
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 26 February 2024
Sec. Conservation and Restoration Ecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1339446
This article is part of the Research TopicOpportunities and Challenges for Wild Bee ConservationView all 13 articles
 James B. Dorey1,2*
James B. Dorey1,2* Olivia K. Davies1Karl N. Magnacca3Michael P. Schwarz1
Olivia K. Davies1Karl N. Magnacca3Michael P. Schwarz1 Amy-Marie Gilpin4
Amy-Marie Gilpin4 Thibault Ramage5Marika Tuiwawa6
Thibault Ramage5Marika Tuiwawa6 Scott V. C. Groom1Mark I. Stevens7,8
Scott V. C. Groom1Mark I. Stevens7,8 Ben A. Parslow7,8
Ben A. Parslow7,8Large parts of the Pacific were thought to host low bee diversity. In Fiji alone, our recent estimates of native bee diversity have rapidly increased by a factor of five (from 4 to >22). Here, we show how including sampling of the forest canopy has quickly uncovered a new radiation of Hylaeus (Hymenoptera: Colletidae) bees in Fiji. We also show that Hylaeus are more common across the Pacific than previously thought and solve one of Charles Michener’s mysteries by linking the previously enigmatic French Polynesian Hylaeus tuamotuensis to relatives in Fiji. We use systematic techniques to describe eight new Hylaeus species in Fiji (n = 6), French Polynesia (n = 1), and Micronesia (n = 1), and discuss impressive dispersal events by this genus. These clades also double the number of Hylaeus dispersals out of Australia from two to four. Our discovery highlights the severe impact of bee sampling methods on ecological interpretations and species discovery, specifically that canopy sampling is needed to correctly assess forest bee diversity even where there is a very long record of sampling. It further highlights the potential for forests to host higher-than-anticipated diversity and conservation value. This has broad methodological and regulatory impacts for land managers seeking to make choices about pollination services and diversity. The new species are Hylaeus (Euprosopoides) chuukensis Dorey, Davies, and Parslow; H. (Prosopisteron) albaeus Dorey, Davies, and Parslow; H. (P.) apertus Dorey, Davies, and Parslow; H. (P.) aureaviridis Dorey, Magnacca, and Parslow; H. (P.) breviflavus Magnacca; H. (P.) derectus Dorey, Davies, and Parslow; H. (P.) navai Dorey, Davies, and Parslow; and H. (P.) veli Dorey, Davies, and Parslow.
A key parameter in island biogeography theory is the distance separating remote islands from potential source populations (MacArthur and Wilson, 1967). Dispersal distances can act as filters, with successively more remote islands experiencing ever-more strict species filtering. However, additional filters also operate, such as taxon-specific capacities for dispersal, similarity in the ecology of source and sink regions, and opportunities to escape local enemies and pathogens (Patiño et al., 2017). Documenting how insular species diversity varies with distance from source regions facilitates our understanding of the relative roles of these filters in the assembly of insular biota.
Until recently, bee diversity in the Southwest Pacific (SWP) was regarded as depauperate, especially eastwards of New Caledonia (Perkins and Cheesman, 1928; Michener, 1979b). This aligns with predictions of decreasing richness over successively increasing distances, moving eastwards, of South Pacific islands from Sahul—Australia, Tasmania, New Guinea, and the surrounding islands (Groom and Schwarz, 2011). However, discerning regional patterns in diversity relies on sampling and taxonomic effort, and studies on bees from the South Pacific have been scarce until very recent times, often comprising privately published or unpublished museum records.
Recent studies have greatly increased the number and diversity of recorded bee species from multiple families in New Caledonia (Pauly and Munzinger, 2003; Barry et al., 2013; Pauly et al., 2015; Zakardjian et al., 2023), which might be expected given the geographical proximity of the archipelago to Sahul. At the same time, 10 years of intensive field work in Fiji has increased the number of described Lasioglossum (Homalictus) Cockerell, 1919 (family Halictidae) species from 4 (Perkins and Cheesman, 1928; Michener, 1979b) to 13 (Dorey et al., 2019) with more than 9 species waiting to be described (Dorey et al., 2020b; Naaz et al., 2022). These recent New Caledonian and Fijian studies show that our understanding of regional insular fauna can radically change perceived biodiversity patterns as sampling efforts increase.
Up until the last decade, only three bee families, Apidae, Megachilidae, and Halictidae, were reliably or recently recorded from the South West Pacific eastwards of Vanuatu (Pauly and Villemant, 2009; Groom and Schwarz, 2011; Naaz et al., 2022). Genetic studies have since shown that all of the apid species, and most of the megachilid species, in this region have been introduced via human agency (Davies et al., 2013; Groom et al., 2014, 2015). Remarkably, there is only one record of an endemic colletid bee in the South Pacific east of Vanuatu, namely, Hylaeus tuamotuensis Michener, 1965 from French Polynesia. Recorded in the 1930s, the species’ provenance was a mystery to Michener (1965), being over 4,000 km south of Hawaii and almost 6,000 km east of Australia. The large Hylaeus radiation in Hawaii is recently derived from a Palearctic ancestor, and not closely related to the Australian or South West Pacific taxa (Magnacca and Danforth, 2006). This dearth of colletid specimens is surprising given that Colletidae comprises the most abundant and diverse family of bees in Sahul.
The bee family Colletidae has a Gondwanan origin with centers of diversity in South America and Australia (Almeida et al., 2012). The colletid subfamily Hylaeinae has an Australian origin, and one molecular-based study (Kayaalp et al., 2013) suggests that a single dispersal event outside of Sahul led to an almost global distribution of the largest hylaeine genus, Hylaeus Fabricius, 1793 [764 species; Ascher and Pickering (2020)], with an additional dispersal to New Zealand. This is a remarkable case of geographical radiation, but limited taxon sampling in the Kayaalp et al. (2013) study does not allow us to understand how the various dispersal steps were taken, nor does it take into account what can be gleaned by examining distributional patterns.
Here, we provide the first rigorous evidence of endemic hylaeine bees in the South Pacific east of Vanuatu. We use morphological and mitochondrial DNA data to describe eight new species from Fiji (n = 6), French Polynesia (n = 1), and Micronesia (n = 1). Despite 10 years of intensive sampling on Fiji, this radiation of bees on Fiji was never described and was detected with uncertain provenance or forgotten in museum drawers. We show that canopy sampling techniques employed at only a few sites in Fiji rapidly revealed new species. We argue that the presence of colletid bees in the Pacific has been largely underestimated and recommend widespread canopy sampling regimes to correct this shortfall globally. We further highlight the massive and global potential for species discovery of bees in trees and the importance of this discovery for biogeographical understandings, forest management, and conservation.
Hylaeus specimens were caught between April 2016 and October 2019 by sweep netting flowers of several plant species. For all but one specimen (2016), samples were only collected using canopy nets (5–11 m) and sweeping off of red-flowering plants.
All contemporary samples were collected in August of 2017 near the summit of Mt Marau (Tahiti). Most specimens were collected on the flowers of the introduced Solanum nigrum L. (Solanaceae) and one was collected in a yellow pan trap that was set for a few hours nearby.
Samples were collected on a single day, 14 April 2014, via sweep netting of flowering vegetation. Samples were collected from the Weno (Xavier College Campus) and Fono Mu islands.
Tissue samples were taken from a single hind leg of each individual that was sequenced and then sent to the Centre for Biodiversity Genomics. Mitochondrial DNA was extracted and sequenced there using the SEQUEL platform, using the methods described by Hebert et al. (2018). The resulting sequences were then checked against the National Center for Biotechnology Information (NCBI) BLAST database to exclude non-target sequences. We retrieved 21 in-group Hylaeus sequences and three out-group Hyleoides Smith, 1853 or Meroglossa Smith, 1853 sequences. Outgroup specimens were identified by OKD using Houston (1975).
We employed the package bModelTest version 1.2.1 (Bouckaert and Drummond, 2017) in the BEAST2 version 2.6.6 (Bouckaert et al., 2019) package in order to determine the best partition schemes for our COI partitions, split into first (SYM/GTR+Γ+I), second (123324+Γ+I+x), and third (TN93+Γ+x) codon positions. Each codon position was assigned a relaxed log normal clock. We assigned three outgroup species—Hyleoides concinna (Fabricius, 1775), Meroglossa impressifrons (Smith, 1853), and M. itamuca (Cockerell, 1910)—and restricted them in the phylogeny according to Almeida and Danforth (2009). All tree priors were linked and assigned a Birth Death process in BEAUti version 2.6.6 (Bouckaert et al., 2019). We used CoupledMCMC version 1.0.2 (Müller and Bouckaert, 2020) with four heated chains, running for 100 million iterations, resampling every 20,000th iteration, and undertook four independent runs in BEAST2 to ensure convergence—as defined by an effective sample size of >200 in Tracer version 1.7 (Rambaut et al., 2018). The log and tree files from these four independent heated runs were combined using LogCombiner version 2.6.6 (Bouckaert et al., 2019). The consensus tree was created in TreeAnnotator version 2.6.6 (Bouckaert et al., 2019) and visualized using FigTree version 1.4.4 (Drummond, 2016).
To describe these bees, we used dissecting microscopes and entered data directly into Lucid version 4. We recorded measurements and converted these into ratios using Excel and then transcribed them into Lucid. Specimens were identified to subgenus using the keys made by Houston (1981) and Michener (2007) by JBD. We checked the availability of our names against the expanded BeeBDC bee taxonomy list (Dorey et al., 2023a; Dorey et al., 2023b) that was generated using Ascher and Pickering (2020). The key to the Micronesian Hylaeus was modified from Krombein (1950). Material is deposited in the Bernice Pauahi Bishop Museum (BPBM), Honolulu, Hawaii, USA, and the South Australian Museum (SAMA), Adelaide, South Australia, Australia.
Images of Fijian and French Polynesian specimens were taken with a Canon EOS 5DSR using a Canon MP-E 65mm f 2.8 1–5x macro (dorsal, lateral, and rear), Leica m205 C microscope with a Leica DFC 500 camera, Nikon 4x plan achromat microscopic, and Nikon 10x plan achromat microscopic lenses. Male sterna 7–8 and genitalia were imaged using a Nikon eclipse 50i with the same camera attached. Images were then stacked using Zerene Stacker (Littlefield, 2017) and then cleaned using Adobe Photoshop and Photoshop Lightroom. Images at the BPBM were taken using a Leica M165c microscope, a DMC5400 camera, and the Leica automontage system. These images were stacked using Helicon Focus (HeliconSoft, 2023).
