 Samuel J. Lane1*†
Samuel J. Lane1*† Isaac J. VanDiest1
Isaac J. VanDiest1 Valerie N. Brewer1†Courtney R. Linkous1Taylor E. Fossett1
Valerie N. Brewer1†Courtney R. Linkous1Taylor E. Fossett1 Christopher G. Goodchild1†
Christopher G. Goodchild1† Kendra B. Sewall1,2
Kendra B. Sewall1,2- 1Department of Biological Sciences, Virginia Tech, Blacksburg, VA, United States
- 2Department of Biological Sciences and School of Neuroscience, Virginia Tech, Blacksburg, VA, United States
Behavioral traits are often the first response to changing environmental conditions, including human induced rapid environmental change. For example, animals living in urban areas are often more aggressive than rural animals. This is especially evident in songbirds; males of several species display elevated aggression in urban habitats. Increased male aggression has been associated with reduced parental care, but the consequences of this trade-off for males, social partners, and offspring in the context of urbanization remains unclear. We explored the effects of increased urban male aggression on the life history traits, parental care, and offspring outcomes of song sparrows (Melospiza melodia). We predicted that urban males would reduce paternal investment and result in urban females providing greater nestling care or reduced fledging success in urban habitats compared to rural. Contrary to our prediction, aggressive urban males did not decrease care but visited the nest more often compared to rural males. Additionally, urban birds had higher nest and fledging success compared to rural, though this was largely due to higher nest predation in rural habitats. Our study is among the first to evaluate trade-offs associated with elevated aggression expressed by urban animals and adds to a growing body of evidence that urban habitats provide benefits to some species.
1 Introduction
Life-history theory is structured around the idea that the evolution of suites of traits meant to maximize an individual’s fitness constrain each other and consequently generate trade-offs (Stearns, 1992; Reznick et al., 2000; Roff and Fairbairn, 2007). With time and energy being limited, behavioral trade-offs occur when animals must strike a balance among behaviors that are energetically costly or require substantial time. The habitat in which an individual lives has been shown to modulate trade-offs between behaviors, because the environmental context determines an animal’s time and energy budget, and the extent to which behaviors constrain each other (Gunnarsson et al., 2005; Holtmann et al., 2017). Resource availability, conspecific density, predator presence, and heterospecific interactions are among the contextual factors that determine an individual’s capacity to maximize fitness linked traits (Linden and Møller, 1989; Martin, 1995), all of which are being altered by global anthropogenic change (McKinney, 2008; Grimm et al., 2008b). Supporting the expansion of species to urban habitats can maintain urban biodiversity as human expansion continues (McKinney, 2002; Grimm et al., 2008b). However, urban habitats present wildlife with an altered abiotic and biotic landscape in which to live and reproduce (Chace and Walsh, 2006; Grimm et al., 2008a) that may modify the trade-offs among life-history traits.
Many species of songbirds have readily adjusted to cities, but breeding adults likely face new or exaggerated challenges not present in their native habitats. One way that animals can adjust to novel conditions is by shifting how they resolve life-history trade-offs. Urban habitats contain anthropogenic predators (e.g., domestic cats) (Loss et al., 2013; Rosenberg et al., 2019) that influence survival rates of both offspring and adults (Baker et al., 2008; van Heezik et al., 2010). These habitats are more fragmented than native habitats with different resource distribution and availability (Saunders et al., 1991; Chace and Walsh, 2006; McKinney, 2006). Finally, obligate brood parasites tend to follow fragmentation and human disturbance (Brittingham and Temple, 1983; Robinson et al., 1995; Chace et al., 2003; Rodewald, 2009; Faaborg et al., 2010) and may be more prevalent in urban communities. Comparing the behaviors of species in which individuals maintain breeding territories in both urban and rural habitats (termed ‘urban adapters’; Blair, 1996; McKinney, 2002; Shochat et al., 2006; Bonier, 2012), can reveal how individuals re-allocate effort into different life-history behaviors in response to novel urban habitats (Blair, 1996; McKinney, 2002; Shochat et al., 2006; Bonier, 2012).
One trade-off that could be shifted in urban habitats is the balance between territoriality and paternal care in male songbirds. During the breeding season, male songbirds choose and defend territories with quality nesting substrates and resources needed to survive and rear young (Stamps and Krishnan, 1997; Garcia and Arroyo, 2002). In theory, the highest quality males should be the most aggressive and best able to defend the highest quality breeding territories (Otter and Ratcliffe, 1996; Garcia and Arroyo, 2002; Scales et al., 2013) leading to increased fitness (Otter and Ratcliffe, 1996). Urban habitats often have fewer available breeding territories and resources compared to rural habitats (Juárez et al., 2020), and urban habitats likely favor a more aggressive phenotype (Duckworth, 2008; Foltz et al., 2015; Juárez et al., 2020). Indeed, one behavioral pattern found in many urban populations (Lowry et al., 2013; Renthlei et al., 2017; Dammhahn et al., 2020) but especially in songbirds, is increased male conspecific aggression (Evans et al., 2010; Fokidis et al., 2011; Scales et al., 2011; Atwell et al., 2014; Davies and Sewall, 2016). While an exaggerated aggressive phenotype may facilitate territory acquisition in urban habitats, investment in aggressive behavior could result in a trade-off with other energetically costly behaviors, such as parental care (Ketterson and Nolan, 1994; Ketterson and Nolan, 1999; Duckworth, 2006; McGlothlin et al., 2007). For socially monogamous urban songbirds, any trade-offs in breeding behaviors (i.e., territoriality and parental care) can have consequences for social partners because mates cooperate in rearing young (Gunnarsson et al., 2005; Holtmann et al., 2017). Thus, if urban habitats drive shifts in how male songbirds resolve life-history trade-offs, there may also be consequences for their female partners and the condition and survival of their offspring (Marzluff, 2001; Duckworth, 2006; Grimm et al., 2008a; Scolozzi and Geneletti, 2012).
Many territorial songbird species establish social partnerships during the breeding season to provide biparental care to their offspring (Westneat, 1987; Arcese, 1989; Griffith et al., 2002). This social monogamy allows partners to split the energetic demands of reproduction, so the burden of care does not fall on one individual (Møller and Birkhead, 1993; Griffith et al., 2002). However, female’s higher energetic investment, and guaranteed genetic relatedness to offspring, means they often show increased reproductive investment compared to males (Westneat, 1987; Arcese, 1989; Griffith et al., 2002). Indeed, prior studies of the consequences of male aggression have highlighted impacts on mates and offspring, including compensatory behaviors and reduced fledging success (Whittingham et al., 1994; Saino and Møller, 1995; Paredes et al., 2005). More aggressive male house finches, for instance, provide less paternal care and females compensate by providing more care (Badyaev and Hill, 2002). While females can often compensate for reduced care by their male partners, harsh environmental conditions that make compensation impossible can result in smaller offspring or reduced fledging success. If increased aggression in urban males constrains their ability to provide paternal care, this could increase the demands on their social partner to provide greater maternal care.
Here, we investigated the effects of urbanization and the associated increase in male aggression on the parental care provided by males, the care provided by their mates, and the condition and fledging success of their offspring by comparing adult behavior and nestling outcomes among wild song sparrows (Melospiza melodia) in urban and rural habitats. Song sparrows are a commonly studied urban adapter that, like many songbird species, are open cup nesters with altricial young, for which biparental parental care is especially vital (Nice, 1943; Smith and Roff, 1980; Roff et al., 2005; Martin and Briskie, 2009). We hypothesized that urban males, which are reliably more aggressive toward conspecifics, may resolve trade-offs between territoriality and paternal care differently than their rural counterparts and provide less care to offspring. We expected that their social partners would compensate to maintain offspring fitness or, if they could not, nestling growth and survival would be reduced. Specifically, we predicted that urban males would provide less care compared to rural males. We also predicted that maternal care would be positively correlated with social partner aggression, and negatively correlated with paternal care. Finally, we predicted urban nesting success would be lower, and urban offspring would be underdeveloped near fledging compared to rural birds.
2 Methods
2.1 Parental behavior
2.1.1 Subjects
To examine the relationship between male aggression, parental visitation, and its effects on social partners, in the breeding seasons of 2018 – 2021 we measured parental care at nests located within 6 previously established sites in Southwestern Virginia, USA that vary along an urban-rural gradient (Davies and Sewall, 2016; Davies et al., 2018). We only monitored nest outcomes and nestling growth in 2017, all additional parental measures began in 2018. To evaluate the levels of urbanization, Davies et al. (2018) used an automated system to assign an urbanization score to each site using the protocols described in Seress et al. (2014) (For detailed descriptions and photos of field sites and for full analysis details see Davies et al., 2018). After we located a nest and established the breeding stage, we measured male territorial aggression in response to a simulated territorial intrusion following Hyman et al. (2004) (see below). We performed behavioral trials between April and July of each breeding season (see supplemental materials for a detailed figure outlining our methods timeline).
