 Stephanie Rehschuh
Stephanie Rehschuh Mathieu Jonard
Mathieu Jonard Martin Wiesmeier
Martin Wiesmeier Heinz Rennenberg
Heinz Rennenberg Michael Dannenmann
Michael Dannenmann- 1Institute of Meteorology and Climate Research–Atmospheric Environmental Research (IMK-IFU), Karlsruhe Institute of Technology (KIT), Garmisch-Partenkirchen, Germany
- 2Earth and Life Institute, Université catholique de Louvain, Louvain-la-Neuve, Belgium
- 3Chair of Soil Sciences, TUM School of Life Sciences, Technische Universität München, Freising-Weihenstephan, Germany
- 4Chair of Tree Physiology, Institute of Forest Sciences, University of Freiburg, Freiburg, Germany
Drought-sensitive European beech forests are increasingly challenged by climate change. Admixing other, preferably more deep-rooting, tree species has been proposed to increase the resilience of beech forests to drought. This diversification of beech forests might also affect soil organic carbon (SOC) and total nitrogen (TN) stocks that are relevant for a wide range of soil functions and ecosystem services, such as water and nutrient retention, filter functions and erosion control. Since information of these effects is scattered, our aim was to synthesize results from studies that compared SOC/TN stocks of beech monocultures with those of beech stands mixed with other tree species as well as monocultures of other tree species. We conducted a meta-analysis including 38 studies with 203, 220, and 160 observations for forest floor (i.e., the organic surface layer), mineral soil (0.5 m depth) and the total soil profile, respectively. Monoculture conifer stands had higher SOC stocks compared to monoculture beech in general, especially in the forest floor (up to 200% in larch forests). In contrast, other broadleaved tree species (oak, ash, lime, maple, hornbeam) showed lower SOC stocks in the forest floor compared to beech, with little impact on total SOC stocks. Comparing mixed beech-conifer stands (average mixing ratio with regard to number of trees 50:50) with beech monocultures revealed significantly higher total SOC stocks of around 9% and a smaller increase in TN stocks of around 4%. This equaled a SOC accrual of 0.1 Mg ha−1 yr−1. In contrast, mixed beech-broadleaved stands did not show significant differences in total SOC stocks. Conifer admixture effects on beech forest SOC were of additive nature. Admixing other tree species to beech monoculture stands was most effective to increase SOC stocks on low carbon soils with a sandy texture and nitrogen limitation (i.e., a high C/N ratio and low nitrogen deposition). We conclude that, with targeted admixture measures of coniferous species, an increase in SOC stocks in beech forests can be achieved as part of the necessary adaptation of beech forests to climate change.
Introduction
European beech (Fagus sylvatica)–the dominant species of the potential natural forest vegetation and the main broad-leaved forest tree species in Central Europe (Ellenberg, 1988; Bolte et al., 2007) is increasingly affected by climate change due to longer, more intense and more frequent summer droughts. This has already led to physiological constrains (Rennenberg et al., 2006) as well as nutrient limitations including impaired ecosystem nitrogen cycling (Geβler et al., 2004; Jonard et al., 2015; Dannenmann et al., 2016). Several studies showed that the admixture of different tree species (e.g., oak, pine, and silver fir) to European beech can reduce drought susceptibility of beech (Pretzsch et al., 2013; Metz et al., 2016; Magh et al., 2018). These admixtures to beech might not only alter soil water dynamics and availability, but also change soil organic carbon (SOC) and nutrient cycling and accumulation in soils, compared to monoculture stands. A targeted selection of tree species has a large potential for SOC accumulation (Mayer et al., 2020) with up to 2–5 fold organic carbon (C) stock increase in the forest floor and up to two fold in mineral soil (Vesterdal et al., 2013). Organic C in soils is relevant for a wide range of soil functions and ecosystem services such as water and nutrient retention, erosion control and climate regulation (Lal, 2006; Lal et al., 2018). In this context, forests are increasingly discussed as option to mitigate greenhouse gas emissions (Hulvey et al., 2013). According to the United Nations framework convention on climate change of 1992 and the Kyoto protocol of 1997, C sequestration in terrestrial sinks by silvicultural management, including a change in tree species, has been accepted to offset carbon dioxide emissions for national carbon budgets.
However, information on admixing tree species to increase forest stand resilience in the context of climate change, and the potential consequences for SOC and total nitrogen (TN) stocks are scarce, but urgently required for forest management and policy. Effects of tree species on SOC and TN stocks have been studied mostly using common garden experiments or retrospective analyses (comparison of paired stands or single trees). Earlier research mostly addressed the differences in SOC stocks of monoculture forest stands (Ovington, 1956; Vesterdal and Raulund-Rasmussen, 1998; Gurmesa et al., 2013). In most cases coniferous and broadleaved stands were compared, with conifers revealing generally higher organic C stocks in the forest floor with rather inconsistent effects in the mineral soil (Vesterdal et al., 2013). Later, also interest in mixed forests increased (Borken and Beese, 2005; Cremer et al., 2016; Dawud et al., 2017). The most recent reviews on the effects of tree species on SOC stocks in temperate forests were conducted in the last decade (Vesterdal et al., 2013; Boča et al., 2014), however, comprehensive summaries on SOC stocks in mixed forest stands do not exist. So far, only the contribution of tree mixtures and tree species diversity to aboveground carbon sequestration have been reviewed (Hulvey et al., 2013).
Due to the high relevance of European beech in Europe, our central aim was to synthesize results from studies that compared SOC and TN stocks of beech monocultures with those of mixed beech stands as well as monocultures of other tree species. Our objectives were to: (1) detect effects of admixing other tree species to beech stands on SOC and TN stocks, (2) compare these stocks under beech monocultures with those of other monocultures, (3) identify soil and climatic factors that indicate the potential for SOC/TN stock increase due to admixture, and (4) identify knowledge gaps and future research needs. We hypothesized that (1) admixture effects on SOC stocks cannot be linearly derived from stocks in the corresponding monoculture stands due to non-additive and synergistic species mixture effects, and (2) that admixture of coniferous trees to beech stands results in larger increases of SOC and TN stocks than admixture of other broadleaved species. Finally, we expected that (3) the admixing effects vary depending on both soil and climate conditions.
Materials and Methods
Data Compilation
A literature search was performed up to 13.12.2019 using different databases (see Supplement 1 for flow diagram of protocol). Besides others, key words used and differently combined were tree species, beech, tree diversity, soil organic carbon, carbon, stocks, storage as well as different countries and different tree species. The articles had to match the following criteria to be included in the meta-analysis: (1) The experiments used paired stands/designs (i.e., common garden experiments or retrospective analyses with adjacent stands on the same parent material, similar history and stand age) of European beech and at least one other tree species/tree species mixture with beech; (2) organic C stocks in the forest floor and/or mineral soil were reported or could be calculated; and (3) sampling designs included replicated measurements (n ≥ 3). In case, TN stocks were reported or could be calculated, we also collected information on this parameter. Data were extracted from tables or graphs by digitalization using GetData Graph Digitizer (version 2.26, Russian Federation). If a study included multiple observations at different sites, each site was regarded as independent study and included in the analysis. We recorded the data only once, if different studies were using the same data/ study site. Original SOC/TN stock data were converted to standard units (Mg ha−1). In case, only SOC/TN concentrations were reported, stocks were calculated using bulk density (BD) and sampling depth. If BDs were not determined (2 cases), they were estimated using the pedotransfer function from Post and Kwon (2000) (Equation 1) based on the organic matter (OM) content.
Sampling depth for SOC stocks of the various studies ranged from 2 to 100 cm (Table 1). The mean sampling depth was 35 cm and for the comparison of SOC/TN stock data from various studies with different soil depths, a depth of 50 cm was selected. Although this does not reflect the full picture of SOC stocks, differences in SOC stocks due to species composition generally occur in the upper mineral soil (Jandl et al., 2014; Jonard et al., 2017). Original SOC/TN stock data presenting stocks for a depth <50 cm (21 studies, see Table 1) were extrapolated to 50 cm using the depth distribution functions from Jobbágy and Jackson (2000, 2001) (Equations 2, 3), where Y is the cumulative proportion of the SOC/TN stock from the soil surface to the depth d (cm). The relative rate of SOC stock decrease with depth (β) was calculated separately for deciduous stands (0.9656), coniferous (0.9673) and mixed beech-coniferous stands (0.9665). The SOC/TN stock in the upper 50 cm (X50) was calculated using β, the original SOC/TN stock (Mg ha−1) (Xd0) and the original soil depth available in individual studies (cm) (d0) (Equation 3).
A test based on available measured profile data revealed that this extrapolation to 50 cm had neither effects on SOC and TN stocks nor on assessment of tree species effects (Supplement 2).
