
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 19 November 2024
Sec. Urban Ecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1457049
This article is part of the Research Topic Urban Biodiversity in the Global South View all 6 articles
 Dickson Anoibi Matthew1*
Dickson Anoibi Matthew1* Samuel Temidayo Osinubi2
Samuel Temidayo Osinubi2 Samuel Tertese Ivande1,3†Soladoye B. Iwajomo1,4
Samuel Tertese Ivande1,3†Soladoye B. Iwajomo1,4 Ulf Ottosson1
Ulf Ottosson1Introduction: The rural-urban gradient serves as a valuable context for investigating the impact of urbanization on biodiversity. While previous studies have demonstrated shifts in bird communities along this gradient, our understanding of the specific impacts of individual urban components such as man-made physical structures and anthropogenic noise along this gradient remains limited, and more so, in Afro-tropical environments.
Method: Employing the point count method, we recorded birds and also determined the levels of anthropogenic noise and physical structures, across fifteen sites along the rural-urban gradient on the Jos Plateau, Nigeria. We then investigated variations in bird communities along the urbanization gradient and assessed the influence of the two urban components -anthropogenic noise and physical structures, on bird populations.
Results: There was a decline in bird abundance and species richness along the urbanization gradient. Similarly, species common to all the urbanization categories (species present at least at one point in rural, suburban, and urban) also exhibited a decrease in abundance. The suburban area showed a greater similarity in bird community composition to the urban area than the rural area. Notably, as the level of urban development increased, numerous bird species associated with undisturbed sites gradually vanished. Both anthropogenic noise and physical structures exhibited significant negative effects on bird abundance and species richness. Interestingly, we did not find evidence to suggest that the impact of anthropogenic noise was dependent on the level of physical structures. Anthropogenic noise had a significant negative relationship with bird abundance and species richness at all levels of physical structures.
Discussion: We provide evidence that bird abundance and species richness respond negatively to urbanization-related increase in anthropogenic noise and physical structures. That the impact of anthropogenic noise on birds was independent of physical structures suggests that birds in reserved areas, including urban green areas with fewer human activities may equally be affected by noise as are birds in human-dominated areas, if noise sources are near. Overall, our research underscores the detrimental consequences of anthropogenic habitat modification, particularly the alteration of structural and acoustic properties, and emphasizes the importance of preserving undisturbed habitats and implementing ecologically mindful urban planning strategies to safeguard bird communities in the Afro-tropics.
Urbanization is the transformation of rural areas and natural landscapes into built-up environments through the development of infrastructure, such as buildings, roads, and utilities. The main driver of urbanization is human population increase (United Nations, Department of Economic and Social Affairs, Population Division, 2018); a process by which an increasing proportion of a population resides in urban areas, leading to the growth and expansion of cities and towns. Urbanization represents a major transformative anthropogenic process shaping terrestrial landscapes worldwide. It alters natural habitats and is considered one of the most significant threats to wildlife, leading to the extinction of native species (Isaksson, 2018).
Although urbanization impacts all biodiversity, birds are a common subject used in studying this impact. They are considered to be a good candidate taxon, because they show predictable population responses to environmental changes and relatively easy to detect and identify (Gregory et al., 2003; Fraixedas et al., 2020). Typically for birds, increased urbanization has significant influence on species richness, diversity, abundance and composition (Benítez-López et al., 2010; Batáry et al., 2018; Callaghan et al., 2019; Neate-Clegg et al., 2023). Although, species richness and diversity are commonly reported to decrease with urbanization (Evans et al., 2018), there are also reports that, for some species, abundance increases with urbanization intensity because they are capable of exploiting available resources within the altered urban landscape (Proppe et al., 2013; Evans et al., 2018; Isaksson, 2018).
The variation in the level of urban development allows classification of settlements into rural, sub-urban and urban land uses. As such, the rural-urban gradient is important for studying the effect of urbanization on birds (Norton et al., 2016; Narango and Rodewald, 2016). Studies along this urbanization gradient – rural-suburban-urban have shown to be more effective in detecting the negative effect of urbanization on bird communities when compared to studies that only compared two levels of urbanization (Batáry et al., 2018).
Two important features of the urban environment are physical structures - concrete buildings, tarred roads and other reflective surfaces; and anthropogenic noise emanating from different sources such as moving vehicles, industry machines, human activities, home gadgets etc (Slabbekoorn and Peet, 2003; Dowling et al., 2012; Matthew et al., 2024). An increase in the density of urban physical structures results in reduced bird species richness because of the destruction and fragmentation of suitable habitats and foraging sites for the birds (Seress and Liker, 2015; Isaksson, 2018). However, some species, especially synanthropic species benefit from the abundant physical structures in urban areas because they take advantage of cavities and edges of buildings as well as structures like telecom towers for roosting and nesting, thereby accounting for increased bird abundance along the urbanization gradient (Mainwaring, 2015).
Most anthropogenic noise is less than 2 kHz (Slabbekoorn and Peet, 2003) and can impair communication in birds through the masking of bird vocalizations occurring in its range (Kleist et al., 2016). Almost all birds utilize vocalization, or other sounds as a channel of communication, which has important functions for survival and reproduction (Winkler, 2001; Osinubi et al., 2012). Many bird species, therefore, tend to avoid noisy areas leading to reduced abundance and richness (Proppe et al., 2013; Kumdet et al., 2021; Ajibola-James et al., 2024). This has been demonstrated in the result of a meta-analysis by Benítez-López et al. (2010) showing a general decrease in bird population densities with increasing proximity to noise sources. Additionally, anthropogenic noise can influence birds to abandon rich habitat patches due to high predation risk driven by the inability of individuals to detect danger in noisy habitats (Slabbekoorn, 2017).
Importantly, anthropogenic noise and physical structures are shown to increase along the urbanization gradient (Dowling et al., 2012, Matthew et al., 2024); studies have also demonstrated shifts in bird communities along this gradient (Benítez-López et al., 2010; Evans et al., 2018; Callaghan et al., 2019; Neate-Clegg et al., 2023). However, the impact of urbanization, including the specific impact of anthropogenic noise and physical structures, on bird communities has received relatively less attention in Africa (Awoyemi and Ibáñez-Álamo, 2023) despite the continent’s high rate of urbanization (United Nations, Department of Economic and Social Affairs, Population Division, 2018). As such, our understanding of their singular as well as interactive effects on birds remain limited. In this study, we evaluated the effect of anthropogenic noise and physical structures on bird communities along an urbanization gradient in Nigeria, an Afrotropical environment. Specifically, 1) we compared bird abundance, species richness and composition among settlements along the urbanization gradient, 2) we determined the effects of anthropogenic noise and physical structures on the abundance and species richness of birds and 3) we determined the combined effect of the above predictors on the abundance and richness of birds.
We carried out bird surveys in Jos (latitude 09°55′00″N, and longitude 08°53′25″E), Plateau State, Nigeria (Figure 1), from October to December (late wet season to early dry season), 2020. Jos is the largest human settlement on the Jos Plateau and the capital city of Plateau State. According to the national 2006 census, Jos had a population of about 900,000 residents (Federal Republic of Nigeria, 2012). Recent findings revealed that the city had marked differences in land-use types and urbanization levels that depict a rural-to-urban gradient (Matthew et al., 2024).
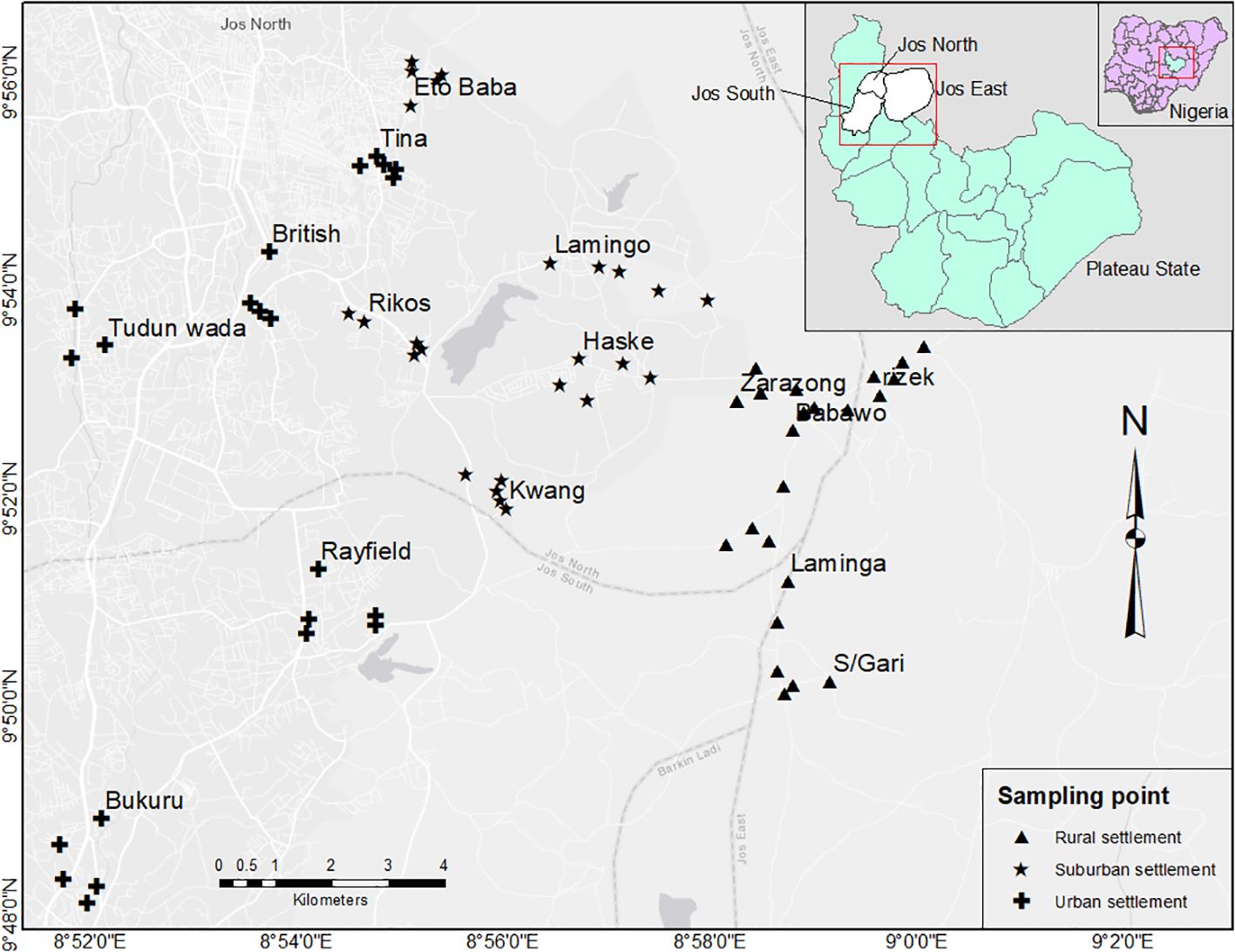
Figure 1. Study area map, showing sampling points along the rural-urban gradient in Jos). Five point count stations were sampled at each of 5 rural sites (Zarazong, Babawo, Rizek, Laminga, SabonGari), 5 suburban sites (Kwang, Haske, Lamingo, Eto Baba) and 5 urban sites (Burkuru, Rayfield, Tudun Wada, British, Tina), making a total of 25 points for each of the three urbanization categories and a total of 75 sampling points across the rural-urban gradient.
