 Christophe Dupichaud1*Bertrand Lefebvre1Claire H. Milne2Rich Mooi3Martina Nohejlová4Renaud Roch5Farid Saleh6*Samuel Zamora7,8
Christophe Dupichaud1*Bertrand Lefebvre1Claire H. Milne2Rich Mooi3Martina Nohejlová4Renaud Roch5Farid Saleh6*Samuel Zamora7,8- 1Unité Mixte de Recherche (UMR) 5276 Laboratoire de Géologie de Lyon, Terre, Planétes, Environnement (LGLTPE), Univ Lyon, Univ Lyon 1, Ecole Normale Supérieure de Lyon (ENSL), Centre National de la Recherche Scientifique (CNRS), Villeurbanne, France
- 2Department of Classics, University of California, Berkeley, Berkeley, CA, United States
- 3Department of Invertebrate Zoology and Geology, California Academy of Sciences, San Francisco, CA, United States
- 4Czech Geological Survey, Prague, Czechia
- 5Jurassica Museum, Porrentruy, Switzerland
- 6Institute of Earth Sciences (ISTE), University of Lausanne, Geopolis, Lausanne, Switzerland
- 7Instituto Geológico y Minero de Espana (IGME-CSIC), Zaragoza, Spain
- 8Grupo Aragosaurus-Instituto Universitario de Investigación en Ciencias Ambientales de Aragón (IUCA), Universidad de Zaragoza, Zaragoza, Spain
In the Lower Ordovician of Morocco, solutan echinoderms are relatively common, locally abundant, geographically widespread (Central and Eastern Anti-Atlas), and biostratigraphically long-ranging (late Tremadocian–mid Floian) components of the Fezouata Biota. The lower part of the Fezouata Shale (late Tremadocian) yielded one specimen, here tentatively identified as a Castericystis-like syringocrinid, with exceptionally preserved internal soft parts presumably corresponding to the distal region of the gut. Most solutan remains from the Fezouata Shale are assigned to Plasiacystis mobilis, which was already known from the late Tremadocian of France and the Darriwilian of the Czech Republic and United Kingdom. Several isolated dististeles provisionally identified as Plasiacystis sp. may belong to large individuals of P. mobilis or a new taxon. Several specimens of late Tremadocian solutans from the Fezouata Shale, morphologically intermediate between Minervaecystis vidali (late Tremadocian, France) and Plasiacystis mobilis, are assigned to Nimchacystis agterbosi gen. et sp. nov. The diagnosis of the syringocrinid family Minervaecystidae is modified to include the four genera Minervaecystis, Nimchacystis, Pahvanticystis, and Plasiacystis, all characterised by an ovoid theca, a laterally inserted feeding appendage, and a twisted, flattened dististele. Minervaecystids are interpreted as active epibenthic detritus feeders, using their dististele to crawl on soft substrates. The newly described solutan taxa highlight that the benthic communities of Fezouata Biota are more diverse than what was previously described.
Introduction
The Early Ordovician represents a critical interval in the diversification of echinoderms, with the transition from low diversity, relatively cosmopolitan assemblages dominated by eocrinoids, glyptocystitid rhombiferans, and stylophorans (Furongian–mid Tremadocian), to more diverse faunas (late Tremadocian and onwards) characterised by the onset of new classes, typical of Sepkoski’s Palaeozoic Evolutionary Fauna (e.g., asteroids, crinoids, diploporitans, and ophiuroids) (Sumrall et al., 1997; Lefebvre et al., 2013). Morocco (Anti-Atlas), along with France (Montagne Noire) and western USA (Nevada, Utah), is one of the few places in the world yielding abundant echinoderm faunas documenting this major evolutionary transition (Sprinkle and Guensburg, 2004; Lefebvre et al., 2013; Lefebvre et al., 2016). While faunas from North America (Laurentia) occur in carbonate-dominated substrates from low latitudes, France and Morocco (Gondwana) offer a good opportunity to understand such transitions at higher latitudes in siliciclastic-dominated substrates. The Anti-Atlas is also the only region known so far where exceptionally preserved soft parts (ambulacral system and gut) were documented in Early Ordovician echinoderms (Lefebvre et al., 2019; Saleh et al., 2021; Dupichaud and Lefebvre, 2022). Exceptionally preserved soft parts can be particularly important in the case of extinct classes, such as solutans, whose anatomy and phylogenetic positions within the phylum Echinodermata are debated (see, e.g., David et al., 2000; Smith, 2005; Zamora and Rahman, 2014; Lefebvre et al., 2019). Solutans are considered either basal, pre-radial echinoderms (Smith, 2005; Rahman and Lintz, 2012; Zamora and Rahman, 2014) or regular blastozoans, closely related to gogiid eocrinoids (e.g., David et al., 2000; Lefebvre and Lerosey-Aubril, 2018; Nohejlová and Lefebvre, 2022).
The class Soluta is a small, long-lived clade of echinoderms (Drumian–Emsian) characterised by a single feeding appendage, a more or less flattened theca, and a stem-like appendage (homoiostele) used for permanent attachment to the sea floor in plesiomorphic forms (e.g., Coleicarpus; Ubaghs and Robison, 1988; Daley, 1996) but generally modified into a locomotory device in most taxa (e.g., Sprinkle and Guensburg, 1997; Rahman and Lintz, 2012; Noailles et al., 2014). The fossil record suggests a Laurentian origin for solutans, with all their earliest occurrences so far documented exclusively in North America (Lefebvre and Lerosey-Aubril, 2018). During the Furongian–Tremadocian interval, solutans expanded to South China (Zamora et al., 2013) and high-latitude Gondwanan regions, such as the Anti-Atlas (Lefebvre et al., 2016; Dupichaud and Lefebvre, 2022) and the Montagne Noire (Thoral, 1935; Ubaghs, 1970; Dupichaud et al., 2023).
In the Anti-Atlas, the Fezouata Shale (Tremadocian–Floian) yielded extremely abundant remains of solutans (Lefebvre et al., 2016; Saleh et al., 2022a), some of which had putative soft parts preserved (Saleh et al., 2021; Dupichaud and Lefebvre, 2022). However, while the morphology and systematics of solutans from the Montagne Noire have been thoroughly investigated (Ubaghs, 1970; Dupichaud et al., 2023), no comparable work has been performed so far on those from the Fezouata Shale. Consequently, the aim of this paper is to present the first detailed systematic overview of solutans from the Fezouata Biota and to discuss their taphonomy, palaeoecology, and palaeobiogeographic affinities.
Geological setting
In the Anti-Atlas, remains of Early Ordovician solutans were found in distinct stratigraphic levels of the Fezouata Shale in several localities belonging to three distinct geographic areas (Figures 1B, C, D). Most solutan occurrences are situated in the Ternata Plain, north of Zagora, in the Central Anti-Atlas. In this area, the Fezouata Shale corresponds to a ca 900-m-thick, relatively monotonous siliciclastic sequence consisting mainly of siltstones, but with intercalated sandstone levels towards the top, near the transition with the overlying Zini Formation (Destombes et al., 1985; Vaucher et al., 2017). In the Zagora area, the Fezouata Shale corresponds to a storm-dominated, tide-modulated succession deposited on a relatively shallow shelf that dipped gently to the North West (in modern orientation) (Vaucher et al., 2017; Saleh et al., 2020). In the Ternata plain, the stratigraphic position of each solutan-bearing locality could be determined based on a precise biostratigraphic scheme using acritarchs, chitinozoans, conodonts, and graptolites (Lefebvre et al., 2018).
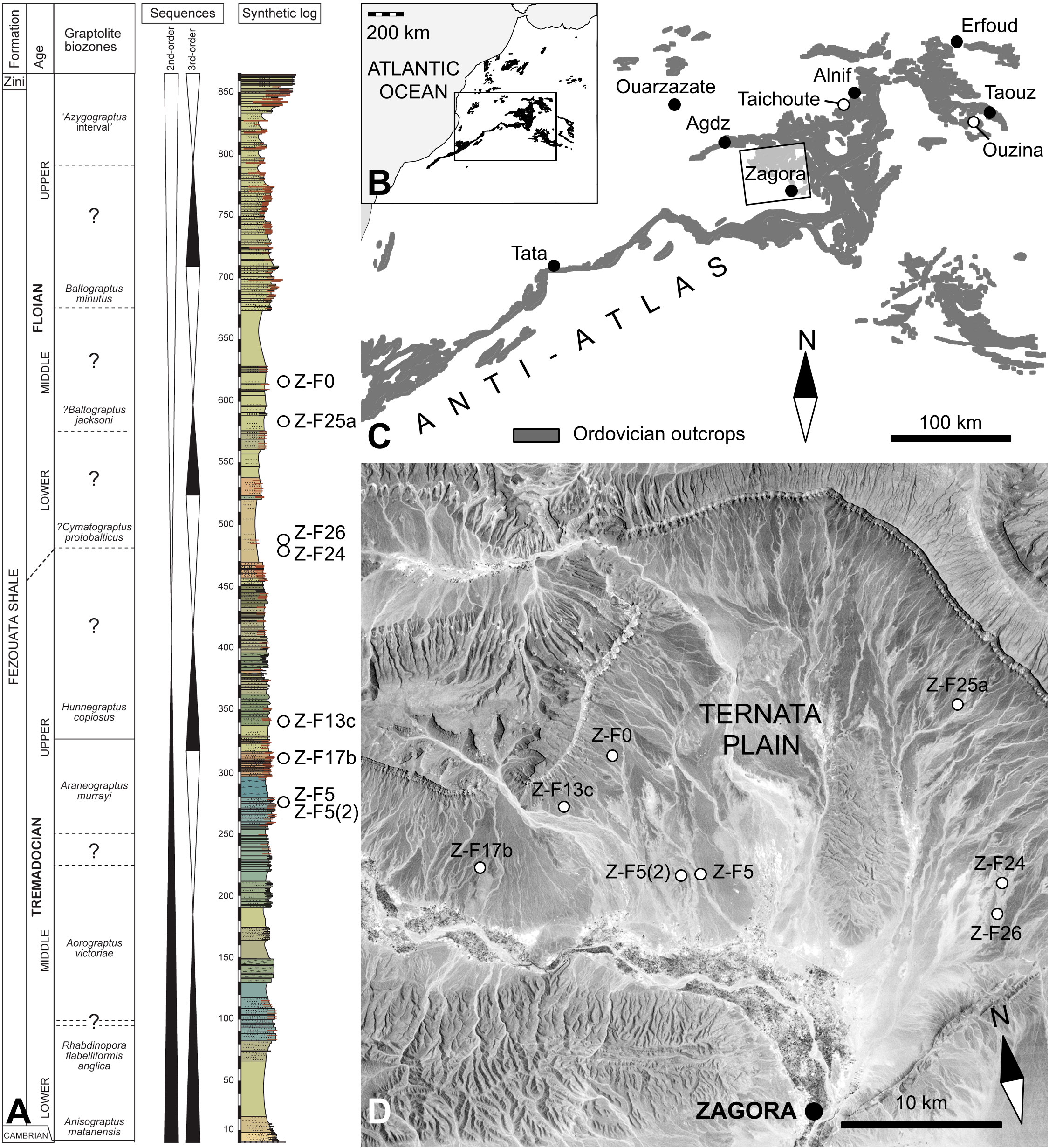
Figure 1 Stratigraphic and geographic occurrences of solutan echinoderms in the Fezouata Shale (Lower Ordovician), Anti-Atlas, Morocco. (A) Stratigraphic position of solutan localities in the Ternata plain (Zagora area, Central Anti-Atlas). Graptolite biozonation, eustatic sequences, and synthetic log (left columns) are based on Lefebvre et al. (2016); Lefebvre et al. (2018). Colours on the log correspond to those of the rocks on the outcrop. (B) Simplified geological map of northwestern Africa showing the distribution of Ordovician rocks and the location of the Anti-Atlas. (C) Simplified geological map of the Anti-Atlas showing the distribution of Ordovician rocks and the location of the three main areas yielding Early Ordovician solutan remains: Ternata plain (Zagora area), western Maïder (Taichoute), and Taouz Massif (Ouzina). (D) Landsat view of the Ternata plain (Zagora area, Central Anti-Atlas) showing the location of all main Early Ordovician solutan localities mentioned in the text and specified on the log; photograph courtesy of the U.S. Geological Survey.
In the Zagora area, the stratigraphically oldest solutan remains (Sagenograptus murrayi Zone, late Tremadocian) were found in green micaceous siltstones exposed at Oued Beni Zoli (Z-F5; Figures 1A, D, 2D) and the nearby locality of Tinzouline (Z-F5[2]; Figures 1A, D). Both sites yielded a particularly abundant and diverse fauna dominated by gastropods (Pelecyogyra), rhombiferans (Macrocystella), and trilobites (Ampyx, Asaphellus, Ceraurinella, Colpocoryphe, Euloma, Parabathycheilus, and Toletanaspis) (Ebbestad, 2016; Martin et al., 2016; Kouraïss et al., 2019). Bivalves (Redonia), brachiopods, cephalopods (Polymeres), conulariids, eocrinoids (Balantiocystis), graptolites, hyolithids, solutans, somasteroids, and stylophorans are minor elements of the assemblage (Kröger and Lefebvre, 2012; Lefebvre et al., 2016). Solutans, like most other organisms from Z-F5, are preserved intact or slightly disarticulated, suggesting in-situ burial by storm deposits at or slightly below the storm wave base (Martin et al., 2016; Kouraïss et al., 2019).

Figure 2 Diversity of preservation. (A) MHNM.15690.216, Plasiacystis sp. from the Z-F24 locality (early Floian), preserved in storm-generated accumulations. (B) MPZ.2022.927, Plasiacystis mobilis from the Taichoute locality (middle Floian), preserved in a silico-aluminous concretion. (C) UCBL-FSL.713058, Nimchacystis agterbosi from the Z-F17b locality (late Tremadocian), preserved in fine beige siltstones. (D) MHNM.15690.200, Plasiacystis mobilis from the Z-F5 locality (late Tremadocian), preserved in green micaceous siltstones. Scale bars = 10 mm. Photographs by Vincent Perrier.
In recent years, several fully articulated specimens of solutans were found in fine beige siltstones at Tignit (Z-F17b), associated with a low-diversity assemblage comprising the enigmatic fossil Webbyites felix Muir and Gutiérrez-Marco, 2023. This level is probably laterally equivalent to the nearby Z-F6 locality (Oued Ouaoufraout), which similarly yielded several specimens of Webbyites, and was assigned to the uppermost part of the S. murrayi Zone (late Tremadocian) (Muir and Gutiérrez-Marco, 2023). The preservation of fully articulated specimens of solutans and Webbyites implies that they were buried by distal storm deposits in an otherwise quiet, relatively distal setting, below the storm wave base (Saleh et al., 2020; Saleh et al., 2021). The precise locality and stratigraphic position of the single known specimen of solutan with exceptionally preserved soft parts (Figure 3) are unknown. However, the associated lithology (fine beige siltstones, Figure 2C) strongly suggests that it was found in the same stratigraphic interval as Z-F6 and Z-F17b, i.e., in the uppermost part of the lower interval with exceptional preservation identified by Lefebvre et al. (2018).

