 Jaime A. Garizábal-Carmona1,2
Jaime A. Garizábal-Carmona1,2 Jefry S. Betancur3
Jefry S. Betancur3 Sergio Montoya-Arango4
Sergio Montoya-Arango4 Laura Franco-Espinosa5N. Javier Mancera-Rodríguez1*
Laura Franco-Espinosa5N. Javier Mancera-Rodríguez1*- 1Semillero de Ecología Urbana, Grupo de Investigación Ecología y Conservación de Fauna Silvestre, Departamento de Ciencias Forestales, Universidad Nacional de Colombia, Medellín, Colombia
- 2Corporación Merceditas, Medellín, Colombia
- 3Grupo de Investigación de Ecología y Evolución de Vertebrados, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, Colombia
- 4Faunativa S.A.S., Medellín, Colombia
- 5Independent Researcher, Medellín, Colombia
Introduction: Categorizing species according to their frequencies across urbanization levels and identifying some species traits that explain this variation could be a valuable tool for focusing conservation efforts, particularly in biodiversity hotspots with high endemism rates. This study proposes a semi-quantitative and multi-scale protocol to categorize bird species as urban avoiders, utilizers, and dwellers, based on their frequencies at different urbanization levels. Additionally, it evaluates the relationships of these categories with altitudinal ranges, trophic guilds, and foraging strata.
Methods: We performed bird counts in 124 points located within urban areas, and in 15 points located in non-urban areas of a Colombian Northern Andean city (Medellín and surroundings). Each urban point was assigned to urbanization levels based on 200, 500, and 1,000 m buffers categorized as high (67–100% of built cover), moderate (34–66% of built cover), or low (0–33% of built cover).
Results: We categorized 103 bird species: 49 as urban avoiders, 31 as urban utilizers, and 23 as urban dwellers. The two recorded Colombian endemic species and seven near-endemics were categorized as urban avoiders, with only one near-endemic species categorized as an urban utilizer (the other three were data deficient). Furthermore, most bird species with exclusive Andean distribution were categorized as urban avoiders (78.57%). Urban avoiders had narrower altitudinal ranges (1,969 ± 524 m) than utilizers (2,287 ± 592m) and dwellers (2,569 ± 654m), and they had the largest proportion of frugivorous and frugivorous-insectivorous species, while urban dwellers had a greater proportion of omnivorous species. Overall, bird species with exclusive Andean distribution and narrow altitudinal ranges are the most threatened by urban sprawl, irrespective of their trophic guild or foraging strata.
Discussion: This study emphasizes the importance of protecting native forest remnants in urban surroundings for conserving native Andean bird species, as urban green spaces in high-density cities may not sufficiently support their long-term survival and reproduction. Also, it highlights the need to identify conservation priorities based on local biodiversity patterns, taking into account that species-specific urban tolerance depends on particular landscape dynamics and species regional pools.
1 Introduction
More than half of world human population now resides in cities, with urban sprawl increasing significantly in recent decades (Seto et al., 2011). This expansion has adverse effects on natural ecosystems and biodiversity (Pauchard et al., 2006; McKinney, 2008; Aronson et al., 2014). Conservation concerns regarding urban sprawl increase in mountain biodiversity hotspots such as Tropical Andes, due to the high density of human population established in places with a high rates of endemism (Cincotta et al., 2000; Rahbek et al., 2019). These urban regions are often overlooked by conservation efforts that primarily focus on pristine ecosystems (Mcdonald et al., 2008; Buchanan et al., 2011). Consequently, in biodiverse Neotropical countries such as Colombia, research efforts often prioritize less human-perturbed ecosystems (Martin et al., 2012; Arbeláez-Cortés, 2013), while conservation strategies tend to concentrate on endangered species at the national scale (Renjifo et al., 2020).
As urban population continue to grow, the importance of biodiversity conservation within urban ecosystems becomes increasingly evident (Miller and Hobbs, 2002; Mcdonald et al., 2009; Puppim de Oliveira et al., 2011; Chin et al., 2022). However, in northern South America, knowledge gaps persist regarding the effects of urbanization on biodiversity, representing a limitation for effective conservation efforts (Ortega-Álvarez and MacGregor-Fors, 2011). Moreover, identifying the consequences of habitat loss and environmental pressures on specific species can be challenging, especially when only a few species in the assemblage are considered endangered at national or global levels. To address these challenges, Fischer et al. (2015) proposed a theoretical framework for categorizing bird species based on their tolerance to urbanization. This framework distinguishes between urban dwellers (i.e., high tolerance), urban utilizers (i.e., intermediate tolerance), and urban avoiders (i.e., low tolerance). Categorizing species according to this framework could help to identify conservation priorities within urban regions. Furthermore, identifying the features that explain urban tolerance can aid in predicting which species are most vulnerable to urbanization pressures. This knowledge can inform the design of conservation strategies focused on specific species or environmental characteristics that mitigate the impacts of urbanization (Goddard et al., 2010; Aronson et al., 2017; Threlfall et al., 2017).
Giving the significance of multiple spatial scales in modulating urban biodiversity patterns (e.g., Jokimäki and Kaisanlahti-Jokimäki, 2003; Conole and Kirkpatrick, 2011; Concepción et al., 2015; Xie et al., 2016; Tryjanowski et al., 2017; Sidemo-Holm et al., 2022), this study conducted in urban and non-urban areas of a Northern Andean city (Medellín and surroundings) aimed to: (i) propose a semi-quantitative and multi-scale protocol to categorize bird species as urban avoiders, utilizers, and dwellers, based on their frequencies at different urbanization levels, and (ii) evaluate relationships of these categories with altitudinal ranges, trophic guilds, and foraging strata of bird species. We hypothesized that bird species with wider altitudinal ranges would exhibit higher urban tolerance, potentially link to the ability to tolerate greater environmental heterogeneity (Ruggiero, 2001). Similarly, we hypothesized that bird species with certain trophic guilds, such as omnivorous and granivorous, and foraging strategies encompassing the use of multiple strata might exhibit greater urban tolerance, due to the related capacity of exploiting diverse and novel food resources, reflecting higher ecological plasticity (Evans et al., 2011; González-Lagos and Quesada, 2017). Therefore, we predicted that bird species with narrower altitudinal ranges, more specialized diets, and limited foraging strata would be categorized as urban avoiders, whereas those with broader altitudinal ranges, more generalized diets, and flexible foraging strata would be categorized as urban utilizers or dwellers.
2 Materials and methods
2.1 Study area
The city of Medellín and nine adjacent municipalities (referred to as “Medellín” hereafter) are located in northern central Andes of Colombia (6.26029 North, −75.574139 West), with an urban area mainly established between 1,400 and 1,700 m.a.s.l (Schnitter et al., 2006) (Figure 1). Medellín is one of the most densely populated cities in northern South America with approximately 21,000 people/km2 (Parés-Ramos et al., 2013), with an overall human population of approximately four million people. Our study area represented the urban core and adjacent non-urban areas of Medellín, which were defined using a buffer of 2,000 m from the city limits.
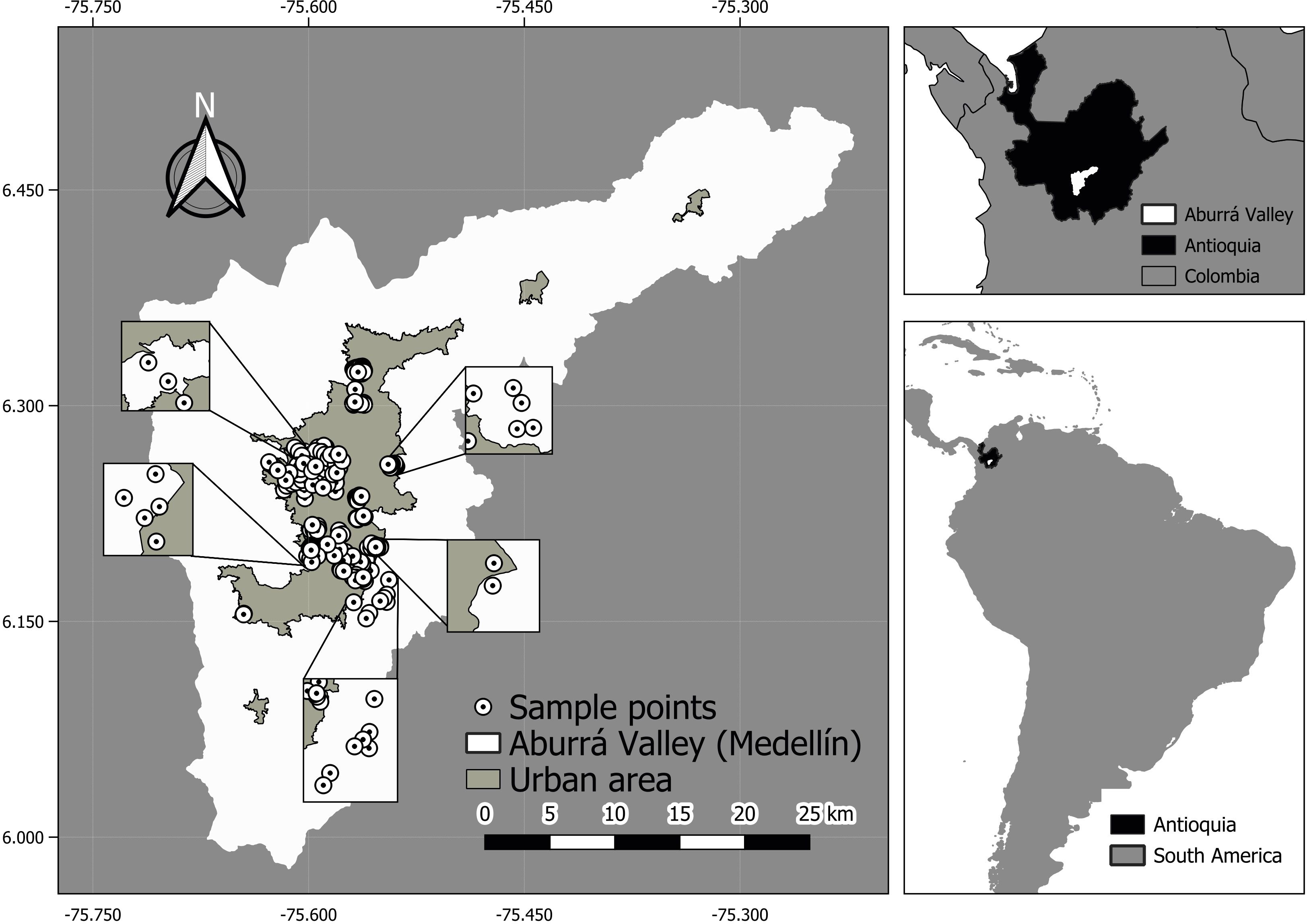
Figure 1. Study area and sampled points located across urban and non-urban areas of Medellín and surrounding municipalities, Colombian northern Andes.
