
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change, 06 June 2019
Sec. Tropical Forests
Volume 2 - 2019 | https://doi.org/10.3389/ffgc.2019.00029
This article is part of the Research TopicImproving Environmental Sustainability in Oil Palm: Linking Science, Policy, & Practice across the TropicsView all 18 articles
 Christopher R. Woodham1,2*†
Christopher R. Woodham1,2*† Anak Agung Ketut Aryawan3†Sarah H. Luke4Paul Manning1,5Jean-Pierre Caliman3Mohd Naim3
Anak Agung Ketut Aryawan3†Sarah H. Luke4Paul Manning1,5Jean-Pierre Caliman3Mohd Naim3 Edgar C. Turner4
Edgar C. Turner4 Eleanor M. Slade1,6
Eleanor M. Slade1,6Oil palm plantations are a major agricultural land use in Southeast Asia. In the coming decades large areas of mature oil palm will be cleared and replanted. To inform more sustainable long-term production in this globally important crop, it is crucial we understand how replanting impacts ecosystem functions and services. We investigated whether several production-relevant ecosystems functions (dung removal, soil mesofauna feeding activity, herbivory, herbivore predation, and seed predation), and the simultaneous delivery of all functions (ecosystem multifunctionality), vary between recently-replanted oil palm (1–4 years) and mature oil palm (23–30 years) areas. Following new in-country and Roundtable on Sustainable Palm Oil (RSPO) guidelines, riparian buffers of mature oil palm, in which subsequent natural regrowth is allowed, are being preserved during the replanting cycle in plantations that lack natural forest reserves. We investigated whether or not mature oil palm riparian buffers maintain levels of ecosystem functioning beneficial for palm oil production. Only one function (herbivory) differed between mature and replanted areas, with higher levels of herbivory found in recently replanted oil palm. There was no difference in ecosystem multifunctionality between mature and recently-replanted oil palm. Mature oil palm riparian buffers were found to be valuable for maintaining lower levels of herbivory than recently-replanted oil palm. However, no other functions, nor ecosystem multifunctionality, differed between the mature oil palm riparian buffers and recently-replanted oil palm. The results of this study suggest that replanting has limited impacts on the ecosystem functions we considered. Furthermore, they suggest mature oil palm riparian buffers do not have negative impacts on production-relevant ecosystem functions in oil palm landscapes.
Oil palm plantations are a major agricultural land use in Southeast Asia (Foster et al., 2011; Food Agricultural Organization of the United Nations, 2017). Malaysia and Indonesia are the largest palm oil producing countries globally (Food Agricultural Organization of the United Nations, 2017; Meijaard et al., 2018), with oil palm being of great socioeconomic importance in these countries (Turner et al., 2008; Foster et al., 2011). However, the expansion of oil palm in Southeast Asia has contributed toward the widespread and large-scale conversion of forested land and has been a major factor driving biodiversity loss (Edwards et al., 2010; Meijaard et al., 2018).
Large scale oil palm planting began in the region in the mid-1980s and large areas of mature oil palm plantation are due to be cleared and replanted in the coming decade as the palms reach the end of their productive lifespan (Snaddon et al., 2013). In Malaysia alone, over 1 million ha are expected to be cleared and replanted in the coming years (Snaddon et al., 2013). The clearing of mature oil palm for replanting, which is achieved through clear-felling, leads to a loss of vertical vegetation structure and canopy cover, and consequently a hotter, drier and more variable microclimate (Luskin and Potts, 2011). Replanted areas of oil palm have been found to contain lower frog species richness than mature oil palm (Kurz et al., 2016), while other studies have demonstrated that the composition of bird and ant communities vary with oil palm age (Desmier De Chenon and Susanto, 2006; Wang and Foster, 2016).
A potential strategy for maintaining structure and habitat complexity within oil palm plantations is the preservation and restoration of riparian buffers—strips of non-production habitat present along waterways in cultivated areas. Riparian buffers are a legal requirement in many Southeast Asian countries, and are a requirement of Roundtable on Sustainable Palm Oil (RSPO) and Indonesian Sustainable Palm Oil (ISPO) certified plantations (Barclay et al., 2017; Roundtable on Sustainable Palm Oil (RSPO), 2018). There are beneficial hydrological impacts of riparian buffers (Tabacchi et al., 2000; Mayer et al., 2007) and forested riparian reserves have also been shown to be effective for conserving biodiversity associated with tropical forests (Gray et al., 2014, 2015; de la Peña-Cuéllar et al., 2015; Luke et al., 2019).
All research to date has focused on riparian buffers that are fragments of natural forest. However, many plantations—including many that were planted before the advent of sustainability guidelines—lack forested riparian buffers, or have forested buffers that are highly degraded. The replanting of oil palm provides an opportunity for the creation and restoration of riparian buffers where they are currently absent or degraded. In these cases, RSPO guidelines suggest two major approaches to restoring mature oil palm riparian buffers: (1) the passive approach—in which natural regrowth occurs without any intervention, and (2) the active approach—enrichment planting of tree species within the buffers (Barclay et al., 2017; Lucey et al., 2018). In the first of these options, mature palms are commonly retained in riparian areas after replanting in order to provide habitat structure and shading to facilitate passive regrowth. Therefore, mature oil palm riparian buffers—riparian strips of mature oil palm that have not been cleared for replanting—are likely to become a common landscape feature during the widespread replanting of oil palm (Snaddon et al., 2013; Barclay et al., 2017).
An important consideration when quantifying ecosystem functioning is that ecosystems typically provide numerous functions and services, a concept known as ecosystem multifunctionality (Isbell et al., 2011; Maestre et al., 2012; Byrnes et al., 2014; Lefcheck et al., 2015). Conserving the multifunctional nature of agroecosystems is an important component of sustainable agriculture (Renting et al., 2009). Loss of ecosystem multifunctionality caused by management-associated perturbations can consequently lead to loss of agricultural productivity (Bommarco et al., 2013), and the loss of services with important effects for the wider landscape (e.g., water purification and carbon sequestration). Despite this studies that consider the impacts of replanting and riparian buffers on ecosystem functioning are particularly lacking. We know of no studies that consider the effects of oil palm replanting on ecosystem functioning, and the effects of riparian buffers on functions, along with the potential for spillover of beneficial functions into the surrounding plantation, remains poorly understood (Edwards et al., 2014; Gray and Lewis, 2014; Gray et al., 2014, 2016). Therefore, as a first step toward effectively managing the large-scale replanting that will occur in the coming decade, it is important that we improve our understanding of how replanting, and the use of mature oil palm riparian buffers as a potential conservation strategy, affect ecosystem functioning.