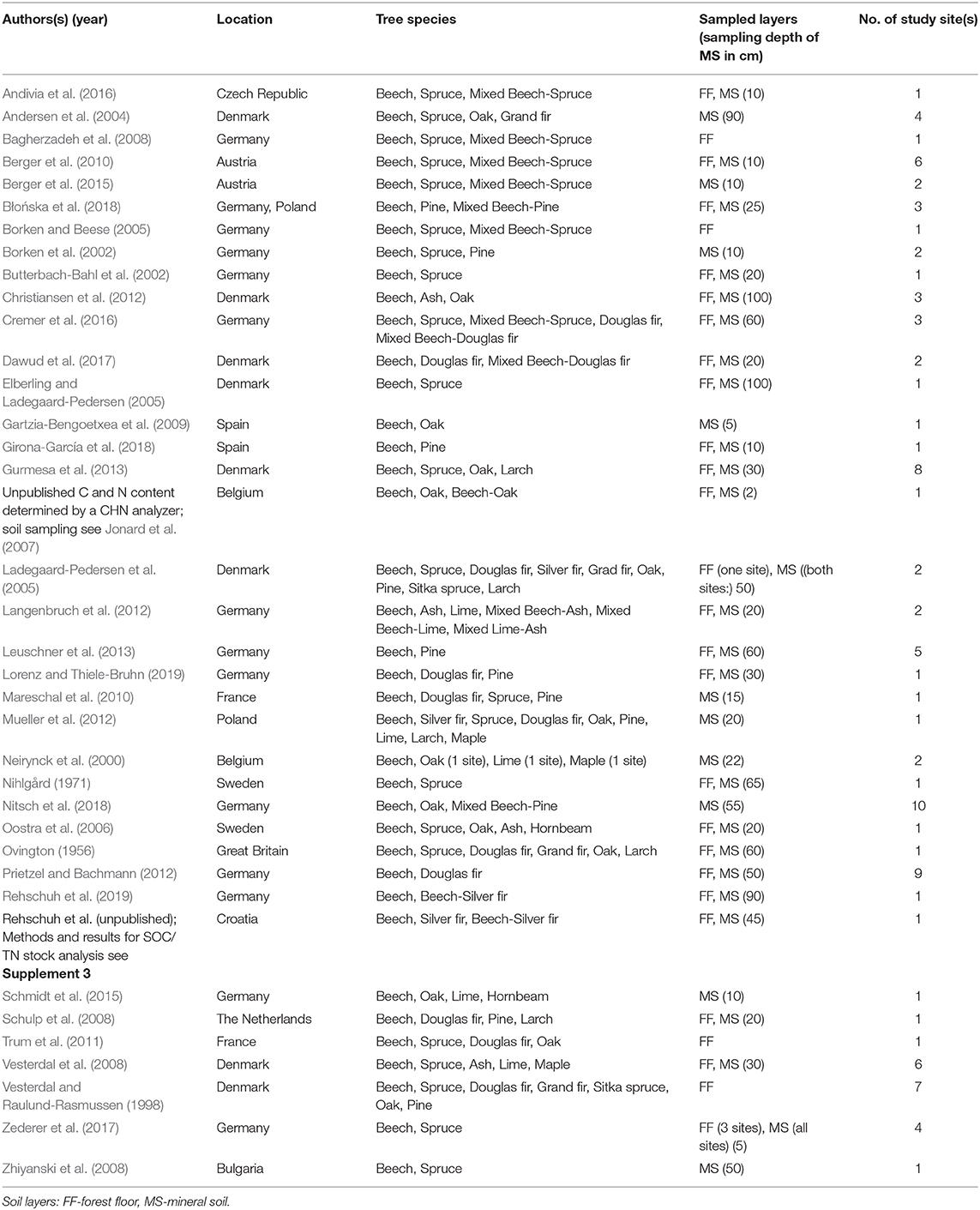
Table 1. References included in the SOC stock database for meta-analysis.
Studies were also included when only organic C stocks for the forest floor or the mineral soil were reported. From reports presenting both, forest floor plus mineral soil, total SOC stocks were calculated. The dataset was split into three subsets: forest floor only, mineral soil only (50 cm), and total soil profile to 50 cm. Thus, we obtained a meta-data set of 203, 220, and 160 observations for forest floor, mineral soil and total soil profile from 38 studies, respectively (list of data sources see Table 1). European beech and comparisons with other species, were investigated from its northern to it's southern distribution limit. However, studies in Mediterranean and boreal climate region were very limited (two studies in the Mediterranean) (Table 1). Data were collected from 13 countries, with dominating research in Germany and Denmark (Figure 1).
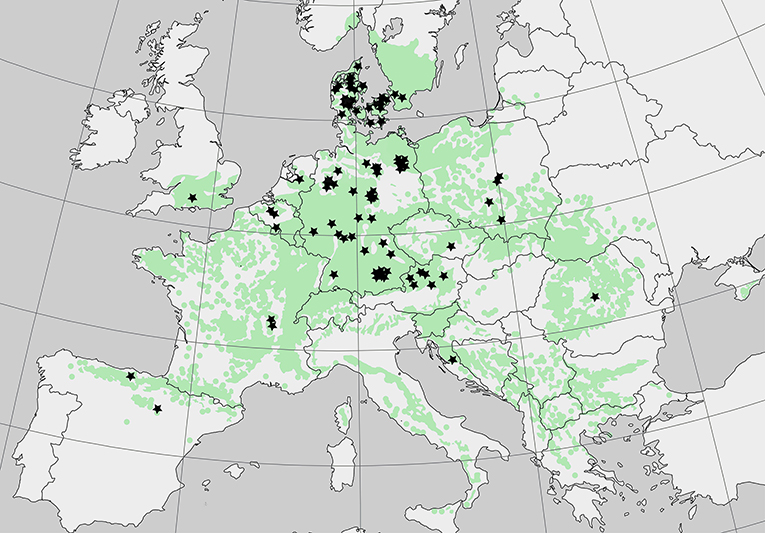
Figure 1. Distribution of forest sites included in the meta-analysis (n = 102) illustrated by stars. The green color indicates the potential natural distribution area of European beech. Source: http://www.euforgen.org/species/fagus-sylvatica/.
We compiled information of SOC/TN stocks for 12 different tree species and seven types of mixtures with beech, each compared with monocultures of European beech (Table 1). Information of little studied species (e.g., Larix decidua) were merged with information on species of the same genus (e.g., Larix kaempferi) under their genus name (larch), if species effects showed the same trend. Species included are as follows (English names are used in text and figures): Norway and Sitka spruce (Picea abies & P. sitchensis), Douglas fir (Pseudotsuga menziesii), silver and Grand fir (Abies alba & A. grandis), larch (Larix decidua & L. kaempferi), pine (Pinus sylvestris; P. contorta; P. nigra), ash (Fraxinus excelsior), sycamore maple (Acer pseudoplatanus), lime (Tilia platyphyllos & T. cordata), oak (Quercus robur & Q. petraea), and hornbeam (Carpinus betulus). Main research from larch was on Japanese larch (Larix kaempferi), from oak on pedunculate oak (Quercus robur) and from spruce on Norway spruce (Picea abies). The age of the forest stands ranged between 27 and 160 years. If not stated otherwise, “broadleaved” refers to “broadleaved other than beech.” From the 14 studies investigating mixed stands, eight indicated the mixing composition: The percentage of beech in the mixtures ranged between 40 and 60%, with an average of 49%. Since the information on the mixture composition was limited, the dataset did not allow us to derive conclusions for different mixing ratios.
For each study we compiled meta-data as predictor variables, i.e., information regarding location, climate, stand, and soil properties (Supplement 1). For unification, we transferred the various soil taxonomies from national soil classifications used by the individual studies to the international classification of the World Reference Base for Soil Resources (IUSS Working Group WRB, 2015). Soil texture was summarized in four categories from sandy to loamy clay/clayey loam. Furthermore, the percentage of the sand content was extracted. If only the soil texture class was given, the percentage of sand was estimated using the soil texture triangle. We selected the months May to October as an approach to represent growing season climate and to calculate the drought index for each of this month (Equation 4). The drought index was later averaged for the whole 6 months period.
Information on nitrogen deposition was collected from external sources (e.g.,1 for Belgium or2 for Germany).
Data Calculation, Statistical Analysis and Presentation
Since standard deviations were mostly not presented in the desired soil depth and a sum of standard deviations from each single soil layer would have created even a higher bias, an unweighted meta-analysis was used as has been common in previous SOC stock studies (Guo and Gifford, 2002; Li et al., 2012; Boča et al., 2014). For all publications included in the meta-analysis, the response ratio was calculated as the absolute annual difference of SOC stocks between the various tree species and mixtures in comparison to European beech monoculture (in the following shortened by using the phrase “tree-species-induced differences in SOC stocks”) (Equation 5). In addition, the percentage differences in SOC stock were calculated.
As the number of observations was relatively scarce for some tree species, we combined results to different groups (conifers, broadleaved, mixed beech-conifers, and mixed beech-broadleaved). The difference between the SOC stock of the control (beech monoculture) and the treatment (various other tree species and mixtures) was considered significant at p < 0.05 when the 95% confidence interval did not include 0% difference (i.e., no change) in SOC stocks.
Multivariate models were developed to explore the impact of potential soil and environmental controls on tree-species-induced organic C stock differences in the forest floor and the mineral soil. To correct the percentage differences in SOC stocks for heteroscedasticity, values were log transformed. For the selection of the most relevant variables to be included in the model, we used multiple stepwise regressions with Akaike information criterion (AIC) for best model selection. The following potential predictor variables were considered for the extent of tree-species-induced SOC stock differences: stand type (categorical), forest floor/mineral soil organic C stock of the reference beech monoculture stand, forest floor/mineral soil C/N ratio, forest floor organic C stock of the respective stand, sand content in the mineral soil, nitrogen deposition, stand age of the paired stands, mean annual temperature (MAT) and mean annual precipitation (MAP) (all numeric).
Statistical analyses and data presentation were performed using R 3.5.3 (R Core Team, 2017). To test for normal distribution and homogeneity of variances the Shapiro-Wilk test and the Levene test were used at p < 0.05. The one-way analysis of variance (ANOVA) followed by the Tukey post hoc test (for normally distributed data) and the Kruskal–Wallis test followed by the Bonferroni post hoc test (for non-normally distributed data) was applied to compare mean values among various groups with p < 0.05 set as threshold for significant differences.
