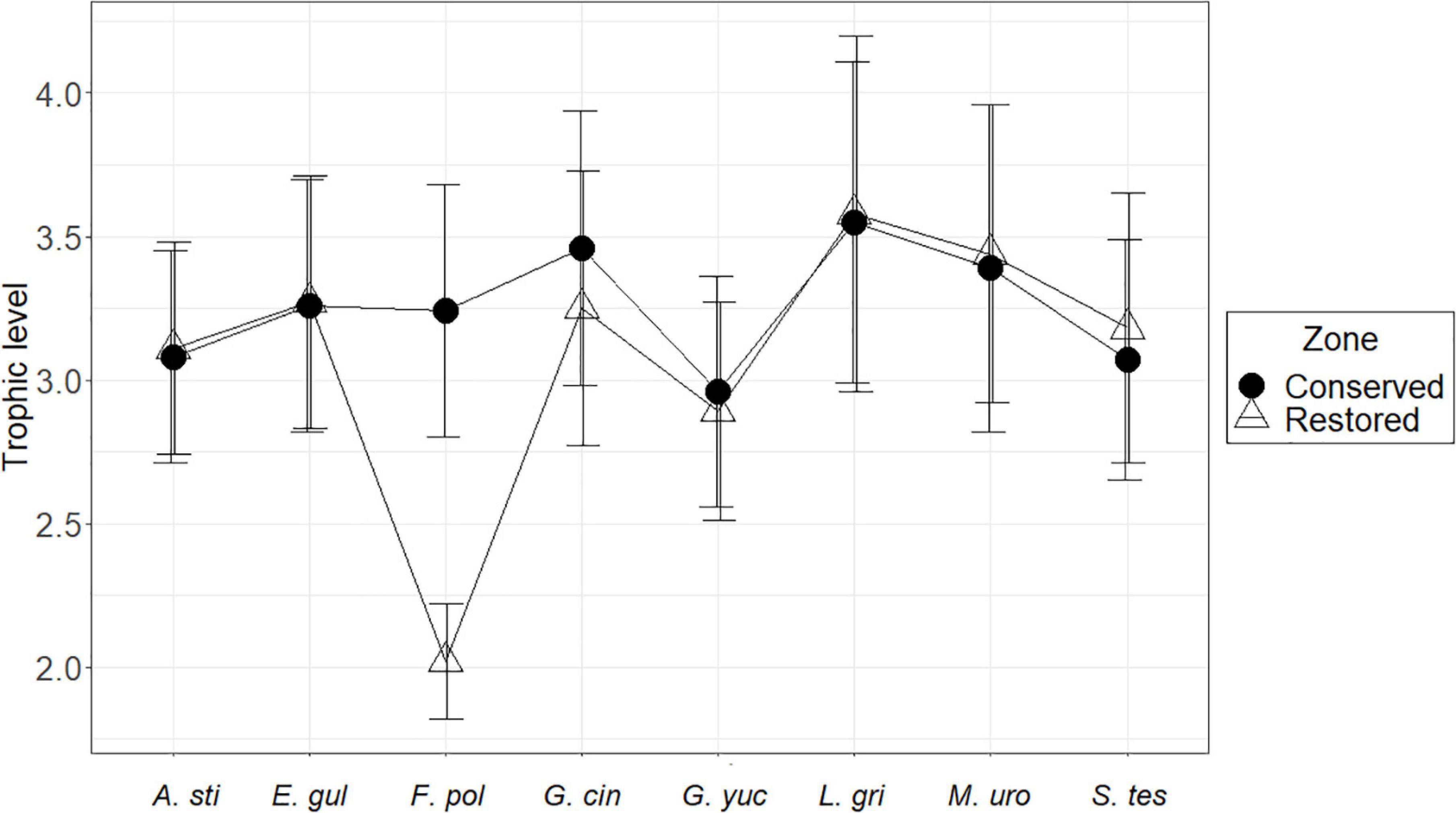
Figure 4. Trophic level (mean and standard deviation) of the fish species analyzed; the conserved area is represented by the circles, and the restored area by the triangles. The name of the fish species is written in code; the first letter corresponds to the genus and the next three to the species.
