
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change, 19 February 2020
Sec. Fire and Forests
Volume 3 - 2020 | https://doi.org/10.3389/ffgc.2020.00013
This article is part of the Research TopicImpact of Fire on BiodiversityView all 4 articles
 Giselda Durigan1,2*
Giselda Durigan1,2* Natashi A. L. Pilon2
Natashi A. L. Pilon2 Rodolfo C. R. Abreu3,4
Rodolfo C. R. Abreu3,4 William A. Hoffmann3
William A. Hoffmann3 Marcio Martins5
Marcio Martins5 Bruno F. Fiorillo6
Bruno F. Fiorillo6 Alexsander Z. Antunes7Ana Paula Carmignotto8
Alexsander Z. Antunes7Ana Paula Carmignotto8 Jonas B. Maravalhas9
Jonas B. Maravalhas9 Jésica Vieira9
Jésica Vieira9 Heraldo L. Vasconcelos9
Heraldo L. Vasconcelos9Although savannas are fire-adapted ecosystems, prescribing fire for biodiversity conservation remains controversial at least in some regions where savannas occur. Faced with uncertainty, many decision makers and even scientists are still reluctant to prescribe fire for conservation purposes in fire-prone ecosystems, invoking the precautionary principle. Knowledge gaps on the ideal fire regime, such as how and when to burn, and especially the fear of biodiversity losses, are among the main arguments against fire management applied to remnants of native savanna vegetation. To inform this debate, we assessed the impact of prescribed fires on diversity of plants (different growth forms), ants, frogs, lizards, birds, and small mammals, in savannas and grasslands of the Brazilian Cerrado. We assessed the existing species richness, composition, and abundance in areas subjected to long periods of fire suppression and compared to that observed over a short period after prescribed dry-season fires, within each group of plants and animals. Whenever possible, we carried out separate analyses for grassland and savanna. Burning did not significantly reduce species richness of any of the groups analyzed, but had a positive effect on richness of graminoids in grassland. When analyzed at the species level, abundance of most animal groups did not show consistent responses to fire, except for a decrease in some frog populations in grasslands. Forbs, graminoids, and subshrubs increased in abundance after fire in grassland areas, though in savanna areas, abundance of forbs, and subshrubs tended to decline after fire. Species composition changed little in response to fire as indicated by low levels of dissimilarity between burned and unburned areas. These results confirm the high resilience of Cerrado biota to fire, as expected for savanna ecosystems in general. Besides, we demonstrate here that the risk of biodiversity losses cannot justify the objections to the use of prescribed fire for conservation purposes in the Cerrado.
Fire has been a natural process for hundreds of millions of years, controlling the spatial distribution of biomes and maintaining the structure and functioning of fire-prone ecosystems (Scott, 2000; Bond and Keeley, 2005), long before humans arose (Scott, 2000). Fire has therefore been a significant evolutionary force for species living in tropical grassy biomes (Simon et al., 2009; Keeley et al., 2011; Simon and Pennington, 2012), resulting in diverse adaptations and an overall dependence on fire in many cases (Pilon et al., 2018; Fidelis et al., 2019). Despite evidence that mesic savannas need fire to maintain their structure and biodiversity (Bond and Parr, 2010; Andersen et al., 2012; Abreu et al., 2017; Rosan et al., 2019), prescribing fire is still controversial in many areas (Driscoll et al., 2010; Fidelis and Pivello, 2011; Pivello, 2017). Arguments against fire management result from the historical mistakes in the use of fire, from current conservation policies aimed at mitigation of climate change, as well as from academia, invoking the Precautionary Principle. This principle states that preventive action should be taken against a potential danger even if there is scientific uncertainty whether the danger is real or not (Hansson, 2017). The “preventive action” has been, therefore, suppressing fire by all means, even without evidence that prescribed fires would cause damage to the ecosystems.
Prehistoric humans learned to use fire as one of the first tools to manage their world (Bond and Keeley, 2005) and have since modified natural fire regimes directly and indirectly in different ways since then (Pivello, 2011; Archibald, 2016). Especially in the last millennium, human use of fire has intensified in association with deforestation and land clearing, which were often followed by environmental degradation (Dean, 1997; Perlin, 2005). Indeed, forest fires have been widely used as a proxy for tropical deforestation (Nelson and Chomitz, 2011). For this reason, and because fire is directly associated with carbon emissions, global warming, putative biodiversity losses, and damage to private properties (Gill, 2012), it has been widely seen as something bad that should be avoided, a vision strongly reinforced by the media (Paveglio et al., 2011).
The bias for ecosystem management aimed at carbon sequestration and storage (Tilman et al., 2000; Yessoufou, 2017) or favoring forest ecosystems and tree species (Veldman et al., 2015a,b) has led to the misperception that fire suppression is always positive. Even when fire is recognized to have a natural role, suppression is continued due to insufficient information regarding the best time or frequency to burn. However, the costs of waiting for evidence-based prescriptions must also be considered, since fire suppression can lead to huge biodiversity losses (Bowman, 2000; Bond and Parr, 2010; Abreu et al., 2017) and other negative consequences. In Brazilian savannas, recent studies have shown dramatic changes in vegetation structure due to fire suppression, with negative impacts on the water-related ecosystem services (Honda and Durigan, 2016; Oliveira et al., 2017), and on biodiversity, by eliminating species of ants and plants typical of open habitats (Pinheiro et al., 2016; Abreu et al., 2017). Prescribed burnings have been considered, therefore, as a management tool to avoid the negative consequences of woody encroachment and biome shift (Durigan and Ratter, 2016; Rosan et al., 2019) as well as to mitigate the huge carbon emissions and biodiversity losses of uncontrolled high intensity fires (Silveira et al., 1999; Russell-Smith et al., 2017). The effects of prescribed fires on biodiversity in the Brazilian savanna, however, are still poorly understood.
Although fire effects on Cerrado biodiversity have deserved attention in the last decades (e.g., Sato and Miranda, 1996; Cavalcanti and Alves, 1997; Briani et al., 2004; Cintra and Sanaiotti, 2005; Costa et al., 2013; Maravalhas and Vasconcelos, 2014; Camargo et al., 2018; Mendonça et al., 2015a), the few available studies are unevenly distributed in space and concentrated in the core area of the biome, hindering generalizations, and predictions valid for the whole range of environmental conditions within the Cerrado (Arruda et al., 2018). Additionally, most studies were carried out after unplanned wildfires, not comparing burned and unburned sites or periods before and after fire, thus not allowing conclusions about biodiversity losses due to fire. Furthermore, most biodiversity studies address a single taxonomic group, sometimes with contrasting results among sites or opposite tendencies compared to other savannas. For plants, community studies addressing fire effects on all components of Cerrado vegetation are lacking. Information about biodiversity responses to habitat changes caused by fire for a wide range of organisms is desirable to better inform management decisions (Bond and Archibald, 2003; Driscoll et al., 2010).
In this study, we adopted a multi-taxonomic approach to address fire effects on species diversity at the southern limit of Cerrado distribution, a region where these effects have rarely been experimentally assessed. We evaluated the short-term effects of prescribed burning on diversity of five functional groups of plants (graminoids, forbs, subshrubs, shrubs, and trees) and five taxonomic groups of animals (ants, frogs, lizards, birds, and small mammals) in a Brazilian savanna area that had been protected from fire for about three decades. We aimed to answer the following questions: (i) What is the response of plant and animal diversity to prescribed fires? (ii) Do distinct groups respond differently? (iii) Is there a risk of prescribed fires causing biodiversity losses in the Cerrado?
The study was carried out at Santa Bárbara Ecological Station (SBES), located at the southern limit of the savanna biome in Brazil (Águas de Santa Bárbara, state of São Paulo), within the coordinates 22°46′-22°51′ S and 49°10′-49°16′ W, with altitudes ranging from 600 to 680 m above sea level. The protected area preserves 2,715 ha of Cerrado vegetation encompassing a mosaic of grasslands, savannas, and forests. Climate in the region is classified as Köppen Cfa-type, hot, humid with a dry winter (Alvares et al., 2013), annual rainfall is around 1,300 mm, and mean monthly temperature of 18°C for the coldest month and over 22°C for the hottest month. Soil characteristics exhibit little variation in the study area, being deep oxisols with high sand and low nutrient content, high saturation of aluminum, and low soil water holding capacity (Abreu et al., 2017). Over the past three decades, there was a remarkable increase in tree biomass in the whole area, associated with fire-suppression policies, resulting in loss of open habitats and a major decrease in the number of plant and ant species in areas encroached by forest, particularly among savanna specialists (Abreu et al., 2017).
After about 30 years of fire suppression policy, we initiated an experiment of fire management at SBES, across the gradient from open grasslands to forest-type vegetation. The experiment was replicated in three sites within SBES, located at least 1.7 km from each other, with each site encompassing the whole vegetation gradient (grassland, savanna, and forest), and including burned and unburned (control) areas of each vegetation type. Hereafter, we treat these three sites as blocks, to make clear that all experiments described below were made within these blocks. The fire history over the 30 years before starting the experiment slightly differs among blocks and among vegetation plots. In one block, no fire was recorded before the first prescribed fire. In the second block, there was a single wildfire (in 2011) which affected the entire block. Within the third block, some vegetation plots had no fire, some were burned just once (2011) and one plot is in a small grassland patch that was burned several times in the 30-year period (the last time in 2011). Over the last 4 years before the first prescribed fire, therefore, none of the experimental blocks was burned. Historically, fire was used in the region every 2 years (Mendonça et al., 2015b), generally in the middle of the dry season, to renew pastures and to reduce woody growth, a common practice throughout the Cerrado for centuries (Dias, 2006). Therefore, fire at our experimental blocks occurred at a much lower frequency in the last decades than historically in the region. Prescribed fires were applied block by block, on different days, in the middle of the austral winter, before the peak of drought, in 2015, 2016, and 2017. In general, fire started around 10h00 a.m., under weather conditions recorded in the experimental blocks of wind speed below 5 km h−1, relative air humidity ranging from 45 to 80%, and temperature never surpassing 25°C.
All experiments described for the different groups were carried out in the blocks above described. Each block encompasses a tree biomass gradient from grassland to forest, including burned and unburned areas (controls). Forest vegetation (the cerradão) was not included in this study because prescribed fires were unsuccessful due to the low flammability of this system. Total area burned varied among blocks, but exceeded 35 ha in 2015, reaching about 70 ha in 2016 and 2017.
Sampling methods and periods of observations differed among taxonomic and functional groups as necessary. In all cases, our sampling for this study was limited to examining the effect of a single fire and does not provide information about the consequences of repeated burning over long periods of time. For ants and plants, sampling units were 20 m × 50 m plots (0.1 ha each) distributed in the three blocks, representing savanna (cerrado sensu stricto, typical savanna) and grassland (open savanna, dominated by grasses, forbs, and subshrubs, interspersed with occasional trees). Data were collected before the first experimental fires and several months after fire (six for ants, eight for plants) in the same plots. Birds were sampled immediately and 2 months after the second experimental fire, in burned and unburned patches of savanna and grassland within the blocks. Amphibians and reptiles were also sampled after the second fire, over 1 year, using pitfall traps installed in burned and unburned grassland patches in two blocks. Small mammals were sampled over 1 year in the same pitfall traps, after the third prescribed fire. Detailed description of sampling procedure for each group is presented in the following topics. For each group, we assessed differences or changes in species richness, abundance, and composition, in burned areas compared to unburned areas within the first year after fire, and separately by vegetation type (savanna or grassland) when possible. Methods used for data analyses are presented below.
