
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 07 December 2023
Sec. Conservation and Restoration Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1254462
This article is part of the Research TopicLarge-Scale Dam Removal and Ecosystem RestorationView all 25 articles
 Olivier Dézerald1*†
Olivier Dézerald1*† Jean-Marc Roussel1†
Jean-Marc Roussel1† Elven Lanoë1
Elven Lanoë1 Thibaut Beauverger2,3Alan Bazin2,3Héctor Rodríguez-Pérez4,5Simon Dufour6†
Thibaut Beauverger2,3Alan Bazin2,3Héctor Rodríguez-Pérez4,5Simon Dufour6† Ivan Bernez1†
Ivan Bernez1† Christophe Piscart4†
Christophe Piscart4†Introduction: Ecological restoration through dam removals receives increasing attention from scientists, environmental managers and policy makers. However, most dam removal projects focus on small structures (< 10 m in height) and on few ecosystem compartments at a time (e.g. river morphology, reservoir sedimentation, aquatic invertebrates, terrestrial vegetation), but rarely aquatic and riparian ecosystems simultaneously.
Methods: We explored the joint taxonomic recovery (temporal patterns in α- and β-diversity) of three communities after the removal of the Vezins dam (36 m in height; Sélune River, France): aquatic benthic invertebrates, riparian invertebrates, and the riparian vegetation. These communities were monitored yearly, during three years after the dam removal, on sites located within (n = 3; restored sites) and outside (n = 3; two upstream and one downstream; non-impounded sites) the former reservoir.
Results: Results showed a fast recovery of the three ecological communities, as patterns in α-diversity between restored and non-impounded sites were not necessarily different from each other 1.5 years after complete reservoir dewatering. The mean number of species or morphospecies (± standard deviation) reached up to 28.2 ± 5.2, 17.0 ± 2.3 and 77.5 ± 11.2 for the aquatic invertebrates, and the riparian vegetation and invertebrates, respectively. Relative to the sampled area, the riparian invertebrates were the most diversified of all ecological communities with > 500 taxa (i.e., pooling all sites and years). In addition, in some restored sites, α-diversity kept increasing over time while species turnover (β-diversity) remained high after three years for all ecological communities suggesting a transient recovery (i.e., still facing temporal changes in species diversity and composition). This recovery was mediated by the identity of the ecological community as inter-annual changes in α- and β-diversity of the riparian vegetation were less pronounced compared to those of aquatic and terrestrial invertebrates. This recovery depended also on site-specific features as the most distant restored site from the former dam had more time for recovery following the slow reservoir dewatering and through increased sedimentation in the downstream site.
Discussion: Differential patterns of recovery in α- and β-diversity found in this study are discussed in light of species functional traits and ecosystem functioning.
Increasing societal concerns towards biodiversity and water has shed new light on a widespread but overlooked driver of rivers’ ecosystem functioning: the barriers to free flow. Barriers to free flow have played an important role in modern human history to control water for irrigating crops, drinking water, generating hydropower and preventing floods. Recent estimates, most likely far below reality, point to at least 1.6 million instream barriers in Europe (Belletti et al., 2020), and only a few remaining very long and free-flowing rivers worldwide (Grill et al., 2019). In addition, although most of those barriers are less than a couple of centuries old, many began to be outdated with unsafe structures and low benefit-cost ratios. Consequently, those barriers, mainly dams, are frequently dismantled, thus drawing attention from the scientific community (Hart et al., 2002; O’Connor et al., 2015; Ryan Bellmore et al., 2017; Habel et al., 2020) and leading to the emergence of a new field in restoration ecology: “the science of dam removal” (Poff and Hart, 2002).
Dam removal projects offer unique opportunities to explore the ecological mechanisms underlying the recovery of rivers after the long-term press disturbances induced by dams. The ecological consequences of dam removals on habitat fragmentation through longitudinal processes are well-documented, with particular emphasis on fish and aquatic macroinvertebrate communities. For instance, fish communities rapidly recolonize upstream areas following small dam removals (Hogg et al., 2015; Kornis et al., 2015; Bubb et al., 2021), although communities may not necessarily recover immediately after the removal (< 3 years; Catalano et al., 2007; Stanley et al., 2007; Poulos and Chernoff, 2017). Long-term trends also agree on overall positive effects of river restoration on fish populations (Birnie-Gauvin et al., 2017) and fish communities (Whittum et al., 2023). Aquatic macroinvertebrate communities show more mitigated responses to dam removal compared to fish with first decreases in diversity, most likely due to high downstream sedimentation (Chiu et al., 2013; Renöfält et al., 2013; Rubin et al., 2017), followed by increases in diversity over time (Orr et al., 2008; Kil and Bae, 2012; Mahan et al., 2021). Yet, macroinvertebrate communities may not recover completely even after three years following dam removals (Hansen and Hayes, 2012; Renöfält et al., 2013; Poulos et al., 2019; Mahan et al., 2021). In addition, increasing evidence supports strong influence of taxa identity (Sethi et al., 2004; Tszydel et al., 2009), seasonality (Sullivan and Manning, 2017), geomorphic features (Tullos et al., 2014) and habitat characteristics (Pollard and Reed, 2004; Claeson and Coffin, 2016; Poulos et al., 2019) on patterns of macroinvertebrate recovery in stream following dam removal.
Removing dams not only affect aquatic habitats longitudinally but also terrestrial habitats through both longitudinal and lateral processes. For instance, recent evidence supports strong longitudinal patterns in riparian vegetation communities following dam removals (Brown et al., 2022) with pioneer plant species tracking changes in geomorphic and hydrologic features in downstream new coastal habitats (Foley et al., 2017) and in newly dewatered habitats closest to the river channel (Lisius et al., 2018; Ravot et al., 2020). A study on an exposed reservoir due to dam maintenance confirms an important shift in species composition from mesic to xeric plant communities within less than four years (Auble et al., 2007). In restored sites, plant recovery was usually fast (Laslier et al., 2019) and was positively affected by mycorrhizal and ectomycorrhizal fungi (Cortese and Bunn, 2017) while being threatened by non-native and competitive plant species (Orr and Stanley, 2006; Tullos et al., 2016). In turn, only few studies have investigated the effects of dam removals on the terrestrial fauna (e.g., megafauna, birds; Stephens, 2017; McCaffery et al., 2018; McCaffery et al., 2020). Concerning macroinvertebrates, some evidence suggest strong declines in spiders within four years post-removal (Sullivan et al., 2018). Altogether, the primary literature on dam removal highlights three important limitations to our understanding of mechanisms at play during community recovery. First, most studies focus on local processes following the removal of small dams (< 10 m in height; but see Foley et al., 2017). More specifically, compared to small dams, tall dams may have stronger impacts on the hydromorphological features (e.g., increased sediment load in impounded sites, large spatial extent of influence) of rivers, and thus may trigger lagged responses and longer recovery of ecological communities. Second, most studies focus on one ecological community (e.g. aquatic invertebrates, terrestrial vegetation), mainly aquatic ones (Doyle et al., 2005; Carlson et al., 2018), while investigating multiple ecological communities should provide more accurate responses of ecosystems to dam removals (Thomson et al., 2005; Chang et al., 2017; Cook and Sullivan, 2018; Atristain et al., 2023). Third, we lack an integrated view of spatial (longitudinal and lateral processes) and temporal (inter-annual processes) patterns of recovery. It is particularly difficult to find appropriate control sites when comparing restoration processes due to high historical contingencies and ecological idiosyncrasies. In addition, very often we lack information on the state of ecosystems before removals. We therefore advocate to move further away from restored vs. control and before vs. after treatment designs.
Changes in α- and β-diversity, i.e., the spatial variations in local and regional species composition (Whittaker, 1965), offers unparalleled insights into the mechanisms (i.e., species sorting, species competition, dispersal limitation) supporting meta-community dynamics (López-Delgado et al., 2020). These changes can also inform biodiversity-based conservation decisions by identifying sink and source populations of colonizers for enhanced recovery (Socolar et al., 2016). Therefore, exploring temporal changes in α- and β-diversity of multiple ecological communities following a dam removal may then help dissecting spatial-temporal patterns of recovery. In the present study, we aimed to address all three above-mentioned limitations from dam removal studies. We explored temporal changes in patterns of α- and β-diversity in both terrestrial and aquatic environments by monitoring multiple ecological communities (i.e., aquatic benthic invertebrates, riparian invertebrates, and the riparian vegetation) from one to three years after the removal of the tall Vezins dam (36 m in height; Sélune river, France). The sampling sites, both within and outside the former reservoir, were selected to reflect a longitudinal upstream-to-downstream gradient along the river thus moving further away from the common restored vs. control study design. In addition, since some study sites were previously under water (i.e., precluding meaningful before vs. after comparisons), we started the monitoring right after the dam removal thus moving away from a before vs. after study design. Based on knowledge gained from previous studies, we specifically tested two main hypotheses. Our first hypothesis (H1) states that taxonomic recovery is driven by rapid inter-annual increase in α-diversity in restored sites. Indeed, evidence shows that the diversity of invertebrate and plant communities can increase in just a few month up to less than four years even though transient decreases can be observed right after the dam removal or dewatering (Auble et al., 2007; Foley et al., 2017; Carlson et al., 2018). Our second hypothesis (H2) stipulates that β-diversity remain high between distant sites with gradual changes in species composition from upstream to downstream sites and across years of sampling. This second hypothesis follows a pattern found in a companion study using the same ecosystems but occurring before the dams were removed (Rodríguez-Pérez et al., 2021). Last, we briefly discussed how spatial-temporal patterns in α- and β-diversity can be community-dependent (e.g., invertebrates vs. vegetation, aquatic vs. terrestrial) due to their conspicuous differences in life history traits, which integrate environmental changes over different time scales (Southwood, 1977; Townsend and Hildrew, 1994; Doyle et al., 2005).
