 Jeff Licht
Jeff Licht Nicholas G. Smith
Nicholas G. Smith- 1School for the Environment, University of Massachusetts Boston, Boston, MA, United States
- 2Department of Biological Sciences, Texas Tech University, Lubbock, TX, United States
Climate change and land management are altering forest fire frequency and intensity worldwide. In some Northeast U.S. forests, pitch pine (Pinus rigida Miller) is not suffering from presence but rather a lack of wildfire events. In their absence, prescribed fire is being used to diminish fuel loads, open canopies and reduce competition. Pyrogenic carbon (PyC) produced by the fires may also improve soil moisture retention and plant physiological processes. Where the application of prescribed fire is not feasible due to nearby human populations, we reason prescribed fire PyC could be replaced by anthropogenic PyC product to provide similar soil benefits. We tested this hypothesis with pitch pine seedlings at a site absent overstory planted in submerged tree pots with control and PyC-imbued soils. Investigators found anthropogenic and forest PyC fostered similar growth, soil moisture retention and photosynthetic intrinsic water use efficiency, both significantly higher than unamended soils. We conclude anthropogenic subsurface PyC soil amendment provides a conservation management tool for enhancing benefits in ecosystems where prescribed fire is not a viable option in northerly forests in the U.S.
Introduction
Forest fire frequency and intensity are being altered by climate change, notably in northern latitudes (Liu et al., 2019). Pitch pine (Pinus rigida Miller) species, adapted to wildfire perturbance (Patterson and Backman, 1988; Parshall et al., 2003), face increased hardship from an absence of stand-replacing fire (Foster and Motzkin, 2003; Nowacki and Abrams, 2008) which no doubt contributes to pressure on pitch pine vigor (Patterson et al., 1983). Warmer temperatures and changes in precipitation patterns might possibly tip the balance toward greater competitiveness except for the fact that pitch pine competitors are not as dependent on open canopies (Fischer et al., 2013), are more resistant (Lesk et al., 2017) to southern pine beetle (Dendroctonus frontalis) and appear to be more adaptive and resilient to climate change (Swanston et al., 2018).
In place of forest fire, prescribed fire (Little, 1953; Neill et al., 2007) is employed to open overstories, reduce fuel availability and eliminate competition from other species. Prescribed fire pyrogenic carbon (PyC), essentially charcoal, disperses into Oa-Ab horizons (Neill et al., 2007) leaving little doubt of its indelible mark on juvenile trees. Much is known about the effects of forest fire and prescribed fire PyC (Doerr et al., 2018). Notably, prescribed fire (Foereid et al., 2015; Michelotti and Meisel, 2015) produces highly reactive and highly sorbent material that enrich forest soils (Thomas and Gale, 2015; Mitchell et al., 2016). However, less understood are anthropogenic PyC impacts on forest soil properties and tree growth (Ge et al., 2019), especially juvenile trees. Some studies focus on PyC and tree germination processes (McElligott, 2011; Ledig et al., 2015, but few if any describe post-germination impacts and none on seedling growth of a particular species pitch pine.
PyC efficacy is measured in terms of soil moisture sorption and retention (Licht et al., 2017), which influence growth potential. Forest PyC has been found to temporarily disrupt water holding capacity in Oa-Ab horizon soil pores as a function of short-term hydrophobicity (Mikita-Barbato et al., 2015). However, we suspect long-term moisture holding capacity over the length of the study increases as carbonates from anthropogenic PyC mineralize, resulting in increased total P sorption (Chintala et al., 2014) and C coalescence into pools (Preston and Schmidt, 2006). Researchers find forest soils react quickly to PyC dispersal based on substantial soil surface area (Sigmund et al., 2017)—exponentially greater than bare soil—which accommodates greater nutrient uptake and improves soil moisture retention (Licht et al., 2017). Where wildfire is no longer experienced, such as in coastal sand plain pitch pine forests (Lee et al., 2018), and prescribed fire is employed, there is a need to determine if alternative sources may be found to replicate positive impacts of prescribed fire PyC. One prominent benefit is ecophysiological; it results from nutrient mobilization and moisture retention. As an increase in photosynthetic intrinsic water use efficiency or iWUE (Renninger et al., 2013; Carlo et al., 2016), iWUE is derived from stable isotope analysis (stated as per mils: parts per thousand, notated as (°/oo) and quantifies the efficiency of carbon capture to water loss at the leaf level (Farquhar et al., 1989; Lajtha and Michener, 1994).
Where forest fire is absent and prescribed fire is unavailable for use, mechanical thinning is an option which fosters open canopies and reduces forest floor or grassland fuel (Anderson, 2017). This bioturbation method (Abney et al., 2019) is akin to other types of disturbance (e.g., animal, human, or climate pressures). Thinning is found to generate temporary nutrient turnover following soil disturbance (Saursaunet et al., 2018) without the associated risks of fire close to human settlement. We conduct an analysis which determines it is a much more expensive intervention ($3,000 US/0.404 h) than prescribed fire ($300 US/0.404 h), but may be used on a limited basis (cost estimates based on a report by US Fish and Wildlife Service, Hadley MA, USA, 2013). Indeed, tree removal and soil disturbance may be as, or more, effective at controlling pitch pine populations than prescribed fire (Lee et al., 2018).
In this paper, we examine prescribed fire and anthropogenic PyC impact on subsurface water and nutrient supply changes to pitch pine growth and iWUE. Our aim is to take PyC results and frame a “package” of anthropogenic PyC conservation potential for small, but critically and vitally important pitch pine populations where prescriptive fire is not an option in the Northeast U.S.
Materials and Methods
Study and Extraction Sites
A study site (42°39'18.23 “N 71°35'01.34”W) in Middlesex County, MA was used; its environmental and meteorological characteristics are similar to those of a pitch pine community 14.5 km southeast from the forest (42°22'51.57“N 71°28'22.60”W) where a prescribed fire was carried out on site in the same county (Figure 1). The forest site elevation was 82 m; the experimental site elevation was 90 m.
Figure 1. Map of locations of extraction and experimental field sites in eastern MA, USA; *Boston, MA, the state capital.
Materials
Soil extractions from the forest were taken from the “O-A” horizon at 20 cm depth at random locations in burned and unburned portions of the 20.23 h forest using a narrow shovel. Untreated (absent charcoal) soils were characterized as Histosol (85%), Freetown (8%), and Scarboro (7%) according to NRCS SoilWeb. The dehydrated bulk density was +/– 2 g cm−3, estimated using the Kettler et al. (2001) rapid HMP method with 3% aqueous solution and 0.053 mm sieve to collect the sand fraction and settled silty fraction following decantation of clay followed by calculations of different fractions (clay = 3.4%, silt = 29.8%, and sand = 66.8%). Three soil treatments were created using (1) forest soil absent forest charcoal, (2) forest soil blended (9–11% v/v) with forest charcoal PyC and (3) forest soil absent charcoal blended (10% v/v) with anthropogenic PyC.
