
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
HYPOTHESIS AND THEORY article
Front. Ecol. Evol. , 22 April 2024
Sec. Evolutionary Developmental Biology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1343346
This article is part of the Research Topic The Future of the Homology Concept under Novel Evolutionary Paradigms View all 5 articles
 Torben Göpel1,2,3*
Torben Göpel1,2,3*Morphology, the scientific discipline dealing with description and comparison of organismal form, is one of the oldest disciplines in biology and traditionally strongly linked to the concept of homology. With morphological data being used and morphological knowledge being applied in other (younger) biological disciplines, morphology has often been degraded to an only auxiliary discipline or a mere set of methods serving those other disciplines. While this notion has been wrong all along, the last decades have seen a renaissance of morphology mostly due to significant leaps in imaging techniques and the advent of 3D digital data. Modern large-scale morphological endeavors in what is called phenomics and new means of functional analyses underline the fruitfulness of morphological research. Furthermore, morphology has been revisited on a conceptual level leading to a “re-philosophication” of morphology acknowledging its nature as explanatory science. Based on Richter and Wirkner’s research program of Evolutionary Morphology, this essay expands the conceptual framework to identify entities and processes vital for morphology as independent scientific discipline. With no unified homology concept in sight (and maybe not even desired), following the emergence of bio-ontologies in morphology, a plea is made for conceptual explicitness which acknowledges the plurality of homology concepts but enables intersubjective transfer.
Morphology is one of the oldest disciplines in biology utilizing perception, description, categorization and comparison (of biological forms) which are basic processes of human cognition. In the last decades morphology has been degraded by some to being only an auxiliary methodology, a tool used for other biological disciplines (Ghiselin, 1980, 2006), while others pointed out the epistemic value of morphology (Sudhaus, 2007; Scholtz, 2010; Wirkner and Richter, 2010; Richter and Wirkner, 2014). Advocates of the position that morphology is a merely descriptive, non-explanatory endeavor, which has “so little to contribute” (Ghiselin, 1980, p. 181; Ghiselin, 2006) attribute all gain of knowledge to other disciplines, first and foremost to evolutionary biology, population genetics and systematics (Ghiselin, 2006 and literature therein). Ghiselin’s argument is grounded in a distinction of the terms morphology and anatomy where the former is the mere description and only the latter includes comparison and assessment to causal explanations (Ghiselin, 2006). This distinction of the two terms, however, is anything but undisputed and anatomy is often regarded a sub-discipline of morphology restricted to internal structures (Hanser et al., 2000). Already, Goethe (1817), who introduced the term “Morphologie”, stated that there is more to morphology than the descriptive level:
“Die Erfahrung muß uns vorerst die Theile lehren, die allen Thieren gemein sind, und worin diese Theile verschieden sind. Die Idee muß über dem Ganzen walten und auf eine genetische Weise das allgemeine Bild abziehen” (Goethe, 1817, p. 151).
Experience must first teach us the parts which are common to all animals, and in what these parts are different. The idea must rule over the whole and draw the general picture in a genetic way. (own translation)
“Indem wir jenen Typus aufstellen und als eine allgemeine Norm […] denken, setzen wir in der Natur eine gewisse Konsequenz voraus, wir trauen ihr zu, daß sie in allen einzelnen Fällen nach einer gewissen Regel verfahren werde.” (Goethe, 1817, p. 171).
By establishing the type and thinking of it as a general norm […], we assume a certain consistency in nature. We trust it to proceed according to a certain rule in all individual cases. (own translation)
Goethe distinguished between (1) describing and comparing parts (which might be referred to as “morphemes”; see below), (2) comparing these parts and thus the organisms, and (3) the search for the type which points towards a common cause for our morphological observations which is inherently natural. Ghiselin (2006) argues that pure morphology is grounded in idealistic, typological morphology based on Goethe and thus cannot contribute to evolutionary understanding. It is necessary, however, to depict Goethe’s view of morphology in the context of the pre-Darwinian time in which it was developed (Richter and Wirkner, 2013). Goethe clearly stated that morphological observations are caused by a rule/law of nature towards which the type hints (Goethe, 1817, p.171). His concept of a type should therefore be understood as an auxiliary construct in lack of a proper understanding of the predicted certain rule of nature (“gewisse[n] Regel”; Goethe, 1817, p.171) leading to morphological disparity which since Darwin (1859) is broadly accepted to be descent with modification. As will be shown below, Goethe’s basic conceptualization of morphology still applies after more than 200 years recognizing the distinct processes of (1) description and (3) explanation with (2) comparison as the essential connection (Figure 1).
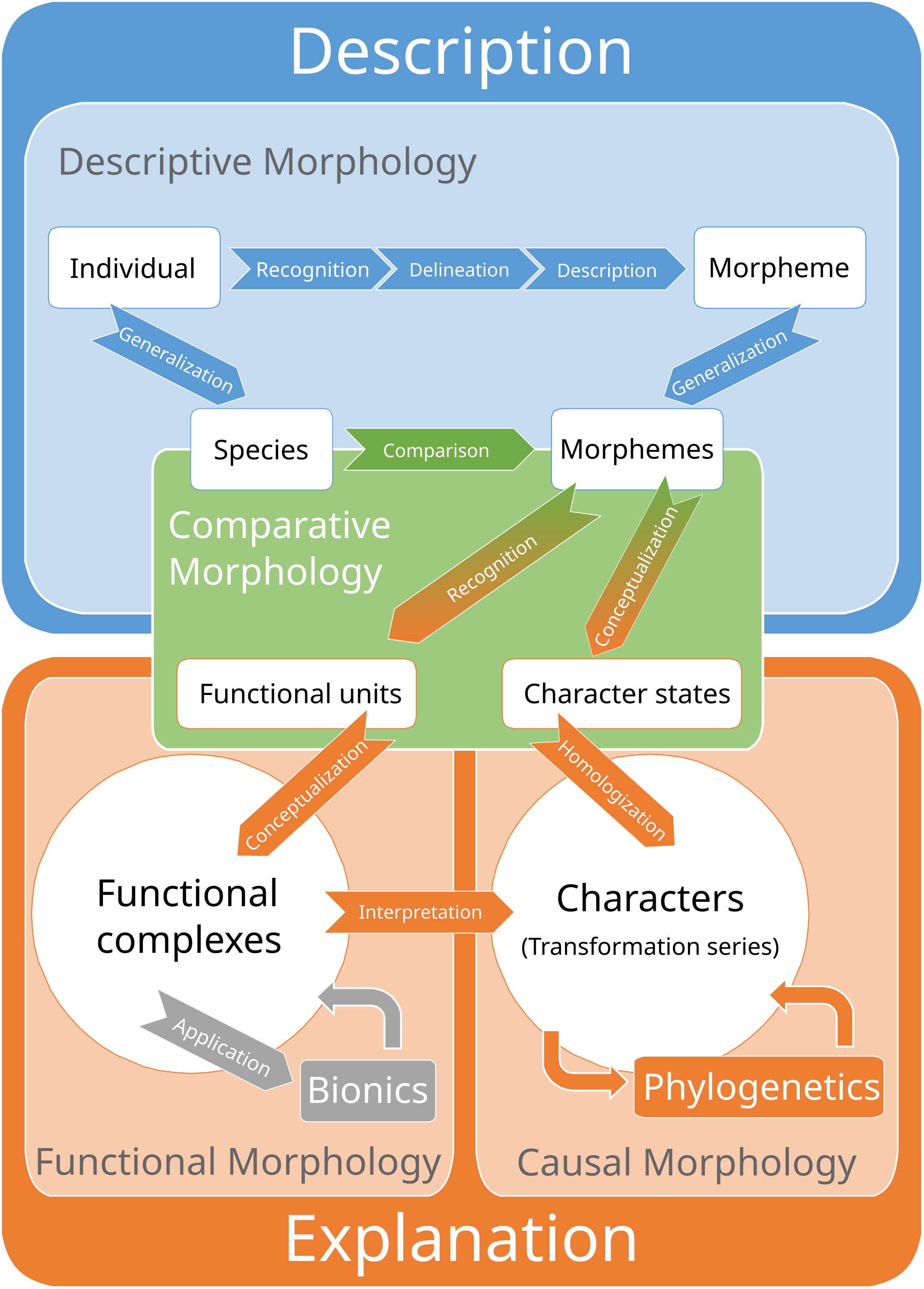
Figure 1 Conceptual framework of evolutionary morphology with assignment of sub-disciplines to the descriptive and explanatory level. White-filled elements represent the entities and arrows represent processes important in the respective sub-discipline. Based on the research program for Evolutionary Morphology by Richter and Wirkner (2014).