The aims of this study were to: (1) assess the impacts of oil palm replanting on ecosystem functions and multifunctionality; and (2) assess the importance of mature oil palm riparian buffers for maintaining ecosystem functions, and multifunctionality, within newly replanted oil palm landscapes. We hypothesized: (1) more beneficial levels of production-relevant ecosystem functions will be maintained in mature oil palm than replanted oil palm; and (2) mature oil palm riparian buffers within replanted areas will maintain more beneficial levels of production-relevant ecosystem functioning than replanted areas without mature oil palm riparian buffers.
Data collection took place in oil palm estates owned and managed by Pt Ivo Mas Tunggal, a subsidiary company of Golden Agri Resources, with technical advice from Sinar Mas Agro Resources and Technology Corporation Research Institute (SMARTRI) in Riau, Sumatra, Indonesia (0°55′56″ N, 101°11′62″ E). This area is dominated by oil palm, human settlements and human infrastructure. There are no extensive areas of natural habitat locally. The nearest large area of forest is over 20 km from the sampling sites. All sampling sites were located adjacent to waterways, separated by at least 1.3 km (Figure S1), and were located within one of three vegetation types: (1) recently replanted oil palm (1–4 years) with young oil palm planted up to the edge of the waterway (hereafter “replanted oil palm”), (2) recently replanted oil palm (1–4 years) with a 50 m wide mature oil palm riparian buffer (hereafter “replanted oil palm with a riparian buffer”); and (3) mature oil palm plantation, with mature oil palm (23–30 years) planted up to the edge of the waterway (hereafter “mature oil palm”) (Figure 1, Table S2). The buffers in this study are being restored under the passive approach. As replanting was fairly recent, the mature palm trees within the buffers were still alive and growing. There were five sampling sites in each vegetation type, giving N = 15 sites in total (Figure S1). At each sampling site plots were located along a transect that ran perpendicular to the waterway (Figure 1), with plots positioned 25, 50, and 500 m into the oil palm. Distance into the oil palm was measured from the edge of the adjacent waterway. The center of each plot was marked by the position of a focal oil palm at the specific distance on the transect. All data was collected within a 5 m radius of the focal oil palm. All data collection took place between 5th August and 1st September 2016.
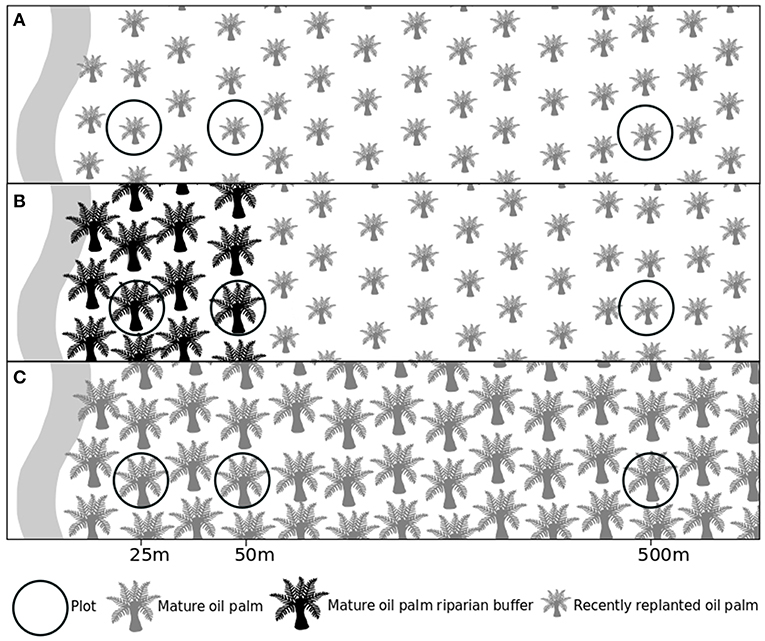
Figure 1. Diagram of sampling sites. Black circles represent plots positioned along a transect running perpendicular to the waterway. (A) Replanted oil palm; (B) Replanted oil palm with a mature oil palm riparian buffer; and (C) Mature oil palm.
Five ecosystem functions relevant to palm oil production were measured: dung removal, soil mesofauna feeding activity, herbivory, herbivore predation, and seed predation (see Table 1 for our justification of the choice of functions).

Table 1. Justification of the ecosystem functions measured and their impacts on ecosystem service provision.
Fresh cattle dung (no more than 1 h old) was collected and homogenized. One dung sample of 700 g was placed in the center of each plot and left in the field for 48 h before the remaining dung was collected and reweighed. Large polystyrene plates raised using wooden skewers protected the dung samples from rain. Three evaporation controls (dung fully enclosed in mesh, so it was not accessible to invertebrates) were placed in each of the vegetation types and reweighed after 48 h. The mass lost through evaporation was then subtracted from the original mass of dung (700 g) to account for moisture changes occurring over the 48-h period. The difference between this evaporation controlled mass and the mass of dung remaining after 48 h allowed an estimation of dung removal (Table S3).
Bait lamina (Terra Protecta GmbH, Berlin, Germany) were used to measure soil mesofauna feeding activity (Kratz, 1998; Hamel et al., 2007). We followed the methods of Tao et al. (2016). Bait lamina are plastic sticks with dimensions: 120 × 1 × 6 mm, and 16 apertures of 1.5 mm per stick, with 5 mm spacing between apertures. The bait lamina were inserted into the ground, perpendicular to the soil surface, such that the top aperture of each bait lamina was just below the soil surface. The apertures of the bait lamina were filled with a standardized bait consisting of cellulose powder, bran flakes and active carbon in the ratio 70:27:3. Six bait lamina were placed within each plot in a 3 × 2 grid, with a 20 cm spacing between each stick. The bait lamina were left for 6 days before being collected. Perforation of the bait in each aperture was recorded by holding the stick up to the light, with partial or complete perforation being recorded as evidence of feeding activity.
Levels of herbivory on oil palm fronds were measured by sampling the focal palm within each plot. We followed the methods of Foster et al. (2014). A middle-aged frond was cut from each of the focal oil palms. This was the 17th frond for mature oil palms and the 9th frond for young palms. Twenty leaflets were then collected at regular intervals from the full length of each of the cut fronds—with 10 leaflets collected from each side of the frond. The leaflets were photographed against a whiteboard. These images were visually analyzed for herbivore damage. This does not include damage by rhinoceros beetles. ImageJ software was then used to determine the area of each leaflet that had been damaged by herbivores and also the total area of each leaflet. Proportion herbivory was calculated by dividing the leaflet area damaged by herbivores by the total leaflet area (Schneider et al., 2012).