Our initial Fijian Hylaeus (Prosopisteron) albaeus sp. nov. specimen was caught in April 2016 by sweep netting a single flowering Metrosideros sp. (Myrtaceae) tree at Rakiraki on the northern coast of Viti Levu (Figure 1A). That specimen was captured at a height of ~3 m. Further targeted low strata sampling in that area in the following 2 years did not yield more Hylaeus specimens, but cyclone Winston had removed much vegetation in the region. Our next successful collection of Hylaeus (Prosopisteron) specimens were during April of 2019 on the Fijian island of Taveuni and at 875 meters above sea level (m asl) under Des Voeux Peak, 22 and three samples of Hylaeus (Prosopisteron) apertus sp. nov. and Hylaeus (Prosopisteron) veli sp. nov., respectively, were collected on a single red-flowering Metrosideros collina var. collina (Forst.) A.Gray tree, where the lower-branches were sampled at heights of 4–7 m (Figures 1B, E). We obtained further samples of Hylaeus (Prosopisteron) in October 2019 from the telecom tower escarpments ~3 km west of Nadarivatu at 898–1,072 m asl, from sweeps of a few red-flowering mistletoes at a height of 3–7 m, Decaisnina forsteriana (Schult.) Barlow. Here, we collected eight and three specimens of Hylaeus (Prosopisteron) derectus sp. nov. and Hylaeus (Prosopisteron) navai sp. nov., respectively (Figures 1C, D). However, the Hylaeus were much less abundant than Lasioglossum. Additionally, a nearby small (2–3 m tall) flowering M. c. var. collina only hosted Lasioglossum.
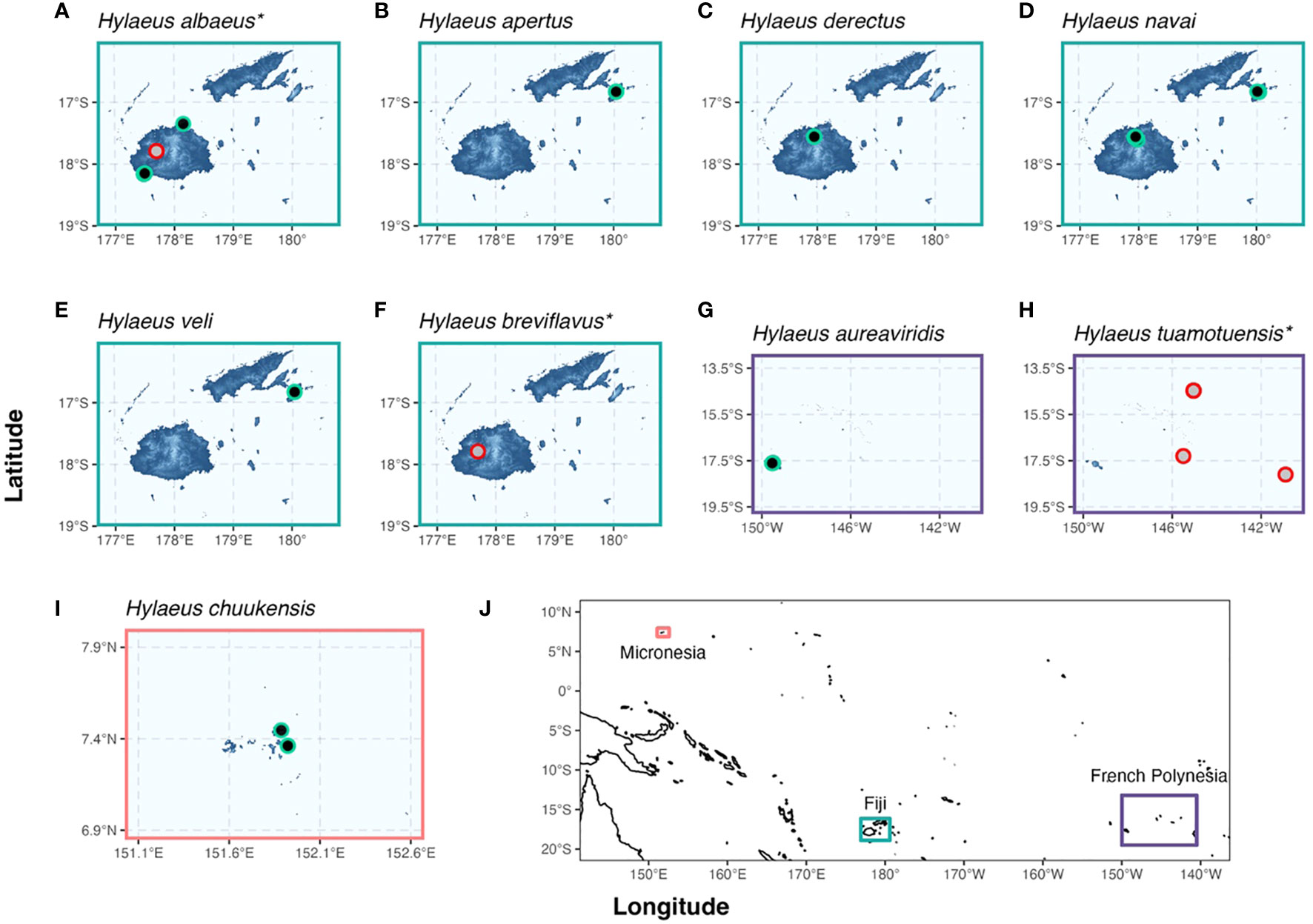
Figure 1 Collection locality maps of the examined Hylaeus species. The Fijian Hylaeus—Hylaeus albaeus sp. nov. (A), H. apertus sp. nov. (B), H. derectus sp. nov. (C), H. navai sp. nov. (D), H. veli sp. nov. (E), and H. breviflavus sp. nov. (F)—are surrounded by green, the French Polynesian species—H. aureaviridis sp. nov. (G) and H. tuamotuensis (H)—are surrounded by purple, and the Micronesian species—H. chuukensis sp. nov. (I)—is surrounded by salmon. The inset (J) shows the entire study area. Asterisks, and gray and red points (A, F, H) indicate that at least some coordinates are georeferenced from general localities.
Only one successful collection event was made in French Polynesia, returning six Hylaeus (Prosopisteron) aureaviridis sp. nov. from a Solanum nigrum and one from a yellow pan trap (Figure 1G). An additional Hylaeus observation has been made on iNaturalist by davidfl22 on 30 July 2023 (https://www.inaturalist.org/observations/176048893), which was also observed on Metrosideros collina.
We collected three specimens (two male and one female) of Hylaeus (Euprosopoides) chuukensis sp. nov. from Chuuk, Micronesia (Figure 1I). A male was collected from the Xavier College Campus (7.447, 151.887) and a further male and a female were collected from Fono Mu Islet (7.362, 151.923).
Our COI BEAST tree returned a well-supported phylogeny, where only a single node (between two Fijian spp.) had a posterior probability under 0.95 (Figure 2). Within-species variation was very low, and many nodes were quite deep relative to the base of the tree and the outgroup (Figure 2). While one clade contained only Fijian representatives, one was mixed with Fijian and French Polynesian representatives, and the Micronesian species formed its own clade (Figure 2).
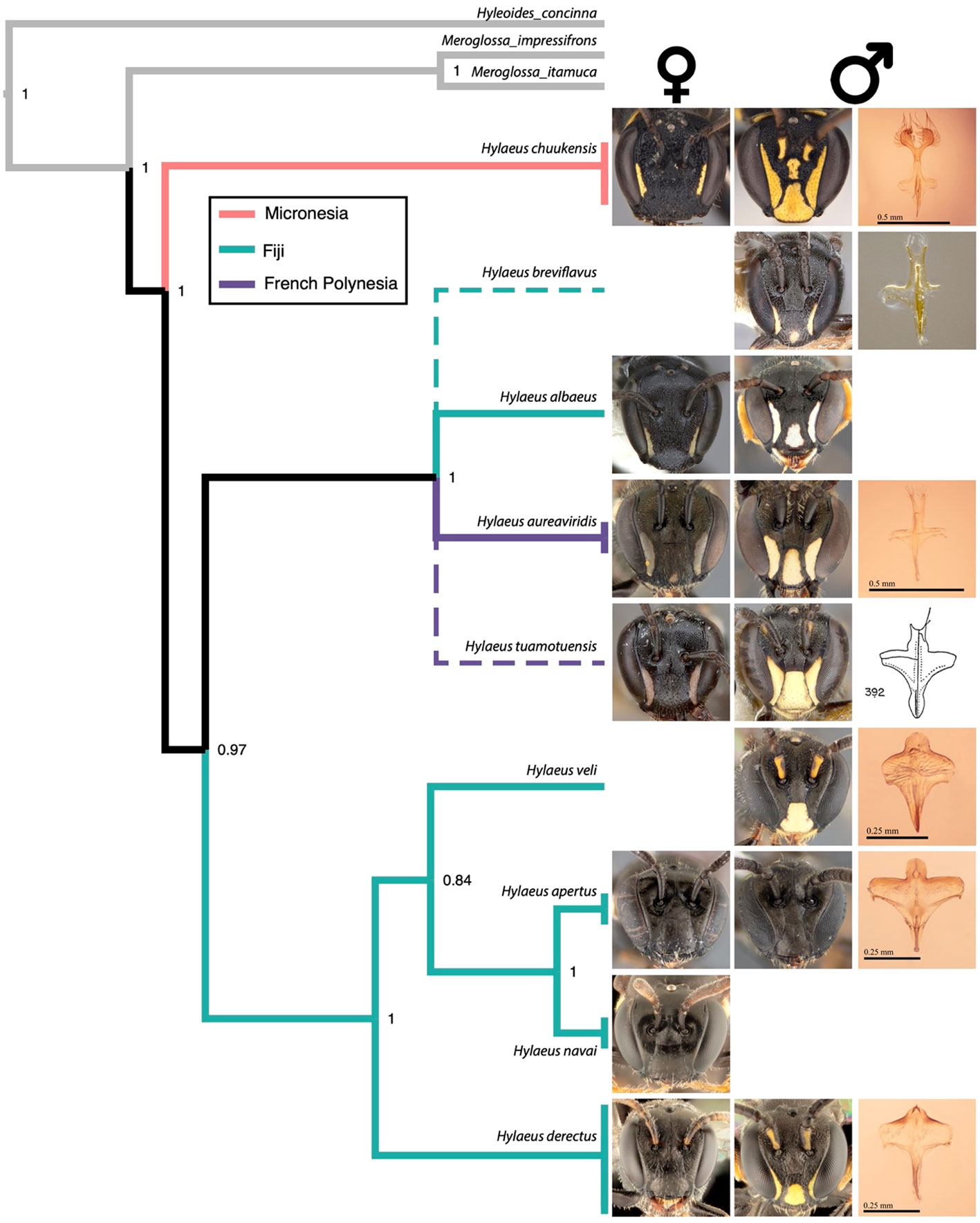
Figure 2 The phylogeny of the Micronesian (salmon), Fijian (green), and French Polynesian (purple) Hylaeus. The outgroup (gray) contains three Australian Hylaeinae from different genera, the Micronesian Hylaeus is of the subgenus Euprosopoides, while the remaining Hylaeus are of the subgenus Prosopisteron. Posterior supports are indicated at nodes, and dashed lines indicate inferred positioning from male genitalic characters. Images show the female faces (left), male face (middle), and male sternite 8 (right) for each species, where specimens exist. Note the bifurcation on posterior lobe (top) of sternite 8 for the middle Fiji–French Polynesia clade compared to the simple apex on the lower Fiji-only clade. Within-species genetic variation was essentially non-existent and so the terminals were flattened. The sternite 8 line drawing of H. tuamotuensis Michener, 1965 is reproduced from Michener (1965).