Discussion
Changes in fish diet are highly related to the richness and abundance of the prey species at a spatial and temporal scale (Hinojosa-Garro et al., 2013). This is also related to environmental seasonal changes and by anthropic activities, where a majority of fish present opportunistic behavior in order to take advantage of peaks in prey abundance (Demopoulos and Smith, 2010). Mangrove deforestation may be due to changes in hydrologic patterns that affect the ecosystem, altering the diversity, biotic interactions, and trophic webs (Shinnaka et al., 2007). Trophic webs connect the resources of the ecosystem with the inhabiting organisms, and through its study, which provide a holistic vision on the state of the ecosystem as compared with information that results only from species richness and abundance (Villéger et al., 2010; Thompson et al., 2012; Ellis and Bell, 2013; Vander Zanden et al., 2016). In the species analyzed in this study, there were interspecific differences identified in the diets, and most of these species fed on zoobenthic organisms, a trophic guild largely reported in marine and estuarine fish within mangrove ecosystems and favored because of high secondary productivity (Elliott et al., 2007; Arceo-Carranza et al., 2013; Dolbeth et al., 2013; Palacios-Sánchez et al., 2019).
Most of the fish inhabiting these estuarine environments are characterized as having generalist-type diets that are flexible in taking advantage of temporal peaks in prey abundance, such that the diet in these estuarine species reflects the variety and type of available resources in its environment (Livingston, 1984; Elliott et al., 2007). This was found in the fish diets from the Sian Ka’an zone, primarily in F. polyommus, a species registering a shift in its diet and taking advantage of the abundance of microcrustaceans such as amphipods and tanaidaceans. This shift in diet suggests an altered state in the environment caused by anthropic activities, in this case, the road interrupting the hydrologic flow and decreasing the vegetation cover related to mangroves, and microhabitats for fish prey existing in the restored zone.
Many of estuarine and marine species using the estuaries in early phases of development are considered generalist. This is found to be related with the ontogenetic development and change in the morphoanatomical characteristics of fish, which allows them to shift from one food source to another, functioning like opportunistic species that exploit the available resources found in the surroundings, making them capable of responding to biotic and abiotic changes, either in a disturbed zone with less diversity in prey, or in a conserved zone that present higher numbers in potential prey (Wooton, 1990; Bellwood et al., 2006). This trophic flexibility is common in estuarine fish, with species temporally exploiting the peaks in prey abundance that exist, however, when there are disturbances that alter the trophic webs and lower the trophic levels; some fish respond by consuming prey at lower trophic levels, such as F. polyommus that is secondary consumer (Poot Salazar et al., 2005) in the conserved zone (feeding zoobenthos), and primary consumer in the restored zone feeding on detritus and phytobenthos. Contrarily, for a marine fish such as L. griseus, a species associated with mangroves in their juvenile phase, feed on microcrustaceans, and upon migrating to the reef zones, the food source is based on macrocrustaceans and fish, additionally increasing its trophic level (Faunce and Serafy, 2008). Similarly, S. testudineus can change its diet according to the available resources, between microcrustaceans and mollusks (Palacios-Sánchez and Vega-Cendejas, 2010; Chi-Espinola and Vega-Cendejas, 2013), but they remain at the same trophic level and zoobentophagous guild.
Nonetheless, there are species that have one food guild, and one type of prey, better adapted to a specialist strategy, such as the juveniles of A. stipes, with a diet based on the consumption of zoeas larvae, of decapods and copepods (Vaslet et al., 2015). Atherinomorus stipes is related with mangrove areas in the Mexican–Caribbean; notwithstanding, its presence is limited by ecosystem alterations, more so, if there is an increase in turbidity of the water, limiting foraging and hindering its encounter with preferred prey (Nash et al., 2017). This high turbidity in water observed for the restored zone probably explains the low abundance of this specie.