To monitor parental visitation by focal males and their social partners, we captured adult birds with mist nets between 0500- and 1115-hours from March through July of each year. Of the 288 song sparrow nests monitored during this project (166 urban and 119 rural), we were able to monitor paternal care at 86 nests (67 urban and 19 rural), and maternal care at 54 nests (38 urban and 16 rural). At 43 nests, both the male and female were captured and only these nests were included in the social partner behavior analysis. At all other nests, we were only able to capture either the male or female. We captured males by targeting them immediately after simulated territorial intrusions, while we captured females when they were leaving or returning to the nest. To avoid nest abandonment, we allowed all active nests to reach at least mid- to late-incubation before capture, did not attempt to catch a social pair (a) for more than 120 min/day, (b) during inclement weather, or (c) on sequential days. We took morphological measurements from the adults and banded them with a unique color band. Before releasing each bird, we fitted a passive integrated transponder (PIT) tag (2.12x10mm; CYNTAG, Inc; Item#: 601205-2248) to the tarsometatarsus of each adult following the methods established in Bridge and Bonter (2011) to monitor parental visitation. The PIT tags are small coils of magnetic wire programed with a unique 10-digit code that can be read by any radio frequency identification (RFID) system (Bridge and Bonter, 2011; Bridge et al., 2019; Farr et al., 2021). We gave every bird a unique combination of color bands and multicolored heat shrink tubing incasing the PIT tags (Figure 1A) to allow us to visually identify individuals using binoculars.

Figure 1 A PIT tag attached to the tarsometatarsus of a song sparrow (A) and an antenna fitted around a song sparrow nest (B) during the 2018 field season. When the magnetic coil built into the PIT tag disrupts the magnetic field created by the antenna the duration of the visit is recorded, time stamped and stored by the microprocessor hidden nearby in a waterproof container.
2.1.2 Male aggression
To examine the tradeoff between male aggression and parental care, we quantified male aggression in 140 males between 2018 and 2021. Of these males, we were also able to monitor parental behavior in 61 (46 urban and 15 rural males). During aggression trials we located focal males based on location of spontaneous singing relative to the active nest and/or in response to brief (< 1 min) playback. Once located, we exposed a male to 6 min of playback with one of 6 3D printed song sparrow mounts placed at the approximate center of the territory (For details on the design and construction of the 3D printed models please see Beck et al., 2019). Each playback consisted of 2 song types recorded from the same male designed to approximate a male’s natural song (Hyman et al., 2004; Davies and Sewall, 2016). To assess male territorial aggression during each playback, we recorded distance to speaker/mount, number of low amplitude songs (soft songs), and number of wing waves each minute (Hyman et al., 2004; Akçay et al., 2011; Davies and Sewall, 2016; Davies et al., 2018). To measure the average distance to the speaker during the trials, we recorded the bin distance from the speaker (0-2, 2-4, 4-8,8-16, and >24m) every 5 seconds for the entire 6 min behavioral trail. During data extraction average distance was calculated following methods in Peters et al. (1980) (Hyman et al., 2004; Davies et al., 2018). We combined all the territorial response data into a single ‘territorial aggression score’ using Principal Component Analysis (PCA; See supplementary materials for table of loadings). Male aggression is repeatable within individuals during a breeding season (Davies and Sewall, 2016), even though territorial aggression declines over the season. Therefore, we used a single trial to quantify aggression but include day of year in all analysis.
2.1.3 RFID and parental visitation
To monitor parental nest visitation rates in 2018 – 2021, we placed a RFID system at the nest (for details on function, form, and construction of RFID systems for monitoring wild bird populations see Adelman et al., 2014; Bridge et al., 2019) on the days leading up to nestling hatch. The system consisted of an antenna wrapped in camouflage electrical tape (for waterproofing) that was carefully placed around the external diameter of the nest (Figure 1B). The antenna was attached to a battery-powered microprocessor (E. Bridge; GEN 2 RFID Reader and Data Logger, 2012; Bridge and Bonter, 2011) with a 2 GB card (hereafter referred to as the “board”). The board and battery were housed in a waterproof container that was hidden near the nest. When the PIT tag disrupts the magnetic field created by the antenna, it is powered on and transmits a unique identification number which is recorded by the board as long as the PIT tag is within its range (here the range is the internal area of the nest cup plus a few centimeters above, and below the bottom of the nest cup). Batteries for the RFID system were changed every 2 – 3 days, and parental visitation was monitored for 24 hours a day throughout the breeding attempt.
This RFID system was set with a read interval of 6 seconds and allows for continuous fine-scale monitoring of parental visitation during the entire nestling period. All RFID data were inspected visually for quality and accuracy. We used videos to spot-check RFID data by visually recording behavior and matching it to data recorded by the boards. We considered a lag time of >1 min between reads of a PIT tag number as a male leaving the nest. This was marked as an ‘OFF’ time in the data. To calculate the number of nest visits made by an individual, we summed the number of OFF visits recorded in a day. Females move frequently when on the nest during the early nestling stage, so we increased the OFF time to >2 min to be more conservative in analyzing female visitations. To calculate “total time” an individual provided care for a given day, we subtracted the first and last read for that individual for the day. We used the number of visits and the “total time” for each day to calculate visitation rate (visits/(time of final visits in the day – time of first visit of the day)) for each parent each day of the nestling period. Additionally, we calculated the average duration of each nest visit (total time on the nest/visits) per day. In total, we measure parental visitation in 76 males (57 urban and 19 rural) and 54 females (38 urban and 16 rural). Of these adults were able to capture both social partners at 43 nests (32 urban and 11 rural). Not all nests survived the breeding attempts, and some nests were located, or parents were captured after hatch. To account for variation in monitoring duration, nestling age, and day of year were included as fixed-effects covariates in all analysis.
2.2 Monitoring nesting success and offspring development
2.2.1 Nest searching and monitoring
To examine the effects of urbanization on the fitness of wild song sparrows, we located and monitored nests in urban and rural habitats throughout 5 breeding seasons. In the spring and summers of 2017 to 2021 we searched for and monitored song sparrow nests at the previously mentioned field sites. We searched habitat types evenly (2 days searching urban, 2 days searching rural repeated across the field season) to control for nest searching effort. Nest searching was conducted primarily in the early morning (0500 to 1100) during peak parental behavior, from early May until late July in 2017, and from early March to late July in 2018- 2021. For analysis we only included one nest from each pair that was closest to the mean lay date for the year. In total, we found and monitored 166 nests in urban habitats and 119 nests in rural habitats.
We located nests primarily through behavioral observations of adults and systematic searching of known territories and nesting substrates (Martin and Geupel, 1993). When we located nests during incubation, a minimum of two eggs were candled to establish incubation stage and target hatching date. After we located a nest, we limited the amount of time spent at the nests and number of visits to the nests to reduce nest abandonment. We only visited nests to establish vital developmental stages (lay date, onset of incubation, and hatch date), monitor offspring development, and conduct adult behavior assays and capture. We used previously published life-history data for this species to estimate dates for events (i.e., nest building, lay and incubation duration) that occurred before a nest was found (Smith and Roff, 1980; Sogge and Van Riper, 1988).
We defined nest success as at least one song sparrow nestling fledging from the nest and nest failure as complete nest predation (all eggs gone from the nest, all nestlings gone while still altricial), parental abandonment, or an anthropogenic form of nest destruction. Fledging success was the number of nestlings that fledged from the nest. If we did not record nest success or failure on the RFID system, we estimated the date of nest completion as the midpoint between the last two nest visits (Mayfield, 1961). We controlled for stage location bias (nests found in late stages overestimate nesting success), by calculating daily nest survival rates and nest survival rates for each habitat using Mayfield estimations (Mayfield, 1961; Jehle et al., 2004). We visually inspected all final clutches for brood parasitism by brown-headed cowbirds (Molothrus ater), using previously established methods to distinguish host eggs from brown-headed cowbirds (Smith and Arcese, 1994). If a nest was found after hatch, we used visual identification and morphometrics to identify brood parasitism (Pyle, 1997).
