 Nicolas Lavesque
Nicolas Lavesque João M. M. Nogueira2†
João M. M. Nogueira2†- 1CNRS, Univ. Bordeaux, EPOC, UMR 5805, Station Marine d’Arcachon, Arcachon, France
- 2Laboratório de Poliquetologia, Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil
- 3Univ. Bordeaux, CNRS, INRAE, Univ. La Rochelle UMS 2567 POREA, Pessac, France
- 4Australian Museum Research Institute, Australian Museum, Sydney, NSW, Australia
- 5Department of Biological Sciences, Macquarie University, Sydney, NSW, Australia
Five new species of Terebellides are described from coastal and deep waters of Papua New Guinea, using both morphology and molecular tools (for four species). Terebellides elenae n. sp. is characterized by the absence of both a glandular lateral region on TC3 and papillae on margins of the branchial lamellae and by the presence of partially fused branchial lobes with conspicuous fifth lobe and dorsal rounded projections until TC6. Terebellides fauchaldi n. sp. has a very large glandular lateral region on the third thoracic chaetiger (TC3), a fifth branchial lobe and partially fused branchial lobes, and conspicuous dorsal rounded projections on TC2–6. Terebellides madeep n. sp. is characterized by a thin glandular region on TC3 and by four free branchial lobes. Terebellides oculata n. sp. is one of the only two species in the world to have eyespots. Finally, T. papillosa n. sp. has geniculate chaetae on TC6 and TC7 and bears a large number of papillae. A majority-rule consensus tree using the 16S gene and an identification key for all Terebellides species described from the Central Indo-Pacific region are provided.
Introduction
Trichobranchids are tubicolous, deposit-feeding polychaetes found from shallow to shelf depths (Hutchings and Peart, 2000). Within the family Trichobranchidae Malmgren, 1866, the genus Terebellides Sars, 1835 is very diverse with 86 currently valid species (Read and Fauchald, 2023). For a long time, the type species T. stroemii Sars, 1835 was reported worldwide and considered cosmopolitan (Hutchings and Kupriyanova, 2018; Hutchings and Lavesque, 2020). The redescription and designation of a neotype by Parapar and Hutchings (2014) and the recent use of molecular tools (Nygren et al., 2018) led to the description of approximately 30 new species, especially from Europe (Parapar et al., 2016a; Lavesque et al., 2019; Parapar et al., 2020a; Barroso et al., 2022), Africa (Parapar et al., 2020b), and the Indo-Pacific region (Parapar et al., 2016b, 2016c; Hsueh and Li, 2017; Zhang and Hutchings, 2018).
However, we suggest that the diversity within this genus is still underestimated. In the Central Indo-Pacific region, sensu Spalding et al. (2007), 16 species have been described: Terebellides akares Hutchings, Nogueira and Carrerette, 2015; T. baliensis Hsueh and Li, 2017; T. ectopium Zhang and Hutchings, 2018; T. ehlersi McIntosh, 1885; T. guangdongensis Zhang and Hutchings, 2018; T. hutchingsae Parapar, Moreira and Martin, 2016; T. intoshi Caullery, 1915; T. jitu Schüller and Hutchings, 2010; Terebellides jorgeni Hutchings, 2007; T. kowinka Hutchings and Peart, 2000; T. narribri Hutchings and Peart, 2000; T. woolawa Hutchings and Peart, 2000; and T. yangi Zhang and Hutchings, 2018. Two other species were described from the region, but T. sieboldi Kinberg, 1867 is considered a nomen dubium (Schüller and Hutchings, 2013) and T. ypsilon Grube, 1878 is considered indeterminable, as the type material only contains a few chaetae and some epidermis, the original description is brief, and the figures are somewhat schematic (Hutchings and Peart, 2000).
Since 2010, four sampling campaigns have been undertaken in Papua New Guinea to explore the coastal and deep-sea biodiversity of this region, considered a marine biodiversity hotspot: BIOPAPUA 2010, PAPUA NIUGINI 2012, MADEEP 2012, and KAVIENG 2014 (Pante et al., 2012; Corbari et al., 2019). These cruises were conducted by the Muséum National d’Histoire Naturelle (MNHN) and the Institut de Recherche pour le Développement (IRD), in collaboration with the University of Papua New Guinea (UPNG). Thousands of polychaetes belonging to many families were collected and stored in the MNHN collection and are available for examination and description, including new species. For example, these samples allowed us to recently describe three deep-sea species of Marphysa Quatrefages, 1866 (Lavesque et al., 2022). During a workshop organized by MNHN in 2021 to sort these samples, numerous spaghetti worms (i.e., terebellids and trichobranchids) were found, and molecular analysis confirmed the presence of a large number of undescribed species which will be described in a series of papers including this one. In this study, we describe five new species of Terebellides from coastal and deep waters, using both morphological and molecular tools. The use of molecular data (such as the 16S gene) is essential as many cryptic species of Terebellides have very restricted distributions (Nygren et al., 2018; Lavesque et al., 2021). Finally, an identification key for the species of Terebellides from the Central Indo-Pacific region is provided.
Material and methods
Sampling and morphological analyses
Specimens were collected in Papua New Guinea during the PAPUA NIUGINI cruise in 2012 (https://expeditions.mnhn.fr/campaign/papuaniugini) and the MADEEP cruise in 2014 (https://expeditions.mnhn.fr/campaign/madeep) (Figure 1). All materials were sampled by a dredge or a beam trawl, sorted on board RV Alis and fixed in 80% ethanol. A few parapodia were removed from several specimens for molecular analysis. Specimens were examined under a Nikon SMZ25 stereomicroscope and a Nikon Eclipse Ci microscope and photographed with a Nikon DS-Ri 2 camera. Measurements were made with the NIS-Elements Analysis software.
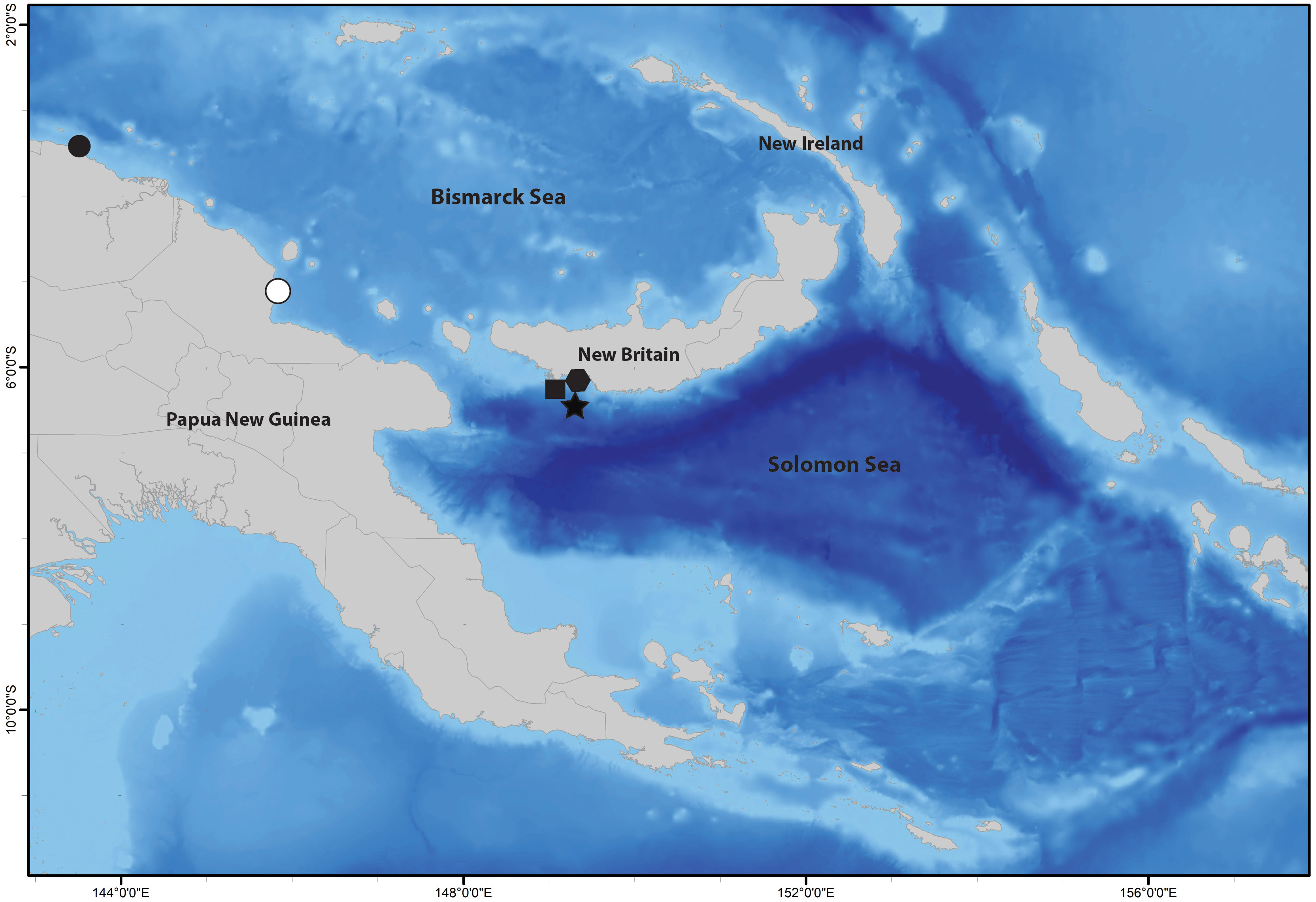
Figure 1 Type localities of Terebellides species described in this study, in Papua New Guinea: Terebellides elenae n. sp. (black circle), Terebellides oculata n. sp. (white circle), Terebellides papillosa n. sp. (black square), Terebellides fauchaldi n. sp. (black hexagon), and Terebellides madeep n. sp. (black star).
Concerning the morphological classifications, the different types of branchiae follow Parapar et al. (2016a); the types of thoracic uncini follow Parapar et al. (2020b), and finally, the different types of abdominal uncini follow Parapar et al. (2020a). Concerning thoracic uncini, an additional fifth type has been identified during this study: “type 5,” with RvC = 1/0.7, with four to five mid-sized teeth above the main fang, surmounted by two rows of short denticles and an upper crest of several small denticles. Methyl green, which can be washed out, was used to observe glandular areas.
Dehydrated specimens used for examination by scanning electron microscopy (SEM) were prepared by critical point drying, coated with gold, and examined and photographed with a Hitachi TM3030.
The studied material is deposited at the Muséum National d’Histoire Naturelle, Paris (MNHN).
Molecular data and analyses
Extraction of DNA was done with Maxwell (Promega, Madison, USA), an automated DNA/RNA isolation, with Maxwell® RSC Blood DNA kit, following the protocol supplied by the manufacturers. Approximately 400 bp of the 16S rDNA gene was amplified, using the primers 16Sannf (GCGGTATCCTGACCGTRCWAAGGTA) (Sjölin et al., 2005) and 16SbrH (CCGGTCTGAACTCAGATCACGT) (Palumbi et al., 1991). Approximately 600 bp of the COI (cytochrome c oxidase subunit I) gene was amplified using the primers LCO1490 and HCO2198 (Folmer et al., 1994). Polymerase chain reaction (PCR) was performed with GoTaq® G2 Flexi DNA Polymerase Kit in 20 μl mixtures containing 4 μl of 5X Green GoTaq® Flexi Reaction Buffer (final concentration of 1×), 1.2 μl of MgCl2 (25 mM) solution, 0,4 μl of PCR nucleotide mix (final concentration of 0.2 mM each dNTP), 0.2 μl of each primer (final concentration of 1 μM), 0.1 μl of Taq DNA Polymerase (5 U/μl), 1 μl of template DNA, and 12.9 μl of nuclease-free water. The PCR amplification conditions were 94°C/600 s–(94°C/60 s–59°C/30 s–72°C/90 s) * 40 cycles–72°C/600 s for 16S and 94°C/600 s–(94°C/40 s–44°C/40 s–72°C/60 s) * 5 cycles–(94°C/40 s–51°C/40 s–72°C/60 s) * 35 cycles–72°C/300 s–4°C for COI. PCR success was verified by electrophoresis in a 1% p/v agarose gel stained with ethidium bromide. The amplified products were sent to Eurofins Genomics (Ebersberg, Germany) Company to complete double strain sequencing, using the same set of primers as used for PCR. Overlapping sequence (forward and reverse) fragments were merged into consensus sequences.
Thirty-one 16S sequences were downloaded from GenBank or obtained during this study, with 30 sequences belonging to Terebellides species, while the one remaining belongs to a closely related genus (Trichobranchus) used as an outgroup (Table 1). All 16S sequences were aligned in Geneious Prime 2023.0.1 using the MAAFT (Katoh et al., 2002) plugin and default settings. The maximum likelihood analysis was performed in IQ-TREE 2.2.0 (Trifinopoulos et al., 2016) with the best-fitting evolutionary model GTR+F+G4 selected. Bootstrap support was estimated using an ultrafast bootstrap algorithm (UFBoot) (Minh et al., 2013) for 1,000 replicates. Pairwise Kimura two-parameter (K2P) genetic distance was performed using MEGA version 7.0.26. All sequences obtained in this study were deposited in GenBank.