The Sélune is a 91 km-long river located in northwest France, it drains a watershed of 1,106 km2, and flows into the Mont Saint Michel bay (Figure 1A; Rodríguez-Pérez et al., 2021). The climate is temperate oceanic with a mean annual precipitation of 800 mm and a mean annual temperature of 10°C. The Sélune river flows across a patchy landscape, mainly composed of crops, pastures and livestock farming, with the presence of a few forested and urbanized areas. Two hydropower dams, the Roche-qui-Boit (built in 1919; 16 m in height) and the Vezins (built in 1932; 36 m in height) were located at 26 km and 30 km from the Sélune’s mouth, respectively (Figure 1). The emptying of the Vezins’ reservoir started in May 2017 and was complete in May 2019 (Figures 1B–E). Note that the reservoir was filled again in October/November 2018 after decisions related to water and sediment management issues during the winter period (Figure 1F). The dismantling of the Vezins dam started in 2019 and ended in late 2020 while the Roche-Qui-Boit dam was completely removed in 2023 after gradual decreases in water levels (i.e., free flowing water since June 2022; Figure 1F).
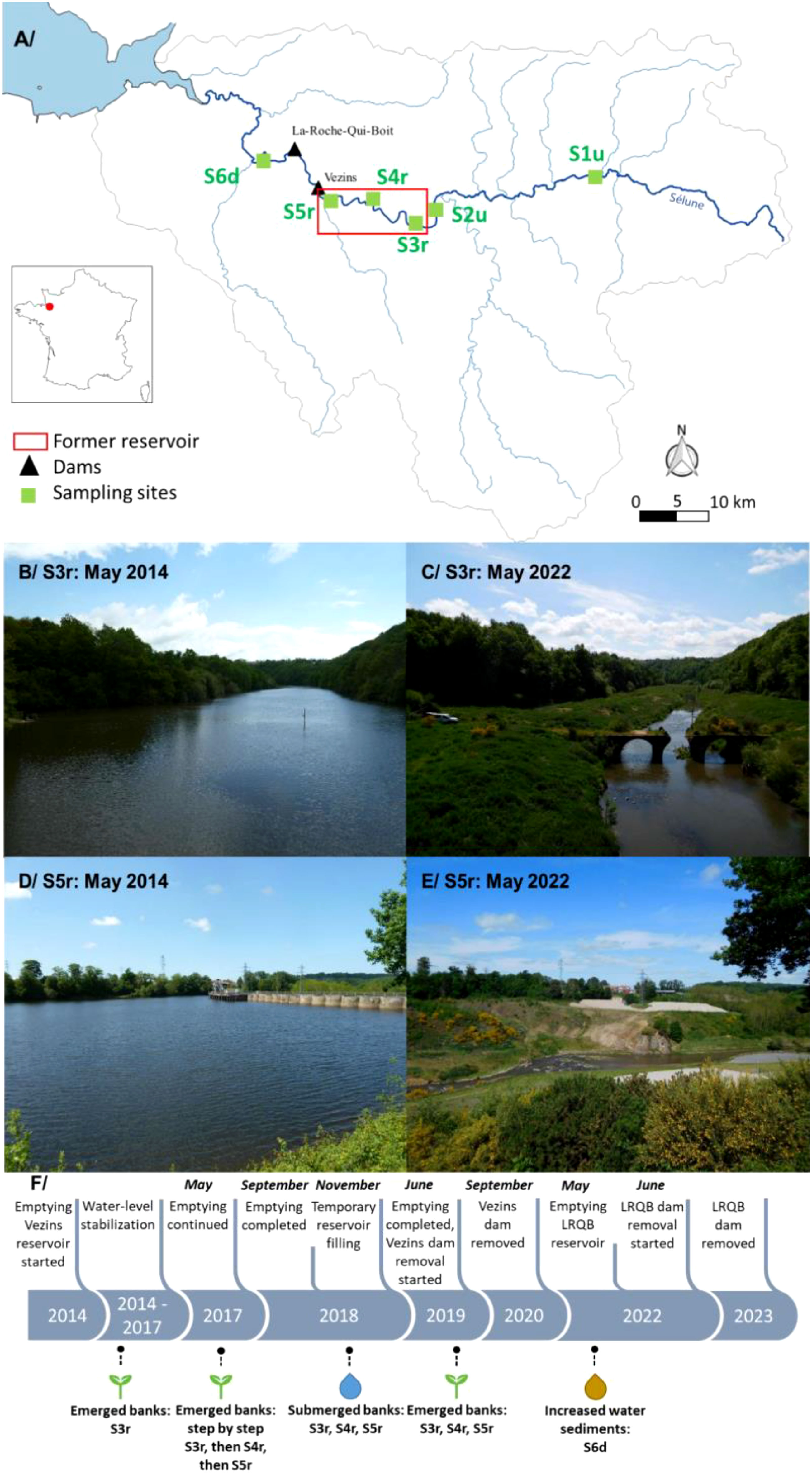
Figure 1 Environmental context of the Sélune river. (A) spatial locations of sampling sites. Pictures displaying the effect of the dam removal in S3r (B, C) and S5r (D, E) between 2014 (B, D) and 2022 (C, E). (F) Timeline of events related to the removal of the Vezins and La-Roche-Qui-Boit (LRQB) dams. Photos credits: Observatoire photographique des paysages de la Sélune - Université Paris Nanterre et SMBS.
Six sampling sites were selected and monitored once a year in late spring/beginning of Summer for three years (i.e., 2020, 2021, and 2022) after the end of the reservoir emptying (spring 2019). This sampling period was chosen, assuming that it maximizes peaks of species richness and abundances of all studied ecological communities (i.e., plants and animals, aquatic and terrestrial; Sullivan and Manning, 2017). Two sites were located upstream the former Vezins’ reservoir: S1u (latitude: 48.599043; longitude: -0.958010083) and S2u (latitude: 48.57171721; longitude: -1.116205727; Figure 1). Three restored sites were located within the former dam reservoir: S3r (latitude: 48.56789852; longitude: -1.146223939), S4r (latitude: 48.57532161; longitude: -1.178982545), and S5r (latitude: 48.57576597; longitude: -1.221868824). The last site was located downstream all dams: S6d (latitude: 48.59570284; longitude: -1.2938635). The sites S3r and S5r were located in the shallowest and deepest part of the former reservoir, while S4r was at intermediate depth. Beside constraints in sampling efforts, those sites were selected to reflect potential upstream-downstream patterns in species richness in aquatic communities that could influence the recovery of communities within the former dam reservoir (S3r-S5r). In addition, they were selected because the terrestrial habitats in those sites reflect the typical landscape in the Sélune basin with S1u, S2u and S6d being close to crops, pastures and livestock farms (i.e., intermediate human activities between forest and urban patches) while S3r, S4r and S5r being located in a relatively more steep-sided river valley (i.e., typical of the former dam reservoir) also located nearby crops, pastures and livestock farms. Note, that it may be challenging to select sampling sites in dam removal projects in an attempt to optimize the trade-off between statistical robustness and processing time and costs. This is particularly important as funding opportunities and successes fluctuate over time, while dam removal projects may last for decades (Birnie-Gauvin et al., 2017). We therefore initiated a simple sampling procedure (i.e., six sites sampled once a year at the same period) that could easily be maintained in the long run, especially in cases of funding shortages.
Benthic macroinvertebrates, hereafter ‘aquatic invertebrates’, were sampled using a Surber net sampler (collecting area: 0.05 m2; mesh size: 500 µm). Terrestrial macroinvertebrates, hereafter ‘riparian invertebrates’, were sampled using a suction sampler (13 cm in diameter; mesh size: 500 µm). To mimic the collecting area of the aquatic invertebrates (0.05 m2), four suctions were performed over 10 seconds to constitute one sample of riparian invertebrates. Then six samples of aquatic and riparian invertebrates per site and date were collected to reflect dominant habitats occurring at the sites. Samples were preserved in the field with 96° ethanol and stored until processed in the laboratory. Samples were sorted under binoculars and identified to the lowest taxonomic resolution (i.e., species or morphospecies level), except for some taxa (e.g., nematods, water mites, midges, flies), which were identified at the family or order level. We will hereafter use the terms ‘taxa’ or ‘richness’ or ‘taxonomic richness’, in reference to this diversity of taxonomic resolutions in our database. The sampling of aquatic invertebrates in S1u, S2u and S3r was not done in 2021.