We provide brief species descriptions based on parts of the dichotomous key and full image plates (Figures 3–11). However, we also provide (i) a version of the manuscript with full-length descriptions embedded and an (ii) interactive Lucid key in our FigShare repository (https://doi.org/10.25451/flinders.24481231). There, we also share all data associated with the descriptions including (iii) all collection data (including georeferenced museum specimens), (iv) csv outputs from Lucid, (v) the R-code used to produce figures and manipulate data, (vi) BEAST2 run files and outputs, (vii) summary box plots of all measurements, and (viii) the GenBank submission file. While we do not provide written descriptions of male internal characters here (see FigShare), we do provide all images and note that they can be an excellent identification aid.
Family Colletidae Lepeletier de Saint Fargeau, 1841.
Subfamily Hylaeinae Viereck, 1916.
Genus Hylaeus Fabricius, 1793.
Subgenus Prosopisteron Cockerell, 1906.
1. Face with paraocular marks present … 2.
Paraocular area unmarked … 6.
2. Paraocular and clypeal marks contiguous, at least ventrally (Figure 9A); pronotal lobe yellow; mandible yellow; Tuamotu Islands … H. tuamotuensis Michener, 1965.
Paraocular and clypeal marks distinctly separated (Figures 3 and 5–7; pronotal lobe black; mandible variable … 3.

Figure 3 Hylaeus albaeus Dorey, Davies, and Parslow sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). (Scale bar, A-C, F-H = 1.00 mm; D, E = 0.25 mm.)

Figure 4 Hylaeus apertus Dorey, Davies, and Parslow sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). (Scale bar, A-C, F-H = 1.00 mm; D, E = 0.25 mm.)

Figure 5 Hylaeus aureaviridis Dorey, Magnacca, and Parslow sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). (Scale bar, A-C, F-H = 1.00 mm; D, E = 0.25 mm.)

Figure 6 Hylaeus breviflavus Magnacca sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), lateral head (D) (scale bar = 0.50 mm), genitalia (E), and sternite 7 (F) (scale bar = 0.25 mm).

Figure 7 Hylaeus derectus Dorey, Davies, and Parslow sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). (Scale bar, A–C, F, G, and H = 1.00 mm; D = 0.50 mm; E = 0.25 mm.)
3. Head and mesosoma tinged with submetallic green; clypeal mark large, extending nearly to posterior clypeal margin (Figures 5A–C); Tahiti … H. aureaviridis sp. nov.
Head and mesosoma mostly black; clypeal mark small … 4.
4. Scape marked with yellow; supraclypeal area reticulate, without distinct striae; posterior margin of clypeus straight; face in profile strongly convex ventrally; metasoma without hair bands (Figures 7A–C); Fiji … H. derectus sp. nov.
Scape dark brown to black; supraclypeal area striate; face in profile flatter, not strongly convex ventrally; T1 with prominent apicolateral bands of white setae … 5.
5. Facial markings white; pronotal lobe white; gena sparsely punctured (Figure 3); Fiji … H. albaeus sp. nov.
Facial markings yellow; pronotal lobe brown; gena closely punctured (Figure 6); Fiji … H. breviflavus sp. nov.
6. Face, mandible, and pronotum black; scutum with open punctures; anterior third of dorsal propodeum weakly rugose (Figures 4A–C); Fiji … H. apertus sp. nov.
Face with a large clypeal mark, mandible yellow, and pronotal lobe with a yellow spot; scutum with close to dense punctures; anterior half or more of dorsal propodeum rugose (Figures 10A–C); Fiji … H. veli sp. nov.
1. Mask with two pale patches … 2.
Mask with zero or three pale patches … 3.
2. Pronotal collar black with two lateral yellow patches; scutellum reticulate with very fine punctures; T2–4 with translucent yellow margins and no apical bands of setae; mandible with an anterior stripe along its length (Figures 9F, G); Tuamotu Islands … H. tuamotuensis Michener, 1965.
Pronotal collar black; scutellum shinning with conspicuous close punctures; T2–4 margin not clearly differentiated; T1–2 with posterolateral fascia of white setae; mandible all black (Figures 3F, G); Fiji … H. albaeus sp. nov. or H. breviflavus sp. nov.
3. Mask with three patches; head and mesosoma mostly black with submetallic green tinge; posterior margins of tergites with a distinct and broad paler margin; scutum and scutellum with dense and short erect hairs (Figures 5F, G); Tahiti … H. aureaviridis sp. nov.
Face entirely black; head and mesosoma mostly black, no metallic tinge; posterior margin of tergites usually black or if there is a paler margin it is narrow; scutum and scutellum with close-open partly erect hairs … 4.
4. Propodeum dorsal face reticulate and anterior half or more rugulose; T2 reticulate, clypeus posterior margin straight (Figures 7F–H); Fiji … H. derectus sp. nov.
Propodeum dorsal face reticulate and only anterior third rugulose; T2 shining medially, clypeus posterior margin concave … 5.
5. Medial groove on frons is partly poorly defined before meeting median ocellus; supraclypeal suture width:length ratio ≥ 13; gena reticulate but impunctate (Figures 8B–D); Fiji … H. navai sp. nov.
Medial groove on frons clearly meets median ocellus; supraclypeal suture width:length ratio ≤ 10; gena reticulate with sparse punctures (Figures 4F, G); Fiji … H. apertus sp. nov.
Materials examined. Holotype 1♂ Rakiraki hotel, Viti Levu, Fiji, −17.3603, 178.1537, 3 m asl, 2016/04/01, MP Schwarz, swept from Metrosideros collina var. collina, CFJRR_NH9, (BPBM: 18008). Paratypes 1♀1♂ Sigatoka Prov., Sigatoka Sand Dunes N.P., Viti Levu, Fiji, −18.16, 177.5, 100 m asl, 2002/12/13, M Irwin, E Schlinger, M Tokota‘a, Malaise trap, FJ-6B Malaise (BPBM: ♂ FBA 026760; ♀ FBA 026755); 1♂ 1♀ Sigatoka Prov., Sigatoka Sand Dunes N.P., Viti Levu, Fiji, −18.16, 177.5, 100 m asl, 2003/12/13, M Irwin, E Schlinger, M Tokota‘a, Malaise trap, FJ-6C Malaise (BPBM: ♂FBA 035899; ♀FBA 035880); 4♀ Sigatoka Sand Dunes N.P., malaise 1.1 km SSW of Volivoli Vlg., Viti Levu, Fiji, −18.1694, 177.4847, 55 m asl, 2003/11/15, E Schlinger, M Tokota‘a, Malaise trap, FJVL6b_M02_16 (BPBM:♀ FBA 063181, FBA 063184, FBA 063195, FBA 064760).
Male diagnosis. In combination, mask has three widely separated white marks; head and body are otherwise mostly black (not submetallic); supraclypeal area striate; and metasoma with distinct posterolateral setal bands on T1. Very similar to H. breviflavus sp. nov., distinguished by the white face marks and the scutum with moderately close puncture but with distinct interspaces, the surface reticulate and somewhat dull.
Male description. Head mask with three white patches, clypeal mark moderately small, not extending dorsally much over half length of clypeus, paraocular marks narrow and not or barely reaching ventral margin of antennal sockets; scape black; mandible mostly white, brown apex; gena reticulate, sparse punctures; frons densely punctured and rugulose and medial groove clearly meets median ocelli; supraclypeal area linear striae and small sparse punctures; paraocular area reticulate, sparse shallow punctures along eye margin or becoming punctured posterior of antennae; clypeal posterior margin convex; supraclypeal suture width: supraclypeal suture length (SSW : SSL) 7.39. Mesosoma pronotal collar black, yellow spot on pronotal lobe; scutum close-dense punctures and dense, very short, prostrate hairs, appearing velvety; metanotum with moderately dense, short, erect setae; lateral propodeum reticulate, close-open punctures; dorsal propodeum reticulate, anterior third weakly rugose. Metasoma T1 black, T2–4 black and posterior margin paler; T5–6 dark brown or posterior margin paler; T7 dark brown. T2 distinctly more convex in lateral view than other terga. T1 with a distinct fascia of white setae laterally along posterior margin; T2 with bands present but sparser, posterior terga lacking bands, with evenly spaced, moderately long erect setae.
Female diagnosis. Separated from other species here by the combination of two yellow face patches in the paraocular areas; pronotal collar lacking lateral yellow markings; and T1–2 with posterolateral seta bands. Not distinguishable from H. breviflavus sp. nov.
Female description. Head mask two patches consisting of narrow longitudinal stripes in paraocular area, not or barely reaching ventral margin of antennal sockets; scape and mandible black; gena closely punctured; frons close small punctures, smooth above, striate below and medial groove clearly meets median ocelli; supraclypeal area linear striae with small sparse punctures; paraocular area striate with dense, small punctures; clypeal posterior margin straight, SSW : SSL 1.6. Mesosoma pronotal collar black; scutum reticulate, small close punctures; lateral propodeum reticulate, strigate-rugulose dorso-posteriorly; dorsal propodeum reticulate, anterior third rugose. Metasoma black. T1 with a distinct fascia of white setae laterally along posterior margin; T2 with bands present but sparser, posterior terga lacking bands, with evenly spaced, moderately long erect setae.