Plants were sampled in the 0.1-ha plots, including three in grassland (one per block) and nine in savanna plots (three per block), before the first prescribed fires, carried out in 2015, and 1 year later (8 months after fire). Within each plot, we measured the stem diameter at breast height (dbh) of all trees with dbh ≥ 5 cm. Trees, treelets, and shrubs with 1 cm ≤ dbh < 5 cm were measured within ten 5-m × 5-m subplots distributed regularly throughout each plot. Woody plants smaller than 1 cm dbh (trees, treelets, and shrubs), as well as subshrubs, lianas, dwarf palms, and the herbaceous community (grasses, forbs, vines, and sedges) were sampled within 40 1-m × 1-m subplots evenly spaced on a grid within each plot. All individuals sampled within these plots were identified to species level in the field or collected for subsequent identification. To quantify changes in vegetation structure due to fire, we calculated tree basal area and tree density (dbh ≥ 5 cm) before the prescribed burns and again after fire in the 12 plots, on the basis of trees top-killed by fire. We calculated average values (percent changes) for grassland and for savanna plots.
Plant species were classified into five functional groups, with some underrepresented growth forms merged: trees (including treelets, large palms, and lianas), shrubs (including dwarf palms), subshrubs, forbs (including vines), and graminoids (grasses and sedges). Species were also classified according to habitat preference into two groups: (i) savanna specialists and (ii) generalists, species that occur in both savannas and forests (see Abreu et al., 2017).
Ants were collected in the nine 0.1-ha plots of savanna (three in each block) approximately 6 months before the first prescribed fire (winter 2015) and again 6 months after fire. Sampling in grasslands took place 6 months after fire, when we sampled three burned plots (one in each block) and three control, unburned plots (one in each block). Five 2.5-m × 2.5-m grids were established along the borders of each vegetation plot, keeping a minimum distance of 20 m between any two sampling grids. Four pitfall traps were set in each grid, totaling 20 traps per plot. Pitfall traps consisted of small plastic cups (250 ml, 8.5 cm high, and 7.8 cm in diameter) buried in the ground and partially filled with water and detergent. Traps remained in operation for 48 h and their contents were combined within grids. All ant specimens collected were sorted to morphospecies and, whenever possible, identified to species level using available taxonomic keys or through comparison with specimens previously identified by experts and deposited at the Zoological Collection of the Federal University of Uberlândia, Brazil. The proportion of species that are regarded as savanna/grassland specialists was estimated based on information provided by Vasconcelos et al. (2018).
A complete survey of birds observed in Santa Barbara Ecological Station over a long period before fire, 2008–2013, was carried out by Lucindo et al. (2015). For the present study, we recorded birds in the burned and adjacent unburned areas within the experimental blocks (savanna and grassland vegetation), after the second fire (winter 2016), by walking transects during and immediately after fire (10 h in August), and two months later (8 h in October), in the apex of the bird reproductive season. We sampled three transects (10 m wide, at least 200 m distant from each other) in burned areas and three transects in unburned areas in each sampling period. We walked slowly, at a speed of about 1 km h−1, registering each individual bird observed within fixed distances of 10 m on both transect sides. Since fire was patchy in some areas, for each observation we recorded if the bird was in burned or unburned vegetation, and also the habitat type – if it was savanna or grassland. Binoculars (8 × 40) and a digital camera with 42× optic zoom were used to identify the birds observed. We focused on resident species, excluding from analysis birds recorded only flying over the area, vultures and raptors attracted to the prescribed burning areas, and migratory species recorded only after October, e.g., Lesser Elaenia Elaenia chiriquensis Lawrence, 1865, Fork-tailed Flycatcher Tyrannus savana Daudin, 1802, and Plumbeous Seedeater Sporophila plumbea Wied, 1830. Species nomenclature followed Piacentini et al. (2015). We categorized species as savanna specialists or generalists on the basis of Motta-Júnior et al. (2008).
Frogs and lizards (the latter herein including amphisbaenians) were sampled in grassland of two blocks, each with a burned and an unburned sampling unit. Frogs and lizards were sampled over 1 year after the second prescribed burning (winter 2016). Sampling occurred from August 2016 to July 2017 for 10 days each month, totaling 3,840 pitfall trap-nights. Each of the four sampling units (two unburned, control units and two burned units) had two arrays of pitfall traps 60 m apart, each comprising a 40-m long line of four 100-L buckets and a plastic drift fence about 60–70 cm high, buried 10 cm into the soil. Vouchers were collected for identification at species level and will be deposited in the Museu de Zoologia (MZUSP), Universidade de São Paulo, São Paulo, and CFBH collection, Universidade Estadual Paulista, Rio Claro, Brazil. Young individuals of Physalaemus cuvieri and P. centralis are very difficult to identify in the field and thus both species were here considered together as Physalaemus cuvieri/centralis. Almost all species of frogs and lizards sampled in this study are grassland and savanna specialists (e.g., Brasileiro et al., 2005; Araujo et al., 2013, 2014). The few exceptions are habitat generalists that can be found in areas of forest as well as in disturbed areas (e.g., Ameiva ameiva among lizards and Physalaemus cuvieri among frogs).
Small mammals, herein represented by marsupials, rodents, and armadillos were sampled in grasslands of two blocks, in burned and adjacent unburned sampling units. In each of the four sampling units (two burned and two unburned control units) small mammals were surveyed by two arrays of pitfall traps (as described for frogs and lizards; see above). The mammals were sampled over 1 year after the third prescribed burning (winter 2017), from August 2017 to July 2018, in a 10-day monthly survey, totaling 3840 pitfall trap-nights, with similar efforts in burned (1920 pitfall trap-nights) and unburned (control) areas (1920 pitfall trap-nights). Vouchers were collected for identification at species level and will be deposited in the mammal collection of the Museu de Zoologia da Universidade de São Paulo (MZUSP). The taxonomic classification and nomenclature follow Gardner (2008) for armadillos, Voss and Jansa (2009) for marsupials, and Patton et al. (2015) for rodents. Species surveyed were classified as grassland/savanna specialists or as generalists—occurring on both open and forested habitats—based on habitat selectivity data from previous studies in the Cerrado (Marinho-Filho et al., 2002; Carmignotto et al., 2012, 2014) as well as from the study area (Carmignotto, pers. data).
We evaluated if prescribed fires affected species richness of each group by evaluating overall decline in species number, and by testing whether there was a decline in abundance and/or a decline in species number after accounting for a change in abundance.
To test for the overall effect of fire across all taxa we used paired t-tests comparing the log10-transformed number of species or the log10-transformed abundances of graminoids, forbs, subshrubs, shrubs, trees, ants, frogs, lizards, birds, and small mammals between the burned and unburned areas. To compare richness within groups after controlling for abundance, we generated individual-based rarefaction curves (Colwell et al., 2012) using the default options of EstimateS version 9.1.0 (Colwell, 2013). Rarefaction curves with non-overlapping 95% confidence intervals were considered significantly different (Colwell et al., 2012).
To quantify species turnover caused by fire, we calculated the Simpson index, which measures the degree of compositional differences between two areas (or periods) independently of differences in species richness between the areas (Koleff et al., 2003). We determined nestedness-resultant dissimilarity between the burned and unburned areas by calculating the difference between the Sørensen—which also measures the compositional similarity between two samples but considers differences in sample size, i.e., species richness—and Simpson indexes (Baselga, 2010). For calculation of the dissimilarity indexes we included all species recorded in each sampling period, except the singleton species (i.e., species that had just one individual recorded in any given sampling period). This was because the presence or absence of these latter species in a given sampling period often represents a sampling artifact and therefore the inclusion of singletons in our analyses would inflate true turnover rates. For all the analyses above, grassland and savanna were treated as distinct datasets, to check if responses differed between these vegetation types. This was done for most groups, except for frogs, lizards and small mammals, which were sampled only in grasslands.
Vegetation structure, indicated by tree basal area and tree density, was characterized before and after fire, to better illustrate the habitat and to quantify changes due to fire. Before fire, grassland plots had a mean tree basal area (±standard error) of 1.85 ± 0.80 m2 ha−1, and 217 ± 69 trees per hectare (dbh ≥ 5 cm), while savanna had a mean basal area of 7.93 ± 0.72 m2 ha−1 and 1,448 ± 127 trees per hectare. In grasslands, fire reduced the basal area by 29.4 ± 13.1% (reduction of 0.37 ± 0.12 m2 ha−1) and the density of adult trees by 23.3 ± 5.1% (47 ± 12 trees top-killed by fire per hectare). In the savanna plots, basal area was reduced by fire by 8.6 ± 1.3% (a decrease of 0.63 ± 0.09 m2 ha−1) and tree density by 5.6 ± 1.2% (73 ± 14 trees top-killed by fire per hectare).
Across all taxa and blocks, we recorded 435 species before fire (64.7% savanna/grassland specialists) and 423 after fire (66.2% savanna/grassland specialists, Table 1). Ants and birds were the most species-rich animal groups, in both grassland and savanna. Forbs were the most speciose group of plants in grasslands, whereas trees were the most diverse group in savannas. The proportion of species that are specialists of savanna and grassland habitats varied widely among taxa, ranging from 100% of the recorded species of subshrubs to only 11% of birds (Table 1). For most of the plants and animal groups, however, the open habitats specialists represented at least 60% of the sample, with ants (32%) and birds (11%) as exceptions (Table 1).
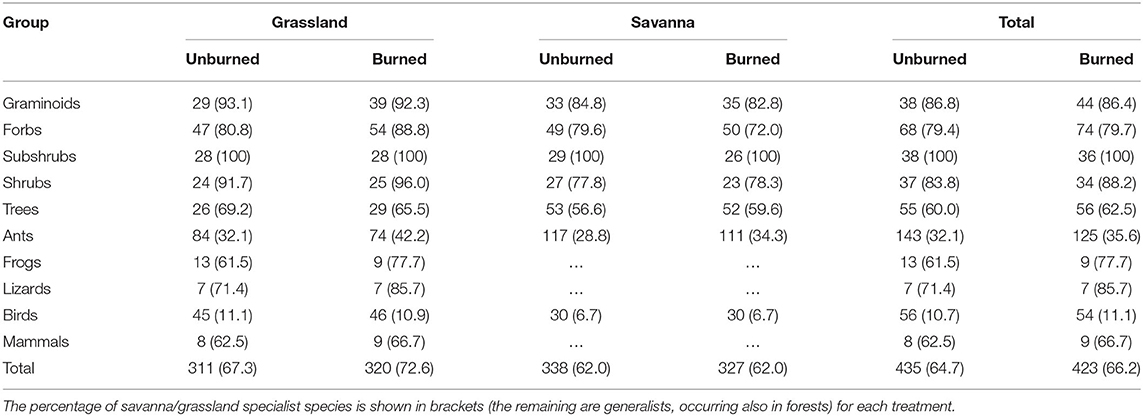
Table 1. Number of species recorded, for each group of plants and animals, in each vegetation type (grassland or savanna) in burned and unburned areas, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
Burned grasslands tended to have more species of graminoids, forbs, and trees, and fewer species of ants and frogs than those that were not burned, whereas burned savannas had slightly fewer species of subshrubs, shrubs, and ants (Table 1). The percentage of open-habitat specialists in burned grasslands increased for forbs, shrubs, ants, frogs, lizards, and mammals, and, in burned savannas, trees and ants showed higher proportions of specialists. Rarefaction curves, however, showed no difference in species richness between burned and unburned areas for any group (Figures 1, 2), with the exception of graminoids in grassland, where species richness increased after fire. Similarly, when we analyzed the responses of all groups together (i.e., when we tested if the number of species from each group found in the burned plots was equivalent or not to that found in the unburned plots) no significant differences were detected (Figure 3; paired t-tests, P > 0.05, Table S1). This was true whether we included all species or only the savanna/grassland specialists (Figure 3). However, considering only savanna/grassland specialists, we found that species richness in the grasslands (but not in savannas) was greater after than before the fire (Table S1).