Site were representative of different urban land uses from rural areas to the city center. Rural areas were characterized by relatively lower densities of people (≤142 individuals/Km2), buildings and road networks. Land use type is predominantly farmlands and with scattered building interspersed by open lands and natural vegetation. Suburban areas were characterized by intermediate population (between 142-800 people/Km2), intermediate building and road network densities and mainly included areas just outside of busier urban centers. Urban areas were characterized by high population density (>1000 people/Km2), high density of buildings and human-created structures, little open space, dense road networks, and high commercial activities. Population density information were sourced from (https://citypopulation.de/en/nigeria/admin/plateau/) and (https://www.arcgis.com/apps/mapviewer/index.html?layers=bcaa2926caf54782b360bd46305cecf1). According to Matthew et al. (2024), physicals structures (mostly buildings and pavements) along this gradient ranged from 0.49% in rural area to 99% in the urban area (Figure 1).
We used a stratified-random design for this study. Rural, suburban and urban areas were identified in the Greater Jos Metropolis (this includes Jos North, Jos South, and Jos East Local Government Areas). Five point count stations were sampled at each of 5 rural sites (Zarazong, Babawo, Rizek, Laminga, SabonGari), 5 suburban sites (Kwang, Haske, Lamingo, Eto Baba) and 5 urban sites (Burkuru, Rayfield, Tudun Wada, British, Tina), making a total of 25 points for each of the three urbanization categories and a total of 75 sampling points across the rural-urban gradient (Figure 1).
Each point was surveyed just once and for 10 minutes between the hours of 0630 - 1100. Points were located between 200 - 500 m apart in order to reduce the chance of double counting and pseudo replication (Morelli et al., 2017). During counts at each sampling point, all bird species seen and heard withing a 50 meters radius around the sampling points were counted and recorded (Danjuma et al., 2012). We sticked to 50 meters radius to minimize problems of detectability associated with increasing distance of birds from the observer. We also avoided collecting bird data on rainy days or days with strong wind. To avoid any bias due to observer ability, all sampling points were surveyed by the same observer (DAM) who is a trained ornithologist.
In this study we considered physical structures to include both horizontal and vertical man-made urban structures such as buildings tarred roads, concrete and interlocking floors (Dowling et al., 2012; Isaksson, 2018). We determined the percentage of physical structures within a 200 m radius around the points where birds were counted. GPS coordinates of sampling points were exported from the Garmin Etrex 20 GPS device onto the BaseCamp software and saved as a single file in the GPS exchange format (GPX). The data was thereafter cleaned using the MapSource software and converted into a CSV file in excel 2016. We then obtained a 30 m resolution satellite image (Landsat 8) of the study area for the year 2020 from the Earth Explorer website (earthexplorer.usgs.gov). In ArcGIS 10.8, we overlaid the shape file of the study area extracted from the map of Nigeria on the satellite image and carried out supervised image classification as follows. Training samples were generated by selecting multiple pixels of the different land-use types and land features -vegetation, water body, impervious surfaces like roads and concrete floors, and building from the composite layer of bands 1-7 of the satellite image. The mapping software ArcGIS then used these training samples to categorize the area in the shape file into the respective categories of land-use types and land features (Richards, 1986). Afterwards, we grouped all land use types and land features into physical structures and others since we were only interested in physical structures. Still in ArcGIS, we imported coordinates of the sampling points overlaid on the classified image, and converted them into a shape file and created a 200-m radius buffer around each coordinate. Finally, we calculated the percentage of physical structures within each buffer using the “calculate geometry” function (Rhodes et al., 2023; Hao et al., 2024). The percentage was calculated by dividing the number of pixels of physical structures by the total number of pixels and multiplied by 100.
At every sampling point, we recorded anthropogenic noise in decibel (dB) at 1.5 m height with a calibrated Extech 407730 digital sound level meter for three minutes from the four cardinal directions (N, E, S, W) (Slabbekoorn and Smith, 2002; Matthew et al., 2024). We set the noise meter to “C weighting” and “slow” mode when recording. This setting is good at capturing most of the low-frequency background noise and automatically averages the noise levels in case of abrupt high levels. We then took the mean of the noise levels from the four cardinal directions for each point.
We used the R program version 4.3.3 (R Core Team, 2024) for all analyses. For objective 1: testing the variation in general bird abundance and abundance of birds encountered at least at one point in all the urban categories (hereafter called common birds), bird species richness and bird species composition along the urbanization gradient; we used a generalized linear model (GLM) with the Poisson error distribution because the response variables were count data. There was overdispersion in the models with abundance as a response variable, therefore, we used the quasipoisson error distribution for the GLM analyses having abundance as a response variable. We maintained the use of the Poisson error distribution where there was no overdispersion, as was the case with all the GLMs having species richness as the response variable. The post hoc test, emmeans from the emmeans package in R was used for pairwise comparison between levels of categorical predictor variables. Furthermore, we used analysis of similarity (ANOSIM) with the adonis function in R to test the similarity in bird species composition between and among settlements. Thereafter, we visualized the result of ANOSIM using a Non-metric Multidimensional Scaling (NMDS) analysis. For objective 2, to test the individual effect of anthropogenic noise and physical structures; we first checked and confirmed weak correlation between anthropogenic noise and physical structure (r = 44.29). We then included both variables in the models for bird abundance and species richness. Therefore, one model was run for testing abundance and another model for species richness. Before testing the second objective, we first used a GLM with gaussian family to investigate variation in values of anthropogenic noise and physical structures across the settlement gradient (rural, suburban and urban). To test objective 3, the interaction effect of anthropogenic noise and physical structure on bird abundance and richness, we ran a GLM model with interaction of anthropogenic noise and physical structure. However, to be able to determine and visualize the relationship between one predictor variable (anthropogenic noise) and the response variables within each category of the other predictor variable (physical structure), we used a k-means clustering analysis to categorized physical structures into two categories (high and low) (Dowling et al., 2012) as determined by Within-Cluster Sum of Squares (WSS) and silhouette plots from the “factorextra” package in R. The reason for using k-means clustering was to avoid the ambiguity that may result when the variables are grouped by mere settlement classification (Matthew et al., 2024). Simple classification of study sites into rural and urban may not fully describe the variation in the physical and acoustic habitat properties between sites for reason of overlaps in values of the variables (Zhan et al., 2021). K-means clustering is able to group the values into categories irrespective of settlement classification by pooling similar values from all settlements around a given mean and creating non-overlapping levels of the variables.
A total of 1214 birds of 68 species in 37 families and 14 orders were recorded in this survey (species list can be found in Supplementary Table S1). The total count of birds was 524 (rural), 393 (suburban) and 297 (urban). Bird abundance ranged from 4 to 43 in the rural, 3 to 30 in the suburban and 4 to 28 in the urban. Species richness was 54 in the rural, 43 in the suburban and 31 in the urban (see Figure 2 for species accumulation curve). Furthermore, the species richness at sampling points ranged from 4 to 19 for rural, 3 to 11 for suburban and 2 to 11 for urban. Compared to rural settlement, the bird abundance was significantly lower in suburban (GLM: Est = -0. 2877 ± 0.13, t = -2.126, df = 73, p = 0.037, R2 = 0.22) and urban (GLM: Est = -0.527 ± 0.15, t = -3.578, df = 73, p = 0.001, R2 = 0.22) settlements respectively, Figure 3A). However, pairwise comparison with a post-hoc test revealed no significant difference in the abundance of birds between suburban and urban settlements (p = 0.348) Relatedly, bird species richness was significantly lower in suburban (GLM: Est = -0.271 ± 0.09, z = -2.972, df = 73, p = 0.003, R2 = 0.36) and urban (GLM: Est = -0.481 ± 0.09, z = -4.893, df = 73, p < 0.001, R2 = 0.36) settlements respectively compared to the rural settlement (Figure 3B) which had a mean abundance of 10.64 ± 4.27. Although species richness was higher in the suburban settlement, compared to the urban settlement, the observed difference was not statistically significant (p = 0.225).