The sharing of food resources reduces competition between fish; however, in estuarine environments, many species share from a pool of prey, more so, in the juvenile phases; thus, in theory, the amount of food available for each individual is affected by the consumption of individuals co-occurring in nursery zones. Saulnier et al. (2020) and Day et al. (2020) reported that marcrobenthos produced in these zones directly affect the density of juvenile organisms, which relates to zones that are better conserved and have mangrove habitats, and submerged grasses that provide feeding zones. Fish, therefore, are important elements that serve in quantifying diversity, abundance, and composition of prey found in conserved and impacted environments, and in this case, those that are found in the process of restoration.
An alteration in hydrology, because of anthropogenic actions in estuarine ecosystems, drives a change in the fish assemblages, whether in its taxonomic composition or in functional aspects such as the food guilds and trophic levels (Baptista et al., 2015; Arceo-Carranza et al., 2016; Soria-Barreto et al., 2021). This was reflected in differences in the diets presented by resident species from the mangrove; for example, in F. polyommus, the altered hydrology affected the vegetation and microhabitats for available prey; thus, it presented a diet based on detritus and macrophytes in the restored zone, resources that are characteristic for ecosystems in secondary succession (Harrington and Harrington, 1982).
This study provides evidence on the impact that is generated in the trophic webs (species richness and prey abundance in diets) due to anthropic activities (Bosire et al., 2008). The interruption of water flow caused by road construction in this area of the Mexican–Caribbean affects not only the mangrove cover, but also the faunal diversity (benthonic and nektonic), including the predator–prey interactions of the fish communities present (Bosire et al., 2008; Chen and Ye, 2011). Shifting to alternative prey provokes changes in the vertical structure of the food web decreasing the number of trophic levels, modifying the structure and function of the biological communities in these coastal wetlands. According to some studies on mangrove restoration, there is a gradual shift to more heterogeneous food sources after several years of the restoration process (Loch et al., 2020).
Finally, the results of this work show that trophic ecology of fish are a useful tool, especially in prey abundance, trophic levels, and trophic guilds, key points to know the ecological functions of the community and thus determining and monitoring restoration processes. This garners relevance when there is an attempt to find ecological indicators that show the advances in restoration projects, and in so doing, establish the functional success of the restoration of coastal wetlands. In this case, F. polyommus, being an opportunistic organism, is ideal to know the food resources in the conserved and restored mangroves.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
LH-M and DA-C contributed in the idea and design of the study. LH-M performed the analysis and writing of the manuscript. DA-C and LE-V contributed in the writing and criticism of the intellectual content. All authors contributed to the discussion, review, and approval of the final manuscript.
Funding
The authors would like to thank the Program “Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT-UNAM) project code: IN216219. To CONACYT for the scholarship granted to LH-M (number 491060), as the results presented in this study is part of a Master’s thesis at PCMyL, UNAM.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to thank J. O. Valdez Iuit for field support and collection of samples. The authors would also like to thank Comisión Nacional de Areas Naturales Protegidas (CONANP) and Omar Ortíz Moreno, director of the Sian Ka’an Biosphere Reserve for the field work facilities within “El Playón,” Sian Ka’an.