Besides urban green spaces and other green cover within city limits, urban core represented areas occupied by built cover such as buildings, roads, and other human infrastructure that reached up to 75% of the city by 2015 (Paniagua-Villada et al., 2024). These areas across the city were assigned to categorical values proposed by MacGregor-Fors (2011) when measuring urbanization levels: highly developed (67–100% of built cover, i.e., high urbanization level), moderately developed (34–66% of built cover, i.e., intermediate urbanization level), or low urban developed areas (0–33% of built cover, i.e., low urbanization level). These percentages were estimated based on land cover reclassification, using the Tasseled cap index, using bands on brightness, greenness, and wetness to differentiate between built, grass, and tree cover (Dymond et al., 2002; Samarawickrama et al., 2017). This cover reclassification used a 10 × 10 m resolution satellite image taken on 22 February 2019 by Sentinel-2 (USGS EROS Archive). Non-urban areas corresponded to sparsely developed areas dominated by lawns, herbaceous croplands, cultivated human-consumed herbaceous plants, exotic tree plantations, and some native forest remnants.
2.2 Bird surveys
To assess bird species categorization based on species frequency, we designed a sampling strategy following recommendations by Bibby et al. (1998) and Sutherland (2006). We used data from 139-point counts situated between 1,486 and 2,351 m.a.s.l. in both urban (n = 124) and non-urban areas of Medellín (n = 15). All points were located at least 200 m from each other to avoid bird re-counting. The dataset we used resulted from a posteriori data compilation of sampling conducted between January 2014 and June 2019, including both dry and rainy seasons. Each point had a 25 m fixed radius and was visited for 10-min, four times within the same week, exclusively during favorable weather conditions (particularly no rain), within the time frame of 06:00 to 10:00. Total observation effort was 5,560 minutes (556 total visits). As bird surveys were always diurnal and included months when migratory species were absent, we excluded migratory and nocturnal species, as well as overflying individuals that weren’t directly using the sampling habitats. Species that were found in only one sampling point were also excluded, as they were considered accidental, along with bird species found in non-urban areas whose altitudinal distribution did not coincide with the altitude of the urban core (i.e., species whose maximum altitudinal distribution was under 1,400 m.a.s.l. or whose minimum altitudinal distribution was above 1,700 m.a.s.l.). The last were excluded because no evidence of urban tolerance could be interpreted from their absence within the city, which could be better explained by their altitudinal distribution.
2.3 Distribution, conservation status, trophic guilds, and foraging strata
We assigned altitudinal ranges according to Ayerbe-Quiñones (2018), endemism to Chaparro-Herrera et al. (2024), migratory status to Naranjo et al. (2012), and conservation status to Renjifo et al. (2014). We assigned trophic guilds and foraging strata categories based on Wilman et al. (2014), which is a compilation of bird ecological traits at the species level based on The Handbook of the Birds of the World (del Hoyo et al., 1992). They provided a data frame specifying percentage of diets and foraging strata at 10% increments. Based on this information, we assigned categories as follows: only one diet when some item had ≥ 80%, two simultaneous diets when there were two items with at least 30% each, and Omnivorous when there were three or more types of food and none of them had ≥ 80%. Similarly, we assigned a single foraging stratum when one stratum had ≥ 80%, two simultaneous strata when there were two with at least 30% each, and Multiple when there were three or more strata present and none of them had ≥ 80%.
2.4 Categorization protocol and data analysis
To categorize bird species as urban avoiders, urban utilizers, or urban dwellers, sensu Fischer et al. (2015), we considered four criteria and rated them on a scale from 1 to 3. We assigned values based on species frequencies at each point, according to comparisons between urban and non-urban areas frequencies, and frequencies at each urban point according to its urbanization level at 200, 500, and 1,000 m buffers. The lowest value (1) was assigned to the presumably least tolerant species (most frequent in sites with lower urbanization levels or non-urban areas), while the highest value (3) was assigned to the presumably most tolerant species for each evaluated criterion at each spatial scale (most frequent in sites with higher urbanization levels). The intermediate value (2) was assigned to species that are presumably moderate in their tolerance to urbanization (most frequent in sites with intermediate urbanization levels or similar values across the urbanization gradient) (see Table 1).
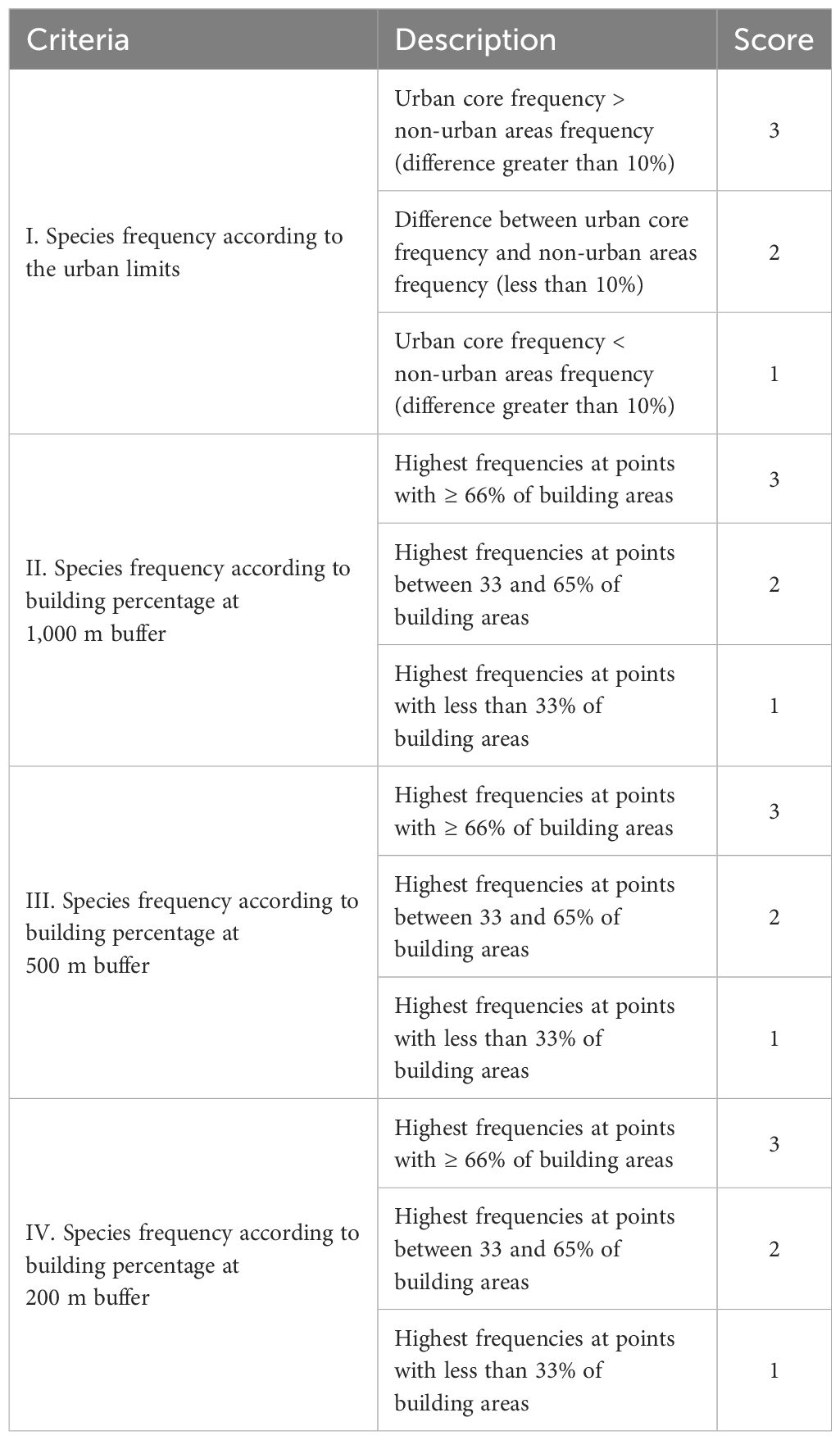
Table 1. Categorization criteria and scores for bird species according to frequencies at 139-point counts sampled across urban and non-urban areas of Medellín and surrounding municipalities, Colombian northern Andes.