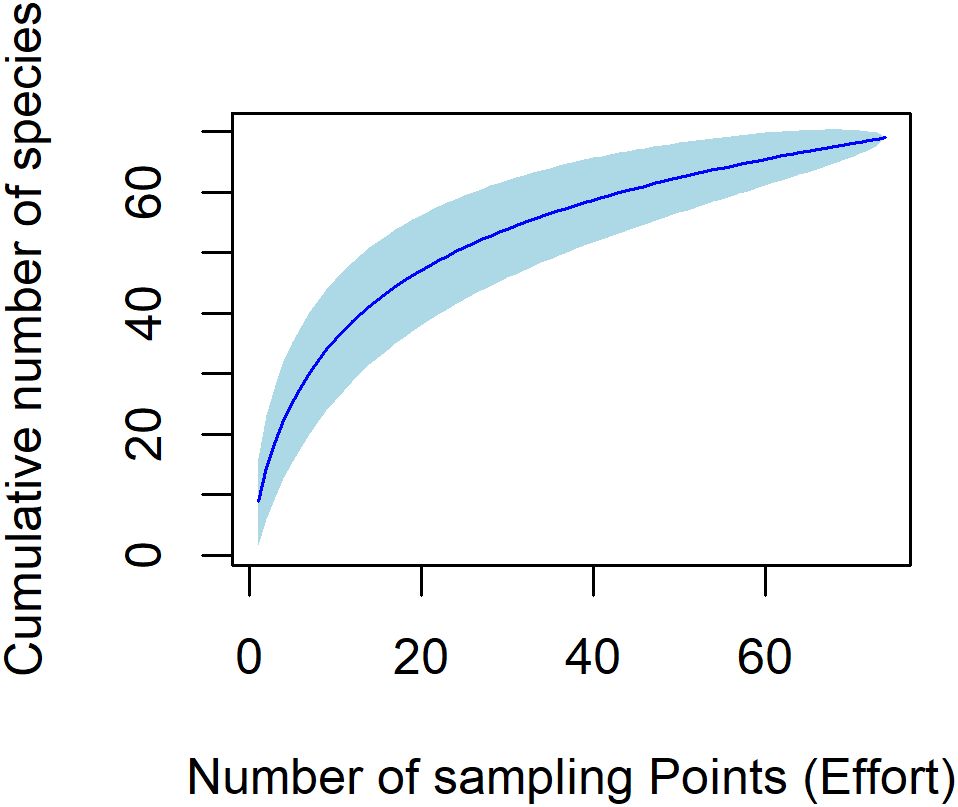
Figure 2. Species accumulation curve for all the 75 sampling stations across all sites. A total of 68 species were encountered in the study.
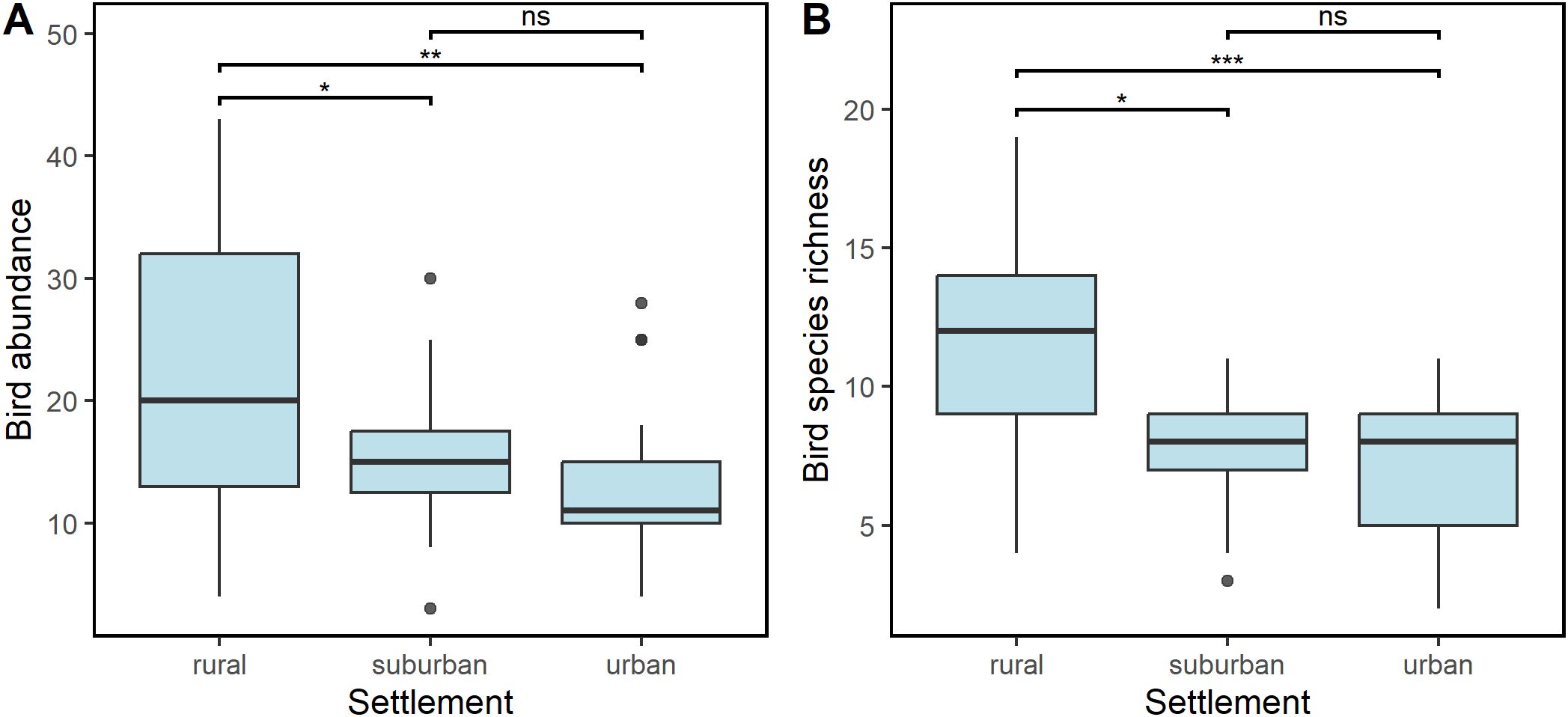
Figure 3. Variation in bird abundance (A) and bird species richness (B) across urbanization gradient. Bird abundance was significantly higher in rural area than suburban and urban areas. However,the result of a post-hoc test of pairwise comparison between suburban and urban areas with TukeyHSD revealed no significant difference between them (p = 0.275). The species richness was significantly higher in rural area than suburban and urban areas. However,the result of a post-hoc test of pairwise comparison between suburban and urban areas with TukeyHSD revealed no significant difference between them (p = 0.107). * p <0.05, ** p < 0.01, *** p < 0.001.
Of the 68 species recorded in the survey, only 17 were common to all settlements (i.e., species present at least at one point in each of rural, suburban and urban areas). The overall abundance of these common species was lower in urban and suburban settlements compared to rural settlement (Figure 4). The observed difference between rural and urban settlements was significant (GLM: Est = -0.354 ± 0.14, t = -2.454, df = 73, p = 0.016, R2 = 0.07), suburban was not significantly different from the rural settlement (GLM: Est = -0.178 ± 0.14, t = -1.310, df = 73, p = 0.194, R2 = 0.07). Furthermore, a pairwise comparison between suburban and urban settlements did not show a significant difference (p = 0.462).
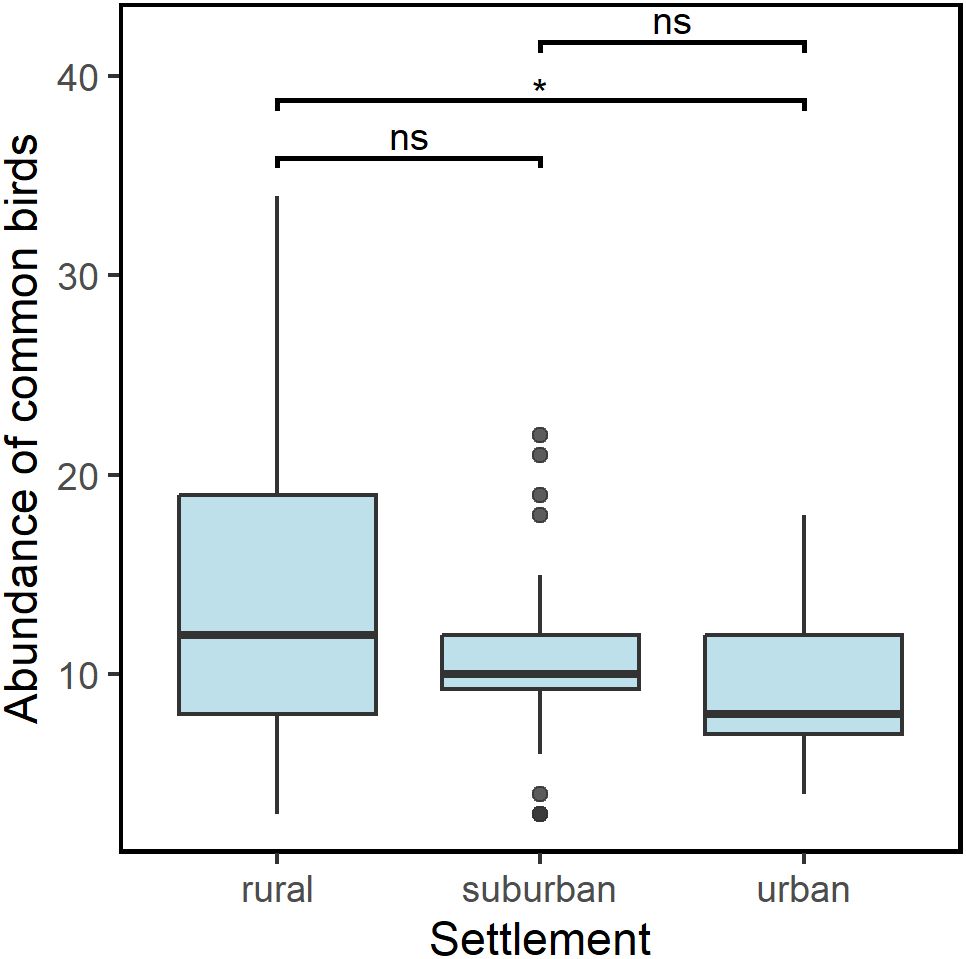
Figure 4. Abundance of birds common to all settlements across the urbanization gradient. Significant difference was only found between rural and urban areas. * p <0.05; ns, p value not significant.
The number of bird species uniquely observed at the different settlements generally decreased along the gradient from rural to urban (see Table 1 for a list of unique species and the number of individuals recorded for each settlement); while 16 species were observed in the rural settlement only, 7 and 3 species were observed in only suburban and urban settlements respectively. Although there was a general decrease in the abundance of the common birds, some species like the Laughing Dove Stretopilia senegalensis and Speckled Pigeon Columbia guinea, showed peak abundance in suburban settlements; a few other species such as Western Plantain Eater Crinifer piscator, Yellow-billed Kite Milvus aegyptius, Red-billed Firefinch Lagonosticta senegala and Bronze Mannikin Spermestes cuculata, showed an increasing trend from rural to urban settlements (Figure 5). However, none of the observed variation in the number of individual species across the gradient was significant (Supplementary Table S2).