In the post-Darwinian era, morphology has continued to search for causal factors. Riedl (2000) defines morphology as the science of biological shape (Gestalt) and their deciphering (see also Richter and Wirkner, 2013). Implication of findings from developmental biology (Haeckel, 1866; Spemann, 1901) and functional morphology (Bock and von Wahlert, 1965; Seilacher, 1970) complemented the comparative approaches. Thus, morphology seeks to explore and explain disparity of biological forms (Scholtz, 2010) in all its aspects, which therefore includes also mechanistic (development), biomechanical (function) and phylogenetic (evolution) causes (Love, 2003, 2006; Richter and Wirkner, 2013, 2014; Sudhaus, 2007; Wirkner and Richter, 2010). Morphology exceeds the descriptive level (although descriptions constitute the primary data basis) and acts as explanatory science (Bock, 1991; Riedl, 2000; Richter and Wirkner, 2014). Since Darwin (1859) showed that evolution – as descent with modification – is the overarching driving force for all biological diversity, morphology can only be thought of as an evolutionary morphology (Richter and Wirkner, 2013, 2014). Under this paradigm that all explanation of biological form is evolutionary in some way, the terms morphology and evolutionary morphology may even be used as synonyms when referring to the scientific discipline, although the latter is more explicit as to what this discipline encompasses. Every morphological effort after the mere description points towards (in the end evolutionary) causalities: Comparative morphology compares either within or between species. Comparison within a species allows for identification of intraspecific variability which, among others, is important for the conceptualization of character states (Pigliucci, 2001; Wirkner and Richter, 2010; Richter and Wirkner, 2014; Wagner, 2014; Göpel and Richter, 2016). Differences between individuals of the same species can be either genetically determined (e.g. polymorphism) or subject to phenotypic plasticity. Although unclear in most cases, assessment on this question is of importance as only inherited features can be hypothesized to constitute a putative character state of said species (Richter and Wirkner, 2013; Wagner, 2014). Comparison between species considers differences and correspondences to formulate hypotheses on homology (Richter, 2005; Szucsich and Wirkner, 2007; Wirkner and Richter, 2010). The homology hypotheses derived from species comparison yields morphological characters and character states suitable for further investigation, e.g. phylogenetic analyses and/or reconstruction of character transformation (e.g. character mapping; Swofford and Maddison, 1992). Functional morphology investigates the biomechanical attributes and properties of biological forms (function sensu Bock and von Wahlert, 1965) and observes the biological role which these form–function complexes fulfill in nature. Besides the mere working principle (Drack et al., 2018; Drack and Betz, 2022) derived from the form, functional morphology, via investigation of the biological role, observes morphology acting in the environment of the organism (Bock and von Wahlert, 1965). This way, functional morphology approaches e.g. adaptation as causal factor for biological form (but see also e.g. Gould and Lewontin, 1979 for an expansion of the adaptationist programme). The ultimate level of explanation, i.e. evolution, is reached in the reconstruction of evolutionary transformation (Sudhaus, 2007; Wirkner and Richter, 2010; Richter and Wirkner, 2014). Phylogenetic relationships enable reconstruction of ancestral character states to derive hypotheses on the sequence of character state transformations (Swofford and Maddison, 1987; Wirkner and Richter, 2010).
In practice, reconstruction of ancestral states and character state transformation is performed either using phylogenetic hypotheses that have been derived from (solely or among others) said morphological characters (e.g. Wirkner and Richter, 2010) or by mapping morphological characters onto phylogenetic hypotheses inferred from other (mostly molecular) data (e.g. Shen et al., 2023). The first approach is straightforward as the inference of ancestral states is part of the inference of the tree (Richter, 2005).
Solely relying on molecular data for inference of phylogenetic hypotheses followed by mapping of morphological characters onto these trees has become a common practice given the disproportionately large datasets available and actually processable in phylogenomics. However, this practice can be criticized for different reasons: For instance, (1) there is no general primate of molecules over the organism’s morphology. Morphology does not per se follow molecular evolution as in many cases morphological entities interact with the environment and thus are subject to selective pressure and evolutionary processes can often only be understood with integration of morphological or other phenotypic data. And there might even be functionally impossible scenarios of character transformation resulting from solely relying on molecular phylogenies (Sudhaus, 2007). (2) Epistemologically, a phylogenetic hypothesis inferred from molecular data serves as explanatory hypothesis for the data included in the inference only, therefore rendering reconstruction of ancestral states for characters not used for the inference in the first place invalid (Fitzhugh, 2014, 2016).
While the first criticism is certainly true, mapping of morphological characters might not directly lead to other phylogenetic hypotheses but can hint towards the interesting contradictions between morphological and molecular data. Discovered functional impossibility or implausibility can spark reconsideration of the initial phylogenetic hypotheses. The second point focusing on the epistemology of abductive inference in general, while formally correct, misses the aspect that a phylogenetic hypothesis is a hypothesis on past population splitting events dealing with groups of integral organisms. Phylogeny is a singular historic process, and the single characters of an organism cannot have a phylogeny independent of that organism’s phylogeny. Thus, all characters logically must follow the phylogeny of the organisms that they are a part of, e.g. there cannot be a phylogenetic history of duck bills that is logically independent and different from the phylogeny of ducks. Character mapping uses the phylogenetic hypothesis in a “given this scenario”-approach, i.e. if this was the phylogeny of the studied taxa (and therefore all their genotypic and phenotypic traits), what would be the implications on the sequences of evolutionary transformation of certain characters (Göpel and Wirkner, 2018).
These sequences of character transformation provide a strictly historical perspective (Wirkner and Richter, 2010; Richter and Wirkner, 2014) and can then be complemented with other levels of causality (e.g. development, function/biological role, selection, population genetics; Sudhaus, 2007), to derive comprehensive scenarios of evolutionary transformation of characters (Hennig, 1966).