Figure 3 Photograph of CASG 72938 showing preserved soft tissues as dark masses. (A) Part. (B) Counterpart. Scale bar = 10 mm. Photographs by Vincent Perrier.
Disarticulated remains of solutans (dististeles and isolated thecal plates) are relatively common at locality Z-F13c, which is situated in the same area as Z-F6 and Z-F17b (western part of the Ternata plain; Figures 1A, D), but in stratigraphically younger levels belonging to the Hunnegraptus copiosus Zone (latest Tremadocian) (Lefebvre et al., 2016). The pistachio micaceous siltstones of Z-F13c also yielded a relatively diverse benthic assemblage consisting of disarticulated skeletal remains belonging to bivalves, brachiopods, conulariids (Eoconularia), and trilobites (Ampyx, Asaphellus, and Geragnostus). Taphonomic features suggest relatively proximal, well-oxygenated environmental conditions at, or more likely above, the storm wave base (Martin et al., 2016).
The next two solutan localities, Z-F24 and Z-F26, are geographically (southeastern part of the Ternata plain) and stratigraphically (Cymatograptus? protobalticus Zone, early Floian) very close to each other (Figures 1A, D). However, these localities have yielded distinct assemblages, probably reflecting different environmental conditions. In Z-F24, solutan remains (mostly isolated dististeles) are abundant in beige micaceous siltstones, associated with a relatively low diversity assemblage comprising mostly cephalopod phragmocones (Destombesiceras, Rioceras)? and large trilobite elements (cephalons and pygidiums), along with some rare bivalves (Ekaterodonta and Redonia), crinoids (Iocrinus), edrioasteroids (Argodiscus), eocrinoids, gastropods (Carcassonnella), hyolithids, and mitrates (Anatifopsis) (Kröger and Lefebvre, 2012; Lefebvre et al., 2016; Figure 2A). The strong disarticulation and sometimes fragmentation of most skeletal remains, as well as their occurrence in lenticular levels, are indicative of storm-generated accumulations generated in relatively proximal settings. This interpretation is further supported by the frequent current-induced orientation of cephalopod and hyolithid conchs (Kröger and Lefebvre, 2012). In contrast, the extremely fine, micaceous beige siltstones of Z-F26 (Toumiat) have yielded a particularly diverse benthic assemblage dominated by bivalves (e.g., Babinka, Coxiconchia, Ekaterodonta, Praenucula, and Redonia), gastropods (Lesueurilla), tergomyans (Carcassonnella), and trilobites (e.g., Ampyx, Asaphellus, Parabathycheilus, and Toletanaspis) (Ebbestad, 2016; Martin et al., 2016; Polechová, 2016). Other faunal elements comprise relatively common cephalopods (Destombesiceras), eocrinoids (Balantiocystis), and cornutes (Sokkaejaecystis) associated with rare asterozoans, brachiopods, conulariids, crinoids (Ramseyocrinus)?, edrioasteroids (Argodiscus), graptolites, mitrates (Balanocystites), and solutans (Kröger and Lefebvre, 2012; Lefebvre et al., 2016). Most skeletal remains are fully articulated, with some bivalves preserved in a butterfly position (Polechová, 2016). These taphonomic features are suggestive of quiet environmental conditions, below the storm wave base.
Rare, isolated dististeles of Plasiacystis collected in the beige micaceous siltstones of Z-F0 (Jbel Bou Zeroual; Figures 1A, D) and Z-F25a (Bou Chrebeb; Figures 1A, D) represent the youngest known occurrences of solutans in the Ternata plain (?Baltograptus jacksoni Zone, middle Floian). These two sites have yielded relatively similar, extremely abundant, and very diverse benthic assemblages dominated by bivalves (e.g., Babinka, Coxiconchia, and Redonia), gastropods (Lesueurilla and Sinuites), tergomyans (Carcassonnella and Thoralispira), and trilobites (Agerina, Ampyx, Asaphellus, Colpocoryphe, Euloma, Parabathycheilus, Prionocheilus, and Toletanaspis) (Ebbestad, 2016; Martin et al., 2016; Polechová, 2016). The associated fauna includes relatively common brachiopods, cephalopods (Polymeres), conulariids (Archaeoconularia and Eoconularia), cornutes (Sokkaejaecystis), eocrinoids (Balantiocystis), graptolites, and hyolithids, as well as rare demosponges (Pirania), edrioasteroids (Argodiscus), anomalocystitid mitrates, and possibly also the pterobranch()? W. felix (Kröger and Lefebvre, 2012; Botting, 2016; Lefebvre et al., 2016; Muir and Gutiérrez-Marco, 2023). In both sites, the occurrence of exceptionally preserved organisms (e.g., demosponges and Webbyites) associated with articulated remains of skeletonised taxa (e.g., trilobites) suggests relatively quiet environmental conditions at or slightly below the storm wave base (Martin et al., 2016).
In the Anti-Atlas, Early Ordovician solutan remains were also found in two other areas outside of the Ternata plain. In the western Maïder, at Taichoute (approximately 18 km SW of Alnif; Figure 1C), a concretion-bearing level (i.e., assemblage a1 in Saleh et al., 2022a) yielded a Plasiacystis-dominated assemblage (Figure 2B) that could be assigned to the middle Floian based on the presence of the graptolite Baltograptus deflexus (Saleh et al., 2022a). Other faunal elements include brachiopods, cephalopods (Bathmoceras), gastropods (Lesueurilla), mitrates (Anatifopsis), glyptocystitid rhombiferans (Macrocystella), somasteroids, and trilobites. Contrary to the situation in localities from the Ternata plain (see above), fossil remains in Taichoute are preserved three-dimensionally in silico-aluminous concretions. Most solutans are intact and complete (feeding appendage, theca, and homoiostele), though frequently slightly disarticulated and collapsed. Their preservation suggests that concretions were probably initiated around decaying carcasses exposed on the sea floor in relatively quiet, distal environments, below the storm wave base (Loi and Dabard, 1999; Loi and Dabard, 2002; Saleh et al., 2022a).
Similarly preserved solutan remains were found in the Taouz Massif (eastern Anti-Atlas) at Destombes’ locality 1540 (Figure 1C; Ouzina, in Havlíček, 1971). Destombes (2006) proposed an early Floian age for this concretion-bearing level. However, contrary to the situation in Taichoute, the Ouzina assemblage is largely dominated by the glyptocystitid rhombiferan Macrocystella bohemica, while Plasiacystis mobilis is relatively rare (Lefebvre et al., 2016). Other members of the Ouzina assemblage include brachiopods (Plectorthis), cephalopods, conulariids, hyolithids (Cavernolites and Elegantilites), mitrates (Anatifopsis), somasteroids, tergomyans (Thoralispira), and trilobites (e.g., Ampyx and Asaphidae) (Havlíček, 1971; Marek, 1983; Destombes, 2006; Lefebvre et al., 2016). As in Taichoute, it is very likely that this assemblage was autochthonous and associated with relatively distal environmental conditions below the storm wave base.
Materials and methods
Due to preservation as external and internal moulds (imprints) in the rock, fossils were cast with latex to reveal their original aspect. Latex casts were whitened with ammonium chloride (NH4Cl) sublimated for observation and imaging purposes. Fossils and latex casts were photographed using a Canon EOS 5SD R equipped with a Canon 100-mm macro lens and a Canon EOS 70D equipped with an EF-S 60-mm macro lens. Latex casts were observed and drawn with a stereomicroscope Zeiss Discovery.V8 equipped with a camera lucida for drawings. Drawings were reworked digitally with the software Krita v5.1.5. Measurements were made with the software ImageJ.
Over 150 specimens were observed based on material belonging to the following public institutions: Cadi-Ayyad University, Marrakesh, Morocco (AA); Natural History Museum, London, UK (BMNHUK); geological collections of the California Academy of Sciences, San Francisco, USA (CASG); Muséum d’Histoire Naturelle, Marseille, France (MHNM); Muséum d’Histoire Naturelle, Nantes, France (MHNN); Musée des Confluences, Lyon, France (ML); Museo de Ciencias Naturales de la Universidad de Zaragoza, Zaragoza, Spain (MPZ); Senckenberg Museum, Frankfurt, Germany (SMF); palaeontological collections of Lyon 1 University, Villeurbanne, France (UCBL-FSL); and Yale Peabody Museum, New Haven, USA (YPM). For comparative purposes, additional material belonging to the National Museum, Prague, Czech Republic (NMP), and several private collections in the Czech Republic, France, and Morocco was also considered.
Systematic palaeontology
Phylum Echinodermata Klein, 1778
Class Soluta Jaekel, 1901
Remarks: The systematic subdivision of solutans into two main orders (Dendrocystitida and Syringocrinida) based on the morphology of their homoiostele follows Noailles et al. (2014). Similarly, descriptive morphological terms used below are those used in all recent descriptions of solutans (e.g., Lefebvre et al., 2012; Rahman and Lintz, 2012; Noailles et al., 2014; Lefebvre and Lerosey-Aubril, 2018; Dupichaud and Lefebvre, 2022; Nohejlová and Lefebvre, 2022; Rozhnov, 2022; Dupichaud et al., 2023; Zamora and Gutiérrez-Marco, 2023). Discussion of the phylogenetic position of solutans within the phylum Echinodermata is beyond the scope of this paper.
Order Syringocrinida Noailles et al. 2014
Family Minervaecystidae Ubaghs, 1970
Emended diagnosis: Family of syringocrinids with ovoid theca lacking lobation or marginal frame. No visible differentiation between the lower and upper thecal sides. Thecal plates thin, smooth, polygonal, tesselated, and irregular in both shape and size. Feeding appendage emerging abruptly on the lateral side of the theca, relatively far from the apex. Small, cone-shaped anal pyramid on opposite, lateral edge of the theca, close to homoiostele insertion.
Remarks: The family Minervaecystidae was originally erected by Ubaghs (1970) to include Minervaecystis vidali (Thoral, 1935), from the Saint-Chinian Formation (late Tremadocian, Montagne Noire, France), as well as isolated homoiosteles from the Whipple Cave Formation (Cambrian Stage 10, Nevada, USA; Ubaghs, 1963). The diagnosis of this family is emended here to include all syringocrinids (i.e., solutans with a flattened, oblique dististele) possessing a large ovoid theca with a laterally inserted feeding appendage. In this expanded definition, the family Minervaecystidae includes the four genera Minervaecystis Ubaghs, 1970, Nimchacystis gen. nov., Pahvanticystis Lefebvre and Lerosey-Aubril, 2018, and Plasiacystis Prokop and Petr, 2003.
The morphology of the earliest known member of the family, Pahvanticystis utahensis (Weeks Formation, Guzhangian, Utah, USA), suggests that minervaecystids probably originated from a Castericystis-like ancestor. Pahvanticystis differs from other minervaecystids in the more anterior insertion of its feeding appendage along the antanal thecal side (Lefebvre and Lerosey-Aubril, 2018). The morphology of the isolated minervaecystid homoiosteles from the late Cambrian of Utah is very close to that of Minervaecystis (i.e., similar proxistele consisting of over 20, regular tetramerous rings; Ubaghs, 1963; Sumrall et al., 1997). However, the absence of thecal remains and precise information on the location of the feeding appendage prevents any identification at generic level. Similarly, as yet undescribed putative minervaecystids have been also reported from the Wah Wah Formation (late Floian) of Utah, a more precise taxonomic assignment is so far impossible due to the absence of information concerning their morphology (Sprinkle and Guensburg, 1993; Dupichaud et al., 2023).
The three minervaecystid genera Minervaecystis, Nimchacystis gen. nov., and Plasiacystis have almost identical thecae and feeding appendages, but they differ strongly in the morphology of their homoiosteles. In Minervaecystis, the proxistele is elongate and consists of numerous (over 20) regular telescopic rings, each made of four plates (Thoral, 1935; Ubaghs, 1970). Its long, flattened dististele is relatively narrow, stem-like, markedly curved, and bears several comb-like spikes along its keeled, concave (right) margin (Ubaghs, 1970; Dupichaud et al., 2023). Plasiacystis is a long-lived genus known from the Saint-Chinian Formation (late Tremadocian, France), the Fezouata Shale (late Tremadocian–middle Floian, Morocco), the Hope Shale (Darriwilian, UK), and the Šarká Formation (Darriwilian, Czech Republic) (Jaekel, 1918; Prokop and Petr, 2003; Lefebvre et al., 2012; Lefebvre et al., 2016; Dupichaud and Lefebvre, 2022; Dupichaud et al., 2023). Its proxistele is not organised into regular rings, being a flexible cylindrical structure made of numerous, imbricate scale-like elements. In Plasiacystis, the dististele corresponds to a short, broad, paddle-like structure, with a wide, rounded distal extremity (Prokop and Petr, 2003; Lefebvre et al., 2012; Dupichaud and Lefebvre, 2022; Dupichaud et al., 2023). Because of the unusual “beaver-tailed” morphology of its dististele, Plasiacystis was originally placed in a distinct family, Plasiacystidae (Prokop and Petr, 2003; Lefebvre et al., 2012). However, the discovery of syringocrinids (Nimchacystis gen. nov.) with a homoiostele morphologically intermediate between those of Minervaecystis and Plasiacystis (see below) suggests instead that they all belong to the same clade. Consequently, Nimchacystis gen. nov. and Plasiacystis are here assigned to the family Minervaecystidae.
The reevaluation of the morphology of Dehmicystis Caster, 1968, based on new material from the lower Ludlow (Silurian) of the Central Iberian Zone (Spain; Zamora and Gutiérrez-Marco, 2023) questions its systematic position within the class Soluta. Dehmicystis was originally described based on a few poorly preserved, strongly flattened, and tectonically distorted specimens from the Hunsrück Shale (Emsian) of Germany (Dehm, 1934; Rahman and Lintz, 2012). Dehmicystis has usually been considered a typical dendrocystitid solutan (see, e.g., Caster, 1968; Rahman and Lintz, 2012; Noailles et al., 2014; Zamora and Gutiérrez-Marco, 2023). However, new morphological data (e.g., large ovoid, non-lobate theca; laterally inserted feeding appendage; keeled dististele; Rahman and Lintz, 2012; Zamora and Gutiérrez-Marco, 2023) suggest either syringocrinid affinities or a morphological convergence of some Siluro-Devonian dendrocystitids with Cambro-Ordovician syringocrinids, possibly resulting from similar modes of life.
Genus Plasiacystis Prokop and Petr, 2003
Type species: Plasiacystis mobilis Prokop and Petr, 2003, by monotypy.
Emended diagnosis. Genus of syringocrinid solutan with very flexible, cylindrical proxistele composed of numerous, unorganised, imbricate, scale-like elements. Proxistele and dististele of nearly equal lengths. Dististele comprising two series of large, opposite plates forming short, wide, rigid, flattened paddle with rounded distal extremity. One side of dististele frequently ornamented with single spike-shaped knob placed on the convex half.