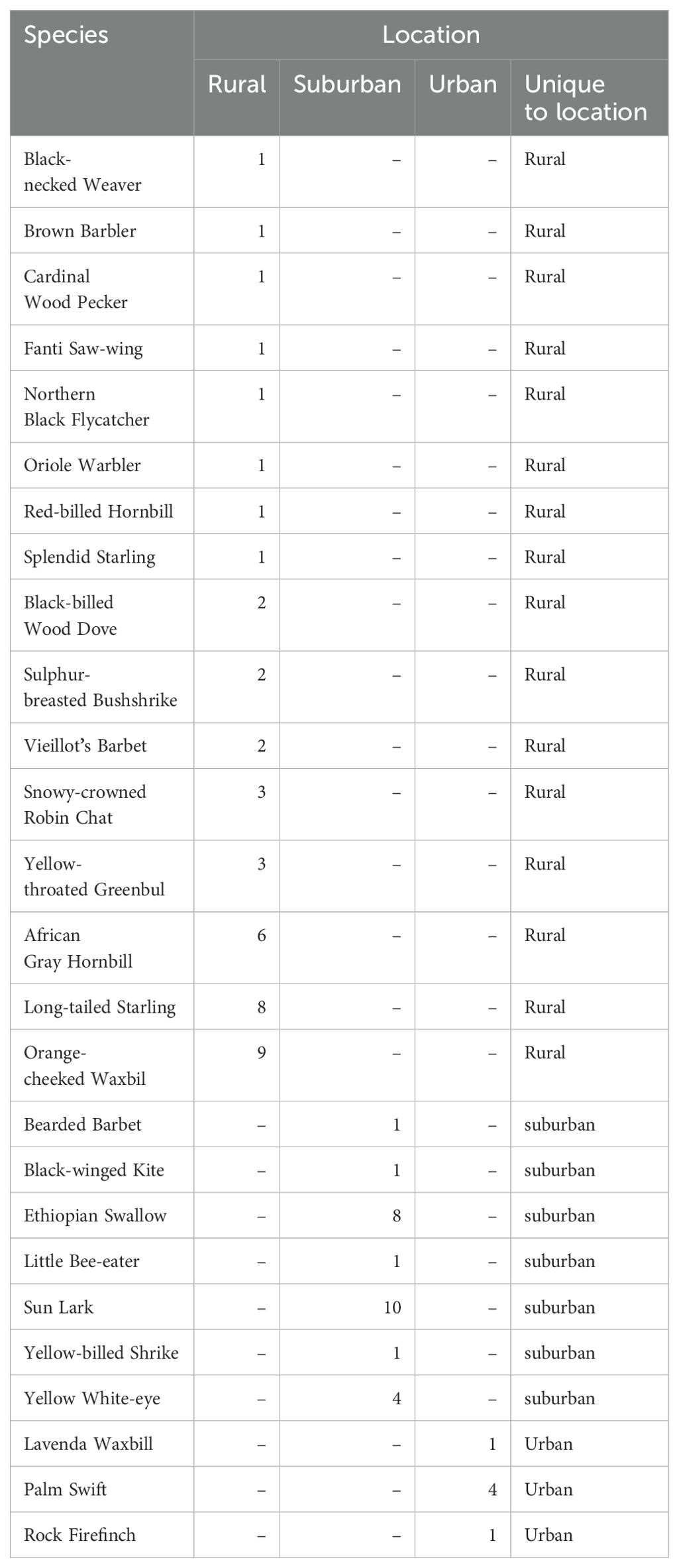
Table 1. List of species and number of individuals recorded uniquely in each settlement.
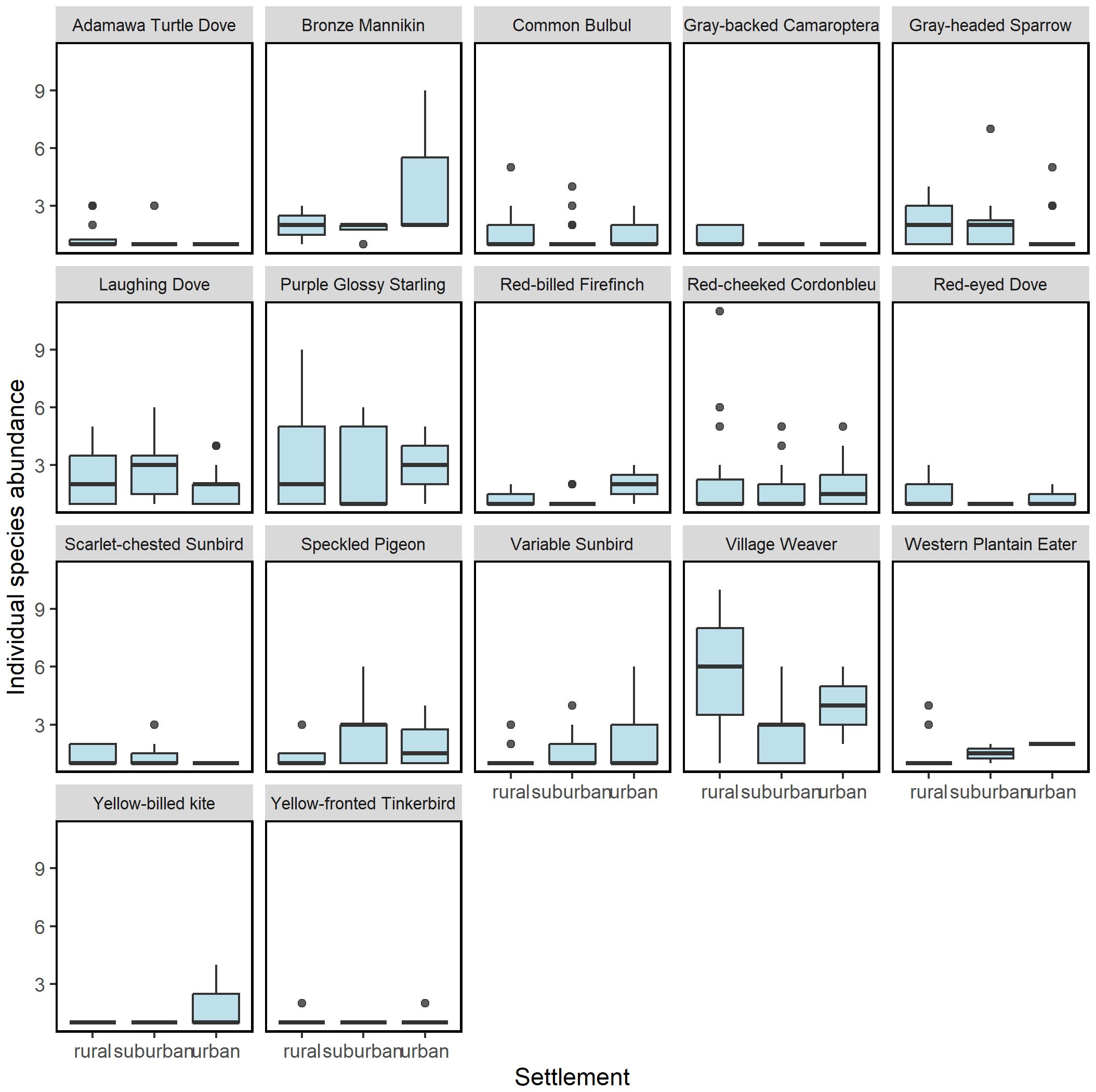
Figure 5. Count of common bird species across urbanization gradient. This figure shows only number of the 17 species observed at least at one point in each of rural, suburban and urban areas, termed common species. The Laughing Dove Stretopilia senegalensis and Speckled Pigeon Columbia guinea, showed peak count in suburban settlements; the Western Plantain Eater Crinifer piscator, Yellow-billed Kite Milvus aegyptius, Red-billed Firefinch Lagonosticta senegala and Bronze Mannikin Spermestes cuculata, were more in urban area. The remaining 11 species were more in rural area.
Table 2 shows the Bray-Curtis dissimilarity indices from ANOSIM of species composition (not for common species but for all species) between settlements. Bird species composition was more similar between suburban and urban settlements than rural versus suburban, and rural versus urban settlements respectively. The ANOSIM showed that the observed difference in bird community composition across settlements was significant (R = 0.20, p = 0.001). This is visualized with non-metric multidimensional scaling (Figure 6).
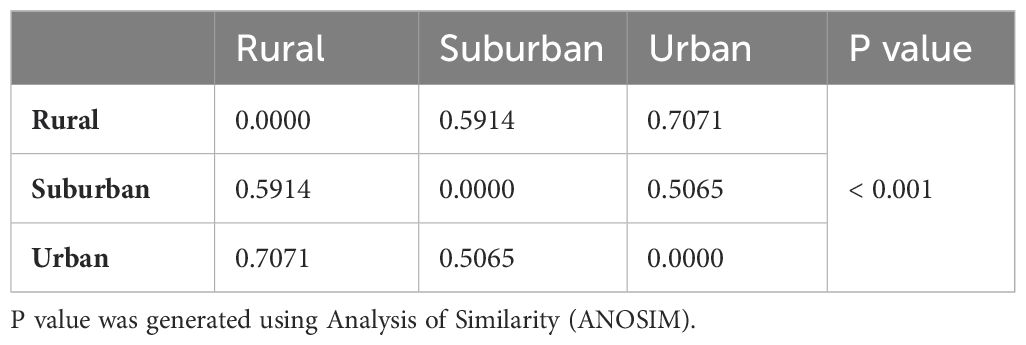
Table 2. Bray-Curtis dissimilarity indices between settlements.