The last two decades have seen new imaging technologies, such as micro-computed X-ray tomography (Wirkner and Richter, 2004; Metscher, 2009) or light-sheet microscopy (Huisken et al., 2004; Weber et al., 2014), providing unprecedented insight into form, function, and development. Meanwhile, the above-mentioned degradation of morphology to a mere tool as well as the reproach that morphology – due to its historical background – was laden with idealistic and typological thinking has also led to a “re-philosophication” of morphology, i.e. a reconsideration of morphology on a conceptual level and more elaborate understanding of the scientific process of morphology (Sudhaus, 2007; Scholtz, 2010; Wirkner and Richter, 2010; Deans et al., 2012; Richter and Wirkner, 2014; Deans et al., 2015).
Not only general concepts and terminology of evolutionary morphology need consideration but also special terminology – e.g. to address certain parts of organisms (i.e. morphemes; Richter and Wirkner, 2014; Göpel and Richter, 2016), their attributes and qualities – needs to be clear and understandable. Vogt et al. (2010) described this as the “linguistic problem of morphology”, a key issue to be solved in order to enable integrative, large scale morphological approaches, often called “phenomics” (Houle et al., 2010; Burleigh et al., 2013; Deans et al., 2015). The linguistic problem consists of three main aspects (Vogt et al., 2010): (1) Morphology lacks a standardized terminology accepted in the morphological community. (2) There is no standardized, formalized method of morphological description. (3) There is a need for a rationale in the delineation of morphological traits (morphemes in descriptive morphology, characters/character states in causal morphology).
One suggestion to overcome the aspects of the linguistic problem is the use of ontologies (Vogt, 2009; Vogt et al., 2010; Burleigh et al., 2013; Deans et al., 2015). Ontologies are computer-readable, semantically structured knowledge bases used as representation of terms and their definitions (together called concepts or classes; Smith, 2004). These concepts usually classify instances, i.e. real individual objects, based on common properties. Furthermore, ontologies encompass relationships between concepts (Mabee et al., 2007; Dececchi et al., 2015). By their computer-readable syntax, ontologies enable machines to draw implications on their own. Furthermore, complex data mining becomes possible (Vogt, 2018).
Ontologies have found their way to life sciences and are popular in biomedical research and functional genetics. Biomedical phenomena like diseases as well as cytological and genetic terms are organized in elaborate ontologies like e.g. the Cell Ontology (Bard et al., 2005) or the Gene Ontology (Ashburner et al., 2000). From biomedicine via developmental genetics, ontologies have covered “classical” morphological fields of knowledge as well (Mabee et al., 2007). Since, a number of working groups have published anatomy ontologies (ontologies containing anatomical and morphological terminology) for their field of research; covering e.g. teleosts (Dahdul et al., 2010), hymenopterans (Yoder et al., 2010), spiders (Ramírez et al., 2007; Ramírez and Michalik, 2014, 2019), arthropod circulatory systems (Wirkner et al., 2017), and plants (Cooper et al., 2013). Besides anatomy ontologies themselves, also formal phenotypic ontologies have been designed to allow for descriptive statements to be formulated. Such formal phenotypic ontologies are among others the Biological Spatial Ontology (Dahdul et al., 2014), containing e.g. anatomical axes, planes and spatial relations, as well as the Phenotypic Quality Ontology (Gkoutos et al., 2004) containing all kinds of qualities to describe properties of anatomical entities (colors, elasticity, etc.).
Additional to defining concepts and linking them to terms, standardized and formalized methods of description, can be approached with ontologies (Deans et al., 2012; Balhoff et al., 2013; Göpel and Richter, 2016; Wirkner et al., 2017; Vogt, 2018). The ultimate goal is to develop an ontology environment in which morphological descriptions themselves can be done in a computer-readable, semantic way which then can also be used for tree inference or character mapping alternative to traditional character matrices (Vogt, 2018). Another advantage of an ontology-based descriptive morphology is the detachment of the descriptions from the evolutionary interpretations providing the “raw” morphological descriptions as observation statements which then are not conflated with explanatory accounts like statements on homology or biological role/adaptation (Göpel and Richter, 2016; Wirkner et al., 2017).
With new conceptual frameworks of morphology as a discipline as well as new tools and ways to describe morphology, central concepts such as homology face the challenge of whether they are still any meaningful. And as anyone familiar with the theoretical literature can attest, suggested answers to the homology question cover the entire spectrum between unifying concepts (Ramsey and Siebels Peterson, 2012; Ballego-Campos et al., 2023), specifying the meaning (Fitzhugh, 2006; Szucsich and Wirkner, 2007; Wagner, 2014; Fitzhugh, 2016; Göpel and Richter, 2023), diversifying the term towards different concepts (Vogt, 2017; Ochoterena et al., 2019), and omitting the term homology altogether (Cracraft, 2023).
As mentioned above, evolutionary morphology, just like any other science, can be subdivided into the descriptive level and the explanatory level (Figure 1). In descriptions, the researcher mainly deals with statements of fact about the status quo of his research object. Descriptive statements give answers to what-questions. The explanatory level of morphology, in contrast, seeks reasons for the described status quo (Bock, 1991, 2000; Sudhaus, 2007; Richter and Wirkner, 2014). Explanatory statements give answers to why-questions (Bock, 1991; Fitzhugh, 2008). These answers, especially in regard of evolutionary transformations, often refer to past events (Hennig, 1966; Bock, 2000; Sudhaus, 2007; Fitzhugh, 2008) which cannot be observed and thus remain hypotheses. The contrast between description and explanation is fundamental in science (Duhem, 1908). To delineate description from explanation regarding its terminology, the term “morpheme” (pl. “morphemes”) has been introduced to refer to the unit of morphological description (Richter and Wirkner, 2013; Göpel and Richter, 2016; Richter et al., 2020, 2014; Wirkner et al., 2017). Previously, terms mostly used in this context were “character” and “structure”, both of which have certain disadvantages. “Character” as term is used in morphology, taxonomy and phylogenetic systematics (Richter and Wirkner, 2014). However, the meaning or concept behind this term differs greatly between the three disciplines. In taxonomy, character refers to characteristic, observable features of populations or species in order to distinguish these from other such groups of organisms. Taxonomic characters are results of comparison and serve in differential diagnosis (Göpel and Richter, 2016). In phylogenetic systematics, characters are units of evolution (Houle, 2001; Richter and Wirkner, 2014; Wagner, 2014; Göpel and Richter, 2016). Although character also is used regularly in the sense of character state (which then refers to the species level), characters are seen as transformation series comprising the respective character states (Hennig, 1966; Grant and Kluge, 2004; Wirkner and Richter, 2010; Göpel and Richter, 2016). It is clear, however, that either one of the phylogenetic definitions of the term character refers to evolutionary units and thus units of explanation. By using the term character to refer to the units of both descriptive morphology and evolutionary interpretation, description and explanation might be conflated and the substantial intellectual and inferential processes that happen between description of morphology and reconstruction of character evolution can easily be overlooked. The term morpheme, borrowed from linguistic morphology (Hockett, 1961), has been introduced as a terminological means to facilitate delineation of descriptive from explanatory statements (Richter and Wirkner, 2014; Göpel and Richter, 2016; Wirkner et al., 2017). Using a separate term for descriptive units also reflects the biological reality that not every material part, i.e. morpheme, of an organism is also its own inheritable evolutionary entity, i.e. character (state) (Göpel and Richter, 2016, 2023; Richter and Wirkner, 2014). For instance, morphemes which develop as result of phenotypic plasticity (e.g. certain arteries in the vascular system of the parthenogenetic marbled crayfish; Scholz et al., 2021), can of course be described but are not inheritable themselves and therefore cannot be seen as evolutionary unit. In such cases, the evolutionary unit to which a character refers may rather be the ability to produce such morphemes via phenotypic plasticity than the artery itself.