The riparian vegetation was characterized at each site using 18 quadrats (1 x 1 m). For each quadrat, the species’ identities and relative percent cover were determined, in situ, by the same person across all three years of sampling. Quadrats were located either close to (n = 9) or further away from (n = 9; > 15 m depending on field topography) the stream to integrate local heterogeneities in site conditions. Note that terrestrial communities, the riparian vegetation and invertebrates, were all sampled within 20 m from the river channel. The information from 12 quadrats in S1u 2020 were lost while nine quadrats from S6d were not done in 2021. The exact location of quadrats (i.e., riparian vegetation) and collecting areas (i.e., aquatic and terrestrial invertebrates) could vary by just a few meters at most from year to year.
To test for changes in α-diversity across years and sites, we used generalized linear models (GLMs; stats-package) with the taxonomic richness as response variable fit with either Poisson or Negative Binomial families. The year and site identities and two-way interactions were used as predictors. Pairwise comparisons were evaluated using Tukey method (emmeans-package). One model was built for each ecological community (i.e., aquatic invertebrates, riparian invertebrates, riparian vegetation). Models were checked for overdispersion (performance-package), and model fit was graphically evaluated. The normality of residuals was assessed using a Shapiro test (stats-package).
To explore changes in β-diversity across years and sites, we used a permutational multivariate analysis of variance (PERMANOVA; RVAideMemoire-package; Hervé, 2020). The Jaccard dissimilarity index (vegan-package; Oksanen et al., 2019) was calculated on a matrix with the presence/absence of species across years and sites (rows). The year and site identities, and two-way interactions were used as predictors. Since PERMANOVA may fail to appropriately identify potential intra-group variability (e.g., year and site), we performed the analysis of homogeneity of multivariate group dispersions (PERMDISP; vegan-package) with similar model construction as the PERMANOVA. Results from both PERMANOVA and PERMDISP were graphically displayed using Principal COordinates analysis (PCO; ecodist-package; Goslee and Urban, 2007). Since our study design follows a longitudinal upstream-to-downstream gradient rather than a restored vs. control design, the influence of dam removal on α- and β-diversity can be highlighted by significant ‘site:year’ interactions and by further exploring significant pairwise differences across sites (i.e., upstream, restored, downstream) and within restored sites across years following GLM, PERMANOVA, and PERMDISPs analyses. All statistical analyses were done with the R software (R Core Team, 2020).
After three years of sampling and across the six sites, i.e., pooling all years and sites, the riparian invertebrates were the most diversified of all ecological communities with 530 species distributed into 92 families and 29 orders. The riparian vegetation displayed the second highest number of species (n = 158) distributed into 41 families and 23 orders. Last, we found 110 species within aquatic invertebrates with 70 families and 19 orders. Regardless of the year of sampling, any given sampling site displayed between 53 and 69 aquatic species, between 37 and 93 species of plants, and between 236 and 350 species of riparian invertebrates. In the different sites, the number of families varied from 39 to 52, from 17 to 35, and from 64 to 77, aquatic invertebrates, riparian vegetation and riparian invertebrates, respectively. Similarly, the number of orders were in the range 13-17, 11-20 and 20-25 for the aquatic invertebrates, riparian vegetation and riparian invertebrates, respectively. Overall, we collected between one (i.e., multiple taxa) and 13,145 (i.e., Simuliidae spp) aquatic invertebrate specimens (mean ± SD: 455 ± 1,709) and between one (i.e., multiple taxa) and 5,236 (i.e., Entomobryomorpha sp2) terrestrial invertebrate specimens (mean ± SD: 64 ± 294). The number of specimens for the riparian vegetation could not be estimated.
Overall, the mean richness varied (± standard deviation) from 14.2 (± 3.66) to 28.2 (± 5.23), from 3.9 (± 1.41) to 17.0 (± 2.35) and from 23.5 (± 13.8) to 77.5 (± 11.2) for the aquatic invertebrates, and riparian vegetation and invertebrates, respectively (Figure 2). The richness was significantly influenced by the interaction between site and year of sampling for the aquatic and riparian invertebrates (p < 0.0001 and p < 0.0001, respectively; Table 1) but not for the riparian vegetation (p = 0.16; Table 1; Figure 2). However, when the interaction is not included in the riparian vegetation model, the site and year effects became significant (Chi2 = 117.5, p < 0.0001 and Chi2 = 18.4, p < 0.0001, respectively; results not shown).
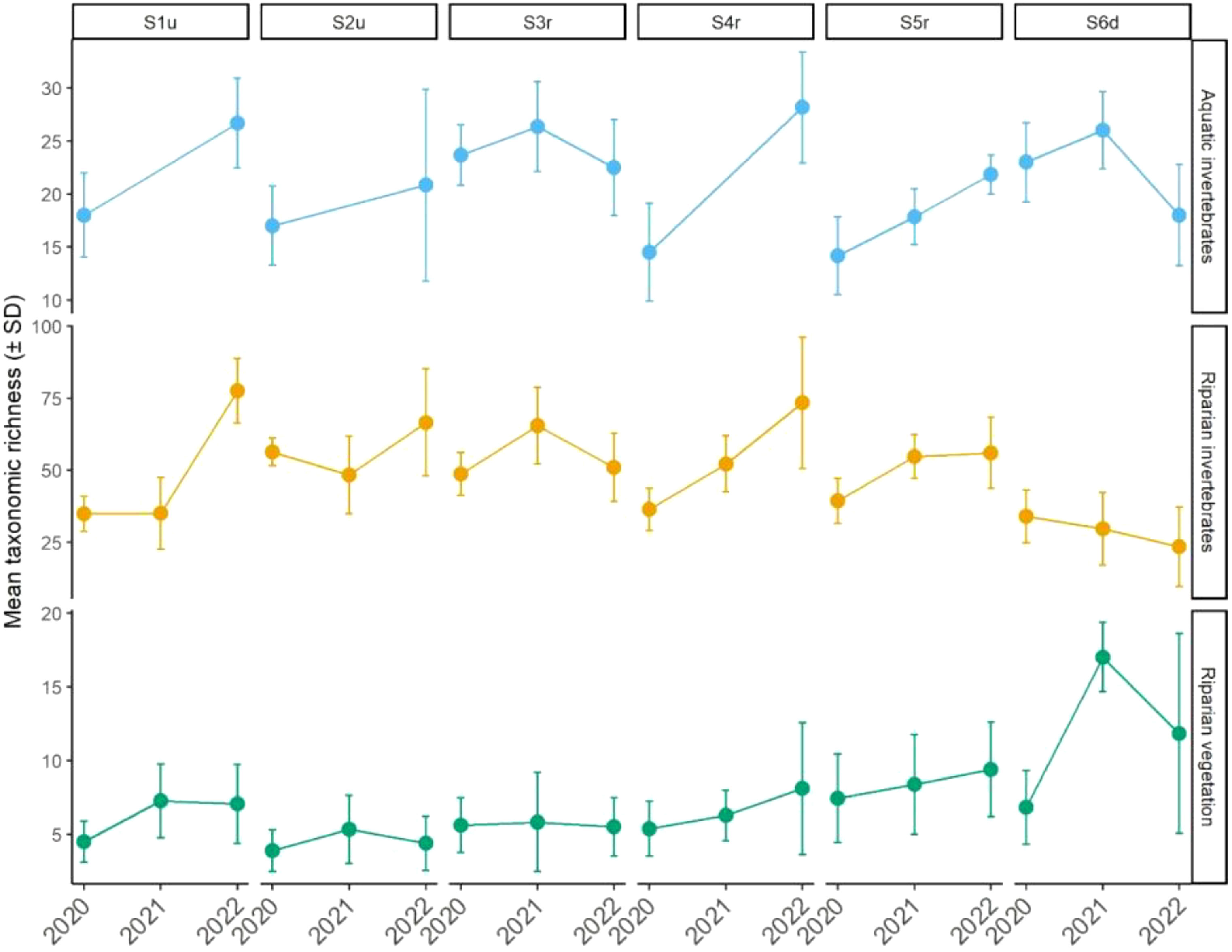
Figure 2 Changes in mean taxonomic richness of the aquatic invertebrates, the riparian vegetation and the riparian invertebrates from upstream to downstream sites and over time (see Table 2 for slope significance).