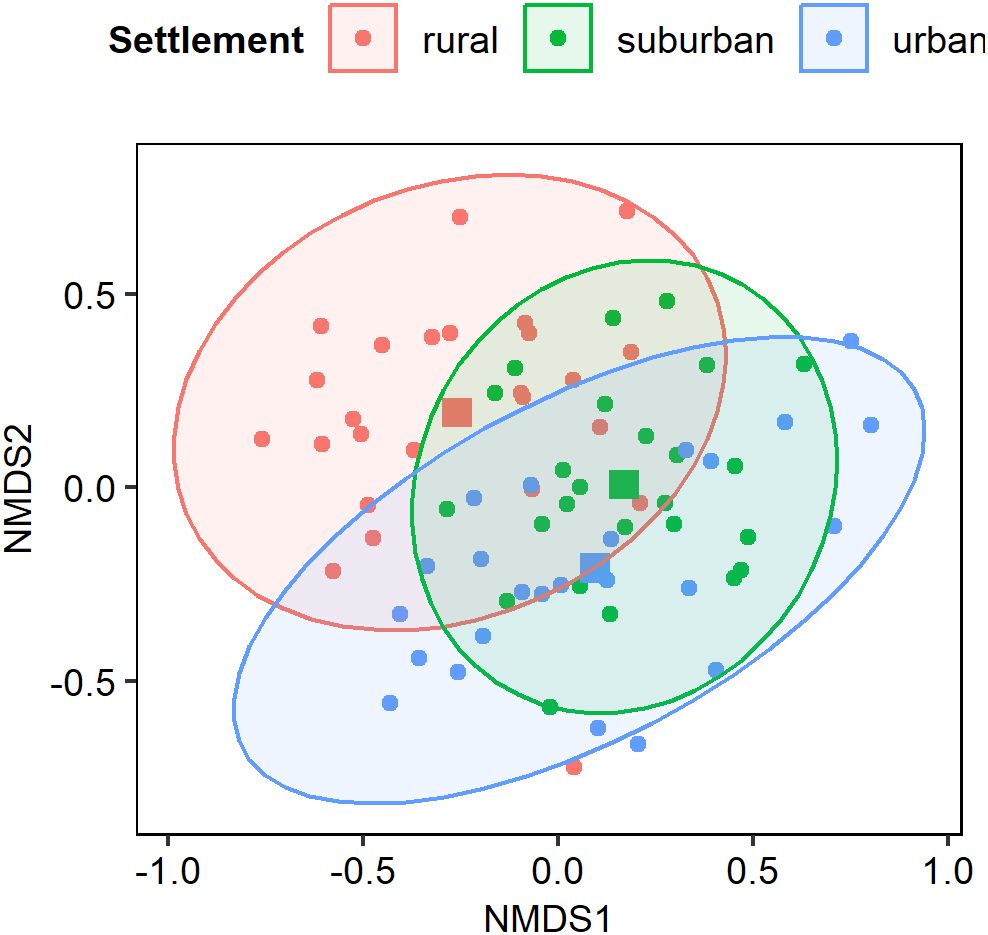
Figure 6. Result of NMDS of bird community composition across settlements along the urbanization gradient. Stress = 0.2102. Species composition in suburban settlement was more similar to urban settlement than the rural settlement. The centroids for the settlements are represented with square shapes and circular points represent species.
The levels of anthropogenic noise and physical structures differed across settlement types. They significantly increased from rural to urban settlement along the urbanization gradient (GLM: Anthropogenic noise: Chisq = 20.794, df = 2, p < 0.0001, R2 = 0.25 Figure 7A; percentage physical structure: Chisq = 84.75, df = 2, p < 0.0001, R2 = 0.57; Figure 7B).
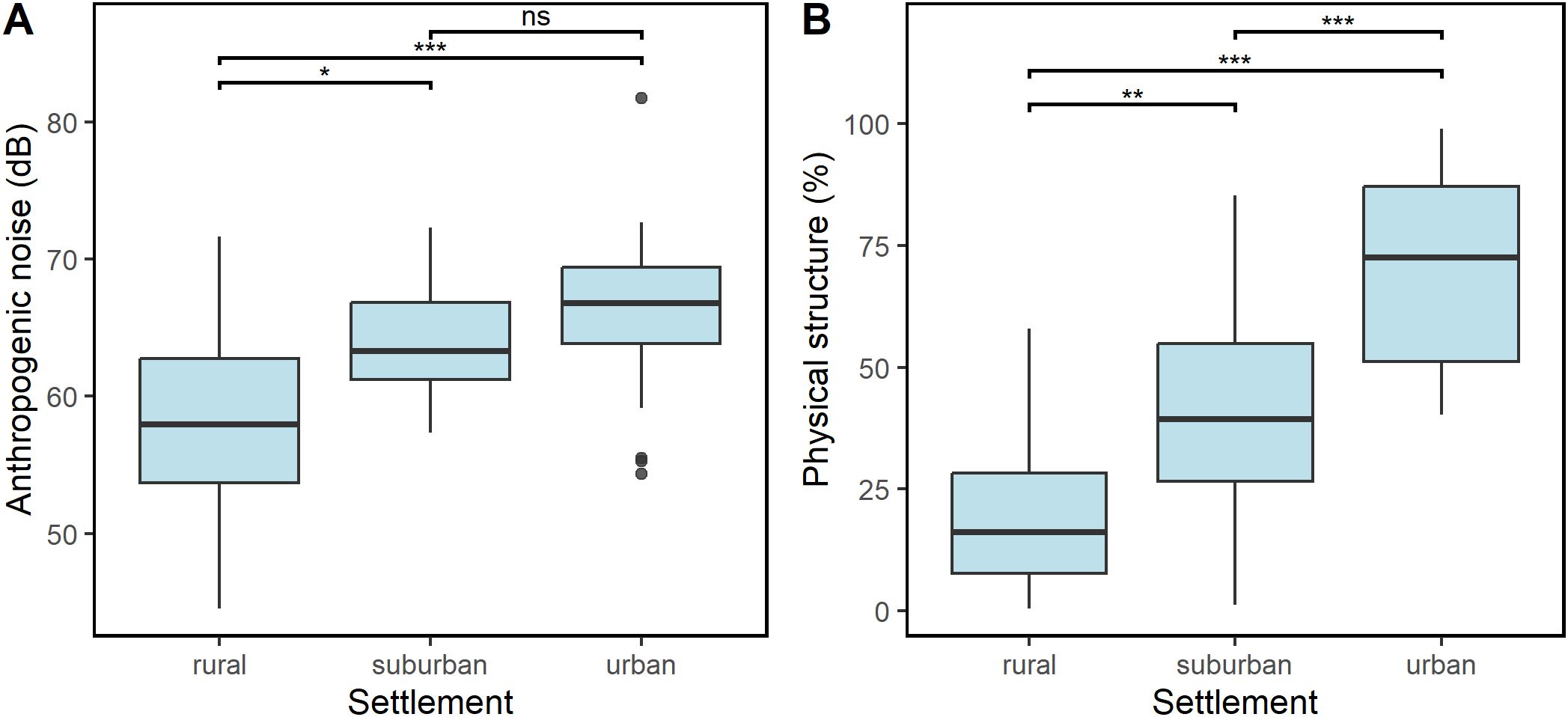
Figure 7. Variation in ambient noise (A) and percentage cover of physical structures (B) along the urbanization gradient. There were significant differences in the levels of anthropogenic noise and physical structure across the settlements.The difference in ambient noise between suburban and urban settlements was not significant. * p <0.05, ** p < 0.01, *** p < 0.001.
Bird abundance had a negative and significant relationship with anthropogenic noise (GLM: Est = -0.025 ± 0.51, t = -2.843, df = 2, p = 0.006, R2 = 0.32, Figure 8A) and physical structure (GLM: Est = -0.005 ± 0.01, t = -2.670, df = 2, p = 0.009, R2 = 0.32Figure 8B). Similarly, as shown in Figures 9A, B, bird species richness had a negative and significant relationship with anthropogenic noise (GLM: Est = -0.209 ± 0.01, z = -3.176, df = 2, p = 0.001, R2 = 0.44) and physical structure (GLM: Est = -0.007 ± 0.01, z = -3.176, df = 2, p = 0.001, R2 = 0.44) respectively. The observed decrease in bird abundance and species richness with anthropogenic noise and physical structure were not affected by the level of urbanization (Supplementary Figure S1).
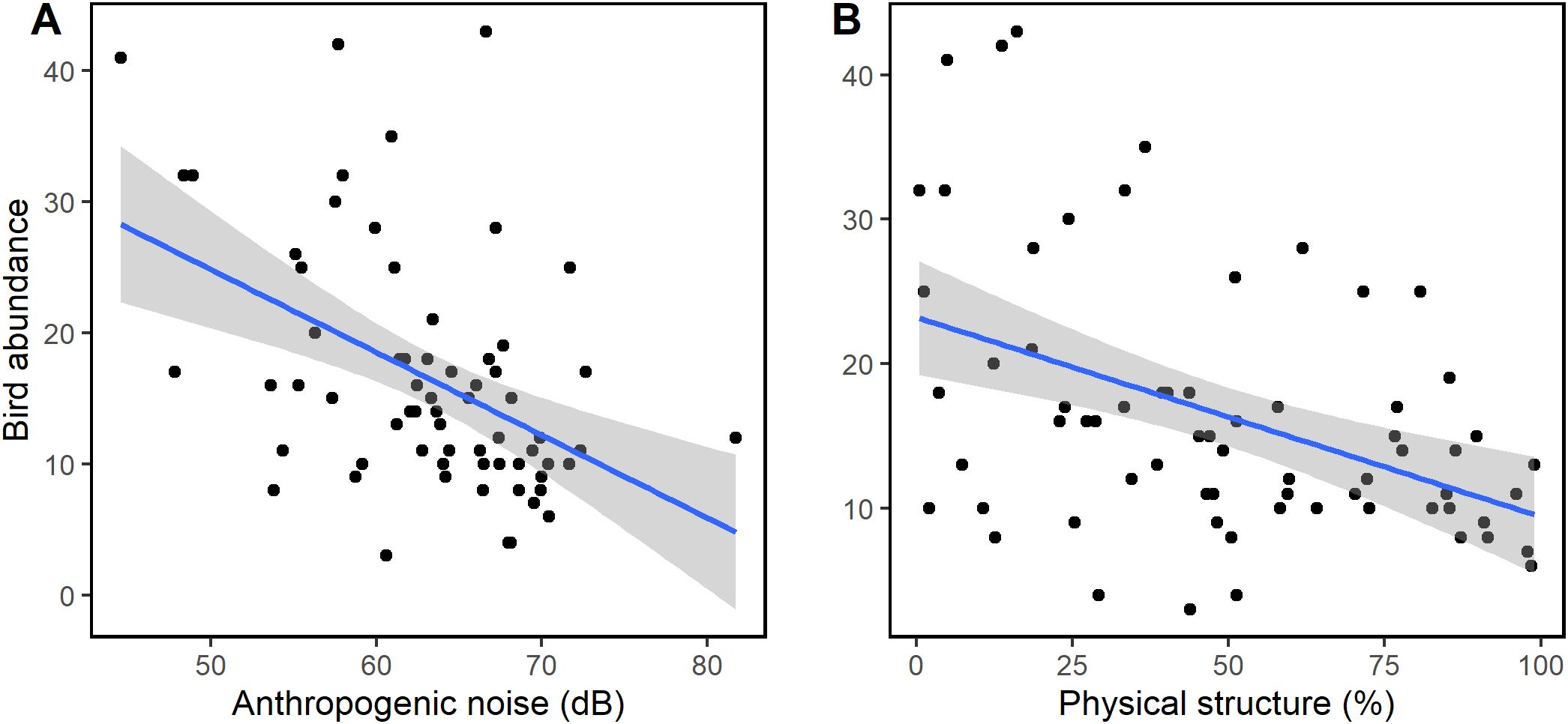
Figure 8. Relationship between (A) ambient noise, (B) physical structure and general bird abundance. Significant decrease of bird abundance with increasing levels of anthropogenic noise and physical structure was observed.