Remarks: The genus Plasiacystis was originally described as a very unusual solutan with a large, polyplated, flattened theca, conspicuously elongate proxistele made of numerous scale-like elements, a short paddle-like dististele, and, possibly, no feeding appendage (Prokop and Petr, 2003). This initial reconstruction was based on relatively abundant material preserved in siliceous concretions from the Šarká and Dobrotivá formations (Darriwilian) of the Prague Basin, Czech Republic. However, the reevaluation of this material showed that it was a mixture of at least three different echinoderm taxa (Lefebvre et al., 2012). The generic name Plasiacystis was retained for the holotype and some other specimens, all from the Šarká Formation and corresponding to regular solutans with a feeding appendage, relatively ovoid theca, and distinctive paddle-like dististele (Lefebvre et al., 2012). Other portions of the original reconstruction of Plasiacystis were assigned to another, yet undescribed solutan (theca only) and to a rhenopyrgid edrioasteroid (proxistele only) (Lefebvre et al., 2012).
Plasiacystis mobilis Prokop and Petr, 2003
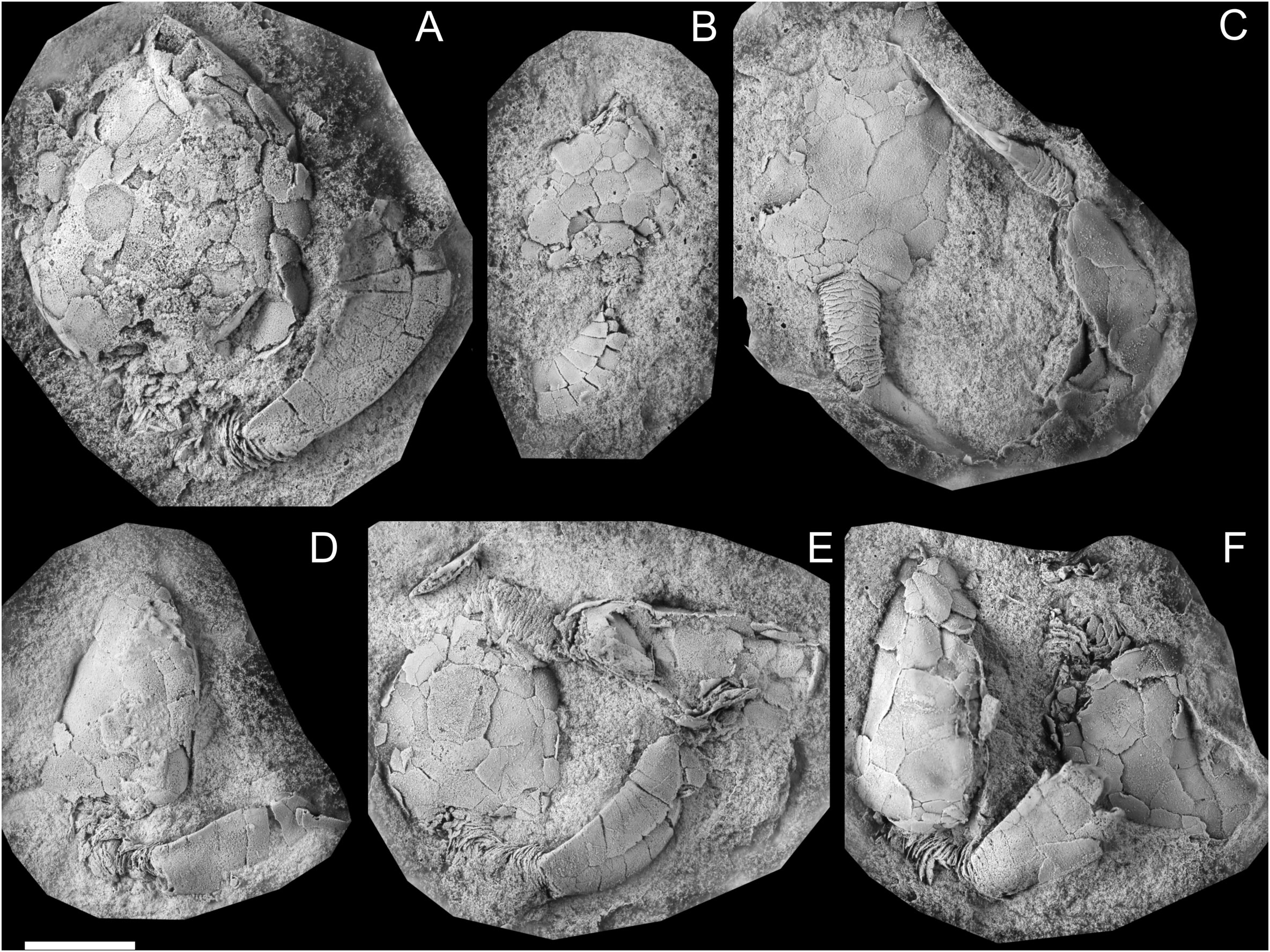
Figure 4 Whitened casts of Plasiacystis mobilis occurring in concretions from Taichoute (middle Floian). (A) MPZ.2022.927. (B) MPZ.2023.251. (C) ML20.269363 showing two specimens in the same concretion. (D) ML20.269394. (E) ML20.269360 showing two specimens in the same concretion. (F) ML20.269308 showing two specimens in the same concretion. Scale bar = 10 mm. Photographs by Vincent Perrier.
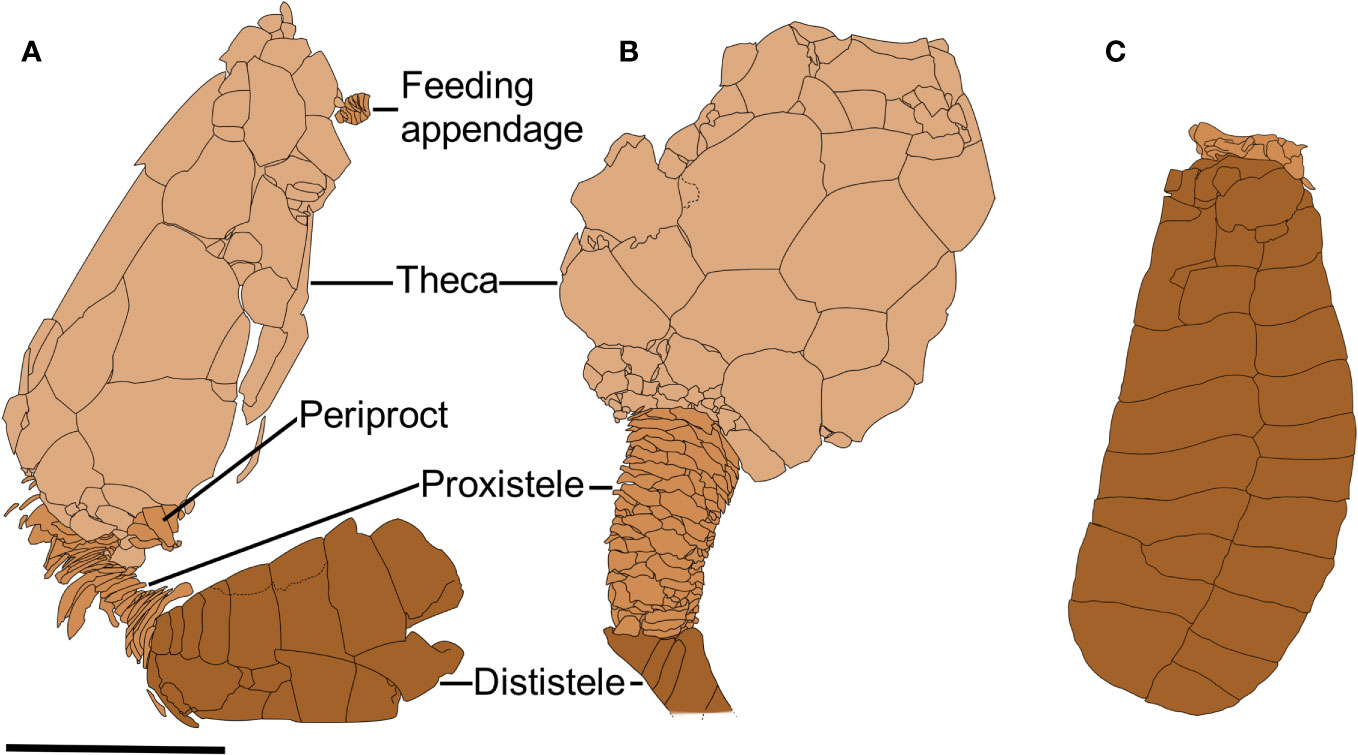
Figure 5 Observation drawings of Plasiacystis mobilis preserved in concretions from Taichoute (middle Floian). (A) ML20.269306 showing a specimen with both its periproct and a part of its feeding appendage. (B) ML20.269363 showing a nearly intact proxistele. (C) ML20.269402 showing an isolated dististele. Scale bar = 10 mm.
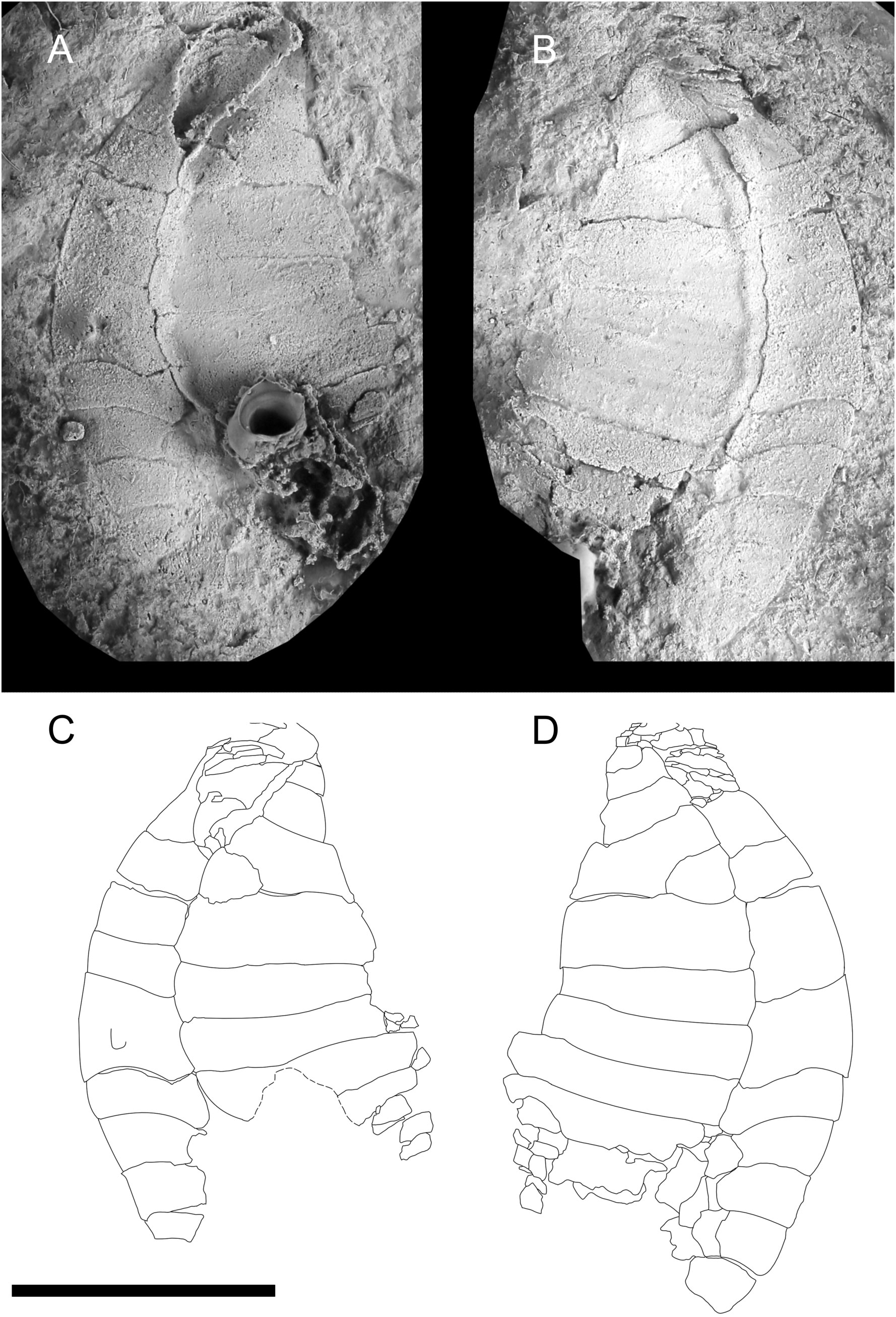
Figure 6 The only specimen of Plasiacystis mobilis found so far in France (Montagne Noire, late Tremadocian). (A) Whitened cast of the UCBL-FSL.711925 counterpart showing the ornamental knob. (B) Whitened cast of the UCBL-FSL.711925 part. (C) Observation drawing of the UCBL-FSL.711925 counterpart showing the ornamental knob. (D) Observation drawing of the UCBL-FSL.711925 part. Scale bar = 10 mm. Photographs by Vincent Perrier.
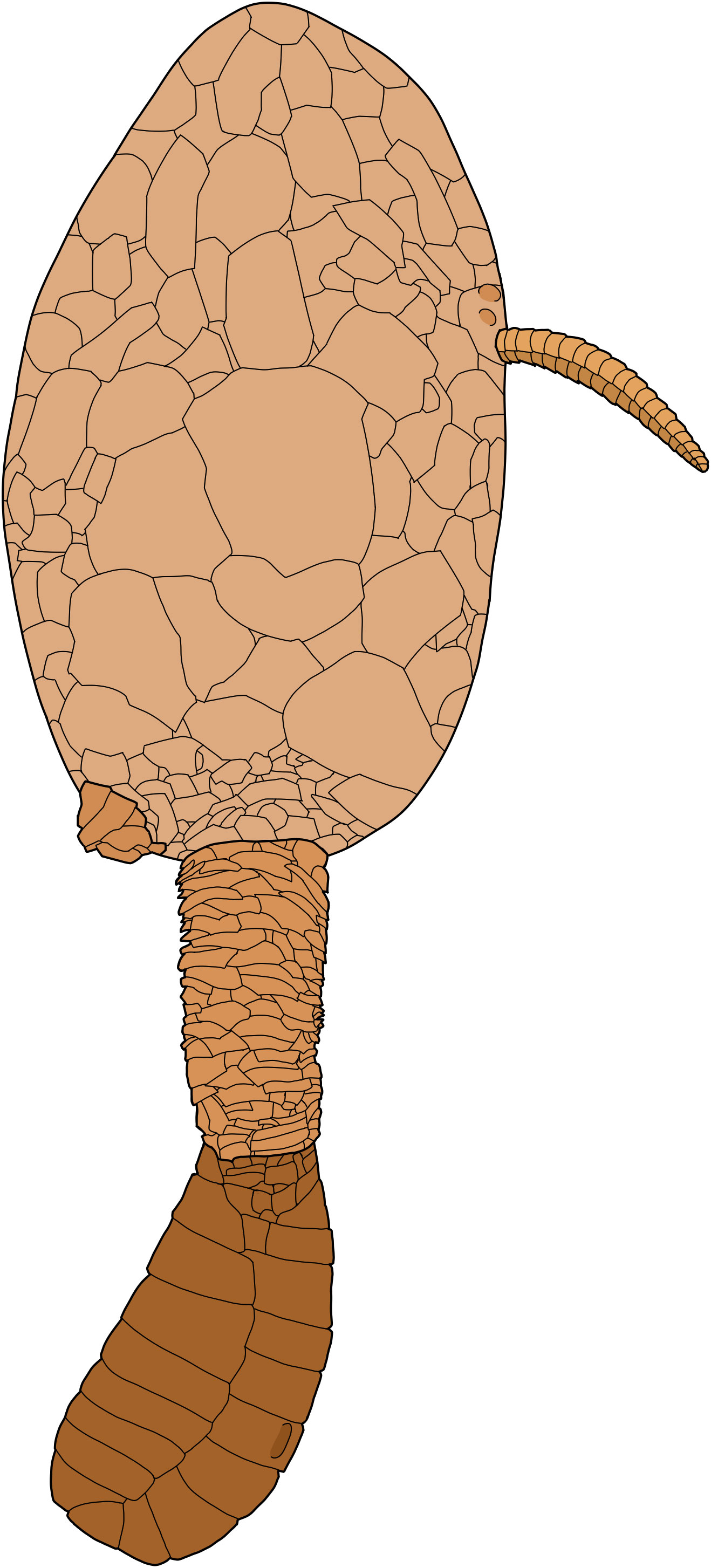
Figure 7 Reconstruction of the underside of Plasiacystis mobilis.