Minimum and maximum total scores were 4 and 12, respectively, and thus, we divided by equal numerical ranges to assign the final category as follows: urban avoider (4 to 6), urban utilizer (7 to 9), and urban dweller (10 to 12). Urban avoiders would represent species that inhabit exclusively or mainly in non-urban environments, with only isolated records within urban environments (i.e., without urban populations). Urban utilizers would represent species that inhabit both urban and non-urban environments, but still depend on metapopulation dynamics (i.e., urban populations depending on non-urban populations). Finally, urban dwellers would represent species that inhabit mainly in urban environments, where their populations thrive (i.e., long-term urban populations without any dependence on non-urban populations).
We plotted incidence-based rarefaction curves using the “iNEXT” R package (Hsieh et al., 2016) and verified that surveyed points in non-urban areas (n = 15) and urban areas at 200, 500, and 1,000 m (n = 124) suggested representative sampling (bootstrap = 1,000 repetitions and confidence intervals = 84%, see MacGregor-Fors and Payton, 2013). In urban areas, each spatial scale was independently addressed from the same 124 sampling points at each buffer, as follows: low (n = 49), intermediate (n = 59), and high urbanization levels at 200 (n = 16); low (n = 32), intermediate (76), and high urbanization levels at 500 (n = 16); and low (n = 30), intermediate (n = 81), and high urbanization levels at 1,000 m (n = 13) (Supplementary Figure 1). After validating normal distribution, and testing homoscedasticity and leverage using the “stats” package in R software (R Core Team, 2019), we compared altitudinal ranges among categories of urbanization tolerance using an ANOVA test and Tukey Honest Significant Differences (Tukey-HSD). Tukey-HSD analysis provides p-values after adjustment for the multiple comparisons, reducing the effects of sample size differences between groups (Miller, 1981). Then, we ran a General Linear Model (GLM) with Poisson distribution to test the relation between species tolerances to urbanization (score as the response variable) and altitudinal range (as the explicative variable). The altitudinal range was considered as the difference between the maximum and minimum reported elevations for each species, according to Ayerbe-Quiñones (2018). In the case of categorical variables, we performed the non-parametric Pearson’s Chi-square (X2) and Fisher’s exact tests for evaluating whether trophic guilds or foraging strata explained differences in urbanization tolerance. The last test was performed using the “exact2x2” R package (Fay, 2010) to corroborate statistical relationships for contingency tables with some small frequencies (< 5).
3 Results
We recorded 139 bird species, 103 of which were categorized: 49 as urban avoiders (47.6%), 31 as urban utilizers (30.1%), and 23 as urban dwellers (22.3%) (Supplementary Table 1). We considered the other species as “Data Deficient”. According to our defined criteria, the two recorded Colombian endemic species (Hypopyrrhus pyrohypogaster and Ortalis columbiana), and seven near-endemic species (Chlorostilbon melanorhynchus, Ramphocelus flammigerus, Saucerottia saucerottei, Forpus conspicillatus, Pheugopedius mystacalis, Stilpnia vitriolina, and Tangara labradorides) were categorized as urban avoiders. Only one near-endemic species (Thamnophilus multistriatus) was categorized as urban utilizer and the other three species with narrow geographic distribution were considered Data Deficient (Cyanocorax affinis, Cyclarhis nigrirostris, and Saltator atripennis; also, near-endemics). Most species categorized as urban avoiders (33 of 49: 67.35%) have an exclusive Andean distribution, comparing with 8 of 31 species categorized as urban utilizers (25.81%) and 1 of 23 species categorized as urban dwellers (4.35%) (Table 2). In addition, the only two exotic species recorded were categorized as utilizer (Bubulcus ibis) or as dweller (Columba livia). The latter was more frequent in highly developed areas (> 66% covered by buildings or impervious surfaces).
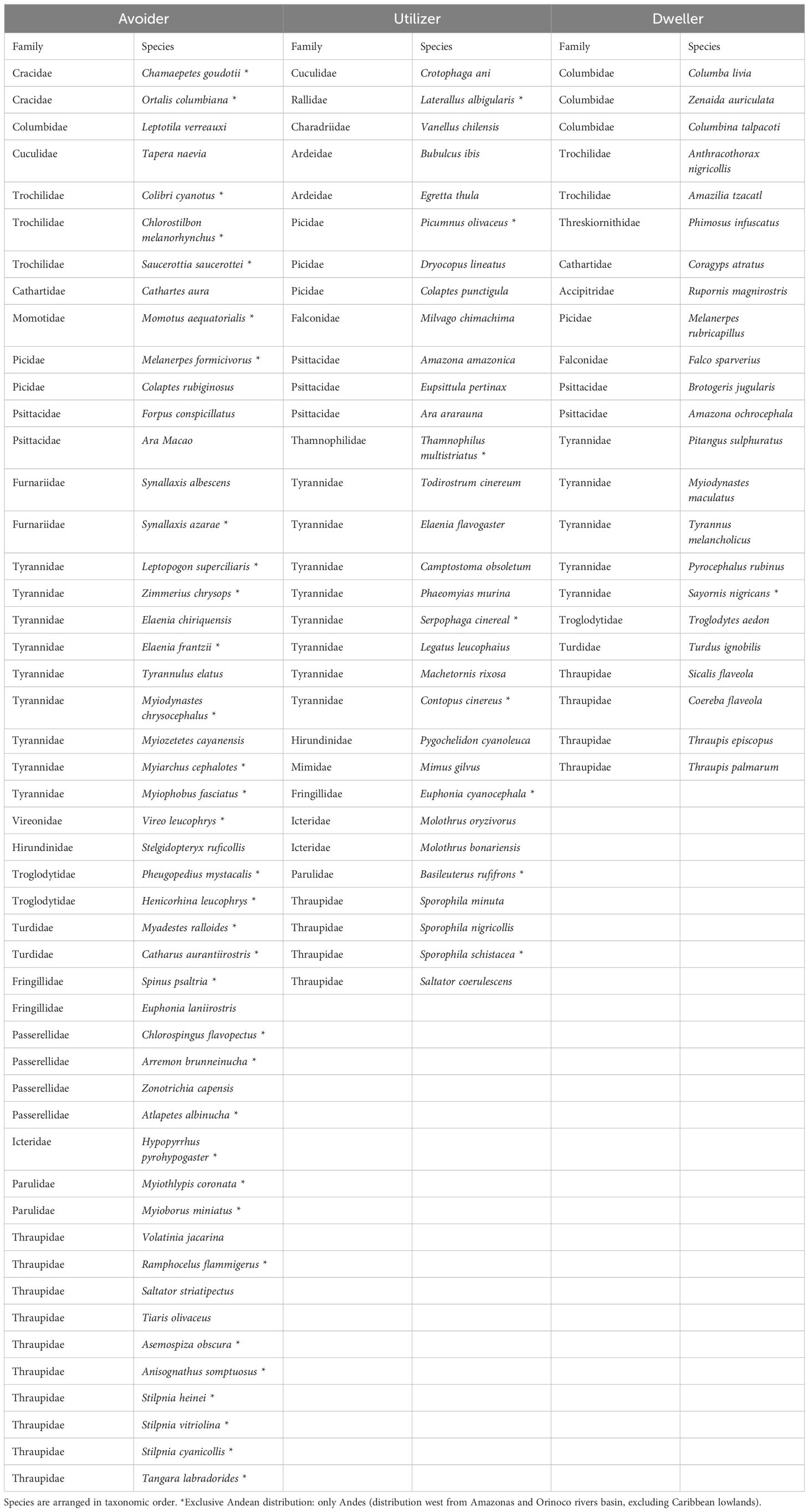
Table 2. Bird species categorization to dweller, utilizer or avoider, based on species frequency and data collected at 139-point counts located in urban (n = 124) and non-urban areas (n = 15) of Medellín and surrounding municipalities, Colombian northern Andes, between 2014 and 2019.