Etymology. This species is named after the distinctive white spots on the face of at least the male where albaeus is Latin for white. Hence, they are the white-spotted Hylaeus.
Distribution. Known from Rakiraki (3 m asl), and Sigatoka (100 m asl) in Viti Levu, Fiji (Figure 1A).
Remarks. This species is very similar to H. breviflavus sp. nov., and it is possible they may prove to be conspecific. However, face mark coloration is usually a consistent character within species of Hylaeus. They are also widely separated physically and ecologically, with H. albaeus sp. nov. taken at the coast and lowlands, and H. breviflavus sp. nov. at moderate elevations. Females are associated based on morphological similarities, such as a strong band of hair on the lateral edges of T1, and the collection of the specimens together. Females associated with the males of H. albaeus sp. nov. and H. breviflavus sp. nov. cannot be distinguished. Further collections of both are required and we raise the need for further sequencing efforts of these two species and both sexes.
Materials examined. Holotype 1♂ Des Voeux track, Taveuni, Viti Levu, Fiji, −16.83622, −179.97303, 872 m asl, 2019/04/29 13:12, JB Dorey, swept from Metrosideros collina var. collina (BPBM: 18009). Paratypes 4♂ 6♀ Des Voeux track, Taveuni, Viti Levu, Fiji, −16.83622, −179.97303, 872 m asl, 2019/04/29 13:12–13:42, JB Dorey, swept from Metrosideros collina var. collina (SAMA: ♂ 32-035991, 32-035992, 32-035993, 32-035994; ♀ 32-035985, 32-035990, 32-035986, 32-035987, 32-035988, 32-035989). Other materials 4♂ 7♀ Des Voeux track, Taveuni, Viti Levu, Fiji, −16.83622, −179.97303, 872 m asl, 2019/04/29 13:12–13:42, JB Dorey, swept from Metrosideros collina var. collina (BPBM: ♂ 19FJ54, 19FJ60, 19FJ65, 19FJ71; ♀ 19FJ55, 19FJ57, 19FJ64, 19FJ66, 19FJ69, 19FJ73, 19FJ75).
Male diagnosis. Punctation of the scutum is sparser laterally than medially (more or less even in most other species); the metasoma is predominantly smooth and polished with few setae; and, excluding legs, it is entirely black.
Male description. Head mask none; scape and mandible black; gena reticulate, sparse punctures; frons reticulate, open small punctures and medial groove clearly meets median ocelli; supraclypeal area reticulate or reticulate-rugulose; paraocular area reticulate, sparse shallow punctures along eye margin; clypeal posterior margin straight or concave; SSW : SSL 7.5–38. Mesosoma pronotal collar black, some brown on pronotal lobe; scutum reticulate, large open-close punctures and close-open, short, partly erect hairs; metanotum with a posterior row of long simple hairs and short setae elsewhere; lateral propodeum reticulate and can have close-open punctures; dorsal propodeum reticulate, anterior third (sometimes weakly) rugose. Metasoma black or dark brown. T2–3 with very faint lateral hairbands, nearly hairless medially.
Female diagnosis. In combination, has no face patches and medial groove on frons clearly meets median ocellus. The pronotal lobes are marked with yellow.
Female description. Head mask none; scape black; mandibles black, apex sometimes brown; gena reticulate, sparse punctures; frons reticulate, reticulate-rugulose, open-sparse small punctures and medial groove clearly meets median ocelli; supraclypeal area reticulate; paraocular area reticulate, sparse shallow punctures along eye margin; clypeal posterior margin concave, SSW : SSL 6.5–9.75. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, small open punctures and close-open, short, partly erect hairs; metanotum with a sparse posterior row of long simple hairs, nearly hairless elsewhere; lateral propodeum reticulate and can have close-open punctures; dorsal propodeum reticulate, anterior third (sometimes weakly) rugose. Metasoma T1–3 black or dark brown; T2–3 posterior margins of can be paler; T4–7 black. T2–3 with very faint lateral hairbands, nearly hairless medially.
Etymology. This species is named for the lack of a mask for both sexes. The name is derived from the Latin apertus (open), and hence, they are the open-faced Hylaeus.
Distribution. Only known from Mt De Voeux (875 m asl), Taveuni, Fiji (Figure 1B).
Materials examined. Holotype 1♂ Near the summit of the Mt Marau, Tahiti, French Polynesia, −17.609041, −149.533164, 1,409 m asl, 2017/08/27, T Ramage, sweep net from Solanum nigrum or caught in nearby yellow pan trap, PFnG9Gm (BPBM: 18010). Paratypes 1♀ Tahiti, Near the summit of the Mt Marau, French Polynesia, −17.609041, −149.533164, 1,409 m asl, 2017/08/27, T Ramage, sweep net from Solanum nigrum or caught in nearby yellow pan trap, PFnG9Gf (SAMA: 32-036001); 1♀ Mt. Marau, Tahiti, French Polynesia, 1,300–1,400 m asl, 1984/08/28, G Paulay (BPBM); 1♀ Mt. Marau, Tahiti, French Polynesia, 1,409 m asl, 1977/06/29, PD Ashlock (BPBM); 1♂ 1♀ Fare Ata, Aorai Trail, Tahiti, French Polynesia, 1,800 m asl, 1977/03/09, WC Gagne (BPBM).
Male diagnosis. In combination, mask has three yellow patches (clypeus and paraocular areas); clypeal mark large; and head and body mostly black with faint tinges of submetallic green.
Male description. Head mask three yellow patches, clypeal mark extending nearly entire length and width, only narrow margins black, paraocular marks ventrally filling in area between eye and clypeus, dorsally extending nearly to ventral margin of antennal sockets; scape and mandible black; gena reticulate can have sparse punctures; frons densely punctured and rugulose and medial groove clearly meets median ocelli; supraclypeal area reticulate-rugulose or reticulate with distinct striae; paraocular area reticulate and sparse shallow punctures along eye margin or punctured posterior of antennae; clypeal posterior margin convex; SSW : SSL 0.95. Mesosoma pronotal collar black, yellow patch on pronotal lobe, scutum reticulate, small open or punctures, dense short erect hairs; metanotum with posterior row of branched hairs, can occur elsewhere; lateral propodeum reticulate or reticulate, strigate-rugulose dorso-posteriorly; dorsal propodeum reticulate, anterior third weakly rugose. Metasoma black or dark brown. T2–3 hairbands absent or very faint, sparse short hairs medially.
Female diagnosis. In combination, mask has three pale patches (paraocular stripes and a small spot at clypeal apex) and head and body are mostly black with submetallic green reflections.
Female description. Head mask three small pale patches, paraocular marks moderately narrow, extending dorsally about to ventral margin of antennal sockets, clypeus with a small ventral mark (probably sometimes absent); scape and mandible black; gena reticulate, sparse punctures; frons open to densely punctured and rugulose and medial groove clearly meets median ocelli; supraclypeal area reticulate-rugulose; paraocular area reticulate, sparse shallow punctures along eye margin or open small punctures; clypeal posterior margin straight or convex; SSW : SSL 2.75. Mesosoma pronotal collar, yellow patch on pronotal lobe; scutum open-close punctures and dense short erect hairs; metanotum with sparse, elongate branched setae, can occur in a posterior row; lateral propodeum very finely reticulate or reticulate; dorsal propodeum reticulate, anterior third weakly rugose. Metasoma dark brown, posterior margins paler. T2–3 hairbands absent or very faint, sparse short hairs medially.
Etymology. This species is named for both its yellow face patches and submetallic green sheen. Hence, we combine the Latin aurae (golden) and viridis (green) to become the golden-green Hylaeus.
Distribution. Known from Tahiti, near the summit of the Mt Marau and Fare Ata, Aorai Trail (1,300–1,800 m asl), French Polynesia (Figure 1G).
Remarks. The metallic coloration is faint but distinct, especially on the mesonotum. This character is unusual among Hylaeus, particularly extra-Australian species.
Materials examined. Holotype 1♂ Nausori Highlands, Viti Levu, Fiji, 500–600 m asl, 1970/10/01, NLH Krauss (BPBM: 18011). Paratypes 2♀ Nausori Highlands, Viti Levu, Fiji, 500–600 m asl, 1970/10/01, NLH Krauss (BPBM).
Male diagnosis. Has three small yellow patches on its face where the clypeal mark is present only as a small spot, and supraclypeal area striate. Nearly identical to H. albaeus sp. nov. but the mask is yellow rather than white and the scutum has dense punctures without distinct interspaces, the surface microcarinulate and somewhat shiny. The clypeal mark is also smaller than any of the specimens of H. albaeus sp. nov., but only one male specimen is known.
Male description Head mask three small yellow patches, clypeal mark small, less than one-third length of clypeus, paraocular marks narrow and not or barely reaching ventral margin of antennal sockets, scape black; mandibles black and brown; gena closely punctured; frons densely punctured and rugulose and medial groove poorly defined before median ocelli; supraclypeal area linear striae and small close punctures; paraocular area striate, becoming punctured dorsal of antennae; SSW : SSL 0.95. Mesosoma pronotal collar black; scutum shining, small close punctures, dense, very short, prostrate hairs, appearing velvety; metanotum with dense, short, appressed tomentose setae; lateral propodeum reticulate, strigate-rugulose dorso-posteriorly; dorsal propodeum reticulate, anterior third rugose. Metasoma T1 dark brown; T2–4 dark brown and posterior margin paler; T5–7 black. T1 with a distinct fascia of white setae laterally along posterior margin; T2 with bands present but sparser, posterior terga lacking bands, with evenly spaced, moderately long erect setae.
Female diagnosis and description. See H. albaeus sp. nov.
Etymology. This species is named for the small yellow patches on the male’s face, particularly the little clypeal patch. The name is from the Latin brevi (small) and flavus (yellow); hence, they are the little yellow-spotted Hylaeus.
Distribution. Only a locality provided as “Nausori Highlands, Viti Levu, Fiji” (Figure 1F).
Remarks. See the remarks for H. albaeus.
Materials examined. Holotype 1♂ Near Nadarivatu, Viti Levu, Fiji, −17.568, 177.953, 904 m asl, 2019/10/19, JB Dorey, DE18 (BPBM: 18012). Paratypes 1♂ 3♀ Near Nadarivatu, Viti Levu, Fiji, −17.5682, 177.9527, 898 m asl, 2019/10/18 14:45, JB Dorey, sweep net from Decaisnina forsteriana (BPBM: ♂ 19JDFJ4ii; ♀ 19JDFJ5a, 19JDFJ5b, 19JDFJ5i); 1♂ 2♀ Near Nadarivatu, Viti Levu, Fiji, −17.5682, 177.9527, 898 m asl, 2019/10/14 14:28, JB Dorey, sweep net from Decaisnina forsteriana (SAMA: ♂ 32-035997; ♀ 32-035995, 32-035996).