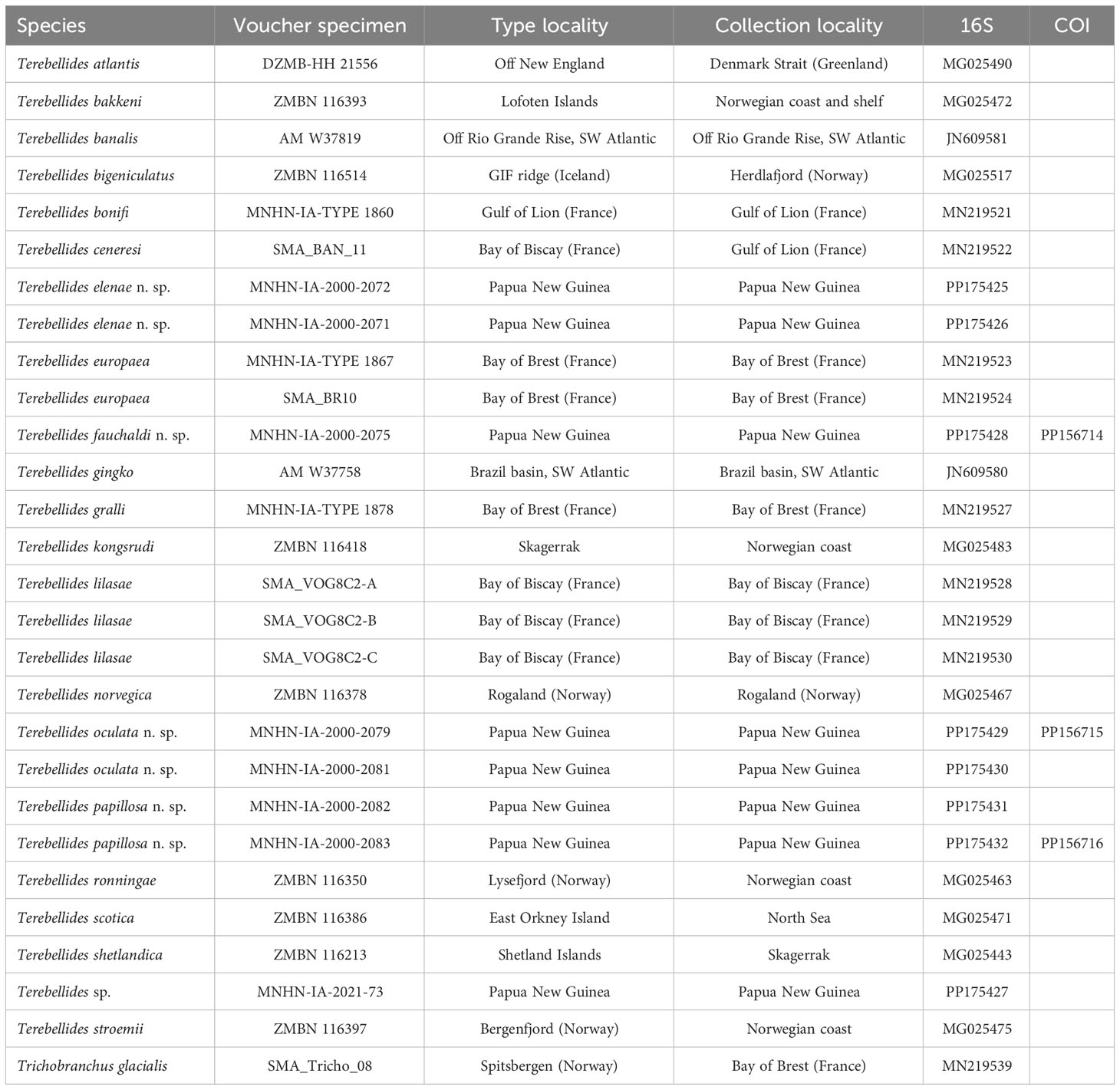
Table 1 Terminal taxa used in the molecular part of the study (16S and COI genes), with voucher specimens, type localities, collection localities, and GenBank accession numbers for both genes.
Results
Taxonomy
Family Trichobranchidae Malmgren, 1866.
Genus Terebellides Sars, 1835.
Type species: Terebellides stroemii Sars, 1835.
Diagnosis (emended from Hutchings et al., 2021) Transverse prostomium attached to the dorsal surface of the upper lip; basal part as a thick crest, eyespots rarely present; distal part extending along the upper lip until near the anterior border of the lip. Buccal tentacles of two types, uniformly cylindrical and expanded at the tips, spatulate. Peristomium forming lips; lips expanded, circular upper lip, distal margin convoluted; expanded lower lip, scoop-shaped, with a large marginal lobe. Segment 1 short, conspicuous all around or only visible ventrally; following anterior segments with lobes as low ventral collars, frequently protruding laterally for short extension on segments 2–4, at least. Branchiae as single, stalked, four to five lobed structure, inserted mid-dorsally on segments 2–3 or 2–4, lobes in two pairs, each with multiple lamellae with ciliary tracts and rows of ciliated papillae; a fifth anterior lobe often present. Notopodia beginning on segments 3 or 4, usually 3, extending for 17–18 segments, until segment 20; narrowly winged notochaetae in both rows throughout. Neuropodia beginning from segments 7 or 8, usually 8; sessile thoracic neuropodia, uncini emerging directly from the body wall; first pair of neuropodia or first two pairs, when beginning from segment 7, with subdistally bent, distally tapered spines, from segment 9 thoracic neurochaetae as avicular uncini, hood below the main fang absent, crest with relatively few transverse rows of secondary teeth; abdominal neuropodia as foliaceous pinnules, bearing avicular uncini, with rows of secondary teeth on top and lateral to the main fang. Nephridial papillae only on segment 3, genital papillae, if present, normally on segments 6–7, at bases of notopodia, posterior and dorsal. Pygidium smooth to slightly crenulate.
Terebellides elenae n. sp.
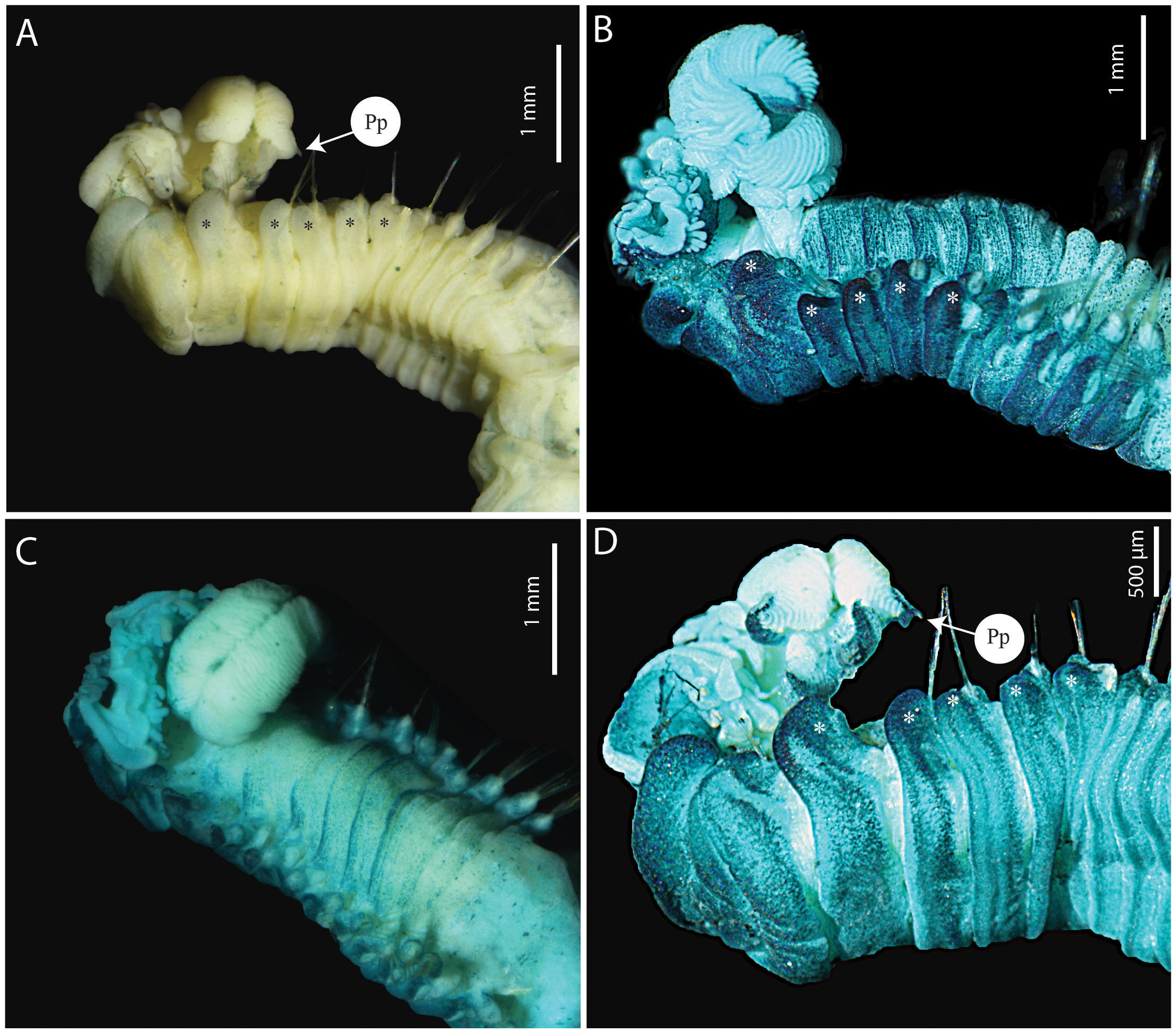
Figure 2 Terebellides elenae n. sp. (A, C–D) holotype MNHN-IA-2000-2071 (B) paratype MNHN-IA-2000-2073. (A) Anterior part, ventro-lateral view. (B) Anterior part, lateral view, MG staining. (C) Anterior part, dorsal view, MG staining. (D) Anterior part, ventro-lateral view, MG staining. Pp, posterior projection. Stars show dorsal rounded projections.
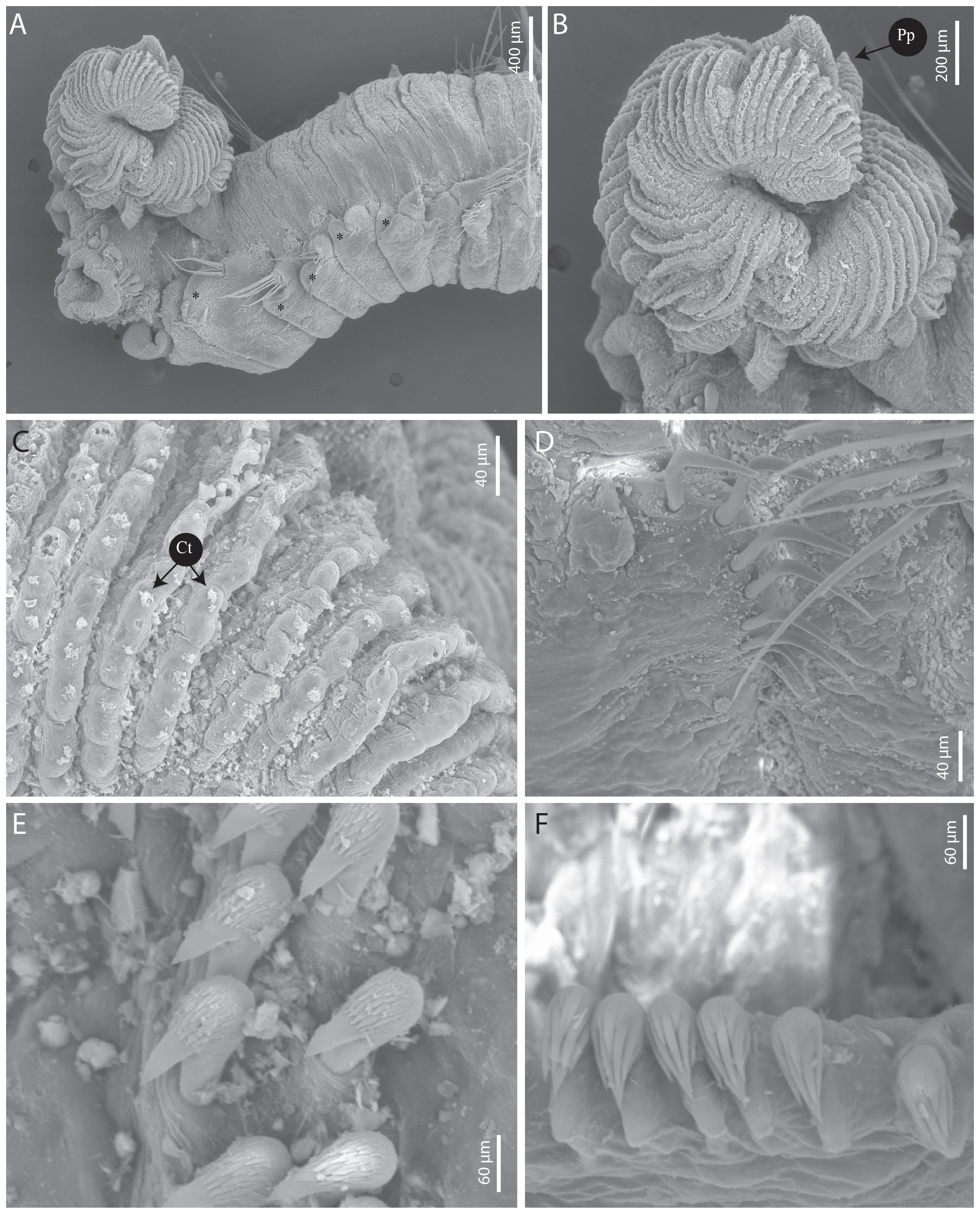
Figure 3 Terebellides elenae n. sp. (A–E) paratype SEM MNHN-IA-2000-2073. (A) Anterior part, dorso-lateral view. (B) Branchial lobes, dorsal view. (C) Branchial lamellae, dorsal view. (D) Geniculate chaetae (TC6). (E) Uncini, thoracic chaetigers. (F) Uncini, abdominal chaetigers. Ct, ciliary tufts; Pp, posterior projection. Stars show dorsal rounded projections.