Herbivore predation was measured following the methods of Foster et al. (2014). Fresh oil palm fronds were collected and cut into six-leaflet sections, with each six-leaflet frond section having three leaflets on either side of the midrib. Six recently-killed mealworms were subsequently attached to each frond section using a cyanoacrylate adhesive. One frond section was raised into the canopy in each plot using string and left for 24 h before being lowered from the canopy. The number of mealworms removed from the frond was then recorded.
Sunflower seeds were used for measuring seed predation. Ten shelled sunflower seeds were placed on a paper plate in each plot. Large polystyrene plates, raised using wooden skewers, protected the seeds from rain. The seed plates remained in the field for 24 h before the number of seeds removed was recorded. Due to time restrictions, seed predation data were only collected for sites from two vegetation types: mature oil palm and replanted oil palm.
To investigate ecosystem multifunctionality, we used the averaging approach (Maestre et al., 2012; Byrnes et al., 2014). For a discussion of the advantages and limitations of the averaging approach see Byrnes et al. (2014) and Manning et al. (2018). This approach produces a relative multifunctionality index using the mean of standardized ecosystem function values. For standardization, the “desirable” direction of each function must be consistent. First, we determined whether high levels of each ecosystem function were likely to have positive or negative impacts on the provision of ecosystem services valuable to oil palm plantation managers (Table 1). High levels of dung removal, soil mesofauna feeding activity and herbivore predation were considered to be desirable; whereas high levels of herbivory were considered undesirable. Therefore, we transformed the proportion herbivory results by reflecting each value about 0 (multiplying by −1), and then adding the maximum observed proportion herbivory to each of the reflected values. This ensured that the minimum transformed herbivory value was zero, with a high transformed herbivory value being considered beneficial. Seed predation results were not used to calculate multifunctionality, as seed predation data were only collected for two of the three vegetation types. The values for each ecosystem function were standardized by expressing each individual observation as a proportion of the maximum value for that function. The average multifunctionality index was then calculated by taking the mean of the standardized values for the four functions. Multifunctionality was only calculated for plots for which there were data for all four ecosystem functions, resulting in five of the fifty plots being excluded.
To test for differences in levels of ecosystem functioning between mature oil palm and replanted oil palm, we fitted linear models to the data. The model fits were examined and did not display any pattern in the residuals vs. the fitted values, and the normal plots were close to linear. Individual models were produced for each ecosystem function and for the index of multifunctionality. Vegetation type was the explanatory variable in these models. Response variables were proportions and so were arcsine square-root transformed to meet model assumptions. The seed predation data did not meet the model assumptions, even after transformation, and so a Mann–Whitney U-test was used to test for differences in the proportion of seeds removed between mature and replanted oil palm. Observations from each of the different distances within a site were pooled for this analysis as at a large spatial scale both habitats (mature oil palm and replanted oil palm) are homogenous with distance into the oil palm.
To investigate whether ecosystem functions, and multifunctionality, are maintained within mature oil palm riparian buffers, we looked for interactions between distance into oil palm and the three vegetation types (mature oil palm, replanted oil palm with a riparian buffer, and replanted oil palm). If mature oil palm riparian buffers are maintaining ecosystem functioning then we expect there to be an interaction between distance into oil palm and vegetation type. Linear mixed effects models were fitted to the data, with distance into oil palm and vegetation type included as fixed effects and sampling site included as a random effect on the model intercept. For each function, and for our index of multifunctionality, model selection was carried out using Akaike Information Criterion (AIC) values. When one model had an AIC value more than two units lower than all other models, this was selected as the best model (Mazerolle, 2006; Symonds and Moussalli, 2011). If multiple models had AIC values within two units of each other the simplest model (the model with the fewest terms) was selected as the best model (Mazerolle, 2006; Symonds and Moussalli, 2011). We calculated fixed-effects R2 and total R2 values for each of the models using the “r.squaredGLMM” function (Nakagawa and Schielzeth, 2013; Johnson, 2014) from the package MuMIn (Barton, 2018).
All analyses were carried out in R version 3.5.1 (R Core Team, 2018) and R studio version 1.1.456 (RStudio Team, 2015), using the packages lme4 (Bates et al., 2015), MASS (Venables and Ripley, 2002), MuMIn (Barton, 2018), and dplyr (Wickham and Francois, 2015). The packages ggplot2 (Wickham, 2009) and cowplot (Wilke, 2016) were used for producing figures.
Of the five ecosystem functions measured, only herbivory showed significant differences between mature oil palm and replanted oil palm, with significantly higher levels of herbivory found in replanted oil palm (Figure 2, Table 2). The observed mean proportion herbivory was over 3.5 times higher in replanted oil palm, although the absolute values of herbivory were very small [Replanted 0.00659 (SE = 0.00166), Mature 0.00175 (SE = 0.000441), Figure 2C]. There was no significant difference in average multifunctionality between mature oil palm and replanted oil palm (Figure 2F, Table 2).
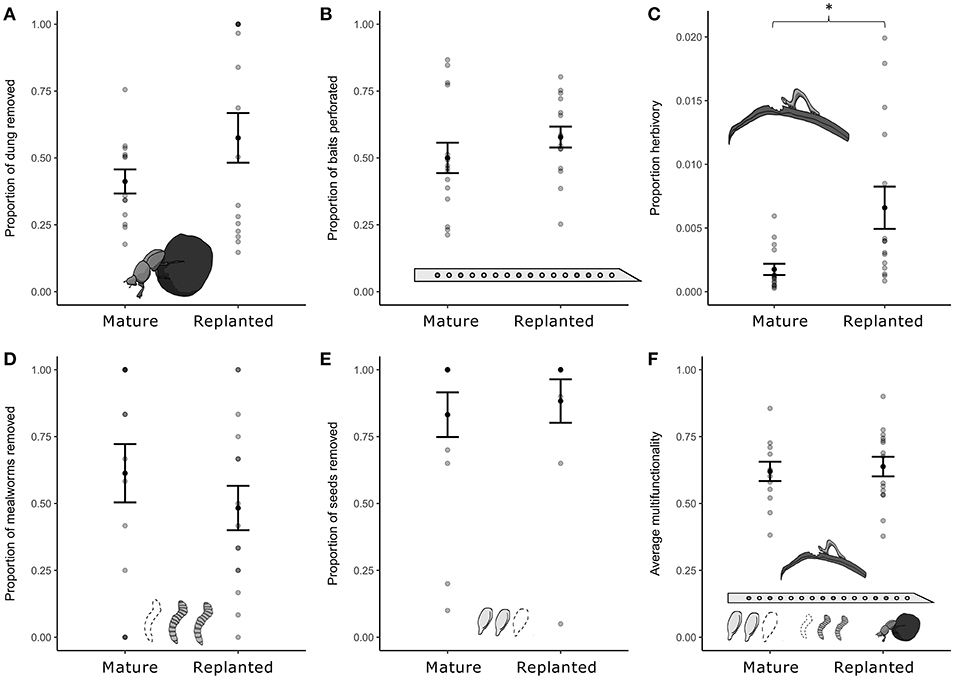
Figure 2. The effect of oil palm replanting on (A) dung removal; (B) soil mesofauna feeding activity; (C) herbivory; (D) herbivore predation; (E) seed predation; and (F) average multifunctionality. Plots show mean ± SE. Mature denotes mature oil palm, and Replanted denotes replanted oil palm. Stars denote significant differences (*P < 0.05). Note the scale on the y-axis of plot (C) ranges from 0 to 0.020.