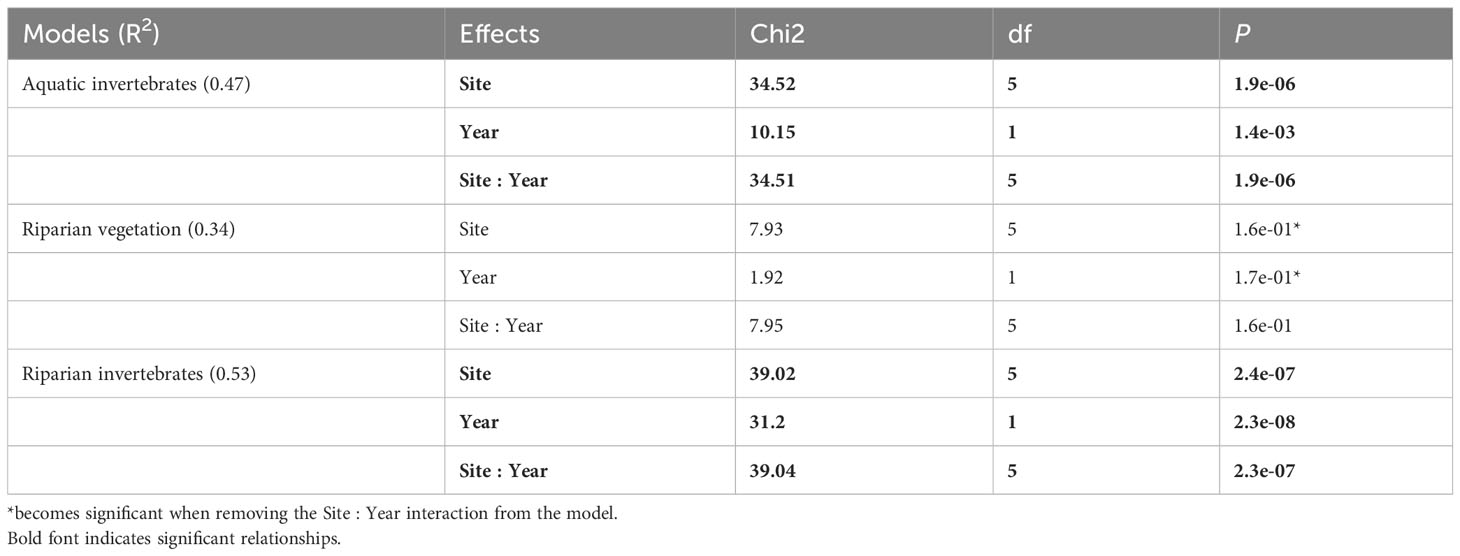
Table 1 Results of GLMs evaluating the effects of the sampling sites, years and two-way interactions on the taxonomic richness.
In 2020, approximately 1.5 years after the beginning of the reservoir dewatering, levels of taxonomic richness of all communities in restored sites (i.e., S3r-S5r) were not significantly different from that of sites located outside the former reservoir (i.e., S1u, S2u, S6d; Table S1). The richness of all ecological communities increased significantly over time in the restored site S4r (Table 2; Figure 2). Similar patterns were found in S5r for the aquatic and riparian invertebrates but the relationship was not significant for the riparian vegetation (Table 2; Figure 2). In the upstream of all restored sites (S3r), none of the communities showed significant changes in richness over time (Table 2; Figure 2). We also found a significant increase in taxonomic richness over time in S1u for the aquatic and riparian invertebrates and in S6d for the riparian vegetation (Table 2). Last, a significant decrease in richness of the riparian invertebrates was found in S6d (Table 2; Figure 2).
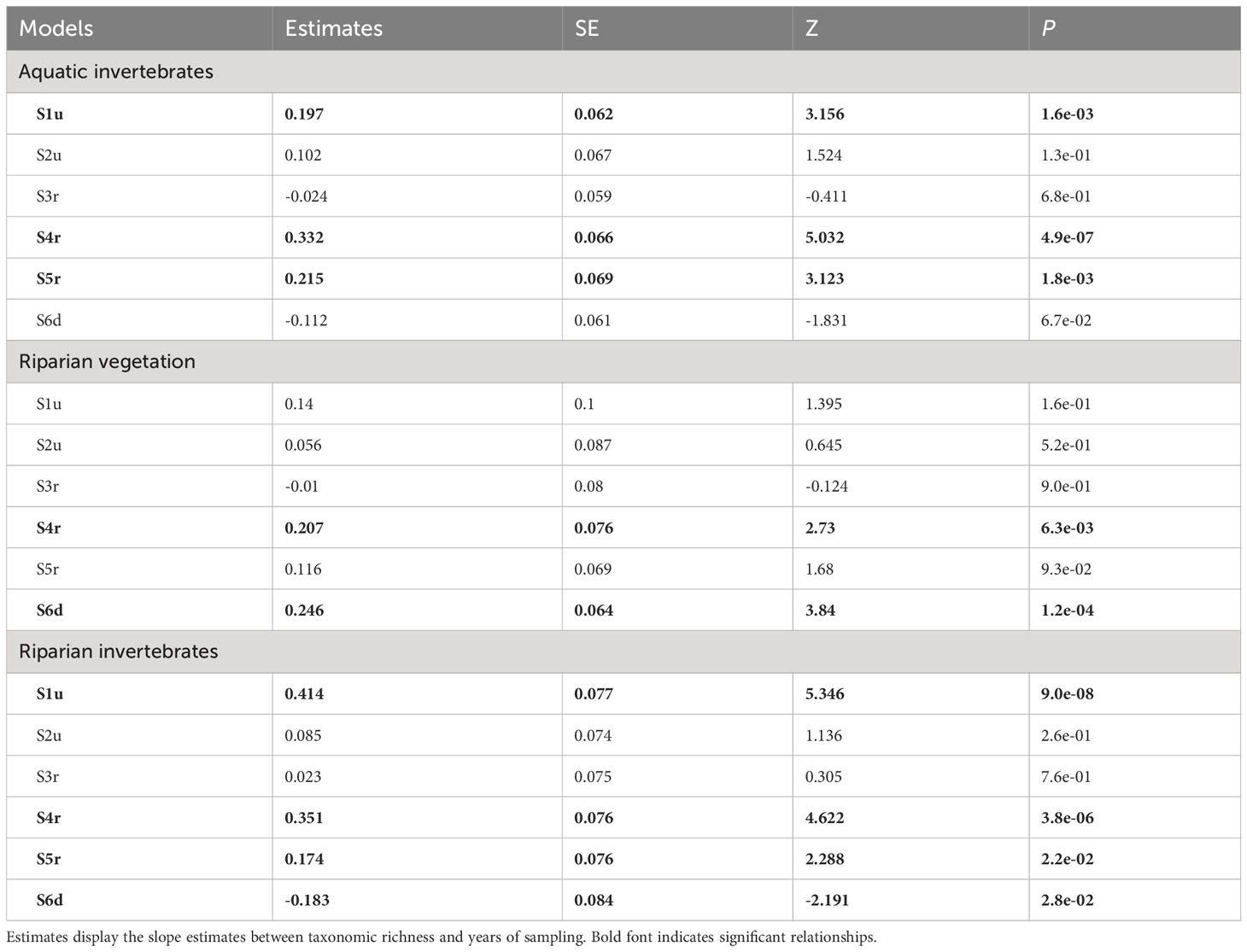
Table 2 Contrasts of GLMs evaluating the effects of the sampling sites, years and two-way interactions on the taxonomic richness of three ecological communities (see Table 1 for associated models and Figure 2 for visualizations).
All three ecological communities (i.e., riparian vegetation, aquatic and riparian invertebrates) showed significant differences in taxonomic turnover (Jaccard’s dissimilarity index on presence/absence data) across sites and years (p < 0.0001; Table 3; Figure 3). Posthoc tests after the PERMANOVA indicates that the taxonomic turnover of each community varied significantly across all years (Figures 3A–C; Table S2) and sites (Figures 3D–F; Table S2), except for the turnover in taxa identity between S2u and S4r of the aquatic invertebrates, which was not significant (Figure 3D; see more details about pairwise comparisons across years and sites in Table S3). The temporal turnover in taxa of all ecological communities indicated that the year 2020 differed from 2021 and 2022 (Figures 3A–C). For the aquatic invertebrates, the average intra-site and intra-year Jaccard dissimilarity in restored sites decreased by 0.053 ± 0.10 between 2020 and 2022 (e.g., pairwise comparisons of all samples within S3r in 2020), while it increased by 0.010 ± 0.12 and 0.17 ± 0.04 for the riparian vegetation and riparian invertebrates respectively (Table S4). In 2022, the average intra-site dissimilarity between three restored sites remained high for all ecological communities: the aquatic invertebrates (0.49 ± 0.06), the riparian vegetation (0.74 ± 0.07), and the riparian invertebrates (0.79 ± 0.05). The average intra-site and inter-year dissimilarity in restored sites (e.g., pairwise comparisons of all samples between S3r - 2020 and S3r - 2022) were the highest and lowest for the riparian (0.88 ± 0.01) and aquatic invertebrates (0.63 ± 0.05), respectively (intermediate values for the riparian vegetation: 0.86 ± 0.10; Table S4). In addition, the first PCoA axis of the riparian vegetation was positively correlated with upstream sites (S1u, S2u and S3r) and negatively with downstream ones (S4r to S6d) suggesting some signal of the river continuum (Figure 3E). The second PCoA axis of the riparian vegetation discriminates the communities found in restored sites (S3r-S5r; negatively correlated with PCoA2) from those found in other sites (S1u, S2u and S6d; positively correlated with PCoA2; Figure 3E).