Forest charcoal PyC consists of charcoal particulate extracted from a shallow (<12 cm) burned duff and screened (<10 mm) for blending as treatment F. To determine pyrolysis temperature of the forest charcoal we conducted a fire event simulation with several pitch pine seedlings at an outdoor experiment site. Using a K-type probe (4.8 mm in diameter and 30 cm long) and data logger (EXTECH EA11) we measured a high temperature parameter where the probe was attached 88 cm above the tree base. A mean pyrolysis maximum temperature (Tmax) between 409 and 415°C was recorded 9–12 s after exposure of the bark to a steady flame. We ascertained these findings were well below crown recordings (Tmax = 650–700°C. reported by A. Richardson, Harvard Forest) and ground layer (surface; Tmax = 550–600°C reported from an experimental site in the New Jersey pine barrens, Foereid et al., 2015).
Retort PyC (Mitchell et al., 2016; Licht et al., 2017) was produced in an Adam-style retort (Sullivan Farm, Sullivan, New Hampshire USA). To gauge production temperature, a built-in retort probe and data logger continuously recorded temperature ever 6 h (reported in Licht et al., 2017) achieving Tmax in the 590–615°C range. The Tmax results were consistent with experiments described by others (Mitchell et al., 2013; Ghorbel et al., 2015). Retort PyC was also pulverized in a small blender and screened to <10 mm particles.
Experiments
At the field site, twenty-four 32 cm depth planting holes were excavated one meter apart under an open canopy. Excavated field soil was set aside. Three soil treatments were created comprising treatments labeled: no treatment = NT, soil plus forest PyC = PyCF and soil plus anthropogenic PyC = PyCA. Eight 2.5 L pots (9.6 cm wide ×30 cm depth; CP413CH Steuwe and Sons Inc., Tangent OR USA) were filled with either 1,420 mL NT, 1,280 mL NT, and 142 mL PyCF and 1,280 mL NT and 142 mL PyCA. Thus, there were 8 pots for each treatment, a total of 24 (N = 24) unpooled replicates. Each pot was planted with a single 3 year pitch pine seedling (Pinelands Nursery, Princeton, NJ) and then plunged in planting holes. Seedlings were irrigated immediately after planting and left to acclimate (to stabilize according to moisture, nutrient, CEC, pH, etc.). No further irrigation was furnished during the balance of the experiments. Daytime temperatures averaged 21.66°C during the mid-day of the growing season. Fall-winter-early spring temperatures ranged from 15°C down to −13.3°C. Surface winds were variable year-round, averaging ≤ 7 km h−1. Year-round humidity averaged 56%. Average annual precipitation was 113.69 cm. As described below, soil, needle samples, and growth measurements were obtained on an annual basis for 3 years.
Field Measurements
Plant Height, Stem Diameter, and Root Length
In the fall of each year, three individuals in each treatment group were randomly selected for tree biometrics. Seedling height was measured from ground level. Primary stem diameter at 5 cm height was measured using a ProSkit electronic digital caliper (Amelia, VA, USA). Root length (along a coarse to fine gradient) was also measured. To accomplish this measurement, trees were carefully excavated and then reinstalled in each planting hole.
Laboratory Measurements
Gravimetric Moisture Retention
We performed a dry down-to-moisten up bench test resulting in a post-hoc analysis of extracted soil sample moisture retention. An extra set of three pots per treatment per year pots were placed in planting holes which allowed us to extract aliquots that endured the same mineralization over 3 years—after measurement the soils were returned. Identical volume samples (175 mL) were extracted and dried at 100°C for 2 days, then placed in a 300 mL plastic cup to which 70 mL of H2O was added. These were measured for gravimetric moisture retention on a wet basis utilizing the Shields method to conduct these experiments as in Licht et al. (2017). Net moisture retention as a subset of soil moisture evaporation (ψg) was determined over a 10 week period as net evaporative loss or adsorption to surfaces (Qi et al., 2018).
Carbon and Nitrogen Stable Isotopes, Elemental Analysis
Soil δ13C (isotopic) measurements were acquired for five randomly selected individuals per treatment per year following extraction of three soil types each year in early summer, over a span of 3 years. Leaf δ13C (isotopic) measurements were also obtained on fully expanded leaves (needle cluster) in 2015, 2016, and 2017 at peak PAR. Sample fascicles were separated and dried for 2 days at 60°C and then ground in a SPEX ball mill (Metuchen, NJ, USA). Ground samples were fed to a Perkins Elmer Elemental Analyzer ECS 4100 (Waltham, MA, USA) coupled with a Thermo Delta (Waltham, MA, USA) V+ IR-MS continuous flow isotope ratio mass spectrometer with a universal triple collector. Combustion gasses were separated on a gas chromatograph column, passed through a diluter and reference gas box, and introduced into the spectrometer. Measured δ13C (Farquhar et al., 1989) was used as a proxy for photosynthetic intrinsic water use efficiency (iWUE).
Foliar Nutrient
Fifty milliliter needle samples of five randomly selected individuals per treatment per year were extracted from seedlings and dried for 2 days at 60°C. Then they were ground in a SPEX ball mill (Metuchen, NJ, USA), sieved to <10 mm, and submitted for standard plant tissue nutrient analysis using a TJA Model 975 AtomComp ICP-AES (Thermo Jarrell-Ash Corp., Franklin, MA) and Leco CN-2000 Carbon-Nitrogen Analyzer (Leco Corp., St. Joseph, MI). A standard 5 mL trace-metal-grade HNO3 treatment was employed, then refluxed on a hotblock at 80°C for 2 h, diluted to 25 mL with 0.45 micron PTFE syringe filters and acidified using 5% HNO3. Analysis was focused on inorganic fractions (Ca, P, K, Mg, Al, and Zn). Foliar C and N were calculated from elemental analysis.
Soil Nutrient
Repeated tests of 240 mL volume samples of untreated and treated soils were subjected to Modified Morgan digestive analysis using inductively coupled plasma spectroscopy (ICP) to measure micronutrient concentrations, proton activity of a 1:1 slurry to measure pH. The sum of extractable K+, Ca2+, Mg2+, and Na+ were used to gauge soil cation exchange capacity (CEC) measured using Formic acid (HCO2H). Repeated analyses of 240 mL volume samples were performed to determine inorganic and organic chemistry of pyrolysis charcoals. Tests were conducted using Modified Morgan digestive analysis following the procedure of Licht et al. (2017). Due to low sample size, these data were not analyzed using the statistical methods indicated below, but results indicate that soil Ca, P, Mg, K, and Zn all tended to increase following PyC amendments (Table 4). Soil C and N were calculated from elemental analysis.