Male diagnosis. In combination, mask has three patches; posterior margin of clypeus is straight; clypeus in profile strongly convex; scape marked with yellow ventrally; and scutum deeply and strongly punctate.
Male description. Head mask three yellow patches, clypeal mark large, extending approximately 2/3 distance to dorsal margin of clypeus, paraocular marks narrow and pointed dorsally, not reaching antennal sockets; scape yellow anteriorly; mandibles black with paler apex; gena reticulate, sparse punctures; frons densely punctured and rugulose and medial groove clearly meets median ocelli; supraclypeal area reticulate or reticulate-rugulose; paraocular area reticulate, becoming punctured posterior of antennae; clypeal posterior margin straight; SSW : SSL −15. Mesosoma pronotal collar black, yellow spot on pronotal lobe; scutum reticulate, large, deep open-close punctures and dense short erect hairs or close-open, short, partly erect hairs; metanotum with short setae medially and can have long setae in a posterior row, can be branched; lateral propodeum reticulate, close-open punctures; dorsal propodeum anterior half or more rugose. Metasoma black or dark brown, posterior margins can be paler. T2–3 with faint lateral bands and nearly hairless medially.
Female diagnosis. In combination, mask has no patches (face is black); posterior margin of clypeus is straight; and clypeus in profile strongly convex.
Female description. Head mask none; scape brown, paler anteriorly; mandibles black and brown; gena reticulate, sparse punctures; frons reticulate-rugulose, sparse punctures, open small punctures and medial groove partly poorly defined before median ocelli; supraclypeal area reticulate-rugulose or reticulate, distinct striae; paraocular area reticulate, sparse shallow punctures along eye margin; clypeal posterior margin straight; SSW : SSL 46–76. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, close punctures, close-open, short, partly erect hairs; metanotum with short setae medially and can have long setae in a posterior row, can be branched; lateral propodeum reticulate, close-open punctures; dorsal propodeum anterior half or more rugose. Metasoma black or dark brown and posterior often margin paler. T2–3 with faint lateral bands and nearly hairless medially.
Etymology. This species is named for the straight posterior margin of the clypeus in both sexes from the Latin derectus (straight). Hence, they are the straight-faced Hylaeus.
Distribution. Only known from near Nadarivatu (898–904 m asl), Viti Levu, Fiji (Figure 1C).
Remarks. The strongly convex clypeus is somewhat reminiscent of H. crabronoides (Perkins, 1899) of Hawaii, but the two are not otherwise similar.

Figure 8 Hylaeus navai Dorey, Davies, and Parslow sp. Nov. Female lateral habitus (A), face (B), dorsal mesosoma (C), and lateral head (D) (scale bar = 0.50 mm).
Materials examined. Holotype 1♀ Near Nadarivatu, Viti Levu, Fiji, −17.5682, 177.9527, 898 m asl, 2019/10/18 15:35, JB Dorey, sweep net from Decaisnina forsteriana, 19JDFJ7i (BPBM: 18013). Paratypes 2♀ Mt Nadarivatu, Viti Levu, Fiji, −17.576245, 177.935436, 1,072 m asl, 2019/10/19, JB Dorey, sweep net from Decaisnina forsteriana (BPBM: ♀ DE146; SAMA: ♀ 32-035998); 1♀ Naitasiri Prov., Navai Village, Viti Levu, Fiji, −17.616, −177.983, 700 m asl, 2003/07/15, E. Schlinger, FJ-11A Malaise, (BPBM: FBA 029757); 1♀ Cakaudrove Prov., Soqulu House in Soqulu Estate, Viti Levu, Fiji, −16.833, −180.000, 140 m asl, 2002/11/21, E. Schlinger, Malaise 1, (BPBM: FBA 099896); 1♀ Cakaudrove Prov., 5.3 km SE Tavuki Vlg. Mt. Devo, Viti Levu, Fiji, −16.841, −179.968, 1,064 m asl, 2002/11/17, Schlinger, M Tokota‘a, Malaise 3, (BPBM: FBA 134592).
Female diagnosis. In combination, has no face patches; propodeum dorsal face reticulate and only anterior third rugulose; and medial groove on frons is partly poorly defined before meeting the median ocellus. The head is short and broad (wider than long), and the body overall has weak reticulate microsculpture and open punctation, thus appearing quite shiny.
Female description. Head mask none; scape and mandibles black; gena reticulate; frons reticulate, open small punctures and medial groove partly poorly defined before median ocelli; supraclypeal area reticulate; paraocular area reticulate, sparse shallow punctures along eye margin; clypeal posterior margin concave; SSW : SSL 13–26. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, small open punctures; scutum close-open, short, partly erect hairs, few much longer erect hairs or so few hairs as to appear hairless; metanotum with a posterior row of long simple setae, almost hairless medially; lateral propodeum reticulate; dorsal propodeum reticulate, anterior third weakly rugose. Mesosoma black. T2–3 with faint lateral bands and nearly hairless medially.
Etymology. This species is named in recognition of the people of Navai village who have made a large contribution to the taxonomy and understanding of the Fijian bee fauna. Meli Naiqama has acted many times as our guide and helped collect bees, his family (especially his parents, Esira and Paulini Senimasi) has hosted and fed us over many trips, and the whole village has always made us feel very welcome! Hence, they are Navai’s Hylaeus.
Distribution. Known from near Navai Village (700 m asl) and Nadarivatu (898–1,072 m asl) on Viti Levu, Fiji and Soqulu House (140 m asl) and Mt De Voeux (1,064 m asl), Taveuni, Fiji (Figure 1D).

Figure 9 Hylaeus tuamotuensis Michener, 1965. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). [Scale bar, A-C, F-H = 1.00 mm; D, E are reproduced from Michener (1965) with no scale.]
Hylaeus tuamotuensis Michener, 1965: 123.
Materials examined. Holotype 1♂ Tukuhora, Anaa I., Tuamotu Islands, −17.3, −145.5 [georeferenced], 1934/5/13, EC Zimmerman (BPBM). Other materials 10♂ 1♀ Teavaroa to Opakari, Takaroa Atoll, French Polynesia, −14.47, −145.04 [georeferenced], 0–2 m asl, 1984/06/29, G.A. Samuelson, mostly on Euphorbia atoto (BPBM); 1♀ Boring Bay, Hao Island, French Polynesia, −18.1, −140.9 [georeferenced], 1934/06/19, EC Zimmerman (BPBM).
Male diagnosis. In combination, mask has three yellow patches (clypeus and supraclypeal areas); head and body are otherwise mostly black (not submetallic); and the supraclypeal area has striae.
Male description. Head mask three yellow patches, clypeus completely yellow and paraocular areas mostly yellow, terminating around anterior margin of antennal sockets; scape black, lighter underside; mandible yellow, brown apex; gena reticulate; frons small close punctures, almost touching but not rugulose and medial groove clearly meets median ocelli; supraclypeal area faint sinuate striae; paraocular area striate, becoming punctured dorsal of antennae; clypeal posterior margin convex. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, small open punctures, dense short erect hairs; metanotum posterior row of long branched hairs, some hairs medially; lateral propodeum reticulate, strigate-rugulose dorsoventrally; dorsal propodeum reticulate, anterior third weakly rugose. Mesosoma black. T2–3 lateral hairbands absent or very faint.
Female diagnosis. In combination, mask has two patches; metasoma is mostly black; and the pronotal colour has two yellow patches.
Female description Head mask two large pale patches almost reaching anterior margin of antennal socket; scape black, lighter underside; mandible black with yellow streak; gena reticulate; frons small close punctures, almost touching but not rugulose and medial groove clearly meets median ocelli; supraclypeal area faint sinuate striae, paraocular area striate, becoming punctured dorsal of antennae; clypeal posterior margin convex. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, small open punctures, dense short erect hairs, and scattered additional setae about twice as long; metanotum posterior row of long branched hairs, some hairs medially; propodeum reticulate, anterior third weakly rugose. Metasoma T1–6 black and posterior margin paler; T7 black. T2–3 lateral hairbands absent or very faint.
Distribution. Known from Tukuhora, Anaa Island; Takaroa Island; and Boring Bay, Hao Island, Tuamotu Islands, French Polynesia (Figure 1H).

Figure 10 Hylaeus veli Dorey, Davies, and Parslow sp. Nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), lateral head (D) (scale bar = 0.50 mm), genitalia (E), and sternite 7 (F) (scale bar = 0.25 mm).
Materials examined. Holotype 1♂ Des Voeux track, Taveuni, Fiji, −16.83622, −179.97303, 875 m asl, 2019/04/19 13:37, MI Stevens, MC Elmer, sweep net from Metrosideros collina var. collina (BPBM: 18014). Paratypes 2♂Des Voeux track, Taveuni, Fiji, −16.83622, −179.97303, 875 m asl, 2019/04/19 13:37, JB Dorey, sweep net from Metrosideros collina var. collina (SAMA: 32-035999; BPBM: 19FJ72).
Male diagnosis. Mask has one yellow patch (on clypeus); scutum and scutellum reticulate-rugulose; and mandible and anterior of scape almost all yellow.
Male description. Head mask one yellow patch, covering almost entire clypeus, only very narrow lateral and dorsal margins dark; scape yellow on ventral and medial surfaces; mandible yellow; gena reticulate, sometime with sparse punctures; frons reticulate, large close punctures and medial groove clearly meets median ocelli; supraclypeal area reticulate; paraocular area reticulate, sparse shallow punctures along eye margin or becoming punctured posterior of antennae; clypeal posterior margin straight or concave; SSW : SSL 8–34. Mesosoma pronotal collar black, yellow or brown spot on pronotal lobe; scutum reticulate-rugulose with close-dense punctures and dense short erect hairs or close-open, short, partly erect hairs; metanotum with a posterior row of long simple setae and moderate simple setae elsewhere; lateral propodeum reticulate or reticulate dorso-anteriorly, rugose ventro-posteriorly; dorsal propodeum anterior half or more rugose. Metasoma black or dark brown, posterior margins can be paler. T2–3 with faint lateral hair bands, nearly hairless medially.