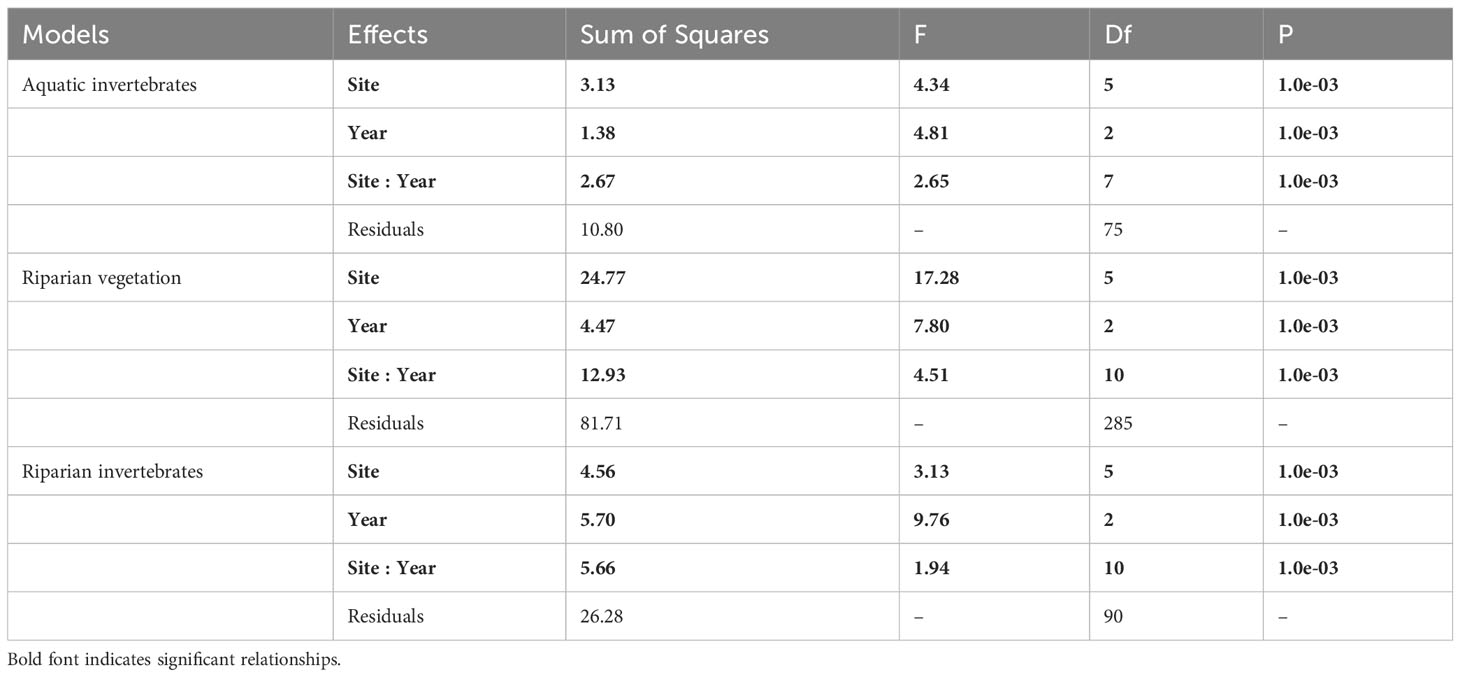
Table 3 Results of PERMANOVAs evaluating the effects of the sampling sites, years and two-way interactions on the pairwise taxonomic dissimilarities (Jaccard’s index on presence/absence matrices).

Figure 3 Principal coordinate analyses displaying the centroids (position of labels) and dispersions around centroids (ellipses) of two sources of variation (i.e., years: A–C; sites: D–F) of three ecological communities (i.e., aquatic invertebrates: A, D; riparian vegetation: B, E; riparian invertebrates: C, F) within the Jaccard’s dissimilarity space. For the sack of visual clarity, results of pairwise comparisons across sites and years are not shown (significant year:site interaction in PERMANOVA and PERMDISP but see Tables S2 and S5, respectively).
The overall variability in taxonomic turnover of aquatic invertebrates across sampling years and sites were not significantly different (posthoc tests after PERMDISP; Table S5; ellipses in Figure 3). In turn, the variability was significantly different between most years (except between 2021 and 2022) and sites (except for S1u-S3r, S2u-S5r, S4r-S6d) for the riparian vegetation (Table S5). Last, the variability in taxonomic turnover of riparian invertebrates was significantly different across all years, and only between S1u-S6d, S2u-S6d, S3r-S6d, S4r-S6d, S5r-S6d (Table S5; see more details pairwise comparisons across years and sites in Table S6).
Although previous analyses on β-diversity were done at the species or morphospecies level (i.e., very rich information; n = 798 taxa in total), we here start exploring main trends in taxonomic turnover using the order level (n = 71 orders in total; Figure 4). Similar exploration of taxonomic turnover can be done at the family (Figure S1) and species/morphospecies levels (Figure S2). Across all six sites, Diptera were amongst the most dominant orders of aquatic organisms, along with Ephemeroptera and Amphipoda (Figure 4). Spatial patterns in plant orders were slightly more complex with more turnover in the most dominant orders from S1u to S6d compared to the aquatic invertebrates. Overall, Poales followed by Rosales were the most dominant orders in S1u, S2u, S3r, S6d, while Asterales and Poales dominated communities in S4r and S5r (Figure 4). Non-impounded sites (S1u, S2u, S6d) can be distinguished from other sites by the presence of Apiales, Ranunculales and Lamiales in the four most abundant orders. The riparian invertebrates showed the highest turnover in the ranks of dominant orders compared to the aquatic invertebrates and the riparian vegetation (Figure 4). Entomobryomorpha dominated communities within restored sites (S3r to S5r), but were also present in the most abundant orders in other sites (S1u, S2u and S6d). Araneae were among the most abundant orders in the restored sites: S3r and S4r.
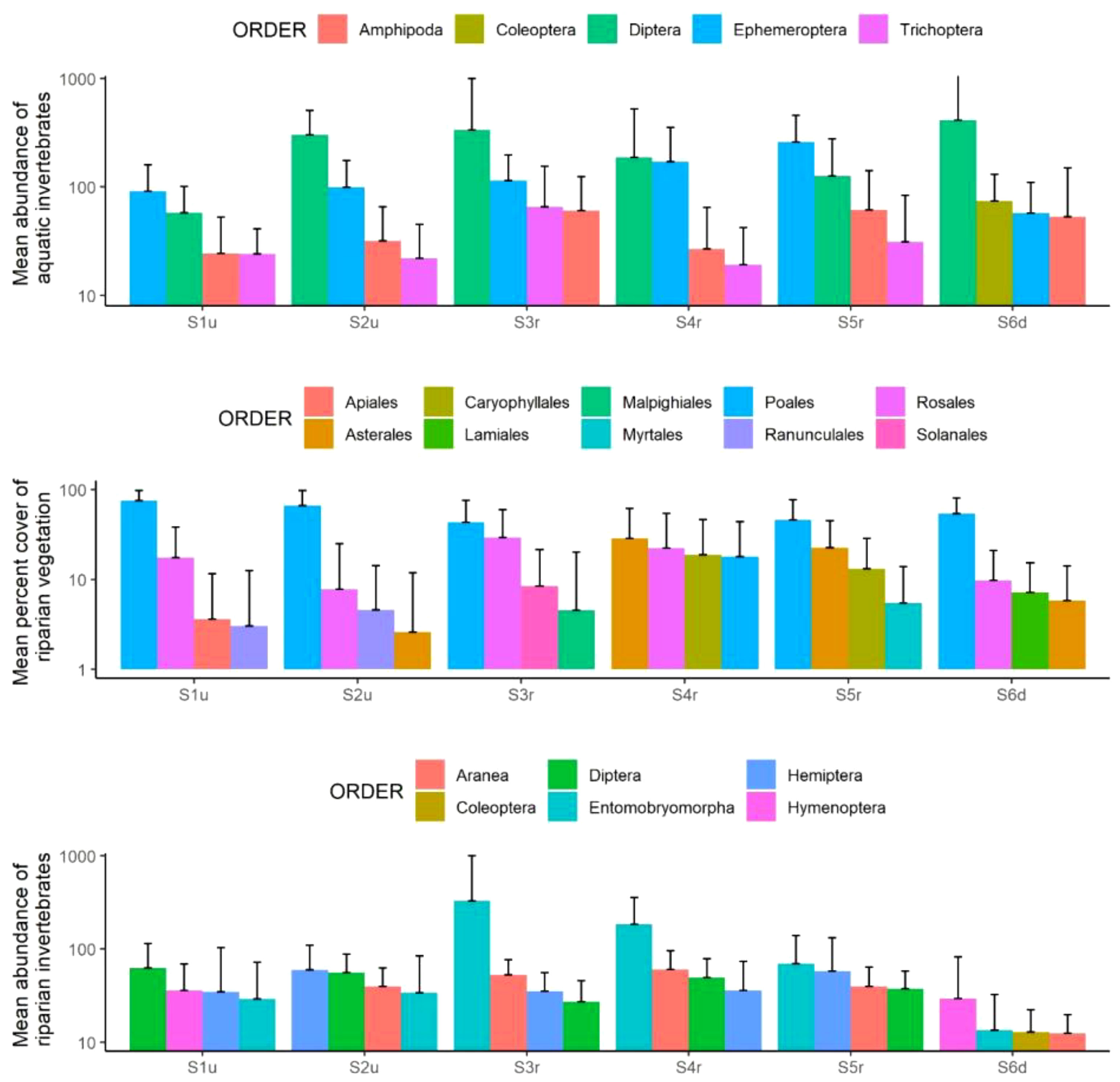
Figure 4 Distribution of the four most abundant orders of aquatic invertebrates, riparian vegetation and riparian invertebrates within each sampling site. Note that y-axes are displayed in the log10 scale.