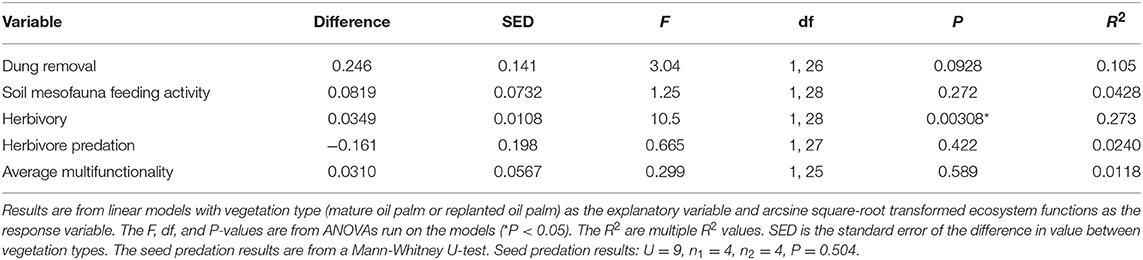
Table 2. Effects of oil palm replanting on ecosystem functions.
The best model for herbivory included the interaction between distance into oil palm and vegetation type (Tables 3, 4). The interaction explained 28.4% of the variation in mean proportion herbivory (Table 3). Mean proportion herbivory was approximately constant with distance in mature oil palm and replanted oil palm (Figures 3A,C). Whereas, in replanted oil palm with a riparian buffer there was an increase in herbivory with distance into oil palm (Figure 3B). There are two data points on Figure 3B that are more than double the value of the other data points at their distance. If these data points are considered outliers and removed from the analysis, the best model for herbivory still includes the interaction between distance into oil palm and vegetation type (Tables S5, S6). This strongly suggests that these two potential outliers are not driving the results of our model selection process.
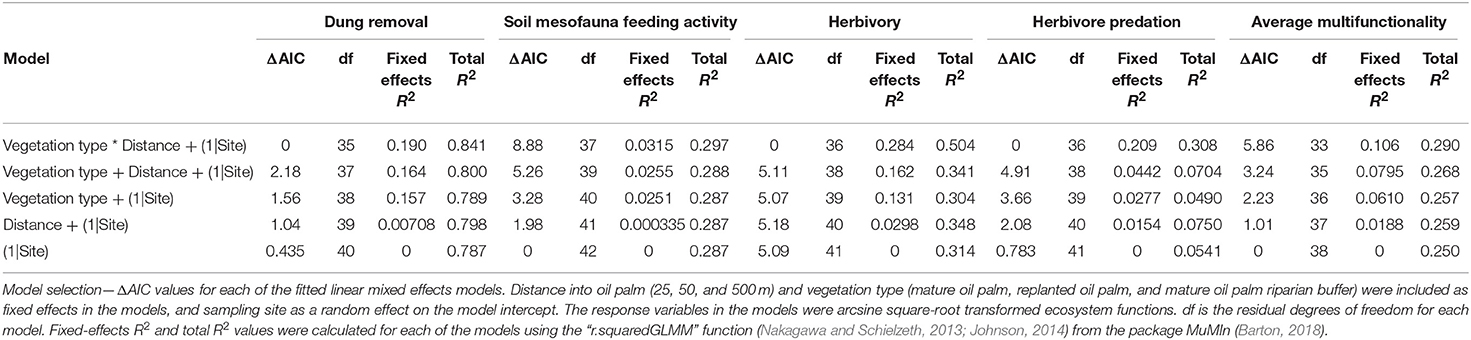
Table 3. Effects of vegetation type and distance into oil palm on individual functions and average multifunctionality.
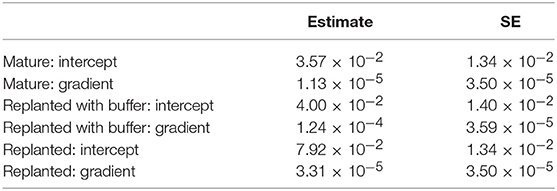
Table 4. Model coefficients for the best herbivory model: Transformed_Herbivory ~ Vegetation Type * Distance + (1|Site).
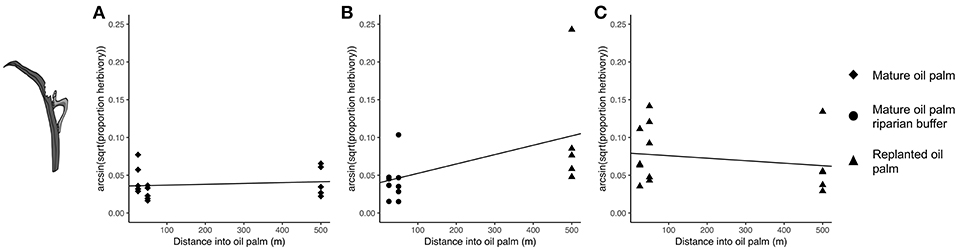
Figure 3. Plot of the best-fitting model for herbivory: Transformed_Herbivory ~ Vegetation Type * Distance + (1|Site). The herbivory data has been arcsine square-root transformed. Point shape denotes vegetation type. (A) Mature oil palm; (B) Replanted oil palm with a mature oil palm riparian buffer; and (C) Replanted oil palm.
In contrast, for dung removal, herbivore predation, soil mesofauna feeding activity and average multifunctionality the best model was the random effect only model (Table 3). Therefore, we found no evidence for effects of vegetation type, distance into oil palm or their interaction on these functions or average multifunctionality (Table 3).