Statistical Analysis
We used analysis of variance (ANOVA) to examine the effect of soil type (after modification: PyCF and PyCA), year, soil type x year on plant growth (height, stem diameter, and root length), soil water retention, and leaf foliar chemistry (% C, % N, C/N molar ratio, δ13C, δ15N). The leaf C/N was log transformed before model fitting to meet normality assumptions. Soil water retention was arcsin square root transformed because it was percentage data. As foliar chemistry measurements were made on the same individual trees across multiple years, we used a repeated measures design using the “lme4” package in R (Bates et al., 2015). We used F-tests and χ2 tests to test for significance in the non-repeated and repeated measure models, respectively. The F-tests were performed using the “anova” function in base R and the χ2 tests were performed using the “Anova” function from the “car” package in R (Fox and Weisberg, 2011). Tukey's HSD was used for post hoc comparison using least square means using the “emmeans” package in R (Lenth, 2019).
Results
Soil treatments PyCF and PyCA increased height in the final 2 years of treatment (soil type by year: P = 0.001; Figure 2 and Table 1). There was a nearly 50% difference in vertical growth favoring trees subject to PyC amendment compared to controls (NT) over 3 years. This was confirmed by significant post-hoc Tukey's tests for treatment effects in years 2 and 3 (P < 0.05 in both cases), but not year 1 (P > 0.05). Heights did not differ between PyCA and PyCF treatments in any year (Tukey's test: P > 0.05). A significant treatment by year interaction (P = 0.018; Figure 2 and Table 1) indicated that PyCF and PyCA increased root length in the second (Tukey's test: P < 0.05) and third (Tukey's test: P < 0.05), but not the first (Tukey's test: P > 0.05) year of the experiment. In all there was a 125% increase due to PyC involvement over the 3 years compared to controls. There was no difference in root length between PyCF and PyCA treatments in any year (Tukey's test: P > 0.05). Stem diameter was unaffected by the treatments in any single year (P > 0.05; Figure 2 and Table 1), but did differ by year (P < 0.001).
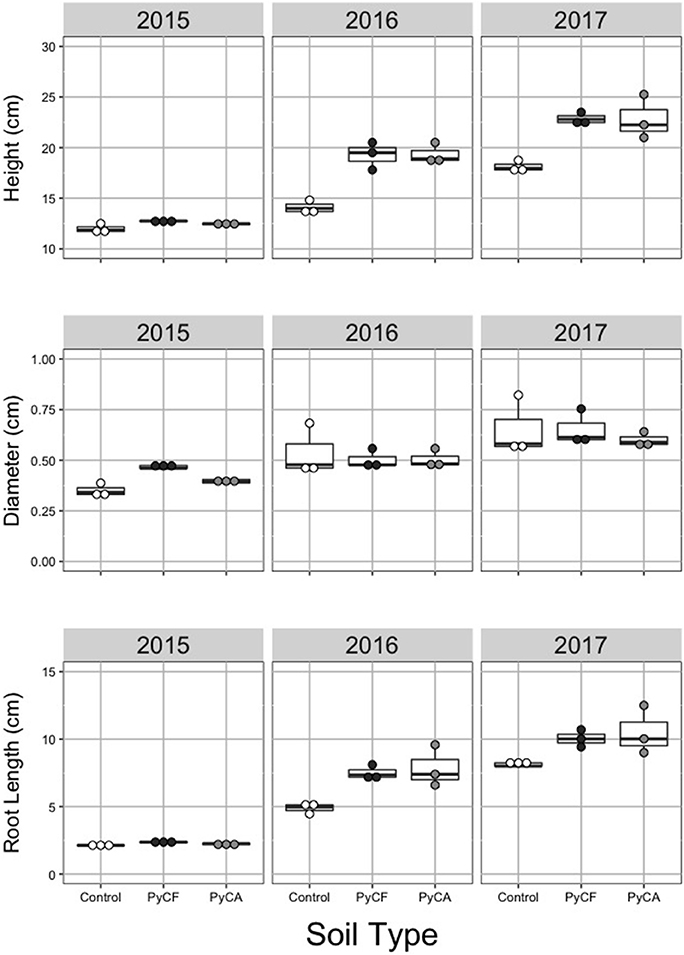
Figure 2. Dot and box plots of plant height (top), diameter (middle), and root length (bottom) for soil-less media controls (white dots) and media amended with forest (PyCF; dark gray dots) and recycled feedstock (PyCA; light gray dots) pyrogenic carbon in 2015, 2016, and 2017. Statistical results can be found in Table 1.
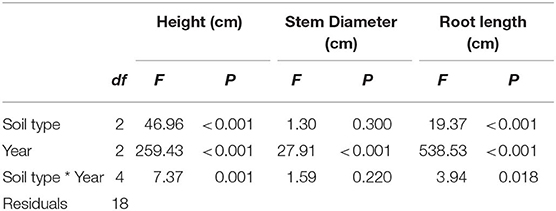
Table 1. Results from analysis of variance (ANOVA) for plant height, stem diameter and root length.
There was a significant interaction between year and treatment on soil moisture retention capacity (P = 0.004; Figure 3 and Table 2). Post-hoc Tukey's tests indicated that retention was significantly higher in the PyC amended soils as compared to the unamended soils in all years (P < 0.05 in all cases). In the first year, soil moisture retention was higher in PyCF than PyCA (Tukey's test: P < 0.05), but PyCF and PyCA had the same retention in the second and third year (Tukey's test: P < 0.05 in both cases).
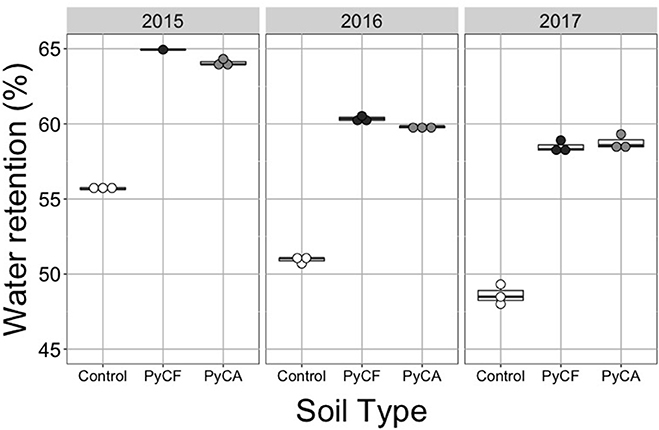
Figure 3. Dot and box plots of soil moisture retention for soil-less media controls (white dots) and media amended with forest (PyCF; dark gray dots) and recycled feedstock (PyCA; light gray dots) pyrogenic carbon in 2015, 2016, and 2017. Statistical results can be found in Table 2.
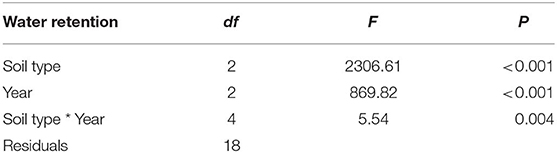
Table 2. Results from analysis of variance (ANOVA) for soil water retention.