In less than 1.5 years after the complete dewatering of the Vezins’ reservoir, the mean taxonomic richness of riparian vegetation, riparian invertebrates and aquatic invertebrates found in restored sites was similar to that of non-impounded sites. This suggests a fast recovery as expected (H1), in line with results from previous studies (Claeson and Coffin, 2016; Lisius et al., 2018; Poulos et al., 2019; Mahan et al., 2021). It contrasts, however, with other studies on macroinvertebrate recovery, which found either null or negative effects of dam removal on taxonomic diversity followed by an increase in diversity in subsequent years (Stanley et al., 2002; Kil and Bae, 2012). We found a negative but nonsignificant effect of the dam removal on the taxonomic richness of the aquatic invertebrates in the downstream site S6d, possibly impacted by increased downstream sedimentation after the removal of the second dam (La-Roche-Qui-Boit; Figure 1F; Mahan et al., 2021). Indeed, the presence of the La-Roche-Qui-Boit dam between the Vezins dam and S6d may have downplayed the adverse effects of increased sediments on aquatic communities in S6r by filtering the sediments coming from the removal of the Vezins dam. Overall, changes in the geomorphic (Tullos et al., 2014), and habitat features (Pollard and Reed, 2004; Claeson and Coffin, 2016) following dam removal should impose strong constraints on local communities, but the intensity of these constraints may depend on the spatial locations of study sites relative to the removed dam.
Contrary to our first hypothesis (H1), the α-diversity of all communities did not continue rising after one year post-removal in S3r, which rather hosted communities with near constant mean taxonomic richness over time. Two interrelated explanations could support this result. First, ecological communities in S3r had more time to colonize the new riverbanks and channel compared to other restored sites because of the slow dewatering (i.e. over several months) of the Vezins’ reservoir. Second, S3r is also the closest of all restored sites to the two upstream non-impounded sites (i.e., S1u and S2u), which may be important sources of colonizers (e.g., hydrochory, active dispersal of winged organisms). Therefore, community recovery may have been enhanced by a rapid organismal colonization at S3r from nearby upstream sites (i.e., upstream non-impounded sites, river tributaries) and in situ seed banks or dormant forms. For instance, the riparian vegetation in S3r displayed the characteristics of a fast colonizing and competitive post-pioneer assemblages, which were dominated by Urtica urtica (stinging nettle) and Phalaris arundinacea (reed canary grass; Figure S2). To help understand ecological recovery, future dam removal studies could identify nearby and in situ pools of colonizers (Ravot et al., 2021), for instance, within the 100 m from river channel (< 20 m in the present study; enhancing lateral recovery; Auble et al., 2007) or by increasing the sampling effort along the river and its tributaries (enhancing longitudinal recovery).
Patterns of recovery depended on ecological community, which is in accordance with our expectations. At the most downstream of the three sites in restoration (S5r), the riparian vegetation displayed no increase in mean taxonomic richness after one year compared to significant increases for both aquatic and riparian invertebrates. This site had a shorter restoration time than S4r and S3r (Figure 1F), and hosted, for instance, more pioneer plant taxa in communities, which then have either been replaced by more persistent taxa or prevented the establishment of additional taxa due to strong competition for limiting space and resources (Foley et al., 2017; Laslier et al., 2019).
In turn, all ecological communities showed significant increases in mean taxonomic richness across all three years of sampling in S4r. Studies on dam removals found similar temporal patterns for macroinvertebrate (Hansen and Hayes, 2012; Renöfält et al., 2013; Poulos et al., 2019; Mahan et al., 2021) and riparian vegetation (Foley et al., 2017; Lisius et al., 2018; Brown et al., 2022). The literature is far less abundant regarding responses of riparian invertebrate communities to dam removal, but some reports suggest strong declines in spiders within four years post-removal (Sullivan et al., 2018). Our results indicate that spiders (Figures 4, S1, S2) were among the most abundant taxa in restored sites. The extent to which taxonomic richness will continue to rise in S4r for all communities and in S5r for aquatic and riparian invertebrates beyond three years is unknown. Altogether these results suggest that the ecological recovery although fast is transient over three years for some communities and restored sites (i.e., context-dependent).
We originally assumed that the upstream-to-downstream gradient would leave an imprint on the recovery of ecological communities in the form of gradual changes in taxonomic composition (β-diversity) from upstream to downstream sites (S1u to S6d; H2; Rodríguez-Pérez et al., 2021). This assumption was supported by the riparian vegetation (supporting H2), displaying an upstream-to-downstream pattern in β-diversity. To a lesser extent, this spatial signal was also found in aquatic communities (i.e., upstream-to-downstream changes correlated with PCoA2 except for S1u; Figure 3F). This upstream-to-downstream pattern in β-diversity suggests that the longitudinal dispersal of communities along the river may play an important role in community recovery to dam removals.
Last, we expected the taxonomic recovery to vary according to the identity of the ecological community. This assumption was also partially supported as we found that all studied communities displayed high compositional differences in 2020 compared to 2021 and 2022 (including restored and non-impounded sites). This result may find an explanation when considering unmeasured but large-scale processes. For instance, Sullivan and Manning (2017) found that seasonality was as much important as local drivers (e.g., daily discharge, water depth and velocity, stream width) in macroinvertebrate responses to a dam removal. In addition, as for α-diversity, we found a strongly significant interaction between sites and years of sampling driving patterns in β-diversity of all ecological communities, suggesting a strong temporal and context-dependence of community recovery. Interestingly, the temporal turnover in taxonomic composition was higher in riparian organisms (i.e., invertebrates and plants) than in aquatic invertebrates. This difference between realms may arise because riparian organisms are influenced by in situ, lateral and longitudinal colonization processes while aquatic organisms are mainly driven by in situ and longitudinal processes. These finding add to the growing evidence of the spatial, temporal and ecological complexity of river recovery following dam removals. This recovery may result from direct and indirect interactions between local and regional deterministic processes along with dispersal limitations of organisms.
Functional ecology offers a powerful framework to better understand the causes and consequences of species-environment relationships (Southwood, 1977; Townsend and Hildrew, 1994; Violle et al., 2007) with countless implications in conservation biology and restoration ecology. It is a common practice in dam removal projects to investigate changes in the functional characteristics of fish communities (Shaffer et al., 2018; Jones et al., 2023), aquatic macroinvertebrate communities (Pollard and Reed, 2004; Hansen and Hayes, 2012; Kil and Bae, 2012; Poulos et al., 2019; Mahan et al., 2021), and riparian vegetation (Foley et al., 2017). In the present study, we explored taxonomic changes of these three communities as a first step to report ecosystem recovery after dam removal. Yet, rapid evaluations of taxonomic lists in our study (Figures 4, S1, S2) corroborate some evidence found in previous studies. For instance, collector-gatherers, represented by Chironomidae (Diptera) and Baetidae (Ephemeroptera), were among the most dominant taxa in newly restored channels (S3r to S5r; Mahan et al., 2021). In addition, pioneer plant taxa rapidly colonized the newly emerged riverbanks (e.g., Salicaceae, Polygonaceae, Asteraceae; Foley et al., 2017). A non-exhaustive review of the literature on the use of functional traits in dam removal studies highlighted, however, three main trends: most studies evaluate trophic-related traits (but see Tullos et al., 2014; Sullivan and Manning, 2017) excluding potentially important traits related to community recovery (e.g., dispersal capacities, flood-resistant seeds, dormant forms, reproduction type, life history traits), and functional information is scarce regarding many ecological communities (e.g., riparian vegetation, riparian invertebrates) thus preventing from a more integrated vision of ecosystem functioning under recovery. Exploring trophic relationships or food webs among organisms provides such an integrated picture of ecosystem functioning. Injecting food-web perspectives into the science of dam removal is, however, a daunting challenge. Only very few attempts explored how dam removal may affect cross-ecosystem trophic interactions (Sullivan et al., 2018). We therefore push future dam removal studies to further investigating the joint recovery of multiple ecological communities from both aquatic and terrestrial realms through the lens of trait-based and food-web ecology.