Holotype: NMP.L.13216.
Type level: Šarká Formation, early–middle Darriwilian (Middle Ordovician).
Type locality: Osek, near Rokycany, Prague Basin, Czech Republic.
Additional new material, localities, and horizons: The studied material from the Fezouata Shale, Anti-Atlas (Morocco), includes three specimens from Oued Beni Zoli (Z-F5; Figures 1A, D), S. murrayi Zone, late Tremadocian (MHNM.15690.200 and ML20.269401–402), 12 specimens from the hill N of Ouaoufraout (Z-F13c; Figures 1A, D), H. copiosus Zone, late Tremadocian (AA.CNOc.OI.5–6, AA.CNOc.OI.9, AA.CNOc.OI.12, AA.CNOc.OI.14, AA.CNOc.OI.18–20, AA.CNOc.OI.22, AA.CNOc.OI.24, AA.CNOc.OI.31, and AA.CNOc.OI.45), two specimens from Toumiat (Z-F26; Figures 1A, D), C.? protobalticus Zone, early Floian (AA.TMT.OI.78–79), five specimens from Ouzina (Taouz Massif; Figure 1C), C.? protobalticus Zone, early Floian (UCBL-FSL.711645, UCBL-FSL.711648–649, ML20.268923, and ML20.269072), one specimen from Bou Chrebeb (Z-F25a; Figures 1A, D),?B. jacksoni Zone, middle Floian (UCBL-FSL.712446), eight specimens from Jbel Bou Zeroual (Z-F0; Figures 1A, D),?B. jacksoni Zone, middle Floian (AA.JBZ.OI.137, AA.JBZ.OI.141, AA.JBZ.OI.145–147, AA.JBZ.OI.202, ML20.269378, and YPM.IP.520778), 90 specimens from Taichoute (Western Maïder; Figure 1C),?B. jacksoni Zone, middle Floian (AA.TAI2.OI.1, UCBL-FSL.712954, UCBL-FSL.712956, UCBL-FSL.712958–959, UCBL-FSL.713593, ML20.269255, ML20.269265, ML20.269267, ML20.269271, ML20.269275–277, ML20.269283, ML20.269301–309, ML20.269311–360, ML20.269362–375, MPZ.2022.926–927, and MPZ.2023.251), and three specimens from unknown localities of the Ternata Plain (BMNHUK.EE.8727, MHNN.P.045588, and YPM.IP.371).
Diagnosis: As for genus, by monotypy.
Description: On average, specimens ca 50 mm long from the apex of the theca to the tip of dististele. Homoiostele approximately half the total length of the body, ca 28 mm long on average. Dististele representing approximately 60% of homoiostele length, ca 18 mm long on average.
Theca ovoid, slightly elongate, lacking lobes or a marginal frame (Figure 4). No visible differentiation between the upper and lower sides. Thecal plates numerous, irregular, polygonal, tesselate, almost smooth, flat or slightly convex, generally larger around the middle part of theca, and smaller around homoiostele insertion (Figures 4, 5B).
Feeding appendage stemming laterally, at an approximately one-third distance to the apex, along the antanal thecal side (Figures 4F, 5A; Type specimen NMP.L.29120). Apical tip and cover plates not observed.
Gonopore and hydropore situated on one or two plates, near insertion of feeding appendage (see, e.g., specimens UCBL-FSL.711949, ML20.269319, ML20.269324, and ML20.269337). Right of homoiostele insertion, small cone-shaped periproct, slightly bulging outward, made of thin, elongate plates arranged in a circle around the anal opening (Figures 4E, F, 5A).
Homoiostele inserted in the middle part of the posterior margin (i.e., opposite thecal apex) and divided into two distinct parts. Proxistele cylindrical, made of many thin, scale-shaped, imbricate elements of irregular sizes (Figures 4C, 5B). Dististele short, flattened, bean-shaped, with rounded tip (Figures 4A, D, E, 5C). Paddle-like portion of homoiostele made of two rows of opposite, tightly sutured, polygonal plates of irregular size. Dististele transversely thicker along longitudinal contact between opposite rows and thinner abaxially along its sharp lateral edges (Figures 4A, D, E, 5C). Longitudinal thickening of dististele enclosing narrow cylindrical lumen (see, e.g., specimens ML20.269321 and ML20.269325). In approximately 75% of observed specimens, short, smooth, spike-shaped knob borne by one plate of convex half of dististele (Figures 4B, 6). Knob highly variable in position.
Remarks: The particularly abundant material collected in the Fezouata Shale can be assigned to solutans (single feeding appendage) and more precisely to syringocrinids (flattened, oblique dististele). Moreover, the lateral insertion of the feeding appendage on the antanal thecal side and the morphology of the periproct (small, cone-shaped anal pyramid) both support a placement within the family Minervaecystidae. The possession of an unorganised proxistele made of numerous imbricate platelets and the possession of a dististele modified into a short, wide, bean-shaped paddle are diagnostic features of the genus Plasiacystis. Comparison of the Fezouata specimens of Plasiacystis with the original type material of P. mobilis did not reveal any significant morphological differences. Consequently, the Moroccan material is here assigned to P. mobilis, thus confirming the particularly long stratigraphic range of this taxon (late Tremadocian–middle Darriwilian; see Lefebvre et al., 2012; Lefebvre et al., 2016; Dupichaud et al., 2023). Moreover, the particularly abundant and well-preserved material from the Anti-Atlas brings new data on previously undocumented (e.g., gonopore and hydropore) or poorly known morphological features of P. mobilis (e.g., anal pyramid and feeding appendage) (see Lefebvre et al., 2012). The Moroccan material also makes it possible to document the occurrence of intraspecific variability with respect to the presence and position of the ornamentation (spike-shaped knob) on the dististele.
Plasiacystis sp.
Figures 2A, 8, 9
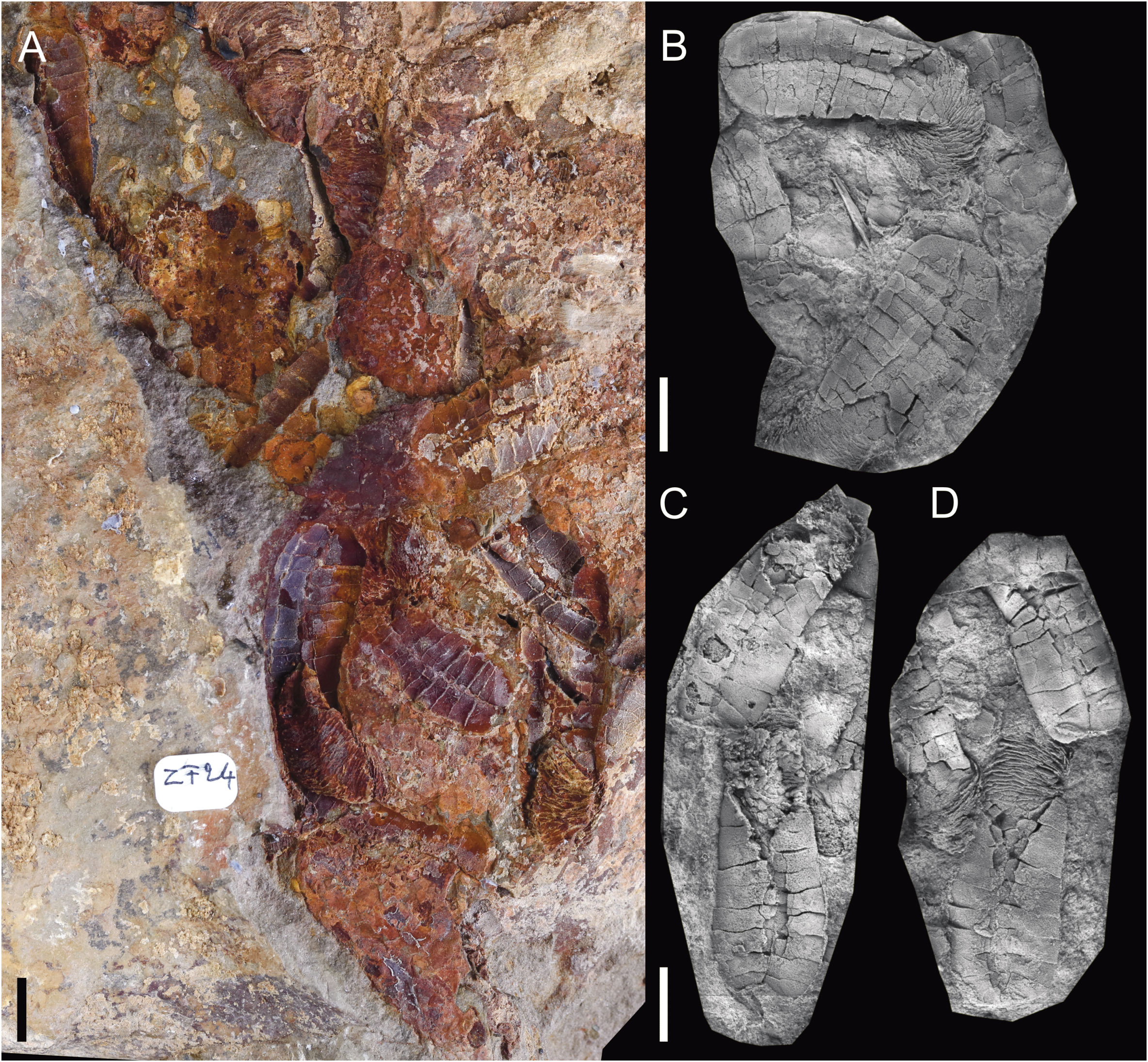
Figure 8 Plasiacystis of the Z-F24 locality; storm-generated accumulations (early Floian). (A) Photograph of MHNM.15690.224 showing the accumulation of disarticulated steles and thecae. (B) Whitened cast of MHNM.15690.214. (C) Whitened cast of MHNM.15690.222 part. (D) Whitened cast of MHNM.15690.222 counterpart. Scale bars = 10 mm. Photographs by Vincent Perrier.
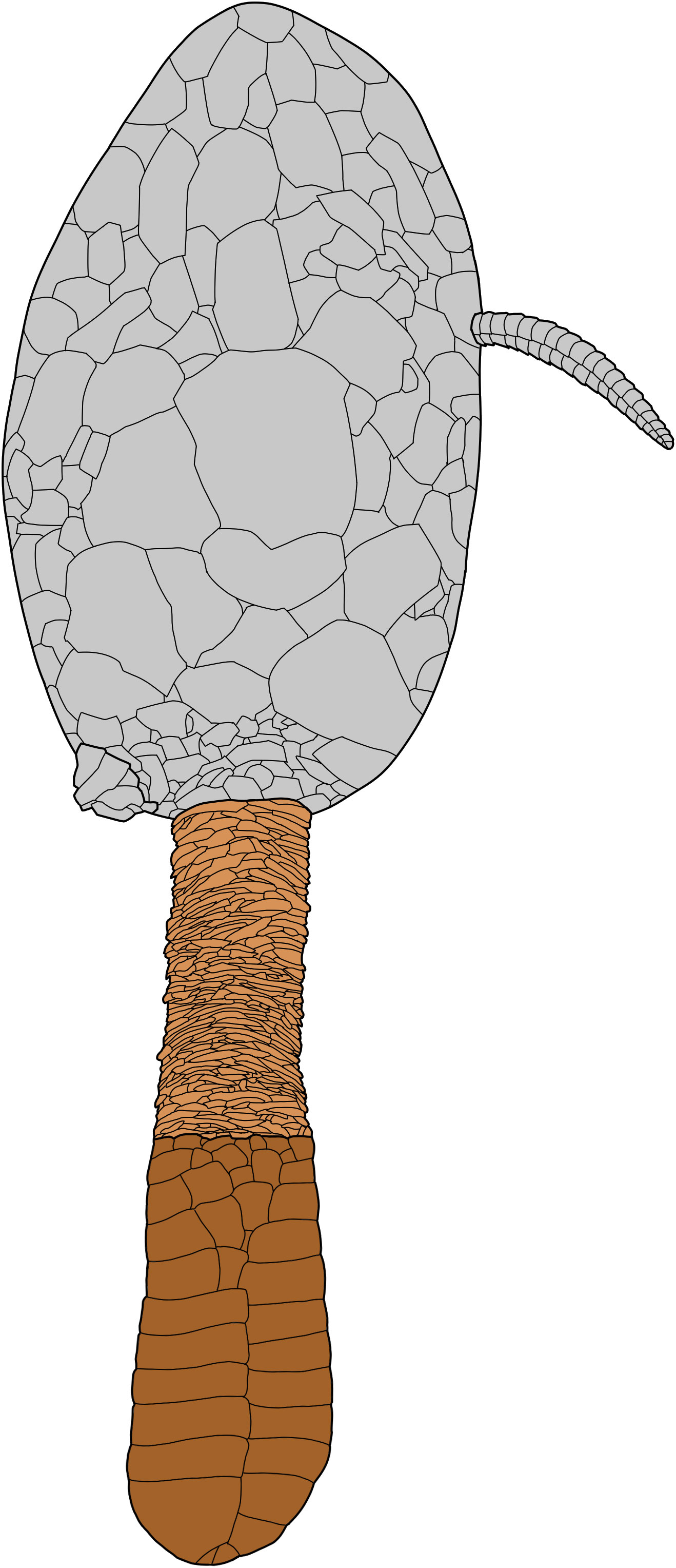
Figure 9 Reconstruction of the underside of Plasiacystis sp. Grey areas extrapolated from Plasiacystis mobilis.