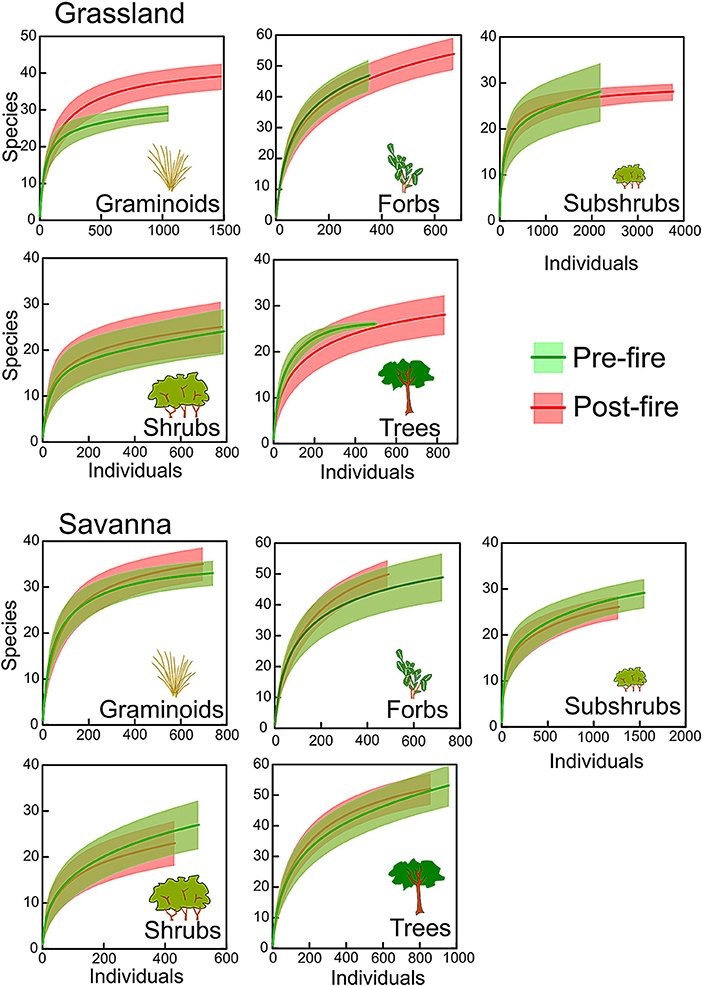
Figure 1. Individual-based rarefaction curves (with 95% confidence intervals) comparing species richness of different plant groups between the unburned (green) and burned plots (red) in grassland and savanna vegetation types, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
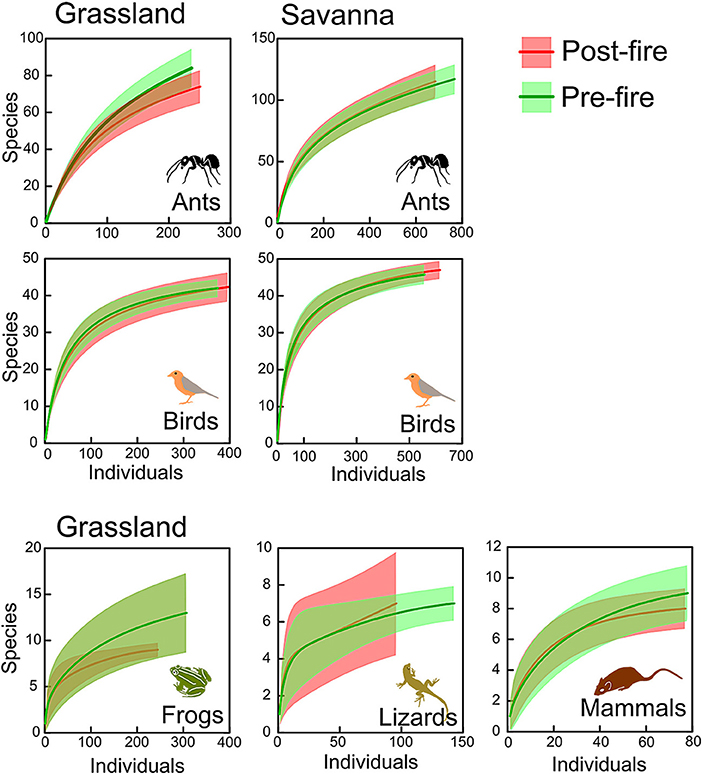
Figure 2. Individual-based rarefaction curves (with 95% confidence intervals) comparing species richness of different animal groups between the unburned (green) and burned areas (red) in grassland and savanna vegetation types, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
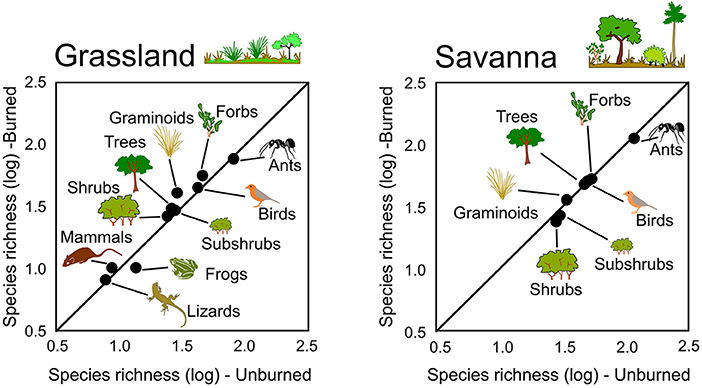
Figure 3. Species richness (log10-transformed data) of different plant and animal groups in unburned and burned areas for grassland and savanna vegetation types. The line represents the 1:1 equivalence line, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
We evaluated, for each group separately, if there was a consistent change in species abundances (i.e., if most species decreased or increased in abundance in response to fire) (Figures 4, 5). In savanna, most species of forbs and subshrubs declined in abundance after fire (paired t-test, P < 0.05; Figure 4, Table S1). In contrast, in the grasslands, abundance of most species of forbs, graminoids, and subshrubs increased with fire (paired t-test, P < 0.05, Figure 4, Table S1), whereas abundance of most frog species declined (paired t-test, P = 0.017, Figure 5, Table S1).
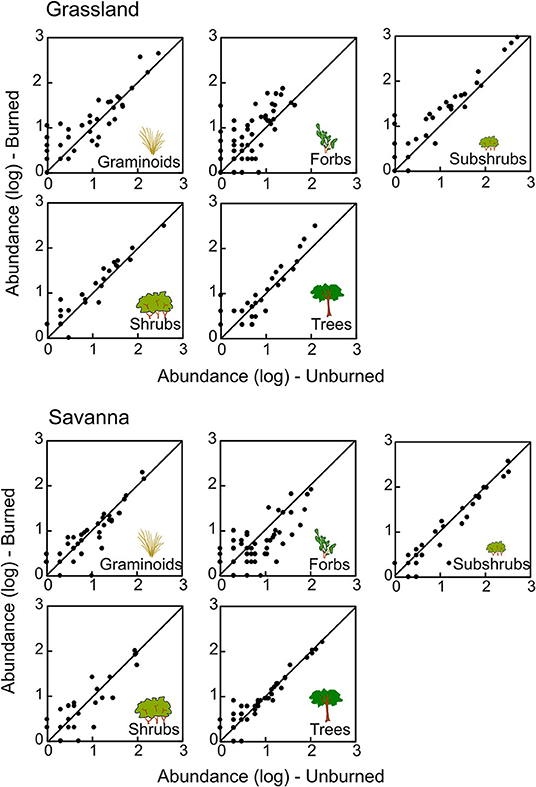
Figure 4. Abundance (log10-transformed data) of different plant groups sampled in unburned and burned plots of grassland and savanna. Each point represents one species. The line represents the 1:1 equivalence line, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
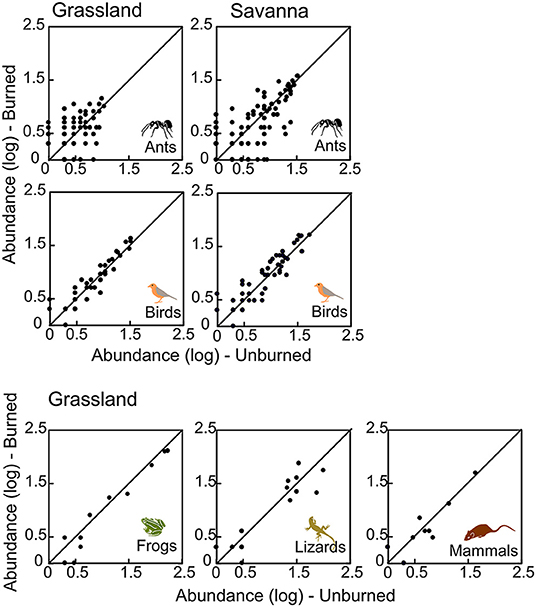
Figure 5. Abundance (log10-transformed data) of different animal groups sampled in unburned and burned areas of grassland and savanna. Each point represents one species. The line represents the 1:1 equivalence line, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
Overall, species composition changed little in response to fire as indicated by very low levels of dissimilarity (< 0.1 in most cases) between burned and unburned areas (Figure 6). Changes tended to be more marked in grasslands than in savannas. The dissimilarity between burned and unburned savannas tended to be caused mainly by species replacement (turnover), but in grasslands, nestedness was the main component of change in species composition (Figure 6). In grasslands, therefore, either new species appeared (as was the case for all plant groups) or some existing species were not sampled after fire (as was the case for frogs, mainly).
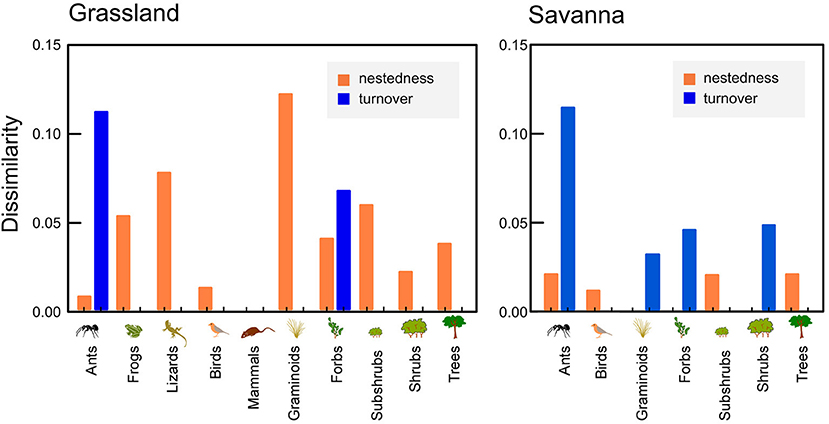
Figure 6. Degree of dissimilarity in species composition (due to turnover or nestedness) between burned and unburned areas for different plant and animal groups in grasslands and savannas, at Santa Bárbara Ecological Station, state of São Paulo, Brazil.