Etymology. This species is named for the veli of Fijian folklore who are powerful little people associated with forests. Accounts of the veli are varied and they were often seen in a positive light, but they could also be dangerous, for example, if you chopped down their favorite trees (Tomlinson, 2016). Hence, the name is meant to invoke a sense of responsibility for protecting these new forest-specialist species and their trees. Hence, they are veli’s Hylaeus.
Distribution. Only known from De Voeux Peak (875 m asl), Taveuni, Fiji (Figure 1E).
Family Colletidae Lepeletier de Saint Fargeau, 1841.
Subfamily Hylaeinae Viereck, 1916.
Genus Hylaeus Fabricius, 1793.
Subgenus Euprosopoides Michener, 1965.
1. Abdomen usually ferruginous in part or entirely; yellow markings more extensive than in other species, pronotal band complete, space between lateral margin of clypeus and inner eye margin entirely yellow, females with yellow spots on clypeus, scutum anterolaterally and axillae; almost all the punctures on scutum uniformly subcontiguous; Carolines (Yap) … H. yapensis (Yasumatsu, 1942).
Abdomen never ferruginous, occasionally dark brown on basal segments, but usually black with obscure metallic green or blue reflections; yellow markings much less extensive, pronotal band always interrupted in middle, space between lateral margin of clypeus and inner eye margin not entirely yellow (needs confirmation in H. rotensis), females without yellow markings on clypeus, scutum, or axillae; punctures of thorax separated (needs confirmation in H. rotensis) … 2.
2. Scutellum and metanotum entirely black; female unknown; Marianas (Rota) … H. rotensis (Yasumatsu, 1942).
Scutellum and metanotum with large yellow marks … 3.
3. Metasoma metallic blue or violet in both sexes; yellow mark on scutellum covering only the posterior third (male) or half (female); supraclypeal mark present in males; some punctation of thorax, at least proximal to parapsidal lines, separated by more than the width of a puncture (especially in females); Chuuk archipelago … H. chuukensis sp. nov.
Metasoma black, sometimes brownish anteriorly; yellow mark on scutellum covering posterior 60%–100%; supraclypeal mark present or absent in males; punctation of thorax separated by a little less than or as much as the width of a puncture … 4.
4. Male: upper margin of yellow mark on clypeus irregular, supraclypeal mark present; yellow mark on scutellum larger than on metanotum, the anterior margin of the mark on the former straight. Female: pronotal collar not produced above level of anterior part of scutum, scarcely notched in middle; yellow mark on scutellum covering the entire disk except narrow anterior margin. Marianas (Guam) … H. guamensis (Cockerell, 1914).
Male: upper margin of yellow mark on clypeus rounded, the supraclypeal mark absent ()?; yellow mark on metanotum as large as that on scutellum, the anterior margin of the mark on the latter with a median notch. Female: pronotal collar strongly produced above level of anterior part of scutum and with a broad, deep notch in middle; yellow mark on scutellum less extensive, covering only the posterior half or two-thirds of disk; anterior metasomal segments sometimes brownish; Carolines (Palaus) … H. hirticaudus Cockerell, 1939.

Figure 11 Hylaeus chuukensis Dorey, Davies, and Parslow sp. nov. Male lateral habitus (A), face (B), dorsal mesosoma (C), genitalia (D), and sternite 7 (E). Female lateral habitus (F), face (G), and dorsal mesosoma (H). (Scale bar, A–C, F–H = 1.00 mm; D, E = 0.50 mm.)
Materials examined. Holotype 1♂ Chuuk, Weno, Xavier College Campus, Micronesia, 7.447, 151.887, 39 m asl, 2014/04/14, SVC Groom, (BPBM: 18007). Paratypes 1♂1♀ Chuuk, Fono Mu Islet, Micronesia, 7.362, 151.923, 0 m asl, 2014/04/14, SVC Groom (SAMA: ♂32-38374; ♀ 32-38373).
Male diagnosis. Abdomen strongly metallic blue or violet; pronotal band incomplete; punctures on scutum dense, but interspaces often larger than punctures proximal to parapsidal lines; yellow patch on scutellum much wider than on metanotum and filling the posterior third. May be the only species with yellow patches anteriorly on the scapes. Falls out of Houston (1981)’s Australian Euprosopoides key at couplet #4 because it has a metallic blue abdomen and the first recurrent vein of forewing lacks a stub-like branch.
Male description. Head mask four yellow patches, clypeus yellow except narrow lateral and dorsal margins (touching paraocular marks ventrally), supraclypeal area with a separate mark, and paraocular areas broadly yellow, extending dorsal of antennal sockets, dorsally narrowing to a point; scape yellow anteriorly; mandible black, apex brown; gena closely punctured; frons large close punctures and medial groove partly poorly defined before median ocelli; supraclypeal area large open punctures, very faint striae; paraocular area large close to sparse punctures; clypeal posterior margin straight; SSW : SSL 33. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum reticulate, open-close punctures and close-open, long, mostly erect hairs; metanotum posteriolateral row of long and branched setae, shorter branched setae elsewhere; lateral propodeum very finely reticulate; dorsal propodeum coarsely areolate, distinct posterior carina. Metasoma black or submetallic blue. T2–3 hairbands absent but with short setae covering.
Female diagnosis. Abdomen strongly metallic blue or violet; pronotal band incomplete; punctures on scutum dense, but interspaces often larger than punctures proximal to parapsidal lines and medially; yellow patch on scutellum slightly wider than on metanotum and filling less than the posterior half; without clypeal marking; pronotal collar produced above the anterior part of scutum and deeply notched medially. Falls out of Houston (1981)’s Australian Euprosopoides key at couplet #4 because it has a metallic blue abdomen and the first recurrent vein of forewing lacks a stub-like branch.
Female description. Head mask two small linear yellow patches not reaching anterior margin of paraocular area but extending posterior to the anterior margin of the antennal sockets; scape and mandible black; gena shining with sparse to close punctures; frons large close punctures and medial groove clearly meets median ocelli; supraclypeal area large close punctures; paraocular area large close to sparse punctures; clypeal posterior margin indistinct or obscured. Mesosoma pronotal collar black, yellow patch on pronotal lobe; scutum large open-close punctures, open medially and close-open, short, partly erect hairs; metanotum posteriolateral row of long and branched setae, shorter branched setae elsewhere; lateral propodeum very finely reticulate; dorsal propodeum coarsely areolate, distinct posterior carina. Metasoma black or submetallic blue. T2–3 hairbands absent or very faint but with short setae covering.
Etymology. Of the 12 already described species in Hylaeus (Euprosopoides), 8 are Australian and 4 are found in the Pacific. The four Pacific species are H. (E.) guamensis (Cockerell, 1914) from Guam, Northern Mariana Islands; H. (E.) rotensis (Yasumatsu, 1939) from Rota, Northern Mariana Islands; H. (E.) hirticaudus Cockerell, 1939 from Palau; and H. (E.) yapensis (Yasumatsu, 1942) from Yap in Micronesia (Ascher and Pickering, 2020). Hence, we follow this tradition and the name chuukensis is from the island group where this species occurs in Chuuk, Micronesia. Hence, they are Chuuk’s Hylaeus.
Distribution. The Chuuk archipelago (0–39 m asl), Micronesia (Figure 1I).
By identifying and describing eight new species, we advance our understanding of this fascinating and speciose bee genus. We show that Hylaeus is more numerous in the Pacific than previously suspected and that more work in this region is urgently needed.
We show that Micronesia has at least one additional species, found in Chuuk (Hylaeus chuukensis sp. nov.), bringing the regional total to five Hylaeus (Euprosopoides) species, with no two from the same island group. With the use of canopy nets in 2019 in Fiji, we were able to collect four additional species from red-flowering plants, but only at heights of >3 m (even if known food plants occurred below this height). We describe these four species that form a monophyletic Fijian clade; two on the main island of Viti Levu (H. derectus sp. nov. and H. navai sp. nov.) and three on the island of Taveuni (H. apertus sp. nov., H. navai sp. nov., and H. veli sp. nov.). On each island, we essentially collected these species on a single collection event and locality. However, H. navai sp. nov. was also collected using Malaise traps in 2002 and 2003. According to our phylogeny and bee dissections (especially the bifurcation of S8; Figure 2), one of our clades has two representatives in Fiji (H. albaeus sp. nov. and H. breviflavus sp. nov.), and two in French Polynesia (H. aureaviridis sp. nov. and H. tuamotuensis; Figure 2).
While the Micronesian and French Polynesian specimens were collected using standard methods, we only actively collected one Fijian Hylaeus specimen without a canopy net (H. albaeus sp. nov.). We could not relocate H. albaeus sp. nov. after its initial collection despite targeted, but standard, efforts over the following 2 years. However, this species has been collected in Malaise traps on five other occasions at two localities (~87 Malaise trap days). All of these collections were made in relatively cleared regions on the dry (western) side of Viti Levu and in regions that have been heavily sampled for Lasioglossum Curtis, 1833 in the past 10 years. We suggest that these Malaise collections, and possibly our 2016 collection, were of bees that were moving between stands of trees. Taken together, this indicates an extreme affinity of this Hylaeus clade for canopy life with only rare vagrants being collected at lower floral resources; despite a decade of targeted bee sampling in Fiji (Naaz et al., 2022).
Having two species in each of Fiji and French Polynesia might indicate natural long-distance dispersals between the two archipelagos, almost certainly via the intervening archipelagos (Figures 1, 2). This contrasts with assisted movement by the Austronesian peoples who used large ships for east–west return voyages that carried dozens of people, livestock, and plants for trading and settling of new territories (Thomas, 2021). This supports the idea that Hylaeus bees can be successful very-long-distance dispersers (many hundreds of kilometers at a time) and that Michener’s mystery, H. tuamotuensis, most likely island hopped from Fiji (~3,000 km). The massive dispersal of this Hylaeus clade in the Pacific indicates that it is likely very widespread and speciose throughout the whole region.
Our findings also contrast with those by Poulsen and Rasmussen (2020) who suggested that, compared to mainland species, most island bees should be of moderate size (10–17 mm); however, their analyses were mostly restricted to non-endemic island species. In comparison, the Fijian (µ = 3.9 mm), French Polynesian (µ = 4.2 mm), and Micronesian (µ = 7.4 mm) Hylaeus, as well as the Fijian Lasioglossum (µ = 5.2 mm), are all minute to small bees and represent pre-human long-distance dispersals. Hence, our results support patterns observed by Michener (1979a) that smaller bees might be more easily dispersed by wind. Additionally, these Hylaeus are likely stem-nesters that could also disperse via rafting.