In linguistics, “morpheme” traditionally refers to the smallest meaning-bearing object (Hockett, 1961), whereas in biological morphology, the term is suggested to describe any part of an organism that can be part of a morphological description (Richter and Wirkner, 2014). Without adopting the restriction of being the smallest entity, morphemes can be delineated at any given level of granularity from the macroscopic to subcellular scale. Thus, a morpheme can also be part of other morphemes at higher levels of organization, e.g. a phalanx is a morpheme and is part of the morpheme finger, which in turn is part of the morpheme hand. In plant morphology, for instance, the morpheme petal is part of the morpheme flower etc. In ontologies used to organize concepts and terms referring to organismal morphology/anatomy (often referred to as anatomy ontologies; Dahdul et al., 2010; Yoder et al., 2010; Mungall et al., 2012; Costa et al., 2013; Wirkner et al., 2017; Ramírez and Michalik, 2019), ontology concepts generally refer to classes of morphemes, i.e. morphemes of individual organisms are the instances that share defining features and thus are given the same term (e.g. “heart” or “brain”; Göpel and Richter, 2016; Wirkner et al., 2017). Due to the hierarchical nature of ontologies, classes of higher hierarchical level (superclasses) are classes of classes of morphemes and group morphemes with more general correspondences; e.g. the classes “arm” and “leg” may both belong to a superclass “limb”. This also shows the relatedness but incongruence of the concepts behind morphemes, ontology concepts and (phylogenetic) characters: To apply Sereno’s (Sereno, 2007) formalism, a morpheme can be either variable or locator in a character statement (Göpel and Richter, 2016; Göpel and Wirkner, 2018). While morphemes always are actually existing parts of an organism, characters (and character states) can refer to either the state of existence of such corresponding parts (absent/present or neomorphic characters; Sereno, 2007) or a certain property of these morphemes (multi-state or transformational characters; Sereno, 2007). In the former case, ontology concepts of anatomy ontologies are used to refer to the character, whereas in the latter case, ontological relations or ontology concepts referring to qualities (e.g. colors or shapes; instead of entities) refer to the actual character (Göpel and Richter, 2016; Wirkner et al., 2017; Göpel and Wirkner, 2018).
Ideally, delineation and description of a morpheme should avoid conflation with the non-observable explanatory level. Therefore, descriptive morphology, i.e. the description of morphemes, should not make any statements on (evolutionary/phylogenetic) homology (Richter and Wirkner, 2014; Wirkner et al., 2017; Göpel and Richter, 2023). Descriptions free of homology assumptions can serve as universally valid data basis for all following research questions independent of whether prevailing phylogenetic hypotheses and thus homology hypotheses change. In a second step then, comparative and causal morphology can derive and test homology hypotheses with the primary morphological data, i.e. the description of morphemes, unaltered. It should be noted that in practice, descriptive and comparative statements often are combined and even homology assumptions (be they explicitly stated or just implied). This might pose a problem if changes in e.g. phylogenetic interpretation render these aspects wrong as the mere descriptive statements would have to be extracted from the convoluted description-comparison statements later to be interpreted in a new phylogenetic context.
Based on the research program for evolutionary morphology by Richter and Wirkner (2013, 2014), the conceptual framework of morphology as science has been (further) developed; the relationships between the entities and processes in evolutionary morphology are summarized in Figure 1. In descriptive morphology, the individual organism is the primary object in focus and its morphemes are recognized, delineated and described (Figure 1; Richter and Wirkner, 2013, 2014). From the individual level, a process of generalization to the species level takes place (Sudhaus and Rehfeld, 1992; Frost and Kluge, 1994), leading to generalized descriptions of morphemes (Göpel and Richter, 2016). In comparative morphology, morphemes of different species are compared to provide the transition to the explanatory level (Figure 1). Studying the function of morphemes leads to the recognition of functional units (Schwenk, 2001) and the conceptualization of functional complexes (Figure 1) acting together (e.g. the two podomeres of a lobster claw). Knowledge from functional morphology can be exported to be applied in bionics which in turn can also enlighten functional understanding of biological morphology in the process of reverse-engineering (Nyakatura et al., 2019).
The comparison of generalized descriptions of morphemes on e.g. species level also allows for conceptualization of character states as inherited units of evolution (Wirkner and Richter, 2010; Richter and Wirkner, 2014; Göpel and Richter, 2016; Göpel and Wirkner, 2018). Homologization, the generation of hypotheses on homology between character states (Hennig, 1966; Richter, 2017; Göpel and Richter, 2023), assigns different character states to the same character based on a hypothesis of common origin (Hennig, 1966). It must be noted here, however, that the term “homologization” only makes sense if some evolutionary concept of homology is applied. In post-Darwinian evolutionary morphology this is generally the case (with some exceptions, see e.g. Vogt, 2017), although what “homology” describes in detail might be different (see below). The analysis of characters as evolutionary transformation series is the core of causal morphology (Figure 1). Different from Richter and Wirkner (Richter and Wirkner, 2013), the term causal morphology here refers to the analysis of ultimate causes (Mayr, 1961) only and albeit both explanatory, is distinct from functional morphology. The latter indeed is of explanatory value and is vital in adaptational interpretation of character state transformations but operates on the proximate level of explanation (Mayr, 1961; Bock, 2000). The evolutionary interpretation of character state transformation in causal morphology happens in form of reciprocal illumination/research cycles (Hennig, 1966; Kluge, 1997) by application of phylogenetic methods and reconsideration and reconceptualization of character (state) concepts shown to be in conflict with other such conceptualizations (Figure 1; Kluge, 1997).
In causal morphology, the character analysis involving the research cycles between character (re)conceptualization and inference of phylogenetic hypothesis provides hypotheses on What has happened in the past to causally account for Why individuals share certain traits in the present (Bock, 2000, 2007; Fitzhugh, 2008, 2016; Hennig, 1966). Functional morphology informs about the biological role and the adaptive value of functional complexes (Bock and von Wahlert, 1965; Bock, 2000) and thus provides interpretations as to Why some traits prevailed in the past while others did not. This integration of the functional/proximate and the historical/ultimate perspectives of explanation (Bock, 2000, 2007; Mayr, 1961) renders what is here called causal morphology the integrative core of evolutionary morphology.
This framework as such applies for every semaphoront, i.e. individuals in a certain state and during a certain (brief) period in their life-time (Hennig, 1966), that can be investigated and thus covers developmental aspects as well.