2.2.2 Offspring condition
To examine the effect of urbanization, and possible indirect effects it has on parental nest visitation, and on offspring condition we compared nestling body condition just prior to fledging (Day 9 and 10) between habitat types. Across all years we measured 155 urban nestlings and 36 rural nestlings from 61 urban nests and 14 rural nests. We measured nestling song sparrow mass, and tarsus just before fledging (Day 9; Pyle, 1997). To ensure accuracy, each measurement was taken 5 times, the highest and lowest measurement were dropped, and an average was taken from the remaining 3. If we located a nest after the clutch had hatched, measurements were taken on the day it was found and nestling age was estimated from morphometric variables (Pyle, 1997; Jonsomjit et al., 2007).
2.3 Statistical analysis
2.3.1 Behavior
We conducted statistical analyses using R (R Core Team, 2021, v. 4.0.4). We analyzed all parental visitation and male aggression data using linear mixed effects models (LMM). We fitted all LMM’s and generalized linear mixed effects models (GLMM) using the package “lme4” (Bates et al., 2015) and we checked the model assumptions using the “performance” package (Lüdecke et al., 2021). We tested the significance of fixed effects from the LMMs using the lmerTest (Kuznetsova et al., 2017) package, which estimates degrees of freedom (df) with the Satterthwaite approximation. A full summary of results from all models presented in this paper are included in the supplemental materials. We analyzed the effect of habitat on paternal visitation, maternal visitation, and the correlation between paternal visitation and aggression in separate LMMs using a data set that included all adults monitored during this project. In each model, visits/hour (by either the male or female depending on the model) was used as the response variable. For the two models examining paternal and maternal visitation, habitat type was a categorical fixed-effect. To examine the relationship between paternal visitation and aggression we included male aggression score (PC1, which explained 53.19% of the variation in behavior, see Supplementary Table S1, model results) and habitat type as the predictor variables. To examine the relationship between maternal visitation and male aggression we included maternal visits/hour calculated for each day as the response variable with social partner’s aggression score and habitat type as the predictor variable with an interaction indicated. We created a final LMM to examine the relationship between social partner parental visitation that only included the nests from which we had parental visitation from both social partners (41 pairs). For this model, we averaged male and female social partners visitation rates for each day, then included female visitation as the predictor variable, with male visitation and habitat type as fixed-effects. For all behavioral analysis we included age of the nestlings (day 1 – 10), total number of nestlings in the nest (including brood parasites), Julian day, and year sampled as fixed-effect covariates. Parental visitation did not vary with the presence of a brood parasite in the nest, and the covariate was dropped from all parental visitation models, but brood parasite nestlings were included in all brood sizes. We included the unique nest ID as the random effect in each model to control for autocorrelation within the nest and for repeated measure of continuous recording of parental visitation.
2.3.2 Nest outcomes and nestling condition
To analyze patterns of nest success, nest predation, and brood parasitism by brown-headed cowbirds between urban and rural habits we used separate linear models (LMs) fitted to a binomial distribution. For each model we included nest success or failure, presence or absence of nest predation, and presence or absence of brood parasitism as binomial response variables. We used habitat type, and year as fixed-effect covariates in the models. For the nest success and nest predation models, we also included the presence or absence of brood parasitism as fixed-effect covariates. We further analyzed fitness measures of fledging success and hatching success between urban and rural habitat using generalized linear models (GLMs) fitted to a negative binomial distribution using the Mass (Venables and Ripley, 2002) package. In each model, fledging success or hatching success was the response variable, with habitat type, the presences of brood parasitism, lay initiation date and year included as fixed-effect covariates. We did the same analysis on the subset of nests that were successful, except the data was fitted to a normal distribution. Finally, we examined a commonly used fitness metric, lay date, between habitat types using a LM (Winkler et al., 2020). We included Julian lay date as the response variable and habitat type, brood parasitism, and year as fixed-effect covariates in the model. For this model, we used a data set that included the first breeding attempt for each pair, rather than the closest breeding attempt to the mean lay date.
To determine if there were any differences in nestling condition near fledging between habitat types, we used two separate LMMs with morphological measures (either nestling tarsus length or mass) as the response variable in each model. We included habitat type and nestling age as fixed-effects. Julian date, number of nestlings in the nest, brood parasitism, and year were also included as fixed-effect covariates. To control for autocorrelation between nestlings in the same nest, we included the unique nest ID as the random effect.
3 Results
3.1 Male aggression and parental visitation between urban and rural habitats
Male nest visitation rates significantly increased with the total number of nestlings present in the nest (β = 0.43 ± 0.11, t490.09 = 3.78, p = 0.0002) and nestling age (β = 0.16 ± 0.03, t552.09 = 5.59, p = <0.0001) across habitat types. However, independent of brood size and nestling age, urban males visited the nest significantly more often compared to rural males (β = 0.96 ± 0.46, t79.09 = 2.07, p = 0.04; Figure 2; Random effects: among-nest variance = 2.30, residuals variance = 2.60). Contrary to our predictions, we found no relationship between male aggression and nest visitation rates through the brooding period (β = 0.09 ± .62, t54.35 = 0.15, p = 0.89; Figure 3) nor did we find a relationship between male aggression and maternal visitation rates (β = -0.81 ± 0.67, t29.12 = -1.20, p = 0.24; Random effects: among-nest variance = 0.70, residuals variance = 2.04). While urban males visited the nest more frequently than rural males, urban females did not. There was a trend for increased maternal visitation in urban habitats (β = 0.66 ± 0.33, t48.39 = 1.99, p = 0.05; Figure 4; Random effects: among-nest variance = 0.64, residuals variance = 1.94) and a significant negative relationship between female visitation rates and nestling age (β = -0.08 ± 0.03, t315.98 = -2.60, p = 0.01), but female visitation rates did not vary with the number of nestlings present in the nest (β = 0.08 ± 0.12, t223.87 = 0.62, p = 0.53). Finally, there was a significant positive relationship between daily social partner nest visitation rates across habitat types (β = 0.33 ± 0.05, t231.20 = 7.17, p = <0.0001; Random effects: among-nest variance = 0.56, residuals variance = 1.53; Figure 5) and there was no difference in this relationship between habitats.
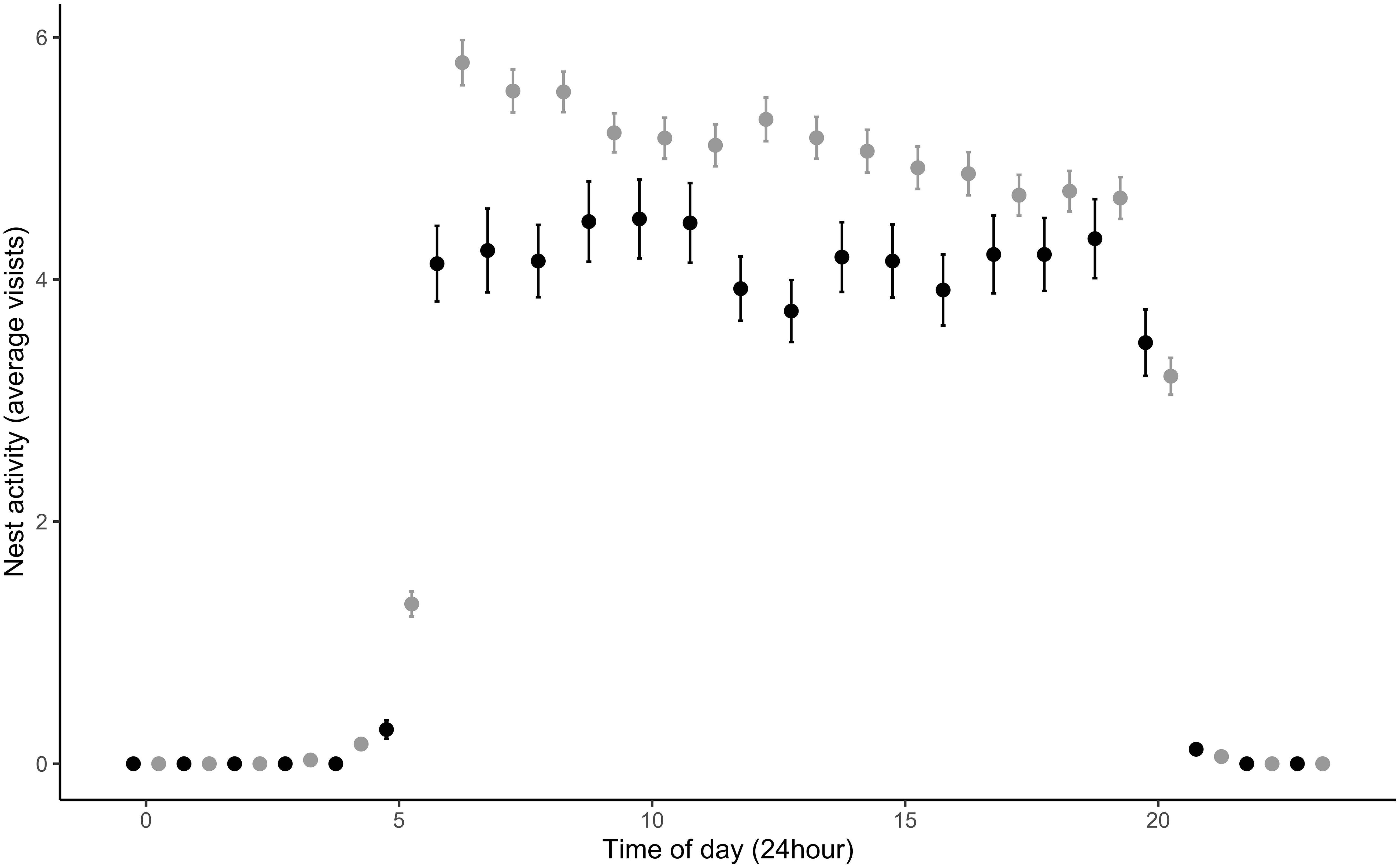
Figure 2 Paternal visitation between urban (gray) and rural (black) habitats during the nestling period (day 0 – 10). Urban males (67) visited the nest significantly more often (5.66 ± 0.11 times/hour) during nestling provisioning compared to rural males (19) that visited the nest on average 4.70 ± 0.18 times/hour. During these visits, urban males spent on average 7.60 ± 0.31 minutes per visits compared to rural, who spent on average 7.72 ± 1.04 minutes per visit. Each time point is averaged from 388 urban observation (days) and 92 rural observations (days).