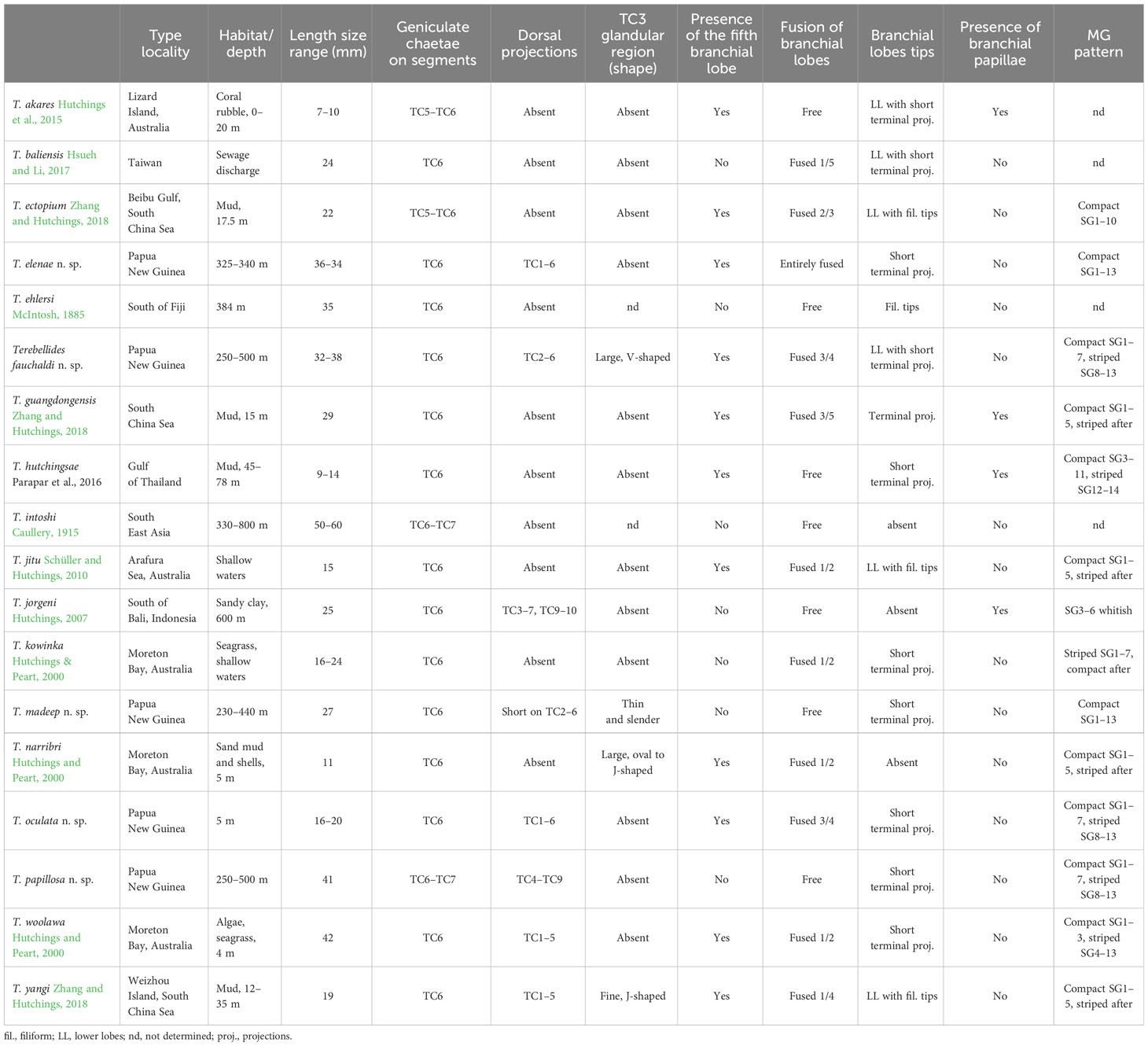
Table 2 Distinguishing characters of species of Terebellides (Polychaeta: Trichobranchidae).
Zoobank registration LSID: lsid:zoobank.org:act:F3C2E3DE-9D7E-4893-AB24-3A7C1AC9D16E.
Type material.
Holotype.
MNHN-IA-2000-2071, complete, some parapodia used for molecular analysis, South Pacific Ocean, Papua New Guinea, Kairiru Island, PAPUA NIUGINI, DW4047, 3.35°S, 143.46°E, depth 325–340 m, coll. December 2012.
Paratypes.
All specimens from the same collection site as the holotype. MNHN-IA-2000-2072, anterior part only, some parapodia used for molecular analysis; MNHN-IA-2000-2073, complete, in three parts, mounted for SEM.
Description (based on holotype, with variation in parentheses for paratypes).
Mid-size species, holotype 26.8 mm long (34 mm) and 1.8 mm wide (1.7 mm). Body tapering posteriorly, segments increasingly shorter and more compacted toward the pygidium. Preserved specimens whitish.
Prostomium compact; eyespots absent; a large upper lip surrounding the mouth; buccal tentacles of two types, uniformly cylindrical and with expanded tips, spatulate, most of them lost (Figures 2A–D, 3A). Lower lip expanded below the upper lip (Figures 2A, B). SGs1 and 2 only visible ventrally; following segments with lobes as ventral collars, lateral lappets on SG5–9 (TC3–7) continuing ventrally; conspicuous dorsal rounded projections on SG3–8 (TC1–6), more developed on SG3–4 (TC1–2); white lateral areas on SG3–4 (TC1–2) present, lacking glandular lateral region on SG5 (TC3) (Figures 2A, B, D).
Branchiae arising as a single structure from SG3, reaching SG8, as a single elongate and annulated mid-dorsal stalk, with two pairs of almost entirely fused lobes on top and anterior branchial projection (fifth lobe) (Figures 2A, C, D). Dorsal lobes with approximately 50 packed lamellae lacking papillar projections on margins, but with ciliated tufts (type 2); dorsal and ventral lobes terminating with short pointed papillae (Figures 2A–D, 3A–C).
Eighteen pairs of thoracic notopodia (SG3–20), first two pairs well developed, notochaetae from TC1 (SG3) about the same size as those from subsequent notopodia (Figures 2A–C, 3A). All notochaetae simple capillaries, arranged in two rows, anterior row shorter. Neuropodia as sessile pinnules from TC6 (SG8) to pygidium. First thoracic pair of neuropodia (TC6) with six sharply bent geniculate chaetae, with acute tips (Figure 3D), subsequent thoracic neuropodia with approximately 30–45 uncini per torus arranged in two regular rows, uncini type 5, RvC = 1/0.7, four to five mid-sized teeth above the main fang surmounted by two rows of short denticles and upper crest of several minute denticles (Figure 3E); uncini arranged in single rows from SG9 (TC7); approximately 30–35 pairs of abdominal neuropodia, as erect paddle-shaped pinnules, each with approximately 35 uncini present at the margin; type 2 uncini, RvC = 1/0.9, four to five teeth above the main fang, surmounted by rows of four to five short teeth and two rows of shorter denticles (Figure 3E).
Two pairs of short nephridial papillae posterior to the base of notopodia of SG6–7 (TC4–TC5). Pygidium rounded, with thick margins.
Methyl green staining pattern: the first 11–13 segments stain solid (Figure 4); ventral faces of branchial lobes stain dark blue; posterior neuropodia with ventral glandular region (Figures 2B–D).
Etymology.
This species is dedicated to our friend Dr. Elena Kupriyanova, senior taxonomist at the Australian Museum, for her contribution to the taxonomy of polychaetes, especially the Serpulidae.
Habitat.
Between 325 and 340 m deep, mud.
Type locality.
South Pacific Ocean, Papua New Guinea, West of Kairiru Island.
Remarks.
In the Central Indo-Pacific region, considering the species which lack both a glandular lateral region on TC3 (SG5) and papillae on margins of branchial lamellae, and also share the presence of partially fused branchial lobes with conspicuous fifth lobe, specimens of T. elenae n. sp. are similar to those of T. woolawa and T. jitu. However, individuals of T. elenae n. sp. differ from both these species by having a well-developed first thoracic chaetiger, which is reduced in T. woolawa and T. jitu. Terebellides elenae n. sp. also has almost entirely fused branchial lobes, whereas the lobes are fused over 50% of their length in the other two species. Furthermore, T. elenae n. sp. has dorsal rounded projections until SG8 (TC6), which are absent in T. jitu. Finally, Terebellides elenae n. sp. is a deep-sea species (350 m deep), while individuals of both T. woolawa and T. jitu have shallow to moderate depth distributions (0–40 m depth for T. woolawa, 150 m for T. jitu).
Terebellides fauchaldi n. sp.
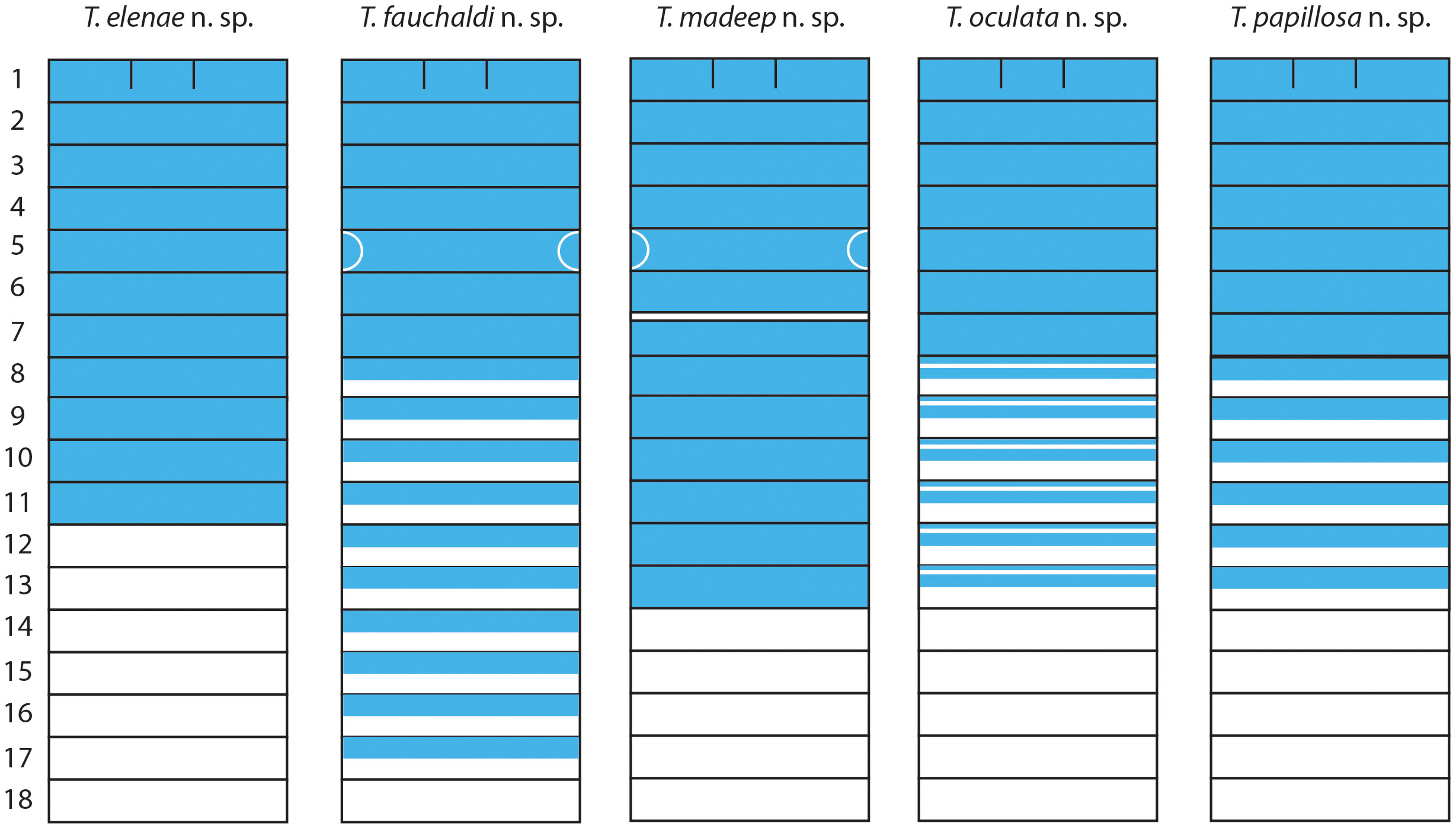
Figure 4 Methyl green staining patterns in ventral view of new Terebellides species described in this study. Numbers refer to segments.