Against the background of the conceptual epistemic framework of evolutionary morphology presented in Figure 1, the general place of homology in morphology seems conveniently undisputed. This is, however, far from being true and what is understood by “homologization” (or homology assessment) differs greatly between authors. Even, for instance, within a phylogenetic consensus that homology is a relationship between character states substantial differences can be found. Since there is a vast and ever-growing body of literature on homology and reviews of homology concepts which try to unify or make their case which definition is to be preferred, I will not reheat what has been elaborated in great detail already. The question about what homology is should, however, be complemented by the question how we adequately treat homology as term and concept(s) so ubiquitous in our daily work. With no unification in sight, obviously no authority forcing a certain concept upon us, and maybe not even the desire/use for unification (Cracraft, 2023; Gouvêa and Brigandt, 2023; Minelli, 2023), I attempt to make a case for conceptual explicitness similar to what is advocated by the ontology community.
To help illustrate, two homology concepts which are both grounded in the same Hennigian view of characters as evolutionary transformation series shall serve as example (Figure 2): The first concept of homology strictly follows Hennig’s (1966) wording: “Different characters [states] that are to be regarded as transformation stages of the same original character [state] are generally called homologous” (Hennig, 1966, p.93). As Hennig defined, homology is thus to be interpreted as a relationship between different character states of the same character, thus emphasizing evolutionary sameness (or character identity sensu Wagner, 2014) over similarity (Richter, 2016; Göpel and Richter, 2016, 2023). In contrast, the second concept puts emphasis on equivalence of morphological patterns and thus a relationship between the manifestations of the same character state of a phylogenetic character (Szucsich and Wirkner, 2007; Szucsich et al., 2013). While both views are grounded in the same phylogenetic environment and both describe an evolutionary relationship of common origin of character states, their use of the term homology is substantially different and in fact mutually exclusive. A unification not only appears impossible but also would water down both concepts to be less meaningful. Both homology concepts explicitly leave out the respective other relationship which they then refer to as “character state identity” (for the identity relationship between the manifestations of the same character state; Richter, 2017) and “isoseriality” (for the relationship between different character states of the same transformation series; Szucsich et al., 2013), respectively (Figure 2). Neither approach is superior. In fact, both views apply the very same two central concepts: (1) a relationship between different character states of the same characters, (2) relationship between different manifestations of the same character state. They only differ as to for which of the two concepts the term “homology” is to be used.
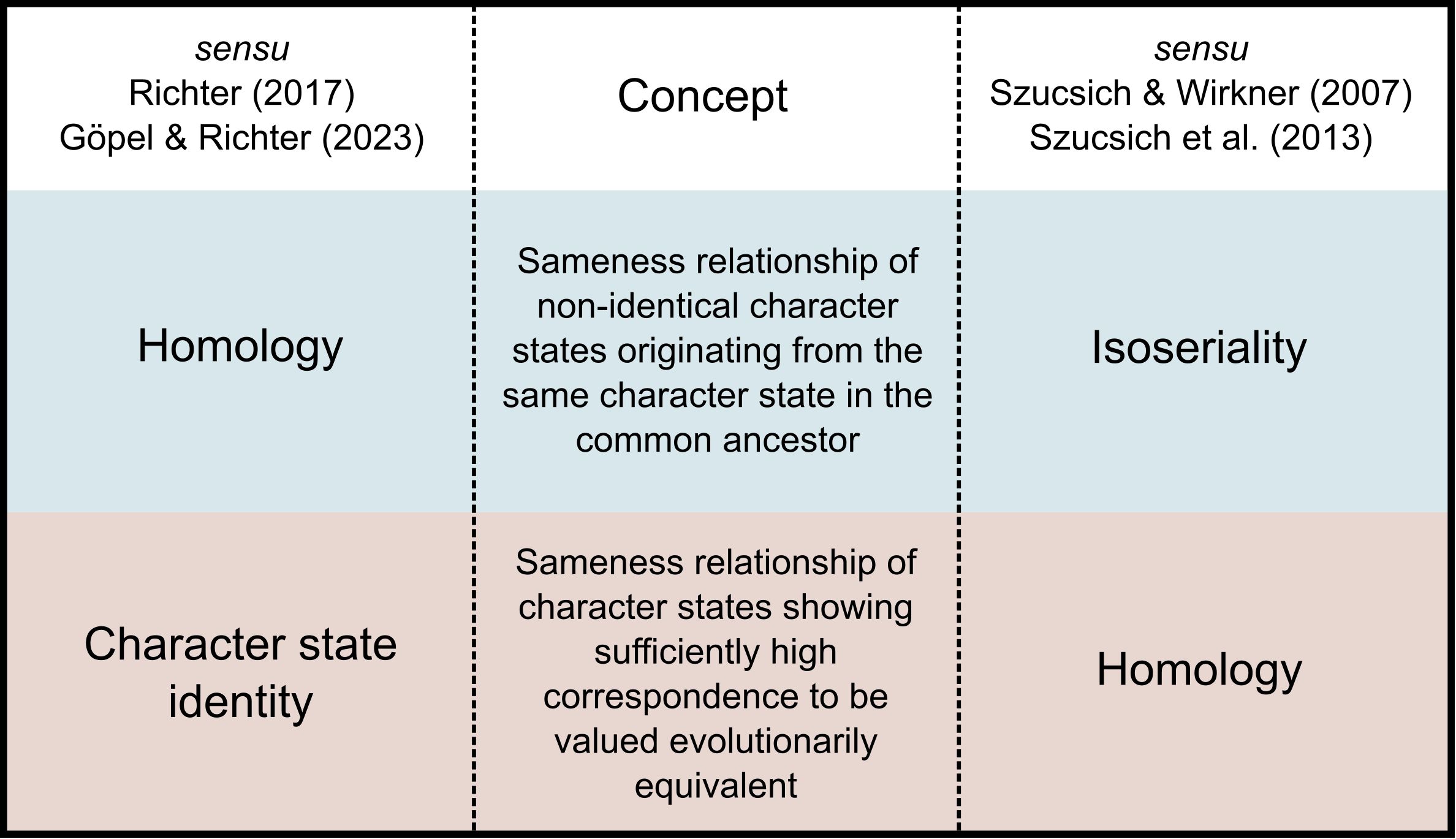
Figure 2 Conceptual consensus and terminological discord. In this example, the concepts applied by both groups of authors are the same, however, which concept is assigned the term “homology” differs. Note that the wording of both concepts is not directly taken from either reference to rather circumscribe the underlying meaning.