References
Amundsen, P. A., Gabler, H. M., and Staldvik, F. J. (1996). A new approach to graphical analysis from stomach contents data – modification of the Costello (1990) method. J. Fish Biol. 48, 607–614. doi: 10.1111/j.1095-8649.1996.tb01455.x
Arceo-Carranza, D., Gamboa, E., Teutli-Hernández, C., Badillo-Alemán, M., and Herrera-Silveira, J. A. (2016). Los peces como indicador de restauración de áreas de manglar en la costa Norte de Yucatán. Rev. Mex. Biodiv. 87, 489–496. doi: 10.1016/j.rmb.2016.03.001
Arceo-Carranza, D., Vega-Cendejas, M. E., and Hernández de Santillana, M. (2013). Day and night trophic variations of dominant fish species in a lagoon influenced by freshwater seeps. J. Fish Biol. 82, 54–68. doi: 10.1111/j.1095-8649.2012.03463.x
Baptista, J., Martinho, F., Nyitrai, D., Pardal, M. A., and Dolbeth, M. (2015). Long-term functional changes in an estuarine fish assemblage. Mar. Pollut. Bull. 97, 125–134. doi: 10.1016/j.marpolbul.2015.06.025
Bellwood, D. R., Wainwright, C. J., Fulton, C. J., and Hoey, A. S. (2006). Functional versatility supports coral reef biodiversity. Proc. Biol. Sci. 273, 101–107. doi: 10.1098/rspb.2005.3276
Bernardino, A. F., Oliveira Gomes, L. E., Louise Hadlich, H., Andrades, R., and Barreto Correa, L. (2018). Mangrove clearing impacts on macrofaunal assemblages and benthic food webs in a tropical estuary. Mar. Pollut. Bull. 126, 228–235. doi: 10.1016/j.marpolbul.2017.11.008
Blaber, S. J. M. (2000). Tropical Estuarine Fishes: Ecology, Exploitation and Conservation. Hoboken, NJ: Blackwell Science Ltd, 372.
Bosire, J. O., Dahdouh Guebas, F., Walton, M., Crona, B. I., Lewis, R. R., Field, C., et al. (2008). Functionality of restored mangroves: a review. Aquat. Bot. 89, 251–259. doi: 10.1016/j.aquabot.2008.03.010
Brown, S. C., Bizarro, J. J., Cailliet, G. M., and Ebert, D. A. (2012). Braking with tradition: redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fish. 95, 3–20. doi: 10.1007/s10641-011-9959-z
Castro-Aguirre, J. L., Schmitter-Soto, J. J., and Espinosa Pérez, H. S. (1999). Ictiofauna Estuarino-Lagunar y Vicaría de México. Ciudad de México: Limusa.
Chen, G. C., and Ye, Y. (2011). Restoration of Aegiceras corniculatum mangroves in Jiulongjiang estuary changed macro-benthic faunal community. Ecol. Eng. 37, 224–228. doi: 10.1016/j.ecoleng.2010.10.003
Chi-Espinola, A. A., and Vega-Cendejas, M. E. (2013). Hábitos alimenticios de Sphoeroides testudineus (Perciformes: Tetraodontidae) en el Sistema lagunar de Ría Lagartos, Yucatán, México. Rev. Biol. Trop. 61, 849–858.
Costello, M. J. (1990). Predator feeding strategy and prey importance: a new graphical analysis. J. Fish. Biol. 36, 261–263. doi: 10.1111/j.1095-8649.1990.tb05601.x
Day, L., Le Bris, H., Saulnier, E., Pinsivy, L., and Brind’Amour, A. (2020). Benthic prey production index estimated from trawl survey supports the food limitation hypothesis in coastal fish nurseries. Estuar. Coast. Shelf Sci. 235:106594. doi: 10.1016/j.ecss.2020.106594
De Dios Arcos, C., Badillo Alemán, M., Arceo Carranza, D., and Chiappa Carrara, X. (2019). Feeding ecology of the waterbirds in a tropical mangrove in the southeast Gulf of México. Stud. Neotrop. Fauna Environ. 55, 1–9. doi: 10.1080/01650521.2019.1682232
Demopoulos, A. W. J., and Smith, C. R. (2010). Invasive mangroves alter macrofaunal community structure and facilitate opportunistic exotics. Mar. Ecol. Prog. Ser. 404, 51–67. doi: 10.3354/meps08483
Dolbeth, M., Cardoso, P., Grilo, T., Raffaelli, D., and Pardar, M. A. (2013). Drivers of estuarine benthic Species distribution patterns following a restoration of a seagrass bed: a functional trait analyses. Mar. Pollut. Bull. 72, 47–54. doi: 10.1016/j.marpolbul.2013.05.001
Elliott, M., Whitfield, A. K., Potter, I. C., Blaber, S. J. M., Cyrus, D. P., Nordlie, F. G., et al. (2007). The guild approach to categorizing estuarine fish assemblages: a global review. Fish Fish. 8, 241–268. doi: 10.1111/j.1467-2679.2007.00253.x
Ellis, W. L., and Bell, S. S. (2013). Intertidal fish communities may make poor indicators of environmental quality: lessons from a study of mangrove habitat modification. Ecol. Ind. 24, 421–430. doi: 10.1016/j.ecolind.2012.07.008
Enchelmaier, A. C., Babcock, E. A., and Hammerschlag, N. (2020). Survey of fishes within a restored mangrove habitat of a subtropical bay. Estuar. Coast. Shelf Sci. 244:106021. doi: 10.1016/j.ecss.2018.11.009
FAO (2002). Species Identification Guide for Fishery. Special Publication No. 5. Rome: American society of ichtyologist and Herpetologists.