Foliar C/N differed by year (P = 0.024), but was unaffected by soil treatment (P > 0.05) and the soil type by year interaction (P > 0.05; Figure 4 and Table 3). There was a significant interaction between year and soil type for % C (P <0.027, Table 3), but post hoc Tukey's tests did not indicate a soil treatment effect in any of the 3 years (P > 0.05 in all cases). There were no significant year or soil type effects for % N (P> 0.05 in all cases; Table 3).
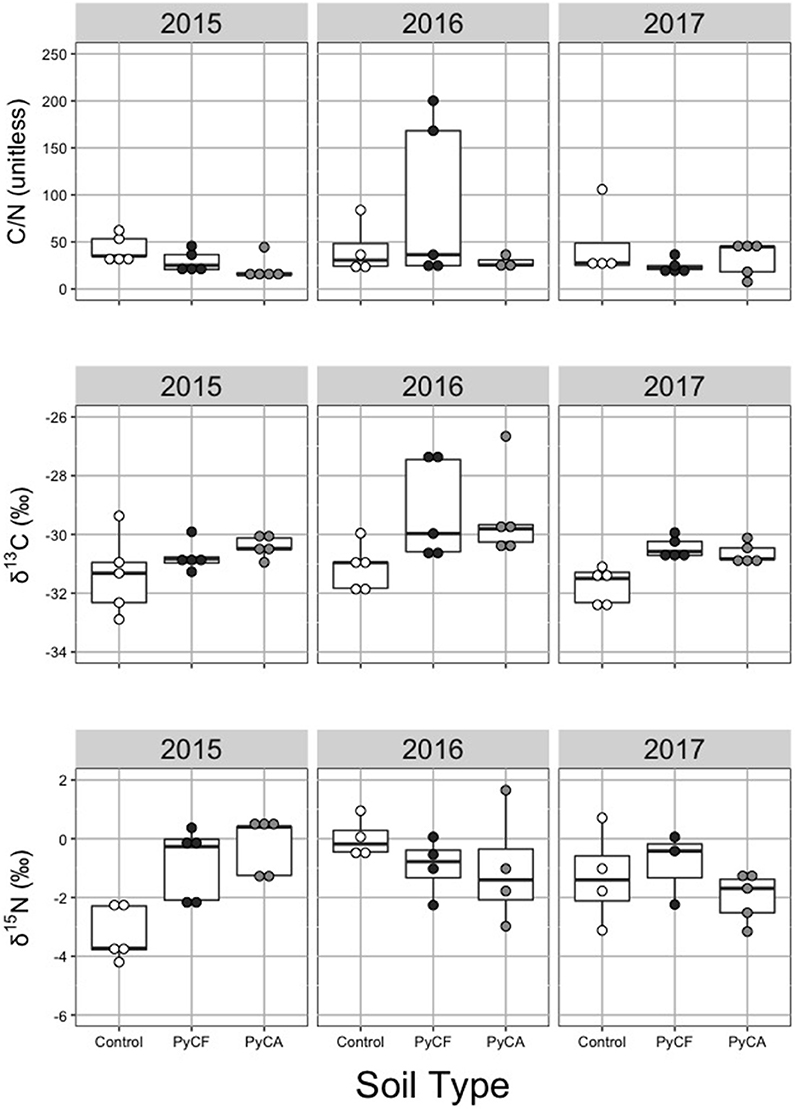
Figure 4. Dot and box plots of leaf carbon to nitrogen ratios (C/N; top), 13C discrimination (δ13C; middle), and 15N discrimination (δ15N; bottom) for soil-less media controls (white dots) and media amended with forest (PyCF; dark gray dots) and recycled feedstock (PyCA; light gray dots) pyrogenic carbon in 2015, 2016, and 2017. Statistical results can be found in Table 3.
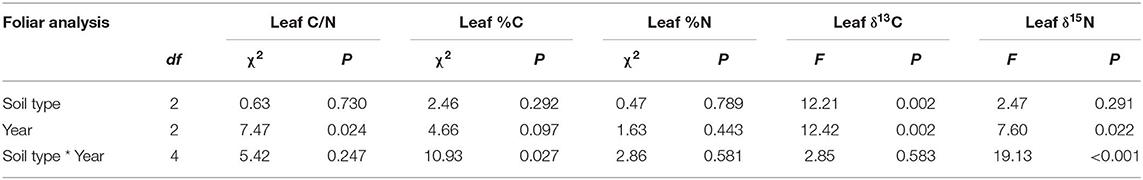
Table 3. Results from the mixed model analysis of variance for leaf carbon to nitrogen ratios (C/N), 13C discrimination (δ13C), and 15N discrimination (δ15N).
We found PyCF and PyCA treatments significantly increased foliar δ13C across all 3 years (P = 0.002, Figure 4 and Table 3). Results indicated increased photosynthetic iWUE (less negative δ13C) in those treatments compared to the NT treatment. Tukey's test found no significant difference between PyCF and PyCA treatment (P > 0.05). There were no significant differences in δ15N across any treatment.
Due to a limited sample size (N = 3), soil and needle mineral data were not analyzed using the statistical methods indicated above, but mean and SD values are reported. Results indicate soil and foliar Ca, P, Mg, K, and Zn tended to increase following PyC amendments (Table 4).
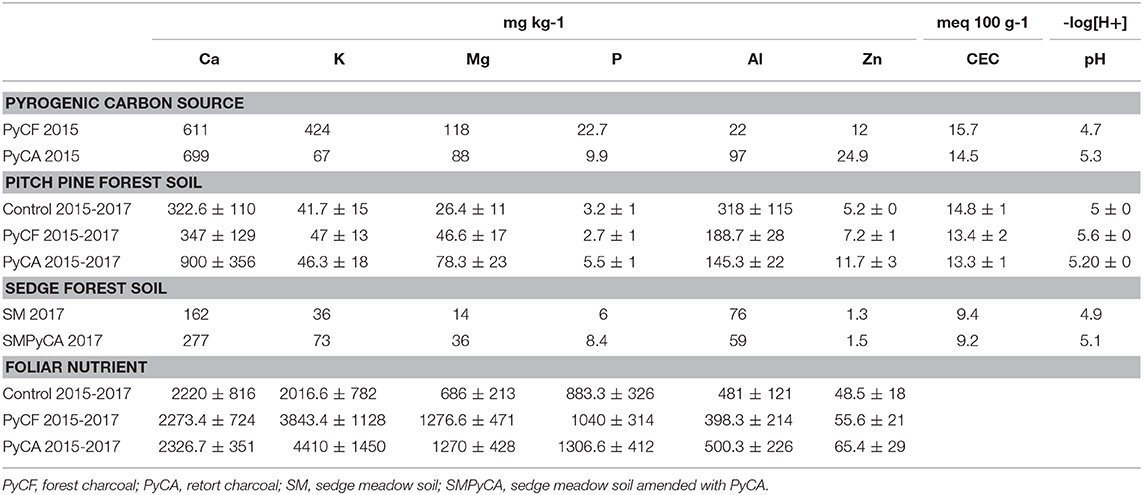
Table 4. PyC, Pitch pine forest soil, Sedgemeadow forest soil, and Foliar Nutrient data were extracted, analyzed, and reported between 2015 and 2017 using mean ± SD.