The most abundant bee species in Fiji, Lasioglossum (Homalictus) fijiense (Perkins and Cheesman, 1928), has been shown to have undergone a massive and sudden population size increase following the arrival of humans on the archipelago (Dorey et al., 2021a). Because L. (H.) fijiense favors open ground for nesting and is a super-generalist pollinator, this expansion was attributed to the broad clearing and slash-and-burn agriculture of the Lapita and post-Lapita peoples (Dorey et al., 2021a). Prior to this time, Fiji was likely dominated by hardwood forest (Roos et al., 2016). Tropical hardwood forests in Fiji today have dense canopies with cool, dark, and damp understories that might not suit either ground-nesting (Lasioglossum) or cavity-nesting (Hylaeus) bees. However, by using forest canopies, Hylaeus might avoid the cool, moldy, and quickly decaying nesting substrates of forest understories. This might have additional benefits, such as easy access to early sunlight and nearby floral resources. Hence, we predict that the opposite pattern (population size decrease) might be observed in the Fijian Hylaeus and that they are likely vulnerable to both ancient and contemporary clearing of forests. This stressor is additional to the climate-change vulnerability that has been identified for the endemic Lasioglossum species (Dorey et al., 2020b).
In Fiji alone, the Lasioglossum diversity has increased from 4 (Michener, 1979b), to 13 (Dorey et al., 2019), to 22 (Dorey et al., 2020b), with current estimates from molecular data at ~30 species (unpublished data). We have only just started to scratch the surface of the Hylaeus radiation’s true diversity in the Pacific. Our Hylaeus sampling efforts are incredibly sparse in Micronesia, French Polynesia, and even in Fiji. Between Fiji and French Polynesia, there are hundreds of islands and islets (e.g., the intervening archipelagos Tonga, Samoa, Cook Islands, Wallis, and Futuna). It is then reasonable to assume that there are many more Hylaeus species to be discovered and described across the Pacific. They need to be found and described before we can even consider conserving them.
Perhaps our most important finding is related to the methods that are broadly used to sample bees around the world. We empirically show that a decade of sampling bias has led to a gross misunderstanding of a region’s pollinator fauna. We provide evidence for the importance of forests for pollinators and that they can host a unique fauna not readily captured by standard sampling techniques (Ulyshen et al., 2023). Similar disparities have been observed in other studies. For example, sampling bias, canopy specialization, and even red-flower specialization has been shown in the very rarely collected, but widespread, hylaeine bee, Pharohylaeus lactiferus Cockerell, 1910, that was not collected for almost 100 years of Australian bee sampling until canopy sampling was employed (Dorey, 2021). Another study found that the American Augochlora pura (Say, 1836) (Halictidae) was 40 times more abundant in the canopy than understory, and even excluding this species abundance, richness and Shannon’s diversity were all higher (Ulyshen et al., 2010). Additionally, Urban-Mead et al. (2021) found that between canopy and understory, (i) bee abundance did not differ (but this changed between years), (ii) richness did not vary, but (iii) Hill-Shannon diversity was higher in the canopy. In this study and Dorey (2021), active canopy sampling was employed while Ulyshen et al. (2010) used flight-intercept traps and all found strong indications of strata-dependent patterns. The weaker patterns found by Urban-Mead et al. (2021) may be due to the use of yellow, white, and blue bee bowls. That canopy-specialist Hylaeus appear to prefer red flowers deviates from the long-held belief that bees do not see, or often forage on, red-flowering plants (von Frisch, 1914). However, Horridge (1998) argued that “bees see red” and that red flowers would be particularly high-contrast on green backgrounds, as in a forest canopy.
Such sampling bias might have large impacts on biological understandings and land management. For example, Orr et al. (2021) undertook excellent modeling of global bee diversity patterns but had to control for forestation; otherwise, patterns of increasing net primary productivity and bee diversity were inverted. This indicated that forest cover might be bad for bee diversity on a macroecological scale. While we do not refute this finding, we do highlight that sampling bias away from canopy sampling has the potential to impact such a pattern, particularly as forest canopies can be almost inaccessible for sampling. Current evidence, especially from the northern hemisphere, suggests that open forests can be beneficial for wildflower and bee abundance (Hanula et al., 2016). However, this evidence likely stems from the almost exclusive use of understory sampling techniques that will be biased towards understory bee taxa—although we show here that, under the right environmental conditions, Malaise traps may detect some canopy vagrants. This has implications for thinning, clearing, and burning management techniques and recommendations that would otherwise ignore the importance of forest and canopy pollinators (Dorey et al., 2021b; Ulyshen et al., 2023). We add to a growing discussion about bee systematic sampling (Prendergast et al., 2020; Prendergast and Hogendoorn, 2021), temporal (Dorey et al., 2020a), and strata biases (Dorey, 2021).
Despite a decade of intensive and widespread low-strata sampling across Fiji, only one Hylaeus specimen was collected using standard active-sampling techniques until canopy sampling was employed. Firstly, we show that, including our Hylaeus (Euprosopoides) and Hylaeus (Prosopisteron), there have been at least four dispersals of Hylaeus out of Australia instead of two as suggested by Kayaalp et al. (2013). However, greater work in the region is required and will allow a higher-resolution examination of hylaeine dispersal patterns. Secondly, in contrast to the Fijian Lasioglossum (Homalictus) fijiense, these bees are canopy specialists and are therefore expected to be vulnerable to both ancient and contemporary anthropogenic habitat destruction. We recommend that further sequencing and analyses are required on the demographic patterns of this new bee clade to assess possible impacts. We also postulate that, like the endemic Lasioglossum species, these tropical hylaeines might be vulnerable to changing climates. Finally, we highlight that much greater sampling and sequencing efforts in canopies across the South Pacific will lead to further discoveries. We demonstrate and argue the need for better application of canopy sampling and understory sampling methods.
Our understanding of the Pacific bee fauna as a whole continues to rapidly grow, to inform theory and conservation, and to surprise. We therefore emphasize the need for greater research funding in the region as a whole and for the training and support of local experts to continue and expand upon this work.
All data and code associated with this article are made available via our FigShare — https://doi.org/10.25451/flinders.24481231 and our sequence data are additionally made available via GenBank — SUB13956663 (OR780414-OR780432; iii and viii on FigShare). Zoobank LSID urn:lsid:zoobank.org:pub:583AF470-5B12-41CF-8957-CE4971B68C59.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because ethics approval is not required for work on insects.
JD: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Project administration, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. OD: Conceptualization, Data curation, Methodology, Project administration, Writing – review & editing. KM: Conceptualization, Data curation, Methodology, Validation, Visualization, Writing – review & editing. MPS: Conceptualization, Funding acquisition, Supervision, Writing – review & editing. A-MG: Validation, Writing – review & editing. TR: Data curation, Validation, Visualization, Writing – review & editing. MT: Data curation, Resources, Validation, Writing – review & editing. SG: Data curation, Funding acquisition, Writing – review & editing. MIS: Funding acquisition, Writing – review & editing. BP: Conceptualization, Data curation, Methodology, Software, Validation, Visualization, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Fieldwork was supported by a Federal Australian Government New Colombo Plan project (NCPST Fiji 15482 to JD, MPS, MT, and MIS); Australian Government Research Training Program scholarships (to OD); AJ and IM Naylon PhD Scholarship (to JD); and Playford Trust (to JD and OD). The Waterhouse Club funded fieldwork in Micronesia (to MPS, SG, and MIS).
The authors would like to thank the Ministry of iTaukei Affairs for access to Navai village. We would like to warmly thank and recognise Meli Naiqama, his family, and the village of Navai for their continued help during field work in Fiji. Additionally, we would like to thank Matthew Elmer and Carmen da Silva for their support during our joint Fijian field work. This work was completed under permit number 1755-11. We are also extremely grateful to Ben Robinson and all the members of the Waterhouse Club (https://waterhouseclub.com/) and the Conservation Society of Pohnpei for their assistance with collections across Micronesia.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
BPBM, Bernice Pauahi Bishop Museum; SSW, Supraclypeal Suture Width; SSL, Supraclypeal Suture Length; SAMA, South Australian Museum.
Almeida E. A. B., Danforth B. N. (2009). Phylogeny of colletid bees (Hymenoptera: Colletidae) inferred from four nuclear genes. Mol. Phylogenet. Evol. 50, 290–309. doi: 10.1016/j.ympev.2008.09.028
Almeida E., Pie M., Brady S., Danforth B. (2012). Biogeography and diversification of colletid bees (Hymenoptera: Colletidae): emerging patterns from the Southern End of the World. J. Biogeogr. 39, 526–544. doi: 10.1111/j.1365-2699.2011.02624.x
Ascher J. S., Pickering J. (2020). Discover Life bee species guide and world checklist (Hymenoptera: Apoidea: Anthophila). Available online at: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (Accessed 2022/07/27).
Barry J. D., Jérôme M., Alain P., Gordon M. (2013). Flower-visiting records of the native bees of New Caledonia. Ann. Missouri Bot. Garden. 99, 19–43. doi: 10.3417/2010076
Bouckaert R. R., Drummond A. J. (2017). bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 17, 42. doi: 10.1186/s12862-017-0890-6
Bouckaert R., Vaughan T. G., Barido-Sottani J., Duchêne S., Fourment M., Gavryushkina A., et al. (2019). BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PloS Comput. Biol. 15, 1–11. doi: 10.1371/journal.pcbi.1006650
Davies O. K., Groom S. V. C., Ngo H. T., Stevens M. I., Schwarz M. P. (2013). Diversity and origins of Fijian leaf-cutter bees (Megachilidae). Pac. Sci. 67, 561–570. doi: 10.2984/67.4.7
Dorey J. B. (2021). Missing for almost 100 years: the rare and potentially threatened bee Pharohylaeus lactiferus (Hymenoptera, Colletidae). J. Hymenoptera. Res. 81, 165–180. doi: 10.3897/jhr.81.59365
Dorey J. B., Fagan-Jeffries E. P., Stevens M. I., Schwarz M. P. (2020a). Morphometric comparisons and novel observations of diurnal and low-light-foraging bees. J. Hymenoptera. Res. 79, 117–144. doi: 10.3897/jhr.79.57308
Dorey J. B., Fischer E. E., Chesshire P. R., Nava-Bolaños A., O’Reilly R. L., Bossert S., et al. (2023a). A globally synthesised and flagged bee occurrence dataset and cleaning workflow. Sci. Data. 10, 1–17 doi: 10.1038/s41597-023-02626-w
Dorey J. B., Groom S. V. C., Freedman E. H., Matthews C. S., Davies O. K., Deans E. J., et al. (2020b). Radiation of tropical island bees and the role of phylogenetic niche conservatism as an important driver of biodiversity. Proc. R. Soc. B.: Biol. Sci. 287, 20200045. doi: 10.1098/rspb.2020.0045
Dorey J. B., Groom S. V. C., Velasco-Castrillón A., Stevens M. I., Lee M. S. Y., Schwarz M. P. (2021a). Holocene population expansion of a tropical bee coincides with early human colonisation of Fiji rather than climate change. Mol. Ecol. 30, 1–18. doi: 10.1111/mec.16034
Dorey J. B., O’Reilly R. L., Bossert S., Fischer E. E. (2023b). BeeBDC: an occurrence data cleaning package”. R package version 1.0.1. (South Australia: CRAN and in Adelaide)
Dorey J. B., Rebola C. M., Davies O. K., Prendergast K. S., Parslow B. A., Hogendoorn K., et al. (2021b). Continental risk assessment for understudied taxa post catastrophic wildfire indicates severe impacts on the Australian bee fauna. Global Change Biol. 27, 6551–6567. doi: 10.1111/gcb.15879
Dorey J. B., Schwarz M. P., Stevens M. I. (2019). Review of the bee genus Homalictus Cockerell (Hymenoptera: Halictidae) from Fiji with description of nine new species. Zootaxa 4674, 1–46. doi: 10.11646/zootaxa.4674.1.1
Drummond A. J. (2016)FigTree version 1.4.3. Available online at: http://tree.bio.ed.ac.uk/software/figtree/.