Here we assessed the effects of prescribed fires on species richness, abundance and composition of five groups of plants and five groups of animals within the same protected area in the southern limit of the Brazilian Cerrado. We found negligible effects of fire, with species richness unaltered, except in one of the 10 groups (graminoids increased after fire in grasslands). Slight changes in abundance and species composition were observed within some groups, differing in directions (increasing or decreasing) or processes (turnover or nestedness) between vegetation types (grassland and savanna). However, there were no significant changes in species richness between burned and unburned areas. We must highlight that our study was limited to the effects of a single fire, and that we burned our experimental areas under controlled conditions. Therefore, our results cannot be extrapolated to uncontrolled firestorms under extreme weather conditions or to annually burned ecosystems, both cases deserving specific investigation.
Our results are in accordance with most studies quantifying the effects of fire on tropical savanna biodiversity, which found little or no effect (van Wilgen et al., 2007; Frizzo et al., 2011), corroborating the adaptation of Cerrado biodiversity to fire, both among plants (e.g., Coutinho, 1982, 1990; Simon and Pennington, 2012) or animals (e.g. Vieira and Marinho-Filho, 1998; Layme et al., 2004; Prada and Marinho-Filho, 2004). Recent studies, however, have shown fire influences other than changes in richness, such as body conditions of lizards, which depend on habitat preferences of different species (Costa et al., 2019). Such differences deserve further investigation for other groups.
In our study, a single prescribed burning after a long period without fire caused substantial changes in vegetation structure, with tree density 23% lower in grasslands and 6% lower in savannas, and a tree basal-area reduction of 30% in grasslands and 9% in savannas, mostly due to top killing adult trees. Structural changes were more pronounced in grasslands than in savannas, due to the greater abundance of grasses in the former, which are the main fuel for fire. Thus, the expected role of fire in decreasing the density and biomass of arboreal vegetation tends to increase the structural differences if all phytophysignomies of the Cerrado mosaic are submitted to the same burning frequency. Most studies addressing fire effects on Cerrado plant communities have focused on trees, in general showing high stem mortality, biomass reduction and even diversity loss, with fire acting as a filter eliminating non adapted species (Durigan et al., 1994; Sato and Miranda, 1996; Hoffmann and Moreira, 2002; Medeiros and Miranda, 2005; Hoffmann et al., 2009; Lopes et al., 2009). While these studies demonstrate the efficacy of prescribed burns in avoiding woody encroachment and biome shifts, their results have been often interpreted as negative effects of fire on plant communities, since non-woody species have been disregarded. However, periodic fires are considered essential for maintaining floristic composition and open vegetation types in Brazilian savanna (Coutinho, 1990; Moreira, 2000; Fidelis and Pivello, 2011; Pivello, 2011, 2017). In addition, despite the decrease in aerial biomass and the remarkable changes in vegetation structure shortly after fire, woody vegetation of savannas is demographically resilient to fire (Higgins et al., 2007) and recovery can occur in a few years after fire, even in woodland savannas (Eiten, 1972; Furley et al., 2008). For the ground layer, the intense post-fire photosynthetic activity offsets carbon emissions due to burning in about 12 months (Santos et al., 2003).
Although changes in vegetation structure represent modifications in habitat for plants and animals, fire-driven changes in plant communities of tropical savannas due to fire are much more about structure than species composition or biodiversity (Higgins et al., 2007; van Wilgen et al., 2007; Lebbink et al., 2018), since fire does not eliminate typical savanna species. No biodiversity losses due to fire have been consistently observed in savannas for most animal groups, such as arthropods (but see Morais and Benson, 1988; Andersen and Müller, 2000; Maravalhas and Vasconcelos, 2014; Vasconcelos et al., 2017), birds (Cavalcanti and Alves, 1997; Corbett et al., 2003; Mills, 2004; Reis et al., 2016), mammals (Vieira, 1999; Prada and Marinho-Filho, 2004; Beale et al., 2018), reptiles (Griffiths and Christian, 1996; Corbett et al., 2003; Langford et al., 2007; Morais et al., 2011; Costa et al., 2013), and amphibians (Corbett et al., 2003; Langford et al., 2007; Morais et al., 2011). Despite the pattern of low impact on richness observed in most studies, however, there are some exceptions when fire effects are analyzed at a small scale, comparing different pieces of the vegetation mosaic. Costa et al. (2019) found half the lizard species favored in unburned plots while the other half was favored in burned plots. These results indicate that maintaining the vegetation mosaic can enhance beta and gamma diversity, a recommendation supported also by Andersen et al. (2012), who found advantage for small mammals in preserving some large infrequently burned areas.
When abundance within animal groups is considered in our study, we found significantly fewer individuals in burned areas only for frogs in grasslands, despite no difference found in the savannas or in richness in any habitat. For the other groups, the balance between increase and decrease of species populations resulted in no net changes in total abundance. During fires in Brazilian savannas, small animals like frogs, lizards, snakes, and small mammals seek refuge underground by getting into termite mounds and burrows made by armadillos and burrowing rodents (Coutinho, 1990; Faria and Araujo, 2004; Frizzo et al., 2011; Costa et al., 2013) or seeking for refuge in unburned patches nearby. The lack of marked decreases in the richness and abundance for most small animals (<1 kg of mass) after fires may reflect a combination of factors, like the apparent lack of direct effects of fire on them (inferred from the great difficulty of finding dead animals just after fire), the common use of underground retreats during fire (Coutinho, 1990; Vieira and Marinho-Filho, 1998), and the insectivorous and granivorous habits of most vertebrates we sampled (Pianka and Vitt, 2003; Wells, 2007; Frizzo et al., 2011; Vieira and Briani, 2013). In fact, many arthropods also use underground retreats during fire (M. Martins, unpubl. observations) and thus may stay in burned areas of Cerrado after fire. Furthermore, herbivorous insects also may stay in burned areas because the vegetation begins to sprout or germinate in just a few days after fire. Indeed, the massive flowering (Prada et al., 1995; Vieira et al., 1996) and seed production after fire (Pilon et al., 2018) attract insects and represent a considerable food increase to reptiles, birds, and mammals in post fire areas (Prada and Marinho-Filho, 2004; Frizzo et al., 2011; Vieira and Briani, 2013). Thus, food shortage may not be a problem for insectivorous animals after fire (Lepesqueur et al., 2012). Additionally, indirect positive effects of fire on fauna can be expected due to changes in vegetation, favoring movement of animals across the landscape (Henriques et al., 2000), such as birds and large mammals, attracted to the vegetation patches vigorously resprouting.
The most controversial faunal group in terms of response to fire in the literature has been the small mammals, for which we found no changes due to fire in species composition, richness, or abundance over a year after fire. Negligible effects were observed by Layme et al. (2004) in the abundance of a rodent species in Amazonian savannas. Other studies in Brazilian savannas have shown a quick decline followed by rapid recovery and increase in abundance and diversity over time (Vieira, 1999; Henriques et al., 2000, 2006; Briani et al., 2004). Conversely, in Australian savannas, relevant losses of small mammals after fire have been observed, but this has been attributed to predation by feral cats and invasive frogs, which is easier after fire (Firth et al., 2006; Woinarski et al., 2010; Lawes et al., 2015). Actually, under certain conditions, small mammals can be severely affected by fire and require attention (Andersen et al., 2005, 2012), especially when predators are favored by fire, which is the case of feral cats in Australia. Long-term studies have been recommended to assess fire impacts on small mammals, since diversity and abundance within this group can reach their maximum in the first 2 years after fire, decreasing over time if fire is suppressed (Briani et al., 2004; Henriques et al., 2006 in Brazilian savanna). This population dynamics can be understood, primarily, as a direct effect of the input of resources after fire, that allows increase in population sizes in the next generations (Vieira, 2003; Henriques et al., 2006; Vieira and Briani, 2013). However, to understand the impacts of fire on small mammals, we must understand how their assemblages are structured in the Brazilian Cerrado. For instance, studies at both local and regional scales have shown that small mammals are composed of savanna specialists and forest dwellers at roughly similar proportions, with few generalist species (Carmignotto et al., 2012, 2014). Since fire alters vegetation structure, in the sense of reducing arboreal biomass and increasing herbaceous layer, these changes directly affect the small mammals, favoring open habitat dwellers and consequently decreasing forest specialists in burned areas (Henriques et al., 2006; Camargo et al., 2018).
Among plant groups, the effects of fire were slightly more visible than for animal groups. We found change in species richness, but only for graminoids in grassland areas, with colonization by new species triggered by fire. The arrival of 10 new species of grasses was likely favored by two fire effects: (i) intensifying sexual reproduction (Eiten, 1972; Coutinho, 1977, 1990; Le Stradic et al., 2015; Pilon et al., 2018) and (ii) clearing the ground and thus favoring seed dispersal and germination (Coutinho, 1990; Fidelis et al., 2012), since both processes are constrained by the vast amount of biomass accumulated after a long period without fire. Studies on grasses and forbs of tropical savannas have shown that the effect of fire reducing woody biomass and cover is remarkably beneficial for the ground layer plant communities (Sheuyange et al., 2005; Zimmermann et al., 2010). This has been already observed in temperate savannas and grasslands, where fire is essential for the maintenance of open canopy and thus perpetuating the herbaceous plant communities (Morgan, 1998; Sparks et al., 1998; Overbeck et al., 2005; Fidelis et al., 2012). Moreover, in Cerrado grasslands, long-term fire suppression also reduced aboveground biomass of graminoids, forbs, and subshrubs (Oliveras et al., 2013).
We also observed changes in abundance for some groups of plants, and the response to fire was different between grasslands and savannas in the direction of changes in species populations. While the abundance of most species of forbs and graminoids increased in grassland, clearly favored by fire, the abundance of forbs and subshrubs declined in savanna, where fire was not strong enough and light is nearly certainly a limiting resource for sun-demanding small plants. After long periods without fire, forbs and graminoids likely spent their reserves to survive in the shade (Pinheiro et al., 2016), reducing their ability to sprout or to colonize from seeds when fire returned to the system (Zimmermann et al., 2010).
Overall, community composition of most plant and animal groups underwent very small changes due to fire. For plants, changes were a little more pronounced in grasslands, where the arrival of new species after fire (and no species losses) indicates that the pre-fire community represents a nested subset of the post-fire community. In contrast, changes in species composition of graminoids, forbs, and subshubs in savannas were exclusively due to species turnover—rather than nestedness—following fire. Ants showed the highest level of species turnover among the animal groups. However, all ant species that appeared or disappeared following fire were rare in our sampling, suggesting that at least part of the detected turnover may simply be an artifact of incomplete sampling. Changes in ant species composition are likely to be driven by changes in the vegetation structure rather than by direct effects of fire (Andersen et al., 2012). In fact, long-term experimental burning of cerrado vegetation showed to significantly alter the composition of ant species (Maravalhas and Vasconcelos, 2014).