Faunce, C. H., and Serafy, J. E. (2008). Growth and secondary production of an eventual reef fish during mangrove residency. Estuar. Coast. Shelf Sci. 79, 93–100. doi: 10.1016/j.ecss.2008.03.006
Froese, R., and Pauly, D. (eds) (2019). FishBase. World Wide Web electronic Publication Version. Available online at: www.fishbase.org, (accessed April, 2021)
Hale, R., Mac Nally, R., Blumstein, D. T., and Swearer, S. E. (2019). Evaluating where and how habitat restoration is undertaken for animals. Rest Ecol. 27, 775–781. doi: 10.1111/rec.12958
Hamilton, S. E., and Casey, D. (2016). Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 25, 729–738. doi: 10.1111/geb.12449
Harrington, R. W. Jr., and Harrington, E. S. (1982). Effects on fishes and their forage organisms of impounding a Florida salt marsh to prevent breeding by salt marsh mosquitoes. Bull. Mar. Sci. 32, 523–531.
Hernández-Mendoza, L. C. (2020). Ictiofauna Como Indicador de Recuperación de las Funciones Ecológicas en Una Zona de Manglar en la reserva de la biosfera Sian Ka’an, Quintana Roo. Ph.D. thesis. Yucatán: Universidad Nacional Autónoma de México.
Herrera-Silveira, J. A., Cortes Balam, O., Valdez Iuit, J., Ramírez-Ramírez, J., Serna, M. A., Mariño, T. I., et al. (2015). Implementación de Medidas de Adaptación Para Reducir la Vulnerabilidad Ante Los Impactos del Cambio Climático de la Comunidad de Punta Allen (Reserva de la Biosfera de Sian Ka’an) A Través de la Rehabilitación de un Ecosistema de manglar en el humedal “El Playón”. CINVESTAV, Amigos de Sian Ka’an, ANP de Sian Ka’an, INECC, IMTA -CNA. Punta Allen: GEF.
Herrera-Silveira, J. A., Teutli Hernández, C., Zaldívar Jiménez, A., Pérez Ceballos, R., Cortés Balam, O., Osorio Moreno, I., et al. (2014). Programa Regional Para la Caracterización y el Monitoreo de Ecosistemas de Manglar del Golfo de México y el Caribe Mexicano: Península de Yucatán. CINVESTAV-ECOPEY/CONABIO, FB1307-FN009/08. Informe Final. México D.F: CONABIO.