3.1 Altitudinal range and urban tolerance
Bird species exhibited altitudinal ranges of 2,221 ± 654 m, with significant differences observed between categories (F = 9.02, df = 2, p < 0.05). The Tukey-HSD test suggested not overlapping altitudinal ranges between urban avoiders and dwellers (adjusted p < 0.001), which represented the tolerance extremes across the urbanization gradient (low and high tolerance, respectively), and between urban avoiders and utilizers (adjusted p-value < 0.05). However, we found no significant differences between urban utilizers and dwellers (adjusted p = 0.18) (Figure 2). On average, urban avoiders had altitudinal ranges of 1,969 ± 524 m, whereas utilizers had altitudinal ranges of 2,287 ± 592 m and dwellers of 2,569 ± 645 m. Tolerance to urbanization of bird species increased as a function of altitudinal ranges (GLM: β = 0.0401 ± 0.0008, p < 0.001; R2 = 0.16), with higher ranges suggesting higher urbanization tolerance.
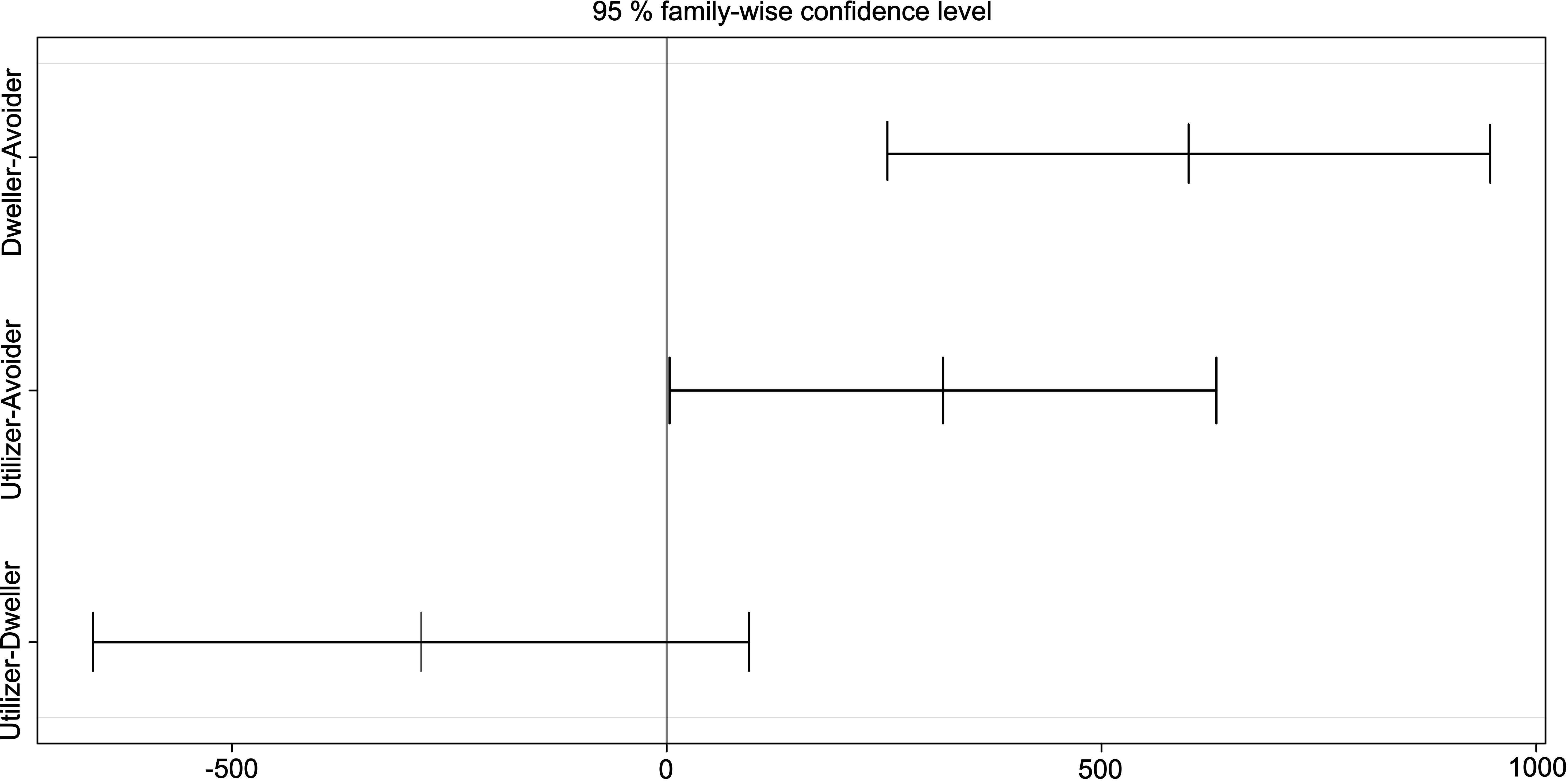
Figure 2. Tukey’s HSD test plot using altitudinal ranges and urban tolerance categories (avoider, utilizer, and dweller), for bird resident species assemblages across urban and non-urban areas of Medellín and surrounding municipalities, Colombian northern Andes. Analysis based upon data collected at 139-point counts located in urban (n = 124) and non-urban areas (n = 15), between 2014 and 2019.
3.2 Trophic guilds and foraging strata according to urban categorization
We assigned more than half of categorized bird species to Insectivorous (32.04%) or Omnivorous (26.21%). Others trophic guilds were represented by Frugivorous-Insectivorous (14.56%), Granivorous (7.77%), and Frugivorous (6.80%). The rest of trophic guilds were represented by five or fewer bird species (maximum 4.85% each). Urban avoiders had more Insectivorous (30.61%) and Omnivorous (24.49%), and the largest proportion of Frugivorous (10.20%) and Frugivorous-Insectivorous (20.41%). Contrastingly, urban utilizers had the largest proportion of Insectivorous (41.94%), while urban dwellers had the largest proportion of Omnivorous (34.78%), and the fewer of Insectivorous (21.74%) and Frugivorous-Insectivorous (4.35%) (Figure 3A). Regarding foraging strata, the most common among bird species was Multiple (35.92%), followed by Ground (15.53%), Understory-Midstory (10.68%), and Midstory-Canopy (10.68%), with the rest grouping nine or fewer species (maximum 8.74% each). Differences between categories in foraging strata percentages were less clear than in trophic guilds, with urban avoiders and dwellers including more species assigned to the Multiple strata than urban utilizers. Urban avoiders showed the largest proportion of Understory-Midstory (Figure 3B), and urban utilizers had the largest proportion of species assigned to Ground, Ground-Understory and Understory-Midstory (Figure 3B). Otherwise, dwellers had the fewest species assigned to Ground-Understory and the highest to Understory (Figure 3B). However, after evaluating statistical differences between categories, urban tolerance of bird species did not depend on trophic guild (X2 = 21.61, df = 18, p-value = 0.25, and p-value of Fisher’s exact test = 0.18) or foraging strata (X2 = 19.84, df = 18, p-value = 0.35, and p-value of Fisher’s exact test = 0.50).