Species richness, for any group of plants or animals, is directly related with the mosaic of habitats present in a region, with beta diversity (i.e., diversity among distinct habitats) greatly surpassing alpha diversity (i.e., diversity within one type of habitat), pointing to the importance of open and forested formations for species richness and composition in the Cerrado (Silva and Bates, 2002; Durigan and Ratter, 2006; Nogueira et al., 2009, 2011; Carmignotto and Aires, 2011; Santos-Filho et al., 2012; Amaral et al., 2017). So, the homogenization of vegetation structure, as seen in the case of increased woody encroachment in the absence of fire, affecting grassland/savanna specialists (Vieira and Marinho-Filho, 1998; Vieira, 1999; Henriques et al., 2000, 2006; Briani et al., 2004; Maravalhas and Vasconcelos, 2014; Abreu et al., 2017), will disrupt community structure resulting in biodiversity losses. Studies of open formations around the world also highlight the importance of habitat heterogeneity for maintaining biodiversity of the landscape (Andersen et al., 2012; Beale et al., 2018; McCleery et al., 2018; Bond, 2019; Costa et al., 2019). At our study area, open grasslands and savannas have particular importance for conservation at local and regional scales. These habitats have been extirpated from most regions of São Paulo state (Durigan and Ratter, 2006), and as a consequence many species of plants (Durigan et al., 2018) and animals dependent upon open, grassy habitats (Carmignotto and Monfort, 2006; Duarte and Vogliotti, 2009; Lemos and Azevedo, 2009) are currently extinct or under threat, and are restricted to a few scattered localities in protected areas (see Percequillo and Kierulff, 2009; São Paulo, 2018). The long-term fire suppression policies applied to protected areas in Brazil have contributed to the encroachment of woody vegetation, which results in the loss of biodiversity in these areas, with open habitats and their peculiar species disappearing (Abreu et al., 2017).
A well-planned fire management program should set as the primary goal for the maintenance of different vegetation types, with variable structure, which is a surrogate for biodiversity at different scales. That depends on defining fire frequency according to the expectation of biomass decrease in distinct patches of the landscape. It is still controversial if mimicking the “historical fire regime” will maintain that mosaic and result in successful conservation (Freeman et al., 2017). And there is little evidence that pyrodiversity (varying fire season, frequency, intensity etc.) begets biodiversity (Parr and Andersen, 2006; Davies et al., 2012; Farnsworth et al., 2014; Maravalhas and Vasconcelos, 2014). Instead, evidence points to a binary fire strategy – burned and unburned patches sharing the space – as sufficient to maintain savanna biodiversity. Preserving patches of unburned ecosystems can guarantee refuge for fire-sensitive species which could not survive if the whole landscape is simultaneously or too frequently burned. On the other hand, burning some patches frequently would guarantee survival and successful reproduction of plant and animal species adapted to open and grassy vegetation.
Fire has been suppressed for long periods in many countries that adopted misinformed conservation strategies, but these strategies have been revised on the basis of evidence (Caillault et al., 2015). Using fire to manage tropical savannas has been increasingly supported by ecological studies, especially in Africa and Australia (Andersen et al., 1998, 2005; Biggs, 2002, 2003; Furley et al., 2008; van Wilgen, 2009; Woinarski and Legge, 2013; Caillault et al., 2015; Archibald, 2016). There is a global consensus among savanna ecologists that burning is essential for maintaining heterogeneity and biodiversity of tropical savannas (Coutinho, 1982, 1990; Bond and Keeley, 2005; Andersen et al., 2012; Maravalhas and Vasconcelos, 2014; Durigan and Ratter, 2016; Pivello, 2017; Schmidt et al., 2018). On the other hand, fire suppression has shown to be ecologically and economically unsustainable (Bowman et al., 2013), eventually resulting in catastrophic firestorms that cause huge carbon emissions and biodiversity losses (Silveira et al., 1999; Bond and Archibald, 2003; França, 2010; Pivello, 2011; Batista et al., 2018; Fidelis et al., 2018). In the last years, fire experiments have increased in Brazil (Dias and Miranda, 2010; Rissi et al., 2017; Schmidt et al., 2018), including our experiment presented here. Here we demonstrate that prescribed fires do not cause losses in species richness of plants and animals, and even bring gains in richness and abundance of plant species in Cerrado grasslands. These results confirm the resilience of the Cerrado biota to fire, as expected for savanna ecosystems in general, therefore supporting the use of fire management for conservation and restoration purposes in the Cerrado. We argue that prescribed burns should be applied not only to avoid uncontrolled wildfires, but also to maintain open vegetation types and their biodiversity as fundamental pieces of the Cerrado mosaic, thus enhancing beta and gamma diversity, threatened by homogenization due to fire suppression or by modified fire regimes.
The datasets generated for this study are available on request to the corresponding author.
The animal study was reviewed and approved by Comissão de Ética no Uso de Animais (CEUA), Instituto de Biociências, Universidade de São Paulo.
GD, WH, HV, and MM designed research. GD, NP, RA, WH, MM, BF, AA, AC, JM, JV, and HV performed research. HV and JM analyzed data. GD led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for submission.
This material is based upon work supported by the National Science Foundation under Grant Number DEB1354943 to WH; NP was funded by FAPESP grant #2016/17888-2, MM, BF, and AC were funded by FAPESP grants #2015/21259-8 and #2018/14091-1, and GD was funded by CNPq grant #303179/2016-3.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Márcio S. Suganuma, Geissianny B. Assis, Mário G. B. Cava, Éliton R. Silveira, Flaviana M. Souza, Elmo Koch, Paula Rocha Lemos, Mirela Alcolea, Giovana Ribeiro Felício, Karine Yumi Tominaga, Mariana Dias Guilardi, Bárbara Armando Godinho, Carolina Henkes Inamassu, Julia dos Santos Gutierres, Guilherme Zamarim Rezende, Luciana de Oliveira Furtado, Evelin de Campos da Costa, Jorge Henry Maciel dos Santos, Nathany Biela, Carolina Farhat, and Rafael Menegucci for assistance with data collection, and the employees of the Instituto Florestal for logistical assistance. The research project was carried out under COTEC research license #26108-008.476/2014, CETESB fire license #035354, and ICMBio license #50658-1.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2020.00013/full#supplementary-material
Abreu, R. C. R., Hoffmann, W. A., Vasconcelos, H. L., Pilon, N. A. L., Rossatto, D. R., and Durigan, G. (2017). The biodiversity cost of carbon sequestration in tropical savanna. Sci. Adv. 3:e1701284. doi: 10.1126/sciadv.1701284
Alvares, C. A., Stape, J. L., Sentelhas, P. C., Moraes, G., Leonardo, J., and Sparovek, G. (2013). Köppen's climate classification map for Brazil. Meteorol. Z. 22, 711–728. doi: 10.1127/0941-2948/2013/0507
Amaral, A. G., Munhoz, C. B., Walter, B. M., Aguirre-Gutiérrez, J., and Raes, N. (2017). Richness pattern and phytogeography of the Cerrado herb–shrub flora and implications for conservation. J. Veg. Sci. 28, 848–858. doi: 10.1111/jvs.12541
Andersen, A. N., Braithwaite, R. W., Cook, G. D., Corbett, L. K., Williams, R. J., Douglas et al. (1998). Fire research for conservation management in tropical savannas: introducing the Kapalga fire experiment. Aust. J. Ecol. 23, 95–110. doi: 10.1111/j.1442-9993.1998.tb00708.x
Andersen, A. N., Cook, G. D., Corbett, L. K., Douglas, M. M., Eager, R. W., Russell-Smith, J., et al. (2005). Fire frequency and biodiversity conservation in Australian tropical savannas: implications from the Kapalga fire experiment. Austral Ecol. 30, 155–167. doi: 10.1111/j.1442-9993.2005.01441.x
Andersen, A. N., and Müller, W. J. (2000). Arthropod responses to experimental fire regimes in an Australian tropical savannah: ordinal-level analysis. Austral Ecol. 25, 199–209. doi: 10.1046/j.1442-9993.2000.01038.x
Andersen, A. N., Woinarski, J. C., and Parr, C. L. (2012). Savanna burning for biodiversity: fire management for faunal conservation in Australian tropical savannas. Austral Ecol. 37, 658–667. doi: 10.1111/j.1442-9993.2011.02334.x
Araujo, C., Nascimento, B., Maffei, F., Moya, G., Corrêa, D., and Santos, S. (2014). Lizards from Estação Ecológica de Santa Bárbara, a remnant of Cerrado in the state of São Paulo, Brazil. Check List 10:1038. doi: 10.15560/10.5.1038
Araujo, C. D., Corrêa, D. T., and Santos, S. M. (2013). Anurans of the Estação Ecológica de Santa Bárbara, a remnant of open Cerrado formations in the state of São Paulo. Biota Neotrop. 13, 230–240. doi: 10.1590/S1676-06032013000300026
Archibald, S. (2016). Managing the human component of fire regimes: lessons from Africa. Phil. Trans. R. Soc. B. 371:20150346. doi: 10.1098/rstb.2015.0346
Arruda, F. V. D., Sousa, D. G. D., Teresa, F. B., Prado, V. H. M. D., Cunha, H. F. D., and Izzo, T. J. (2018). Trends and gaps of the scientific literature about the effects of fire on Brazilian Cerrado. Biota Neotrop. 18:e20170426. doi: 10.1590/1676-0611-bn-2017-0426
Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Global Ecol. Biogeogr. 19, 134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Batista, E. K. L., Russell-Smith, J., França, H., and Figueira, J. E. C. (2018). An evaluation of contemporary savanna fire regimes in the Canastra National Park, Brazil: Outcomes of fire suppression policies. J. Environ. Manage. 205, 40–49. doi: 10.1016/j.jenvman.2017.09.053
Beale, C. M., Mustaphi, C. J. C., Morrison, T. A., Archibald, S., Anderson, T. M., Dobson, A. P., et al. (2018). Pyrodiversity interacts with rainfall to increase bird and mammal richness in African savanas. Ecol. Lett. 21, 557–567. doi: 10.1111/ele.12921
Biggs, H. C. (2002). Proposed policy for the ecosystem management of fire in the Kruger National Park. Unpublished Policy Document, Kruger National Park (Revised 2005). Available online at: https://www.sanparks.org/ (accessed August 03, 2019).
Biggs, H. C. (2003). “Integration of science: success, challenges, and the future,” in The Kruger Experience: Ecology and Management of Savanna Heterogeneity, eds J. du Toit, K. H. Rogers, and H. C. Biggs (New York, NY: Island Press), 468.
Bond, W. J., and Archibald, S. (2003). Confronting complexity: fire policy choices in South African savanna parks. Int. J. Wildland Fire 12, 381–389. doi: 10.1071/WF03024
Bond, W. J., and Keeley, J. E. (2005). Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 20, 387–394. doi: 10.1016/j.tree.2005.04.025
Bond, W. J., and Parr, C. L. (2010). Beyond the forest edge: ecology, diversity and conservation of the grassy biomes. Biol. Conserv. 143, 2395–2404. doi: 10.1016/j.biocon.2009.12.012
Bowman, D. M. J. S. (2000). Australian Rainforests: Islands of Green in the Land of Fire. Cambridge: Cambridge University Press.