Zoobank registration LSID: lsid:zoobank.org:act:CE32F17D-6E76-4F4D-BDCC-F3915B34A0BE.
Type material.
Holotype.
MNHN-IA-2000-2074, complete, South Pacific Ocean, Papua New Guinea, West New Britain, MADEEP CP4329, 6.133°S, 149.16°E, depth 250–500 m, coll. May 2014.
Paratypes.
MNHN-IA-2000-2075, complete, some parapodia used for molecular analysis, posterior part mounted for SEM, South Pacific Ocean, Papua New Guinea, West New Britain, MADEEP CP4331, 6.116°S, 149.2°E, depth 260 m, coll. May 2014. Paratype SEM MNHN-IA-2000-2076, anterior part only, mounted for SEM, South Pacific Ocean, Papua New Guinea, West New Britain, MADEEP CP4333, 6.116°S, 149.16°E, depth 220–440 m, coll. May 2014. MNHN-IA-2000-2077, complete, in two parts, South Pacific Ocean, Papua New Guinea, West New Britain, MADEEP CP4329, 6.133°S, 149.16°E, depth 250–500 m, coll. May 2014.
Description (based on holotype, with variation in parentheses for paratypes).
Relatively large species, holotype 38.2 mm long (32.3–38.2 mm) and 3.2 mm wide (2.6–4.3 mm). Body tapering posteriorly with segments becoming increasingly shorter and more compacted toward the pygidium.
Prostomium compact; eyespots absent. Upper lip large, surrounding mouth with many buccal tentacles of two types, uniformly cylindrical and with expanded tips, spatulate; lower lip forming an expanded structure below the upper lip (Figures 5A–D, 6A, C). SGs1 and 2 short, SG1 only visible ventrally, SG2 visible all around (Figures 5A–D, 6A, C). Lateral lappets on SG3–7 (TC1–5), largest on TC3, forming a ventral collar, decreasing in size posteriorly (Figures 5A–D, 6A, C); conspicuous dorsal rounded projections on SG4–8 (TC2–6), largest on SG5 (TC3) and partially recovering notopodia of SG4 (TC2) (Figures 5A–D, 6A, C). Presence of a large V-shaped glandular lateral region on SG5 (TC3) (Figures 5A, B, D).

Figure 5 Terebellides fauchaldi n. sp. (A–D) paratype MNHN-IA-2000-2076. (A) Anterior part, antero-ventral view. (B) Anterior part, dorsal view. (C) Anterior part, ventral view. (D) Anterior part, antero-ventral view, MG staining. Drp, dorsal rounded projections; Glr, glandular lateral region.
Branchiae arising as a single structure from SG3 (TC1), reaching SG7 (TC5), consisting of a single elongate and annulated stalk placed mid-dorsally, two pairs of lobes, fused for approximately 3/4 of length, lower pair thinner; anterior branchial projection (fifth lobe) present (Figures 5A, B, 6A, B). Upper lobes with approximately 75 tightly packed lamellae (Figures 5A, B, 6A, B); papillar projections or ciliary tufts on margins of branchial lamellae both absent (type 1) (Figures 6A, B). Distal region of the upper lobes without projections, lower lobes with short terminal pointed tips.
Eighteen pairs of thoracic notopodia (SG3–20). First pair shorter than the subsequent ones; notochaetae from SG3 (TC1) much shorter than the ones from the subsequent notopodia, and transversally aligned (Figures 5A, B, 7A, C). All notochaetae simple capillaries, arranged in two rows, anterior row shorter. Neuropodia present as sessile pinnules from TC6 (SG8) to the last segment; uncini arranged in single rows from SG9 (TC7). First thoracic pair of neuropodia (SG8) provided with four (five) bent acute tipped, geniculate chaetae (Figure 6D; one chaeta bifid, but probably due to abnormality); all subsequent thoracic neuropodia with approximately 10–15 uncini per torus arranged in irregular rows, type 1 uncini with RvC = 4/1, with three to four teeth above the main fang, surmounted by a row of short denticles and upper crest of several minute denticles (Figure 6E). Approximately 32 abdominal neuropodia (24) as erect paddle-shaped pinnules, with approximately 30 uncini along the anterior margin, uncini of type 1, with RvC = 1/0.7, with three to five pointed teeth above the main fang, surmounted by a row of short pointed teeth and upper crest of minute teeth (Figure 6F).
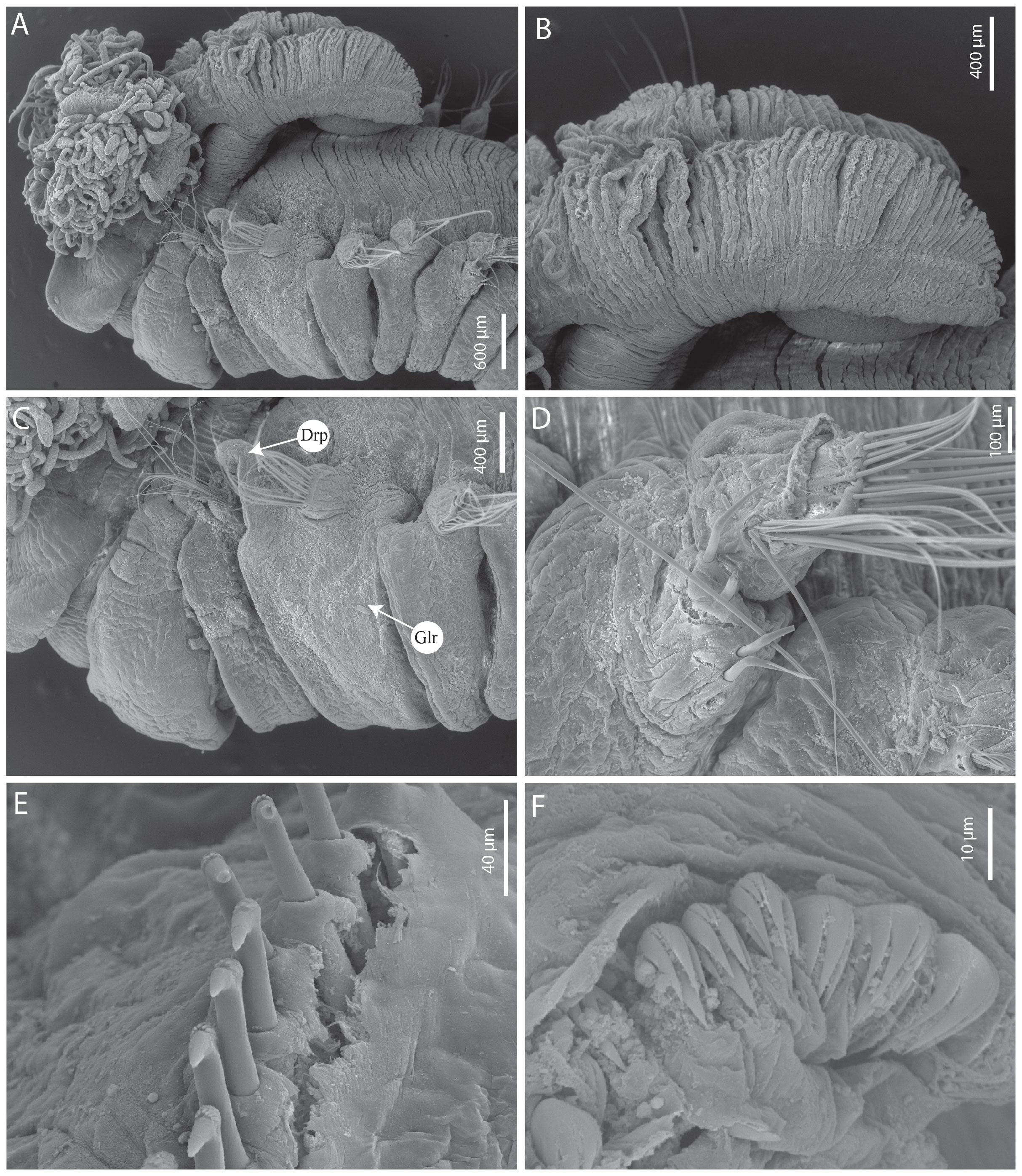
Figure 6 Terebellides fauchaldi n. sp. (A–E) paratype SEM MNHN-IA-2000-2076 (F) paratype MNHN-IA-2000-2075. (A) Anterior part, lateral view. (B) Branchial lobes, lateral view. (C) Anterior part, lateral view. (D) Geniculate chaetae, TC6. (E) Uncini, thoracic chaetiger. (F) Uncini, abdominal chaetiger. Drp, dorsal rounded projection; Glr, glandular lateral region.
Three pairs of short nephridial papillae, located latero-posteriorly to the base of each notopodium of SG4–6 (TC2–TC4). Pygidium crenulated (rounded), as funnel-like depression.
Methyl green staining pattern: the first seven segments stain solid; striped from SG8 (Figure 4); a large V-shaped glandular lateral region on SG5 (TC3) remains white (Figure 5D).
Etymology.
This species is dedicated to Kristian Fauchald, friend of PH and JMMN, for his great contribution to polychaetes taxonomy and for inspiring many taxonomists such as NL.
Habitat.
Between 250 and 500 m depth, mud with a lot of plant debris and sunken wood.
Type locality.
South Pacific, Papua New Guinea, New Britain, Ainto Bay.
Remarks.
Considering the species present in the Central Indo-Pacific region, Terebellides fauchaldi n. sp. is similar to T. madeep n. sp., T. narribri, and T. yangi regarding the presence of a glandular lateral region on TC3. However, T. fauchaldi n. sp. differs from T. madeep n. sp. by the presence of a fifth branchial lobe and partially fused branchial lobes, which are completely free from each other in the latter species. Both species differ from each other in the shape of the glandular region, which is very large in T. fauchaldi n. sp. and thin in T. madeep n. sp. Finally, T. fauchaldi n. sp. has conspicuous dorsal rounded projections on SG4–8 (TC2–6), extremely large on SG5 (TC3), while T. madeep n. sp. only has short dorsal rounded projections on TC2–6.
Terebellides fauchaldi n. sp. differs from T. narribri by the presence of short terminal tips on branchial lower lobes and conspicuous rounded dorsal projections on thoracic chaetigers, whereas such branchial tips and dorsal projections are both absent in T. narribri.
Finally, T. fauchaldi n. sp. differs from T. yangi mainly by the presence of short terminal branchial tips on lower lobes instead of filamentous tips as in T. yangi. Both species can also be differentiated by the shape of the glandular lateral region on SG5 (TC3), which is thin and J-shaped in T. yangi and very large and V-shaped in T. fauchaldi n. sp.
Terebellides madeep n. sp.
Zoobank registration LSID: lsid:zoobank.org:act:7E8EA1A5-0089-4DB1-9A2F-84E066C8824B.
Type material.
Holotype.
MNHN-IA-2000-2078, complete, two parapodia (TC15 and TC29) mounted for SEM, South Pacific Ocean, Papua New Guinea, New Britain, Ainto Bay, MADEEP DW4328, 6.116°S, 149.16°E, depth 230–440 m, coll. May 2014.
Description.
Mid-size species, holotype 27.1 mm long and 2.7 mm wide. Body tapering posteriorly, segments increasingly shorter and more compacted toward the pygidium. Preserved specimen whitish.
Prostomium compact; eyespots absent; a large upper lip surrounding the mouth, buccal tentacles uniformly cylindrical and with expanded tips, spatulate, mostly lost (Figures 7A–C). Lower lip forming an expanded structure below the upper lip (Figure 7A). SGs1 and 2 visible all around; following segments with lobes as ventral collars; lateral lappets on SG3–8 (TC1–6), continuing ventrally in SG4–15 (TC2–13); short dorsal rounded projection on SG4–8 (TC2–6); thin and slender glandular lateral region on SG5 (TC3) (Figures 7A–C).
Branchiae arising as a single structure from SG2, reaching SG7 (TC5), consisting of a single elongate and annulated stalk placed mid-dorsally, with two pairs of lobes completely free from each other from the base, upper lobes with approximately 45 packed lamellae without papillar projections with ciliary tufts on the margins (type 1) (Figures 7A–C), all lobes terminating by short digitiform papillae; anterior branchial projection (fifth lobe) absent (Figures 8A–C).