To test for interactive effects of species mixtures, we analyzed experimental designs that included a beech monoculture, a mixed beech stand and a monoculture of the admixed species. The Friedman test, a non-parametric paired test, followed by the Wilcoxon-Nemenyi-McDonald-Thompson post hoc test was used to test for significant differences between the SOC stocks of the mixed stand, beech monoculture and the monoculture of the admixed species. To test for the type of mixing effect (additive vs. non-additive) we compared the expected SOC stocks for the mixture (calculated based on the proportions for the corresponding monocultures) with the observed SOC stocks for the mixtures. From the regression patterns mixing effects can be derived (Figure 2). After assessing data distribution and homogeneity of variance, the paired t-test allowed to test whether the expected SOC stock was significantly different from the observed SOC stock.
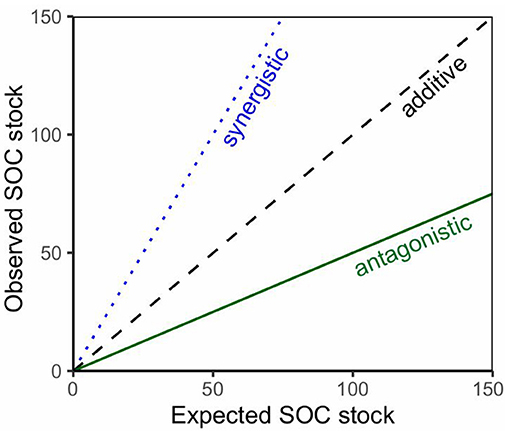
Figure 2. Mixing effects derived from the relation of expected SOC stocks of mixed forest stands (based on calculated stocks using the corresponding monocultures) and observed SOC stocks for the mixed stands.
Results
We found generally more observations comparing beech monoculture with other monocultures than comparing beech monoculture with mixed-beech stands (Supplement 4). Especially for spruce and oak, mixtures with beech were rarely studied: While there were ~40 observations comparing spruce and oak monocultures with beech monocultures, only 10–13 and one observations compared beech vs. spruce-beech and oak-beech mixed stands, respectively. Generally, tree-species induced effect sizes (expressed as % SOC stock difference) were largest in the forest floor (Supplement 5). Because of the small size of the forest floor C pool compared to the mineral soil C pool, annual changes in SOC accrual for the entire soil profile (Mg ha−1 yr−1 of C) were however mostly dominated by effects in the mineral soil (Figure 3).
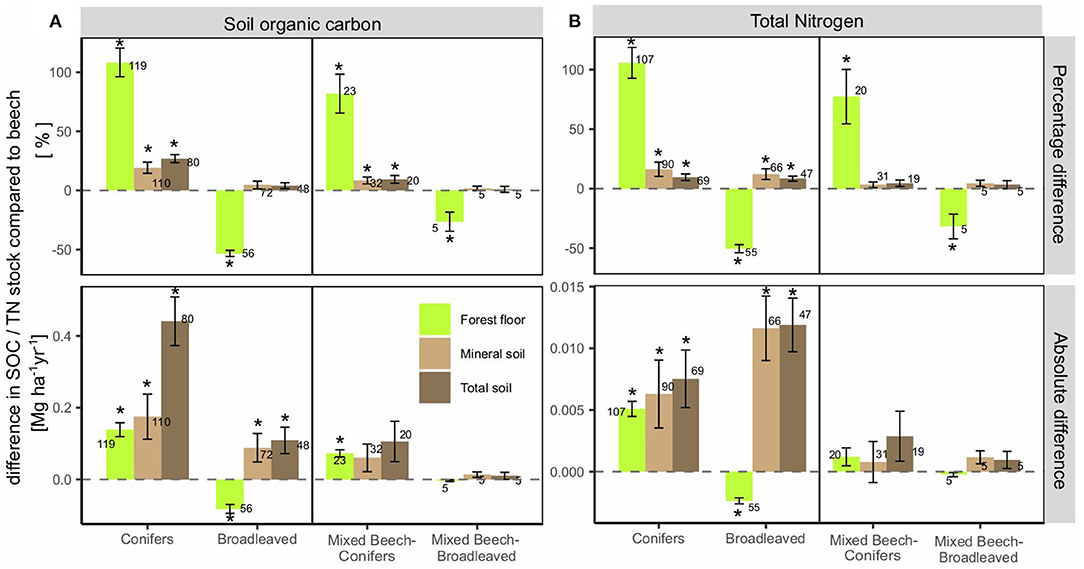
Figure 3. Average percentage and absolute annual differences of (A) SOC and (B) TN stocks summarized for monocultures (conifers, broadleaved) and mixtures with beech (mixed beech-conifers/mixed beech- broadleaved) compared to beech monocultures. Average mixing ratio: 50%/50%. Numbers next to the bars are the number of observations. Error bars show the standard error. Asterisks indicated significant differences compared to beech monocultures. Note that in some cases the difference for the total soil is not the sum of those for the forest floor and mineral soil, since not all studies included observations for both layers.
Tree Type Effects on SOC Stocks
Conifer monocultures showed significantly higher SOC accumulation in all soil layers when compared with beech monocultures (Figure 3A). Broadleaved monoculture stands other than beech had substantially less organic C in the forest floor compared to beech, but a larger SOC accrual in the mineral soil and total soil profile (50 cm) (0.09 Mg C ha−1 yr−1, p < 0.05). Comparing mixed stands to beech monocultures generally revealed similar effects as indicated from comparing monocultures (Figure 3A). Mixed beech-conifer forests had on average 9% more SOC in the total soil profile, which translated into an increased SOC accrual of 0.1 Mg C ha−1 yr−1 (Figure 3A). In contrast, mixtures of beech with other broadleaved tree species on average showed similar SOC stocks as compared to beech monocultures.
Effects of Single Tree Species on SOC Stocks
Among the coniferous tree species, monocultures of larch, spruce, Douglas fir as well as silver/Grand fir showed the largest increases in total SOC stocks compared to beech monoculture, while only pine monocultures had similar SOC stocks as beech monoculture (Figure 4, Supplement 5A). Coniferous forest floor organic C stocks were generally considerably higher when compared to beech with up to 200% or 0.32 Mg ha−1 yr−1 in larch forests, that was also the case for mineral organic C stocks that revealed up to 45% or 0.30 Mg ha−1 yr−1 higher stocks (silver & Grand fir stands) (Figure 4). Monoculture stands of oak, ash, lime, and maple stored significantly less organic C in the forest floor than beech monocultures (41–72%), but more in the mineral soil (up to 28%), which overall slightly higher SOC stocks in broadleaved monocultures other than beech (Figure 4, Supplement 5A).

Figure 4. Average absolute annual differences of SOC stocks of monoculture stands compared to beech monoculture. Numbers next to the bars are the number of observations. Error bars show the standard error. Asterisks indicated significant differences compared to beech monoculture. Note that in some cases the difference for the total soil is not the sum of those for the forest floor and mineral soil, since not all studies included observations for both layers.
European beech stands mixed with the conifers spruce, Douglas fir, pine and silver/Grand fir showed considerably higher organic C stocks in the forest floor (2–101%, 60% on average, Figure 5, Supplement 5B) compared to beech monoculture. Up to doubled organic C stocks were found in forest floor of mixed beech-spruce and mixed beech-Douglas fir stands compared to beech monocultures (Supplement 5B). In mineral soil, these effects were most pronounced for pine. This resulted in considerably higher annual differences in total SOC sequestration, in particular for mixed beech-pine stands (increase of 0.36 Mg ha−1 yr−1 of C) (Figure 5). Mixed beech-broadleaved stands are much less studied with the scarce data available indicating marginal SOC changes for mixtures of beech with oak, ash and lime, thereby confirming patterns observed based on comparisons of monocultures (Figure 3A).
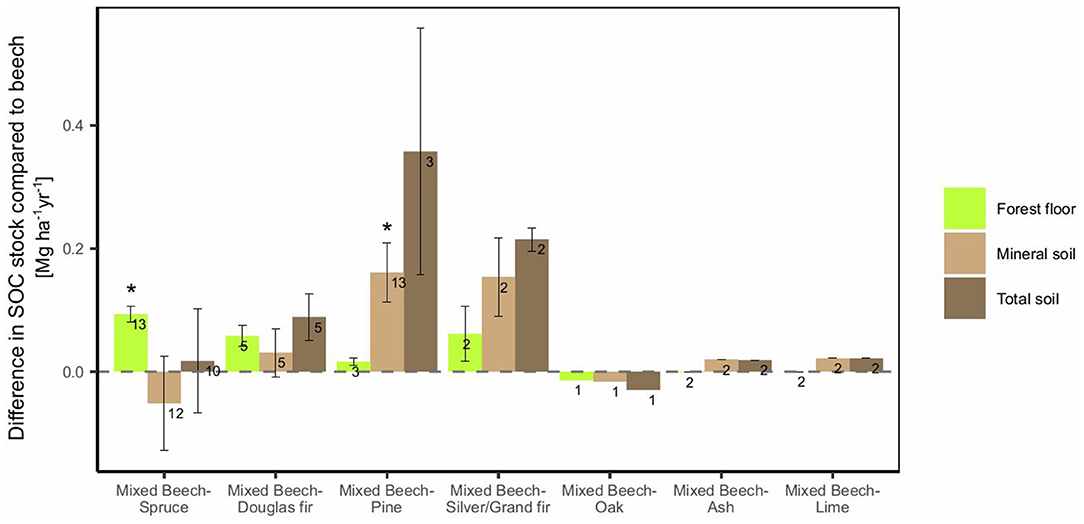
Figure 5. Average absolute annual differences of SOC stocks of mixed forest stands compared to beech monocultures. Numbers next to the bars are the number of observations. Error bars show the standard error. Asterisks indicated significant differences compared to beech monocultures. Note that in some cases the difference for the total soil is not the sum of those for the forest floor and mineral soil, since not all studies included observations for both layers.