Discussion
Our study provides what we understand is the first comprehensive analysis of dual PyC effects on soil moisture, pitch pine seedling growth and iWUE as a function of subsurface implementation using a planting hole technique. In all, we examined root depth, seedling height, stem diameter, soil moisture retention, nutrient sorption and photosynthetic intrinsic water use efficiency at a site free of overstory and controlled resource competition. This investigation considers effectiveness of PyC intervention as the first step toward side-by-side planting hole PyC tests of pitch pine and their locust, spruce, white pine and red pine competitors. In doing so, we aspire to establish a theoretical basis for testing transferability of this “technology” to other species which may face decline owing to absence of fire or prescribed fire. We speculate future experiments could involve jack pine (Pinus banksiana) with which pitch pine sometimes overlap sympatrically in the Great Lakes region, Maine and Canada.
The scope of this investigation is limited to seedlings, not regeneration (Lee et al., 2018) or germination studies but at termination of the study we obtained results which allowed us to speculate as to what will transpire when these 6 year-old trees are more mature (say 10 to 15 years old). A previous model (Meigs et al., 2011) provides direction as to how one might scale up PyC application in mixed forest tests. We considered the cost for an application in a predominantly pitch pine forest. For example, for a 0.404 h area planted with 400 pitch pine seedlings (on 3 m centers) the authors assume an anthropogenic PyC production cost of approximately $105 (production and shipping)—about a third the cost of prescribed fire activity in the same space (based on estimates derived from International Biochar Initiative, Sonoma, CA, USA). A follow-on investigation of PyC influence on pitch pine germination could contrast broadcast methods (suprasurface vs. subsurface dispersal into Oa-Ab horizons) as well as the extent to which germination numbers increase in masticated or unmasticated soils (subject to treatment conditions).
We did not predict a soil moisture gradient change over 3 years; those results were consistent with previously reported findings that moisture retention stabilizes during pyromineral-organic decomposition (Licht et al., 2017). We reason part of the rationale for this premise rests on assumptions that sandy and perhaps gravelly soils found along the Eastern U.S. coast treated with PyC produce a similar reaction in carbonate sorption properties leading to a concomitant reduction in hydrophobicity (Alcañiz et al., 2018). As a function of post-settlement stabilization (irrespective of PyC source), we determine increased solubility of remaining carbonates leads eventually to uptake of other minerals as reported earlier (Licht et al., 2017). Questions still arise however as to PyC influence on disruptions in moisture or nutrients (Gaskin et al., 2007) tied to over-resistance to thermal and microbial decomposition (Spokas, 2010). It is accepted by those studying PyC effects that C decomposition and disaggregation is spurred by P changes occurring well after initial soil incubation. Likewise, when molar ratios change, C and N pools may combine to defeat anisohydric, or drought stress, effects (Parker et al., 2001). Part of this response likely stems from greater C and N volatization (Doerr et al., 2018) from forest fire or prescribed fire more than anthropogenic charcoals; we contemplate this may be explained by differences in temperature and pyrolytic duration. Previous investigation suggests foliar P is found in juvenile foliar tissue following forest fire (Schaffers, 2002). In our study, we found increases in foliar P in seedlings exposed specifically to PyCA. Another potential difference could be ascribed to a greater overabundance of N2 (Aber et al., 1989) which is not consumed in retort pyrolysis vs. fire pyrolysis—however all of these speculations lack experimental confirmation. Unfortunately, due to a limitation in mineral loss/gain findings, a congruent set of implications for PyC mechanics eluded us.
Past research examined production (Tmax) temperature (which affects condensation of volatile compounds) and air temperature, but did not report oxygen availability (Santin et al., 2017), which might explain differences in sorption properties (Jindo et al., 2012). If future research determines differences between feedstocks are not a major hurdle to building a plausible methodology, this would further indemnify investment in research of PyC soil management in forest ecology and conservation. Despite these upbeat assumptions, there are admittedly obstacles to implementation based on dissimilarities in PyC feedstock and production methods on the one hand and differences between anthropogenic and forest PyC effects in a given ecosystem. Allowing for the enigmatic nature of char feedstock characteristics, we were presented with another impediment to evolving a lucid, linear connection between feedstock selection, production, post-installation effects and mechanics (Mia et al., 2017).
It was easier to assign an ecophysiological value to PyC amendment. We found a more modest increase (about 12%) in iWUE fostered by PyCF or PyCA treatments than other investigators (Carlo et al., 2016), who reported a 22% increase in pitch pine photosynthetic iWUE based on PyCF alone. There are bound to be anomalies which arise between PyC analytes for a considerable number of reasons. However, if we extrapolate from the Carlo study, we may intuit higher iWUE (lower negative) resulted from greater soil moisture holding capacity or a considerable difference in tree maturity, as just two explanations. Our dual iWUE results were similar to those in another study (Renninger et al., 2013) which supports the concept that both PyCs we studied yield similar influence on tree physiology. Unfortunately, due to cost constraints, we were unable to add more replicates to this aspect of the research; however the statistical analyses used are robust to low sample size.
Reports of pitch pine response following mechanical thinning indicate scarification increases seedling recovery as a function of soil mineral turnover (North et al., 2015; Lee et al., 2018). Thus, a future key matchup between forest and anthropogenic PyC sorption of soil nutrients should comprise a comparison of disturbance (unrelated to PyC) vs. non-disturbance. It should be acknowledged that although mineral deposits are exposed following mechanical disturbance, one caviat to the thinning method is an associated problem with soil compaction (Zisa et al., 1980).
Some research has shown a relation between forest charcoal-imbued soil and ectomycorrhizal response (Warnock et al., 2007). Early findings (Hobbie et al., 1999) suggest fire effects sway pitch pine root fungi aggregation. The function of soil microbes (Nannipieri et al., 2003), in relation to pitch pine hosts, is discussed according to N-fixing bacteria found inside the fungi mantle (Güerena et al., 2013). In the case of prescribed fire, fungal responses to repeated high burn temperature and post-burn hydrophobia (Williams and Jackson, 2007) also add to what is known about mycorrhizae presence. Indeed, advances in the study of connections between prescribed fire and fungi continue (Luo et al., 2017) along with a search for genetic similarities between disjunct pine populations.