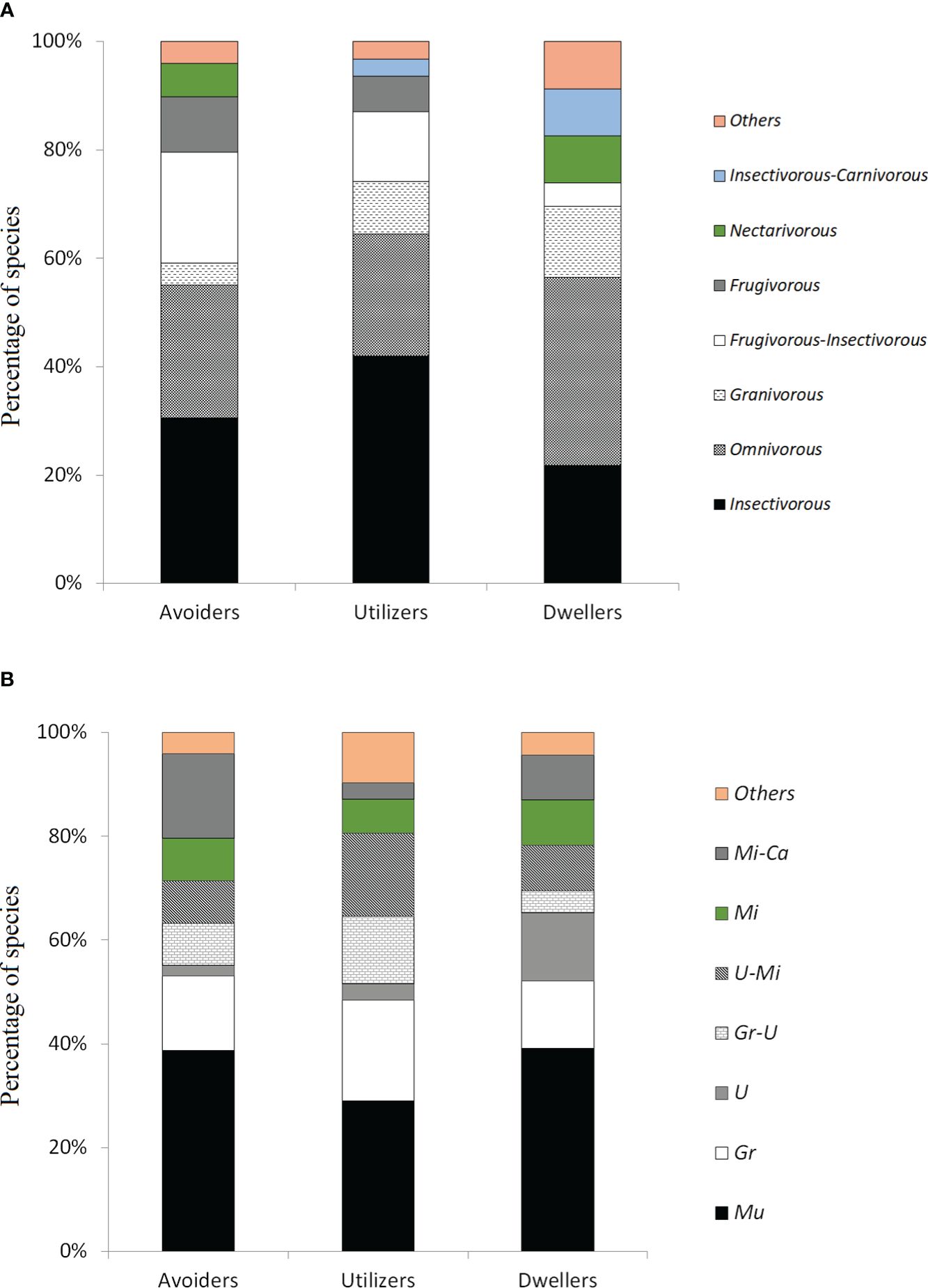
Figure 3. Percentage of bird species by trophic guild (A), and by foraging strata (B) according to urban categorization: urban avoiders (n=49 spp.), utilizers (n=31 spp.) and dwellers (n=23 spp.). Mu: Multiple, Gr: Ground, U: Understory Gr-U: Ground-Understory, U-Mi: Understory- Midstory, Mi: Midstory, Mi-Ca: Midstory-Canopy; “Others” includes: Canopy, Water, and Water-Ground.
4 Discussion
The bird species richness we recorded in this study is a relatively high number compared to other urban studies worldwide (Mills et al., 1989; Tryjanowski et al., 2017; Callaghan et al., 2019), and also most Neotropical ones (Charre et al., 2013; de Castro Pena et al., 2017; Lees and Moura, 2017; Leveau et al., 2017; Carvajal-Castro et al., 2019). Nevertheless, in most Neotropical cities facing similar biogeographic and socioeconomic scenarios, bird assemblages have been described after urbanization has already transformed the natural landscape. As a result, the least tolerant species to urbanization have already disappeared from most developed areas (Stiles, 1990; Biamonte et al., 2011; Escobar-Ibáñez and Macgregor-Fors, 2016). This suggests that a significant proportion of the regional species pool of Medellín and its surrounding landscape has already been filtered out from urban areas, which was indirectly confirmed by the presence of bird species recorded exclusively in non-urban areas (i.e., urban avoiders), aligning with findings from other studies in the region that employed a space-for-time substitution approach (Garizábal-Carmona et al., 2023). Therefore, the decrease of biodiversity may not solely be a matter of the number of species, but rather which species are disappearing, especially as urbanization tends to homogenize landscapes and biotas (McKinney, 2006).
As predicted, we observed a positive relationship between bird species’ urban tolerance and altitudinal ranges, consistent with findings from other cities (Bonier et al., 2007). In Medellín, urban avoiders included the two recorded endemic species and seven near-endemic ones, as well as other native species exclusively distributed across the Tropical Andes. This aligns with regional biodiversity patterns, where species with the smallest geographic ranges tend to peak in mountain ecosystems (Rahbek et al., 2019). The composition of urban avoiders in Medellín encompassed bird species with diverse diets and foraging strata, which may partially explain why these traits were less conclusive in our analysis. Based on these results, it becomes necessary to design complementary strategies for species with varying urban tolerance at both landscape and local scales, as suggested by other authors (Melles et al., 2003; Shwartz et al., 2008; Kong et al., 2010). In Medellín, conserving Andean forest remnants in the city surroundings would likely be crucial for maintaining viable populations of urban avoiders, as creating suitable habitats within the city, especially in terms of patch size, would be challenging (Garizábal-Carmona and Mancera-Rodríguez, 2021). Interventions aimed at enhancing urban green cover at both landscape and local levels may prove to be more efficient for targeting urban utilizers and some avoider species that can be found within the city, especially at sites with higher representation of native vegetation and less human intervention (Garizábal-Carmona and Mancera-Rodríguez, 2021). However, in both cases, it would be important to consider other characteristics in tropical urban regions, such as socioeconomic inequality, where these human-related dynamics also play a pivotal role (Villaseñor et al., 2024).
The rapid landscape transformation and loss of native forest remnants across most of the Northern Andean region (Quintero et al., 2017), underscores the urgency of rapidly identifying species that persist in the regional pool but face high risks of local extinctions driven by urbanization. Urban categorization of species could serve as a key tool in developing more efficient local and regional conservation priorities, by focusing on certain species or taxonomical groups beyond the conservation status at national or global levels. For instance, bird families such as Cotingidae, Furnariidae, Passerellidae, Grallaridae, Odontophoridae, and Rhynocryptidae, which are predominantly rare within urban areas, but fairly common and highly diverse across less perturbated ecosystems of Colombian Andes (Hilty and Brown, 1986; Castaño-Villa and Patiño-Zabala, 2007; Ayerbe-Quiñones, 2018), could benefit from this approach. Therefore, urban tolerance categories can provide urban planners with valuable tools to concentrate conservation efforts on these most vulnerable birds and the environmental characteristics that would support their presence within cities or their surroundings (e.g., Aronson et al., 2017; Threlfall et al., 2017; Beaugeard et al., 2021).
Additionally, understanding the use of food resources and foraging strata could provide valuable insights for making informed decisions regarding vegetation management, such as selecting appropriate plants and determining how they should be managed (Chong et al., 2014; Aronson et al., 2017; Campos-Silva and Piratelli, 2021). However, our study revealed a weak relationship between these traits and urban tolerance, indicating the need for future research to focus on functional diversity and evaluate direct relationships between site-scale characteristics and functional traits. Given that our analysis was constrained to landscape scales (≥ 200 m) and relied on literature-based assessments of diets and foraging strata (Wilman et al., 2014), our ability to draw conclusions regarding these issues was limited. Furthermore, patterns related to diet and foraging strata were inconclusive, probably due to the lack of control for functional differences related to body size (Ortega-Álvarez and MacGregor-Fors, 2009; Estevo et al., 2017), as well as the absence of direct information on other highly variable traits, given the low representation of Neotropical birds in global open data (e.g., Tobias et al., 2022). Secondary information could be a limitation because species commonly found in urban ecosystems exhibit flexibility in behavioral traits (Shochat et al., 2006) and their foraging strategies are influenced by factors such as habitat use and human presence (McKinney, 2006), which are often unknown for most Neotropical species in urban environments (González-Lagos and Quesada, 2017; Rega-Brodsky et al., 2022).
Urban utilizers, which could play a crucial role in biodiversity conservation within highly dense urban landscapes due to their moderate urban tolerance, exhibited a wide range of ecological traits. However, our findings revealed that most utilizer species were native birds typically inhabiting open green areas thriving in both urban and non-urban environments. Utilizer species in our system were frequent at large and medium-size green spaces (between 5 and 50 ha), as well as in some small-size green spaces (< 5 ha) dominated by grass or clustered urban trees (i.e., > 50% of the area). This aligns with findings from other studies indicating that bird assemblages in the Colombian Andes are increasingly dominated by widely distributed Neotropical species favored by agricultural and urban sprawl (Avendaño et al., 2013). Indeed, regarding altitudinal ranges, utilizer species exhibited similarities to dweller species, while both groups were distinct from urban avoiders.