As becomes obvious from the usage of homology and similar terms in the literature, the linguistic problem of morphology (Vogt et al., 2010) does not only apply to terms used to refer to morphological entities but in fact to the very terms and concepts building the conceptual framework of morphology. Homology, a term so central and so frequently used, is a polyseme given as label to numerous different concepts (Brigandt, 2003; Ballego-Campos et al., 2023; Gouvêa and Brigandt, 2023); incongruence of homology statements is inevitable. Attempts of unification will unlikely gain universal acceptance. While authors often refer to the definitions by Owen (1847), there is no formal primate for older/the oldest definitions (Göpel and Richter, 2023). With the use of ontologies conceptual explicitness can help communicate what is meant when a certain term is used. As has been exemplified using the term “paramere” for a certain anatomical structure in Hymenoptera (Figure 1 in Yoder et al., 2010), previous connections of a term with a definition need to be taken into account. Just as with such terms and concepts used to address specific anatomical structures, conceptual and epistemic terms like homology and the different homology concepts could be incorporated into ontologies to have an unambiguous reference for each concept used. In fact, there is already a Homology Ontology (Roux and Robinson-Rechavi, 2010), which however, is rather general and hardly covers the vast amount of phylogenetic/evolutionary definitions of homology and their fine differences. Nonetheless, this would be a way to define concepts and add synonyms as “sensus” (Yoder et al., 2010; Wirkner et al., 2017) and allow for any publication to explicitly refer to a specific concept of homology. Furthermore, semantic, ontology-based morphology representations could analyze homology relations based on the concepts and independent of the term used and could even be extended to include other terms/concepts and relationships like synapomorphy (Hennig, 1966) or homogeny (Lankester, 1870). Conceptual explicitness regarding such central ideas acknowledges the inevitable plurality of homology concepts that has developed over time but enables intersubjective knowledge transfer and might therefore be helpful beyond the homology problem.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
TG: Writing – original draft, Investigation, Funding acquisition, Conceptualization.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work has been funded by third-party grants in the form of a PhD scholarship from the Konrad-Adenauer-Stiftung and a postdoc fellowship from the German Research Foundation (DFG; grant no. GO 3341/1-1). The author’s current position has been provided by the Alexander von Humboldt Foundation in the framework of the Alexander von Humboldt Professorship to J. Huisken, endowed by the German Federal Ministry of Education and Research. The author acknowledges support by the Open Access Publication Funds of the Göttingen University.
I am indebted to my academic mentors, Christian S. Wirkner and Stefan Richter, to spark my interest in both empirical as well as conceptual/theoretical evolutionary morphology. I cordially thank them for discussions and comments on earlier versions of this manuscript. Peter Michalik, Pedro Martinez and one anonymous reviewer provided valuable comments, which is gratefully acknowledged.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ashburner M., Ball C. A., Blake J. A., Botstein D., Butler H., Cherry J. M., et al. (2000). Gene ontology: tool for the unification of biology. Nat. Genet. 25, 25–29. doi: 10.1038/75556
Balhoff J. P., Mikó I., Yoder M. J., Mullins P. L., Deans A. R. (2013). A semantic model for species description applied to the ensign wasps (Hymenoptera: Evaniidae) of New Caledonia. Syst. Biol. 62, 639–659. doi: 10.1093/sysbio/syt028
Ballego-Campos I., Bonifácio S. K. V., Assis L. C. S. (2023). A unified view of homology. Cladistics 39, 398–417. doi: 10.1111/cla.12541
Bard J., Rhee S. Y., Ashburner M. (2005). An ontology for cell types. Genome Biol. 6, 1–5. doi: 10.1186/gb-2005-6-2-r21
Bock W. J. (1991). “Explanations in Konstruktionsmorphologie and evolutionary morphology,” in Constructional morphology and evolution. Eds. Schmidt-Kittler N., Vogel K. (Berlin, Heidelberg, Germany: Springer), 9–29.
Bock W. J. (2000). “Explanations in a historical science,” in Organisms, Genes and Evolution (Franz Steiner Verlag, Stuttgart), 33–42.
Bock W. J. (2007). Explanations in evolutionary theory. J. Zool. Syst. Evol. Res. 45, 89–103. doi: 10.1111/j.1439-0469.2007.00412.x
Bock W. J., von Wahlert G. (1965). Adaptation and the form-function complex. Evolution 19, 269–299. doi: 10.2307/2406439
Brigandt I. (2003). Homology in comparative, molecular, and evolutionary developmental biology: The radiation of a concept. J. Exp. Zool. Part B: Mol. Dev. Evol. 299B, 9–17. doi: 10.1002/jez.b.36
Burleigh J. G., Alphonse K., Alverson A. J., Bik H. M., Blank C., Cirranello A. L., et al. (2013). Next-generation phenomics for the tree of life. PloS Currents. doi: 10.1371/currents.tol.085c713acafc8711b2ff7010a4b03733
Cooper L., Walls R. L., Elser J., Gandolfo M. A., Stevenson D. W., Smith B., et al. (2013). The plant ontology as a tool for comparative plant anatomy and genomic analyses. Plant Cell Physiol. 54, e1–e1. doi: 10.1093/pcp/pcs163
Costa M., Reeve S., Grumbling G., Osumi-Sutherland D. (2013). The Drosophila anatomy ontology. J. Biomed. Semant. 4, 32. doi: 10.1186/2041-1480-4-32. Article 32.
Cracraft J. (2023). It is time to move on from homology in comparative biology. J. Morphol. 284, e21530. doi: 10.1002/jmor.21530
Dahdul W. M., Cui H., Mabee P. M., Mungall C. J., Osumi-Sutherland D., Walls R. L., et al. (2014). Nose to tail, roots to shoots: spatial descriptors for phenotypic diversity in the Biological Spatial Ontology. J. Biomed. Semant. 5, 1–13. doi: 10.1186/2041-1480-5-34
Dahdul W. M., Lundberg J. G., Midford P. E., Balhoff J. P., Lapp H., Vision T. J., et al. (2010). The teleost anatomy ontology: Anatomical representation for the genomics age. Syst. Biol. 59, 369–383. doi: 10.1093/sysbio/syq013
Darwin C. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. (London, United Kingdom, John Murray).
Deans A. R., Lewis S. E., Huala E., Anzaldo S. S., Ashburner M., Balhoff J. P., et al. (2015). Finding our way through phenotypes. PloS Biol. 13 (1), e1002033. doi: 10.1371/journal.pbio.1002033
Deans A. R., Yoder M. J., Balhoff J. P. (2012). Time to change how we describe biodiversity. Trends Ecol. Evol. 27, 78–84. doi: 10.1016/j.tree.2011.11.007
Dececchi T. A., Balhoff J. P., Lapp H., Mabee P. M. (2015). Toward synthesizing our knowledge of morphology: Using ontologies and machine reasoning to extract presence/absence evolutionary phenotypes across studies. Syst. Biol. 64, 936–952. doi: 10.1093/sysbio/syv031
Drack M., Betz O. (2022). A technomorphic conceptualisation of biological “constructions” and their evolution. Vertebrate Zool. 72, 839–855. doi: 10.3897/vz.72.e86968
Drack M., Limpinsel M., de Bruyn G., Nebelsick J. H., Betz O. (2018). Towards a theoretical clarification of biomimetics using conceptual tools from engineering design. Bioinspir. Biomim. 13, 016007. doi: 10.1088/1748-3190/aa967c
Duhem P. (1908). Ziel und Struktur der physikalischen Theorien (Leipzig, Germany: Johann Ambrosius Barth).