The studies that allowed a comparison of beech monoculture with both, mixed stands and monoculture of the admixed species, were analyzed separately. For the forest floor organic C stocks, we found a significant, additive mixing effect for both types of admixtures (conifers, broadleaved species), i.e., the organic C stocks of the mixed stand could be deduced from the mixing ratio (Supplements 6A,D). For the mineral soil, no species composition effect was observed (Supplement 6B) and therefore, no mixing effect. For total SOC stocks, we found no species composition effect and therefore no mixing effect when considering admixtures with broadleaved, but a significant (p < 0.05) and additive species mixing effect for the mixtures with conifers (Supplements 6C,E).
Total N Stocks and C/N Ratios
Tree-species induced differences of TN stocks showed mostly similar patterns as those observed for SOC stocks when compared to beech monoculture (Figure 3B). However, in the mineral and total soil, broadleaved stands showed higher increases in TN stocks compared to beech than conifer stands, which is in contrast to observations of SOC stock differences. The limited number of observations in mixed stands showed TN stock increased in mixed beech-conifer stands by 4% and no change due to admixture of other broadleaved species (Figure 3B).
Strong correlations between the differences of SOC and TN stocks of other monocultures and mixed stands compared to beech were detected (forest floor: R2 = 0.88, mineral soil: R2 = 0.89, total soil: R2 = to 0.69) (Supplements 7A,B). For conifers and mixed beech-conifers, the relative TN stock changes were found to be lower than relative SOC stock changes and the C/N ratio increased in all soil layers (Supplement 7C). For broadleaved species, C/N ratios for the mineral and total soil tended to decrease compared to beech, while mixed beech-broadleaved forest floor C/N ratios were higher than beech C/N ratios.
Controls of Tree-Species Induced SOC Differences
Forest Floor
The multiple stepwise regression analyses showed that the stand type (broadleaved, conifer, mixed beech-conifer, and mixed beech-broadleaved) explained a large part of the tree-species-induced forest floor organic C stock differences. Further, the forest floor organic C stock of the reference beech stand also had predictive power in the multivariate model. In total, 67% of the variance was explained by these factors (Table 2). Differences of organic C stocks between other tree species/mixtures and beech were highest at forest floors with low initial forest floor organic C stocks of 0.4–10 Mg ha−1 and decreased with increasing initial organic C stock (Figure 6A).
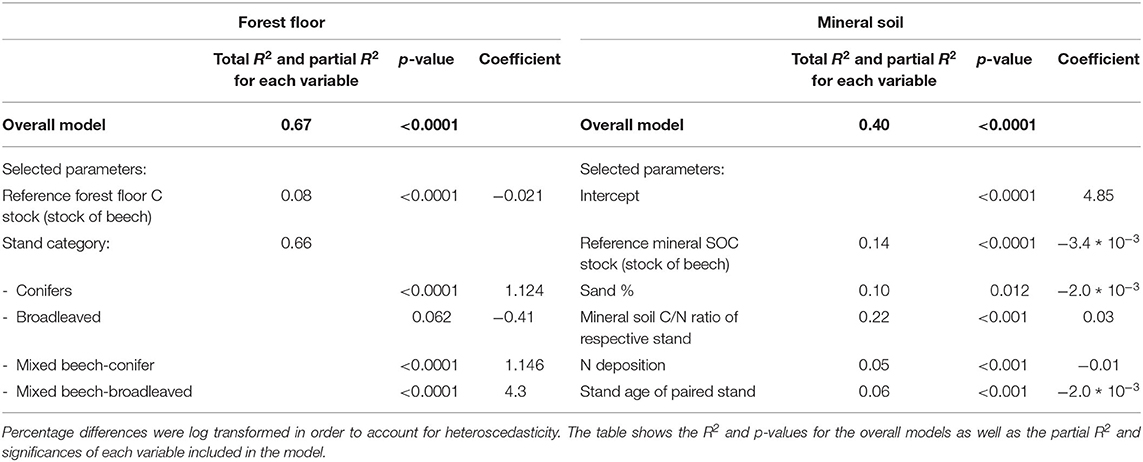
Table 2. Results of the multiple stepwise regression analyses to identify the factors driving the differences in organic C stocks of other tree species/mixtures compared to beech in the forest floor and the mineral soil.
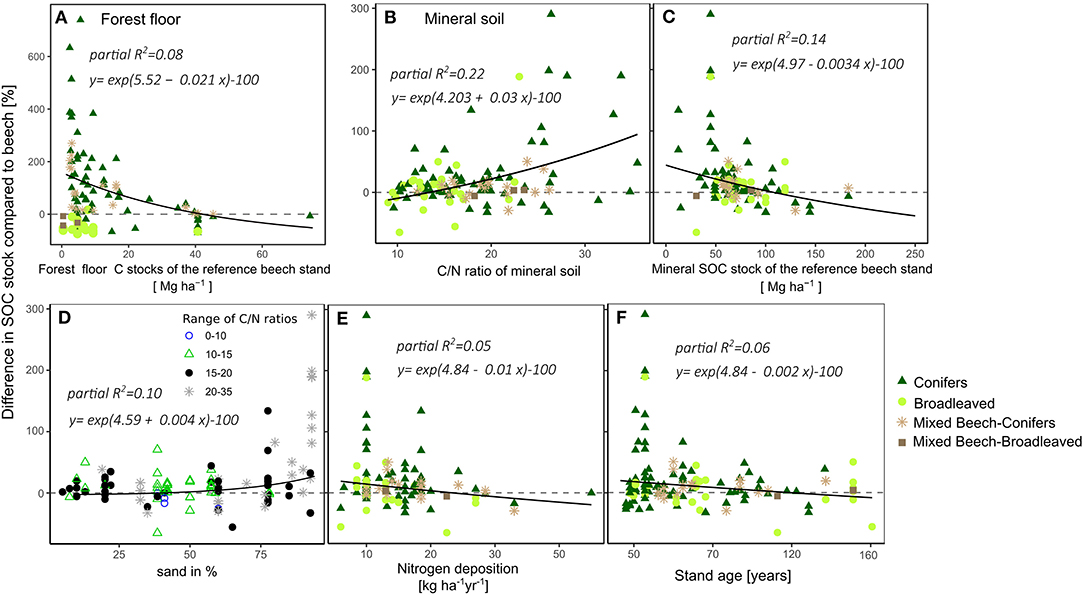
Figure 6. Effects of statistically significant drivers on tree-species-induced differences in forest floor C stocks (A) and mineral SOC stocks (B–F). For forest floor C stock differences, this was the forest floor organic C stock of the reference beech stand (A). For the mineral soil differences, important drivers were (B) the C/N ratio of mineral soil, (C) the reference mineral SOC stock of the beech stand, (D) the soil sand content, (E) nitrogen deposition, and (F) the stand age. The solid lines represent the multivariate model predictions with average values assumed for all of the other variables in the model. The equations were obtained from the full multivariate model using the function: y = exp(4.85 − 3.4 * 10−3 a − 2.0 * 10−3 b + 0.03 c − 0.01 d − 2 * 10−3 e) − 100, with the constants a- Mineral SOC stock of the reference beech stand [Mg ha−1] = 78, b- sand in % = 60, c- C/N ratio of the mineral soil = 16.8, d- Nitrogen deposition [kg ha−1yr−1] = 13.2, e- stand age (years) = 65. The effect of the sand percentage is partly confounded with the other effects. Therefore, the effect of sand content is illustrated using C/N ratios.
Mineral Soil
For the tree-species-induced SOC stock differences in mineral soil, the C/N ratio of the mineral soil of the respective species, SOC stock of mineral soil of the reference beech stand, percentage of sand content in soil, the stand age of the paired stands and nitrogen deposition explained 40% of the variance (Table 2). The mineral soil C/N ratio was with the highest partial R2 the most important predictor of tree-species-induced mineral SOC stock differences. The multivariate model revealed an exponential relation with increasing tree-species-induced SOC stock differences at increasing mineral C/N ratio (Figure 6B). Furthermore, largest differences in SOC stocks due to tree species appeared at stands with a low nitrogen deposition (8–20 kg ha−1yr−1) and at younger forest stands, aged 40–50 years (Figures 6E,F).
Regarding the influence of SOC stocks of beech reference stands, largest tree-species-induced differences in C sequestration were found for low SOC stocks of 12–50 Mg ha−1, with exponentially decreasing differences at increasing beech SOC stock (Figure 6C). Finally, the percentage of sand was an important predictor (partial R2 = 0.1). On sandy soils up to three-fold higher SOC stocks were observed (mean: 57%) for other species or beech mixtures compared to beech monocultures. With a decrease in soil sand content, tree-species-induced differences in SOC stock decreased (Figure 6D). When subdividing the meta-dataset in 4 texture categories from high (sandy) to low (clayey loam/loamy clay) grain sized soils, the largest tree-species-induced SOC stock increase was observed for sandy soils (Supplement 8A). Consequently, also soil type affected tree-species induced SOC changes with Podzols and Regosols showing significantly higher C sequestration rates compared to the other soil types (Acrisol, Cambisol, Chernozem, Histosol, Leptosol, and Luvisol) (Supplement 8C). In this context, legacy effects on SOC stocks were found from the previous land use. Afforested heath and mining lands showed significantly higher C sequestration rate differences of other tree species compared to beech than previous agriculture or forest lands (Supplement 8B).
In contrast to soil parameters, climate parameters, including MAT, MAP, precipitation during growing season and the drought index showed no predictive power for tree-species-induced SOC stock differences (see Table 2 for important parameters).