Groom S. V. C., Hayes S. E., Ngo H. T., Stevens M. I., Schwarz M. P. (2014). Recipe for disruption: multiple recent arrivals of megachilid bees in Pacific archipelagos. J. Insect Conserv. 18, 613–622. doi: 10.1007/s10841-014-9665-1
Groom S. V. C., Schwarz M. P. (2011). Bees in the Southwest Pacific: Origins, diversity and conservation. Apidologie 42, 759–770. doi: 10.1007/s13592-011-0079-8
Groom S. V. C., Tuiwawa M. V., Stevens M. I., Schwarz M. P. (2015). Recent introduction of an allodapine bee into Fiji: a new model system for understanding biological invasions by pollinators. Insect Sci. 22, 532–540. doi: 10.1111/1744-7917.12136
Hanula J. L., Ulyshen M. D., Horn S. (2016). Conserving pollinators in North American forests: a review. Natural Areas. J. 36, 427–439. doi: 10.3375/043.036.0409
Hebert P. D. N., Braukmann T. W. A., Prosser S. W. J., Ratnasingham S., deWaard J. R., Ivanova N. V., et al. (2018). A Sequel to Sanger: amplicon sequencing that scales. BMC Genomics 19, 219. doi: 10.1186/s12864-018-4611-3
HeliconSoft (2023)Helicon focus. Available online at: https://www.heliconsoft.com/heliconsoft-products/helicon-focus/.
Houston T. F. (1975). A revision of the Australian hylaeine bees (Hymenoptera: Colletidae). I. Introductory material and the genera Heterapoides Sandhouse, Gephyrohylaeus Michener, Huleoides Smith, Pharohylaeus Michener, Hemirhiza Michener, Amphylaeus Michener and Meroglossa Smith. Aust. J. Zool. Supplementary. Ser. 23, 1–135. doi: 10.1071/AJZS036
Houston T. (1981). A revision of the Australian hylaeine bees (Hymenoptera: Colletidae). II. Aust. J. Zool. Supplementary. Ser. 29, 1–128. doi: 10.1071/AJZS080
Kayaalp P., Schwarz M. P., Stevens M. I. (2013). Rapid diversification in Australia and two dispersals out of Australia in the globally distributed bee genus, Hylaeus (Colletidae: Hylaeinae). Mol. Phylogenet. Evol. 66, 668–678. doi: 10.1016/j.ympev.2012.10.018
Krombein K. V. (1950). The aculeate hymenoptera of Micronesia II. Colletidae, halictidae, megachilidae, and apidae. Proc. Hawaiian. Entomol. Soc. 14, 101–142.
Littlefield R. (2017). Zerene stacker (Richland, USA). Available at: https://www.zerenesystems.com.
MacArthur R. H., Wilson E. O. (1967). The theory of island biogeography (Princeton, USA: Princeton University Press).
Magnacca K. N., Danforth B. N. (2006). Evolution and biogeography of native Hawaiian Hylaeus bees (Hymenoptera: Colletidae). Cladistics 22, 393–411. doi: 10.1111/j.1096-0031.2006.00119.x
Michener C. D. (1965). A classification of the bees of the Australian and South Pacific Regions (New York: American Museum of Natural History).
Michener C. D. (1979a). Biogeography of the bees. Ann. Missouri Bot. Garden. 66, 277–347. doi: 10.2307/2398833
Michener C. D. (1979b). Genus homalictus in Fiji (Hymenoptera, halictidae). Pac. Insects. 21, 227–234.
Michener C. D. (2007). The bees of the world, 2nd edition (Baltimore, USA: Hopkins University Press).
Müller N. F., Bouckaert R. (2020). Adaptive metropolis-coupled MCMC for BEAST 2. PeerJ. 8, e9473. doi: 10.7717/peerj.9473
Naaz Z. T., Bibi R., Dorey J. B. (2022). Current status of bees in Fiji; geographical distribution and role in pollination of crop plants. Oriental. Insects. 56, 1–27. doi: 10.1080/00305316.2021.1982043
Orr M. C., Hughes A. C., Chesters D., Pickering J., Zhu C.-D., Ascher J. S. (2021). Global patterns and drivers of bee distribution. Curr. Biol. 31, 451–458.e454. doi: 10.1016/j.cub.2020.10.053
Patiño J., Whittaker R. J., Borges P. A. V., Fernández-Palacios J. M., Ah-Peng C., Araújo M. B., et al. (2017). A roadmap for island biology: 50 fundamental questions after 50 years of the theory of island biogeography. J. Biogeogr. 44, 963–983. doi: 10.1111/jbi.12986
Pauly A., Donovan B., Munzinger J. (2015). Les abeilles du genre Homalictus Cockerell 1919 en Nouvelle-Calédonie (Hymenoptera: Apoidea: Halictidae). Belgian. J. Entomol. 34, 1–30.
Pauly A., Munzinger J. (2003). Contribution à la connaissance des Hymenoptera Apoidea de Nouvelle-Calédonie et de leurs relations avec la flore butinée. Annales. la. Société. Entomol. France. (N.S.). 39, 153–166. doi: 10.1080/00379271.2003.10697370
Pauly A., Villemant C. (2009). Hyménoptères Apoidea (Insecta) de l’archipel du Vanuatu. Zoosystema 31, 719–730. doi: 10.5252/z2009n3a16
Perkins R. C. L., Cheesman L. E. (1928). Insects of Samoa and other Samoan terrestrial arthropods pt. I: fasc. 1 (London, UK: British Museum of Natural History).
Poulsen N. R., Rasmussen C. (2020). Island bees: do wood nesting bees have better island dispersal abilities? Apidologie 51, 1006–1017. doi: 10.1007/s13592-020-00778-x
Prendergast K. S., Hogendoorn K. (2021). Methodological shortcomings and lack of taxonomic effort beleaguer Australian bee studies. Austral Ecol. 46, 880–884. doi: 10.1111/aec.12998
Prendergast K. S., Menz M. H. M., Dixon K. W., Bateman P. W. (2020). The relative performance of sampling methods for native bees: an empirical test and review of the literature. Ecosphere 11, e03076. doi: 10.1002/ecs2.3076
Rambaut A., Drummond A. J., Xie D., Baele G., Suchard M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
Roos C. I., Field J. S., Dudgeon J. V. (2016). Anthropogenic burning, agricultural intensification, and landscape transformation in post-Lapita Fiji. J. Ethnobiol. 36, 535–553, 519. doi: 10.2993/0278-0771-36.3.535
Say T. (1836). Descriptions of new North American Hymenoptera, and observations on some already described. Boston J. Nat. His. 1, 210–305.
Tomlinson M. (2016). Little people, ghosts the anthropology of the good. J. Polynesian. Soc. 125, 11–32. doi: 10.15286/jps.125.1.11-32
Ulyshen M., Soon V., Hanula J. (2010). On the vertical distribution of bees in a temperate deciduous forest. Insect Conserv. Diversity 3, 222–228. doi: 10.1111/j.1752-4598.2010.00092.x
Ulyshen M., Urban-Mead K. R., Dorey J. B., Rivers J. W. (2023). Forests are critically important to global pollinator diversity and enhance pollination in adjacent crops. Biol. Rev. 98, 1118–1141. doi: 10.1111/brv.12947
Urban-Mead K. R., Muñiz P., Gillung J., Espinoza A., Fordyce R., van Dyke M., et al. (2021). Bees in the trees: diverse spring fauna in temperate forest edge canopies. For. Ecol. Manage. 482, 118903. doi: 10.1016/j.foreco.2020.118903
von Frisch K. (1914). Der Farbensinn und Formensinn der Bienen. Zool. Jahrbücher.: Abteilung. für. allgemeine. Zologie. und. Physiol. der. Tiere. 35, 1–188.
Yasumatsu K. (1942). Apoidea of Micronesia. III. Records of the Genera Megachile, Heriades, Ceratina and Prosopis. Tenthredo 3, 335–348.
Keywords: Fiji, French Polynesia, Micronesia, Hylaeinae, sampling methods, Lasioglossum, Colletidae, dispersal
Citation: Dorey JB, Davies OK, Magnacca KN, Schwarz MP, Gilpin A-M, Ramage T, Tuiwawa M, Groom SVC, Stevens MI and Parslow BA (2024) Canopy specialist Hylaeus bees highlight sampling biases and resolve Michener’s mystery. Front. Ecol. Evol. 12:1339446. doi: 10.3389/fevo.2024.1339446
Received: 16 November 2023; Accepted: 26 January 2024;
Published: 26 February 2024.
Edited by:
Kris Braman, University of Georgia, United StatesReviewed by:
Holger Heinrich Dathe, Senckenberg Deutsche Entomologische Institut (SDEI), GermanyCopyright © 2024 Dorey, Davies, Magnacca, Schwarz, Gilpin, Ramage, Tuiwawa, Groom, Stevens and Parslow. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: James B. Dorey, amJkb3JleUBtZS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.