Whether or not fire proves to be a long-term solution to southern pine beetle persistence (Lesk et al., 2017), it would be very useful to identify whether forest or prescribed fire play a decisive role in slowing acceleration of blue stain fungi aggregation (Ophiostoma minus) associated with southern pine beetle (Arango-Velez et al., 2018). Following such a study, if results warranted, an analysis of anthropogenic PyC inputs might also be advantageous. Up until the past decade, northward southern pine beetle migration was limited to New Jersey, Long Island NY, Hudson River Valley, and Southeastern MA. Now warming winter temperatures, coupled with the loss of mite predators, forecast greater southern pine beetle aggregation in pitch pine populations in upper New York State and the Northern tier of New England (Lesk et al., 2017). It could be PyC plays a role in plant defense against southern pine beetle given its contributions to higher resistance to drought (Licht and Smith, 2018) but there is no evidence yet that PyC reduces pathogens (Nowak et al., 2015).
Recent climate models anticipate effects of climate change on a host of coastal tree species (Fernandez et al., 2015). Nevertheless, given a paucity of knowledge about PyC mechanics, it is difficult to ascertain the extent to which PyC may be an underutilized resource in the quest for resolving environmental problems and anthropogenic perturbations. It does appear, in the case of pitch pine, at least, PyC elevates productivity where stress is minimal or curbs stress in preference for adding biomass.
Population density is a limitation on prescribed fire if not an antecedent to use of anthropogenic PyC. In New England many fires are set purposefully to reduce forest fuel compared to New York or New Jersey, but total acres consumed by this process are double the rate for states further north (Melvin, 2012). In the Southeast (Chiodi et al., 2018) and Southwest US (Neary et al., 2017), many millions of acres not burned by forest fires require controlled burn management. In these regions exhibiting drier climates than those in the Northeast, forest fire and prescribed fire combine, not just to reduce competition from other tree species, but to combat pest aggression, either southern pine beetle or western pine beetle (Dendroctonus brevicomis LeConte), aided by selective “spot” cutting. Rather than pests or disease, ongoing concerns about the destructiveness of forest fire in Australia (McCaw, 2013), in Spain grasslands (Úbeda et al., 2005) and in other parts of Europe (Lázaro and Montiel, 2010) grow even as efforts continue to employ prescribed fire where human population is less dense.
In this investigation we enterprise preliminary evidence for anthropogenic PyC use based on transformation of lignocellulosic and hemicellulosic wastes which could be achieved economically in vast areas of the world. We intend for our data to signal the potential for novel soil amendment, inclusive of waste conversion, to fit within an armamentarium of tools to resolve conservation and land use management concerns associated with climate warming and irregular storm events.
Data Availability Statement
All data used in this article can be found at: https://github.com/SmithEcophysLab/pitchpine_pyc/releases/tag/v1.0 (doi: 10.5281/zenodo.3600547).
Author Contributions
JL and NS conceived the work, contributed substantially to the interpretation of the data and to drafting the manuscript, gave final approval of the version submitted, and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. JL carried out the field experimentation, conducted preparation for all tests analyzed at laboratories at Boston University, University of Massachusetts, and University of Maine. NS was responsible for conducting statistical analyses and formulating figures and tables.
Funding
NS acknowledges financial support from Texas Tech University.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Experimental anthropogenic PyC was produced at the Sullivan Agricultural Learning Center under the supervision of Steve and Rick Sullivan in Sullivan NH USA.
References
Aber, J., Nadelhoffer, K., Steudler, P., and Melillo, J. (1989). Nitrogen saturation in northern forest ecosystems: excess nitrogen from fossil fuel combustion may stress the biosphere. Bioscience 39, 378–386. doi: 10.2307/1311067
Abney, R., Kuhn, T., Chow, A., Hockaday, W., Fogel, M., and Berhe, A. (2019). Pyrogenic carbon erosion after the rim fire, yosemite national park: the role of burn severity and slope. JGR Biogeosciences 124, 432–449. doi: 10.1029/2018JG004787
Alcañiz, M., Outeiro, L., Francos, M., and Úbeda, X. (2018). Effects of prescribed fires on soil properties: a review. Sci. Total Environ. 613–614, 944–957. doi: 10.1016/j.scitotenv.2017.09.144
Anderson, H. (2017). Ponderosa pine responses to biochar, fertilizer, or mastication on the bitterroot national forest, USA (Graduate student theses, dissertations, and Professional papers), Missoula, MT, United States.
Arango-Velez, A., Chakraborty, S., Blascyk, K., Phan, M., Barsky, J., and El Kayal, W. (2018). Anatomical and chemical responses of eastern white pine (pinus strobus L.) to blue-stain (ophiostoma minus) inoculation. Forests 9:690. doi: 10.3390/f9110690
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Carlo, N., Renninger, H., Clark, K., and Schäfer, K. (2016). Impacts of prescribed fire on Pinus Rigida mill. in upland forests of the Atlantic coastal plain. Tree Phys. 36, 967–982. doi: 10.1093/treephys/tpw044
Chintala, R., Schumacher, T., McDonald, L., Clay, D., Malo, D., Papiernik, S., et al. (2014). Phosphorus sorption and availability from biochars and soil/biochar mixtures. Clean Soil Air Water 42, 626–634. doi: 10.1002/clen.201300089
Chiodi, A. M., Larkin, N. S., and Varner, J. M. (2018). An analysis of Southeastern US prescribed burn weather windows: seasonal variability and El Niño associations. Int. J. Wildland Fire 27, 176–189. doi: 10.1071/WF17132
Doerr, S., Santin, C., Merino, A., Belcher, C., and Baxter, G. (2018). Fire as a removal mechanism of pyrogenic carbon from the environment: effects of fire and pyrogenic carbon characteristics. Front. Earth Sci. 6:127. doi: 10.3389/feart.2018.00127
Farquhar, G., Ehleringer, J., and Hubick, K. (1989). Carbon isotope discrimination and photosynthesis. Ann. Rev. Plant Phys. Plant Mol. Biol. 40, 503–537. doi: 10.1146/annurev.pp.40.060189.002443
Fernandez, I., Schmitt, C., Stancioff, E., Birkel, S., Pershing, A., Runge, J., et al. (2015). Maine's Climate Future: 2015 Update. Orono, ME: University of Maine
Fischer, A., Marshall, P., and Camp, A. (2013). Disturbances in deciduous temperate forest ecosystems of the northern hemisphere: their effects on both recent and future forest development. Biodivers. Conserv. 22, 1863–1893. doi: 10.1007/s10531-013-0525-1
Foereid, B., Lehmann, J., Wurster, C., and Bird, M. (2015). Presence of black carbon in soil due to forest fire in the New Jersey pine barrens. J. Earth Sci. Eng. 5, 91–97. doi: 10.17265/2159-581X/2015.02.001
Foster, D., and Motzkin, G. (2003). Interpreting and conserving the openland habitat of coastal New England: insights from landscape history. For. Ecol. Manag. 185, 127–150. doi: 10.1016/S0378-1127(03)00251-2
Fox, J., and Weisberg, S. (2011). An R Companion to Applied Regression, 2nd Edn. Thousand Oaks, CA: Sage Publications, Inc.