Figure 7 Terebellides madeep n. sp. (A–E) holotype MNHN-IA-2000-2078. (A) Anterior part, lateral view, MG staining. (B) Anterior part, dorso-lateral view. (C) Anterior part, dorsal view. (D) Anterior part, ventral view, MG staining. (E) Uncini, thoracic chaetiger. (F) Uncini, abdominal chaetiger. Glr, glandular lateral region; Pp, posterior projection; Wb, white band.
Eighteen pairs of thoracic notopodia (SG3–20). Notochaetae from SG3 (TC1) about the same size as those from subsequent notopodia, notopodia all longitudinally aligned. Notochaetae simple capillaries, arranged in two rows, anterior row shorter. Neuropodia present as sessile pinnules from TC6 (SG8) to the last segment; uncini arranged in single rows from SG9 (TC7); first thoracic pair of neuropodia (SG8) with five short geniculate chaetae contracted into the body wall and difficult to see; subsequent thoracic neuropodia with approximately 15–20 uncini per torus arranged in two irregular rows, type 1 uncini with RvC = 2/1, three to four teeth above the main fang, followed by a row of short denticles and upper crest with several minute denticles (Figure 7E). Approximately 35 abdominal neuropodia (24) as erect paddle-shaped pinnules, with approximately 35 uncini along the anterior margin, type 1A uncini with RvC = 1/0.7, three to four teeth above the main fang, followed by three to five short teeth on the first row, two denticles on the second row and upper crest with several minute denticles (Figure 7F).
Pygidium rounded, as a funnel-like depression, with slightly crenulated margins.
Methyl green staining pattern: the first 13 segments stain solid, except SG7 (TC5) with a thin white antero-ventral band (Wb); ventral faces of the lobes stain dark blue; glandular lateral region on SG5 (TC3) white (Figures 7A, B, 4).
Etymology.
This species is named after the MADEEP scientific expedition, during which the type material was collected.
Habitat.
Between 230 and 440 m depth, mud.
Type locality.
South Pacific, Papua New Guinea, New Britain, Ainto Bay.
Remarks.
In the Central Indo-Pacific region, considering the presence of a glandular lateral region on TC3, Terebellides madeep n. sp. is similar to T. fauchaldi n. sp., T. narribri, and T. yangi. However, T. madeep n. sp. differs from those of T. fauchaldi n. sp. by the absence of a fifth branchial lobe, which is present in T. fauchaldi n. sp., and by having branchial lobes completely free from each other, which are partially fused in T. fauchaldi n. sp. Members of both species differ from each other in the shape of the glandular region, which is thin in T. madeep n. sp. and very large in T. fauchaldi n. sp. Finally, T. madeep n. sp. has only short dorsal rounded projections on SG4–8 (TC2–6); such projections are larger, especially on SG5 (TC3), for T. fauchaldi n. sp.
Terebellides madeep n. sp. differs from T. narribri in the absence of a fifth branchial lobe, which is present in T. narribri, and in the branchial lobes being completely free from each other, each with short digitiform projections at the tips, whereas the lobes are fused in pairs in T. narribri, lacking projections of any type at the tips. In addition, T. madeep n. sp. has short dorsal rounded projections (lappets) on SG4–8 (TC2–6), which are absent in T. narribri. Finally, the glandular region on SG5 (TC3) is very large in T. narribri and thin in T. madeep n. sp.
Finally, T. madeep n. sp. differs from T. yangi by the absence of a fifth branchial lobe, having all four branchial lobes totally free from each other, while in T. yangi, a fifth branchial lobe is conspicuous and lobes are fused in dorsal and ventral pairs. Also, the dorsal and ventral branchial lobes of T. madeep n. sp. both terminate by short digitiform projections, while in T. yangi, only the lower lobes have filamentous tips.
Terebellides oculata n. sp.
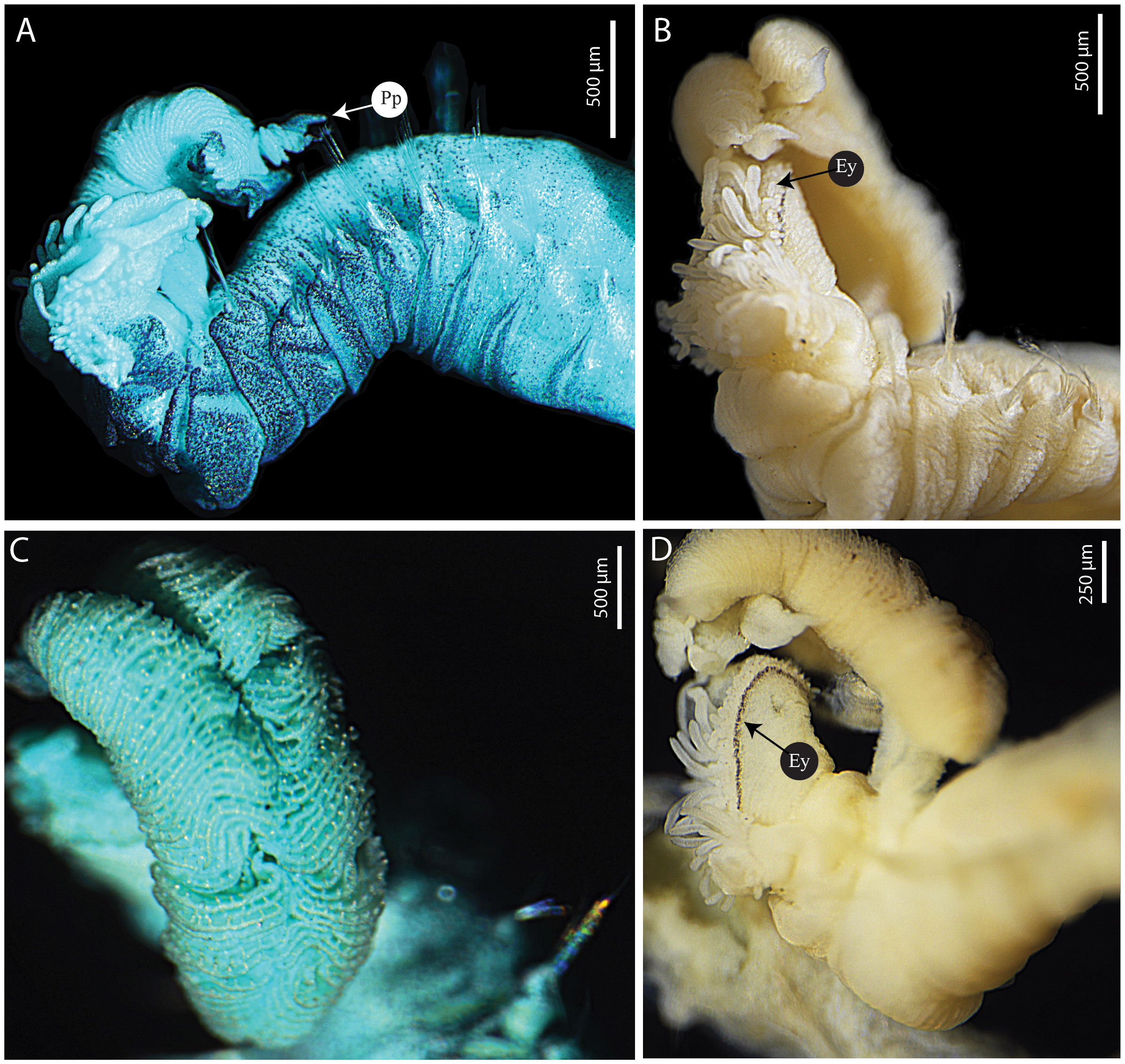
Figure 8 Terebellides oculata n. sp. (A) paratype MNHN-IA-2000-2080 (B–D) holotype MNHN-IA-2000-2079. (A) Anterior part, lateral view, MG staining. (B) Anterior view, fronto-lateral view. (C) Branchial lobes, dorsal view, MG staining. (D) Anterior part, lateral view. Ey, eyes; Pp, posterior projection.
Zoobank registration LSID: lsid:zoobank.org:act:CD39DA15-A0D6-4557-B2C9-950079DEF249.
Type material.
Holotype.
MNHN-IA-2000-2079, complete, some parapodia used for molecular analysis, South Pacific Ocean, Papua New Guinea, Nagada Harbour, PAPUA NIUGINI PD55, 5.15°S, 147.78°E, depth 5 m, coll. November 2012.
Paratypes.
All specimens from the same collection site as the holotype. Paratypes MNHN-IA-2000-2080, complete, mounted for SEM; MNHN-IA-2000-2081, complete, some parapodia used for molecular analysis.
Description (based on holotype, with variation in parentheses for paratypes).
Relatively small species, holotype 14.3 mm long (16.8–19.8 mm) and 1.0 mm wide (1.0–1.1 mm). Body tapering posteriorly, segments increasingly shorter and more compacted toward the pygidium.
Prostomium compact; eyespots present dorsally, surrounding the anterior margin of the prostomium (Figures 8B, D); upper lip large, with convoluted margin, surrounding mouth, with many buccal tentacles (Figures 8A, B, D). Buccal tentacles short, uniformly cylindrical with expanded tips, spatulate (Figures 8A, B, D). Lower lip forming an expanded structure below the upper lip, with transverse ridges (Figure 8B). SGs1 and 2 short, SG1 only visible ventrally, SG2 visible all around (Figures 8A, B, D). Lateral lappets on SG4–8 (TC1–6), largest on SG3 (TC1) and progressively shorter posteriorly (Figures 8A, B, D, 9A). Dorsal rounded projection on SG3–8 (TC1–6), more visible on SG4–5 (TC2–3) (Figures 8A, B, D, 9A). Glandular lateral region on SG5 (TC3) absent (Figures 8A, B, D).
Branchiae arising as a single structure from SG3 (TC1), reaching SG8 (SG6), consisting of a single elongate and annulated stalk placed mid-dorsally (Figures 8A, D, 9A, C), two pairs of lobes, and anterior branchial projection (fifth lobe) (Figures 6A, C, 8B, D); lobes fused for approximately 3/4 of length, lower pair thinner, upper lobes with approximately 65 tightly packed lamellae each (Figures 8A–D, 9A, C, D); ciliated tufts close to the margins of the lamellae present (type 2), visible under a stereomicroscope (Figures 8A–D, 9A, C, D); distal regions of the upper and lower lobes with pointed projections (Figures 8A, D, 9A, C).
Eighteen pairs of thoracic notopodia (SG3–20). First pair shorter than the subsequent notopodia; notochaetae from TC1 about the same size as those from the subsequent notopodia, and dorsally aligned (Figures 8A, B); all notochaetae simple capillaries, arranged in two rows, those from the anterior row slightly shorter. Neuropodia present as sessile pinnules from TC6 (SG8) to the last segment, uncini arranged in single rows from SG9; first pair of thoracic neuropodia (SG8) provided with five to seven bent acute tipped, geniculate chaetae (Figure 9B); subsequent thoracic neuropodia all with approximately 12–15 uncini per torus arranged in one irregular row (Figure 9E); uncini of type 5, with RvC = 1/0.7, four to five mid-sized teeth above the main fang, surmounted by two rows of minute denticles and an upper crest of several small denticles (Figure 9E); approximately 31 abdominal pairs of neuropodia (22), as erect paddle-shaped pinnules, entire margin provided with approximately 15–20 type 2 uncini, with RvC = 1/0.9, with four to five pointed teeth above the main fang, surmounted by a row of short pointed teeth and an upper crest of minute teeth (Figure 9F).