Hinojosa-Garro, D., Arceo Gómez, J., Zambrano, L., and Escalera Vázquez, L. H. (2013). Fish diet composition in permanent and semi-permanent pools in tropical wetlands of the Yucatán Peninsula. Neotrop. Ichthyol. 11:4. doi: 10.1590/S1679-62252013000400016
Kovalenko, K. E., Johnson, L. B., Brady, V. J., Ciborowski, J. J. H., Cooper, M. J., Gathman, J. P., et al. (2019). Hotspots and bright spots in functional and taxonomic fish diversity. Freshw. Sci. 38, 480–490. doi: 10.1086/704713
Laegdsgaar, P., and Johnson, C. (2001). Why juvenile fish utilize mangrove habitats? J. Exp. Mar. Biol. Ecol. 257, 229–253. doi: 10.1016/S00220981(00)00331-2
Le Guen, C., Tecchio, S., Dauvin, J. C., De Roton, G., Lobry, J., Lepage, M., et al. (2019). Assessing the ecological status of an estuarine ecosystem: linking biodiversity and food-web indicators. Estuar. Coast. Shelf Sci. 228:106339. doi: 10.1016/j.ecss.2019.106339
Livingston, R. J. (1984). Trophic response of fishes to habitat variability in coastal seagrass systems. Ecology 65, 1258–1275. doi: 10.2307/1938332
Loch, J. M. H., Walters, L. J., and Cook, G. S. (2020). Recovering trophic structure through habitat restoration: a review. Food Webs 25:e00162. doi: 10.1016/j.fooweb.2020.e00162
McAlpine, C., Caterrall, C. P., Nally, R. M., Lindenmayer, D., Reid, J. L., Holl, K. D., et al. (2016). Integrating plant-and animal-based perspectives for more effective restoration of biodiversity. Front. Ecol. Environ. 14:37–45. doi: 10.1002/16-0108.1
Miller, R. R. (2009). Peces Dulceacuícolas de México. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO).
Mumby, P. J. (2006). Connectivity of reef fish between mangroves and coral reefs: algorithms for the design of marine reserves at seascape scales. Biol. Conserv. 128, 215–222. doi: 10.1016/j.biocon.2005.09.042
Nagelkerken, I., Blaber, S. J. M., Bouillon, S., Green, P., Haywood, M., Kirton, L. G., et al. (2008). The habitat function of mangroves for terrestrial and marine fauna: a review. Aquat. Bot. 89, 155–185. doi: 10.1016/j.ejmech.2020.112957
Nagelkerken, I., Dorenbosch, M., Verberk, C. E. P., Cocheret de la Morinière, E., and Van der Velde, G. (2000). Importance of shallow-water biotopes of a Caribbean bay for juvenile coral reef fishes: patterns in biotope association, community structure and spatial distribution. Mar. Ecol. Prog. Ser. 202, 175–192. doi: 10.3354/meps202175
Nash, M. C., Kraczkowski, M. L., and Chernoff, B. (2017). Insight into the population structure of hardhead silverside, Atherinomorus stipes (Teleostei: Atherinidae), in Belize and the Florida Keys using nd2. Ecol. Evol. 7, 9503–9517. doi: 10.1002/ece3.3457
Palacios-Sánchez, S. E., and Vega-Cendejas, M. E. (2010). Cambios alimenticios en tres especies de Sphoeroides (Tetraodontiformes: Tetraodontidae) posterior al huracán Isidoro en Bocana de la Carbonera, Sureste del Golfo de México. Rev. Biol. Trop. 58, 1223–1235. doi: 10.15517/RBT.V58I4.5407
Palacios-Sánchez, S. E., Vega-Cendejas, M. E., Hernández-de-Santillana, J. M., and Aguilar-Medrano, R. (2019). Anthropogenic impacts in the nearshore fish community of the Yucatan coastal corridor. A comparison of protected and unprotected areas. J. Nat. Conserv. 51:125721. doi: 10.1016/j.jnc.2019.125721
Palmer, M. A., Ambrose, R. F., and Poff, N. L. (1997). Ecological theory and community restoration ecology. Rest Ecol. 5, 291–300. doi: 10.1046/j.1526-100X.1997.00543.x
Pauly, D., Froese, R., Sa-a, P., Palomares, M. L., Christensen, V., and Rius, J. (2000). TrophLab. Available online at: www.fishbase.org/download/TrophLab2K.zip (accessed November 25, 2019).
Poot Salazar, A. V., Canto Maza, W. G., and Vega Cendejas, M. E. (2005). Hábitos alimenticios de Floridichthys polyommus Hubbs, 1936 (Pisces: Cyprinodontidae) en dos sistemas lagunares costeros. Hidrobiologica 15, 183–194.