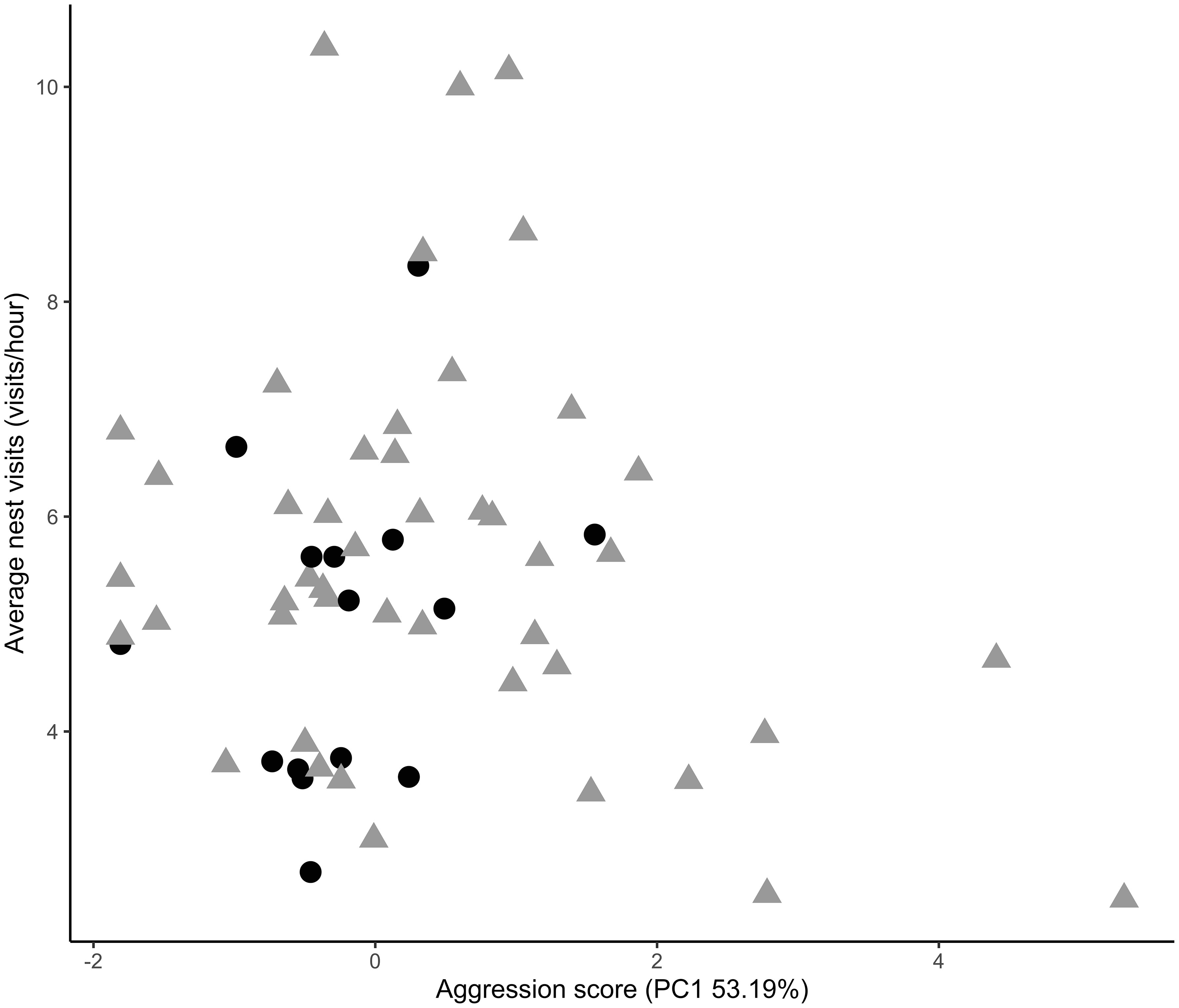
Figure 3 Male aggression in urban (triangles) and rural (circles) song sparrows was not correlated with paternal care. Urban males (46) had an average aggression score of 0.49 ± 0.08 compared to rural (15) that had an average aggression score of -0.40 ± 0.09.
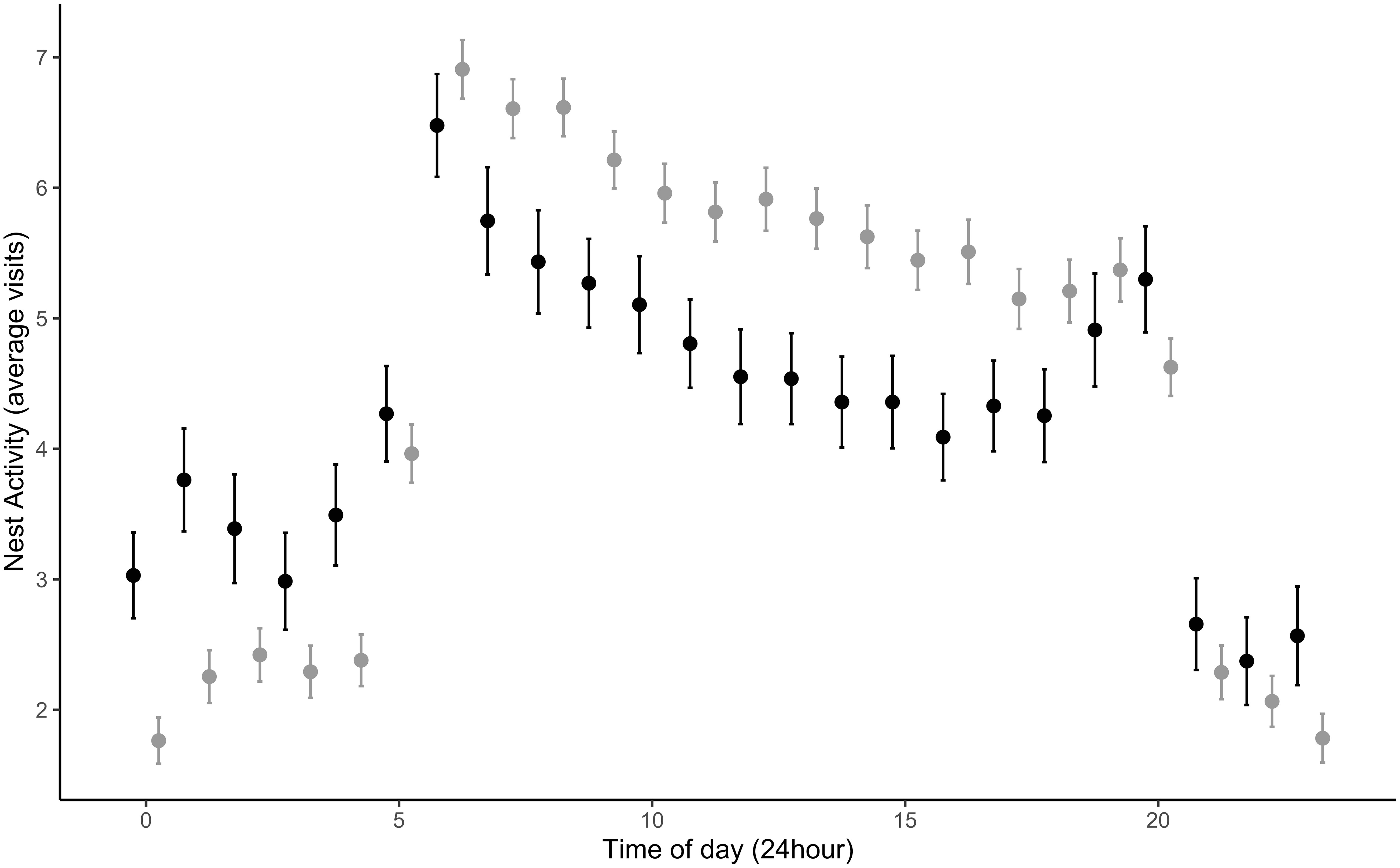
Figure 4 Maternal visitations varied between urban (gray) and rural (black) habitats during the nestling period (day 0 – 10) depending on time of day. Urban females appear to increase nest activity compared to rural during the day, and the opposite occurs at night. Urban females (38) visited the nest on average 5.84 ± 0.10 times/hour during nestling provisioning compared to rural females (16) that visited the nest on average 5.24 ± 0.16 times/hour. During these visits, urban females spent on average 41.94 ± 2.89 minutes per visits compared to rural, who spent on average 55.99 ± 4.76 minutes per visit. Each time point is averaged from 216 urban observation (days) and 67 rural observations (days).