Bowman, D. M. J. S., Murphy, B. P., Boer, M. M., Bradstock, R. A., Cary, G. J., Cochrane, M. A., et al. (2013). Forest fire management, climate change, and the risk of catastrophic carbon losses. Front. Ecol. Environ. 11, 66–68. doi: 10.1890/13.WB.005
Brasileiro, C. A., Sawaya, R. J., Kiefer, M. C., and Martins, M. (2005). Amphibians of an open cerrado fragment in Southeastern Brazil. Biota Neotrop. 5, 93–109. doi: 10.1590/S1676-06032005000300006
Briani, D. C., Palma, A. R. T., Vieira, E. M., and Henriques, R. P. B. (2004). Post-fire succession of small mammals in the Cerrado of central Brazil. Biodivers. Conserv. 13, 1023–1037. doi: 10.1023/B:BIOC.0000014467.27138.0b
Caillault, S., Ballouche, A., and Delahaye, D. (2015). Where are the ‘bad fires’ in West African savannas? Rethinking burning management through a space–time analysis in Burkina Faso. Geogr. J. 181, 375–387. doi: 10.1111/geoj.12074
Camargo, A. C. L., Barrio, R. O. L., Camargo, N. F., Mendonça, A. F., Ribeiro, J. F., Rodrigues, C. M. F., et al. (2018). Fire affects the occurrence of small mammals at distinct spatial scales in a neotropical savana. Eur. J. Wildlife Res. 64:63. doi: 10.1007/s10344-018-1224-8
Carmignotto, A. P., and Aires, C. C. (2011). Mamíferos não voadores (Mammalia) da Estação Ecológica Serra Geral do Tocantins. Biota Neotrop. 11, 307–322. doi: 10.1590/S1676-06032011000100029
Carmignotto, A. P., Bezerra, A. M., and Rodrigues, F. H. (2014). Nonvolant small mammals from a southwestern area of Brazilian Cerrado: diversity, habitat use, seasonality, and biogeography. Therya 5, 535–558. doi: 10.12933/therya-14-197
Carmignotto, A. P., and Monfort, T. (2006). Taxonomy and distribution of the Brazilian species of Thylamys (Didelphimorphia: Didelphidae). Mammalia 70, 126–144. doi: 10.1515/MAMM.2006.013
Carmignotto, A. P., Vivo, M. D., and Langguth, A. (2012). “Mammals of the Cerrado and Caatinga: distribution patterns of the tropical open biomes of Central South America,” in Bones, Clones and Biomes - The History and Geography of Recent Neotropical Mammals, eds B. D. Patterson and L. P. Costa (Chicago: University of Chicago Press), 307–350.
Cavalcanti, R. B., and Alves, M. A. (1997). Effects of fire on savanna birds in central Brazil. Ornitol. Neotrop. 8, 85–87.
Cintra, R., and Sanaiotti, T. M. (2005). Fire effects on the composition of a bird community in an Amazonian savanna (Brazil). Braz. J. Biol. 65, 683–695. doi: 10.1590/S1519-69842005000400016
Colwell, R. K. (2013). EstimateS: Statistical Estimation of Species Richness and Shared Species From Samples. Version 9. User's Guide and Application. Available online at: http://purl.oclc.org/estimates (accessed July 20, 2019).
Colwell, R. K., Chao, A., Gotelli, N. J., Lin, S. Y., Mao, C. X., Chazdon, R. L., et al. (2012). Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. J. Plant Ecol. 5, 3–21. doi: 10.1093/jpe/rtr044
Corbett, L. K., Andersen, A. N., and Müller, W. J. (2003). “Terrestrial vertebrates,” in Fire in Tropical Savannas: The Kapalga Experiment, eds A. N. Andersen, G. D. Cook, and R. J. Williams (New York, NY: Springer), 126–152.
Costa, B. M., Pantoja, D. L., Sousa, H. C., Queiroz, T. A., and Colli, G. R. (2019). Long-term, fire-induced changes in habitat structure and microclimate affect Cerrado lizard communities. Biodivers. Conserv. 6, 1–23. doi: 10.1007/s10531-019-01892-8
Costa, B. M., Pantoja, D. L., Vianna, M. C., and Colli, G. R. (2013). Direct and short-term effects of fire on lizard assemblages from a Neotropical Savanna hotspot. J. Herpetol. 47, 502–510. doi: 10.1670/12-043
Coutinho, L. M. (1977). Aspectos ecológicos do fogo no cerrado: II. As queimadas e a dispersão de sementes. Bol. Bot. 5, 57–64. doi: 10.11606/issn.2316-9052.v5i0p57-63
Coutinho, L. M. (1982). “Ecological effects of fire in Brazilian cerrado,” in Ecology of Tropical Savannas, eds B. J. Huntley and B. H. Walker (Berlin: Springer-Verlag), 273–291. doi: 10.1007/978-3-642-68786-0_13
Coutinho, L. M. (1990). “Fire in the ecology of the Brazilian Cerrado,” in Fire in the Tropical Biota: Ecosystem Processes and Global Challenges, ed J. G. Goldammer (Berlin: Springer Verlag), 82–105.
Davies, A. B., Eggleton, P., van Rensburg, B. J., and Parr, C. L. (2012). The pirodiversity hypothesis: a test with savanna termite assemblages. J. Appl. Ecol. 49, 422–430. doi: 10.1111/j.1365-2664.2012.02107.x
Dean, W. (1997). With Broadax and Firebrand: the Destruction of the Brazilian Atlantic Forest. Oakland: University of California Press.
Dias, B. F. S. (2006). “Degradação ambiental: os impactos do fogo sobre a biodiversidade do cerrado,” in Dimensões Humanas da Biodiversidade: O Desafio de Novas Relações Homem-Natureza No Século XXI, eds I. Garay and B. Becker (Petrópolis, Editora Vozes), 187–213.
Dias, B. F. S., and Miranda, H. S. (2010). “O Projeto Fogo,” in Efeitos do Regime do Fogo Sobre a Estrutura de Comunidades de Cerrado: Resultados do Projeto Fogo, ed H. S. Miranda (Brasília: Ibama), 15–22.
Driscoll, D. A., Lindenmayer, D. B., Bennett, A. F., Bode, M., Bradstock, R. A., Cary, G. J., et al. (2010). Fire management for biodiversity conservation: key research questions and our capacity to answer them. Biol. Conserv. 143, 1928–1939. doi: 10.1016/j.biocon.2010.05.026
Duarte, J. M. B., and Vogliotti, A. (2009). “Ozotoceros bezoarticus (Linnaeus, 1758),” in Fauna Ameaçada de Extinção no Estado de São Paulo – Vertebrados, eds P. M. Bressan, M. C. M. Kierulff, and A. M. Sugieda (São Paulo: Fundação Parque Zoológico de São Paulo: Secretaria do Meio Ambiente), 69.
Durigan, G., Leitão-Filho, H. F., and Rodrigues, R. R. (1994). Phytosociology and structure of a frequently burnt cerrado vegetation in SE-Brazil. Flora 189, 153–160. doi: 10.1016/S0367-2530(17)30582-0
Durigan, G., Pilon, N. A. L., Assis, G. B., Souza, F. M., and Baitello, J. B. (2018). Plantas Pequenas do Cerrado: Biodiversidade Negligenciada. São Paulo: Secretaria do Meio Ambiente.
Durigan, G., and Ratter, J. A. (2016). The need for a consistent fire policy for Cerrado conservation. J. Appl. Ecol. 53, 11–15. doi: 10.1111/1365-2664.12559
Durigan, G., and Ratter, J. A. (2006). Successional changes in cerrado and cerrado/forest ecotonal vegetation in western São Paulo State, Brazil, 1962-2000. Edinb. J. Bot. 63, 119–130. doi: 10.1017/S0960428606000357
Faria, R. G., and Araujo, A. F. B. (2004). Sintopy of two Tropidurus lizard species (Squamata: Tropiduridae) in a rocky cerrado habitat in central Brazil. Braz. J. Biol. 64, 775–786. doi: 10.1590/S1519-69842004000500007
Farnsworth, L. M., Nimmo, D. G., Kelly, L. T., Bennet, A. F., and Clarke, M. F. (2014). Does pyrodiversity beget alpha, beta or gamma diversity? A case study using reptiles from semi-arid Australia. Divers. Distrib. 20, 663–673. doi: 10.1111/ddi.12181
Fidelis, A., Alvarado, S. T., Barradas, A. C. S., and Pivello, V. R. (2018). The year 2017: megafires and management in the cerrado. Fire 1:49. doi: 10.3390/fire1030049
Fidelis, A., Blanco, C. C., Müller, S. C., Pillar, V. D., and Pfadenhauer, J. (2012). Short-term changes caused by fire and mowing in Brazilian Campos grasslands with different long-term fire histories. J. Veg. Sci. 23, 552–562. doi: 10.1111/j.1654-1103.2011.01364.x
Fidelis, A., and Pivello, V. R. (2011). Deve-se usar o fogo como instrumento de manejo no Cerrado e Campos Sulinos? Biodiversidade Bras. 2, 12–25.
Fidelis, A., Rosalem, P., Zanzarini, V., Camargos, L. S., and Martins, A. R. (2019). From ashes to flowers: a savanna sedge initiates flowers 24 h after fire. Ecology 100:e02648. doi: 10.1002/ecy.2648
Firth, R. S., Woinarski, J. C., Brennan, K. G., and Hempel, C. (2006). Environmental relationships of the brush-tailed rabbit-rat, Conilurus penicillatus, and other small mammals on the Tiwi Islands, northern Australia. J. Biogeogr. 33, 1820–1837. doi: 10.1111/j.1365-2699.2006.01543.x
França, H. (2010). Os Incêndios de 2010 nos Parques Nacionais do Cerrado. Technical report. Available online at: http://www.ufabc.edu.br/index.php?option=com_content&view=article&id=4109:professora-da-ufabc-mapeia-queimadas-em-unidades-de-conservacao-do-cerrado&catid=587:2010&Itemid=183 (accessed September 15, 2019).
Freeman, J., Kobziar, L., Rose, E. W., and Cropper, W. (2017). A critique of the historical-fire-regime concept in conservation. Conserv. Biol. 31, 976–985. doi: 10.1111/cobi.12942
Frizzo, T. L., Bonizario, C., Borges, M. P., and Vasconcelos, H. (2011). Uma revisão dos efeitos do fogo sobre a fauna de formações savânicas do Brasil. Oecol. Aust. 15, 365–379. doi: 10.4257/oeco.2011.1502.13
Furley, P. A., Rees, R. M., Ryan, C. M., and Saiz, G. (2008). Savanna burning and the assessment of long-term fire experiments with particular reference to Zimbabwe. Prog. Phys. Geog. 32, 611–634. doi: 10.1177/0309133308101383
Gardner, A. L. (2008) Mammals of South America (Vol. 1) – Marsupials, Xenarthrans, Shrews and Bats. Chicago: The University of Chicago Press.