Discussion
Additive Admixing Effects on SOC and TN Stocks
Tree species admixing effects on SOC stocks could differ from those deduced from comparison of monoculture stands due to non-linear interactions e.g., in litter decomposition. Borken and Beese (2005) investigated two kinds of mixtures of beech with spruce and found similar effects on forest floor organic C stocks, no matter if 30 or 70% spruce were admixed to beech. From this single study, it may be assumed that the mixing ratio does not have large impact on forest floor organic C stocks, possibly due to synergistic or antagonistic interactions. In the present meta-analysis however, additive species mixing effects were observed for the forest floor organic C stocks of mixed stands (Supplements 6A,D, Figure 5). Based on these observations, our hypothesis (1) on synergistic mixture effects cannot be confirmed for the forest floor organic C stocks.
Considering the mineral soil, for mixtures with beech (1) intermediate values between the monoculture stands (Jonard et al., 2007; Cremer et al., 2016), (2) largest SOC stocks in mixed stands (coniferous-beech stands) (Andivia et al., 2016; Błońska et al., 2018; Rehschuh et al., 2019, unpublished), and (3) no detectable differences between beech monoculture and mixed stands (Langenbruch et al., 2012; Berger et al., 2015; Dawud et al., 2017) were observed in individual experimental studies. Synergistic effects of mixed stands on mineral SOC could be due to a more complex structure of the stand and greater density of crowns and rooting systems due to the exploiting of deeper soil layers by beech fine roots when mixed e.g., with spruce (Bolte and Villanueva, 2006; Cremer et al., 2016) with increased belowground biomass and an increased root turnover (Andivia et al., 2016; Błońska et al., 2018). No effects of the admixtures of other tree species to beech were attributed to the prevention of any larger niche differentiation and complementarity because of relatively similar tree species traits (Dawud et al., 2017). The largely additive effects observed in this synthesis point to an overall relatively low importance of such non-linear mixing effects. Consequently, our hypothesis (1) can also not be confirmed for the mineral and total soil, since the SOC stocks of the mixtures can be additively derived from the corresponding monoculture stocks.
Tree-Species Induced SOC and TN Stock Differences
Forest Floor
Our study revealed higher forest floor organic C stocks in mixed beech-conifer stands but lower forest floor OC stocks in beech stands mixed with other broadleaved tree species (Figure 4). That conifers reveal higher organic C stocks in forest floors compared to broadleaved stands such as beech (Prescott and Vesterdal, 2013) is related not only to the more recalcitrant litter (Scheu et al., 2003) with higher lignin and lower nutrient concentrations (especially calcium) (Raich and Tufekcioglu, 2000; Hobbie et al., 2006; Ahmed et al., 2016), but also to the lower faunal and microbial activity due to lower pH-values (Hobbie et al., 2006).
For broadleaved stands, also previous studies reported species-induced differences in forest floor organic C stocks. Vesterdal et al. (2013) showed that relatively low organic C stocks were present under ash, lime and maple, whereas beech and oak were species with organic C stocks intermediate between conifers and other broadleaved species. Beech litter was found to have highest lignin contents and highest C/N ratios with ~53 out of 5 deciduous tree species studied (amongst lime, maple, ash) (Jacob et al., 2010). In the present study, oak forest floors showed with 28 out of 29 observations faster litter decomposition and, hence, lower organic C stocks than beech.
Generally, tree-species induced differences in forest floor organic C stocks can be either due to different litter production or different litter decomposition. Although trees produce similar (Andivia et al., 2016) or even higher litter amounts in broadleaved stands (Butterbach-Bahl et al., 2002), organic C stocks are higher under stands with conifers intermixed, pointing to the importance of processes related to litter decomposition (Augusto et al., 2015). Slower decomposition and, therefore, slower biogeochemical cycles are observed, when litter with lower nutrient concentrations (usually from conifers) are admixed to higher quality litter and vice versa (Chapman et al., 1988). Jonard et al. (2006) found a positive relationship between the forest floor litter mass and the proportion of the most recalcitrant litter. In a litter bag experiment with differently mixed litter of 3–5 broadleaved tree species, Jacob et al. (2010) showed that decomposition rates decreased with increasing proportion of beech, pointing to poorest litter quality of beech leaves among the broadleaved species studied. These results further show that decomposition rather depends on the tree species identity than on tree species diversity (Jacob et al., 2010; Dawud et al., 2016). However, Joly et al. (2017) reported that tree species diversity indirectly affects decomposition due to modified micro-environmental conditions. Apparently, litter fall dynamics and distribution also play an important role for litter decomposition (Rothe and Binkley, 2001; Dawud et al., 2017). In sum, we provide evidence that effects of admixing other tree species to beech forests rather exerts additive effects on forest floor organic C stocks, that can be deduced based on the litter quality of admixed tree species and the mixing ratio. Consequently, conifers are most suitable to increase forest floor organic C stocks of beech stands.
Mineral and Total Soil
The mineral soil showed smaller relative (expressed in % change) but larger absolute (expressed in Mg ha−1 yr−1) tree-species induced changes compared to the forest floor (Figures 4, 5, Supplement 5). This is due to larger residence time, resistance to disturbance, and overall pool sizes of mineral soil compared to forest floor organic C stocks (Hedde et al., 2008; Vesterdal et al., 2013). On average, we detected a plus in SOC sequestration in the mineral soil of 0.2 and 0.1 Mg ha−1 yr−1 for conifers and other broadleaved species, respectively (Figure 3A) compared to beech. These results are generally in line with the findings of Boča et al. (2014), who found conifers compared to beech dominated stands to have on average 26 and 19% more organic C in the forest floor and mineral soil, respectively. Also, Vesterdal et al. (2013) summarized that in temperate climate more mineral SOC is stored under ash, maple, lime, and ulm than under beech. Besides leaf litter inputs, root debris, rhizodeposition (Kuzyakov and Domanski, 2000; Jandl et al., 2007; Trum et al., 2011; Schleuß et al., 2014) and microbial-derived compounds (Angst et al., 2018) are important C inputs into mineral SOC pools, while C output is controlled by decomposition and leaching processes. Usually it is assumed that largest differences due to tree species occur in the top mineral soil because of the great influence of litter incorporation. However, in ancient forests of >230 years also significant differences were found in the subsoil (Nitsch et al., 2018). Generally, the soil organic matter (SOM) of subsoil horizons might be more dynamic than previously thought, possibly driven by root C input (Tefs and Gleixner, 2012). Land use legacy can affect SOC stocks (Li et al., 2012) and thus is a potential confounding factor in meta-analyses comparing different forest stands. Here we minimized such bias by the selection of adjacent paired stands only, and by refusing studies which reported such differences in previous land use. Because only few studies included in the present meta-analysis revealed SOC stocks of the lower subsoil (see Table 1), it was not possible to analyze SOC stocks for deeper soil layers >0.5 m. However, the role of the subsoil in tree-species induced SOC stock differences is assumed to be rather limited (Jandl et al., 2014). Nonetheless, root chemistry is essential. Angst et al. (2019) found that SOC stabilization under different tree species mainly varies due to differences in tissue chemical composition, especially of roots. Tree species with roots rich in nitrogen, but low in calcium contents and low in recalcitrant compounds such as lignin are thought to increase SOM stability (e.g., European larch, Scots pine, and Douglas fir) (Angst et al., 2019). This is because easily decomposable SOM promotes microbial induced stabilization of SOC via organo-mineral compounds (Cotrufo et al., 2013). Therefore, for the selection of tree species to be admixed to beech, in addition to quantifying SOC stocks, also the root litter quality should be considered.
It has been frequently discussed, if there is a difference in the vertical allocation of SOC stocks between conifers and broadleaved species rather than a difference in total SOC stocks. It is suggested that less C in the mineral soil offsets higher C stocks in the forest floor and vice versa (Vesterdal et al., 2008, 2013; Wiesmeier et al., 2012; Prescott and Vesterdal, 2013). In the present study we show that this is possibly evident for broadleaved species other than beech as well as mixed beech-broadleaved stands, showing a slightly negative relation of forest floor organic C stocks and mineral SOC stocks (Supplement 9). However, for coniferous, beech monoculture and mixed-beech-coniferous stands this assumption was not confirmed. Apparently, increases in forest floor organic C are not generally offset by lower mineral SOC storage, but accompanied by larger SOC stocks in mineral soil as well. Around 80% of the conifer stands observed showed higher C stocks in the mineral soil than beech stands. Also in the meta-analysis of Boča et al. (2014), forest floor and mineral organic C patterns were not closely linked when testing forest floor organic C as predictor for mineral SOC stocks. From these contrasting findings, it appears that different species groups have to be distinguished for assumptions on vertical SOC distribution and total stocks.
The present meta-analysis shows that coniferous and other broadleaved stands can sequester significantly more C in the total soil profile than beech stands. Mixed beech-broadleaved stands only showed small differences in total SOC compared to beech. However, mixtures of beech with conifers showed considerably higher total annual SOC accumulation, on average 0.1 Mg ha−1 yr−1 with pine, Douglas fir and silver fir as potential tree species to be intermixed in existing beech forests. Thus, hypothesis (2) of this study was confirmed. For the adaption of beech forests to climate change, admixtures of coniferous tree species could enhance soil C sequestration.
While the admixture of conifers has positive effects on C stocks, nonetheless, tradeoffs should also be mentioned here, such as a decrease in soil pH (Augusto et al., 2015) and a decrease in earthworm biomass and diversity (Ammer et al., 2006). Further, there might be undesired impacts on the soil biological net methane sink, which decreases slightly due to conifer admixture to beech (Menyailo and Hungate, 2003; Rehschuh et al., 2019).