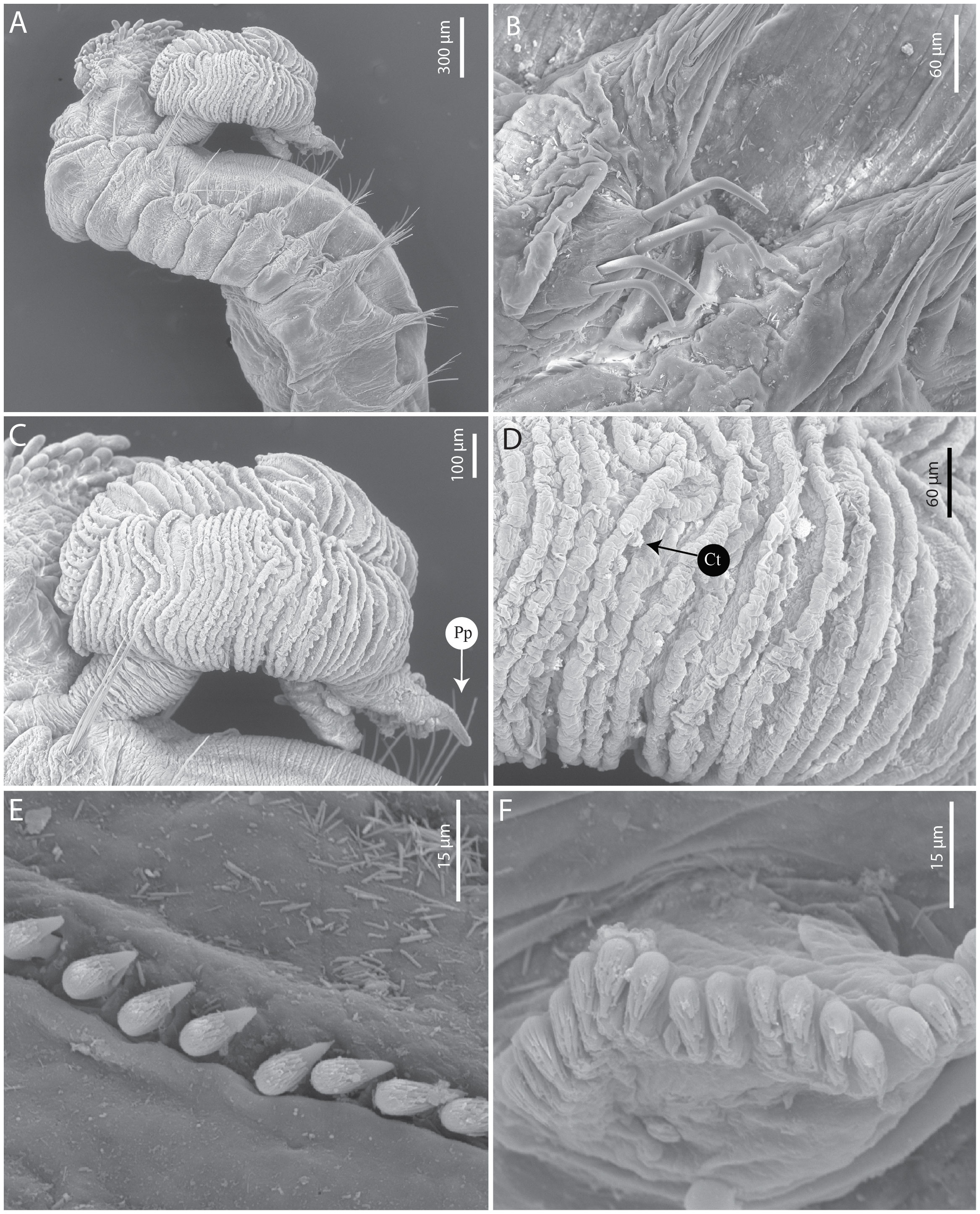
Figure 9 Terebellides oculata n. sp. (A–F) SEM paratype MNHN-IA-2000-2080. (A) Anterior part, lateral view. (B) Geniculate chaetae (TC6). (C) Branchial lobes, lateral view. (D) Branchial lamellae. (E) Uncini, thoracic chaetiger. (F) Uncini, abdominal chaetigers. Ct, ciliary tuft; Pp, posterior projection.
Two pairs of short nephridial papillae, located latero-posteriorly to the base of notopodia of SG6–7 (TC4–5). Pygidium crenulated (rounded), as funnel-like depression.
Methyl green staining pattern: the first seven segments stain solid; striped from SG8 to SG13, each stripe with a white transverse line (Figure 4); terminal projections of branchial lobes stain blue, anterior dorsum with small blue dots (Figure 8A); last segments with blue areas posterior to the neuropodia.
Etymology.
The species is named for the presence of eyespots, a rare characteristic among the species in this genus.
Habitat.
Coastal (harbor), 5 m depth.
Type locality.
South Pacific, Papua New Guinea, Nagada Harbour.
Remarks.
To date, the presence of eyes has only been described in another species of this genus: T. jitu. Although these eyespots may fade over time, they are still visible on the holotype and paratype of T. jitu, collected in 2005 and 1972, respectively. They are also visible on the type material of T. oculata n. sp., which was sampled in 2012. The diagnosis of the genus has been slightly modified to accommodate this character (see Diagnosis above).
Terebellides oculata n. sp. differs from T. jitu by the presence of dorsal rounded projections on the first six thoracic chaetigers, which are absent for T. jitu. The two species also differ by the shape of the thoracic uncini which belong to type 2 for T. jitu and to type 5 for T. oculata n. sp. Finally, T. oculata n. sp. differs by the presence of ciliated tufts close to the margins of the branchial lamellae, which are absent for T. jitu.
With the absence of a glandular region on the third thoracic chaetiger and with a branchia characterized by the absence of a fifth branchial lobe, the partial fusion of the other lobes and the absence of branchial papillae, T. oculata n. sp. is similar to other shallow water species, T. baliensis, T. kowinka, and T. woolawa (Table 2). However, T. oculata n. sp. differs from all these species by the presence of eyespots and from T. baliensis and T. kowinka by the presence of dorsal rounded projections on the first chaetigers. Finally, T. oculata n. sp. differs from T. woolawa by the methyl green pattern which is solid on the first seven segments for T. oculata n. sp. and on the first three segments only for T. woolawa.
Terebellides papillosa n. sp.
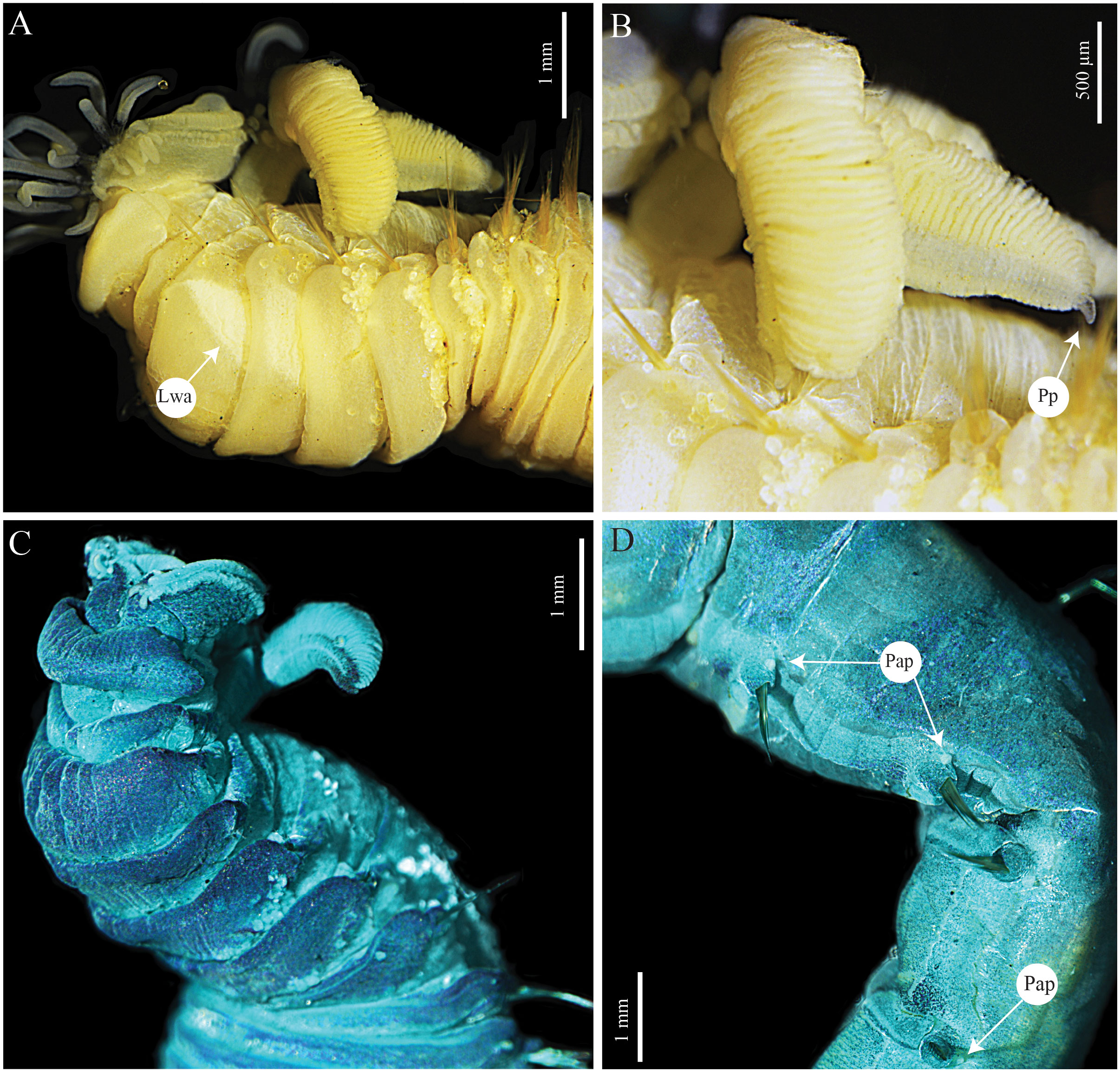
Figure 10 Terebellides papillosa n. sp. (A, B) holotype MNHN-IA-2000-2082 (C, D) paratype SEM MNHN-IA-2000-2083. (A) Anterior part, lateral view. (B) Anterior part, dorso-lateral view. (C) Anterior part, ventro-lateral view, MG staining. (D) Thoracic chaetigers TC14–18, lateral view. Lwa, lateral white areas; Np, nephridial papillae; Pp, posterior projections.
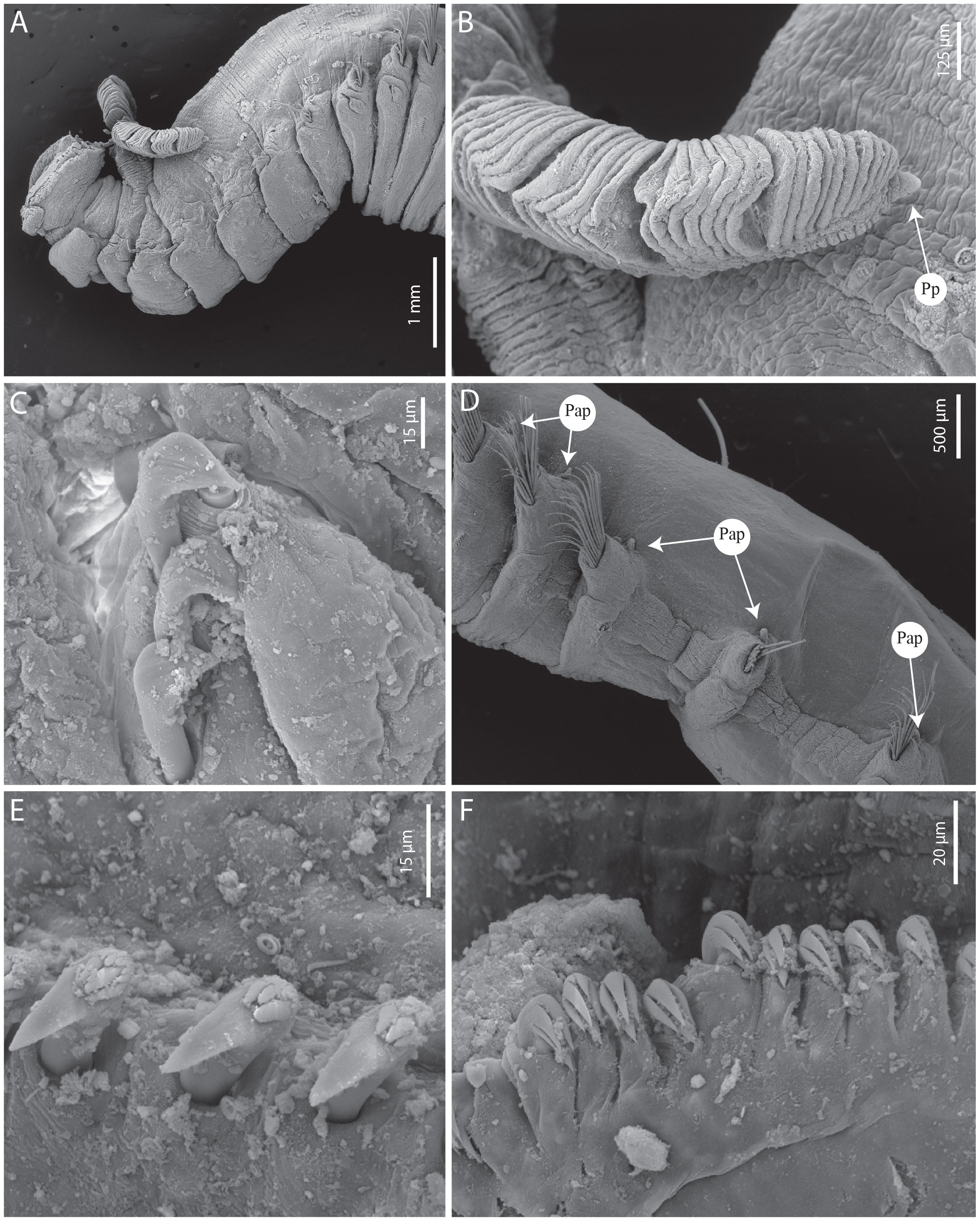
Figure 11 Terebellides papillosa n. sp. (A–E) SEM paratype MNHN-IA-2000-2083. (A) Anterior part, lateral view. (B) Branchial lobe, dorso-lateral view. (C) Geniculate chaetae (TC6), lateral view, MG staining. (D) Thoracic chaetigers (TC10–14), lateral view. (E) Uncini, thoracic chaetiger. (F) Uncini, abdominal chaetiger. Np, nephridial papillae; Pp, posterior projections.
Zoobank registration LSID: lsid:zoobank.org:act:F65A569E-52D8-4A5F-A4DD-D8427FD1E7E2.
Type material.
Holotype.