Urban dwellers comprised bird species widely distributed across the city, including areas where trees or grass represented less than 25%. These species were typically associated with isolated vegetation or related to buildings or places with high human activities, as it is a common trait for species considered highly tolerant to urbanization (Evans et al., 2011; Sol et al., 2014; Barnett et al., 2023; Neate-Clegg et al., 2023). Most of the species categorized as urban dwellers in our study are widely distributed Neotropical native birds found in lowlands, a pattern observed in other Colombian Andean cities (Carvajal-Castro et al., 2019; Echeverry-Galvis et al., 2023). Notably, the only non-native species categorized as an urban dweller was Columba livia (Rock Dove), which is usually one of the most common bird species across all studied Neotropical cities (e.g. Ortega-Álvarez and MacGregor-Fors, 2011; Sanz and Caula, 2015; Bellocq et al., 2017). These urban dweller species are also prevalent in the bird assemblages of Caribbean cities and other Neotropical cities located below 500 m.a.s.l., primarily found west of the Andes (Barbosa de Toledo et al., 2012; Avendaño et al., 2013; Elías Domínguez-López and Ortega-Álvarez, 2014; Sanz and Caula, 2015; de Castro Pena et al., 2017). They typically increase in frequency when rapid environmental changes occur, as a result of the increasing emerging novel resources and habitats, including bird feeders (Clergeau et al., 1998; Conole and Kirkpatrick, 2011; Kowarik, 2011; Møller et al., 2015; Ouyang et al., 2018). Therefore, these highly tolerant species are likely to benefit the most from continued urban sprawl across the Tropical Andes.
The most common resident Neotropical species, categorized as urban dwellers or utilizers, are likely to include highly or moderately tolerant species to urbanization. However, the vulnerability of some low-tolerance resident Neotropical species to urbanization, categorized as urban avoiders, data deficient, or those that were absent in our surveys, requires most robust sampling designs that minimize spatial and temporal biases, and improve sampling balance across environmental conditions (i.e., more similar sampling sizes). This could be achieved, for example, by increasing sampling efforts across less perturbed areas in city adjacencies or across new emergent Andean cities. However, despite challenges in understanding the proximate and ultimate drivers of bird tolerance to urbanization, our study provides valuable insights into the responses of certain bird species to urbanization in highly biodiverse urbanized landscapes. The categorization method proposed in our study could serve as a useful tool for identifying the most vulnerable species to urbanization, thereby facilitating strategies to mitigate the loss of beta-diversity and phylogenetic diversity (La Sorte et al., 2007; Blair and Johnson, 2008; Ferenc et al., 2014; Morelli et al., 2016; Leveau et al., 2017). However, addressing this conservation challenge remains complex, particularly in mountain regions like the northern Andes, where densely urban sprawl intersects with unique and still understudied biodiversity hotspots (Cincotta et al., 2000; Bax and Francesconi, 2019; Rahbek et al., 2019).
Data availability statement
The data presented in the study are deposited in the Zenodo repository, accession link, as open source, at: https://zenodo.org/records/12763590.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because our research is based on observational records only, without any intervention or direct manipulation of wild animals.
Author contributions
JG-C: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. JB: Data curation, Formal analysis, Investigation, Visualization, Writing – review & editing. SM-A: Data curation, Validation, Writing – review & editing. LF-E: Data curation, Validation, Writing – review & editing. NM-R: Conceptualization, Investigation, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Acknowledgments
We thank the executive director of Corporación Merceditas, Rómulo Agudelo, for administrative and logistic support, and Paubla Otálvaro for field support. We also thank Rubén Ortega-Álvarez and Duván Canal for comments that help us to improve this manuscript in preliminary versions, and two reviewers who gave us comments in a previous submitting process. Finally, we thank Natalia Ruiz for GIS support.
Conflict of interest
Author JG-C was employed by Corporación Merceditas. Author SM-A was employed by Faunativa S.A.S.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1432340/full#supplementary-material
References
Arbeláez-Cortés E. (2013). Knowledge of Colombian biodiversity: published and indexed. Biodiversity Conserv. 22, 2875–2906. doi: 10.1007/s10531-013-0560-y
Aronson M. F. J., La Sorte F. A., Nilon C. H., Katti M., Goddard M. A., Lepczyk C. A., et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. 281, 1–8. doi: 10.1098/rspb.2013.3330
Aronson M. F. J., Lepczyk C. A., Evans K. L., Goddard M. A., Lerman S. B., MacIvor J. S., et al. (2017). Biodiversity in the city: key challenges for urban green space management. Front. Ecol. Environ. 15, 189–196. doi: 10.1002/fee.1480
Avendaño J. E., Cortés-Herrera J. O., Briceño-Lara E. R., Rincón-Guarín D. A. (2013). Crossing or bypassing the Andes: a commentary on recent range extensions of cis-Andean birds to the West of the Andes of Colombia. Orinoquia 17, 207–214. doi: 10.22579/20112629.18
Ayerbe-Quiñones F. (2018). Guía ilustrada de la avifauna Colombiana (An illustrated field guide to the birds of Colombia) (Cali, Colombia: WSC).
Barbosa de Toledo M. C., Donatelli R. J., Teixeira Batista G. (2012). Relation between green spaces and bird community structure in an urban area in Southeast Brazil. Urban Ecosyst. 15, 111–131. doi: 10.1007/s11252-011-0195-2
Barnett S. C., van Dongen W. F. D., Plotz R. D., Weston M. A. (2023). Leash status of approaching dogs mediates escape modality but not flight-initiation distance in a common urban bird. Birds 4, 277–283. doi: 10.3390/birds4030023
Bax V., Francesconi W. (2019). Conservation gaps and priorities in the Tropical Andes biodiversity hotspot: Implications for the expansion of protected areas. J. Environ. Manage. 232, 387–396. doi: 10.1016/j.jenvman.2018.11.086
Beaugeard E., Brischoux F., Angelier F. (2021). Green infrastructures and ecological corridors shape avian biodiversity in a small French city. Urban Ecosyst. 24, 549–560. doi: 10.1007/s11252-020-01062-7
Bellocq M. I., Leveau L. M., Filloy J. (2017). “Urbanization and bird communities: spatial and temporal patterns emerging from Southern South America,” in Ecology and conservation of birds in urban environments. Eds. Murgui E., Hedblom M. (Cham, Switzerland: Springer International Publishing), 35–54. doi: 10.1007/978-3-319-43314-1
Biamonte E., Sandoval L., Chacón E., Barrantes G. (2011). Effect of urbanization on the avifauna in a tropical metropolitan area. Landscape Ecol. 26, 183–194. doi: 10.1007/s10980-010-9564-0
Bibby C., Jones M., Marsden S. (1998). Expedition field techniques: bird surveys (London: Expedition Advisory Centre, Royal Geographical Society).
Blair R. B., Johnson E. M. (2008). Suburban habitats and their role for birds in the urban-rural habitat network: Points of local invasion and extinction? Landscape Ecol. 23, 1157–1169. doi: 10.1007/s10980-008-9267-y
Bonier F., Martin P. R., Wingfield J. C. (2007). Urban birds have broader environmental tolerance. Biol. Lett. 3, 670–673. doi: 10.1098/rsbl.2007.0349
Buchanan G. M., Donald P. F., Butchart S. H. M. (2011). Identifying priority areas for conservation: A global assessment for forest-dependent birds. PloS One. 6, 1–10. doi: 10.1371/journal.pone.0029080
Callaghan C. T., Major R. E., Cornwell W. K., Poore A. G. B., Wilshire J. H., Lyons M. B. (2019). A continental measure of urbanness predicts avian response to local urbanization. Ecography. 42, 1–11. doi: 10.1111/ecog.04863
Campos-Silva L. A., Piratelli A. J. (2021). Vegetation structure drives taxonomic diversity and functional traits of birds in urban private native forest fragments. Urban Ecosyst. 24, 375–390. doi: 10.1007/s11252-020-01045-8
Carvajal-Castro J. D., María Ospina-L A., Toro-Ló pez Y., Pulido-G A., Ximena Cabrera-Casas L., Guerrero-Peláez S., et al. (2019). Birds vs bricks: Patterns of species diversity in response to urbanization in a Neotropical Andean city. PloS One. 14, e0218775. doi: 10.1371/journal.pone.0218775
Castaño-Villa G. J., Patiño-Zabala J. C. (2007). The composition of avian communities in fragmented forest in Santa Elena region, Central Colombian Andes. Boletín Científico Museo Hist. Natural. 11, 47–60.
Chaparro-Herrera S., Lozano M., Echeverry-Galvis M. A. (2024). Listado de las aves Endémicas y Casi-Endémicas de Colombia: Evaluación 2013-2023. Ornitología Colombiana 25, 34–45. doi: 10.59517/oc.e580
Charre G. M., Zavala Hurtado J. A., Néve G., Ponce-Mendoza A., Corcuera P. (2013). Relationship between habitat traits and bird diversity and composition in selected urban green areas of Mexico city. Ornitol. Neotrop. 24, 275–293.