Gaskin, J., Speir, A., Morris, L., Ogden, K., Harris, K., Lee, D., et al. (2007). “Potential for pyrolysis char to affect soil moisture and nutrient retention status of a loamy sand soil,” in Proceedings of the Georgia Water Resources Conference (Athens, GA: Institute of Ecology, University of Georgia). Available online at: http://hdl.handle.net/1853/48168
Ge, X., Yang, Z., Zhou, B., Cao, Y., Xiao, W., Wang, X., et al. (2019). Biochar fertilization significantly increases nutrient levels in plants and soil but has no effect on biomass of pinus massoniana (lamb.) and cunninghamia lanceolata (lamb.) hook saplings during the first growing season. Forests 10, 612–629. doi: 10.3390/f10080612
Ghorbel, L., Rouissi, T., Brar, S., Lopez-Gonzalez, D., Ramirez, A., and Godbout, S. (2015). Value added performance of processed cardboard and farm breeding compost by pyrolysis. Waste Manag. 38, 164–173. doi: 10.1016/j.wasman.2015.01.009
Güerena, D., Lehmann, J., Hanley, K., Enders, A., Hyland, C., and Riha, S. (2013). Nitrogen dynamics following field application of biochar in a temperate North America maize-based production system. Plant Soil 365, 239–254. doi: 10.1007/s11104-012-1383-4
Hobbie, E., Macko, S., and Shugart, H. (1999). Insights into nitrogen and carbon dynamics of ectomycorrhizal and saprotrophic fungi from isotopic evidence. Oecologia 118, 353–360. doi: 10.1007/s004420050736
Jindo, K., Suto, K., Matsumoto, K., Garcia, C., Sonoki, T., and Sanchez-Monedero, M. (2012). Chemical and biochemical characterization of biochar-blended composts prepared for poultry manure. Biores. Technol. 110, 396–404. doi: 10.1016/j.biortech.2012.01.120
Kettler, T., Doran, J., and Gilbert, T. (2001). Simplified method for soil particle-size determination to accompany soil-quality analyses. Soil Sci. Soc. Am. 65, 849–852. doi: 10.2136/sssaj2001.653849x
Lajtha, K., and Michener, R. (1994). Stable Isotopes in Ecology and Environmental Science. Hoboken, NJ: Blackwell Scientific Publications.
Lázaro, A., and Montiel, C. (2010). “Overview of prescribed burning policies and practices in Europe and other countries,” in Towards Integrated Fire Management – Outcomes of the European Project Fire Paradox, eds J. Sande Silva, F. Rego, P. Fernandes, and E. Rigolot (Joensuu: European Forest Institute Research Report), 137–150.
Ledig, F., Smouse, P., and Hom, J. (2015). Postglacial migration and adaptation for dispersal in pitch pine (Pinaceae). Am. J. Bot. 102, 2074–2091. doi: 10.3732/ajb.1500009
Lee, C., Robinson, G., Robinson, I., and Lee, H. (2018). Regeneration of pitch pine (Pinus rigida) stands inhibited by fire suppression in Albany pine bush preserve, New York. J. For. Res. 30, 233–242. doi: 10.1007/s11676-018-0644-3
Lenth, R. (2019). emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online at: https://CRAN.R-project.org/package=emmeans
Lesk, C., Coffel, E., D'Amato, A. W., Dodds, K., and Horton, R. (2017). Threats to North American forests from southern pine beetle with warming winters. Nat. Clim. Change 7, 713–717. doi: 10.1038/nclimate3375
Licht, J., and Smith, N. (2018). Influence of lignocellulose and hemicellulose PyC on photosynthesis and water use efficiency in seedlings of a Northeast U.S. pine-oak ecosystem. J. Sust. For. 37, 25–37. doi: 10.1080/10549811.2017.1386113
Licht, J., Smith, N., Mitchell, P., and Shields, F. (2017). Impact of lignocellulose and hemicellulose PyCs on soil moisture in low clay soils. J. Plant Nutrit. Soil Sci. 180, 576–584. doi: 10.1002/jpln.201700221
Little, S. Jr. (1953). Prescribed burning as a tool of forest management in the northeastern states. J. For. 51, 496–500. doi: 10.1093/jof/51.7.496
Liu, Z., Ballantyne, A. P., and Cooper, L. A. (2019). Biophysical feedback of global forest fires on surface temperature. Nat. Commun. 10:214. doi: 10.1038/s41467-018-08237-z
Luo, J., Walsh, E., Miller, S., Blystone, D., Dighton, J., and Zhang, N. (2017). Root endophytic fungal communities associated with pitch pine, switchgrass, and rosette grass in the pine barrens ecosystem. Fung. Biol. 121, 478–487. doi: 10.1016/j.funbio.2017.01.005
McCaw, W. L. (2013). Managing forest fuels using prescribed fire–a perspective from southern Australia. For. Ecol. Manag. 294, 217–224. doi: 10.1016/j.foreco.2012.09.012
McElligott, K. (2011). Biochar amendments to forest soils: effects on soil properties and tree growth (Doctoral dissertation). Moscow, ID: University of Idaho.
Meigs, G., Turner, D., Ritts, W., Yang, Z., and Law, B. (2011). Landscape-scale simulation of heterogeneous fire effects on pyrogenic carbon emissions, tree mortality, and net ecosystem production. Ecosystems 14, 758–775. doi: 10.1007/s10021-011-9444-8
Melvin, M. (2012). National Prescribed Fire Use Survey Report. Retrieved from: http://www.stateforesters.org/sites/default/files/publication-documents/2012_National_Prescribed_Fire_Survey.pdf (accessed December 30, 2019).
Mia, S., Singh, B., and Dijkstra, F. (2017). Aged biochar affects gross nitrogen mineralization and recovery: a 15N study in two contrasting soils. GCB Bioenergy 9, 1196–1206. doi: 10.1111/gcbb.12430
Michelotti, L., and Meisel, J. (2015). Source material and concentration of wildfire-produced pyrogenic carbon influence post-fire soil nutrient dynamics. Forests 6, 1325–1342. doi: 10.3390/f6041325
Mikita-Barbato, R., Kelly, J., and Tate, I. I. I. R. (2015). Wildfire effects on the properties and microbial community structure of organic horizon soils in the New Jersey Pinelands. Soil Biol. Biochem. 86, 67–76. doi: 10.1016/j.soilbio.2015.03.021
Mitchell, P., Dalley, T., and Helleur, R. (2013). Preliminary laboratory production and characterization of biochars from lignocellulosic waste. J. Anal. App. Pyrol. 71–78. doi: 10.1016/j.jaap.2012.10.025
Mitchell, P., Simpson, A., Soong, R., and Simpson, M. (2016). Biochar amendment altered the molecular-level composition of native soil organic matter in a temperate forest soil. Environ. Chem. 13, 854–866. doi: 10.1071/EN16001
Nannipieri, P., Ascher, J., Ceccherini, M., Landi, L., Pietramellara, G., and Renella, G. (2003). Microbial diversity and soil functions. Eur. J. Soil Sci. 54, 655–670. doi: 10.1046/j.1351-0754.2003.0556.x
Neary, D., Bentrup, G., and Schoeneberger, M. (2017). “Appendix a: regional summaries: Southwest,” in Agroforestry: Enhancing Resiliency in US Agricultural Landscapes Under Changing Conditions. Gen. Tech. Report WO-96, eds Schoeneberger, M. Michele, Bentrup, Gary, Patel-Weynand, Toral (Washington, DC: US Department of Agriculture, Forest Service), 160–168.