MNHN-IA-2000-2082, complete, some parapodia used for molecular analysis, South Pacific Ocean, Papua New Guinea, New Britain, Ainto Bay, MADEEP CP4329, 6.13°S, 149.16°E, depth 250–500 m, coll. May 2014.
Paratype.
From the same collection site as the holotype, paratype MNHN-IA-2000-2083, complete, some parapodia used for molecular analysis, mounted for SEM.
Description (based on holotype, with variation in parentheses for paratype).
Large species, holotype 41.1 mm long (39.2 mm) and 3.3 mm wide (3.1 mm). Body tapering posteriorly, segments increasingly shorter and more compacted toward the pygidium.
Prostomium compact; eyespots absent; a large upper lip surrounding the mouth, most buccal tentacles lost (Figures 10A–C); buccal tentacles uniformly cylindrical and with expanded tips, spatulate (Figures 10A, B). Lower lip forming an expanded structure below the upper lip (Figures 10A–C, 11A). SG1 well visible but almost completely covered by lobes on SG2; SG2 well visible, following segments with lobes as ventral collars (Figures 10A–C, 11A). Lateral lappets on SG4–9 (TC2–7), continuing ventrally in SG4–10 (Figures 10A–C, 11A); conspicuous dorsal rounded projections on SG 6–11 (TC4–9) (Figures 10A–C, 11A). Absence of a glandular lateral region on SG5 (TC3), but lateral whitish areas on SG3–4 (TC1–2) present, below the notopodia (Figures 10A, B).
Branchiae arising as a single structure from SG2, reaching SG8 (SG6), consisting of a single elongate and annulated stalk placed mid-dorsally (Figures 10A–C, 11A). Two pairs of lobes, not fused, but with one (two) lobe missing (fallen off); anterior branchial projection (fifth lobe) absent; upper lobes with approximately 35 packed foliaceous lamellae; absence of papillar projections or ciliated tufts over the margin of the branchial lamellae (type 1); distal region of both the upper and lower lobes with short digitiform projections (Figures 10A–C, 11A, B).
Eighteen pairs of thoracic notopodia (SG3–20). First two pairs shorter than the subsequent ones; notochaetae from SG3 (TC1) slightly shorter than the ones from the subsequent notopodia, notopodia all aligned vertically (Figures 10A, 11A); notochaetae as simple capillaries, arranged in two rows, those from the anterior row shorter (Figure 11D). Neuropodia present as sessile pinnules from TC6 (SG8) to the last segment; uncini arranged in single rows from SG10; first two pairs of thoracic neuropodia (SG8–9) with four to five short triangular geniculate chaetae, partially embedded in the body wall and difficult to see (Figure 11C); all subsequent thoracic neuropodia with approximately 8–15 uncini per torus arranged in one irregular row, type 1 uncini with RvC = 3/1, two to three teeth above the main fang, surmounted by a row of several minute denticles and an upper crest of several small denticles (Figure 11E). Approximately 30–35 abdominal pairs of neuropodia as erect paddle-shaped pinnules, with approximately 35 uncini on margin; type 1A uncini, with RvC = 1/0.7, three to four pointed teeth above the main fang, surmounted by a row of one to two shorter pointed teeth and an upper crest of minute teeth (Figure 11F).
Presence of papillae from SG11 to the end of the thorax (SG20), small, flattened, one per segment, inserted dorsally to the base of each notopodia. Pygidium rounded, as funnel-like depression, with smooth margins.
Methyl green staining pattern: the first seven segments stain solid; SG8–13 with distinct stripes (Figure 4); ventral faces of branchial lobes stain dark blue (Figure 10C).
Etymology.
The species name refers to the unusual number of papillae on thoracic segments.
Habitat.
Between 250 and 500 m depth, mud with a lot of plant debris and sunken wood.
Type locality.
South Pacific, Papua New Guinea, New Britain, Ainto Bay.
Remarks.
Among the Central Indo-Pacific species with geniculate chaetae on two segments instead of one, Terebellides papillosa n. sp. is similar to T. akares, T. ectopium, and T. intoshi (Table 2).
Terebellides papillosa n. sp. differs from T. akares and T. ectopium by the geniculate chaetae present on SG8 and SG9 instead of SG7 and SG8 for these two species, also by the absence of a fifth branchial lobe and the presence of dorsal rounded projections on the first chaetigers. Terebellides papillosa n. sp. also differs from T. akares by the absence of branchial papillae in the latter species and from T. ectopium by the presence of branchial lobes completely free from each other (Table 2).
Terebellides papillosa n. sp. differs from T. intoshi by the number of thoracic uncini per torus, with 10–15 uncini on each neuropodium compared to patches of 50 uncini present on each T. intoshi neuropodia. This latter species is also characterized by the absence of digitiform distal tips on both the upper and lower branchial lobes, which are present in Terebellides papillosa n. sp. Finally, Terebellides papillosa n. sp. is characterized by a large number of segments with papillae, from SG11 to the end of the notopodia (Figure 11D) and geniculate chaetae with a very unusual triangular shape (Figure 11C; Table 2).
Molecular data
During this study, we managed to obtain eight sequences of 16S and only three for COI. The 16S sequences corresponded to four of the new species described here, with T. madeep n. sp. the only species for which no sequence could be obtained. Another sequence belonging to an unnamed species (MNHN-IA-2021-73), probably new, was acquired, but the specimen was too degraded to be formally described (Figure 12, Table 1). For COI, the three sequences belong to three different species: T. fauchaldi n. sp., T oculata n. sp., and T. papillosa n. sp.
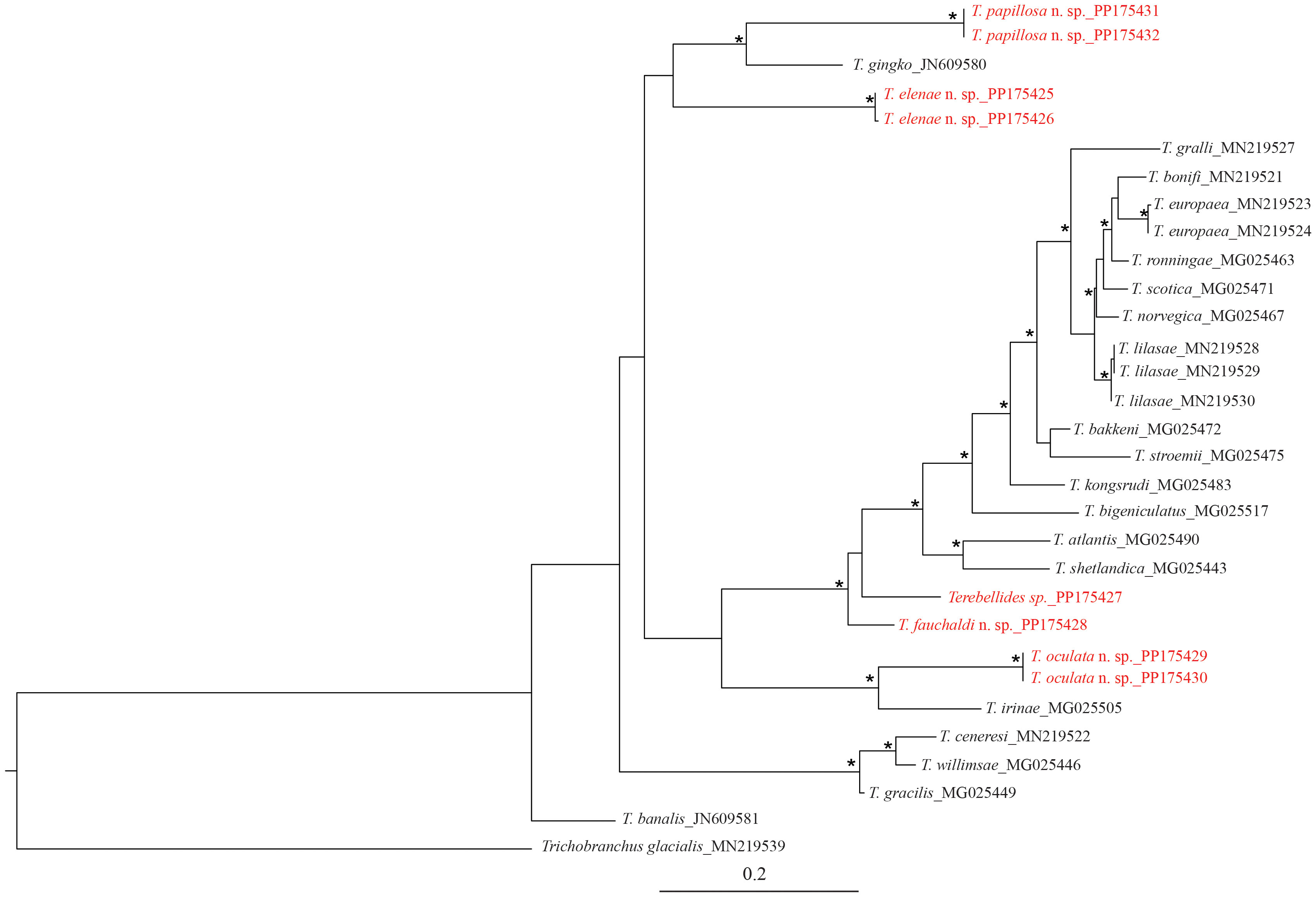
Figure 12 Maximum likelihood tree of Terebellides species based on 16S sequences. Asterisks indicate the bootstrap support values of the ML analysis >80%. Text in red indicates specimens sequenced in this study.
Based on 16S, T. papillosa n. sp. and T. elenae n. sp. constituted a sister group to T. ginko Schüller and Hutchings, 2012. On the other side, T. fauchaldi, T. oculata, and Terebellides sp. (MNHN-IA-2021-73) are grouped with a large clade of European species (Figure 12).
Key to Terebellides species from the Central Indo-Pacific region.
1A. Prostomial eyespsots present…2.
1B. Prostomial eyespots absent… 3.
2A. Dorsal rounded projections present on the first chaetigers; thoracic uncini of type 5; ciliated tufts close to the margins of branchial lamellae present … T. oculata n. sp.
2B. Dorsal rounded projections on the first chaetigers absent; thoracic uncini of type 2; ciliated tufts absent … T. jitu Schuller and Hutchings, 2010.
3A. Geniculate chaetae present on two chaetigers… 4.
3B. Geniculate chaetae present on a single chaetiger… 7.
4A. Geniculate chaetae present on TC5–6… 5.
4B. Geniculate chaetae present on TC6–7… 6.
5A. Branchial lamellae margins with papillae; branchial lobes not fused … T. akares Hutchings et al., 2015.
5B. Branchial lamellae margins without papillae; branchial lobes fused over 2/3 of their length … T. ectopium Zhang and Hutchings, 2018.
6A. Branchial lobes terminating by relatively long filaments; thoracic neuropodia with approximately 50 uncini each; sharply bent and acute geniculate chaetae … T. intoshi Caullery, 1915.
6B. Branchial lobes terminating by short projections; thoracic neuropodia with 10–15 uncini each; geniculate chaetae short and triangular … T. papillosa n. sp.
7A. Glandular region on TC3 present… 8.
7B. Glandular region on TC3 absent… 11.
8A. Fifth branchial lobe present… 9.
8B. Fifth branchial lobe absent … Terebellides madeep n. sp.
9A. Branchial lobes lacking terminal papillae; anterior thoracic chaetigers lacking rounded dorsal projections … T. narribri Hutchings and Peart, 2000.
9B. Branchial tips with terminal papillae; anterior thoracic chaetigers with rounded dorsal projections, more developed on TC3… 10.
10A. Ventral branchial lobes terminating in filamentous tips; glandular region on TC3 thin, J-shaped … T. yangi Zhang and Hutchings, 2018.
10B. Ventral branchial lobes terminating in short terminal projections; glandular region on TC3 large, V-shaped … Terebellides fauchaldi n. sp.
11A. Margins of branchial lamellae with papillae… 12.
11B. Margins of branchial lamellae without papillae… 14.
12A.TC2 smaller than the following ones; dorsal crests present on SG5–6… T. jorgeni Hutchings, 2007.
12B. TC2 the same size as the following chaetigers; dorsal crests on SG5–6 absent…13.
13A. Branchial lobes fused over 3/5 of their length; each thoracic neuropodium with 23–25 uncini … T. guangdongensis Zhang and Hutchings, 2018.
13B. Branchial lobes not fused; thoracic neuropodia with 8–10 uncini each … T. hutchingsae Parapar, Moreira and Martin, 2016.
14A. Branchial lobes fused at least partially… 15.
14B. Branchial lobes totally free from each other… 17.
15A. Fifth branchial lobe present… 16.
15B. Fifth branchial lobe absent … T. kowinka Hutchings and Peart, 2000.
16A. TC1 well developed; branchial lobes almost entirely fused; deep-sea species (350 m depth)… Terebellides elenae n. sp.
16B. TC1 reduced; branchial lobes fused over 50% of their length; intertidal to shallow subtidal … T. woolawa Hutchings and Peart, 2000.
17A. Branchial lobes with filamentous tips … T. ehlersi McIntosh, 1885.
17B. Branchial lobes with pointed tips … T. baliensis Hsueh and Li, 2017.