This study found that recently replanted oil palm (1–4 years) had similar levels of ecosystem functioning to mature oil palm (23–30 years), although levels of herbivory were higher in recently replanted oil palm. Mature oil palm riparian buffers were found to have lower levels of herbivory than the surrounding replanted oil palm. However, for all other ecosystem functions measured, there was no difference in functioning between the riparian buffers and the surrounding replanted oil palm.
The replanting of oil palm had no effect on four of the five ecosystem functions measured, nor did it affect ecosystem multifunctionality. The only ecosystem function affected by oil palm replanting was herbivory, with levels of herbivory over 3.5 times higher in replanted oil palm than in mature oil palm. However, while the relative difference in levels of herbivory was large, the absolute difference was much smaller. Mean proportion herbivory was <0.01 in both mature and replanted oil palm. Thus, herbivory is unlikely to have a short-term impact on oil palm yield (Corley and Tinker, 2016; Denmead et al., 2017), particularly as the rate of photosynthesis does not usually limit growth of young palms, even when leaf area is reduced due to herbivory (Corley and Tinker, 2016). However, the slightly higher levels of herbivory in replanted oil palm could be indicative of higher abundances of herbivorous pests or the loss of predator assemblages in these replanted areas, which could mean that future pest population outbreaks are more likely. When they occur, pest outbreaks cause large yield reductions with large financial costs for oil palm estates (Wood et al., 1973; Corley and Tinker, 2016). Therefore, the potential long-term effects of higher levels of herbivory in replanted oil palm should be actively assessed as part of plantation management strategies.
The results of this study suggest that the replanting of oil palm has only limited effects on the ecosystem functions we considered here, despite clear impacts of replanting on the biodiversity and abiotic conditions of these landscapes (Luskin and Potts, 2011; Kurz et al., 2016). The recently replanted oil palms in our study area were between 1 and 4 years of age (Table S2). Thus, even if there were negative impacts on the ecosystem functions measured immediately after replanting, our results show that ecosystem functioning rapidly recovered. Our results therefore suggest that oil palm ecosystems may be highly resilient to perturbations and that any initial negative impacts of replanting are unlikely to have mid- to long-terms effects on the functions we consider here. The dominance of generalist species in oil palm ecosystems (Gillespie et al., 2012; Fayle et al., 2015; Bukhary et al., 2017) may have a major role in this resilience we have identified, as previous studies have found generalists to be of critical importance for the resilience of ecosystem functioning (Richmond et al., 2005; Palacio et al., 2016).
Mature oil palm riparian buffers have different structural and microclimatic conditions compared to replanted areas and therefore potentially have different levels of ecosystem functions compared to the surrounding plantation. They also have the potential to act as sources of, positive or negative, spillover effects for neighboring areas of oil palm. When considering the value of mature oil palm riparian buffers for conserving ecosystem functioning we must keep in mind that these buffers are distinct from mature oil palm itself. Natural regrowth is permitted in these riparian buffers and on top of this no herbicides, pesticides or fertilizers are applied within the buffers. As a result, the understorey vegetation of these riparian buffers is typically much denser than that in mature oil palm plantations (Figure S4).
Mature oil palm riparian buffers showed reduced levels of herbivory compared to the neighboring replanted oil palm. For herbivory, both the mean proportion herbivory and the variability in mean proportion herbivory were much lower in the mature oil palm riparian buffers than in the neighboring replanted oil palm (although it must be noted that the absolute difference in mean proportion herbivory was small). Management of pest populations in oil palm plantations is of central importance to the industry. Importantly, our results suggest that mature oil palm riparian buffers do not have negative impacts on pest control services in oil palm plantations.
For dung removal, soil mesofauna feeding activity, herbivore predation and average multifunctionality there was no difference in functioning between mature oil palm riparian buffers and the surrounding replanted oil palm. We therefore have no evidence to suggest that mature oil palm riparian buffers conserve beneficial levels of these functions within replanted oil palm landscapes. Crucially, however there is also no evidence that mature oil palm riparian buffers maintain deleterious levels of these functions. This strongly suggests mature oil palm riparian buffers do not have negative spillover effects on ecosystem functioning in neighboring replanted oil palm. This finding is in concordance with findings of studies of spillover effects from natural forest riparian buffers or forest fragments, which have found either no evidence of spillover effects, or evidence of small positive spillover effects (Edwards et al., 2014; Gray and Lewis, 2014; Gray et al., 2014, 2016). However, it must be noted that we only collected data for a subset of the ecosystem functions relevant to oil palm production.
Agroecosystems are the dominant terrestrial ecosystems globally (Ramankutty et al., 2008) and are both major consumers and providers of ecosystem services (Power, 2010). It is therefore crucial that we understand how management practices in agroecosystems affect ecosystem functions, and the supporting and regulating services they underpin. This study has shown that several ecosystem functions in oil palm landscapes are highly resilient to clear felling of old palms, a widely implemented method of replanting (Luskin and Potts, 2011), suggesting that the widespread replanting set to occur in the coming decade may have little impact on this subset of production-relevant ecosystem functions. However, clear-felling large areas of mature oil palm to produce plantations with palms of the same age has been shown to lead to reductions in biodiversity within these agroecosystems (Kurz et al., 2016). Therefore, clearance methods, such as staggered replanting (Luskin and Potts, 2011), that could reduce these biodiversity losses need to be explored.
Current riparian buffer policy in tropical ecosystems has a limited ecological basis due to a lack of available information (see Luke et al., 2019 for a review). The results of this study suggest that mature oil palm riparian buffers do not appear to have negative impacts on the surrounding plantation. These mature oil palm riparian buffers act as the first stage of restoring natural riparian forest in replanted oil palm. Thus, the future growth of trees and understorey plants within these mature oil palm riparian buffers may increase the value of these riparian buffers for conserving ecosystem functioning in the long-term. Importantly, monitoring of the replanted areas and mature oil palm riparian buffers over time is needed to assess how well the buffers establish—with a focus on the impacts of regeneration and the death of the mature palms (an increase in deadwood) on biodiversity and ecosystem functioning.
This study only considered the passive restoration of mature oil palm riparian buffers. Enrichment planting within mature oil palm riparian buffers may be more likely to increase ecosystem functioning. The Riparian Ecosystem Restoration in Tropical Agriculture (RERTA) experiment is investigating the impacts of such enrichment planting on biodiversity and ecosystem functioning (Luke et al. this special issue). Studies, such as RERTA, are needed if we are to improve our understanding of the value mature oil palm riparian buffers have for ecosystem functioning in oil palm landscapes.