Close Coupling of Tree-Species Induced SOC and TN Stock Changes
Differences in SOC and TN stocks compared to beech monocultures were generally closely coupled (Supplements 7A,B). However, there are differences between the groups of tree species types. The relatively higher TN gain in other broadleaved than conifer stands (Figure 3), i.e., a lower C/N ratio of the mineral soil compared to conifers, could be explained by lower C/N ratios in the fresh leaf litter of broadleaved species. Vice versa, Conifers showed significantly higher C/N ratios compared to beech in all soil layers (Supplement 7C), which in the long term could result in progressive nitrogen limitation (Li et al., 2012). From this observation it appears that admixture of conifers to beech would result in higher C/N ratios. Whether this leads to changes in nitrogen availability however remains uncertain, e.g., due to counterbalancing effects of atmospheric N deposition.
Which Beech Sites Are Most Promising to Increase SOC Stocks Due to Admixture of Other Trees?
We assessed a range of predictors of tree-species induced differences in SOC stocks in order to identify indicators for sites, where admixture of other tree species to beech is most promising. Clearly, the admixed or compared tree species were decisive for forest floor organic C gain compared to beech monocultures, thereby reflecting the importance of tree species-associated recalcitrance of litter on C accumulation in forest floors. However, also a range of soil, site and beech stand properties regulated the potential SOC gains due to other tree species. High carbon gain at low forest floor and mineral soil organic C stock of the reference beech stand (Figures 6A,C) might reflect enhanced productivity of admixed tree species and a relatively fast change of forest floor organic C due to production of more recalcitrant litter.
Also with increasing mineral soil C/N ratio (Figure 6B) as well as at stands with lower nitrogen deposition (Figure 6E), larger tree-species induced increases in SOC stocks were found. This might be due to the fact, that SOM decomposition is reduced due to nitrogen limitation and this effect is stronger for species with more recalcitrant woody and leaf debris. Further, largest differences in mineral SOC sequestration rates between various other tree species and beech were found in soils with a sandy texture such as Podzols and Regosols, and sites with the historical land cover/use of heath and mining, that typically go along with low SOC stocks (Figures 6C,D and Supplement 8). Dawud et al. (2017) investigated beech, Douglas fir and mixtures on sandy and clayey soils and found greater impacts of tree species admixture in sandy soils. Also in another study on C sequestration in forest soils, sandy soils showed higher enrichments in C compared to clayey and calcareous soils (Grüneberg et al., 2014). In general, larger SOC stocks and a better stabilization due to associations of C in organo-mineral complexes and aggregation are found in clayey and loamy soils rather than in sandy soils (Jandl et al., 2007; Schleuß et al., 2014; Angst et al., 2018). SOC stocks in the mineral-associated organic matter fraction are mainly controlled by pedogenic properties (Grüneberg et al., 2013). Hence, we assume that clayey and loamy soils buffer tree species effects better than sandy soils because the soil properties might more strongly control SOC stocks than the chemical recalcitrance of litter inputs. In contrast, in sandy soils with relatively low stabilization of organic C in aggregates and organo-mineral associations, the input of chemically recalcitrant litter from coniferous trees as well as a higher productivity will result in higher relative differences of SOC (see also Grüneberg et al., 2014). In sum, on sandy and low carbon soils, such as Podzols, Regosols, previous heath and mining lands, as well as on soils with nitrogen limitation, admixtures of other tree species to beech are most effective in terms of C sequestration, thereby confirming hypothesis (3) part one.
With hypothesis (3) part two, we expected an effect of climate parameters such as MAT, MAP, precipitation during growing season and the drought index on the magnitude of tree-species induced differences, which could not be confirmed (Table 2). Climate is assumed to have a higher effect on mineral SOC stocks than tree species (Vesterdal et al., 2013), i.e., with drier climate, lower microbial activity is expected and thus an enrichment in SOC stocks. A legible effect of climate in our study might be explained by the fact, that most of the studies included were conducted in temperate regions with similar MAP or MAT and only two in the subtropics. Apparently, for the sampled dataset with its relatively narrow climate variability, the effect of tree species on SOC stocks is not substantially influenced by climate parameters.
Conclusion
Based on a meta-analysis we show that the admixture of conifers to beech forests can increase carbon accumulation in soils by on average 9% which corresponds to 0.1 Mg ha−1 yr−1. The highest potential to increase SOC of beech forests is found for soils with sandy texture, low organic matter content, and nitrogen limitation (low N deposition and high C/N ratios). As this meta-analysis highlighted a lack of studies on the most climate-change-sensitive beech stands at the Southern distribution limit, future studies should fill this gap. Furthermore, an improved understanding of C and N stabilization mechanisms in soil, and quantification of both above- and belowground carbon sequestration, are desirable for developing further improved guidelines for forest compositions with high ecosystem carbon sequestration in a changing climate.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
SR, MD, and HR devised the idea of the study. SR extracted the data and conducted the analysis of the results with contribution of MD and MJ. SR wrote the original article with review and editing from all other authors. All authors discussed the results.
Funding
This work was supported by the Bundesanstalt für Landwirtschaft und Ernährung (BLE), Germany, by the Bundesministerium für Ernährung und Landwirtschaft (BMEL) and the Bundesministerium für Umwelt, Naturschutz, Bau und Reaktorsicherheit (BMUB) within the program Waldklimafonds (No. 28W-C-1-069-01), based on the decision of the German Federal Parliament. SR acknowledge support from the Helmholtz Research School MICMoR on Mechanisms and Interactions of Climate Change in Mountain Regions. This article has been funded through the Open Access Publishing Fund of Karlsruhe Institute of Technology.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Jörg Prietzel, Seid Muhie Dawud, Karsten Raulund-Rasmussen, Marcus Schmidt, Thorsten Berger, Miglena Zhiyanski, Inazio Martinez de Arano, and Nahia Gartzia Bengoetxea for their help, further information and additional data, we highly appreciated it. Frederik De Roo and Rüdiger Grote are thanked for help on data collection and processing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2021.606669/full#supplementary-material
Footnotes
References
Ahmed, I. U., Smith, A. R., Jones, D. L., and Godbold, D. L. (2016). Tree species identity influences the vertical distribution of labile and recalcitrant carbon in a temperate deciduous forest soil. For. Ecol. Manage. 359, 352–360. doi: 10.1016/j.foreco.2015.07.018
Ammer, S., Weber, K., Abs, C., Ammer, C., and Prietzel, J. (2006). Factors influencing the distribution and abundance of earthworm communities in pure and converted Scots pine stands. Appl. Soil Ecol. 33, 10–21. doi: 10.1016/j.apsoil.2005.09.005
Andersen, M. K., Raulund-Rasmussen, K., Strobel, B. W., and Hansen, H. C. B. (2004). The effects of tree species and site on the solubility of Cd, Cu, Ni, Pb and Zn in soils. Water Air Soil Pollut. 154, 357–370. doi: 10.1023/B:WATE.0000022991.59456.01
Andivia, E., Rolo, V., Jonard, M., Formánek, P., and Ponette, Q. (2016). Tree species identity mediates mechanisms of top soil carbon sequestration in a Norway spruce and European beech mixed forest. Ann. For. Sci. 73, 437–447. doi: 10.1007/s13595-015-0536-z
Angst, G., Messinger, J., Greiner, M., Häusler, W., Hertel, D., Kirfel, K., et al. (2018). Soil organic carbon stocks in topsoil and subsoil controlled by parent material, carbon input in the rhizosphere, and microbial-derived compounds. Soil Biol. Biochem. 122, 19–30. doi: 10.1016/j.soilbio.2018.03.026
Angst, G., Mueller, K. E., Eissenstat, D. M., Trumbore, S., Freeman, K. H., Hobbie, S. E., et al. (2019). Soil organic carbon stability in forests: distinct effects of tree species identity and traits. Glob. Chang. Biol. 25, 1529–1546. doi: 10.1111/gcb.14548
Augusto, L., De Schrijver, A., Vesterdal, L., Smolander, A., Prescott, C., and Ranger, J. (2015). Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests: spermatophytes and forest functioning. Biol. Rev. 90, 444–466. doi: 10.1111/brv.12119