Material, localities, and horizons: Included material from the Fezouata Shale (Anti-Atlas, Morocco) comprises 16 specimens from the hill North of Toumiat (Z-F24; Figures 1A, D), C.? protobalticus Zone, early Floian (AA.ONT.OI.10, MHNM.15690.211–215, and MHNM.15690.217–226) and one specimen (MHNN.P.045587) from an unknown locality (possibly Taichoute)? preserved in a concretion.
Description: Theca of numerous, irregular, tesselate, smooth, polygonal plates. Thecal outlines and size unknown; only three incomplete, disarticulated thecae could be observed (Figure 8A; specimens MHNM.15690.212, MHNM.15690.224, and MHNM.15690.226). Thecal orifices (anus, gonopore, and hydropore) not observed. Two originally connected portions of the same feeding appendage preserved in one specimen (MHNM.15690.224): one part still inserted into disarticulated theca, and the other in contact with dististele. Proxistele consisting of numerous, irregularly arranged, scale-like imbricate elements (Figures 8B, D). Dististele approximately twice as long as proxistele: on average, proxistele ca 14 mm long, dististele ca 30 mm long (Figure 8). Dististele made of two lateral rows of opposite, rectangular, tightly sutured plates forming flat, straight paddle, with broad and rounded tip (Figure 8).
Remarks: In locality Z-F24, a single storm-generated lens yielded numerous, remarkably large paddle-shaped dististeles, sometimes still in contact with proxisteles, and seldom associated with strongly disarticulated thecae. All available morphological features (theca consisting of numerous, smooth, polyplated plates, unorganised proxistele made of imbricate elements, broad, paddle-shaped dististele with a rounded posterior extremity) strongly support the assignment of these solutans to the genus Plasiacystis. However, their dististeles appear morphologically different from those of P. mobilis: they are proportionately longer (compared to the length of the proxistele), their outlines are straight (instead of bean-shaped as in P. mobilis), and their distal extremity is markedly wider than their anterior margin (whereas both margins are of approximately the same width in P. mobilis). Moreover, no ornamentation is present on the numerous observed dististeles from Z-F24, while most specimens of P. mobilis display a strong spike-shaped knob on their distal paddle. The same morphological features are present in a single large, three-dimensionally preserved, fully articulated incomplete specimen from an unknown concretion-bearing locality (MHNN.P.045587).
A first possible interpretation is to consider that all features departing from the standard morphology of P. mobilis result from allometry and can be explained by the unusually large size of specimens from Z-F24 (Dupichaud and Lefebvre, 2022). The occurrence of morphologically distinct, successive ontogenetic stages has already been seen in some solutans (see, e.g., Noailles et al., 2014). Comparable allometries have been also documented in other Palaeozoic echinoderms, such as eocrinoids (Parsley, 2012; Nohejlová and Fatka, 2016) and stylophorans (Lefebvre et al., 2022). In this interpretation, the unusual morphology of the Z-F24 solutans simply results from their larger size, and all specimens can be assigned to P. mobilis. However, the possibility that these solutans correspond to a distinct, endemic species of Plasiacystis, characterised by a morphologically distinct dististele, cannot be excluded.
Nimchacystis gen. nov.
Derivation of name: From nimcha, a Moroccan curved sword, referring to the shape of the dististele, and the suffix -cystis (ancient Greek), meaning “bag”, commonly used for solutan generic names.
Type species: Nimchacystis agterbosi sp. nov., by monotypy.
Diagnosis: Genus of syringocrinid solutan with very flexible, distally tapering proxistele, composed of more than 20 telescopic cylindrical hexamerous rings, and rigid dististele comprising two sets of large, opposite plates forming elongate, relatively narrow, smooth (unornamented), scimitar-shaped paddle with blunt distal extremity.
Remarks. Minervaecystis, Nimchacystis, and Plasiacystis have almost identical ovoid thecae and laterally inserted feeding appendages, but very different homoiosteles. Nimchacystis shares with Minervaecystis the possession of a well-organised proxistele consisting of more than 20 regular rings, and of a relatively narrow, elongate, curved dististele. For this reason, the first observed specimens of Nimchacystis were assigned to Minervaecystis by Lefebvre and Fatka (2003). However, proximal rings are generally tetramerous in Minervaecystis, while they are hexamerous in Nimchacystis. In Minervaecystis, the width of the stem-like dististele remains more or less constant along its length, while in Nimchacystis, this region forms a broad, elongate blade, tapering both anteriorly and posteriorly. Nimchacystis shares with Plasiacystis the possession of a dististele modified into a stiff paddle-like structure. However, this region is short and broad and with a large, rounded distal extremity in Plasiacystis, whereas it is more elongate and narrower and with a blunt tip in Nimchacystis. In many aspects, the homoiostele of Nimchacystis appears morphologically intermediate between those of Minervaecystis and Plasiacystis (Dupichaud and Lefebvre, 2022). This observation is a strong argument supporting placement of these taxa in the same clade (Minervaecystidae).
Nimchacystis agterbosi sp. nov.
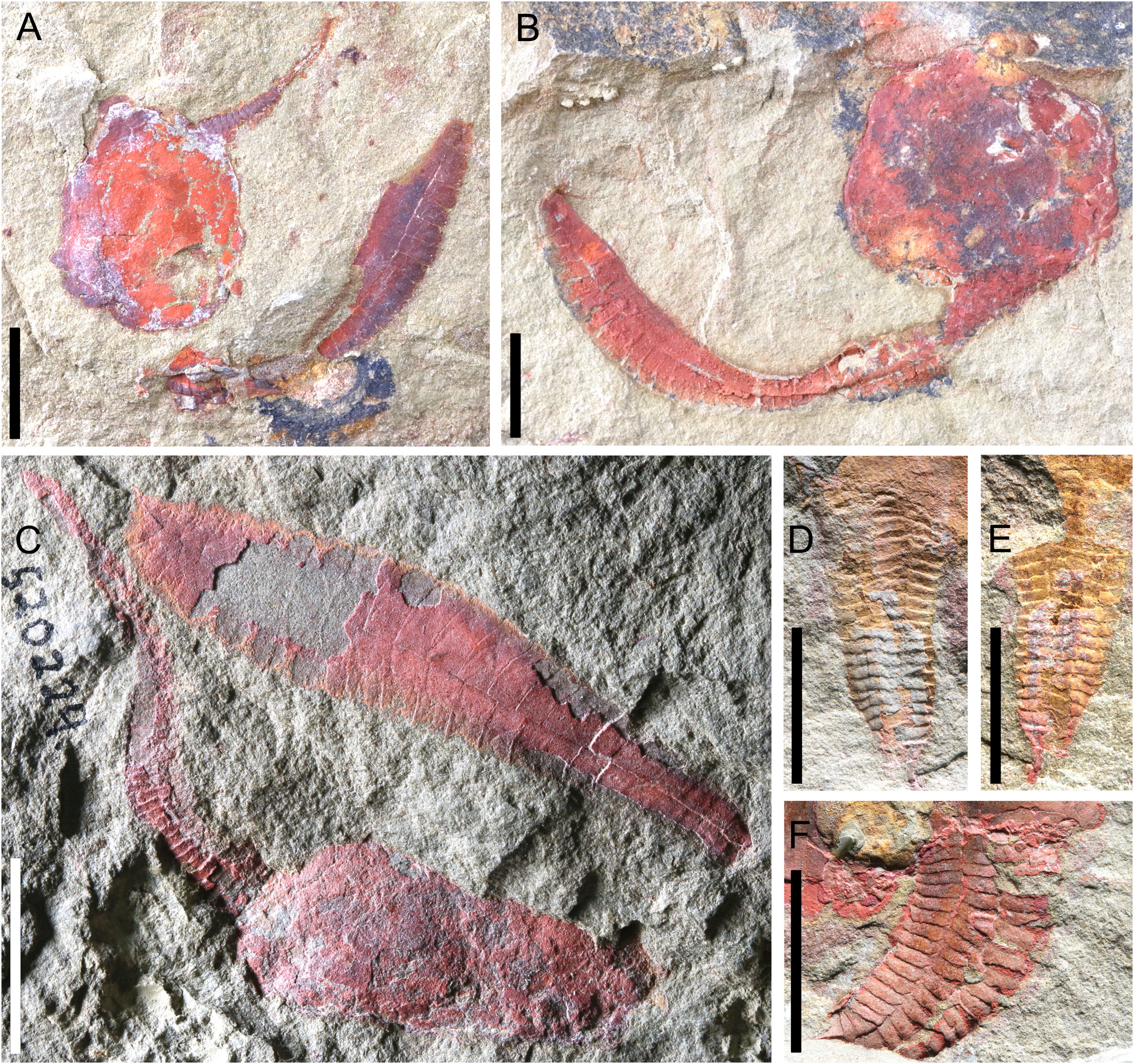
Figure 10 Photographs of Nimchacystis agterbosi from the Z-F17b locality (late Tremadocian). (A) Holotype UCBL-FSL.713120 showing theca, feeding appendage, proxistele, and dististele in near anatomical connectivity as in life. (B) SMF.89671 showing theca and stele in anatomical connection. (C) YPM.IP.520029 showing a large specimen with an apparently complete feeding appendage and dististele. (D) YPM.IP.520031 part showing a complete and well-preserved proxistele. (E) YPM.IP.520031 counterpart showing a complete and well-preserved proxistele. (F) YPM.IP.520370 showing a complete and well-preserved proxistele. Scale bars = 10 mm. Photographs (A) and (B) by Vincent Perrier. Photographs (C) to (F) by Jessica Utrup
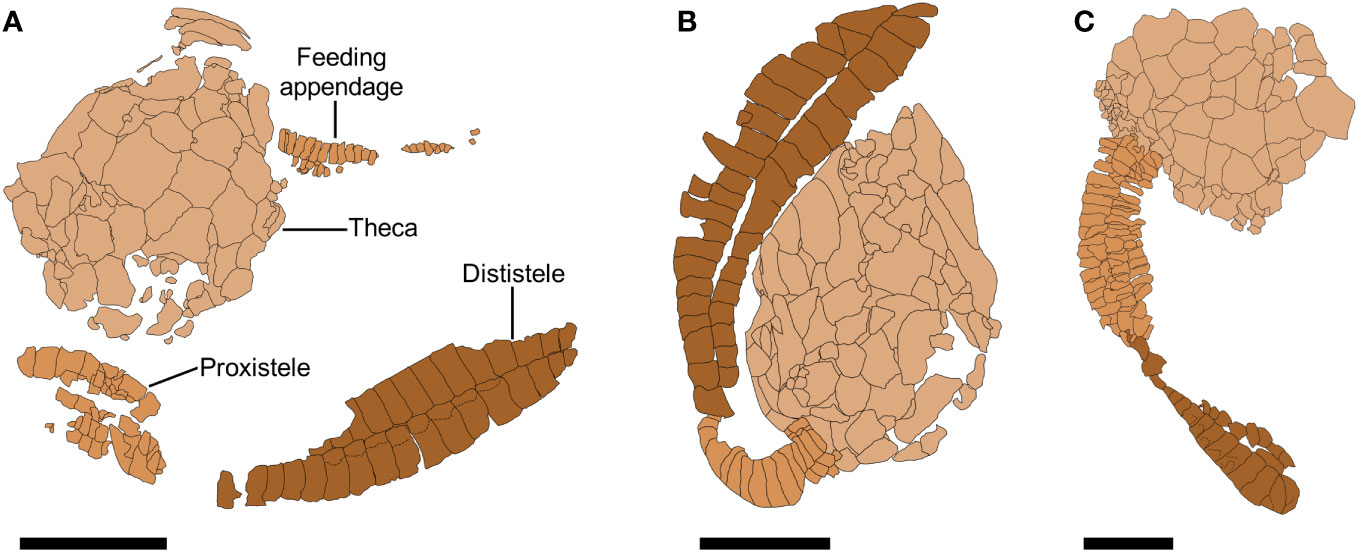
Figure 11 Camera lucida drawings of Nimchacystis agterbosi. (A) Holotype UCBL-FSL.713120 showing theca, feeding appendage, proxistele, and dististele almost in near anatomical connectivity as in life. (B) UCBL-FSL.713058 showing an almost complete dististele. (C) SMF.89671 showing theca and stele in anatomical connection. Scale bars = 10 mm.
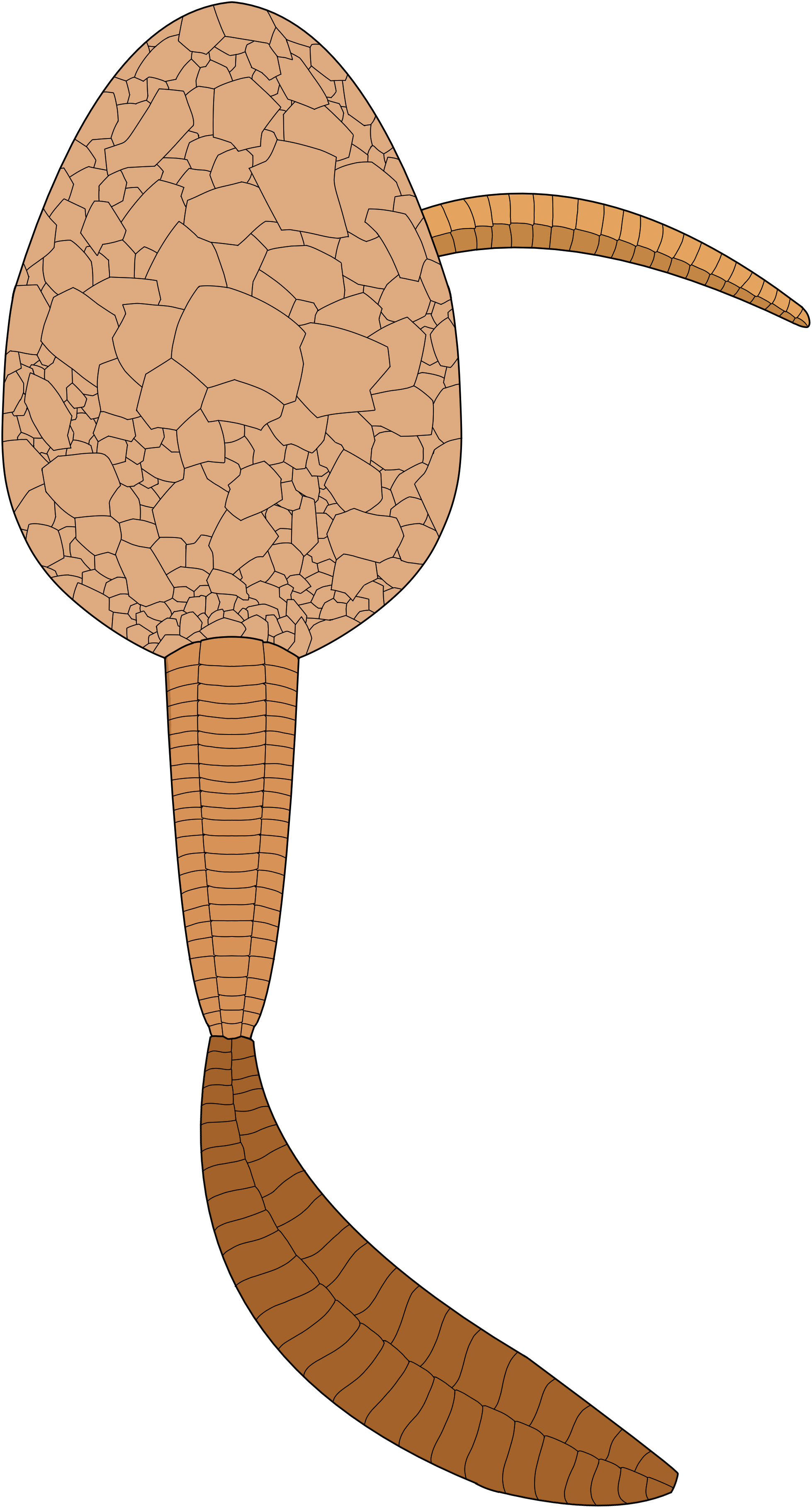
Figure 12 Reconstruction of the underside of Nimchacystis agterbosi.