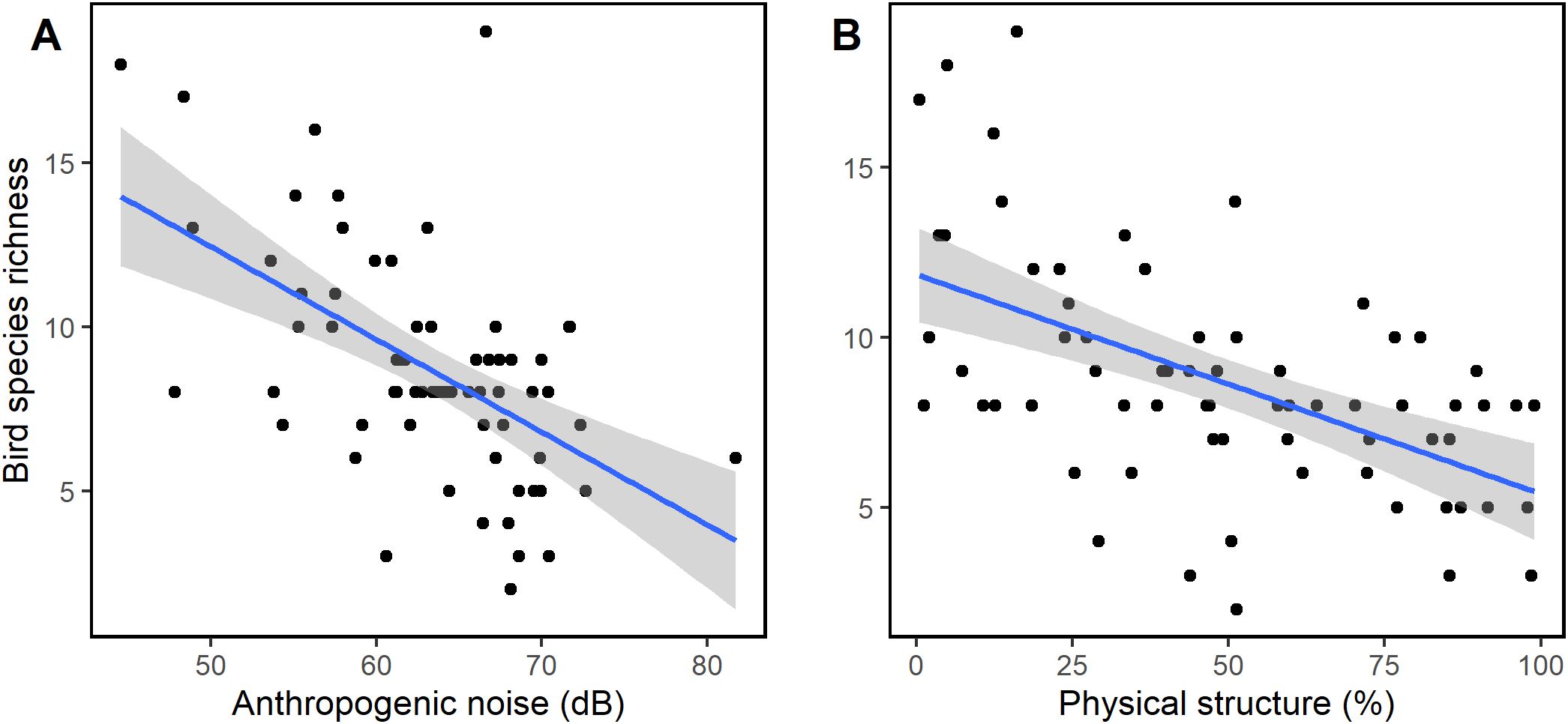
Figure 9. Relationship between (A) ambient noise, (B) physical structure and bird species richness. Significant decrease in bird species richness with increasing levels of anthropogenic noise and physical structure was observed.
The effect of anthropogenic noise on bird abundance and species richness did not seem to be dependent on physical structure as the effect was similar at all levels of physical structures. The interaction models were not statistically significant (GLM: physical structure x Anthropogenic noise: Abundance: Chisq = 2.136, df = 1, p = 0.144, R2 = 0.28, Figure 10A; Species richness: Chisq = 0.713, df = 1, p = 0.399, R2 = 0.38, Figure 10B). Similar trend was observed at all urbanization levels (Supplementary Figure S1).
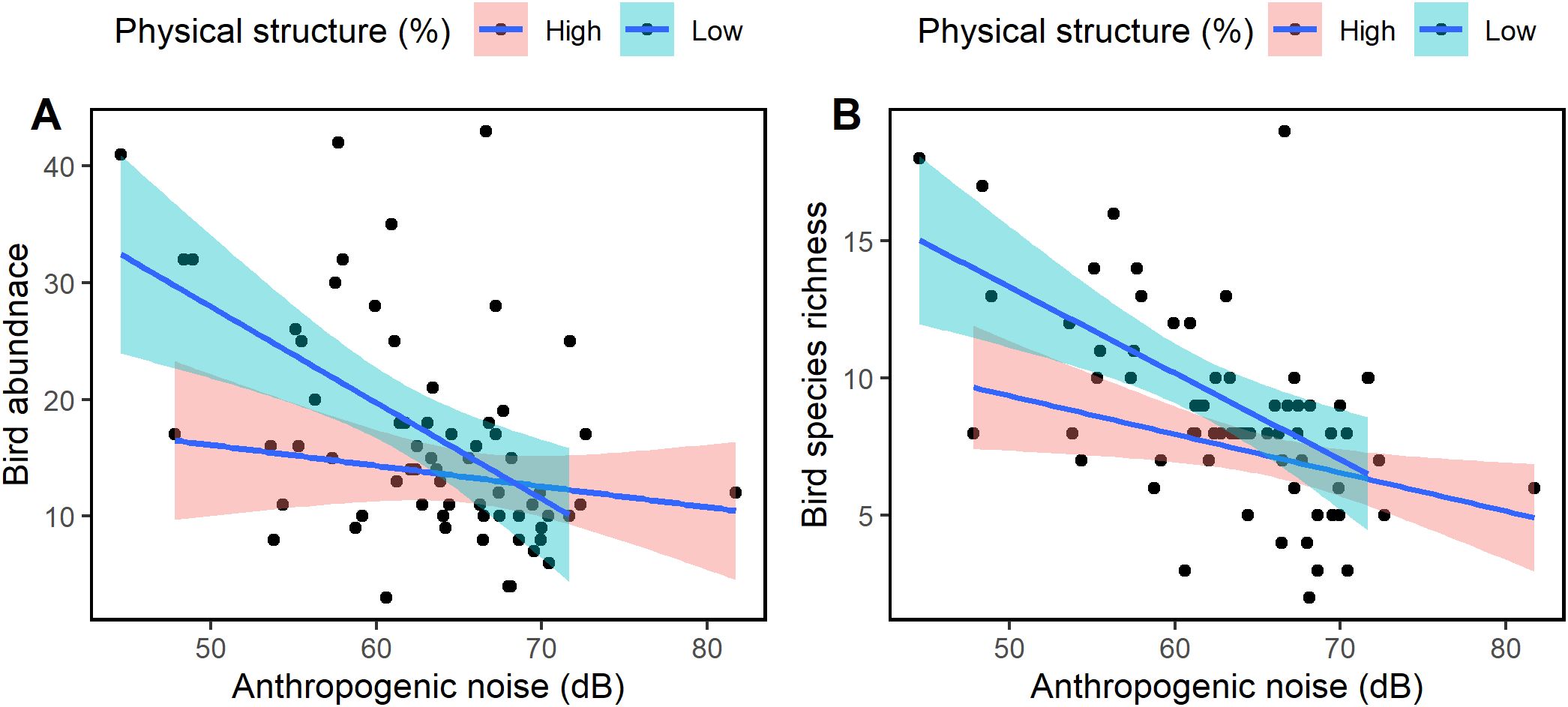
Figure 10. Relationship between (A) anthropogenic noise, (B) bird abundance and species richness at different levels of physical structure. The interaction of anthropogenic noise and physical structure was not significant on bird abundance and species richness. There was decrease of bird abundance and species richness at each level of physical structure.
Our results show that the abundance and richness of avifauna decreased along the rural-urban gradient. This is likely in response to the level of development along the gradient; we observed a significant increase in both anthropogenic noise and percentage cover of physical structure along the urbanization gradient (rural -suburban-urban). Together with climate change, land-use change in the form of urban development is recognized as a major threat to wildlife (Isaksson, 2018). The process of urbanization results in the destruction of natural habitats and introduces human-induced pressure that ultimately affects bird communities (Isaksson, 2018). The observed decrease in bird species richness along the rural-urban gradient in this study implies that some bird species are unable to cope with the alteration being made to their habitat (Isaksson, 2018). We observed that most bird species assumed to be found in undisturbed sites gradually dropped out as the landscape transformed into urban settlements, and this perhaps explains why the rural settlement had the highest number of unique species. Slabbekoorn et al. (2007) recognized that the urban environment is evolutionarily new to birds while Isaksson (2018) acknowledged that a great majority of birds are unable to have their normal life in this altered environment of the urban landscape. Our observation of the decline in species richness along the rural-urban gradient agrees with the reports of several other studies captured in a meta-analysis by Batáry et al. (2018). This relationship is observed across different biogeographical regions (Evans et al., 2018).