Bagherzadeh, A., Brumme, R., and Beese, F. (2008). Temperature dependence of nitrogen mineralization and microbial status in OH horizon of a temperate forest ecosystem. J. Forestry Res. 19, 37–43. doi: 10.1007/s11676-008-0006-7
Berger, T. W., Duboc, O., Djukic, I., Tatzber, M., Gerzabek, M. H., and Zehetner, F. (2015). Decomposition of beech (Fagus sylvatica) and pine (Pinus nigra) litter along an Alpine elevation gradient: decay and nutrient release. Geoderma 251–252, 92–104. doi: 10.1016/j.geoderma.2015.03.024
Berger, T. W., Inselsbacher, E., and Zechmeister-Boltenstern, S. (2010). Carbon dioxide emissions of soils under pure and mixed stands of beech and spruce, affected by decomposing foliage litter mixtures. Soil Biol. Biochem. 42, 986–997. doi: 10.1016/j.soilbio.2010.02.020
Błońska, E., Klamerus-Iwan, A., Lasota, J., Gruba, P., Pach, M., and Pretzsch, H. (2018). What characteristics of soil fertility can improve in mixed stands of scots pine and european beech compared with monospecific stands? Commun. Soil Sci. Plant Anal. 49, 237–247. doi: 10.1080/00103624.2017.1421658
Boča, A., Van Miegroet, H., and Gruselle, M.-C. (2014). Forest overstory effect on soil organic carbon storage: a meta-analysis. Soil Sci. Soc. Am. J. 78:S35. doi: 10.2136/sssaj2013.08.0332nafsc
Bolte, A., Czajkowski, T., and Kompa, T. (2007). The north-eastern distribution range of European beech a review. Forestry 80, 413–429. doi: 10.1093/forestry/cpm028
Bolte, A., and Villanueva, I. (2006). Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur. J. For. Res. 125, 15–26. doi: 10.1007/s10342-005-0075-5
Borken, W., and Beese, F. (2005). Soil respiration in pure and mixed stands of European beech and Norway spruce following removal of organic horizons. Canadian J. Forest Res. 35, 2756–2764. doi: 10.1139/x05-192
Borken, W., Xu, Y.-J., Davidson, E. A., and Beese, F. (2002). Site and temporal variation of soil respiration in European beech, Norway spruce, and scots pine forests. Global Change Biology 8, 1205–1216. doi: 10.1046/j.1365-2486.2002.00547.x
Butterbach-Bahl, K., Rothe, A., and Papen, H. (2002). Effect of tree distance on N2O and CH4 fluxes from soils in temperate forest ecosystems. Plant and Soil 240, 91–103. doi: 10.1023/A:1015828701885
Chapman, K., Whittaker, J. B., and Heal, O. W. (1988). Metabolic and faunal activity in litters of tree mixtures compared with pure stands. Agric. Ecosyst. Environ. 24, 33–40. doi: 10.1016/0167-8809(88)90054-0
Christiansen, J. R., Gundersen, P., Frederiksen, P., and Vesterdal, L. (2012). Influence of hydromorphic soil conditions on greenhouse gas emissions and soil carbon stocks in a Danish temperate forest. Forest Ecol. Manage. 284, 185–195. doi: 10.1016/j.foreco.2012.07.048
Cotrufo, M. F., Wallenstein, M. D., Boot, C. M., Denef, K., and Paul, E. (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 19, 988–995. doi: 10.1111/gcb.12113
Cremer, M., Kern, N. V., and Prietzel, J. (2016). Soil organic carbon and nitrogen stocks under pure and mixed stands of European beech, Douglas fir and Norway spruce. For. Ecol. Manage. 367, 30–40. doi: 10.1016/j.foreco.2016.02.020
Dannenmann, M., Bimüller, C., Gschwendtner, S., Leberecht, M., Tejedor, J., Bilela, S., et al. (2016). Climate change impairs nitrogen cycling in European beech forests. PLoS ONE 11:e0158823. doi: 10.1371/journal.pone.0158823
Dawud, S. M., Raulund-Rasmussen, K., Domisch, T., Finér, L., Jaroszewicz, B., and Vesterdal, L. (2016). Is tree species diversity or species identity the more important driver of soil carbon stocks, C/N Ratio, and pH? Ecosystems 19, 645–660. doi: 10.1007/s10021-016-9958-1
Dawud, S. M., Vesterdal, L., and Raulund-Rasmussen, K. (2017). Mixed-species effects on soil C and N stocks, C/N ratio and pH using a transboundary approach in adjacent common garden douglas-fir and beech stands. Forests 8:95. doi: 10.3390/f8040095
Elberling, B., and Ladegaard-Pedersen, P. (2005). Subsurface CO2 Dynamics in Temperate Beech And Spruce Forest Stands. Biogeochemistry 75, 479–506. doi: 10.1007/s10533-005-3690-9
Ellenberg, H. H. (1988). Vegetation Ecology of Central Europe. Cambridge: Cambridge University Press.
Gartzia-Bengoetxea, N., González-Arias, A., Merino, A., and Martínez de Arano, I. (2009). Soil organic matter in soil physical fractions in adjacent semi-natural and cultivated stands in temperate Atlantic forests. Soil Biol. Biochem. 41, 1674–1683. doi: 10.1016/j.soilbio.2009.05.010
Geβler, A., Keitel, C., Nahm, M., and Rennenberg, H. (2004). Water shortage affects the water and nitrogen balance in central European beech forests. Plant Biol. 6, 289–298. doi: 10.1055/s-2004-820878
Girona-García, A., Badía-Villas, D., and Martí, C. (2018). Changes in Topsoil Properties after Centennial Scots Pine Afforestation in a European Beech Forest (NE Spain). Forests 9:343. doi: 10.3390/f9060343
Grüneberg, E., Schöning, I., Hessenmöller, D., Schulze, E.-D., and Weisser, W. W. (2013). Organic layer and clay content control soil organic carbon stocks in density fractions of differently managed German beech forests. For. Ecol. Manage. 303, 1–10. doi: 10.1016/j.foreco.2013.03.014
Grüneberg, E., Ziche, D., and Wellbrock, N. (2014). Organic carbon stocks and sequestration rates of forest soils in G ermany. Glob Change Biol. 20, 2644–2662. doi: 10.1111/gcb.12558
Guo, L. B., and Gifford, R. M. (2002). Soil carbon stocks and land use change: a meta analysis. Glob. Chang. Biol. 8, 345–360. doi: 10.1046/j.1354-1013.2002.00486.x
Gurmesa, G. A., Schmidt, I. K., Gundersen, P., and Vesterdal, L. (2013). Soil carbon accumulation and nitrogen retention traits of four tree species grown in common gardens. For. Ecol. Manage. 309, 47–57. doi: 10.1016/j.foreco.2013.02.015
Hedde, M., Aubert, M., Decaëns, T., and Bureau, F. (2008). Dynamics of soil carbon in a beechwood chronosequence forest. For. Ecol. Manage. 255, 193–202. doi: 10.1016/j.foreco.2007.09.004
Hobbie, S. E., Reich, P. B., Oleksyn, J., Ogdahl, M., Zytkowiak, R., Hale, C., et al. (2006). Tree speceis effects on decomposition and forest floor dynamics in a common garden. Ecology 87, 2288–2297. doi: 10.1890/0012-9658(2006)87[2288:T.S.E.O.D.A.]2.0.C.O.;2
Hulvey, K. B., Hobbs, R. J., Standish, R. J., Lindenmayer, D. B., Lach, L., and Perring, M. P. (2013). Benefits of tree mixes in carbon plantings. Nat. Clim. Change 3, 869–874. doi: 10.1038/nclimate1862
IUSS Working Group WRB (2015). World Reference Base for Soil Resources 2014, update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. Rome: FAO.
Jacob, M., Viedenz, K., Polle, A., and Thomas, F. M. (2010). Leaf litter decomposition in temperate deciduous forest stands with a decreasing fraction of beech (Fagus sylvatica). Oecologia 164, 1083–1094. doi: 10.1007/s00442-010-1699-9
Jandl, R., Lindner, M., Vesterdal, L., Bauwens, B., Baritz, R., Hagedorn, F., et al. (2007). How strongly can forest management influence soil carbon sequestration? Geoderma 137, 253–268. doi: 10.1016/j.geoderma.2006.09.003
Jandl, R., Rodeghiero, M., Martinez, C., Cotrufo, M. F., Bampa, F., van Wesemael, B., et al. (2014). Current status, uncertainty and future needs in soil organic carbon monitoring. Sci. Total Environ. 468–469, 376–383. doi: 10.1016/j.scitotenv.2013.08.026
Jobbágy, E. G., and Jackson, R. B. (2000). The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10, 423–436. doi: 10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2
Jobbágy, E. G., and Jackson, R. B. (2001). The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 53, 51–77. doi: 10.1023/A:1010760720215
Joly, F.-X., Milcu, A., Scherer-Lorenzen, M., Jean, L.-K., Bussotti, F., Dawud, S. M., et al. (2017). Tree species diversity affects decomposition through modified micro-environmental conditions across European forests. New Phytol. 214, 1281–1293. doi: 10.1111/nph.14452
Jonard, M., André, F., Jonard, F., Mouton, N., Procès, P., and Ponette, Q. (2007). Soil carbon dioxide efflux in pure and mixed stands of oak and beech. Ann. For. Sci. 64, 141–150. doi: 10.1051/forest:2006098
Jonard, M., Andre, F., and Ponette, Q. (2006). Modeling leaf dispersal in mixed hardwood forests using a ballistic approach. Ecology 87, 2306–2318. doi: 10.1890/0012-9658(2006)87[2306:MLDIMH]2.0.CO;2
Jonard, M., Fürst, A., Verstraeten, A., Thimonier, A., Timmermann, V., Potočić, N., et al. (2015). Tree mineral nutrition is deteriorating in Europe. Glob. Chang. Biol. 21, 418–430. doi: 10.1111/gcb.12657
Jonard, M., Nicolas, M., Coomes, D. A., Caignet, I., Saenger, A., and Ponette, Q. (2017). Forest soils in France are sequestering substantial amounts of carbon. Sci. Total Environ. 574, 616–628. doi: 10.1016/j.scitotenv.2016.09.028
Kuzyakov, Y., and Domanski, G. (2000). Carbon input by plants into the soil. Review. J. Plant Nutrition Soil Sci. 163, 421–431. doi: 10.1002/1522-2624(200008)163:4<421::AID-JPLN421>3.0.CO;2-R
Ladegaard-Pedersen, P., Elberling, B., and Vesterdal, L. (2005). Soil carbon stocks, mineralization rates, and CO 2 effluxes under 10 tree species on contrasting soil types. Can. J. Forest Res. 35, 1277–1284. doi: 10.1139/x05-045