CW, AA, SL, and ES designed the study. AA and CW (data collection coordinated by AA and SMARTRI team) collected the data. CW, SL, PM, ET, and ES analyzed the data. CW, SL, PM, ET, and ES wrote the manuscript. AA, JP-C, and MN commented on the manuscript.
AA, J-PC, and MN listed with a SMARTRI affiliation were employed by SMARTRI, a research division of Sinar Mas, at the time of the study. The wider BEFTA Programme is co-funded by Sinar Mas, with matched funds from the Isaac Newton Trust, Cambridge. However, there is an MOU in place that protects the intellectual freedom and data-use for all researchers working on the project. The Programme therefore represents a collaboration between the University of Cambridge and an oil palm company, ensuring that results are readily disseminated to inform best management practices, but maintaining academic independence.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data were collected by, and in collaboration with, Indonesian research staff from SMART Research Institute (SMARTRI). We are extremely grateful to all SMARTRI staff for their invaluable help with the project. We also thank Andrew Hector for his advice on the statistical analysis. We are also grateful to Victoria Pike and Jesse van der Grient and two reviewers for their comments on the manuscript. This project was made possible through funding from SMARTRI; the UK Natural Environment Research Council (NERC), the Heron-Allen Travel Scholarship, Lady Margaret Hall; and the University of Oxford Zoology Department. ES was funded under UK Natural Environment Research Council grant (NE/K016407/1). The BEFTA Programme was funded by The Isaac Newton Trust Cambridge, Golden-Agri Resources, and the UK Natural Environment Research Council grant (NE/P00458X/1). We would like to thank RISTEK for permission for ET and SL to establish the initial BEFTA and RERTA Projects respectively, which seeded this collaboration, and to carry out research in Indonesia (numbers 426/SIP/FRP/SM/XI/2012, 72/EXT/SIP/FRP/SM/IX/2013, 44/EXT/SIP/FRP/SM/IX/2014, 354/SIP/FRP/E5/Dit.KI/X/2016, 66/EXT/SIP/FRP/E5/Dit.KI/IX/2017, and 45/EXT/SIP/FRP/E5/Dit.KI/X/2018).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2019.00029/full#supplementary-material
Azhar, B., Puan, C. L., Aziz, N., Sainuddin, M., Adila, N., Samsuddin, S., et al. (2015). Effects of in situ habitat quality and landscape characteristics in the oil palm agricultural matrix on tropical understory birds, fruit bats and butterflies. Biodivers. Conserv. 24, 3125–3144. doi: 10.1007/s10531-015-1005-6
Barclay, H., Gray, C. L., Luke, S. H., Nainar, A., Snaddon, J. L., and Turner, E. C. (2017). RSPO Manual on Best Management Practices (BMPs) for the Management and Rehabilitation of Riparian Reserves. Available online at: http://www.rspo.org/key-documents/supplementary-materials (accessed May 27, 2019).
Barton, K. (2018). MuMIn:Multi-Model Inference. Available online at: https://cran.r-project.org/package=MuMIn. (accessed May 27, 2019).
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Beynon, S. A., Wainwright, W. A., and Christie, M. (2015). The application of an ecosystem services framework to estimate the economic value of dung beetles to the U.K. cattle industry. Ecol. Entomol. 40, 124–135. doi: 10.1111/een.12240
Bommarco, R., Kleijn, D., and Potts, S. G. (2013). Ecological intensification: harnessing ecosystem services for food security. Trends Ecol. Evol. 28, 230–8. doi: 10.1016/j.tree.2012.10.012
Bukhary, A. A. K., Fauzi, M. M. M., Ruslan, M. Y., Noor Hisham, H., Hazmi, I. R., Abu Hassan, A., et al. (2017). Spatio-temporal dominant omnivorous ant-saproxylic beetle species interactions around epigeal-based microhabitats within different oil palm age stand types. Malayan Nat. J. 69, 19–30.
Byrnes, J. E. K., Gamfeldt, L., Isbell, F., Lefcheck, J. S., Griffin, J. N., Hector, A., et al. (2014). Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods Ecol. Evol. 5, 111–124. doi: 10.1111/2041-210X.12143
Corley, R. H. V., and Tinker, P. B. (2016). The Oil Palm. 5th Edn. Chichester: Wiley Blackwell. doi: 10.1002/9781118953297
de la Peña-Cuéllar, E., Benítez-Malvido, J., Avila-Cabadilla, L. D., Martínez-Ramos, M., and Estrada, A. (2015). Structure and diversity of phyllostomid bat assemblages on riparian corridors in a human-dominated tropical landscape. Ecol. Evol. 5, 903–913. doi: 10.1002/ece3.1375
de Mey, Y., Demont, M., and Diagne, M. (2012). Estimating bird damage to rice in africa: evidence from the Senegal river valley. J. Agric. Econ. 63, 175–200. doi: 10.1111/j.1477-9552.2011.00323.x
Denmead, L. H., Darras, K., Clough, Y., Diaz, P., Grass, I., Hoffmann, M. P., et al. (2017). The role of ants, birds and bats for ecosystem functions and yield in oil palm plantations. Ecology 98, 1945–1956. doi: 10.1002/ecy.1882
Desmier De Chenon, R., and Susanto, A. (2006). Ecological observations on diurnal birds in Indonesian oil palm plantations. J. Oil Palm Res. Special Issue 2006, 122–143.
Doube, B. M., Macqueen, A., and Fay, H. A. C. (1988). Effects of dung fauna on survival and size of buffalo flies (Haematobia Spp.) breeding in the field in South Africa and Australia. J. Appl. Ecol. 25, 523–536. doi: 10.2307/2403841
Edwards, D. P., Hodgson, J. A., Hamer, K. C., Mitchell, S. L., Ahmad, A. H., Cornell, S. J., et al. (2010). Wildlife-friendly oil palm plantations fail to protect biodiversity effectively. Conserv. Lett. 3, 236–242. doi: 10.1111/j.1755-263X.2010.00107.x
Edwards, F. A., Edwards, D. P., Sloan, S., and Hamer, K. C. (2014). Sustainable management in crop monocultures: the impact of retaining forest on oil palm yield. PLoS ONE 9:e91695. doi: 10.1371/journal.pone.0091695
Fayle, T. M., Edwards, D. P., Foster, W. A., Yusah, K. M., and Turner, E. C. (2015). An ant-plant by-product mutualism is robust to selective logging of rain forest and conversion to oil palm plantation. Oecologia 178, 441–450. doi: 10.1007/s00442-014-3208-z
Fincher, G. T. (1975). Effects of dung beetle activity on the number of nematode parasites acquired by grazing cattle. J. Parasitol. 61, 759–762. doi: 10.2307/3279480
Food and Agricultural Organization of the United Nations (2017). FAOSTAT Production Statistics. Rome.