Fitzhugh K. (2006). The philosophical basis of character coding for the inference of phylogenetic hypotheses. Zoo. Scripta 35, 261–286. doi: 10.1111/j.1463-6409.2006.00229.x
Fitzhugh K. (2008). Abductive inference: implications for “Linnean” and “phylogenetic” approaches for representing biological systematization. Evol. Biol. 35, 52–82. doi: 10.1007/s11692-008-9015-x
Fitzhugh K. (2014). Character mapping and cladogram comparison versus the requirement of total evidence: Does it matter for polychaete systematics? Memoirs Museum Victoria 71, 67–78. doi: 10.24199/j.mmv.2014.71.07
Fitzhugh K. (2016). Dispelling five myths about hypothesis testing in biological systematics. Org. Diversity Evol. 16, 443–465. doi: 10.1007/s13127-016-0274-6
Frost D. R., Kluge A. G. (1994). A consideration of epistemology in systematic biology, with special reference to species. Cladistics 10, 259–294. doi: 10.1111/j.1096-0031.1994.tb00178.x
Ghiselin M. T. (1980). The failure of morphology to assimilate Darwinism. In Mayr E., Provine W. (Ed.), The Evolutionary Synthesis: Perspectives on the Un ification of Biology (Cambridge, MA and London, England: Harvard University Press), (pp. 180–193). doi: 10.4159/harvard.9780674865389.c27
Ghiselin M. T. (2006). The failure of morphology to contribute to the modern synthesis. Theory Biosci. 124, 309–316. doi: 10.1016/j.thbio.2005.11.001
Gkoutos G. V., Green E. C., Mallon A.-M., Hancock J. M., Davidson D. (2004). Using ontologies to describe mouse phenotypes. Genome Biol. 6, R8. doi: 10.1186/gb-2004-6-1-r8
Goethe J. W. (1817). Zur Naturwissenschaft überhaupt, besonders zur Morphologie (Stuttgart, Tübingen, Germany, Cotta).
Göpel T., Richter S. (2016). The word is not enough: on morphemes, characters and ontological concepts. Cladistics 32, 682–690. doi: 10.1111/cla.12145
Göpel T., Richter S. (2023). Homologues and homology and their related terms in phylogenetic systematics. Cladistics 39 (3), 240-248. doi: 10.1111/cla.12526
Göpel T., Wirkner C. S. (2018). Morphological description, character conceptualization and the reconstruction of ancestral states exemplified by the evolution of arthropod hearts. PloS One 13, e0201702. doi: 10.1371/journal.pone.0201702
Gould S. J., Lewontin R. C. (1979). The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. London Biol. Sci. 205, 581–598. doi: 10.1098/rspb.1979.0086
Gouvêa D. Y., Brigandt I. (2023). Against unifying homology concepts: Redirecting the debate. J. Morphol. 284, e21599. doi: 10.1002/jmor.21599
Grant T., Kluge A. G. (2004). Transformation series as an ideographic character concept. Cladistics 20, 23–31. doi: 10.1111/j.1096-0031.2004.00003.x
Haeckel E. (1866). Generelle Morphologie der Organismen: Allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von Charles Darwin reformierte Descendenz-Theorie. Band 1: Allgemeine Anatomie. Band 2: Allgemeine Entwicklungsgeschichte (Berlin, Germany, Verlag Georg Reimer ). doi: 10.1515/9783110848281
Hanser H., Ludolph A., Pickenhain L., Reichert H., Spitzer M. (2000). Lexikon der Neurowissenschaft: in vier Bänden (Heidelberg, Germany: Spektrum Akademischer Verlag).
Hennig W. (1966). Phylogenetic Systematics (Urbana, Chicago, Illinois: University of Illinois Press).
Hockett C. F. (1961). Linguistic elements and their relations. Language 37, 29–53. doi: 10.2307/411248
Houle D. (2001). “Characters as the units of evolutionary change,” in The Character Concept in Evolutionary Biology. Ed. Wagner G. P. (San Diego, California: Academic Press), 109–140.
Houle D., Govindaraju D. R., Omholt S. (2010). Phenomics: The next challenge. Nat. Rev. Genet. 11, 855–866. doi: 10.1038/nrg2897
Huisken J., Swoger J., Del Bene F., Wittbrodt J., Stelzer E. H. (2004). Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science 305, 1007–1009. doi: 10.1126/science.1100035
Kluge A. G. (1997). Sophisticated falsification and research cycles: Consequences for differential character weighting in phylogenetic systematics. Zoo. Scripta 26, 349–360. doi: 10.1111/j.1463-6409.1997.tb00424.x
Lankester E. R. (1870). II.—On the use of the term homology in modern zoology, and the distinction between homogenetic and homoplastic agreements. Ann. Magazine Natural History 6, 34–43.
Love A. C. (2003). Evolutionary morphology, innovation, and the synthesis of evolutionary and developmental biology. Biol. Philos. 18, 309–345. doi: 10.1023/A:1023940220348
Love A. C. (2006). Evolutionary morphology and Evo-devo: Hierarchy and novelty. Theory Biosci. 124, 317–333. doi: 10.1016/j.thbio.2005.11.006
Mabee P. M., Ashburner M., Cronk Q., Gkoutos G. V., Haendel M., Segerdell E., et al. (2007). Phenotype ontologies: the bridge between genomics and evolution. Trends Ecol. Evol. 22, 345–350. doi: 10.1016/j.tree.2007.03.013
Mayr E. (1961). Cause and effect in biology. Science 134, 1501–1506. doi: 10.1126/science.134.3489.1501
Metscher B. D. (2009). Micro CT for comparative morphology: Simple staining methods allow high-contrast 3D imaging of diverse non-mineralized animal tissues. BMC Physiol. 9, 11. doi: 10.1186/1472-6793-9-11. Article 11.
Minelli A. (2023). A refreshed approach to homology—Prioritizing epistemology over metaphysics. J. Morphol. 284, e21533. doi: 10.1002/jmor.21533
Mungall C. J., Torniai C., Gkoutos G. V., Lewis S. E., Haendel M. A. (2012). Uberon, an integrative multi-species anatomy ontology. Genome Biol. 13, R5. doi: 10.1186/gb-2012-13-1-r5. Article R5.
Nyakatura J. A., Melo K., Horvat T., Karakasiliotis K., Allen V. R., Andikfar A., et al. (2019). Reverse-engineering the locomotion of a stem amniote. Nature 565, 351–355. doi: 10.1038/s41586-018-0851-2
Ochoterena H., Vrijdaghs A., Smets E., Claßen-Bockhoff R. (2019). The search for common origin: homology revisited. Syst. Biol. 68, 767–780. doi: 10.1093/sysbio/syz013
Owen R. (1847). Report on the archetype and homologies of the vertebrate skeleton. Report of the British Association for the Advancement of Science. 169–340. doi: 10.5962/bhl.title.61890
Pigliucci M. (2001). “Characters and environments,” in The Character Concept in Evolutionary Biology. Ed. Wagner G. P. (San Diego, California: Academic Press), 363–388.
Ramírez M. J., Coddington J. A., Maddison W. P., Midford P. E., Prendini L., Miller J., et al. (2007). Linking of digital images to phylogenetic data matrices using a morphological ontology. Syst. Biol. 56, 283–294. doi: 10.1080/10635150701313848
Ramírez M. J., Michalik P. (2014). Calculating structural complexity in phylogenies using ancestral ontologies. Cladistics 30, 635–649. doi: 10.1111/cla.12075
Ramírez M. J., Michalik P. (2019). The spider anatomy ontology (SPD)—A versatile tool to link anatomy with cross-disciplinary data. Diversity 11, 202. doi: 10.3390/d11100202
Ramsey G., Siebels Peterson A. (2012). Sameness in Biology. Philos. Sci. 79, 255–275. doi: 10.1086/664744
Richter S. (2005). Homologies in phylogenetic analyses - Concept and tests. Theory Biosci. 124, 105–120. doi: 10.1016/j.thbio.2005.09.004
Richter S. (2016). Peter Ax’s views on homology—a comparison with Remane and Hennig. Peckiana 11, 67–75.