Figure 13 Artistic reconstruction of Nimchacystis agterbosi by Christophe Dupichaud.
Derivation of name: The name refers to the non-academic Dutch palaeontologist Ton Agterbos, who collected and made available for study the most complete available specimen (herein designated as holotype) of the new species.
Holotype: UCBL-FSL.713120 (Figures 10A, 11A).
Type level: Fezouata Shale, S. murrayi Zone, late Tremadocian (Lower Ordovician).
Type locality: Tignit (Z-F17b; Figures 1A, D), approximately 26 km NW of Zagora, Ternata plain, Central Anti-Atlas (Morocco).
Additional material, localities, and horizons: Six specimens are from Tinzouline (Z-F5[2]; Figures 1A, D), S. murrayi Zone, late Tremadocian (YPM.IP.520227–231 and YPM.IP.520370). Three other specimens were collected in the lower part of the Fezouata Shale (S. murrayi Zone, late Tremadocian), but from an unknown locality in the Zagora area (SMF.89671–672 and UCBL-FSL.713058). Over 20 additional specimens were observed in private collections, most of them originating either from Tignit (Z-F17b) or from Tinzouline (Z-F5[2]).
Description: On average, specimens ca 73 mm long from thecal apex to the tip of dististele. Theca ovoid, slightly elongated, without evidence of anal lobe or marginal frame. No obvious difference in plating between the lower and upper thecal sides, both made of many polygonal, smooth, tesselate plates of different sizes (Figures 2C, 10A–C). Thecal orifices (anus, gonopore, hydropore) not observed.
Long, distally tapering feeding appendage inserted on left (antanal) thecal side. Feeding appendage 14 mm long in holotype (Figures 10A, 11A), 26 mm long and 3.5 mm wide at contact with theca in YPM.IP.520229 (Figure 10C). Feeding appendage consisting of at least 25 (holotype) and over 50 (YPM.IP.520229) pairs of alternating rectangular flooring plates, and two sets of smaller, opposite rectangular cover plates (Figures 10A, C, 11A).
Homoiostele inserted opposite to thecal apex and representing ca 60% of total body length; homoiostele ca 48 mm long on average. Cylindrical, distally tapering proxistele made of over 20 well-defined, regular telescopic rings, each consisting of six rectangular plates (Figures 10A, B, D–F, 11C). Elongate, curved dististele forming flat, rigid paddle tapering both anteriorly and posteriorly (Figures 2C, 10A, C, 11B, C). Distal paddle ca 30 mm long on average, and forming ca 65% of homoiostele length. Dististele consisting of two opposite rows of plates, each made of approximately 20 smooth (unornamented), rectangular elements (Figures 2C, 10A, C, 11B, C).
Family indet.
Syringocrinid gen. and sp. indet.
Material, locality, and horizon: The single known specimen (CASG 72938; Figure 3) is from an unknown locality of the Ternata plain, Zagora area (Central Anti-Atlas, Morocco). Associated lithology (fine beige siltstones) and preservation (occurrence of pyritised soft tissues within the theca) suggest that the specimen was collected at the top of the lower interval with exceptional preservation of the Fezouata Shale (S. murrayi Zone, late Tremadocian; Lefebvre et al., 2018).
Description: Specimen ca 66 mm long, from feeding appendage distal extremity to tip of dististele (feeding appendage: 6 mm; theca: 16 mm; homoiostele: 44 mm). Theca and associated feeding appendage preserved as extremely faint colourful (oxidised) impressions on rock (Figure 3).
Small, non-lobate theca with regularly convex, ellipsoid outlines. Narrow, elongate (9 mm long), dark, pyritised, three-dimensional intrathecal structure, extending anteriorly from the central part of theca (ca 6 mm away from the insertion of feeding appendage) into small, cone-shaped pyramid posteriorly, next to homoiostele insertion (Figure 3).
Short, poorly preserved portion of wide, segmented feeding arm inserted at the thecal apex (Figure 3).
Homoiostele long, narrow, proximally straight, slightly curved towards sharp tip (Figure 3). Details of homoiostele obscured by preservation as thin, oxidised crust.
Remarks: In spite of the very poor preservation of its skeletal elements, this specimen can be readily identified as a solutan echinoderm, based on the occurrence of a single, stout feeding appendage, a distinct theca, and an elongate stem-like structure (Lefebvre et al., 2016; Saleh et al., 2021; Dupichaud and Lefebvre, 2022). In the lower part of the Fezouata Shale, this type of preservation is typical for echinoderms occurring in levels with exceptional preservation (Lefebvre et al., 2019). Their original calcite skeletons are largely dissolved away, with only their outlines appearing as faint, oxidised impressions on the rock. In contrast, echinoderm soft tissues (ambulacral system and gut) can be preserved as three-dimensionally pyritised structures (Lefebvre et al., 2019; Saleh et al., 2021). In this context, it is very likely that the elongate structure preserved in specimen CASG 72938 represents exceptionally preserved soft parts. Its morphology, position, and posterior connection with a small pyramid (very likely corresponding to the periproct) suggest that this elongate structure corresponds to the hind gut (Saleh et al., 2021; Dupichaud and Lefebvre, 2022). In this specimen, the preferential preservation of the posterior part of the digestive system (the anterior part of the gut is missing) possibly results from its infilling by sediment.
The overall morphology of specimen CASG 72938 (e.g., large, flattened homoiostele) suggests probable syringocrinid affinities. The apical insertion of the feeding appendage excludes its assignment to the family Minervaecystidae, instead supporting affinities with Cambrian-like taxa such as Coleicarpus and Castericystis (Lefebvre et al., 2016; Dupichaud and Lefebvre, 2022). The overall morphology of CASG 72938 strongly recalls that of Castericystis vali Ubaghs and Robison, 1985: similar ellipsoid theca, slightly offset feeding appendage insertion, and long homoiostele with a sharp distal extremity. However, given the poor preservation of the single known specimen from the Fezouata Shale, it is untenable to assign it to the genus Castericystis, which was originally described from the Marjum Formation (Drumian) of Utah (Ubaghs and Robison, 1985; Daley, 1995). However, as yet undescribed Castericystis-like solutans were recently discovered in the Neuville Formation (lower Katian, Upper Ordovician) of Québec, Canada (R. Mooi, pers. obs., 2023). This suggests the persistence through time (at least up to the Late Ordovician) of solutans with Cambrian-like morphologies.
Discussion
Taphonomy
The main indicator for echinoderm taphonomy is the integrity of the calcite skeleton (Ausich, 2001). Fossil echinoderms have been arranged into taphonomic groups, based on experimental analyses of taphonomy conducted on extant taxa (e.g., Meyer, 1971; Kidwell and Baumiller, 1990; Donovan, 1991; Nebelsick, 1995; Kerr and Twitchett, 2004). These taphonomic groups are defined by how resistant their skeletal remains are to post mortem transport and disarticulation (Brett et al., 1997). Following this taphonomic classification, solutans are generally assigned to the “type 1” category, meaning that their skeleton is made of weakly sutured plates that quickly disarticulate after the death of the organism (a few days to a few weeks; Gorzelak and Salamon, 2013) by analogy to modern “type 1” echinoderms such as asteroids and ophiuroids (Lefebvre, 2007; Nohejlová and Lefebvre, 2022). In this context, uncovering extensively contiguous solutan endoskeletal remains implies not only a relatively rapid burial by obrution deposits (e.g., distal storm sediments) soon after or before death but also little or no transport and low bioturbation rates once buried. In the Lower Ordovician of the Anti-Atlas, such taphonomic conditions probably prevailed in most solutan-bearing localities that are generally associated with relatively distal environmental conditions at or below the storm wave base: Z-F0 (Jbel Bou Zeroual), Z-F5 (Oued Beni Zoli), Z-F5(2) (Tinzouline), Z-F17b (Tignit), Z-F26 (Toumiat), Ouzina, and Taichoute. All these localities, in which fully articulated solutans are preserved, can be considered “echinoderm Lagerstätten” sensu Smith (1988).
In contrast, some other localities (e.g., Z-F13c and Z-F24) have yielded solutan dististeles almost exclusively. Associated proxisteles are sometimes preserved, but thecae are generally totally disarticulated into hundreds of isolated plates. Such levels usually occur in more proximal, higher energy, storm-generated accumulation deposits, along with fragments of other invertebrates (e.g., cephalopods and trilobites). Such accumulations are particularly informative because they suggest that different parts of the same individual could follow distinct taphonomic pathways. Clearly, the solutan feeding appendage was the most fragile part of the organism and the most likely to be disarticulated. The more frequent preservation of large portions of proxisteles indicates that this part of the animal was a bit more resistant to disarticulation. However, the storm-generated accumulations of distal paddles of Plasiacystis-like solutans demonstrate that the dististele was the most taphonomically resistant module of the organism. This implies that the dististele of Plasiacystis was a particularly rigid structure made of tightly sutured plates. Following the taphonomic classification of Brett et al. (1997), solutans are assigned to the “type 1” category. However, Plasiacystis-like solutans possess more rigid, decay-resistant modules (e.g., dististele), so part of their anatomy could be assigned to the “type 2” category.
Palaeoecology
The earliest known solutans were permanently sessile (Coleicarpus sprinklei, Wheeler Shale, Drumian) or attached during juvenile stages (C. vali, Marjum Formation, Drumian) (Ubaghs and Robison, 1985; Ubaghs and Robison, 1988; Daley, 1995; Daley, 1996; Zamora et al., 2013; Zamora et al., 2017; Lefebvre and Lerosey-Aubril, 2018). All later solutans are generally considered vagile organisms, even if the presence of a hook at the tip of the homoiostele in some taxa might be related to the persistence of an attached juvenile stage. In this context, minervaecystids provide a remarkable example of an evolutionary trend towards more active mobility, with the transition from long, flattened stem-like appendages (Minervaecystis) towards short, broad, and rigid paddle-like structures (Plasiacystis). The slightly oblique dististele of P. mobilis was the stiffest and most resistant part of the organism (see above). It was thus perfectly designed for crawling (probably backwards, i.e., appendage first) on soft siliciclastic substrates. Its rounded posterior margin suggests the total absence of any attachment structure at juvenile stages (e.g., distal hook or blunt tip) and a permanently free mode of life.
The ovoid theca of minervaecystids is a plesiomorphic feature, inherited from permanently attached solutans (e.g., Coleicarpus). Such a morphology was possibly retained in minervaecystids because it also turned out to be well-suited for active backward crawling on soft substrates by significantly reducing the contact and therefore the resistance to traction exerted by the sediment on smooth, rounded, undifferentiated thecal walls. In contrast, markedly flattened solutan thecae, with contrasting morphologies on their lower and upper sides, possibly represent adaptations to less active epibenthic modes of life (e.g., solutan indet., gen. and sp. nov. in Lefebvre et al., 2012). Such flattened thecal morphologies provide indirect clues that fuel the ongoing debate on the life orientation and feeding posture of solutans (see, e.g., Noailles et al., 2014, and references therein). Direct comparison with other epibenthic organisms suggests that in such solutans, the flat side of their theca was very likely in contact with the substrate, whereas the opposite, convex surface was facing the water column.
Such a comparison is not possible in the case of minervaecystids: their globose, ovoid thecal morphology provides no clues to identify which surface was originally in contact with the substrate. The lack of a flat thecal face in minervaecystids may imply a degree of difference in feeding behaviour with more flattened solutans. As such, minervaecystids could be interpreted as having shown a mixed mode of feeding, allowed by the flexibility of their feeding appendage, being able to switch between bottom-feeding and suspension-feeding. However, their life orientation can be deduced indirectly based on the ornamentation occurring on the homoistele of Minervaecystis. In M. vidali, the concave portion of the dististele bears several comb-like lateral projections (Bather, 1913; Ubaghs, 1970; Dupichaud et al., 2023). Ubaghs (1970) suggested that these structures were originally directed upwards. He interpreted them as fin- or wing-like processes that could help Minervaecystis to swim forward (i.e., theca first; Ubaghs, 1970). This orientation of the comb-like structures (on the left side of the dististele) implies that the periproct was situated left of the homoiostele, and the feeding appendage was inserted, on the opposite, right thecal side (Ubaghs, 1970). In such an orientation, the cover plates (and the underlying ambulacral food groove) are facing away from the substrate, which implies a suspension-feeding mode of life for Minervaecystis (Ubaghs, 1970).
However, the possession of an ovoid theca, a very flexible proxistele that likely housed powerful musculature (by analogy with stylophorans and pleurocystitid rhombiferans; see, e.g., Gorzelak and Zamora, 2016; Lefebvre et al., 2019), and a flattened, oblique dististele, sometimes modified into a rigid paddle (Plasiacystis), suggests that the morphology of minervaecystids was better designed for active backward crawling on soft substrates than for swimming (Prokop and Petr, 2003; Lefebvre and Lerosey-Aubril, 2018). In this interpretation, the comb-like structures occurring on the dististele of Minervaecystis were more likely directed downwards, acting as gripping devices to increase the contact surface between the dististele and the substrate. If this orientation is correct, then the comb-like structures of Minervaecystis were located on the right side of its dististele. In minervaecystids, the anus is therefore situated on the right side of the homoiostele, while the feeding appendage is inserted on the opposite (left) side of the theca. In such an orientation, the cover plates (and the ambulacral food groove) are facing downwards, implying that minervaecystids were detritus feeders. This interpretation is compatible with the exceptional preservation of the hind gut (possibly full of ingested sediment) in the Castericystis-like specimen from the Fezouata Shale (see above; Figure 3). This gut also appears to be partially pyritised (see also Lefebvre et al., 2019). The ingested Fe-rich sediments could have aided the preservation of the hind gut by reducing bacterial activities under oxic conditions (Saleh et al., 2019) and by contributing to the seeding of pyrite precipitation once sulphate-reducing conditions were established (Saleh et al., 2020).