In between spring 2018 and 2019, the emptying of the one-century old, large-dam reservoir of Vezins offered new habitats to aquatic and terrestrial organisms on the Sélune River catchment. One year later, in spring 2020, we found that plants and invertebrates communities, reached levels of taxonomic diversity (α-diversity) similar to what is observed elsewhere on the river and its banksides suggesting a fast recovery. However, after three years post-removal, the α-diversity kept increasing and dissimilarity in taxonomic composition (β-diversity) remained high in some sites, suggesting a fast but transient ecological recovery. These patterns of recovery were strongly influenced by the identity of the ecological community, as well as spatial (e.g., lateral, longitudinal) and temporal (e.g., slow dewatering of former reservoirs) processes. These findings are in line with recent evidence of the strong context-dependent responses of ecological communities to large dam removals. This study initiates a first step within a larger goal to further understanding long-term and ecosystem-wide consequences of large dam removals.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
OD: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JMR: Conceptualization, Methodology, Funding acquisition, Project administration, Writing – review & editing. EL: Writing – review & editing. TB: Writing – review & editing. AB: Writing – review & editing. HR-P: Writing – review & editing. SD: Conceptualization, Funding acquisition, Project administration, Writing – review & editing. IB: Conceptualization, Funding acquisition, Project administration, Writing – review & editing. CP: Conceptualization, Methodology, Funding acquisition, Project administration, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was funded by the Agence de l’Eau Seine-Normandie (N° 1085121 (1) 2019).
We would like to thank Flavie Roussel, Mélissa Horckmans, and Caroline Bergère for their participation in the field campaigns related to this study. We are also sincerely grateful to Laura Soissons, Alban Thomas, and Manuel Chevé, for their unprecedented help in coordinating the Sélune programme. We last want to thank Alain Crave for his generous contribution to this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1254462/full#supplementary-material
Atristain M., von Schiller D., Larrañaga A., Elosegi A. (2023). Short-term effects of a large dam decommissioning on biofilm structure and functioning. Restor. Ecol. 31, e13779. doi: 10.1111/rec.13779
Auble G. T., Shafroth P. B., Scott M. L., Roelle J. E. (2007). Early vegetation development on an exposed reservoir: implications for dam removal. Environ. Manag 39, 806–818. doi: 10.1007/s00267-006-0018-z
Belletti B., Garcia de Leaniz C., Jones J., Bizzi S., Börger L., Segura G., et al. (2020). More than one million barriers fragment Europe’s rivers. Nature 588, 436–441. doi: 10.1038/s41586-020-3005-2
Birnie-Gauvin K., Larsen M. H., Nielsen J., Aarestrup K. (2017). 30 years of data reveal dramatic increase in abundance of brown trout following the removal of a small hydrodam. J. Environ. Manag 204, 467–471. doi: 10.1016/j.jenvman.2017.09.022
Brown R. L., Thomas C. C., Cubley E. S., Clausen A. J., Shafroth P. B. (2022). Does large dam removal restore downstream riparian vegetation diversity? Testing predictions on the Elwha River, Washington, USA. Ecol. App 32, e2591. doi: 10.1002/eap.2591
Bubb D. H., Birnie-Gauvin K., Tummers J. S., Aarestrup K., Jepsen N., Lucas M. C. (2021). Short-term effects of low-head barrier removals on fish communities and habitats. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.697106
Carlson P. E., Donadi S., Sandin L. (2018). Responses of macroinvertebrate communities to small dam removals: Implications for bioassessment and restoration. J. App Ecol. 55, 1896–1907. doi: 10.1111/1365-2664.13102
Catalano M. J., Bozek M. A., Pellett T. D. (2007). Effects of dam removal on fish assemblage structure and spatial distributions in the Baraboo River, Wisconsin. North Amer J. Fish Manag 27, 519–530. doi: 10.1577/M06-001.1
Chang H.-Y., Chiu M.-C., Chuang Y.-L., Tzeng C.-S., Kuo M.-H., Yeh C.-H., et al. (2017). Community responses to dam removal in a subtropical mountainous stream. Aquat Sci. 79, 967–983. doi: 10.1007/s00027-017-0545-0
Chiu M.-C., Yeh C.-H., Sun Y.-H., Kuo M.-H. (2013). Short-term effects of dam removal on macroinvertebrates in a Taiwan stream. Aquat Ecol. 47, 245–252. doi: 10.1007/s10452-013-9439-y
Claeson S. M., Coffin B. (2016). Physical and biological responses to an alternative removal strategy of a moderate-sized dam in Washington, USA. Riv Res. App 32, 1143–1152. doi: 10.1002/rra.2935
Cook D. R., Sullivan S. M. P. (2018). Associations between riffle development and aquatic biota following lowhead dam removal. Environ. Monit Assess. 190, 339. doi: 10.1007/s10661-018-6716-1
Cortese A. M., Bunn R. A. (2017). Availability and function of arbuscular mycorrhizal and ectomycorrhizal fungi during revegetation of dewatered reservoirs left after dam removal. Restor. Ecol. 25, 63–71. doi: 10.1111/rec.12406
Doyle M. W., Stanley E. H., Orr C. H., Selle A. R., Sethi S. A., Harbor J. M. (2005). Stream ecosystem response to small dam removal: lessons from the Heartland. Geomorph 71, 227–244. doi: 10.1016/j.geomorph.2004.04.011
Foley M. M., Warrick J. A., Ritchie A., Stevens A. W., Shafroth P. B., Duda J. J., et al. (2017). Coastal habitat and biological community response to dam removal on the Elwha River. Ecol. Monog 87, 552–577. doi: 10.1002/ecm.1268
Goslee S. C., Urban D. L. (2007). The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Soft 22 (7), 1–19. doi: 10.18637/jss.v022.i07
Grill G., Lehner B., Thieme M., Geenen B., Tickner D., Antonelli F., et al. (2019). Mapping the world’s free-flowing rivers. Nature 569, 215–221. doi: 10.1038/s41586-019-1111-9
Habel M., Mechkin K., Podgorska K., Saunes M., Babiński Z., Chalov S., et al. (2020). Dam and reservoir removal projects: a mix of social-ecological trends and cost-cutting attitudes. Sci. Rep. 10, 19210. doi: 10.1038/s41598-020-76158-3
Hansen J. F., Hayes D. B. (2012). Long-term implications of dam removal for macroinvertebrate communities in Michigan and Wisconsin rivers, United States. Riv Res. App 28, 1540–1550. doi: 10.1002/rra.1540
Hart D. D., Johnson T. E., Bushaw-Newton K. L., Horwitz R. J., Bednarek A. T., Charles D. F., et al. (2002). Dam removal: challenges and opportunities for ecological research and river restoration: we develop a risk assessment framework for understanding how potential responses to dam removal vary with dam and watershed characteristics, which can lead to more effective use of this restoration method. BioSci 52, 669–682. doi: 10.1641/0006-3568(2002)052[0669:DRCAOF]2.0.CO;2
Hervé M. (2020). “RVAideMemoire: testing and plotting procedures for biostatistics,” in R package version 0.9-77. Available at: https://CRAN.R-project.org/package=RVAideMemoire.
Hogg R. S., Coghlan S. M. Jr., Zydlewski J., Gardner C. (2015). Fish community response to a small-stream dam removal in a maine coastal river tributary. Trans. Am. Fish Soc. 144, 467–479. doi: 10.1080/00028487.2015.1007164
Jones A. C., Meiners S. J., Effert-Fanta E., Thomas T., Smith S. C. F., Colombo R. E. (2023). Low-head dam removal increases functional diversity of stream fish assemblages. Riv Res. App 39, 3–20. doi: 10.1002/rra.4063
Kil H. K., Bae Y. J. (2012). Effects of low-head dam removal on benthic macroinvertebrate communities in a Korean stream. Anim. Cel Sys 16, 69–76. doi: 10.1080/19768354.2011.611176
Kornis M. S., Weidel B. C., Powers S. M., Diebel M. W., Cline T. J., Fox J. M., et al. (2015). Fish community dynamics following dam removal in a fragmented agricultural stream. Aquat Sci. 77, 465–480. doi: 10.1007/s00027-014-0391-2
Laslier M., Hubert-Moy L., Corpetti T., Dufour S. (2019). Monitoring the colonization of alluvial deposits using multitemporal UAV RGB-imagery. App Veg Sci. 22, 561–572. doi: 10.1111/avsc.12455
Lisius G. L., Snyder N. P., Collins M. J. (2018). Vegetation community response to hydrologic and geomorphic changes following dam removal. Riv Res. App 34, 317–327. doi: 10.1002/rra.3261
López-Delgado E. O., Winemiller K. O., Villa-Navarro F. A. (2020). Local environmental factors influence beta-diversity patterns of tropical fish assemblages more than spatial factors. Ecology 101, e02940. doi: 10.1002/ecy.2940
Mahan D. C., Betts J. T., Nord E., Dyke F. V., Outcalt J. M. (2021). Response of benthic macroinvertebrates to dam removal in the restoration of the Boardman River, Michigan, USA. PloS One 16, e0245030. doi: 10.1371/journal.pone.0245030
McCaffery R., Jenkins K. J., Cendejas-Zarelli S., Happe P. J., Sager-Fradkin K. A. (2020). Small mammals and ungulates respond to and interact with revegetation processes following dam removal. Food Webs 25, e00159. doi: 10.1016/j.fooweb.2020.e00159
McCaffery R., McLaughlin J., Sager-Fradkin K., Jenkins K. J. (2018). Terrestrial fauna are agents and endpoints in ecosystem restoration following dam removal. Ecol. Restor. 36, 97–107. doi: 10.3368/er.36.2.97
O’Connor J. E., Duda J. J., Grant G. E. (2015). 1000 dams down and counting. Science 348, 496–497. doi: 10.1126/science.aaa9204
Oksanen J., Blanchet G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2019). “vegan: community ecology Package,” in R package version 2.5-6. Available at: https://CRAN.R-project.org/package=vegan.