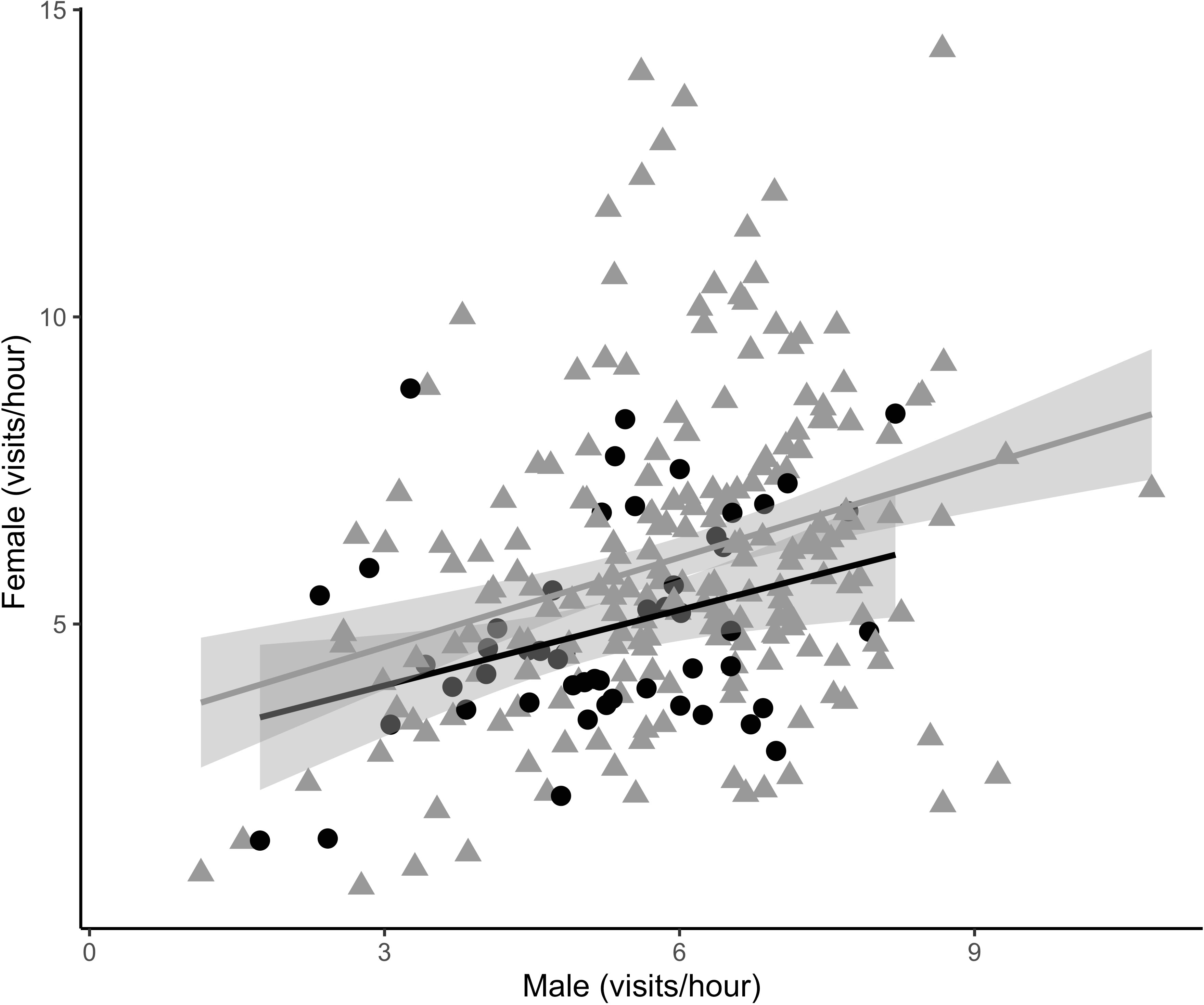
Figure 5 Social partner care was positively corelated during offspring provisioning in both urban (triangles) and rural (circles) habitats. Females (32 urban 11 rural) that visited the nest on average 5.72 ± 0.10 times/hour had social partners (32 urban 11 rural) that visited on nest on average 5.80 ± 0.14 times/hour. Each data point represents the average care provided in a day by the female and her social partner for a total of 274 days of combined parental care data.
3.2 Offspring condition and nest outcomes between urban and rural habitats
There was no effect of habitat type on nestling body mass (β = -0.13 ± 9.65, t 64.99 = -0.01, p = 0.99; Figure 6A) or tarsus length (β = 1.03 ± 6.29, t 63.89 = 0.16, p = 0.87) just before fledge (day 9 and 10). However, nestling mass (β = -1.61 ± 0.52, t 80.28 = -3.13, p = 0.002; Random effects: among-nest variance = 1.59, residuals variance = 2.58; Figure 6B) and tarsus length (β = 0.93 ± 0.33, t 73.27 = -2.81, p = 0.006; Random effects: among-nest variance = 0.82, residuals variance = 0.76) were significantly lower before fledge in nests containing brown headed cowbirds compared to unparasitized nests. Additionally, of nests that survived to fledge and were not predated, nests that were parasitized by brown-headed cowbirds hatched (β = -1.26 ± 0.188, t 85 = -6.69, p = <0.0001) and fledged (β = -1.29 ± 0.23, t 97 = -5.66, p = <0.0001) significantly less song sparrow offspring compared to unparasitized nests.
3.3 The fitness effects of breeding habitat type
Nest success rates were significantly higher in urban habitats compared to rural (β = 1.80 ± 0.32, z279 = 3.15, p = <0.0001) but success rates varied across years (β = -0.34 ± 0.11, z279 = -3.15, p = 0.002). Similarly, hatching success (β = 0.063 ± 0.16, z236 = 3.82, p = 0.0001) and fledging success (β = 1.09 ± 0.21, z203 = 5.22, p = <0.0001) were significantly higher in urban habitats compared to rural. Nest predation rates were significantly higher in rural habitats compared to urban (β = -1.84 ± 0.29, z 284= -6.37, p = <0.0001) and nest predation rates also expressed inter-year variation (β = 0.24 ± 0.33, z284 = 0.79, p = 0.02). Brood parasitism by brown headed cowbirds was significantly higher in urban habitats compared to rural (β = 2.29 ± 0.40, z287 = 5.74, p = <0.0001). We found that lay date was significantly earlier in urban habitats compared to rural (β = -18.69 ± 3.47, t144 = -5.39, p = <0.0001), with rural birds starting to lay on May 27th ± 2.35 days (64 nests with confirmed building or lay dates across 5 years) and urban around May 8thth ± 1.96 days (110 nests with confirmed building or lay dates across 5 years).