In the Lower Ordovician of the Anti-Atlas, minervaecystids (and in particular, Plasiacystis) are geographically widespread (Ternata plain, western Maïder, Taouz Massif), occurring in a large range of environmental conditions (from proximal, storm-influenced settings to more distal, quiet environments) and through a relatively long time interval (late Tremadocian–middle Floian). However, in most localities (e.g., Bou Chrebeb, Jbel Bou Zeroual, Oued Beni Zoli, Tinzouline, and Toumiat) solutans are a minor component of taxonomically diverse assemblages dominated by molluscs (bivalves, cephalopods, gastropods, and tergomyans) and trilobites (Lefebvre et al., 2016). In Ouzina, minervaecystids are also rare, but they are associated with a low diversity, almost monospecific fauna dominated by the glyptocystitid rhombiferan M. bohemica (Lefebvre et al., 2016).
In the Lower Ordovician of the Anti-Atlas, Taichoute (Figure 1C) stands out as the only locality yielding a low-diversity, solutan-dominated assemblage. Taphonomic features of Taichoute solutans (i.e., three-dimensionally preserved, complete, slightly disarticulated or collapsed individuals) suggest that most of them were very likely dead and already decaying on the sea floor when burial occurred (i.e., assemblage a1 in Saleh et al., 2022a). Comparable solutan-dominated assemblages have been described in the middle Cambrian of Utah, USA (Ubaghs and Robison, 1985; Ubaghs and Robison, 1988; Daley, 1995; Daley, 1996) and the Upper Ordovician of the Prague Basin, Czech Republic (Noailles et al., 2014; Nohejlová et al., 2019) and Tafilalt, Morocco (Hunter et al., 2010; Nohejlová and Lefebvre, 2022). The Taichoute echinoderm Lagerstätte provides the first example of a nearly monospecific, dense solutan bed in the Early Ordovician. As is frequently the case in echinoderm dense beds (see Lefebvre, 2007 and references therein), it is possible that the rarity and low diversity of associated benthic invertebrates (e.g., brachiopods, molluscs, and trilobites) result from unfavourable (oligotrophic)? environmental conditions leading to the opportunistic colonisation of the sea floor by gregarious, high-density populations of Plasiacystis (Taichoute) or Macrocystella (Ouzina). The oligotrophic interpretation does not contradict the presence of abundant large carcasses of filter-feeding arthropods in the a2 assemblage of Taichoute (see Saleh et al., 2022a). The a2 assemblage is younger and consists of transported material from more proximal sites, unlike the in-situ, non-transported solutans that colonised the Taichoute seafloor.
Palaeobiogeography
Castericystis, from the Marjum Formation (Drumian, Utah), represents the earliest known syringocrinid solutan (Noailles et al., 2014; Lefebvre and Lerosey-Aubril, 2018). The occurrence of Pahvanticystis in the Weeks Formation (late Guzhangian, Utah; Lefebvre and Lerosey-Aubril, 2018) and of minervaecystid remains in the Whipple Cave (Cambrian Stage 10, Nevada; Ubaghs, 1963; Sumrall et al., 1997) and Wah Wah (late Floian, Utah; Sprinkle and Guensburg, 1993) formations suggests a probable Laurentian origin for minervaecystids.
The onset of relatively cosmopolitan, undifferentiated low-diversity echinoderm faunas in the Furongian–Tremadocian interval probably explains the expansion of minervaecystids into Gondwanan regions, where they are documented as early as the late Tremadocian (S. murrayi Zone) in the Saint-Chinian Formation, Montagne Noire (France: Minervaecystis and Plasiacystis; Thoral, 1935; Ubaghs, 1970; Dupichaud et al., 2023) and the Fezouata Shale, Anti-Atlas (Morocco: Nimchacystis and Plasiacystis; Lefebvre et al., 2012; Lefebvre et al., 2016; Dupichaud and Lefebvre, 2022). The occurrence of closely related minervaecystid taxa, including one shared species (P. mobilis) in the Montagne Noire and the Anti-Atlas, confirms the existence of strong faunal links between these two regions during the Early Ordovician (e.g., Lefebvre et al., 2016; Saleh et al., 2022b). However, Minervaecystis has not so far been documented in Morocco, while this genus is relatively common in coeval deposits from France. Conversely, Nimchacystis is apparently endemic to the Central Anti-Atlas. In the Montagne Noire, Plasiacystis is known from a single isolated dististele, whereas it is geographically widespread and locally abundant in the Anti-Atlas.
Beyond these differences, possibly related to preservational biases and/or subtle differences in environmental conditions, Early Ordovician solutan faunas from the Anti-Atlas and the Montagne Noire make it possible to record a key evolutionary transition within minervaecystids. Within a relatively short time interval (late Tremadocian), their long, flattened, stem-like dististele with lateral comb-like projections (Minervaecystis) was modified into an elongate (Nimchacystis) and then a short, rigid distal paddle (Plasiacystis). This new morphotype, probably triggered by the adoption of a more active, crawling mode of life on soft substrates, persisted at least until the middle Darriwilian: Plasiacystis was documented in the Šarká Formation of the Prague Basin (Czech Republic; Prokop and Petr, 2003; Lefebvre et al., 2012) and the Hope Shale of Shropshire (UK; K. Derstler in Lefebvre et al., 2012). The occurrence of P. mobilis in the Darriwilian of Bohemia confirms the strong faunal links between this region and other high-latitude (peri)Gondwanan areas (Anti-Atlas, Montagne Noire) belonging to the same Mediterranean Province in the Ordovician (Lefebvre et al., 2022 and references therein). The presence of P. mobilis in the Hope Shale is more unexpected because, in the Middle Ordovician, Shropshire was part of Avalonia, which was then an independent micro-continent separated from Gondwana by the opening of the Rheic Ocean. However, several other examples of echinoderm taxa with Gondwanan affinities have been reported in the Middle Ordovician of Avalonia (see, e.g., Jefferies, 1987; Fone, 2003; Lefebvre et al., 2012; Botting et al., 2013; Lefebvre et al., 2022). These shared taxa suggest that they originated from the same pool of organisms before Avalonia rifted away from Gondwana and/or that exchanges were still possible on both sides of the Rheic Ocean in the Middle Ordovician.
Conclusions
The systematic description of solutans from the Fezouata Shale reveals an unexpectedly diverse assemblage comprising a Castericystis-like syringocrinid and at least two minervaecystids (N. agterbosi and P. mobilis) in the Lower Ordovician of the Anti-Atlas (Morocco). The Castericystis-like solutan represents the first evidence of exceptionally preserved soft tissues (part of the digestive system) in this class. The description of Nimchacystis fills the morphological gap between Minervaecystis and Plasiacystis, thus supporting their placement within the same family (Minervaecystidae), along with Pahvanticystis. The abundant and well-preserved material of P. mobilis makes it possible to describe several previously unknown or poorly documented morphological features of this taxon. The new Moroccan material also provides new insights into the palaeobiogeographic dispersion of minervaecystids from Laurentia to high-latitude Gondwana and, subsequently, to Avalonia, as well as their presumed mode of life as active detritus feeders crawling backwards on soft substrates.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
CD: Data curation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. BL: Conceptualization, Data curation, Investigation, Methodology, Supervision, Visualization, Writing – original draft, Writing – review & editing. CM: Writing – review & editing. RM: Writing – review & editing. MN: Methodology, Supervision, Writing – review & editing. RR: Data curation, Methodology, Writing – review & editing. FS: Writing – review & editing, Funding acquisition. SZ: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by Faculty of Geoscience and Environment of the University of Lausanne and the SNF Ambizione grant number PZ00P2_209102.
Acknowledgments
This paper is a contribution to IGCP 735 “Rocks and the Rise of Ordovician Life” (Rocks n’ROL) and to the ANR project “Evolution of the Cambrian-Ordovician Biodiversification Onset Over Space and Time” (ECO-BOOST). FS thanks funding from the Faculty of Geoscience and Environment of the University of Lausanne and the SNF Ambizione grant number PZ00P2_209102. MN was supported by an internal grant from the Czech Geological Survey no. 311410, which is a contribution to the Strategic Research Plan of the Czech Geological Survey (DKRVO/CGS 2023-2027). RM was supported through NSF grant 2036298. SZ was supported by the Spanish Ministry of Science and Innovation (PID2021-125585NB-100), co-financed by the European Regional Development Fund, project “Aragosaurus: Recursos Geológicos y Paleoambientales” (E18_17R) funded by the Government of Aragón. The authors are especially grateful to Marine Fau and Loïc Villier for their comments on an early draft of this paper, as well as to the two reviewers for their careful and insightful reviews. Ton Agterbos, Thierry Castel, Patrick Catto, Vaáclav Micka, Myriam Prost, Roland and Véronique Reboul, Daniel Vizcaïno, Andries Weug, and Ondřej Zicha are warmly thanked for having collected, provided access to, and/or photographed important specimens of solutans from the Lower Ordovician of Morocco. The authors would also like to thank Didier Berthet (Musée des Confluences, Lyon), Denis Demarque, and Serge Régnault (Natural History Museum, Nantes), Khadija El Hariri (Cadi Ayyad University, Marrakech), Tim Ewin (Natural History Museum, London), Yves Laurent, and Pierre Dalous (Natural History Museum, Toulouse), Stéphane Jouve, Anne Médard, and Vincent Poncet (Natural History Museum, Marseille), Rudy Lerosey-Aubril (Senckenberg Museum, Frankfurt), Abel Prieur and Emmanuel Robert (Lyon 1 University, Villeurbanne), and Jessica Utrup (Yale Peabody Museum, New Haven) for access to specimens housed in the public collections over which they have oversight. Vincent Perrier provided invaluable help with photographs.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors BL and FS declared that they were editorial board members of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ausich W. I. (2001). “Echinoderm taphonomy,” in Echinoderm Studies. Eds. Jangoux M., Lawrence J. M. (Rotterdam: Balkema), 171–227.
Bather F. A. (1913). Caradocian cystidea from Girvan. Trans. R. Soc. Edinburgh 49, 359–529. doi: 10.1017/S0080456800003999
Botting J. P. (2016). Diversity and ecology of sponges in the Early Ordovician Fezouata Biota, Morocco. Palaeogeography Palaeoclimatology Palaeoecol. 460, 75–86. doi: 10.1016/j.palaeo.2016.05.018
Botting J. P., Muir L. A., Lefebvre B. (2013). Echinoderm diversity and environmental distribution in the Ordovician of the Builth Inlier, Wales. Palaios 28, 293–304. doi: 10.2110/palo.2012.p12-118r
Brett C. E., Moffat H. A., Taylor W. (1997). Echinoderm taphonomy, taphofacies, and Lagerstätten. In Waters, J. A., and Maples, C. G. (eds), Geobiology of Echinoderms. Paleontological Soc. Papers 3, 147–190. doi: 10.1017/S1089332600000243
Caster K. E. (1968). “Homoiostelea,” in Treatise on Invertebrate Paleontology, part S, Echinodermata, vol. 1 . Ed. Moore R. C. (Boulder: Colorado for Goelogical Society of America Lawrence, Kansas for the University of Kansas Press), S581–S627.
Daley P. E. J. (1995). Anatomy, locomotion and ontogeny of the solute Castericystis vali from the Middle Cambrian of Utah. Geobios 28, 585–615. doi: 10.1016/S0016-6995(95)80214-2
Daley P. E. J. (1996). The first solute which is attached as an adult: a Mid-Cambrian fossil from Utah with echinoderm and chordate affinities. Zoological J. Linn. Soc. 117, 405–440. doi: 10.1111/j.1096-3642.1996.tb01659.x
David B., Lefebvre B., Mooi R., Parsley R. (2000). Are homalozoans echinoderms? An answer from the extraxial-axial theory. Paleobiology 26, 529–555. doi: 10.1666/0094-8373(2000)026<0529:AHEAAF>2.0.CO;2
Dehm R. (1934). Untersuchungen an Cystoideen des rheinischen Unterdevons. Sitzungsberichte Bayerischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Abteilung 1934, 19–43.
Destombes J. (2006). Carte géologique au 1/200 000 de l’Anti-Atlas marocain. Notice explicative. Paléozoïque inférieur : Cambrien moyen et supérieur – Ordovicien – Base du Silurien. Feuille Tafilalt – Taouz. Chapitre E. Notes Mémoires du Service géologique du Maroc 244bis, 1–69.
Destombes J., Hollard H., Willefert S. (1985). “Lower Palaeozoic rocks of Morocco,” in Lower Palaeozoic Rocks of the World. 4. Lower Palaeozoic of North Western - Central Africa. Ed. Holland C. H. (New York: Wiley), 91–336.
Donovan S. K. (1991). “The taphonomy of echinoderms: calcareous multi-element skeletons in the marine environment,” in The processes of fossilization. Ed. Donovan S. K. (London: Belhaven Press), 241–269.
Dupichaud C., Lefebvre B. (2022). “Solutan echinoderms from the Fezouata Shale Lagerstätte (Lower Ordovician, Morocco): diversity, exceptional preservation and palaeobiogeographic implications,” in Abstract book, Second Annual Meeting of IGCP 735, Cadi Ayyad University, Marrakesh, October 19-20 2022. 23–28.
Dupichaud C., Lefebvre B., Nohejlová M. (2023). Solutan echinoderms from the Lower Ordovician of the Montagne Noire (France): new data and palaeobiogeographic implications. Estonian J. Earth Sci. 72, 26–29. doi: 10.3176/earth.2023.80
Ebbestad J. O. R. (2016). Tergomya and Paragastropoda (Mollusca) from the Lower Ordovician Fezouata Formation. Palaeogeography Palaeoclimatology Palaeoecol. 460, 87–96. doi: 10.1016/j.palaeo.2016.01.003
Fone W. (2003). “The first occurence of the mitrate Promitrocystites barrandei (Jaekel 1918) in Great Britain and some questions it raises,” in Programme and abstracts, The Palaeontological Association, 47th Annual Meeting, Leicester, 14th-17th December. 59–60.
Gorzelak P., Salamon M. A. (2013). Experimental tumbling of echinoderms—taphonomic patterns and implications. Palaeogeography Palaeoclimatology Palaeoecol. 386, 569–574. doi: 10.1016/j.palaeo.2013.06.023
Gorzelak P., Zamora S. (2016). Understanding form and function of the stem in early flattened echinoderms (pleurocystitids) using a microstructural approach. PeerJ 4 (e1820), 1–10. doi: 10.7717/peerj.1820
Havlíček V. (1971). Brachiopodes de l'Ordovicien du Maroc. Notes Mémoires du Service Géologique du Maroc 230, 1–135.
Hunter A. W., Lefebvre B., Nardin E., Régnault S., Van Roy P., Zamora S. (2010). “Preliminary report on echinoderm Lagerstätten from the Upper Ordovician of the eastern Anti-Atlas, Morocco,” in Echinoderms: Durham. Eds. Harris L. G., Böttger S. A., Walker C. W., Lesser M. P. (Rotterdam: Balkema), 23–30.