Neill, C., Patterson, W., and Crary, D. (2007). Responses of soil carbon, nitrogen and cations to the frequency and seasonality of prescribed burning in a cape Cod oak-pine forest. Forest Ecol. Manage. 250, 234–243. doi: 10.1016/j.foreco.2007.05.023
North, M., Stephens, S., Collins, B., Agee, J., Aplet, G., Franklin, J., et al. (2015). Reform forest fire management. Science 349, 6254, 1280–1281. doi: 10.1126/science.aab2356
Nowacki, G., and Abrams, M. (2008). The demise of fire and “mesophication” of forests in the eastern United States. Bioscience 58, 123–138. doi: 10.1641/B580207
Nowak, J., Meeker, J., Coyle, D., Steiner, C., and Brownie, C. (2015). Southern pine beetle infestations in relation to forest stand conditions, previous thinning, and prescribed burning: evaluation of the southern pine beetle prevention program. J. For. 113, 454–462. doi: 10.5849/jof.15-002
Parker, J., Fernandez, I., Rustad, L., and Norton, S. (2001). Effects of nitrogen enrichment, wildfire, and harvesting on forest-soil carbon and nitrogen. Soil Sci. Soc. Am. J. 65, 1248–1255. doi: 10.2136/sssaj2001.6541248x
Parshall, T., Foster, D., Faison, E., MacDonald, D., and Hansen, B. (2003). Long-term history of vegetation and fire in pitch pine–oak forests on cape cod, massachusetts. Ecology 84, 736–748. doi: 10.1890/0012-9658(2003)084[0736:LTHOVA]2.0.CO;2
Patterson, W. A., and Backman, A. E. (1988). “Fire and disease history of forests,” in Vegetation History, 3rd Edn. eds B. Huntley and T. Webb (Dordrecht: Kluwer Academic), 603–632.
Patterson, W. A., Saunders, K. E., and Horton, L. J. (1983). Fire regimes of Cape Cod National Seashore. US Department of the Interior, National Park Service, North Atlantic Region, Office of Scientific Studies.
Preston, C., and Schmidt, M. (2006). Black (pyrogenic) carbon in boreal forests: a synthesis of current knowledge and uncertainties. Biogeosci. Discuss. 3, 211–271. doi: 10.5194/bgd-3-211-2006
Qi, Z., Jingfang, S., and Wenwei, L. (2018). A survey about characteristics of soil water retention curve. IOP Conference Series: Earth Environ. Sci. 153:062076. IOP Publishing. doi: 10.1088/1755-1315/153/6/062076
Renninger, H., Clark, K., Skowronski, N., and Schäfer, K. (2013). Effects of a prescribed fire on water use and photosynthetic capacity of pitch pines. Trees 27, 1115–1127. doi: 10.1007/s00468-013-0861-5
Santin, C., Doer, S., Merino, A., Bucheli, T., Bryant, B., Ascough, P., et al. (2017). Carbon sequestration potential and physicochemical properties differ between wildfire charcoals and slow-pyrolysis biochars. Sci. Rep. 7, 1–11. doi: 10.1038/s41598-017-10455-2
Saursaunet, M., Mathisen, K., and Skarpe, C. (2018). Effects of Increased soil scarification intensity on natural regeneration of scots pine pinus sylvestris L. and birch betula spp. L. Forests 9:262. doi: 10.3390/f9050262
Schaffers, A. P. (2002). Soil, biomass, and management of semi-natural vegetation–part II. factors controlling species diversity. Plant Ecol. 158, 247–268. doi: 10.1023/A:1015545821845
Sigmund, G., Hüffer, T., Hofmann, T., and Kah, M. (2017). Biochar total surface area and total pore volume determined by N2 and CO2 physisorption are strongly influenced by degassing temperature. Sci. Total Environ. 580, 770–775. doi: 10.1016/j.scitotenv.2016.12.023
Spokas, K. A. (2010). Review of the stability of biochar in soils: predictability of O: C molar ratios. Carbon Manage. 1, 289–303. doi: 10.4155/cmt.10.32
Swanston, C., Brandt, L., Janowiak, M., Handler, S., Butler-Leopold, P., Iverson, L., et al. (2018). Vulnerability of forests of the Midwest and Northeast United States to climate change. Clim. Change 146, 103–116. doi: 10.1007/s10584-017-2065-2
Thomas, S., and Gale, N. (2015). Biochar and forest restoration: a review and meta-analysis of tree responses. New For. 46, 931–946. doi: 10.1007/s11056-015-9491-7
Úbeda, X., Lorca, M., Outeiro, L. R., Bernia, S., and Castellnou, M. (2005). Effects of prescribed fire on soil quality in Mediterranean grassland (Prades Mountains, north-east Spain). Int. Wildland Fire 14, 379–384. doi: 10.1071/WF05040
Warnock, D. D., Johannes Lehmann, j., Kuyper, T. W., and Rillig, M. C. (2007). Mycorrhizal responses to biochar in soil–concepts and mechanisms. Plant Soil 300, 9–20. doi: 10.1007/s11104-007-9391-5
Williams, J., and Jackson, S. (2007). Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 5, 475–482. doi: 10.1890/070037
Keywords: PyC, prescribed fire, charcoal, forest ecology, soil moisture, water use efficiency, conservation
Citation: Licht J and Smith NG (2020) Pyrogenic Carbon Increases Pitch Pine Seedling Growth, Soil Moisture Retention, and Photosynthetic Intrinsic Water Use Efficiency in the Field. Front. For. Glob. Change 3:31. doi: 10.3389/ffgc.2020.00031
Received: 11 August 2019; Accepted: 02 March 2020;
Published: 31 March 2020.
Edited by:
Warren E. Heilman, United States Forest Service (USDA), United StatesReviewed by:
Melisa Blackhall, Universidad Nacional del Comahue, ArgentinaKenneth L. Clark, Northern Research Station, Forest Service (USDA), United States
Copyright © 2020 Licht and Smith. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeff Licht, amVmZmxpY2h0NjRAZ21haWwuY29t