Saulnier, E., Le Bris, H., Tableau, A., Dauvin, J. C., and Brind’Amour, A. (2020). Food limitation of juvenile marine fish in a coastal and estuarine nursery. Estaur. Coast Shelf Sci. 241:106670. doi: 10.1016/j.ecss.2020.106670
Schmitter-Soto, J. J. (1998). Catálogo de los Peces Continentales de Quintana Roo. Quintana Roo: Guías científicas ECOSUR.
Shinnaka, T., Sano, M., Ikejma, K., Tongnunui, P., Horinouchi, M., and Kurokura, H. (2007). Effects of mangrove deforestation on fish assemblage at Pak Phannang Bay, southern Thailand. Fish Sci. 73, 862–870. doi: 10.1111/j.1444-2906.2007.01407.x
Soria-Barreto, M., Gelabert Fernández, R., Reyna Ramos, H. E., and Brito, R. (2021). The fish community in Gulf of Mexico mangroves, a response to hydrological restoration. Latin Am. Aquat. Res. 49, 507–519. doi: 10.3856/vol49-issue3-fulltext-2635
Thompson, R. M., Brose, U., Dunne, J. A., Hall, R. O. Jr., Hladyz, S., Kitching, R. L., et al. (2012). Food webs: reconciling the structure and function of biodiversity. Trends Ecol. Evol. 27, 689–697. doi: 10.1016/j.tree.2012.08.005
Trexler, J. C., and Goss, C. W. (2009). Aquatic fauna as indicators for Everglades restoration: applying dynamic targets in assesments. Ecol. Indic. 9, 108–119. doi: 10.1016/j.ecolind.2008.11.001
Twilley, R. R., and Day, J. W. (2013). “Mangrove wetlands,” in Estuarine Ecology, eds J. W. Day, B. C. Crump, W. M. Kemp, and A. Yáñez-Arancibia (New York, NY: Wiley-Blaclwell), 165–203.
Vander Zanden, M. J., Olden, J. D., and Gratton, C. (2016). “Food–web approaches,” in Foundations of Restoration Ecology, eds D. A. Falk, M. A. Palmer, and J. B. Zedler (Washington DC:: Island Press), 165–189.
Vaslet, A., Bouchon-Navaro, Y., Harmelin-Vivien, M., Lepoint, G., Louis, M., Bouchon, C., et al. (2015). Foraging habits of reef fishes associated with mangroves and seagrass beds in a Caribbean lagoon: a stable isotope approach. Cienc Mar. 41, 217–232. doi: 10.7773/cm.v41i3.2494
Villéger, S., Ramos Miranda, J., Flores Hernández, D., and Mouillot, D. (2010). Contrasting changes in taxonomic vs. Functional diversity of tropical fish communities after habitat degradation. Ecol. Appl. 20, 1512–1522. doi: 10.1890/09-1310.1
Violle, C., Navas, M. L., Vile, D., Kazakou, E., Fortunel, C., Hummel, I., et al. (2007). Let the concept of trait be functional! OIKOS 116, 882–892. doi: 10.1111/j.0030-1299.2007.15559.x
Keywords: trophic ecology, mangrove restoration, estuarine fish, feeding strategy, trophic guilds, zoobenthivores
Citation: Hernández-Mendoza LC, Escalera-Vázquez L and Arceo-Carranza D (2022) Estuarine Fish Feeding Changes as Indicator to Mangrove Restoration Success in Seasonal Karstic Wetlands. Front. For. Glob. Change 4:743232. doi: 10.3389/ffgc.2021.743232
Received: 17 July 2021; Accepted: 03 December 2021;
Published: 24 January 2022.
Edited by:
Julieta Benitez-Malvido, National Autonomous University of Mexico, MexicoReviewed by:
Morelia Camacho-Cervantes, National Autonomous University of Mexico, MexicoAna L. Lara, Instituto de Ecología (INECOL), Mexico
Copyright © 2022 Hernández-Mendoza, Escalera-Vázquez and Arceo-Carranza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Arceo-Carranza, ZGFyY2VvQGNpZW5jaWFzLnVuYW0ubXg=