4 Discussion
Time and energy are limiting resources for seasonal breeding species that have a narrow window to mate and raise their offspring, and decisions on how individuals allocate these resources can be influenced by the environment. For example, greater male aggression may shift an existing trade-off between paternal care and territorial defense, such that more aggressive males provide less paternal care (Saino and Møller, 1995; Badyaev and Hill, 2002; Pryke and Griffith, 2009). We hypothesized that this trade-off could result in urban male song sparrows, which are reliably more aggressive, providing less paternal care. Decreased male care could, in turn, force females to increase maternal investment. However, if females are unable to compensate for reduced paternal care, offspring condition and survival could decrease. Contrary to the described theoretical framework, we found no evidence that urban male song sparrows sacrificed parental visitation for territorial aggression. Urban males did not have lower visitation rates than rural males at any nestling age or time of day (Figure 2). In fact, we found that the more aggressive urban males visited the nest more often than their rural counterparts and began feeding earlier in the day (Figure 2). Additionally, there was no correlation between male aggression and parental visitation rates for males or females within or across habitats (Figure 6; Lane and Sewall, 2022). This refutes both the idea that urban males are facing a trade-off between territorial defense and parental care, and that urban habitats compromise male parental care. Previously, researchers found that urban male songbirds in this population had better body condition and access to greater fat reserves compared to rural (Goodchild et al., 2022), suggesting that birds in urban habitats are of higher quality than their rural counterparts and are therefore released from constraints experienced by other birds (Reznick et al., 2000). Additionally, urban males likely have different time and energy budgets compared to rural. Our previous research has shown that urban song sparrows have decreased conspecific density and increased territory size compared to rural ones (McKinney, 2006; Davies and Sewall, 2016; Juárez et al., 2020). With larger territories to defend, but decreased conspecific density, urban male song sparrows may be more aggressive yet have fewer territorial encounters with neighbors each day. Though future studies of time budgets for urban and rural birds would be required to evaluate this possibility, we can conclude that neither rural nor urban song sparrows seem to be under sufficient constraint to force a behavioral trade-off between our measures of aggression and paternal care. However, it is important to point out that some of the most aggressive males in the population provided the least amount of care (Figure 1). It may be possible that with an even larger sample size the extremes in behavior would be detected and a trade-off between male aggression and parental care could be established.
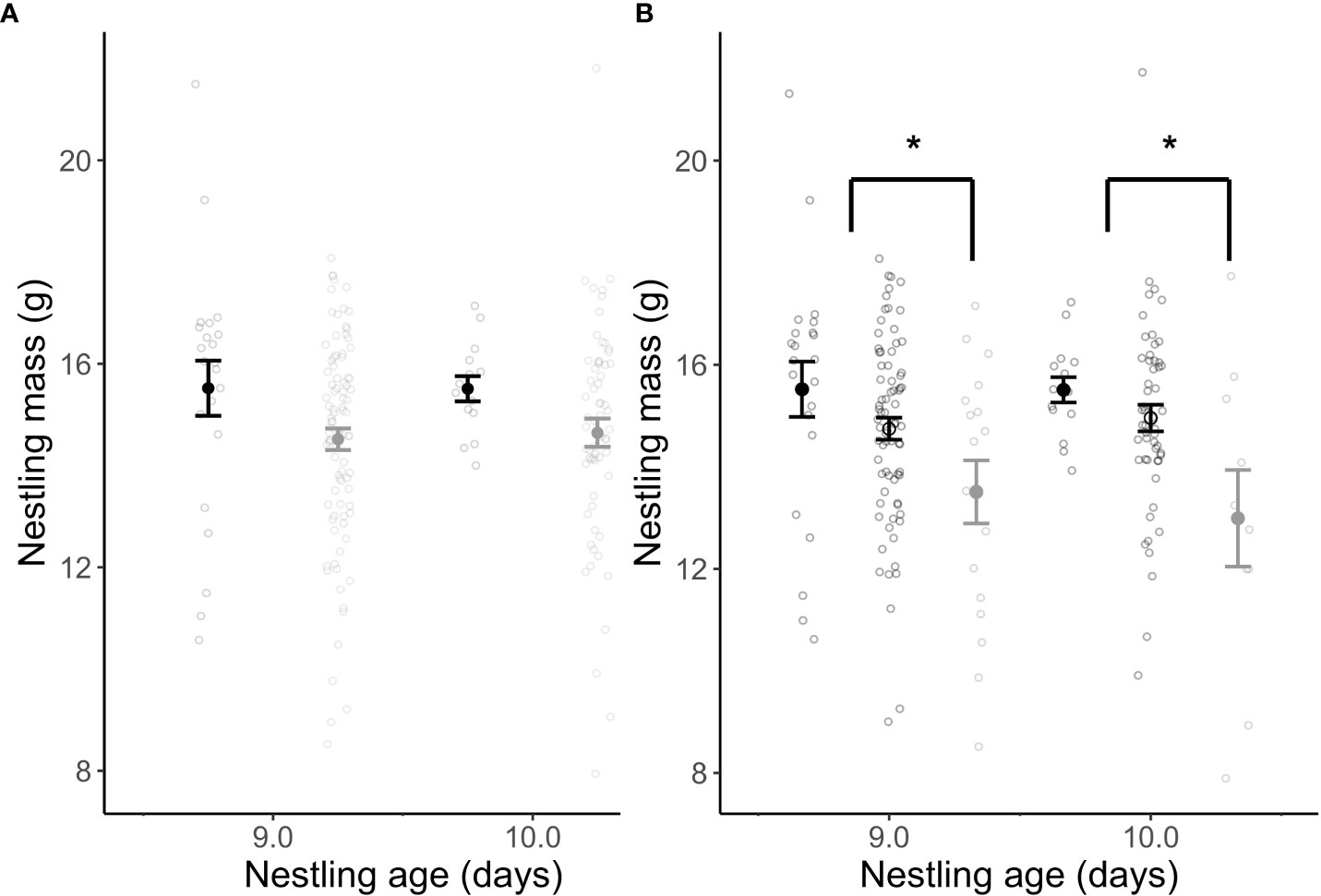
Figure 6 Nestling mass near fledging (Day 9 and 10) between habitat types (A) and in the presence of brood parasitism (B). Song sparrow nestling mass were similar in urban (a, gray) and rural (a, black) nests. However, song sparrow nestling mass is significantly lower in nests containing brown-headed cowbirds compared to those without. Rural (b, closed black) nestlings (15.51 ± 0.34) and urban (b, open black) nestlings (14.83 ± 0.17) were larger on average without brown-headed cowbird nestlings, compared to urban (b, closed grey) nestlings (13.31 ± 0.52) competing with brood parasite nestlings. Only 1 rural nest was documented with cowbird nestlings and was removed from the data for this comparison. The asterisks indicates a significance affect of brood parasitism on nestling growth. Parasitized nestling were significantly smaller compared to nestlings from non parasatized nests.
Female visitation rates were not significantly different across habitats, but there was a nonsignificant trend for urban females to visit the nest at a higher frequency (Figure 4). Specifically, urban females increased visitation rates during the day compared to rural, while rural females are more active at night compared to urban females (Figure 4). We were unable to confirm this night behavior was not an artifact of the RFID systems through video recordings because we did not have access to night vision equipment. However, we would expect to see similar patterns of behavior at night in both habitat types if the RFID systems were causing an artifact. Understanding nocturnal time budgets around species with different life-history strategies, such as cavity or open-cup nesting species, is a current gap in our understanding of animal behavior (Graham et al., 2017; Hau et al., 2017). Future studies examining daily patterns of behavior across urban and rural habitats in species with different life-history traits could determine which behaviors are adaptive under different environmental conditions and confirm or refute our surprising observation of nocturnal activity in rural song sparrows.
Since urban male song sparrows provide as much, if not more, paternal care as their rural counterparts we found no evidence that females were compensating for their mates. In fact, we found a strong positive relationship between parental care provided by social partners (Figure 5). Females that visited the nest more often each day were paired with males that did the same. Collectively, these parental care findings also refute the hypothesis that urban females are compensating for their social partners; rather, song sparrows seem to match their reproductive effort with that of their partner. This behavioral matching raises the hypothesis that song sparrow social pairs express assortative mating and/or matching of parental behaviors within the brooding periods (Wolfenbarger, 1999; Westneat et al., 2011). Our finding contrasts with a previous study in a cavity nesting species that found that urban birds showed decreased coordination in provisioning behavior compared to rural (Baldan and Ouyang, 2020). Rather, male song sparrows at our urban study sites maintain higher territorial aggression and high parental visitation, presumably because time and energy resources allow them to do so.