Jaekel O. (1901). Ueber Carpoideen; eine neue Classe von Pelmatozoen. Z. der Deutschen Geologischen Gesellschaft 52, 661–677.
Jaekel O. (1918). Phylogenie und System der Pelmatozoen. Paläontologische Z. 3, 1–124. doi: 10.1007/BF03190413
Jefferies R. P. S. (1987). “The chordates - a preliminary note,” in Bulletin of the British Museum (Natural History) Geology 41, 285–290.
Kerr T. J. V., Twitchett R. J. (2004). “Experimental decay and disarticulation of Ophiura texturata: implications for the fossil record of ophiuroids,” in Echinoderms: München. Eds. Heinzeller T., Nebelsick J. H. (Rotterdam: Balkema), 439–446.
Kidwell S. M., Baumiller T. (1990). Experimental disintegration of regular echinoids: roles of temperature, oxygen, and decay thresholds. Paleobiology 16, 247–271. doi: 10.1017/S0094837300009982
Klein J. T. (1778). Naturalis Dispositio Echinodermatum. Accessit Lucubratiuncula de Aculeis Echinorum Marinorum et Specilegium de Belemnitis. Edita et Descriptionibus Novisque Inventis et Synonymis Auctorum Auca a Nathanaele Godofredo Leske (Leipzig: Officina Gleditdchiana), 278. pp.
Kouraïss K., El Hariri K., El Albani A., Azizi A., Mazurier A., Lefebvre B. (2019). Digitization of fossils from the Fezouata Biota (Lower Ordovician, Morocco): evaluating computed tomography and photogrammetry in collections enhancement. Geoheritage 11, 1889–1901. doi: 10.1007/s12371-019-00403-z
Kröger B., Lefebvre B. (2012). Palaeogeography and palaeoecology of early Floian (Lower Ordovician) cephalopods from the Upper Fezouata Formation, Anti-Atlas, Morocco. Fossil Rec. 15, 61–75. doi: 10.1002/mmng.201200004
Lefebvre B. (2007). Early Palaeozoic palaeobiogeography and palaeoecology of stylophoran echinoderms. Palaeogeography Palaeoclimatology Palaeoecol. 245, 156–199. doi: 10.1016/j.palaeo.2006.02.021
Lefebvre B., Allaire N., Guensburg T. E., Hunter A. W., Kouraïss K., Martin E. L. O., et al. (2016). Palaeoecological aspects of the diversification of echinoderms in the Lower Ordovician of central Anti-Atlas, Morocco. Palaeogeography Palaeoclimatology Palaeoecol. 460, 97–121. doi: 10.1016/j.palaeo.2016.02.039
Lefebvre B., Derstler K., Sumrall C. D. (2012). A reinterpretation of the solutan Plasiacystis mobilis (Echinodermata) from the Middle Ordovician of Bohemia. Zoosymposia 7, 287–306. doi: 10.11646/zoosymposia.7.1.27
Lefebvre B., Fatka O. (2003). Palaeogeographical and palaeoecological aspects of the Cambro-Ordovician radiation of echinoderms in Gondwanan Africa and peri-Gondwanan Europe. Palaeogeography Palaeoclimatology Palaeoecol. 195, 73–97. doi: 10.1016/S0031-0182(03)00303-1
Lefebvre B., Guensburg T. E., Martin E. L. O., Mooi R., Nardin E., Nohejlová M., et al. (2019). Exceptionally preserved soft parts in fossils from the Lower Ordovician of Morocco clarify stylophoran affinities within basal deuterostomes. Geobios 52, 27–36. doi: 10.1016/j.geobios.2018.11.001
Lefebvre B., Gutiérrez-Marco J. C., Lehnert O., Martin E. L. O., Nowak H., Akodad M., et al. (2018). Age calibration of the Lower Ordovician Fezouata Lagerstätte, Morocco. Lethaia 51, 296–311. doi: 10.1111/let.12240
Lefebvre B., Lerosey-Aubril R. (2018). Laurentian origin of solutan echinoderms: new evidence from the Guzhangian (Cambrian Series 3) Weeks Formation of Utah, USA. Geological Magazine 155, 1190–1204. doi: 10.1017/S0016756817000152
Lefebvre B., Nohejlová M., Martin E. L. O., Kašička L., Zicha O., Gutiérrez-Marco J. C. (2022). “New Middle and Late Ordovician cornute stylophorans (Echinodermata) from Morocco and other peri-Gondwanan areas,” in The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco, vol. 485 . Eds. Hunter A. W., Álvaro J. J., Lefebvre B., Van Roy P., Zamora S. (London: The Geological Society, London, Special Publications), 345–522.
Lefebvre B., Sumrall C. D., Shroat-Lewis R. A., Reich M., Webster G. D., Hunter A. W., et al. (2013). “Palaeobiogeography of Ordovician echinoderms,” in Early Palaeozoic Biogeography and Palaeogeography, vol. 38 . Eds. Harper D. A. T., Servais T. (London, Memoirs: Geological Society), 173–198.
Loi A., Dabard M. P. (1999). Stratigraphic significance of siliceous-argilaceous nodules in Ordovician formations of the Armorican massif (France) and Sardinia (Italy). Acta Universitatis Carolinae Geologica 43, 89–92.
Loi A., Dabard M. P. (2002). Controls of sea-level fluctuations on the formation of Ordovician siliceous nodules in terrigeneous offshore environments. Sedimentary Geology 153, 65–84. doi: 10.1016/S0037-0738(02)00102-1
Marek L. (1983). The Ordovician hyoliths of anti-atlas (Morocco). Sborník Národního Musea v Praze 39, 1–36.
Martin E. L. O., Vidal M., Vizcaïno D., Vaucher R., Sansjofre P., Lefebvre B., et al. (2016). Biostratigraphic and palaeoenvironmental controls on the trilobite associations from the Lower Ordovician Fezouata Shale of the central Anti-Atlas, Morocco. Palaeogeography Palaeoclimatology Palaeoecol. 460, 142–154. doi: 10.1016/j.palaeo.2016.06.003
Meyer D. L. (1971). Post-mortem disintegration of Recent crinoids and ophiuroids under natural conditions. Geological Soc. America Abstracts Programs 3, 645–646.
Muir L. A., Gutiérrez-Marco J. C. (2023). A new species of the problematic organism Webbyites from the Early Ordovician Fezouata Biota of Morocco. Estonian J. Earth Sci. 72, 74–77. doi: 10.3176/earth.2023.24
Nebelsick J. H. (1995). “Actuopalaeontological investigations on echinoids: the potential for taphonomic interpretation,” in Echinoderm Research 1995. Eds. Emson R. H., Smith A. B., Campbell A. C. (Rotterdam: Balkema), 209–214.
Noailles F., Lefebvre B., Kašička L. (2014). A probable case of heterochrony in the solutan Dendrocystites Barrande 1887 (Echinodermata: Blastozoa) from the Upper Ordovician of the Prague Basin (Czech Republic) and a revision of the family Dendrocystitidae Bassler 1938. Bull. Geosciences 89, 451–476. doi: 10.3140/bull.geosci.1475
Nohejlová M., Fatka O. (2016). Ontogeny and morphology of Cambrian eocrinoid Akadocrinus (Barrandian area, Czech Republic). Bull. Geosciences 91, 141–153. doi: 10.3140/bull.geosci.1583
Nohejlová M., Lefebvre B. (2022). “Late Ordovician solutan echinoderms from the western Tafilalt, Morocco,” in The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco, vol. 485 . Eds. Hunter A. W., Álvaro J. J., Lefebvre B., Van Roy P., Zamora S. (London: The Geological Society, London, Special Publications), 523–539.
Nohejlová M., Lefebvre B., Nardin E., Fatka O., Budil P. (2019). “New echinoderm Lagerstätte from the Letná Formation (Sandbian, Upper Ordovician) of Bohemia,” in Abstracts, 10th European Conference on Echinoderms, Moscow, September 16-19, 2019, Vol. 69.
Parsley R. L. (2012). Ontogeny, functional morphology, and comparative morphology of lower (Stage 4) and basal middle (Stage 5) Cambrian gogiids, Guizhou Province, China. J. Paleontology 86, 569–583. doi: 10.1666/10-153R2.1
Polechová M. (2016). The bivalve fauna from the Fezouata Formation (Lower Ordovician) of Morocco and its significance for palaeobiogeography, palaeoecology and early diversification of bivalves. Palaeogeography Palaeoclimatology Palaeoecol. 460, 155–169. doi: 10.1016/j.palaeo.2015.12.016
Prokop R. V., Petr V. (2003). Plasiacystis mobilis, gen. et sp. n., a strange "carpoid" (Echinodermata, ?Homoiostelea: Soluta) in the Bohemian Ordovician (Czech Republic). Sborník Národního muzea (B: Přírodní vědy) [= Acta Musei Nationalis Pragae (B: Natural History)] 59, 151–162.
Rahman I. A., Lintz H. (2012). Dehmicystis globulus, an enigmatic solute (Echinodermata) from the Lower Devonian Hunsrück Slate, Germany. Paläontologische Z. 86, 59–70. doi: 10.1007/s12542-011-0116-y
Rozhnov S. V. (2022). Solutans (Echinoderms): Evolution frozen between torsion and pentaradiality. Paleontological J. 56, 1306–1321. doi: 10.1134/S0031030122110144
Saleh F., Guenser P., Gibert C., Balseiro D., Serra F., Waisfeld B. G., et al. (2022b). Contrasting Early Ordovician assembly patterns highlight the complex initial stages of the Ordovician Radiation. Sci. Rep. 12 (3852), 1–8.
Saleh F., Pittet B., Perrillat J. P., Lefebvre B. (2019). Orbital control on exceptional fossil preservation. Geology 47 (2), 103–106. doi: 10.1130/G45598.1
Saleh F., Pittet B., Sansjofre P., Guériau P., Lalonde S., Perrillat J. P., et al. (2020). Taphonomic pathway of exceptionally preserved fossils in the Lower Ordovician of Morocco. Geobios 60, 99–115. doi: 10.1016/j.geobios.2020.04.001
Saleh F., Vaucher R., Antcliffe J. B., Daley A. C., El Hariri K., Kouraïss K., et al. (2021). Insights into soft-part preservation from the Early Ordovician Fezouata Biota. Earth Sci. Rev. 213 (103464), 1–12. doi: 10.1016/j.earscirev.2020.103464
Saleh F., Vaucher R., Vidal M., El Hariri K., Laibl L., Daley A. C., et al. (2022a). New fossil assemblages from the Early Ordovician Fezouata Biota. Sci. Rep. 12 (20773), 1–10. doi: 10.1038/s41598-022-25000-z
Smith A. B. (1988). Patterns of diversification and extinction in Early Palaeozoic echinoderms. Palaeontology 31, 799–828.
Smith A. B. (2005). The pre-radial history of echinoderms. Geological J. 40, 255–280. doi: 10.1002/gj.1018
Sprinkle J., Guensburg T. E. (1993). “Echinoderm biostratigraphy,” in Paleozoic Biochronology of the Great Basin, Western United States. Open-File Report, vol. 598 . Ed. Taylor M. E. (New York, Oklahoma City: US Geological Survey Open-File Report), 61–63.
Sprinkle J., Guensburg T. E. (1997). “Early radiation of echinoderms,” in Geobiology of Echinoderms The Paleontological Society Papers, vol. 3 . Eds. Waters J. A., Maples C. G., 205–224.
Sprinkle J., Guensburg T. E. (2004). “Crinozoan, blastozoan, echinozoan, asterozoan, and homalozoan echinoderms,” in The Great Ordovician Biodiversification Event. Eds. Webby B. D., Paris F., Droser M. L., Percival I. G. (Columbia University Press), 266–280.
Sumrall C. D., Sprinkle J., Guensburg T. E. (1997). Systematics and paleoecology of Late Cambrian echinoderms from the western United States. J. Paleontology 71, 1091–1109. doi: 10.1017/S0022336000036052
Thoral M. (1935). Contribution à l’étude Paléontologique de l’Ordovicien inférieur de la Montagne Noire et Révision Sommaire de la Faune Cambrienne de la Montagne Noire (Montpellier: Imprimerie de la Charité), 362. p.
Ubaghs G. (1963). Cothurnocystis Bather, Phyllocystis Thoral and an undetermined member of the order Soluta (Echinodermata, Carpoidea) in the uppermost Cambrian of Nevada. J. Paleontology 37, 1133–1142.
Ubaghs G. (1970). Les Echinodermes «Carpoïdes» de l'Ordovicien inférieur de la Montagne Noire (France) (Paris: Editions du CNRS, Cahiers de Paléontologie), 110. p.
Ubaghs G., Robison R. A. (1985). A new homoiostelean and a new eocrinoid from the Middle Cambrian of Utah. Univ. Kansas Paleontological Contributions 115, 1–24.
Ubaghs G., Robison R. A. (1988). Homalozoan echinoderms of the Wheeler Formation (Middle Cambrian) of western Utah. Univ. Kansas Paleontological Contributions 120, 1–17.
Vaucher R., Pittet B., Martin E. L. O., Lefebvre B., Hormière H. (2017). A wave-dominated tide-modulated model for the Lower Ordovician of the Anti-Atlas. Sedimentology 64, 777–807. doi: 10.1111/sed.12327
Zamora S., Deline B., Álvaro J. J., Rahman I. A. (2017). The Cambrian Substrate Revolution and the early evolution of attachment in suspension-feeding echinoderms. Earth-Science Rev. 171, 478–491. doi: 10.1016/j.earscirev.2017.06.018
Zamora S., Gutiérrez-Marco J. C. (2023). Filling the Silurian gap of solutan echinoderms with the description of new species of Dehmicystis from Spain. Acta Palaeontologica Polonica 68, 185–192. doi: 10.4202/app.01054.2023
Zamora S., Lefebvre B., Álvaro J. J., Clausen S., Elicki O., Fatka O., et al. (2013). “Cambrian echinoderm diversity and palaeobiogeography,” in Early Palaeozoic Biogeography and Palaeogeography, vol. 38. (London: Geological Society, London, Memoirs), 157–171.
Keywords: Echinodermata, Soluta, Fezouata Shale, Morocco, Ordovician, systematics
Citation: Dupichaud C, Lefebvre B, Milne CH, Mooi R, Nohejlová M, Roch R, Saleh F and Zamora S (2023) Solutan echinoderms from the Fezouata Shale Lagerstätte (Lower Ordovician, Morocco): diversity, exceptional preservation, and palaeoecological implications. Front. Ecol. Evol. 11:1290063. doi: 10.3389/fevo.2023.1290063
Received: 06 September 2023; Accepted: 04 October 2023;
Published: 23 October 2023.
Edited by:
Haijun Song, China University of Geosciences Wuhan, ChinaReviewed by:
Przemyslaw Gorzelak, Polish Academy of Sciences, PolandJames Thomka, State University of New York College at Plattsburgh, United States
Copyright © 2023 Dupichaud, Lefebvre, Milne, Mooi, Nohejlová, Roch, Saleh and Zamora. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christophe Dupichaud, c.dupichaud@gmail.com; Farid Saleh, farid.saleh@unil.ch