Foster, W. A., Snaddon, J. L., Advento, A. D., Agung, A. A., Barclay, H., Caliman, J. P., et al. (2014). The biodiversity and ecosystem function in tropical agriculture (BEFTA) project. Planter 90, 581–591.
Foster, W. A., Snaddon, J. L., Turner, E. C., Fayle, T. M., Cockerill, T. D., Ellwood, M. D., et al. (2011). Establishing the evidence base for maintaining biodiversity and ecosystem function in the oil palm landscapes of South East Asia. Philos. Trans. Biol. Sci. 366, 3277–3291. doi: 10.1098/rstb.2011.0041
Ghazali, A., Asmah, S., Syafiq, M., Yahya, M. S., Aziz, N., Tan, L. P., et al. (2016). Effects of monoculture and polyculture farming in oil palm smallholdings on terrestrial arthropod diversity. J. Asia Pac. Entomol. 19, 415–421. doi: 10.1016/j.aspen.2016.04.016
Gillespie, G. R., Ahmad, E., Elahan, B., Evans, A., Ancrenaz, M., Goossens, B., et al. (2012). Conservation of amphibians in Borneo: relative value of secondary tropical forest and non-forest habitats. Biol. Conserv. 152, 136–144. doi: 10.1016/j.biocon.2012.03.023
Gray, C. L., and Lewis, O. T. (2014). Do riparian forest fragments provide ecosystem services or disservices in surrounding oil palm plantations? Basic Appl. Ecol. 15, 693–700. doi: 10.1016/j.baae.2014.09.009
Gray, C. L., Lewis, O. T., Chung, A. Y., and Fayle, T. M. (2015). Riparian reserves within oil palm plantations conserve logged forest leaf litter ant communities and maintain associated scavenging rates. J. Appl. Ecol. 52, 31–40. doi: 10.1111/1365-2664.12371
Gray, C. L., Simmons, B. I., Fayle, T. M., Mann, D. J., and Slade, E. M. (2016). Are riparian forest reserves sources of invertebrate biodiversity spillover and associated ecosystem functions in oil palm landscapes? Biol. Conserv. 194, 176–183. doi: 10.1016/j.biocon.2015.12.017
Gray, C. L., Slade, E. M., Mann, D. J., and Lewis, O. T. (2014). Do riparian reserves support dung beetle biodiversity and ecosystem services in oil palm-dominated tropical landscapes? Ecol. Evol. 4, 1049–60. doi: 10.1002/ece3.1003
Hamel, C., Schellenberg, M. P., Hanson, K., and Wang, H. (2007). Evaluation of the “bait-lamina test” to assess soil microfauna feeding activity in mixed grassland. Appl. Soil Ecol. 36, 199–204. doi: 10.1016/j.apsoil.2007.02.004
Isbell, F., Calcagno, V., Hector, A., Connolly, J., Harpole, W. S., Reich, P. B., et al. (2011). High plant diversity is needed to maintain ecosystem services. Nature 477, 199–202. doi: 10.1038/nature10282
Jamian, S., Norhisham, A., Ghazali, A., Zakaria, A., and Azhar, B. (2015). Impacts of two species of predatory Reduviidae on bagworms in oil palm plantations. Insect Sci. doi: 10.1111/1744-7917.12309
Johnson, P. C. (2014). Extension of Nakagawa & Schielzeth's R2GLMM to random slopes models. Methods Ecol. Evol. 5, 944–946. doi: 10.1111/2041-210X.12225
Kamarudin, N., and Wahid, M. B. (2010). Interactions of the bagworm, Pteroma pendula (Lepidoptera: Psychidae), and its natural enemies in an oil palm plantation in Perak. J. Oil Palm Res. 22, 758–764.
Kratz, W. (1998). The bait-lamina test: general aspects, applications and perspectives. Environ. Sci. Pollut. Res. 5, 94–96. doi: 10.1007/BF02986394
Kurz, D. J., Turner, E. C., Aryawan, A. A., Barkley, H. C., Caliman, J. P., Konopik, O., et al. (2016). Replanting reduces frog diversity in oil palm. Biotropica 48, 483–490. doi: 10.1111/btp.12320
Lefcheck, J. S., Byrnes, J. E. K., Isbell, F., Gamfeldt, L., Griffin, J. N., Eisenhauer, N., et al. (2015). Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Nat. Commun. 6:6936. doi: 10.1038/ncomms7936
Linz, G. M., Homan, H. J., Werner, S. J., Hagy, H. M., and Bleier, W. J. (2011). Assessment of bird-management strategies to protect sunflowers. Bioscience 61, 960–970. doi: 10.1525/bio.2011.61.12.6
Lucey, J. M., Barclay, H., Gray, C. L., Luke, S. H., Nainar, A., Turner, E. C., et al. (2018). Simplified Guide: Management and Rehabilitation of Riparian Reserves. Kuala Lumpur: RSPO, RSPO Secretariat Sdn Bhd.
Luke, S. H., Slade, E. M., Gray, C. L., Annammala, K. V., Drewer, J., Williamson, J., et al. (2019). Riparian buffers in tropical agriculture: scientific support, effectiveness and directions for policy. J. Appl. Ecol. 56, 85–92. doi: 10.1111/1365-2664.13280
Luskin, M. S., and Potts, M. D. (2011). Microclimate and habitat heterogeneity through the oil palm lifecycle. Basic Appl. Ecol. 12, 540–551. doi: 10.1016/j.baae.2011.06.004
Maestre, F. T., Castillo-Monroy, A. P., Bowker, M. A., and Ochoa-Hueso, R. (2012). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 100, 317–330. doi: 10.1111/j.1365-2745.2011.01918.x
Manning, P., Slade, E. M., Beynon, S. A., and Lewis, O. T. (2016). Functionally rich dung beetle assemblages are required to provide multiple ecosystem services. Agric. Ecosyst. Environ. 218, 87–94. doi: 10.1016/j.agee.2015.11.007
Manning, P., van der Plas, F., Soliveres, S., Allan, E., Maestre, F. T., Mace, G., et al. (2018). Redefining ecosystem multifunctionality. Nat. Ecol. Evol. 2, 427–436. doi: 10.1038/s41559-017-0461-7
Mariau, D. (2001). The Fauna of Oil Palm and Coconut: Insect and Mite Pests and Their Natural Enemies. Montpellier: CIRAD.