Richter S. (2017). Homology and synapomorphy-symplesiomorphy—neither synonymous nor equivalent but different perspectives on the same phenomenon. Cladistics 33, 540–544. doi: 10.1111/cla.12180
Richter S., Göpel T., Wirkner C. S. (2020). Das Morphem: Beschreibungseinheit der Evolutionären Morphologie. Naturwissenschaftliche Rundschau 73, 181–189.
Richter S., Wirkner C. S. (2013). Objekte der Morphologie. Morphologien. Bildwelten des Wissens 9.2 (Berlin, Germany, Akademie Verlag). 93–97.
Richter S., Wirkner C. S. (2014). A research program for evolutionary morphology. J. Zool. Syst. Evol. Res. 52, 338–350. doi: 10.1111/jzs.12061
Riedl R. (2000). Strukturen der Komplexität. Eine Morphologie des Erkennens und Erklärens (Berlin, Heidelberg, Germany: Springer). doi: 10.1007/978-3-642-56946-3
Roux J., Robinson-Rechavi M. (2010). An ontology to clarify homology-related concepts. Trends Genet. 26, 99–102. doi: 10.1016/j.tig.2009.12.012
Scholtz G. (2010). Deconstructing morphology. Acta Zoo. 91, 44–63. doi: 10.1111/j.1463-6395.2009.00424.x
Scholz S., Göpel T., Richter S., Wirkner C. S. (2021). High degree of non-genetic phenotypic variation in the vascular system of crayfish: a discussion of possible causes and implications. Zoomorphology 140, 317–329. doi: 10.1007/s00435-021-00536-2
Schwenk K. (2001). “Functional units and their evolution,” in The Character Concept in Evolutionary Biology. Ed. Wagner G. P. (San Diego, California: Academic Press), 165–198.
Seilacher A. (1970). Arbeitskonzept zur konstruktions-morphologie. Lethaia 3, 393–396. doi: 10.1111/j.1502-3931.1970.tb00830.x
Sereno P. C. (2007). Logical basis for morphological characters in phylogenetics. Cladistics 23, 565–587. doi: 10.1111/j.1096-0031.2007.00161.x
Shen Y., Yao G., Li Y., Tian X., Li S., Wang N., et al. (2023). RAD-seq data reveals robust phylogeny and morphological evolutionary history of the ornamentally important plant genus, Rhododendron. Hortic. Plant J. doi: 10.1016/j.hpj.2022.11.010
Smith B. (2004). Beyond concepts: Ontology as reality representation. Proceedings of FOIS 2004. International Conference on Formal Ontology and Information Systems, 73–84.
Spemann H. (1901). Entwickelungsphysiologische Studien am Triton-Ei. Arch. Entwickl. Mech. Org. 12, 224–264. doi: 10.1007/BF02152854
Sudhaus W. (2007). Die Notwendigkeit morphologischer Analysen zur Rekonstruktion der Stammesgeschichte. Species Phylogeny Evol. 1, 17–32.
Sudhaus W., Rehfeld K. (1992). Einführung in die Phylogenetik und Systematik (Stuttgart, Germany: Gustav Fischer).
Swofford D. L., Maddison W. P. (1987). Reconstructing ancestral character states under Wagner parsimony. Math. Biosci. 87, 199–229. doi: 10.1016/0025-5564(87)90074-5
Swofford D. L., Maddison W. P. (1992). Parsimony, character-state reconstructions, and evolutionary inferences. In Mayden R.L. (Ed.) Syst. Historical Ecol. North Am. Freshw. Fishes. (Stanford, California: Stanford University Press). pp. 186–223.
Szucsich N. U., Wirkner C. S. (2007). Homology: a synthetic concept of evolutionary robustness of patterns. Zoo. Scripta 36, 281–289. doi: 10.1111/j.1463-6409.2007.00275.x
Szucsich N. U., Wirkner C. S., Pass G. (2013). Deconstructing morphology: reply to Scholtz, (2010). Acta Zoo. 94, 215–217. doi: 10.1111/j.1463-6395.2011.00544.x
Vogt L. (2009). The future role of bio-ontologies for developing a general data standard in biology: Chance and challenge for zoo-morphology. Zoomorphology 128, 201–217. doi: 10.1007/s00435-008-0081-5
Vogt L. (2017). Assessing similarity: On homology, characters and the need for a semantic approach to non-evolutionary comparative homology. Cladistics 33, 513–539. doi: 10.1111/cla.12179
Vogt L. (2018). Towards a semantic approach to numerical tree inference in phylogenetics. Cladistics 34, 200–224. doi: 10.1111/cla.12195
Vogt L., Bartolomaeus T., Giribet G. (2010). The linguistic problem of morphology: Structure versus homology and the standardization of morphological data. Cladistics 26, 301–325. doi: 10.1111/j.1096-0031.2009.00286.x
Wagner G. P. (2014). Homology, Genes, and Evolutionary Innovation (Princeton, New Jersey, USA, Princeton University Press). doi: 10.23943/princeton/9780691156460.001.0001
Weber M., Mickoleit M., Huisken J. (2014). “Light sheet microscopy,” in Methods in cell biology, vol. Vol. 123. (San Diego, California, Academic Press), 193–215.
Wirkner C. S., Göpel T., Runge J., Keiler J., Klußmann-Fricke B. J., Huckstorf K., et al. (2017). The first organ-based ontology for arthropods (Ontology of Arthropod Circulatory Systems - OArCS) and its integration into a novel formalization scheme for morphological descriptions. Syst. Biol. 66, 754–768. doi: 10.1093/sysbio/syw108
Wirkner C. S., Richter S. (2004). Improvement of microanatomical research by combining corrosion casts with MicroCT and 3D reconstruction, exemplified in the circulatory organs of the woodlouse. Microsc. Res. Technique 64, 250–254. doi: 10.1002/jemt.20076
Wirkner C. S., Richter S. (2010). Evolutionary morphology of the circulatory system in Peracarida (Malacostraca; Crustacea). Cladistics 26, 143–167. doi: 10.1111/j.1096-0031.2009.00278.x
Keywords: evolutionary morphology, ontology, homology, concepts, terminology
Citation: Göpel T (2024) Synthesizing foundations in evolutionary morphology and a plea for conceptual explicitness regarding homology concepts. Front. Ecol. Evol. 12:1343346. doi: 10.3389/fevo.2024.1343346
Received: 23 November 2023; Accepted: 29 March 2024;
Published: 22 April 2024.
Edited by:
Roxana Yockteng, Colombian Corporation for Agricultural Research (AGROSAVIA), ColombiaReviewed by:
Peter Michalik, University of Greifswald, GermanyCopyright © 2024 Göpel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Torben Göpel, dG9yYmVuLmdvZXBlbEB1bmktZ29ldHRpbmdlbi5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.