Gill, M. A. (2012). Fire regimes, biodiversity conservation and prescribed-burning programs. Proc. R. Soc. Vic. 124, 1–6. doi: 10.1071/RS12001
Griffiths, A. D., and Christian, K. A. (1996). The effects of fire on the frillneck lizard (Chlamydosaurus kingii) in northern Australia. Aust. J. Ecol. 21, 386–398. doi: 10.1111/j.1442-9993.1996.tb00625.x
Hansson, S. O. (2017). “The precautionary principle,” in Handbook of Safety Principles, eds N. Moller, J. E. Holmberg, and C. Rollenhagen (Wiley Online Library), 258–283.
Henriques, R., Bizerril, M., and Palma, A. (2000). Changes in small mammal populations after fire in a patch of unburned cerrado in Central Brazil. Mammalia 64, 173–185. doi: 10.1515/mamm.2000.64.2.173
Henriques, R. P. B., Briani, D. C., Palma, A. R. T., and Vieira, E. M. (2006). A simple graphical model of small mammal succession after fire in the Brazilian Cerrado. Mammalia 70, 226–230. doi: 10.1515/MAMM.2006.044
Higgins, S. I., Bond, W. J., February, E. C., Bronn, A., Euston-Brown, D. I. W., Enslin, B., et al. (2007). Effects of four decades of fire manipulation on woody vegetation structure in savanna. Ecology 88, 1119–1125. doi: 10.1890/06-1664
Hoffmann, W. A., Adasme, R., Haridasan, M. T., Carvalho, M., Geiger, E. L., Pereira, M. A., et al. (2009). Tree topkill, not mortality, governs the dynamics of savanna–forest boundaries under frequent fire in central Brazil. Ecology 90, 1326–1337. doi: 10.1890/08-0741.1
Hoffmann, W. A., and Moreira, A. G. (2002). “The role of fire in population dynamics of woody plants,” in The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savanna, eds P. S. Oliveira and R. J. Marquis (New York, NY: Columbia University Press), 159–177.
Honda, E. A., and Durigan, G. (2016). Woody encroachment and its consequences on hydrological processes in the savannah. Phil. Trans. R. Soc. B 371:20150313. doi: 10.1098/rstb.2015.0313
Keeley, J. E., Pausas, J. G., Rundel, P. W., Bond, W. J., and Bradstock, R. A. (2011). Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 16, 406–411. doi: 10.1016/j.tplants.2011.04.002
Koleff, P., Gaston, K. J., and Lennon, J. K. (2003). Measuring betadiversity for presence-absence data. J. Anim. Ecol. 72, 367–382. doi: 10.1046/j.1365-2656.2003.00710.x
Langford, G. J., Borden, J. A., Major, C. S., and Nelson, D. H. (2007). Effects of prescribed fire on the herpetofauna of a southern Mississippi pine savanna. Herpetol. Conserv. Bio. 2, 135–143.
Lawes, M. J., Murphy, B. P., Fisher, A., Woinarski, J. C., Edwards, A. C., and Russell-Smith, J. (2015). Small mammals decline with increasing fire extent in northern Australia: evidence from long-term monitoring in Kakadu National Park. Int. J. Wildland Fire 24, 712–722. doi: 10.1071/WF14163
Layme, V. M. G., Lima, A. P., and Magnusson, W. E. (2004). Effects of fire, food availability and vegetation on the distribution of the rodent Bolomys lasiurus in an Amazonian savanna. J. Trop. Ecol. 20, 183–187. doi: 10.1017/S0266467403001263
Le Stradic, S., Silveira, F. A., Buisson, E., Cazelles, K., Carvalho, V., and Fernandes, G. W. (2015). Diversity of germination strategies and seed dormancy in herbaceous species of campo rupestre grasslands. Ausr. Ecol. 40, 537–546. doi: 10.1111/aec.12221
Lebbink, G., Fensham, R., and Cowley, R. (2018). Vegetation responses to fire history and soil properties in grazed semi-arid tropical savanna. Rangeland J. 40, 271–285. doi: 10.1071/RJ17075
Lemos, F. G., and Azevedo, F. C. (2009). “Lycalopex vetulus (Lund, 1842),” in Fauna Ameaçada de Extinção no Estado de São Paulo – Vertebrados, eds P. M. Bressan, M. C. M. Kierulff, and A. M. Sugieda (São Paulo: Fundação Parque Zoológico de São Paulo, Secretaria do Meio Ambiente), 61.
Lepesqueur, C., Morais, H. C., and Diniz, I. R. (2012). Accidental fire in the cerrado: its impact on communities of caterpillars on two species of Erythroxylum. Psyche 1:101767. doi: 10.1155/2012/101767
Lopes, S. D. F., Vale, V. S. D., and Schiavini, I. (2009). Efeito de queimadas sobre a estrutura e composição da comunidade vegetal lenhosa do cerrado sentido restrito em Caldas Novas, GO. Rev. Árvore 33, 695–704. doi: 10.1590/S0100-67622009000400012
Lucindo, A. S., Antunes, A. Z., Kanashiro, M. M., and Dias, M. M. (2015). Birds at santa bárbara ecological station, one of the last cerrado remnants in the state of São Paulo, Brazil. Biota Neotrop. 15:e0155. doi: 10.1590/1676-06032015015514
Maravalhas, J., and Vasconcelos, H. L. (2014). Revisiting the pyrodiversity–biodiversity hypothesis: long-term fire regimes and the structure of ant communities in a Neotropical savanna hotspot. J. Appl.Ecol. 51, 1661–1668. doi: 10.1111/1365-2664.12338
Marinho-Filho, J., Rodrigues, F. H. G., and Juarez, K. M. (2002). “The Cerrado mammals: diversity, ecology, and natural history,” in The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savanna, eds P. S. Oliveira and R. J. Marquis (New York, NY: Columbia University Press), 266–286.
McCleery, R., Monadjem, A., Baiser, B., Fletcher, R., Vickers, K., and Kruger, L. (2018). Animal diversity declines with broad-scale homogenization of canopy cover in African savannas. Biol. Conserv. 226, 54–62. doi: 10.1016/j.biocon.2018.07.020
Medeiros, M. B., and Miranda, H. S. (2005). Mortalidade pós-fogo em espécies lenhosas de campo sujo submetido a três queimadas prescritas anuais. Acta Bot. Bras. 19, 493–500. doi: 10.1590/S0102-33062005000300009
Mendonça, A. F., Armond, T., Camargo, A. C. L., Camargo, N. F., Ribeiro, J. F., Zangrandi, P. L., et al. (2015a). Effects of an extensive fire on arboreal small mammal populations in a neotropical savanna woodland. J. Mammal 96, 368–379. doi: 10.1093/jmammal/gyv038
Mendonça, A. H., Russo, C., Melo, A. C., and Durigan, G. (2015b). Edge effects in savanna fragments: a case study in the cerrado. Plant Ecol. Divers. 8, 493–503. doi: 10.1080/17550874.2015.1014068
Mills, M. S. L. (2004). Bird community responses to savanna fires: should managers be concerned? S. Afr. J. Wildl. Res. 34, 1–11.
Morais, A. R., Côrtes, L. G., and Bastos, R. P. (2011). Queimadas podem alterar as assembléias de anuros? O caso das veredas na estação ecológica serra geral do tocantins. Rev. Biol. Neotrop. J. Neotrop. Biol. 8, 32–39. doi: 10.5216/rbn.v8i2.16856
Morais, H. C., and Benson, W. W. (1988). Recolonização de vegetação de Cerrado após queimada, por formigas arborícolas. Rev. Bras. Biol. 48, 459–466.
Moreira, A. G. (2000). Effects of fire protection on savanna structure in Central Brazil. J. Biogeogr. 27, 1021–1029. doi: 10.1046/j.1365-2699.2000.00422.x
Morgan, J. W. (1998). Importance of canopy gaps for recruitment of some forbs in Themeda triandra-dominated grasslands in south-eastern Australia. Aust. J. Bot. 46, 609–627. doi: 10.1071/BT97057
Motta-Júnior, J. C., Granzinolli, M. A. M., and Develey, P. F. (2008). Aves da Estação Ecológica de Itirapina, estado de São Paulo, Brasil. Biota Neotrop. 8, 207–227. doi: 10.1590/S1676-06032008000300019
Nelson, A., and Chomitz, K. M. (2011). Effectiveness of strict vs. multiple use protected areas in reducing tropical forest fires: a global analysis using matching methods. PLoS ONE 6:e22722. doi: 10.1371/journal.pone.0022722
Nogueira, C., Colli, G. R., and Martins, M. (2009). Local richness and distribution of the lizard fauna in natural habitat mosaics of the Brazilian Cerrado. Austral Ecol. 34, 83–96. doi: 10.1111/j.1442-9993.2008.01887.x
Nogueira, C., Ribeiro, S., Costa, G. C., and Colli, G. R. (2011). Vicariance and endemism in a Neotropical savanna hotspot: distribution patterns of Cerrado squamate reptiles. J. Biogeogr. 38, 1907–1922. doi: 10.1111/j.1365-2699.2011.02538.x
Oliveira, P. T. S., Leite, M. B., Mattos, T., Nearing, M. A., Scott, R. L., Xavier, R. O., et al. (2017). Groundwater recharge decrease with increased vegetation density in the Brazilian cerrado. Ecohydrology 10:e1759. doi: 10.1002/eco.1759
Oliveras, I., Meirelles, S. T., Hirakuri, V. L., Freitas, C. R., Miranda, H. S., and Pivello, V. R. (2013). Effects of fire regimes on herbaceous biomass and nutrient dynamics in the Brazilian savanna. Int. J. Wildland Fire 22, 368–380. doi: 10.1071/WF10136
Overbeck, G. E., Müller, S. C., Pillar, V. D., and Pfadenhauer, J. (2005). Fine-scale post-fire dynamics in southern Brazilian subtropical grassland. J. Veg. Sci. 16, 655–664. doi: 10.1111/j.1654-1103.2005.tb02408.x
Parr, C. L., and Andersen, A. N. (2006). Patch mosaic burning for biodiversity conservation: a critique of the pyrodiversity paradigm. Conserv. Biol. 20, 1610–1619. doi: 10.1111/j.1523-1739.2006.00492.x
Patton, J. L., Pardiñas, U. F. J., and D'Elía, G. (2015). Mammals of South America (Vol. 2) – Rodents. Chicago: The University of Chicago Press.
Paveglio, T., Norton, T., and Carrol, M. S. (2011). Fanning the flames? Media coverage during wildfire events and its relation to broader societal understandings of the hazard. Hum. Ecol. Rev. 18, 41–52.
Percequillo, A. R., and Kierulff, M. C. M. (2009). “Mamíferos, Introdução,” in Fauna Ameaçada de Extinção no estado de São Paulo: vertebrados, eds P. M. Bressan, M. C. M. Kierulff, and A. M. Sugieda (São Paulo: Fundação Parque Zoológico de São Paulo, Secretaria do Meio Ambiente), 33–35.
Perlin, J. (2005). A Forest Journey: The Story of Wood and Civilization. New York, NY: The Countryman Press.