Lal, R. (2006). Enhancing crop yields in the developing countries through restoration of the soil organic carbon pool in agricultural lands. Land Degradation Dev. 17, 197–209. doi: 10.1002/ldr.696
Lal, R., Smith, P., Jungkunst, H. F., Mitsch, W. J., Lehmann, J., Nair, P. K. R., et al. (2018). The carbon sequestration potential of terrestrial ecosystems. J. Soil Water Conserv. 73, 145A−152A. doi: 10.2489/jswc.73.6.145A
Langenbruch, C., Helfrich, M., and Flessa, H. (2012). Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 352, 389–403. doi: 10.1007/s11104-011-1004-7
Leuschner, C., Wulf, M., Bäuchler, P., and Hertel, D. (2013). Soil C and nutrient stores under Scots pine afforestations compared to ancient beech forests in the German Pleistocene: The role of tree species and forest history. Forest Ecol. Manage. 310, 405–415. doi: 10.1016/j.foreco.2013.08.043
Li, D., Niu, S., and Luo, Y. (2012). Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: a meta-analysis. New Phytol. 195, 172–181. doi: 10.1111/j.1469-8137.2012.04150.x
Lorenz, M., and Thiele-Bruhn, S. (2019). Tree species affect soil organic matter stocks and stoichiometry in interaction with soil microbiota. Geoderma 353, 35–46. doi: 10.1016/j.geoderma.2019.06.021
Magh, R.-K., Grün, M., Knothe, V. E., Stubenazy, T., Tejedor, J., Dannenmann, M., et al. (2018). Silver-fir (Abies alba MILL.) neighbors improve water relations of European beech (Fagus sylvatica L.), but do not affect N nutrition. Trees 32, 337–348. doi: 10.1007/s00468-017-1557-z
Mareschal, L., Bonnaud, P., Turpault, M. P., and Ranger, J. (2010). Impact of common European tree species on the chemical and physicochemical properties of fine earth: an unusual pattern. Eur. J. Soil Sci. 61, 14–23. doi: 10.1111/j.1365-2389.2009.01206.x
Mayer, M., Prescott, C. E., Abaker, W. E. A., Augusto, L., Cécillon, L., Ferreira, G. W. D., et al. (2020). Tamm Review: influence of forest management activities on soil organic carbon stocks: a knowledge synthesis. For. Ecol. Manage. 466:118127. doi: 10.1016/j.foreco.2020.118127
Menyailo, O., and Hungate, B. A. (2003). Interactive effects of tree species and soil moisture on methane consumption. Soil Biol. Biochem. 35, 625–628. doi: 10.1016/S0038-0717(03)00018-X
Metz, J., Annighöfer, P., Schall, P., Zimmermann, J., Kahl, T., Schulze, E.-D., et al. (2016). Site-adapted admixed tree species reduce drought susceptibility of mature European beech. Glob. Chang. Biol. 22, 903–920. doi: 10.1111/gcb.13113
Mueller, K. E., Eissenstat, D. M., Hobbie, S. E., Oleksyn, J., Jagodzinski, A. M., Reich, P. B., et al. (2012). Tree species effects on coupled cycles of carbon, nitrogen, and acidity in mineral soils at a common garden experiment. Biogeochemistry 111, 601–614. doi: 10.1007/s10533-011-9695-7
Neirynck, J., Mirtcheva, S., Sioen, G., and Lust, N. (2000). Impact of Tilia platphyllos Scop., Fraxinus excelsoir L., Acer pseudoplatanus L., Quercus robor L. and Fagus sylvatica L. on earthworm biomass and physico-chamical properties of a loamy topsoil. Forest Ecol. Manage. 133, 275–286. doi: 10.1016/S0378-1127(99)00240-6
Nihlgård, B. (1971). Pedological influence of spruce planted on former beech forest soils in Scania, South Sweden. Oikos 22:302. doi: 10.2307/3543854
Nitsch, P., Kaupenjohann, M., and Wulf, M. (2018). Forest continuity, soil depth and tree species are important parameters for SOC stocks in an old forest (Templiner Buchheide, northeast Germany). Geoderma 310, 65–76. doi: 10.1016/j.geoderma.2017.08.041
Oostra, S., Majdi, H., and Olsson, M. (2006). Impact of tree species on soil carbon stocks and soil acidity in southern Sweden. Scand. J. Forest Res. 21, 364–371. doi: 10.1080/02827580600950172
Ovington, J. D. (1956). Studies of the Development of Woodland Conditions Under Different Trees: IV. The ignition loss, water, carbon and nitrogen content of the mineral soil. J. Ecol. 44:171. doi: 10.2307/2257160
Post, W., and Kwon, K. (2000). Soi carbon sequestration and land-use change: processes and potential. Glob. Chang. Biol. 6, 317–327. doi: 10.1046/j.1365-2486.2000.00308.x
Prescott, C. E., and Vesterdal, L. (2013). Tree species effects on soils in temperate and boreal forests: emerging themes and research needs. For. Ecol. Manage. 309, 1–3. doi: 10.1016/j.foreco.2013.06.042
Pretzsch, H., Schütze, G., and Uhl, E. (2013). Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation: drought stress release by inter-specific facilitation. Plant Biol. 15, 483–495. doi: 10.1111/j.1438-8677.2012.00670.x
Prietzel, J., and Bachmann, S. (2012). Changes in soil organic C and N stocks after forest transformation from Norway spruce and Scots pine into Douglas fir, Douglas fir/spruce, or European beech stands at different sites in Southern Germany. Forest Ecol. Manage. 269, 134–148. doi: 10.1016/j.foreco.2011.12.034
R Core Team (2017). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/.
Raich, J. W., and Tufekcioglu, A. (2000). Vegetation and Soil Respiration: Correlations and Controls. Biogeochemistry 48, 71–90. doi: 10.1023/A:1006112000616
Rehschuh, S., Fuchs, M., Tejedor, J., Schäfler-Schmid, A., Magh, R.-K., Burzlaff, T., et al. (2019). Admixing fir to European Beech Forests improves the soil greenhouse gas balance. Forests 10:213. doi: 10.3390/f10030213
Rennenberg, H., Loreto, F., Polle, A., Brilli, F., Fares, S., Beniwal, R. S., et al. (2006). Physiological responses of forest trees to heat and drought. Plant Biol. 8, 556–571. doi: 10.1055/s-2006-924084
Rothe, A., and Binkley, D. (2001). Nutritional interactions in mixed species forests: a synthesis. Can. J. Forest Res. 31, 1855–1870. doi: 10.1139/x01-120
Scheu, S., Albers, D., Alphei, J., Buryn, R., Klages, U., Migge, S., et al. (2003). The soil fauna community in pure and mixed stands of beech and spruce of different age: trophic structure and structuring forces. Oikos 101, 225–238. doi: 10.1034/j.1600-0706.2003.12131.x
Schleuß, P.-M., Heitkamp, F., Leuschner, C., Fender, A.-C., and Jungkunst, H. F. (2014). Higher subsoil carbon storage in species-rich than species-poor temperate forests. Environ. Res. Letters 9:014007. doi: 10.1088/1748-9326/9/1/014007
Schmidt, M., Veldkamp, E., and Corre, M. D. (2015). Tree species diversity effects on productivity, soil nutrient availability and nutrient response efficiency in a temperate deciduous forest. Forest Ecol. Manage. 338, 114–123. doi: 10.1016/j.foreco.2014.11.021
Schulp, C. J. E., Nabuurs, G.-J., Verburg, P. H., and de Waal, R. W. (2008). Effect of tree species on carbon stocks in forest floor and mineral soil and implications for soil carbon inventories. Forest Ecol. Manage. 256, 482–490. doi: 10.1016/j.foreco.2008.05.007
Tefs, C., and Gleixner, G. (2012). Importance of root derived carbon for soil organic matter storage in a temperate old-growth beech forest – Evidence from C, N and 14C content. For. Ecol. Manage. 263, 131–137. doi: 10.1016/j.foreco.2011.09.010
Trum, F., Titeux, H., Ranger, J., and Delvaux, B. (2011). Influence of tree species on carbon and nitrogen transformation patterns in forest floor profiles. Ann. For. Sci. 68, 837–847. doi: 10.1007/s13595-011-0080-4
Vesterdal, L., Clarke, N., Sigurdsson, B. D., and Gundersen, P. (2013). Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manage. 309, 4–18. doi: 10.1016/j.foreco.2013.01.017
Vesterdal, L., and Raulund-Rasmussen, K. (1998). Forest floor chemistry under seven tree species along a soil fertility gradient. Canadian J. Forest Res. 28:12. doi: 10.1139/x98-140
Vesterdal, L., Schmidt, I. K., Callesen, I., Nilsson, L. O., and Gundersen, P. (2008). Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manage. 255, 35–48. doi: 10.1016/j.foreco.2007.08.015
Wiesmeier, M., Spörlein, P., Geuß, U., Hangen, E., Haug, S., Reischl, A., et al. (2012). Soil organic carbon stocks in southeast Germany (Bavaria) as affected by land use, soil type and sampling depth. Glob. Chang. Biol. 18, 2233–2245. doi: 10.1111/j.1365-2486.2012.02699.x
Zederer, D. P., Talkner, U., Spohn, M., and Joergensen, R. G. (2017). Microbial biomass phosphorus and C/N/P stoichiometry in forest floor and A horizons as affected by tree species. Soil Biol. Biochem. 111, 166–175. doi: 10.1016/j.soilbio.2017.04.009
Keywords: tree species, mixed forests, forest management, carbon storage, review
Citation: Rehschuh S, Jonard M, Wiesmeier M, Rennenberg H and Dannenmann M (2021) Impact of European Beech Forest Diversification on Soil Organic Carbon and Total Nitrogen Stocks–A Meta-Analysis. Front. For. Glob. Change 4:606669. doi: 10.3389/ffgc.2021.606669
Received: 15 September 2020; Accepted: 05 January 2021;
Published: 10 February 2021.
Edited by:
Jérôme Laganière, Canadian Forest Service, CanadaReviewed by:
Lukas Kohl, University of Helsinki, FinlandYahya Kooch, Tarbiat Modares University, Iran
Christoph Leuschner, University of Göttingen, Germany
Copyright © 2021 Rehschuh, Jonard, Wiesmeier, Rennenberg and Dannenmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Dannenmann, bWljaGFlbC5kYW5uZW5tYW5uQGtpdC5lZHU=