Song sparrow nestlings in urban and rural habitats had similar body size (tarsus and mass; Figure 6A) before fledging, suggesting that urban habitats are not negatively affecting offspring condition before fledging. However, offspring body mass (Figure 6B) and tarsus length near fledge were significantly smaller in parasitized nests compared to unparasitized nests. Additionally, in the subset of nests that avoided predation, hatching and fledging success of song sparrow nestlings was significantly reduced in parasitized nests compared to unparasitized nests. Obligate brood parasites, such as brown-headed cowbirds, lay their eggs in the nest of a host species, forcing the host to raise the nestling, often to the detriment of the parent and their offspring (Ortega et al., 2005; Faaborg et al., 2010). Nestlings that hatch later often die of malnutrition in nests with brood parasites, and only the largest, most robust host nestlings survive to fledge (Payne, 1977; Remeŝ, 2006). This may explain why nestling body size and mass at fledge was reduced, and why the number of host offspring that were able to hatch and fledge from the nest were significantly lower in nests with brown-headed cowbirds. Collectively, this suggests that urban parental care is adequate, if not elevated, compared to that of rural, but the elevated rates of brood parasitism in urban habitats still compromise their ability to fledge a full clutch of offspring, and reduce nestling body mass and size at fledge.
We found that urban nests had higher daily and annual survival rates across all five years of our study compared to rural (Table 1). Additionally, we found that urban song sparrows had greater nesting, hatching, and fledging success, and began nesting significantly earlier (16 days ± 2.27 days) compared to rural. This higher nestling survival was driven by significantly lower nest predation rates compared to rural. Our findings add to a growing body of evidence that, despite many studies demonstrating increased predator presence in urban habitats, nest predation rates decrease in urban settings compared to rural habitats (Eötvös et al., 2018; Fischer et al., 2012; Gering and Blair, 1999; Shochat et al., 2006). Nest predation may be lower in urban environments because predators can exploit anthropogenic food sources such as trash cans and human food supplementation (i.e., the Predator Paradox hypothesis; Fischer et al., 2020; Seress and Liker, 2015). Alternatively, or additionally, snakes are often important nest predators, and their conspecific density decreases in urban habitats (Klug and Jackrel, 2010). The difference we found in nest predation rates between habitat types could also influence the differences in urban parental visitation rates (Lyon and Montgomerie, 1987; Martin and Ghalambor, 1999; Eggers et al., 2006). Previous research has experimentally shown that increased nest predation pressure decreased nest visitation rates (Ghalambor and Martin, 2002; Mouton et al., 2020). Therefore, it is possible that rural birds limit their visitation rates to their nests to reduce the risk of attracting nest predators. Collectively, these results suggest that some urban generalist species, such as song sparrows, can have greater nesting success in urban habitats compared to rural ones. Resolving the impacts of abiotic aspects of urbanization, nest predation, and nest parasitism risk on parental behavior in songbirds would require future studies that experimentally manipulate these variables using brood size manipulations or manipulations of parental effort.
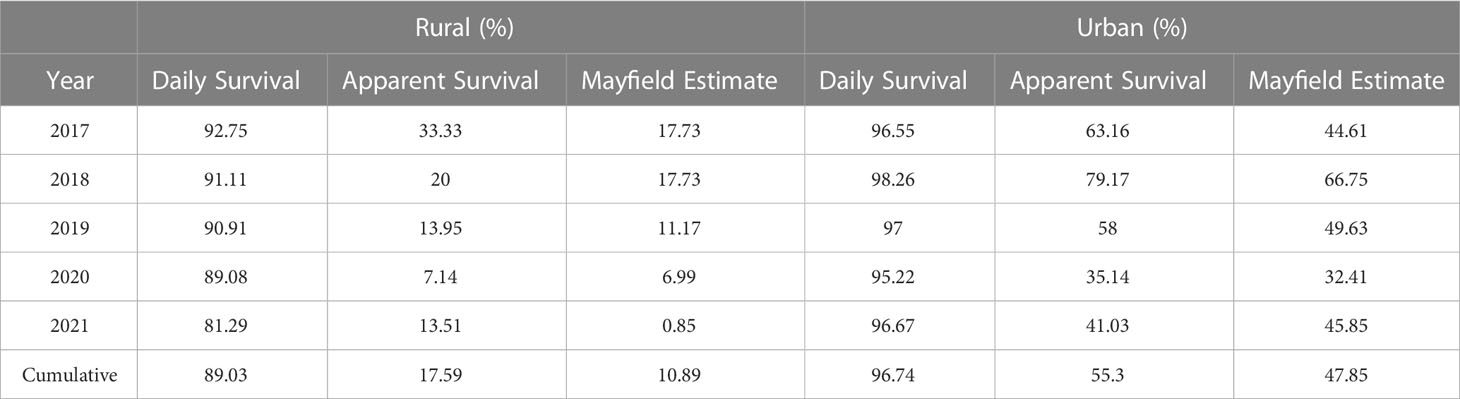
Table 1 Daily, apparent, and Mayfield estimations (Mayfield, 1961) of nest survival rates between urban and rural song sparrows from 2017 – 2021.
5 Conclusions
There is an underlying assumption in many urban ecology studies that urban habitats are detrimental to animal fitness compared to the rural, less disturbed environments. Our data join an increasing number of studies showing this is not the case for all species, and that urban habitats provide benefits to species that are able to invade these novel environments (McKinney, 2002; McKinney, 2008; Chamberlain et al., 2009). We found that more aggressive urban male song sparrows did not sacrifice parental care for territorial aggression. Consequently, their social partners are not suffering increased energetic demands to care for young. This indicates that either song sparrows do not face such a trade-off, or they are released from it in urban habitats. In fact, urban males visited the nest more often than their rural counterparts, a pattern also observed in females though the relationship was not significant. These behavioral differences could be the result of decreased nest predation pressure in urban habitats. Few studies have examined the effects of urbanization on parental care, and these studies have focused on incubation behavior in cavity nesting species (Baldan and Ouyang, 2020; Heppner and Ouyang, 2021; Hope et al., 2022). Hope et al., 2022 found that urban great tits (Parus major) increased overall incubation, yet decreased incubation bout rates compared to rural. Heppner and Ouyang (2021) found the opposite, with urban house wrens (Troglodytes aedon) decreasing incubation time and increasing bout rate. The limited number of studies in this area and their disparate results further illustrates the need for research on the effects of urbanization on parental care, and the importance of including species with diverse life history traits.
We observed a negative effect of brood parasitism on the hatching and fledging success of parasitized nests compared to unparasitized. However, this impact is minimal relative to the much higher nest success in urban habitats and the high nest predation in rural habitats. Understanding the effects of urbanization requires understanding community dynamics, including the interactions between brood parasites, predators, and their hosts. Previous studies found that urban adapters, such as the songbirds in this study, receive benefits such as increased food, water, maintained breeding substrates, and respite from predators (McKinney, 2002; Chamberlain et al., 2009; Shochat et al., 2010; Stracey and Robinson, 2012; Soulsbury and White, 2016; Reynolds et al., 2019). Collectively, our findings align with these studies, and suggest that, if given the space and opportunity to breed and reproduce, some generalist urban adapters and the species that rely on them can flourish in urban environments.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by Institutional Animal Care and Use Committee of Virginia Tech.