Data availability statement
The original contributions presented in the study are publicly available. The type specimens presented in this study are deposited in the MNHN collection, accession numbers MNHN-IA-2000-2071 to MNHN-IA-2000-2083. The 16 sequences (accession numbers: PP175425 to PP175432) and COI sequences (accession numbers: PP156714 to PP156716) are deposited in GenBank.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because DNA analyses were conducted on dead marine invertebrates from museum collections.
Author contributions
NL: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. JN: Funding acquisition, Software, Supervision, Validation, Visualization, Writing – review & editing. GD: Data curation, Formal analysis, Methodology, Software, Validation, Writing – review & editing. PH: Investigation, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. NL and GD have received financial support from the French State in the frame of the “Investments for the future” Programme IdEx Bordeaux, reference ANR-10-IDEX-03-02. JN receives a productivity grant from CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil).
Acknowledgments
Type material was collected during the two MADEEP and PAPUA NIUGINI expeditions. MADEEP deep sea cruise (PIs: Sarah Samadi, Laure Corbari, Karine Olu-Le Roy) took place in April and May 2014 on board RV ‘Alis’ deployed by the Institut de Recherche pour le Développement (IRD). It operated under a Memorandum of Understanding with the University of Papua New Guinea (UPNG), with a permit given by the Papua New Guinea Department of Environment and Conservation (DEC). PIs acknowledge the funding from Agence Nationale de la Recherche (ANR) and National Science Council of Taiwan (ANR TF-DeepEvo 12 ISV7 005 01) and the CNRS Institut Ecologie et Environnement (INEE) (https://doi.org/10.17600/14004000). Madang 2012 expedition (PIs: Philippe Bouchet, Claude Payri, and Sarah Samadi) was part of the “Our Planet Reviewed” PAPUA NIUGINI project organized by the Muséum National d’Histoire Naturelle (MNHN), Pro Natura International (PNI), Institut de Recherche pour le Développement (IRD), and University of Papua New Guinea (UPNG). The organizers acknowledge the funding from the Total Foundation, Prince Albert II of Monaco Foundation, Fondation EDF, Stavros Niarchos Foundation and Entrepose Contracting, in-kind support from the Divine Word University (DWU), and post-expedition support from Agence Nationale de la Recherche (ANR) and National Science Council of Taiwan (ANR TF-DeepEvo 12 ISV7 005 01). The expedition operated under a Memorandum of Understanding with the University of Papua New Guinea (UPNG), with a permit delivered by the Papua New Guinea Department of Environment and Conservation (DEC) (https://doi.org/10.17600/18000841). We also want to thank Stéphane Hourdez, Céline Houbin, Céline Labrune, Jérome Jourde, and Joachim Langeneck for their help in sorting all worms collected during this cruise, during the MNHN workshop in 2021. Finally, we want to thank the three reviewers for their critical reviews of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
COI, cytochrome c oxidase subunit I; MNHN, Muséum National d’Histoire Naturelle (Paris, France); MG, methyl green; RVC, rostrum vs. capitium length ratio; SEM, scanning electron microscope; SG, segment; TC, thoracic chaetiger.
References
Barroso M., Moreira J., Capa M., Nygren A., Parapar J. (2022). A further step towards the characterisation of Terebellides (Annelida, Trichobranchidae) diversity in the Northeast Atlantic, with the description of a new species. ZooKeys 1132, 85–126. doi: 10.3897/zookeys.1132.91244
Caullery M. (1915). Sur les Terebellides Malmgren du Siboga et les Térébelliens voisins. Bull. la Société Zoologique France 40, 111–116.
Corbari L., Frutos I., Sorbe J. C. (2019). Dorotea gen. nov., a new bathyal genus (Amphipoda, Eusiridae) from the Solomon Sea (Papua New Guinea). Zootaxa 4568, 69–80. doi: 10.11646/zootaxa.4568.1.4
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Grube A. E. (1878). Annulata Semperiana. Beiträge zur Kenntniss der Annelidenfauna der Philippinen nach den von Herrn Prof. Semper mitgebrachten Sammlungen. Mémoires l’Académie Impériale des Sciences de St.- Pétersbourg (série 7), Vol. 25. 1–300.
Hsueh P.-W., Li K.-R. (2017). Additions of new species to Thelepus (Thelepodidae), with description of a new Terebellides (Trichobranchidae) from Taiwan. Zootaxa 4244, 429–439. doi: 10.11646/zootaxa.4244.3.10
Hutchings P. (2007). New species of deep-sea Terebellidae and Trichobranchidae (Polychaeta) (sedentary species III). Galathea Rep. 21, 75–90.
Hutchings P., Kupriyanova E. (2018). Cosmopolitan polychaetes – fact or fiction? Personal and historical perspectives. Invertebrate Systematics 32, 1–9. doi: 10.1071/IS17035
Hutchings P., Lavesque N. (2020). I know who you are, but do others know? Why correct names are so important. Zoosymposia 19, 151–163. doi: 10.11646/zoosymposia.19.1.16
Hutchings P., Nogueira J. M. M., Carrerette O. (2015). Telothelepodidae, thelepodidae and trichobranchidae (Annelida, terebelliformia) from lizard island, great barrier reef, Australia. Zootaxa 4019, 240–274. doi: 10.11646/zootaxa.4019.1.12
Hutchings P., Nogueira J. M. M., Carrerette O. (2021). “Terebellidae johnsto,” in Handbook of zoology. A natural history of the phyla of the animal kingdom. Eds. Schmidt-Rhaesa A., Beutel R. G., Glaubrecht M., Kristensen N. P., Prendini L., Purschke G., Richter S., Westheide W., Leschen R. Z. E. (Walter de Gruyter & Co, Berlin), 1–64.
Hutchings P., Peart R. (2000). A revision of the Australian trichobranchidae (Polychaeta). Invertebrate Taxonomy 14, 225–272. doi: 10.1071/IT98005
Katoh K., Misawa K., Kuma K., Miyata T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059–3066. doi: 10.1093/nar/gkf436
Kinberg J. G. H. (1867). “Annulata nova,” in Öfversigt af Königlich Vetenskapsakademiens förhandlingar, vol. 23. (Stockholm), 97–103.
Lavesque N., Daffe G., Glasby C., Hourdez S., Hutchings P. (2022). Three new deep-sea species of Marphysa (Annelida, Polychaeta, Eunicida, Eunicidae) from Papua New Guinea (Bismarck and Solomon seas). Zookeys 122, 81–105. doi: 10.3897/zookeys.1122.89990
Lavesque N., Hutchings P., Daffe G., Nygren A., Londoño-Mesa M. H. (2019). A revision of the French Trichobranchidae (Polychaeta), with descriptions of nine new species. Zootaxa 4664, 151–190. doi: 10.11646/zootaxa.4664.2.1
Lavesque N., Hutchings P., Londoño-Mesa M. H., Nogueira J. M. N. N., Daffe G., Nygren A., et al. (2021). The “Spaghetti Project”: the final identification guide to European Terebellidae (sensu lato) (Annelida, Terebelliformia). Eur. J. Taxonomy 782, 108–156. doi: 10.5852/ejt.2021.782.1593
Malmgren A. J. (1866). Nordiska hafs-annulater. Öfversigt af Kongiliga Veteskaps-Akademiens Förhandlingar 22, 355–410.
McIntosh W. C. (1885). Report on the Annelida Polychaeta collected by H.M.S. Challenger during the years 1873-1876. Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–76. Zoology. 12:1–554. https://biodiversitylibrary.org/page/50688426.
Minh B. Q., Nguyen M. A. T., Von Haeseler A. (2013). Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 30, 1188–1195. doi: 10.1093/molbev/mst024
Nygren A., Parapar J., Pons J., Meißner K., Bakken T., Kongsrud J. A., et al. (2018). A megacryptic species complex hidden among one of the most common annelids in the North East Atlantic. PloS One 13, e0198356. doi: 10.1371/journal.pone.0198356
Palumbi S. R., Martin A., Romano S., McMillan W. O., Stice L., Grabowski G. (1991). The simple fool’s guide to PCR, version 2 (Honolulu: University of Hawaii Zoology Department).
Pante E., Corbari L., Thubaut J., Chan T. Y., Mana R., Boisselier M. C., et al. (2012). Exploration of the deep-sea fauna of Papua New Guinea. Oceanography 25, 214–225. doi: 10.5670/oceanog.2012.65
Parapar J., Capa M., Nygren A., Moreira J. (2020a). To name but a few: Descriptions of five new species of Terebellides (Annelida, Trichobranchidae) from the North-East Atlantic. ZooKeys 992, 1–58. doi: 10.3897/zookeys.992.55977.figure10
Parapar J., Hutchings P. (2014). Redescription of Terebellides stroemii (Polychaeta, Trichobranchidae) and designation of a neotype. J. Mar. Biol. Assoc. United Kingdom 95, 323–337. doi: 10.1017/S0025315414000903
Parapar J., Martin D., Moreira J. (2020b). On the diversity of Terebellides (Annelida, Trichobranchidae) in West Africa, seven new species and the redescription of T. africana Augener 1918 stat. prom. Zootaxa 4771, 1–61. doi: 10.11646/zootaxa.4771.1.1
Parapar J., Moreira J., Gil J., Martin D. (2016c). A new species of the genus Terebellides (Polychaeta, Trichobranchidae) from the Iranian coast. Zootaxa 4117, 321–340. doi: 10.11646/zootaxa.4117.3.2
Parapar J., Moreira J., Martin D. (2016b). On the diversity of the SE Indo-Pacific species of Terebellides (Annelida; Trichobranchidae), with the description of a new species. PeerJ 4, e2313. doi: 10.7717/peerj.2313
Parapar J., Moreira J., O’Reilly M. (2016a). A new species of Terebellides (Polychaeta: Trichobranchidae) from Scottish waters with an insight into branchial morphology. Mar. Biodiversity 46, 211–225. doi: 10.1007/s12526-015-0353-5
Quatrefages A. (1866). “Histoire naturelle des Annelés marins et d’eau douce,” in Annélides et géphyriens., vol. 1. (Librairie Encyclopédique de Roret, Paris).
Read G., Fauchald K. (2023) World polychaeta database. Terebellides sars 1835. Available online at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=129717 (Accessed October 31, 2023).
Sars M. (1835). Beskrivelser og Iagttagelser over nogle maerkelige eller nye i Havet ved den Bergenske Kyst Levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes og Molluskernes classer, med en kort Oversigt over de hidtil af Forfatteren sammesteds fundne Arter og deres Forekommen. Ed. Hallager T. (Bergen, 81 pp). doi: 10.5962/bhl.title.13017
Schüller M., Hutchings P. (2010). New insights in the taxonomy of Trichobranchidae (Polychaeta) with description of a new Terebellides species from Australia. Zootaxa 2395, 1–16. doi: 10.11646/zootaxa.2395.1
Schüller M., Hutchings P. A. (2012). New species of Terebellides (Polychaeta: Trichobranchidae) indicate long-distance dispersal between western South Atlantic deep-sea basins. Zootaxa 3254, 1–31.
Schüller M., Hutchings P. A. (2013). Terebellides from the Southern Ocean with a key to all described species. Zootaxa, 3619, 001–045. doi: 10.11646/zootaxa.3619.1.1
Sjölin E., Erseus C., Källersjö M. (2005). Phylogeny of Tubificidae (Annelida, Clitellata) based on mitochondrial and nuclear sequence data. Mol. Phylogenet. Evol. 35, 431–441. doi: 10.1016/j.ympev.2004.12.018
Spalding M. D., Fox H. E., Allen G. R., Davidson N., Ferdaña Z. A., Finlayson M., et al. (2007). Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57, 573–583. doi: 10.1641/B570707
Trifinopoulos J., Nguyen L. T., Von Haeseler A., Minh B. Q. (2016). W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, W232–W235. doi: 10.1093/nar/gkw256
Keywords: new species, molecular, morphology, taxonomy, Terebellides
Citation: Lavesque N, Nogueira JMM, Daffe G and Hutchings P (2024) Five new species of Terebellides (Annelida, Polychaeta, Trichobranchidae) from Papua New Guinea (Bismarck and Solomon seas). Front. Ecol. Evol. 12:1349362. doi: 10.3389/fevo.2024.1349362
Received: 04 December 2023; Accepted: 30 January 2024;
Published: 12 March 2024.
Edited by:
Paulo Cesar Paiva, Federal University of Rio de Janeiro, BrazilReviewed by:
Rômulo Barroso, Federal University of Bahia (UFBA), BrazilJuan Moreira Da Rocha, Autonomous University of Madrid, Spain
Julio Parapar, University of A Coruña, Spain
Copyright © 2024 Lavesque, Nogueira, Daffe and Hutchings. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicolas Lavesque, bmljb2xhcy5sYXZlc3F1ZUB1LWJvcmRlYXV4LmZy
†ORCID: Nicolas Lavesque, orcid.org/0000-0001-5701-2393
João M. M. Nogueira, orcid.org/0000-0002-8450-4294
Guillemine Daffe, orcid.org/0000-0002-7085-3151
Pat Hutchings, orcid.org/0000-0001-7521-3930