Chin A. T. M., Ruppert J. L. W., Shrestha N., Fortin M.-J. (2022). Urban avian conservation planning using species functional traits and habitat suitability mapping. Land. 11, 1831. doi: 10.3390/land11101831
Chong K. Y., Teo S., Kurukulasuriya B., Chung Y. F., Rajathurai S., Tiang H., et al. (2014). Not all green is as good: Different effects of the natural and cultivated components of urban vegetation on bird and butterfly diversity. Biol. Conserv. 171, 299–309. doi: 10.1016/j.biocon.2014.01.037
Cincotta R. P., Wisnewski J., Engelman R. (2000). Human population in the biodiversity hotspots. Nature. 404, 990–992. doi: 10.1038/35010105
Clergeau P., Savard J. P. L., Mennechez G., Falardeau G. (1998). Bird abundance and diversity along an urban-rural gradient: a comparative study between two cities on different continents. Condor. 100, 413–425. doi: 10.2307/1369707
Concepción E. D., Moretti M., Altermatt F., Nobis M. P., Obrist M. K. (2015). Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale. Oikos. 124, 1571–1582. doi: 10.1111/oik.02166
Conole L. E., Kirkpatrick J. B. (2011). Functional and spatial differentiation of urban bird assemblages at the landscape scale. Landscape Urban Plann. 100, 11–23. doi: 10.1016/j.landurbplan.2010.11.007
de Castro Pena J. C., Martello F., Ribeiro M. C., Armitage R. A., Young R. J., Rodrigues M. (2017). Street trees reduce the negative effects of urbanization on birds. PloS One. 12, 1–19. doi: 10.1371/journal.pone.0174484
del Hoyo J., Elliot A., Sargatal J., Christie D. A. (1992). Handbook of the birds of the world (Barcelona: Lynx Editions).
Dymond C. C., Mladenoff D. J., Radeloff V. C. (2002). Phenological differences in Tasseled Cap indices improve deciduous forest classification. Remote Sens. Environ. 80, 460–472. doi: 10.1016/S0034-4257(01)00324-8
Echeverry-Galvis M. A., Amaya Espinel J. D., Lozano Ramírez P. (2023). Long-term Christmas Bird Counts describe Neotropical urban bird diversity. PloS One 18, E0272754. doi: 10.1371/journal.pone.0272754
Elías Domínguez-López M., Ortega-Álvarez R. (2014). The importance of riparian habitats for avian communities in a highly human-modified Neotropical landscape. Rev. Mexicana Biodiversidad 85, 1217–1227. doi: 10.7550/rmb.43849
Escobar-Ibáñez J. F., Macgregor-Fors I. (2016). Peeking into the past to plan the future: Assessing bird species richness in a neotropical city. Urban Ecosyst. 19, 657–667. doi: 10.1007/s11252-015-0517-x
Estevo C. A., Nagy-Reis M. B., Silva W. R. (2017). Urban parks can maintain minimal resilience for Neotropical bird communities. Urban Forestry Urban Greening 27, 84–89. doi: 10.1016/j.ufug.2017.06.013
Evans K. L., Chamberlain D. E., Hatchwell B. J., Gregory R. D., Gaston K. J. (2011). What makes an urban bird? Global Change Biol. 17, 32–44. doi: 10.1111/j.1365-2486.2010.02247.x
Fay M. P. (2010). Confidence intervals that match Fisher’s exact or Blaker’s exact tests. Biostatistics 11, 373–374. doi: 10.1093/biostatistics/kxp050
Ferenc M., Sedláček O., Fuchs R., Dinetti M., Fraissinet M., Storch D. (2014). Are cities different? Patterns of species richness and beta diversity of urban bird communities and regional species assemblages in Europe. Global Ecol. Biogeog. 23, 479–489. doi: 10.1111/geb.12130
Fischer J. D., Schneider S. C., Ahlers A. A., Miller J. R. (2015). Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv. Biol. 29, 1246–1248. doi: 10.1111/cobi.12451
Garizábal-Carmona J. A., Betancur J. S., Montoya-Arango S., Franco-Espinosa L., Ruíz-Giraldo N., Mancera-Rodríguez N. J. (2023). Bird diversity across an andean city: the limitation of species richness values and watershed scales. Acta Biologica Colombiana 28, 506–516.. doi: 10.15446/abc.v28n3.101974
Garizábal-Carmona J. A., Mancera-Rodríguez N. J. (2021). Bird species richness across a Northern Andean city: Effects of size, shape, land cover, and vegetation of urban green spaces. Urban Forestry Urban Greening 64, 127243. doi: 10.1016/j.ufug.2021.127243
Goddard M. A., Dougill A. J., Benton T. G. (2010). Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol. Evol. 25, 90–98. doi: 10.1016/j.tree.2009.07.016
González-Lagos C., Quesada J. (2017). “Stay or leave? Avian behavioral responses to urbanization in Latin America,” in Avian ecology in latin american cityscapes. Eds. MacGregor-Fors I., Escobar-Ibáñez J. F. (Springer, Cham, Switzerland), 99–123. doi: 10.1007/978-3-319-63475-3
Hilty S. L., Brown W. L. (1986). A guide to the birds of Colombia (Princeton, New Jersey, USA: Princeton University Press).
Hsieh T. C., Ma K. H., Chao A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 7, 1451–1456. doi: 10.1111/2041-210X.12613
Jokimäki J., Kaisanlahti-Jokimäki M. L. (2003). Spatial similarity of urban bird communities: a multiscale approach. J. Biogeog. 30, 1183–1193. doi: 10.1046/j.1365-2699.2003.00896.x
Kong F., Yin H., Nakagoshi N., Zong Y. (2010). Urban green space network development for biodiversity conservation: Identification based on graph theory and gravity modeling. Landscape Urban Plann. 95, 16–27. doi: 10.1016/j.landurbplan.2009.11.001
Kowarik I. (2011). Novel urban ecosystems, biodiversity, and conservation. Environ. pollut. 159, 1974–1983. doi: 10.1016/j.envpol.2011.02.022
La Sorte F. A., McKinney M. L., Pyšek P. (2007). Compositional similarity among urban floras within and across continents: Biogeographical consequences of human-mediated biotic interchange. Global Change Biol. 13, 913–921. doi: 10.1111/j.1365-2486.2007.01329.x
Lees A. C., Moura N. G. (2017). Taxonomic, phylogenetic and functional diversity of an urban Amazonian avifauna. Urban Ecosyst. 20, 1019–1025. doi: 10.1007/s11252-017-0661-6
Leveau L. M., Leveau C. M., Villegas M., Cursach J. A., Suazo C. G. (2017). Bird communities along urbanization gradients: a comparative analysis among three neotropical cities. Ornitol. Neotrop. 28, 77–87. doi: 10.58843/ornneo.v28i0
MacGregor-Fors I. (2011). Misconceptions or misunderstandings? On the standardization of basic terms and definitions in urban ecology. Landscape Urban Plann. 100, 347–349. doi: 10.1016/j.landurbplan.2011.01.013A
MacGregor-Fors I., Payton M. E. (2013). Contrasting diversity values: statistical inferences based on overlapping confidence intervals. PloS One 8, e56794. doi: 10.1371/journal.pone.0056794
Martin L. J., Blossey B., Ellis E. (2012). Mapping where ecologists work: biases in the global distribution of terrestrial ecological observation. Front. Ecol. Environ. 10, 195–201. doi: 10.1890/110154
Mcdonald R. I., Forman R. T. T., Kareiva P., Neugarten R., Salzer D., Fisher J. (2009). Urban effects, distance, and protected areas in an urbanizing world. Landscape Urban Plann. 93, 63–75. doi: 10.1016/j.landurbplan.2009.06.002
Mcdonald R. I., Kareiva P., Forman R. T. T. (2008). The implications of current and future urbanization for global protected areas and biodiversity conservation. Biol. Conserv. 141, 1695–1703. doi: 10.1016/j.biocon.2008.04.025
McKinney M. L. (2006). Urbanization as a major cause of biotic homogenization. Biol. Conserv. 127, 247–260. doi: 10.1016/j.biocon.2005.09.005
McKinney M. L. (2008). Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 11, 161–176. doi: 10.1007/s11252-007-0045-4
Melles S., Glenn S., Martin K. (2003). Urban bird diversity and landscape complexity: species– environment associations along a multiscale habitat gradient. Conserv. Ecol. 7, 5–27. doi: 10.5751/ES-00478-070105
Miller R. G. (1981). Simultaneous statistical inference (New York, NY: Springer). doi: 10.1007/978-1-4613-8122-8
Miller J. R., Hobbs R. J. (2002). Conservation where people live and work. Conserv. Biol. 16, 330–337. doi: 10.1046/j.1523-1739.2002.00420.x
Mills G. S., Dunning J. B., Bates J. M. (1989). Effects of urbanization on breeding bird community structure in southwestern desert. Condor 91, 416–428. doi: 10.2307/1368320
Møller A. P., Díaz M., Flensted-Jensen E., Grim T., Ibáñez-Álamo J. D., Jokimäki J., et al. (2015). Urbanized birds have superior establishment success in novel environments. Oecologia 178, 943–950. doi: 10.1007/s00442-015-3268-8
Morelli F., Benedetti Y., Ibáñez-Álamo J. D., Jokimäki J., Mänd R., Tryjanowski P., et al. (2016). Evidence of evolutionary homogenization of bird communities in urban environments across Europe. Global Ecol. Biogeog. 25, 1284–1293. doi: 10.1111/geb.12486
Naranjo L. G., Amaya J. D., Eusse-González D. E., Cifuentes-Sarmiento Y. (2012). Guía de las Especies Migratorias de la Biodiversidad en Colombia. Aves (A guide to migratory species of Colombian biodiversity: birds) Vol. 1. Ed. M. de A. y D. S./WWF (Bogotá: Ministerio de Ambiente y Desarrollo Sostenible/WWF).