Orr C. H., Kroiss S. J., Rogers K. L., Stanley E. H. (2008). Downstream benthic responses to small dam removal in a coldwater stream. Riv Res. App 24, 804–822. doi: 10.1002/rra.1084
Orr C. H., Stanley E. H. (2006). Vegetation development and restoration potential of drained reservoirs following dam removal in Wisconsin. Riv Res. App 22, 281–295. doi: 10.1002/rra.891
Poff N. L., Hart D. D. (2002). How dams vary and why it matters for the emerging science of dam removal: an ecological classification of dams is needed to characterize how the tremendous variation in the size, operational mode, age, and number of dams in a river basin influences the potential for restoring regulated rivers via dam removal. BioSci 52, 659–668. doi: 10.1641/0006-3568(2002)052[0659:HDVAWI]2.0.CO;2
Pollard A. I., Reed T. (2004). Benthic invertebrate assemblage change following dam removal in a Wisconsin stream. Hydrobiol 513, 51–58. doi: 10.1023/B:hydr.0000018164.17234.4f
Poulos H. M., Chernoff B. (2017). Effects of dam removal on fish community interactions and stability in the Eightmile river system, Connecticut, USA. Environ. Manag 59, 249–263. doi: 10.1007/s00267-016-0794-z
Poulos H. M., Miller K. E., Heinemann R., Kraczkowski M. L., Whelchel A. W., Chernoff B. (2019). Dam removal effects on benthic macroinvertebrate dynamics: a New England stream case study (Connecticut, USA). Sustain 11, 2875. doi: 10.3390/su11102875
Ravot C., Laslier M., Hubert-Moy L., Dufour S., Le Coeur D., Bernez I. (2020). Large dam removal and early spontaneous riparian vegetation recruitment on alluvium in a former reservoir: lessons learned from the pre-removal phase of the Sélune River project (France). Riv Res. App 36, 894–906. doi: 10.1002/rra.3535
Ravot C., Le Cœur D., Dufour S., Bernez I. (2021). Biodiversité végétale précoce de cinq affluents de la Sélune dans la vallée renaturée de Vezin (Normandie), in GOURDAIN P. (ed.), 10ème Colloque du Réseau d’Échange et de Valorisation en Écologie de la Restauration - REVER 10. Paris, 19-21 mars 2019. Naturae 26), 351–361. doi: 10.5852/naturae2021a26
R Core Team (2020). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Renöfält B. M., Lejon A. G. C., Jonsson M., Nilsson C. (2013). Long-term taxon-specific responses of macroinvertebrates to dam removal in a mid-sized swedish stream. Riv Res. App 29, 1082–1089. doi: 10.1002/rra.2592
Rodríguez-Pérez H., Pannard A., Gorzerino C., Pellan L., Massé S., Bouger G., et al. (2021). Ecological consequences of consecutive river damming for three groups of bioindicators. Ecol. Ind. 131, 108103. doi: 10.1016/j.ecolind.2021.108103
Rubin S. P., Miller I. M., Foley M. M., Berry H. D., Duda J. J., Hudson B., et al. (2017). Increased sediment load during a large-scale dam removal changes nearshore subtidal communities. PloS One 12, e0187742. doi: 10.1371/journal.pone.0187742
Ryan Bellmore J., Duda J. J., Craig L. S., Greene S. L., Torgersen C. E., Collins M. J., et al. (2017). Status and trends of dam removal research in the United States. WIREs Water 4, e1164. doi: 10.1002/wat2.1164
Sethi S. A., Selle A. R., Doyle M. W., Stanley E. H., Kitchel H. E. (2004). Response of unionid mussels to dam removal in Koshkonong Creek, Wisconsin (USA). Hydrobiol 525, 157–165. doi: 10.1023/B:HYDR.0000038862.63229.56
Shaffer J. A., Munsch S., Juanes F. (2018). Functional diversity responses of a nearshore fish community to restoration driven by large-scale dam removal. Est Coast. She Sci. 213, 245–252. doi: 10.1016/j.ecss.2018.08.030
Socolar J. B., Gilroy J. J., Kunin W. E., Edwards D. P. (2016). How should beta-diversity inform biodiversity conservation? Tre Ecol. Evol. 31, 67–80. doi: 10.1016/j.tree.2015.11.005
Southwood T. R. E. (1977). Habitat, the templet for ecological strategies. J. Anim. Ecol. 46, 337–365. doi: 10.2307/3817
Stanley E. H., Catalano M. J., Mercado-Silva N., Orr C. H. (2007). Effects of dam removal on brook trout in a Wisconsin stream. Riv Res. App 23, 792–798. doi: 10.1002/rra.1021
Stanley E. H., Luebke M. A., Doyle M. W., Marshall D. W. (2002). Short-term changes in channel form and macroinvertebrate communities following low-head dam removal. J. North Am. Bent Soc. 21, 172–187. doi: 10.2307/1468307
Stephens J. L. (2017). Short-term response of vegetation and the riparian bird community to dam removal on the Rogue River, Oregon. Ecol. Restor. 35, 328–340. doi: 10.3368/er.35.4.328
Sullivan S. M. P., Manning D. W. P. (2017). Seasonally distinct taxonomic and functional shifts in macroinvertebrate communities following dam removal. PeerJ 5, e3189. doi: 10.7717/peerj.3189
Sullivan S. M. P., Manning D. W. P., Davis R. P. (2018). Do the ecological impacts of dam removal extend across the aquatic–terrestrial boundary? Ecosph 9, e02180. doi: 10.1002/ecs2.2180
Thomson J. R., Hart D. D., Charles D. F., Nightengale T. L., Winter D. M. (2005). Effects of removal of a small dam on downstream macroinvertebrate and algal assemblages in a Pennsylvania stream. J. North Am. Bent Soc. 24 (1), 192–207. doi: 10.1899/0887-3593(2005)024<0192:EOROAS>2.0.CO;2
Townsend C. R., Hildrew A. G. (1994). Species traits in relation to a habitat templet for river systems. Fresh Biol. 31, 265–275. doi: 10.1111/j.1365-2427.1994.tb01740.x
Tszydel M., Grzybkowska M., Kruk A. (2009). Influence of dam removal on trichopteran assemblages in the lowland Drzewiczka River, Poland. Hydrobiol 630, 75–89. doi: 10.1007/s10750-009-9781-1
Tullos D. D., Collins M. J., Bellmore J. R., Bountry J. A., Connolly P. J., Shafroth P. B., et al. (2016). Synthesis of common management concerns associated with dam removal. JAWRA J. Am. Wat Res. Assoc. 52, 1179–1206. doi: 10.1111/1752-1688.12450
Tullos D. D., Finn D. S., Walter C. (2014). Geomorphic and ecological disturbance and recovery from two small dams and their removal. PloS One 9, e108091. doi: 10.1371/journal.pone.0108091
Violle C., Navas M.-L., Vile D., Kazakou E., Fortunel C., Hummel I., et al. (2007). Let the concept of trait be functional! Oikos 116, 882–892. doi: 10.1111/j.0030-1299.2007.15559.x
Whittaker R. H. (1965). Dominance and diversity in land plant communities: numerical relations of species express the importance of competition in community function and evolution. Science 147, 250–260. doi: 10.1126/science.147.3655.250
Keywords: ecological restoration, metacommunity, riparian vegetation, macroinvertebrate, habitat fragmentation and modification, river continuum
Citation: Dézerald O, Roussel J-M, Lanoë E, Beauverger T, Bazin A, Rodríguez-Pérez H, Dufour S, Bernez I and Piscart C (2023) Fast but transient recovery of aquatic and terrestrial communities after a large dam removal. Front. Ecol. Evol. 11:1254462. doi: 10.3389/fevo.2023.1254462
Received: 07 July 2023; Accepted: 20 November 2023;
Published: 07 December 2023.
Edited by:
Ming-Chih Chiu, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Sicheng Ao, Chinese Academy of Sciences (CAS), ChinaCopyright © 2023 Dézerald, Roussel, Lanoë, Beauverger, Bazin, Rodríguez-Pérez, Dufour, Bernez and Piscart. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Olivier Dézerald, b2xpdmllci5kZXplcmFsZEBpbnJhZS5mcg==
†ORCID: Olivier Dézerald, orcid.org/0000-0002-9987-9865
Jean-Marc Roussel, orcid.org/0000-0002-6301-3234
Simon Dufour, orcid.org/0000-0001-8268-9371
Ivan Bernez, orcid.org/0000-0002-1833-3212
Christophe Piscart, orcid.org/0000-0002-4054-4542
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.