Mayer, P. M., Reynolds, S. K., McCutchen, M. D., and Canfield, T. J. (2007). Meta-analysis of nitrogen removal in riparian buffers. J. Environ. Qual. 36, 1172–80. doi: 10.2134/jeq2006.0462
Mazerolle, M. (2006). Improving data analysis in herpetology: using Akaike's Information Criterion (AIC) to assess the strength of biological hypotheses. Amphib. Reptil. 27, 169–180. doi: 10.1163/156853806777239922
Md. Said, M. F., and Man, N. (2014). Evaluation of target area concentration (TAC) programme in Malaysia's integrated cattle and oil palm farming. J. Food Prod. Mark. 20, 151–163. doi: 10.1080/10454446.2014.921870
E. Meijaard, J. Garcia-Ulloa, D. Sheil, S. A. Wich, K. M. Carlson, and D. (eds.) Juffe-Bignoli. (2018). Oil Palm and Biodiversity. A Situation Analysis by the IUCN Oil Palm Task Force. Gland: IUCN Oil Palm Task Force. doi: 10.2305/IUCN.CH.2018.11.en
Nakagawa, S., and Schielzeth, H. (2013). A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 4, 133–142. doi: 10.1111/j.2041-210x.2012.00261.x
Nichols, E., Spector, S., Louzada, J., Larsen, T., Amezquita, S., and Favila, M. E. (2008). Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol. Conserv. 141, 1461–1474. doi: 10.1016/j.biocon.2008.04.011
Palacio, R. D., Valderrama-Ardila, C., and Kattan, G. H. (2016). Generalist species have a central role in a highly diverse plant–frugivore network. Biotropica 48, 349–355. doi: 10.1111/btp.12290
Power, A. G. (2010). Ecosystem services and agriculture: tradeoffs and synergies. Philos. Trans. R. Soc. B Biol. Sci. 365, 2959–2971. doi: 10.1098/rstb.2010.0143
R Core Team (2018). R: A Language and Environment for Statistical Computing. Available online at: https://www.r-project.org/ (accessed May 27, 2019).
Ramankutty, N., Evan, A. T., Monfreda, C., and Foley, J. A. (2008). Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Global Biogeochem. Cycles 22:GB1003. doi: 10.1029/2007GB002952
Renting, H., Rossing, W. A., Groot, J. C. J., Van der Ploeg, J. D., Laurent, C., Perraud, D., et al. (2009). Exploring multifunctional agriculture. A review of conceptual approaches and prospects for an integrative transitional framework. J. Environ. Manage. 90(Suppl.), S112–S123. doi: 10.1016/j.jenvman.2008.11.014
Richmond, C. E., Breitburg, D. L., and Rose, K. A. (2005). The role of environmental generalist species in ecosystem function. Ecol. Modell. 188, 279–295. doi: 10.1016/j.ecolmodel.2005.03.002
Roundtable on Sustainable Palm Oil (RSPO) (2018). Principles and Criteria for the Production of Sustainable Palm Oil. Kuala Lumpur.
RStudio Team (2015). RStudio: Integrated Development for R. Boston, MA: RStudio Inc. Available online at: http://www.rstudio.com/
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Slade, E., Burhanuddin, M., Caliman, J.-P., Foster, W., Naim, M., Prawirosukarto, S, Snaddon, J., et al. (2014). Can cattle grazing in mature oil palm increase biodiversity and ecosystem service provision? Plant 90, 655–665.
Slade, E. M., Riutta, T., Roslin, T., and Tuomisto, H. L. (2016). The role of dung beetles in reducing greenhouse gas emissions from cattle farming. Sci. Rep. 6:18140. doi: 10.1038/srep18140
Snaddon, J. L., Willis, K. J., and Macdonald, D. W. (2013). Biodiversity: oil-palm replanting raises ecology issues. Nature 502, 170–1. doi: 10.1038/502170d
Symonds, M. R. E., and Moussalli, A. (2011). A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike's information criterion. Behav. Ecol. Sociobiol. 65, 13–21. doi: 10.1007/s00265-010-1037-6
Tabacchi, E., Lambs, L., Guilloy, H., Planty-Tabacchi, A.-M., Muller, E., and Decamps, H. (2000). Impacts of riparian vegetation on hydrological processes. Hydrol. Process. 14, 2959–2976. doi: 10.1002/1099-1085(200011/12)14:16/17< 2959::AID-HYP129>3.0.CO;2-B
Tao, H.-H., Slade, E. M., Willis, K. J., Caliman, J.-P., and Snaddon, J. L. (2016). Effects of soil management practices on soil fauna feeding activity in an Indonesian oil palm plantation. Agric. Ecosyst. Environ. 218, 133–140. doi: 10.1016/j.agee.2015.11.012
Turner, E. C., Snaddon, J. L., Fayle, T. M., and Foster, W. A. (2008). Oil palm research in context: identifying the need for biodiversity assessment. PLoS ONE 3:e1572. doi: 10.1371/journal.pone.0001572
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics With S, 4th Edn. New York, NY: Springer.
Wang, W. Y., and Foster, W. A. (2016). Ground-foraging ant communities vary with oil palm age. Basic Appl. Ecol. 17, 21–32. doi: 10.1016/j.baae.2015.08.009
Wickham, H., and Francois, R. (2015). dplyr: A Grammar of Data Manipulation. R Package Version 0.4.3.
Keywords: ecosystem function, ecosystem multifunctionality, ecosystem service, palm oil, replanting, resilience, riparian reserve, Sumatra
Citation: Woodham CR, Aryawan AAK, Luke SH, Manning P, Caliman J-P, Naim M, Turner EC and Slade EM (2019) Effects of Replanting and Retention of Mature Oil Palm Riparian Buffers on Ecosystem Functioning in Oil Palm Plantations. Front. For. Glob. Change 2:29. doi: 10.3389/ffgc.2019.00029
Received: 14 November 2018; Accepted: 20 May 2019;
Published: 06 June 2019.
Edited by:
Katharine Anne Abernethy, University of Stirling, United KingdomReviewed by:
Clare Duncan, Deakin University, AustraliaCopyright © 2019 Woodham, Aryawan, Luke, Manning, Caliman, Naim, Turner and Slade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christopher R. Woodham, Y2hyaXN0b3BoZXJ3b29kaGFtQG1lLmNvbQ==
†These authors share joint first authors
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.