Neate-Clegg M. H. C., Tonelli B. A., Youngflesh C., Wu J. X., Montgomery G. A., Sekerciuglu C. H., et al. (2023). Traits shaping urban tolerance in birds differ around the world. Curr. Biol. 33, 1677–1688. doi: 10.1016/j.cub.2023.03.024
Ortega-Álvarez R., MacGregor-Fors I. (2009). Living in the big city: Effects of urban land-use on bird community structure, diversity, and composition. Landscape Urban Plann. 90, 189–195. doi: 10.1016/j.landurbplan.2008.11.003
Ortega-Álvarez R., MacGregor-Fors I. (2011). Dusting-off the file: A review of knowledge on urban ornithology in Latin America. Landscape Urban Plann. 101, 1–10. doi: 10.1016/j.landurbplan.2010.12.020
Ouyang J. Q., Isaksson C., Schmidt C., Hutton P., Bonier F., Dominoni D. (2018). A new framework for urban ecology: an integration of proximate and ultimate responses to anthropogenic change. Integr. Comp. Biol. 58, 915–928. doi: 10.1093/icb/icy110
Paniagua-Villada C., Garizábal-Carmona J. A., Martínez-Arias V. M., Mancera-Rodríguez N. J. (2024). Built vs Green cover: an unequal struggle for urban space in Medellín (Colombia). Urban Ecosys. doi: 10.1007/s11252-023-01443-8
Parés-Ramos I., Álvarez-Berríos N., Aide T. (2013). Mapping urbanization dynamics in major cities of Colombia, Ecuador, perú, and Bolivia using night-time satellite imagery. Land 2, 37–59. doi: 10.3390/land2010037
Pauchard A., Aguayo M., Peña E., Urrutia R. (2006). Multiple effects of urbanization on the biodiversity of developing countries: The case of a fast-growing metropolitan area (Concepción, Chile). Biol. Conserv. 127, 272–281. doi: 10.1016/j.biocon.2005.05.015
Puppim de Oliveira J. A., Balaban O., Doll C. N. H., Moreno-Peñaranda R., Gasparatos A., Iossifova D., et al. (2011). Cities and biodiversity: Perspectives and governance challenges for implementing the convention on biological diversity (CBD) at the city level. Biol. Conserv. 144, 1302–1313. doi: 10.1016/j.biocon.2010.12.007
Quintero E., Benavides A. M., Moreno N., González-Caro S. (2017). Bosques Andinos: Estado actual y retos para su conservación en Antioquia (Andean forests: conservation status and challenges in Antioquia). Eds. Vallejo E.Q., Benavides A. M., Moreno N., Gonzalez-Caro S. (Medellín: Fundación Jardín Botánico de Medellín Joaquín Antonio Uribe- Programa Bosques Andinos (COSUDE).
Rahbek C., Borregaard M. K., Colwell R. K., Dalsgaard B., Holt B. G., Morueta-Holme N., et al. (2019). Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science 365, 1108–1113. doi: 10.1126/science.aax0149
R Core Team (2019). A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available at: http://www.R-project.org/.
Rega-Brodsky C. C., Aronson M. F. J., Piana M. R., Carpenter E.-S., Hahs A. K., Herrera-Montes A., et al. (2022). Urban biodiversity: State of the science and future directions. Urban Ecosyst. 25, 1083–1096. doi: 10.1007/s11252-022-01207-w
Renjifo L. M., Amaya-Villarreal A. M., Butchart S. H. M. (2020). Tracking extinction risk trends and patterns in a mega-diverse country: A Red List Index for birds in Colombia. PloS One 15, 1–19. doi: 10.1371/journal.pone.0227381
Renjifo L. M., Gómez M. F., Velásquez-Tibatá J., Amaya-Villarreal Á.M., Kattan G. H., Amaya-Espinel J. D., et al. (2014). “Libro rojo de aves de Colombia,” in Bosques húmedos de los Andes y la costa pacífica (Red list of Colombian birds: humid forest and pacific coast), vol. I. (Editorial Pontificia Universidad Javeriana, Primera ed. Bogotá, D.C).
Ruggiero A. (2001). Size and shape of the geographical ranges of Andean passerine birds: spatial patterns in environmental resistance and anisotropy. J. Biogeog. 28, 1281–1294. doi: 10.1046/j.1365-2699.2001.00632.x
Samarawickrama U., Piyaratne D., Ranagalage M. (2017). Relationship between NDVI with Tasseled cap Indices : A Remote Sensing based Analysis. Int. J. Innovative Res. Technol. 3, 13–19.
Sanz V., Caula S. (2015). Assessing bird assemblages along an urban gradient in a Caribbean island (Margarita, Venezuela). Urban Ecosyst. 18, 729–746. doi: 10.1007/s11252-014-0426-4
Schnitter P., Giraldo M. L., Patiño J. M. (2006). La ocupación del territorio en el proceso de urbanización del área metropolitana del valle de Aburrá, Colombia (Urbanization process in the Metropolitan Area of Aburrá Valley, Colombia). Rev. electrónica geografía y Cienc. Social. 10, 83–90.
Seto K. C., Fragkias M., Gü Neralp B., Reilly M. K. (2011). A meta-analysis of global urban land expansion. PloS One 6 (8), e23777. doi: 10.1371/journal.pone.0023777
Shochat E., Warren P. S., Faeth S. H., McIntyre N. E., Hope D. (2006). From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 186–191. doi: 10.1016/j.tree.2005.11.019
Shwartz A., Shirley S., Kark S. (2008). How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landscape Urban Plann. 84, 219–229. doi: 10.1016/j.landurbplan.2007.08.003
Sidemo-Holm W., Ekroos J., Reina García S., Söderström B., Hedblom M. (2022). Urbanization causes biotic homogenization of woodland bird communities at multiple spatial scales. Global Change Biol. 28, 6152–6164. doi: 10.1111/gcb.16350
Sol D., González-Lagos C., Moreira D., Maspons J., Lapiedra O. (2014). Urbanisation tolerance and the loss of avian diversity. Ecol. Lett. 17, 942–950. doi: 10.1111/ele.12297
Stiles F. G. (1990). La avifauna de la Universidad de Costa Rica y sus alrededores a través de veinte año (1968-1989) (Birds to the University of Costa Rica and surroundings during 20 year (1968-1989). Rev. Biol. Trop. 38, 361–381.
Sutherland W. J. (2006). Ecologial census techniques: a handbook (Cambridge, UK: Cambridge University Press).
Threlfall C. G., Mata L., Mackie J. A., Hahs A. K., Stork N. E., Williams N. S. G., et al. (2017). Increasing biodiversity in urban green spaces through simple vegetation interventions. J. Appl. Ecol. 54, 1874–1883. doi: 10.1111/1365-2664.12876
Tobias J. A., Sheard C., Pigot A. L., Devenish A. J. M., Yang J., Neate-Clegg M. H. C., et al. (2022). AVONET: morphological, ecological and geographical data for all birds. Ecol. Lett. 25, 581–597. doi: 10.1111/ele.13898
Tryjanowski P., Morelli F., Mikula P., Krištín A., Indykiewicz P., Grzywaczewski G., et al. (2017). Bird diversity in urban green space: A large-scale analysis of differences between parks and cemeteries in Central Europe. Urban Forestry Urban Greening 27, 264–271. doi: 10.1016/j.ufug.2017.08.014
Villaseñor N. R., Muñoz-Pacheco C. B., Escobar M. A. H. (2024). Opposite responses of native and nonnative birds to socioeconomics in a latin american city. Animals 14, 299. doi: 10.3390/ani14020299
Wilman H., Belmaker J., Simpson J., de la Rosa C., Rivadeneira M. M., Jetz W. (2014). Elton Traits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 95, 1–14. doi: 10.1890/13-1917.1
Keywords: biodiversity, bird assemblage, tropical Andes, urbanization, wildlife categorization
Citation: Garizábal-Carmona JA, Betancur JS, Montoya-Arango S, Franco-Espinosa L and Mancera-Rodríguez NJ (2024) Categorizing urban avoiders, utilizers, and dwellers for identifying bird conservation priorities in a Northern Andean city. Front. Ecol. Evol. 12:1432340. doi: 10.3389/fevo.2024.1432340
Received: 13 May 2024; Accepted: 10 July 2024;
Published: 13 August 2024.
Edited by:
Laura Pereyra, CONICET Instituto de Ecorregiones Andinas (INECOA), ArgentinaReviewed by:
Alejandro Schaaf, CONICET Instituto de Ecorregiones Andinas (INECOA), ArgentinaRodrigo Lorenzon, CONICET Santa Fe, Argentina
Copyright © 2024 Garizábal-Carmona, Betancur, Montoya-Arango, Franco-Espinosa and Mancera-Rodríguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: N. Javier Mancera-Rodríguez, bmptYW5jZXJAdW5hbC5lZHUuY28=