Piacentini, V. Q., Aleixo, A., Agne, C. E., Maurício, G. N., Pacheco, J. F., Bravo, G. A., et al. (2015). Annotated checklist of the birds of Brazil by the Brazilian Ornithological Records Committee/Lista comentada das aves do Brasil pelo Comitê Brasileiro de Registros Ornitológicos. Rev. Bras. Ornitol. 23, 91–298. doi: 10.1007/BF03544294
Pianka, E. R., and Vitt, L. J. (2003). Lizards: Windows to the Evolution of Diversity, Vol. 5. Oakland: University of California Press.
Pilon, N. A., Hoffmann, W. A., Abreu, R. C., and Durigan, G. (2018). Quantifying the short-term flowering after fire in some plant communities of a cerrado grassland. Plant Ecol. Divers. 11, 259–266. doi: 10.1080/17550874.2018.1517396
Pinheiro, L. F. S., Kolb, R. M., and Rossatto, D. R. (2016). Changes in irradiance and soil properties explain why typical non-arboreal savanna species disappear under tree encroachment. Aust. J. Bot. 64, 333–341. doi: 10.1071/BT15283
Pivello, V. R. (2011). The use of fire in Brazil: past and present. Fire Ecol. 7, 24–39. doi: 10.4996/fireecology.0701024
Pivello, V. R. (2017). “Fire management for biological conservation in the Brazilian cerrado,” in Savannas and Dry Forests: Linking People with Nature, eds J. Mistry and A. Berardi (Abingdon-on-Thames: Routledge), 141–166.
Prada, M., and Marinho-Filho, J. (2004). Effects of fire on the abundance of Xenarthrans in Mato Grosso, Brazil. Aust. Ecol. 29, 568–573. doi: 10.1111/j.1442-9993.2004.01391.x
Prada, M., Marini-Filho, O. J., and Price, P. W. (1995). Insects in flower heads of Aspilia foliacea (Asteraceae) after a fire in a central Brazilian savanna: evidence for the plant vigor hypothesis. Biotropica 27, 513–518. doi: 10.2307/2388965
Reis, M. G., Fieker, C. Z., and Dias, M. M. (2016). The influence of fire on the assemblage structure of foraging birds in grasslands of the Serra da Canastra National Park, Brazil. An. Acad. Bras. Ciênc. 88, 891–901. doi: 10.1590/0001-3765201620150177
Rissi, M. N., Baeza, M. J., Gorgone-Barbosa, E., Zupo, T., and Fidelis, A. (2017). Does season affect fire behaviour in the Cerrado? Int. J. Wildland Fire 26, 427–433. doi: 10.1071/WF14210
Rosan, T. M., Aragão, L. E., Oliveras, I., Phillips, O. L., Malhi, Y., Gloor, E., et al. (2019). Extensive twenty-first century woody encroachment in South America's savanna. Geophys. Res. Lett. 6594–6603. doi: 10.1029/2019GL082327
Russell-Smith, J., Monagle, C., Jacobsohn, M., Beatty, R. L., Bilbao, B., Millán, A., et al. (2017). Can savanna burning projects deliver measurable greenhouse emissions reductions and sustainable livelihood opportunities in fire-prone settings? Clim. Change 140, 47–61. doi: 10.1007/s10584-013-0910-5
Santos, A. J. B., Silva, G. T. D. A., Miranda, H. S., Miranda, A. C., and Lloyd, J. (2003). Effects of fire on surface carbon, energy and water vapour fluxes over campo sujo savanna in central Brazil. Funct. Ecol. 17, 711–719. doi: 10.1111/j.1365-2435.2003.00790.x
Santos-Filho, M., Frieiro-Costa, F., Ignácio, A. R. A., and Silva, M. N. F. (2012). Use of habitats by non-volant small mammals in Cerrado in Central Brazil. Braz. J. Biol. 72, 893–902. doi: 10.1590/S1519-69842012000500016
São Paulo (2018). Lista das Espécies Ameaçadas de Extinção da Fauna do Estado de São Paulo. São Paulo: Secretaria de Estado do Meio Ambiente.
Sato, M. N., and Miranda, H. S. (1996). “Mortalidade de plantas lenhosas do cerrado sensu stricto submetidas a diferentes regimes de queima,” in Impactos de Queimadas em Áreas de Cerrado e Restinga, eds. H. S. Miranda, C. H. Saito, and B. F. S. Dias (Brasília: ECL/Universidade de Brasília), 102–111.
Schmidt, I. B., Moura, L. C., Ferreira, M. C., Eloy, L., Sampaio, A. B., Dias, P. A., et al. (2018). Fire management in the Brazilian savanna: first steps and the way forward. J. Appl. Ecol. 55, 2094–2101. doi: 10.1111/1365-2664.13118
Scott, A. C. (2000). The pre-quaternary history of fire. Palaeogeogr. Palaeocl. 164, 281–329. doi: 10.1016/S0031-0182(00)00192-9
Sheuyange, A., Oba, G., and Weladji, R. B. (2005). Effects of anthropogenic fire history on savanna vegetation in northeastern Namibia. J. Environ. Manage. 75, 189–198. doi: 10.1016/j.jenvman.2004.11.004
Silva, J. M. C., and Bates, J. M. (2002). Biogeographic patterns and conservation in the South American Cerrado: a tropical savanna hotspot. BioScience 52, 225–233. doi: 10.1641/0006-3568(2002)052[0225:BPACIT]2.0.CO;2
Silveira, L., Rodrigues, F. H. G., Jacomo, A. T. A., and Diniz Filho, J. A. F. (1999). Impact of wildfires on the megafauna of Emas National Park, central Brazil. Oryx 33, 108–114. doi: 10.1046/j.1365-3008.1999.00039.x
Simon, M. F., Grether, R., Queiroz, L. P., Skema, C., Pennington, R. T., and Hughes, C. E. (2009). Recent assembly of the cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. PNAS 106, 20.359–20.364. doi: 10.1073/pnas.0903410106
Simon, M. F., and Pennington, T. (2012). Evidence for adaptation to fire regimes in the tropical savannas of the Brazilian Cerrado. Int. J. Plant Sci. 173, 711–723. doi: 10.1086/665973
Sparks, J. C., Masters, R. E., Engle, D. M., Palmer, M. W., and Bukenhofer, G. A. (1998). Effects of late growing-season and late dormant-season prescribed fire on herbaceous vegetation in restored pine-grassland communities. J. Veg. Sci. 9, 133–142. doi: 10.2307/3237231
Tilman, D., Reich, P., Phillips, H., Menton, M., Patel, A., Vos, E., et al. (2000). Fire suppression and ecosystem carbon storage. Ecology 81, 2680–2685. doi: 10.1890/0012-9658(2000)081[2680:FSAECS]2.0.CO;2
van Wilgen, B. W. (2009). The evolution of fire management practices in savanna protected areas in South Africa. S. Afr. J. Sci. 105, 343–349. doi: 10.4102/sajs.v105i9/10.107
van Wilgen, B. W., Govender, N., and Biggs, H. C. (2007). The contribution of fire research to fire management: a critical review of a long-term experiment in the Kruger National Park, South Africa. Int. J. Wildland Fire 16, 519–530. doi: 10.1071/WF06115
Vasconcelos, H. L., Maravalhas, J. B., and Cornelissen, T. (2017). Effects of fire disturbance on ant abundance and diversity: a global meta-analysis. Biodivers. Conserv. 26, 177–188. doi: 10.1007/s10531-016-1234-3
Vasconcelos, H. L., Maravalhas, J. B., Feitosa, R. M., Pacheco, R., Neves, K. C., and Andersen, A. N. (2018). Neotropical savanna ants show a reversed latitudinal gradient of species richness, with climatic drivers reflecting the forest origin of the fauna. J. Biogeogr. 45, 248–258. doi: 10.1111/jbi.13113
Veldman, J. W., Overbeck, G. E., Negreiros, D., Mahy, G., Le Stradic, S., Fernandes, G. W., et al. (2015a). Where tree planting and forest expansion are bad for biodiversity and ecosystem services. BioScience 65, 1011–1018. doi: 10.1093/biosci/biv118
Veldman, J. W., Overbeck, G. E., Negreiros, D., Mahy, G., Le Stradic, S., Fernandes, G. W., et al. (2015b). Tyranny of trees in grassy biomes. Science 347, 484–485. doi: 10.1126/science.347.6221.484-c
Vieira, E. M. (1999). Small mammal communities and fire in the Brazilian Cerrado. J. Zool. 249, 75–81. doi: 10.1111/j.1469-7998.1999.tb01061.x
Vieira, E. M., Andrade, I., and Price, P. W. (1996). Fire effects on a Palicourea rigida (Rubiaceae) gall midge: a test of the plant vigor hypothesis. Biotropica 28, 210–217. doi: 10.2307/2389075
Vieira, E. M., and Briani, D. C. (2013). Short-term effects of fire on small rodents in the Brazilian Cerrado and their relation with feeding habits. Int. J. Wildland Fire 22, 1063–1071. doi: 10.1071/WF12153
Vieira, E. M., and Marinho-Filho, J. (1998). Pre- and postfire habitat utilization by rodents of Cerrado from Central Brazil. Biotropica 30, 491–496. doi: 10.1111/j.1744-7429.1998.tb00086.x
Vieira, M. V. (2003). Seasonal niche dynamics in coexisting rodents of the Brazilian cerrado. Stud. Neotrop. Fauna Environ. 38, 7–15. doi: 10.1076/snfe.38.1.7.14034
Voss, R. S., and Jansa, S. A. (2009). Phylogenetic relationships and classification of didelphid marsupials, an extant radiation of New World metatherian mammals. B. Am. Mus. Nat. Hist. 322, 1–77. doi: 10.1206/322.1
Woinarski, J. C., Armstrong, M., Brennan, K., Fisher, A., Griffiths, A. D., Hill, B., et al. (2010). Monitoring indicates rapid and severe decline of native small mammals in Kakadu National Park, northern Australia. Wildlife Res. 37, 116–126. doi: 10.1071/WR09125
Woinarski, J. C. Z., and Legge, S. (2013). The impacts of fire on birds in Australia's tropical savannas. EMU 113, 319–352. doi: 10.1071/MU12109
Yessoufou, K. (2017). Potential carbon stock in the Kruger National Park, South Africa. Bot. Lett. 164, 425–432. doi: 10.1080/23818107.2017.1383306
Keywords: cerrado, conservation, fauna, fire impact, fire management, plant functional groups, prescribed burn
Citation: Durigan G, Pilon NAL, Abreu RCR, Hoffmann WA, Martins M, Fiorillo BF, Antunes AZ, Carmignotto AP, Maravalhas JB, Vieira J and Vasconcelos HL (2020) No Net Loss of Species Diversity After Prescribed Fires in the Brazilian Savanna. Front. For. Glob. Change 3:13. doi: 10.3389/ffgc.2020.00013
Received: 24 October 2019; Accepted: 30 January 2020;
Published: 19 February 2020.
Edited by:
Imma Oliveras, University of Oxford, United KingdomReviewed by:
Guarino Rinaldi Colli, University of Brasilia, BrazilCopyright © 2020 Durigan, Pilon, Abreu, Hoffmann, Martins, Fiorillo, Antunes, Carmignotto, Maravalhas, Vieira and Vasconcelos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giselda Durigan, Z2lzZWxkYS5kdXJpZ2FuQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.