We also observed a decrease in abundance along the urbanization gradient implying that urban development also negatively impacts the number of birds. Our result agrees with previous reports of similar studies (Catterall, 2009; Gagne and Fahrig, 2011; Rija et al., 2014). However, some studies have reported contrasting results of an increase in bird abundance along urbanization gradients (Batáry et al., 2018; Rashid et al., 2020; Kurucz et al., 2021). Kurucz et al. (2021) suggested that the increase in bird abundance along the urbanization gradient was primarily due to the dominance of synanthropic species. Surprisingly, we found a general decrease in the abundance of birds that were common to all settlements, including very common synanthropic species like Village Weaver Pleocus cuculatus, Common Bulbul Pycnonotus barbatus and Northern Gray-headed Sparrow Passer griseus. This suggests that there may be a limit to the level of anthropogenic pressure the birds can tolerate, even if they are associated with humans. The various anthropogenic pressures, such as artificial light at night, chemical and noise pollution (Isaksson, 2018), observable in urban areas may affect the physiology, breeding and survival of birds. Furthermore, the observed decrease in bird abundance from this study contrary to other reports may reflect regional differences in bird responses to urbanization. Luck and Smallbone (2010) suggested that it may be difficult to generalize bird response to urbanization because ecological traits and local factors which play significant roles in species responses to the altered environment may differ across regions. For example, some of the studies that reported an increase in bird abundance across the urbanization gradient were conducted in regions that have long history of urbanization, unlike the Afrotropical landscapes that are recently facing a rapid urbanization process, as obtainable in our study area. Therefore, the Afrotropical birds may not have adapted so well to the effect of urbanization. This calls for attention to the rapid transformation of the African environment into the urban landscape. On the other hand, the abundance of some species like Western Plantain Eater, Yellow-billed Kite, Red-billed Firefinch and Bronze Mannikin, increased along the gradient, suggesting differential responses to urbanization by species. Local differences in species responses to urbanization may be due to the life history traits of the species present in the study area, and the local habitat structure (Chamberlain et al., 2017). It may, therefore, be useful to consider the the life history traits of species and the local habitat structure in detail when studying the response of bird communities to urbanization. Although we did not focus on feeding guild response to urbanization in this study, we observed from our data that granivores were the most common, across the settlements. Future studies to look into guild response to urbanization will provide insight into how different species in different guild respond to urbanization. The Speckled Pigeon and Laughing Dove showed peak abundance in the suburban area. Although there was a general decreasing trend in abundance along the urbanization gradient, the observation of some species being more in suburban and urban areas respectively, implies some extent of non-linearity in bird response to urban development (Batáry et al., 2018) and the ability to tolerate and exploit the altered environment by some bird species. We observed that in the urban areas, bird abundance and species richness were highest in places that were less altered or in old residential areas with gardens compared to the highly urbanized places like the city center that lack green areas.
The analysis of similarity using Bray-Curtis distance showed that bird community was more similar between suburban and urban than suburban and rural. This is contrary to the finding of Rashid et al. (2020), where the suburban settlement was closer to rural settlement in species composition than the urban settlement. While the rural areas are more like natural landscapes because of relatively low human disturbance, suburban areas are extensions of urban areas and therefore have similar levels of disturbance as urban areas. This may have resulted in the closer similarity of bird communities observed between suburban and urban settlements. Regarding the species observations unique to each settlement, we recorded the Rock Firefinch Lagonostica sanguinodorsalis, Palm Swift Cypsiurus parvus and Lavenda Waxbill Glaucestrilda caerulescens only in urban settlement. It is worth noting that these species are fairly common and can be found in varieties of habitats and settlements (personal observation). However, in this study we sighted each of them only in urban area and there was only one observation for each of them except the Palm Swift that was observed at two points and had a total of 4 individuals. The observation that Rock Firefinch, Palm Swift and Lavenda Waxbill were only found in the urban area and not in rural or suburban areas where they are expected to be found suggests that they may not be permanently absent from the rural or suburban areas. This is in agreement with Carral-Murrieta et al. (2020), that species may temporarily fly away from or avoid disturbed sites. However, it is unlikely that poor detection was the case in rural and suburban areas. The Rock Firefinch is a vocal species and should have been easier to detect at rural sites which is less noisy. It was seen around rocky outcrop (which is known to be its preferred habitat) in the urban. African Palm swifts are arial species and could not have been easily missed at other sites. The Lavenda Waxbill was seen around grain mill in the urban area.
In this study we surveyed birds once at each point, we recommend that similar studies in the future should consider replicating bird counts to ensure inclusion of species that may be missed during one of the bird counts. Also, given that detection probabilities differ across bird species and landscapes, we recommend that this be considered in future studies as we did not consider it in this study.
Both anthropogenic noise and physical structures significantly increased along the urbanization gradient. They in turn exhibited significant negative influences on bird abundance and species richness at all levels of urbanization. This result suggests that both the structural and acoustic properties of the urban landscape have an important influence on bird communities. The increase in the level of physical structures leads to reverberation and scattering of bird sounds, thereby, impairing communication (Slabbekoorn et al., 2007). In addition, the loss of suitable habitat patches and foraging sites for birds due to increase in man-made physical structure may cause reduced bird abundance and species richness (Seress and Liker, 2015).
Anthropogenic noise has varied effects on bird population. These effects range from the masking of birds’ acoustic signals (Slabbekoorn et al., 2007; Benítez-López et al., 2010; Schroeder et al., 2012; McIntyre, 2013; Senzaki et al., 2016) to physiological stress on the birds (Barber et al., 2010), thereby leading to the reduction in breeding success and survival of birds and overall decline in their population (Habib et al., 2007; Halfwerk et al., 2011; Luo et al., 2015; Mason et al., 2016). On the whole, our result agrees with finding from other studies (Seress and Liker, 2015; Carral-Murrieta et al., 2020).
We did not find evidence to suggest that the effect of anthropogenic noise on bird abundance and richness depended on the level of physical structures. At both low and high levels of physical structure, the effect of noise on bird abundance and species richness was not significantly different. However, the slope due to noise at high levels of physical structures was less steep compared to low levels of physical structure. While the impact of urbanization should not be underestimated, it is possible that, over time, the few bird species that persist in urban environments may exhibit a slower response to the pressures of urbanization (Luck and Smallbone, 2010). This could be due to their ability to adapt and exploit the modified landscape. The observation that noise independently affects bird communities may not particularly favor the concept of land sharing for the conservation of bird species; birds in urban green areas with fewer human activities may equally be affected by noise as are birds in human-dominated areas. This is because noise can affect bird foraging and breeding up to a considerable distance from the noise source (Benítez-López et al., 2010). Many studies have reported decreased abundance and species richness of birds and other animals with noise-related pollution, even in conserved areas (Arévalo and Newhard, 2011; Wiącek and Polak, 2015; Avalos and Bermúdez, 2016; Senzaki et al., 2020; Kumdet et al., 2021; da Silva et al., 2021; Ajibola-James et al., 2024).
This study provides evidence that bird abundance and species richness respond negatively to urbanization-related increases in anthropogenic noise and physical structures. However, birds in reserved areas, including urban green areas with fewer human activities and lower levels of urban development may equally be affected by noise as are birds in human-dominated areas, if noise sources are near. The study underscores the importance of preserving undisturbed habitats and implementing ecologically mindful urban planning strategies to safeguard bird communities in rapidly developing Afrotropical landscapes.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by the A.P. Leventis Ornithological Research Institute's scientific committee. Data was non-invasively collected and no bird was captured or handled. The study was conducted following the local legislation and institutional requirements.
DM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, Software. SO: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing, Software. SI: Conceptualization, Methodology, Supervision, Writing – review & editing. SI: Conceptualization, Validation, Writing – review & editing. UO: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This project was funded by the A. G. Leventis Foundation Scholarship for master’s program awarded to DM.
Discussions with colleagues (Coker Ebenezer, Israel Deji Bolade and Adewale Awoyemi) helped in improving some aspects of the initial draft. We also acknowledge the moral support given by Master students 2020 set of the A.P. Leventis Ornithological Research Institute, during the research. This is paper number 225 from the A.P. Leventis Ornithological Research Institute.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1457049/full#supplementary-material
Ajibola-James O. O., Awotoye O. O., Sonibare J. A., Oroboade J. G. (2024). Assessing the ecological impact of industrial noise pollution on wildlife behavior in Okomu Forest Reserve, Nigeria. Discovery Environ. 2, 58. doi: 10.1007/s44274-024-00092-8
Arévalo J. E., Newhard K. (2011). Traffic noise affects forest bird species in a protected tropical forest. Rev. Bio. Trop. 59, 969–980.
Avalos G., Bermúdez E. (2016). Effect of a major highway on the spatial and temporal variation in the structure and diversity of the avifauna of a tropical premontane rain forest. Rev. Bio. Trop. 64, 1383–1399.
Awoyemi A. G., Ibáñez-Álamo J. D. (2023). Status of urban ecology in Africa: A systematic review. Landscape Urban Planning. 233, 104707. doi: 10.1016/j.landurbplan.2023.104707
Barber J. R., Crooks K. R., Fristrup K. M. (2010). The costs of chronic noise exposure for terrestrial organisms.Trends. Ecol. Evol. 25, 180–189. doi: 10.1016/j.tree.2009.08.002
Batáry P., Kurucz K., Suarez-Rubio M., Chamberlain D. E. (2018). Non-linearities in bird responses across urbanization gradients: A meta-analysis. Glob. Change Biol. 24, 1046–1054. doi: 10.1111/gcb.13964
Benítez-López A., Alkemade R., Verweij P. A. (2010). The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol. Conserv. 143, 1307–1316.
Callaghan C. T., Major R. E., Wilshire J. H., Martin J. M., Kingsford R. T., Cornwell W. K. (2019). Generalists are the most urban-tolerant of birds: a phylogenetically controlled analysis of ecological and life history traits using a novel continuous measure of bird responses to urbanization. Oikos 128, 845–858.
Carral-Murrieta C. O., Garcia-Arroyo M., Marin-Gomez O. H., Sosa-Lopez J. R., MacGregor-Fors I. (2020). Noisy environments: untangling the role of anthropogenic noise on bird species richness in a Neotropical city. Avian Res. 11, 32. doi: 10.1186/s40657-020-00218-5
Catterall C. P. (2009). “Responses of faunal assemblages to urbanisation: global research paradigms and an avian case study,” in Ecology of cities and towns: a comparative approach. Eds. McDonnell M. J., Hahs A. K., Breuste J. H. (Cambridge University Press, New York, New York, USA), 129–155.
Chamberlain D., Kibuule M., Skeen R., Pomeroy D. (2017). Trends in bird species richness, abundance and biomass along a tropical urbanization gradient. Urb. Ecos. 20, 629–638.
Danjuma D. F., Mwansat S. G., Manu A. S. (2012). The effects of forest fragmentation on species richness on the Obudu Plateau, south-eastern Nigeria. Afr. J. Ecol. 51, 32–36.
da Silva B. F., Pena J. C., Viana-Junior A. B., Vergne M., Pizo M. A. (2021). Noise and tree species richness modulate the bird community inhabiting small public urban green spaces of a Neotropical city. Urban Ecosyst. 24, 71–81. doi: 10.1007/s11252-020-01021-2
Dowling J. L., Luther D. A., Marra P. P. (2012). Comparative effects of urban development and anthropogenic noise on bird songs. Behav. Ecol. 23, 201–209. doi: 10.1093/beheco/arr176
Evans B., Reitsma R., Hurlbert A., Marra P. (2018). Environmental filtering of avian communities along a rural-to-urban gradient in Greater Washington, D.C., USA. Ecosphere. 9, e02402. doi: 10.1002/ecs2.2402
Federal Republic of Nigeria. (2012). Federal Republic of Nigeria 2006 population census. Available online at: https://web.archive.org/web/20120305101910/http://www.nigerianstat.gov.ng/nbsapps/Connections/Pop2006.pdf (Accessed April 2, 2024).
Fraixedas S., Lindén A., Piha M., Cabeza M., Gregory R., Lehikoinen A. (2020). A state-of-the-art review on birds as indicators of biodiversity: Advances, challenges, and future directions. Ecol. Indic. 118, 106728. doi: 10.1016/j.ecolind.2020.106728
Gagne S. A., Fahrig L. (2011). Do birds and beetles show similar responses to urbanization? Ecol. Appl. 21, 2297–2312.
Gregory R. D., Noble D., Field R., Marchant J., Raven M., Gibbons D. W. (2003). Using birds as indicators of biodiversity. Ornis Hung. 12-13, 11–24.
Habib L., Bayne E. M., Boutin S. (2007). Chronic industrial noise affects pairing success and age structure of ovenbirds (Seiurus aurocapilla). J. Appl. Ecol. 44, 176–184. doi: 10.1098/rsif.2019.0225
Halfwerk W., Holleman L. J. M., Lessells C. K. M., Slabbekoorn H. (2011). Negative impact of traffic noise on avian reproductive success. J. Appl. Ecol. 48, 210–219. doi: 10.1111/j.1365-2664.2010.01914.x
Hao Z., Zhang C., Li L., Gao B., Wu R., Pei N., et al. (2024). Anthropogenic noise and habitat structure shaping dominant frequency of bird sounds along urban gradients. iScience 27(2). doi: 10.1016/j.isci.2024.109056
Isaksson C. (2018). “Impact of Urbanization on Birds,” in Bird Species. Fascinating Life Sciences. Ed. Tietze D. (Springer, Cham). doi: 10.1007/978-3-319-91689-7_13
Kleist N. J., Guralnick R. P., Cruz A., Francis C. D. (2016). Anthropogenic noise weakens territorial response to intruder's songs. Ecosphere 7, e01259.
Kumdet P. S., Ivande S. T., Dami F. D. (2021). Key drivers of avifauna in greenspace of institutional campuses in a state in Western Africa. Urban For. Urban Green. 61, 127092.
Kurucz K., Purger J. J., Batary P. (2021). Urbanization shapes bird communities and nest survival, but not their food quantity. Glob. Ecol. Cons. 26, e01475. doi: 10.1016/j.gecco.2021.e01475
Luck G. W., Smallbone L. T. (2010). The impact of urbanization on taxonomic and functional similarity among bird communities. J. Biogeogr. 38, 849–906.
Luo J., Siemers B. M., Koselj K. (2015). How anthropogenic noise affects foraging. Glob. Change Biol. 29, 3278–3289.
Mainwaring M. C. (2015). The use of man-made structures as nesting sites by birds: A review of the costs and benefits. J. Nat. Conserv. 25, 17–22.
Mason J. T., McClure C. J. W., Baber J. (2016). Anthropogenic noise impairs owl hunting behavior. Biol. Conserv. 199, 29–32.
Matthew D. A., Ivande S. T., Ottosson U., Osinubi S. T. (2024). Difference in acoustic responses to urbanisation in two African passerines. Ostrich 95, 32–44.
McIntyre E. (2013). Anthropogenic noise and the communication of predation risk between parent and nestling tree swallows (Tachyneta bicolor). Dalhousie University, Halifax, Nova Scotia, 55 pp.
Morelli F., Benedetti Y., Su T., Zhou B., Moravec D., Šímová P., et al. (2017). Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: effects of land use and vegetation structure. Urban For. Urban Green. 23, 84–92. doi: 10.1016/j.ufug.2017.03.009
Narango D. L., Rodewald A. D. (2016). Urban-associated drivers of song variation along a rural–urban gradient. Behav. Ecol. 27, 608–616. doi: 10.1093/beheco/arv197
Neate-Clegg M. H. C., Tonelli B. A., Youngflesh C., Wu J. X., Montgomery G. A., Şekercioğlu Ç.H., et al. (2023). Traits shaping urban tolerance in birds differ around the world. Curr. Biol. 33, 1677–1688.e6. doi: 10.1016/j.cub.2023.03.024
Norton B. A., Evans K. L., Warren P. H. (2016). Urban biodiversity and landscape ecology: patterns processes and planning. Curr. Landsc. Ecol. Rep. 1, 178–192.
Osinubi S. T., Briskie J., Ottosson U., Brown J. A., Chapman H. M. (2012). Characterising calls of the Yellow-breasted Boubou (Laniarus atroflavus) and potential habitat effects. Proc. Pan-African Ornithol. Conf, 14–21.
Proppe D. S., Sturdy C. B., St. Clair C. C. (2013). Anthropogenic noise decreases urban songbird diversity and may contribute to homogenization. Glob. Change Biol. 19, 1075–1084.
Rashid G. M., Butt A., Qadir A., Ali M. H. (2020). Avian assemblage along an urban gradient: diversity, abundance and richness. Ornis Hung. 28, 49–64. doi: 10.2478/orhu-2020-0004
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org.
Rhodes M. L., Ryder T. B., Evans B. S., To J. C., Neslund E., Will C., et al. (2023). The effects of anthropogenic noise and urban habitats on song structure in a vocal mimic; the gray catbird (Dumetella carolinensis) sings higher frequencies in noisier habitats. Front. Ecol. Evol. 11. doi: 10.3389/fevo.2023.125263
Richards J. A. (1986). “Supervised Classification Techniques,” in Remote Sensing Digital Image Analysis (Springer, Berlin, Heidelberg). doi: 10.1007/978-3-662-02462-1_8
Rija A. A., Bugingo A., Said A., Mwamende K. A. (2014). Wet season bird species richness and diversity along urban-rural gradient in Morogoro municipality and surrounding areas, Tanzania. Tanz. J. For. Nat. Conserv. 83, 1–12.
Schroeder J., Nakagawa S., Cleasby I. R., Burke T. (2012). Passerine birds breeding under chronic noise experience reduced fitness. PloS One 7, e39200. doi: 10.1371/journal
Senzaki M., Kadoya T., Francis C. D. (2020). Direct and indirect effects of noise pollution alter biological communities in and near noise-exposed environments. Proc. Biol. Sci. 287, 20200176. doi: 10.1098/rspb.2020.0176
Senzaki M., Yamaura Y., Francis C. D., Nakamura F. (2016). Traffic noise reduces foraging efficiency in wild owls. Sci. Rep. 6. doi: 10.1038/srep30602
Seress G., Liker A. (2015). Habitat urbanization and its effects on birds. Acta Zool. Acad. Sci. 61, 373–408. doi: 10.17109/AZH.61.4.373.2015
Slabbekoorn H. (2017). Anthropogenic noise: impacts on animals. Reference Mod. Life Sci. doi: 10.1016/b978-0-12-809633-8.01100-6
Slabbekoorn H., Peet M. (2003). Birds sing at a higher pitch in urban noise. Nature 424, 267. doi: 10.1038/424267a
Slabbekoorn H., Smith T. B. (2002). Habitat-dependent song divergence in the little greenbul: an analysis of environmental selection pressures on acoustic signals. Evolution. Int. J. Org. Evol. 56, 1849–1858.
Slabbekoorn H., Yeh P., Hunt K. (2007). Sound transmission and song divergence: a comparison of urban and forest acoustics. Condor 109, 67–78. doi: 10.1093/condor/109.1.67
United Nations, Department of Economic and Social Affairs, Population Division (2018). World Urbanization Prospects: The 2018 Revision. Available online at: https://esa.un.org/unpd/wup/Publications (Accessed January 30, 2024).
Wiącek J., Polak M. (2015). Does traffic noise affect the distribution and abundance of wintering birds in a managed woodland? Acta Ornithol. 50, 233–245. doi: 10.3161/00016454AO2015.50.2.011
Winkler H. (2001). “The ecology of avian acoustical signals,” in Ecol. Sens. Eds. Barth F. G., Schmid A. (Springer, Berlin), 79–104. doi: 10.1007/978-3-662-22644-5_5
Keywords: bird community, urbanization, anthropogenic noise, physical structure, rural-urban gradient, Nigeria, Afro-tropics
Citation: Matthew DA, Osinubi ST, Ivande ST, Iwajomo SB and Ottosson U (2024) Structural and acoustic properties of urbanized landscapes adversely affect bird communities in a tropical environment. Front. Ecol. Evol. 12:1457049. doi: 10.3389/fevo.2024.1457049
Received: 30 June 2024; Accepted: 21 October 2024;
Published: 19 November 2024.
Edited by:
Rubén Ortega-Álvarez, National Autonomous University of Mexico, MexicoReviewed by:
Chan Ryul Park, National Institute of Forest Sicence (NIFoS), Republic of KoreaCopyright © 2024 Matthew, Osinubi, Ivande, Iwajomo and Ottosson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dickson Anoibi Matthew, YW5vaWJpbWF0dGhld0BnbWFpbC5jb20=
†Present address: Samuel Tertese Ivande, Global Center for Species Survival, Indianapolis Zoo, Indianapolis, IN, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.