Author contributions
SL and KS conceived the ideas and designed methodology. SL, IV, VB, CL, and TF collected the data. SL and CG analyzed the data. SL, CG, and KS led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Funding
SL received funding from the Virginia Tech Ralph E. Carlson Memorial Scholarship in Ornithology, the Global Change Center at Virginia Tech, the Graduate Student Assembly at Virginia Tech, the Virginia Society for Ornithology, the American Ornithological Society, and the Society for Integrative and Comparative Biology. VB and CL received independent funding from the Multicultural Academic Opportunities Program, the Global Change Center at Virginia Tech and from the Howard Hughes Medical Institute’s 2014 Science Education grant 52008103 to New Mexico State University. This research was further supported by funding from National Science Foundation grant IOS 1652496 and 2114288
Acknowledgments
We thank Radford University, Virginia Tech’s Kentland Farms, and StREAM lab at Virginia Tech for allowing us to perform field research on their premises. We thank WA. Hopkins, J. McGlothlin, and IT. Moore for assistance on project development and comments on manuscript drafts and A. Gamero, and CE. Moore, for assistance in the field.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1234562/full#supplementary-material
References
Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Software 67 (1), 1–48. doi: 10.18637/jss.v067.i01
Adelman J. S., Moyers S. C., Hawley D. M. (2014). Using remote biomonitoring to understand heterogeneity in immune-responses and disease-dynamics in small, free-living animals. Am. Zoologist 54 (3), 377–386. doi: 10.1093/icb/icu088
Akçay Ç., Tom M. E., Holmes D., Campbell S. E., Beecher M. D. (2011). Sing softly and carry a big stick: signals of aggressive intent in the song sparrow. Anim. Behav. 82 (2), 377–382. doi: 10.1016/j.anbehav.2011.05.016
Arcese P. (1989). Intrasexual competition and the mating system in primarily monogamous birds: the case of the song sparrow. Anim. Behav. 38 (1), 96–111. doi: 10.1016/S0003-3472(89)80069-7
Atwell J. W., Cardoso G. C., Whittaker D. J., Price T. D., Ketterson E. D. (2014). Hormonal, behavioral, and life-history traits exhibit correlated shifts in relation to population establishment in a novel environment. Am. Nat. 184 (6), E147–E160. doi: 10.1086/678398
Badyaev A. V., Hill G. E. (2002). Paternal care as a conditional strategy: distinct reproductive tactics associated with elaboration of plumage ornamentation in the house finch. Behav. Ecol. 13 (5), 591–597. doi: 10.1093/beheco/13.5.591
Baker P. J., Molony S. E., Stone E., Cuthill I. C., Harris S. (2008). Cats about town: is predation by free-ranging pet cats Felis catus likely to affect urban bird populations? Ibis 150, 86–99. doi: 10.1111/j.1474-919X.2008.00836.x
Baldan D., Ouyang J. Q. (2020). Urban resources limit pair coordination over offspring provisioning. Sci. Rep. 10 (1), 1–11. doi: 10.1038/s41598-020-72951-2
Beck M. L., Akҫay Ҫ., Sewall K. B. (2019). Male song sparrows modulate their aggressive signaling in response to plumage signals: experiments with 3-D printed models. BioRxiv, 753772. doi: 10.1101/753772
Blair R. B. (1996). Land use and avian species diversity along an urban gradient. Ecol. Appl. 6, 506–519. doi: 10.2307/2269387
Bonier F. (2012). Hormones in the city: endocrine ecology of urban birds. Hormones Behav. 61, 763–772. doi: 10.1016/j.yhbeh.2012.03.016
Bridge E. S., Bonter D. N. (2011). A low-cost radio frequency identification device for ornithological research. J. Field Ornithology 82 (1), 52–59. doi: 10.1111/j.1557-9263.2010.00307.x
Bridge E. S., Wilhelm J., Pandit M. M., Moreno A., Curry C. M., Pearson T. D., et al. (2019). An Arduino-based RFID platform for animal research. Front. Ecol. Evol. 7, 257. doi: 10.3389/fevo.2019.00257
Brittingham M. C., Temple S. A. (1983). Have cowbirds caused forest songbirds to decline? BioScience 33 (1), 31–35. doi: 10.2307/1309241
Chace J. F., Walsh J. J. (2006). Urban effects on native avifauna: a review. Landscape urban Plann. 74 (1), 46–69. doi: 10.1016/j.landurbplan.2004.08.007
Chace J. F., Walsh J. J., Cruz A., Prather J. W., Swanson H. M. (2003). Spatial and temporal activity patterns of the brood parasitic brown-headed cowbird at an urban/wildland interface. Landscape Urban Plann. 64 (3), 179–190. doi: 10.1016/S0169-2046(02)00220-7
Chamberlain D. E., Cannon A. R., Toms M. P., Leech D. I., Hatchwell B. J., Gaston K. J. (2009). Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151 (1), 1–18. doi: 10.1111/j.1474-919X.2008.00899.x
Dammhahn M., Mazza V., Schirmer A., Göttsche C., Eccard J. A. (2020). Of city and village mice: behavioural adjustments of striped field mice to urban environments. Sci. Rep. 10 (1), 1–12. doi: 10.1038/s41598-020-69998-6
Davies S., Beck M. L., Sewall K. B. (2018). Territorial aggression in urban and rural Song Sparrows is correlated with corticosterone, but not testosterone. Hormones Behav. 98, 8–15. doi: 10.1016/j.yhbeh.2017.11.010
Davies S., Sewall K. B. (2016). Agonistic urban birds: elevated territorial aggression of urban song sparrows is individually consistent within a breeding period. Biol. Lett. 12 (6), 20160315. doi: 10.1098/rsbl.2016.0315
Duckworth R. A. (2006). Behavioral correlations across breeding contexts provide a mechanism for a cost of aggression. Behav. Ecol. 17 (6), 1011–1019. doi: 10.1093/beheco/arl035
Duckworth R. A. (2008). Adaptive dispersal strategies and the dynamics of a range expansion. Am. Nat. 172 (S1), S4–S17. doi: 10.1086/588289
Eggers S., Griesser M., Nystrand M., Ekman J. (2006). Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. Proc. R. Soc. B: Biol. Sci. 273 (1587), 701–706. 10.1098/rspb.2005.3373
Eötvös C. B., Magura T., Lövei G. L. (2018). A meta-analysis indicates reduced predation pressure with increasing urbanization. Landscape Urban Plann. 180, 54–59. doi: 10.1016/j.landurbplan.2018.08.010
Evans J., Boudreau K., Hyman J. (2010). Behavioural syndromes in urban and rural populations of song sparrows. Ethology 116 (7), 588–595. doi: 10.1111/j.1439-0310.2010.01771.x
Faaborg J., Holmes R. T., Anders A. D., Bildstein K. L., Dugger K. M., Gauthreaux S. A., et al. (2010). Recent advances in understanding migration systems of New World land birds. Ecol. Monogr. 80 (1), 3–48. doi: 10.1890/09-0395.1
Farr J. J., Haave-Audet E., Thompson P. R., Mathot K. J. (2021). No effect of passive integrated transponder tagging method on survival or body condition in a northern population of Black-capped Chickadees (Poecile atricapillus). Ecol. Evolution. doi: 10.1002/ece3.7783
Fischer J. D., Cleeton S. H., Lyons T. P., Miller J. R. (2012). Urbanization and the predation paradox: the role of trophic dynamics in structuring vertebrate communities. BioScience 62 (9), 809–818. doi: 10.1525/bio.2012.62.9.6
Fokidis H. B., Orchinik M., Deviche P. (2011). Context-specific territorial behavior in urban birds: no evidence for involvement of testosterone or corticosterone. Hormones Behav. 59 (1), 133–143. doi: 10.1016/j.yhbeh.2010.11.002
Foltz S. L., Ross A. E., Laing B. T., Rock R. P., Battle K. E., Moore I. T. (2015). Get off my lawn: increased aggression in urban song sparrows is related to resource availability. Behav. Ecol. 26 (6), 1548–1557. doi: 10.1093/beheco/arv111
Garcia J. T., Arroyo B. E. (2002). Intra-and interspecific agonistic behaviour in sympatric harriers during the breeding season. Anim. Behav. 64 (1), 77–84. doi: 10.1006/anbe.2002.3035
Gering J. C., Blair R. B. (1999). Predation on artificial bird nests along an urban gradient: predatory risk or relaxation in urban environments? Ecography 22 (5), 532–541. doi: 10.1111/j.1600-0587.1999.tb00542.x
Ghalambor C. K., Martin T. E. (2002). Comparative manipulation of predation risk in incubating birds reveals variability in the plasticity of responses. Behav. Ecol. 13 (1), 101–108. doi: 10.1093/beheco/13.1.101
Goodchild C. G., VanDiest I., Lane S. J., Beck M., Ewbank H., Sewall K. B. (2022). Variation in hematological indices, oxidative stress, and immune function among male song sparrows from rural and low-density urban habitats. Front. Ecol. Evol. 97. doi: 10.3389/fevo.2022.817864
Graham J. L., Cook N. J., Needham K. B., Hau M., Greives T. J. (2017). Early to rise, early to breed: a role for daily rhythms in seasonal reproduction. Behav. Ecol. 28 (5), 1266–1271. doi: 10.1093/beheco/arx088
Griffith S. C., Owens I. P., Thuman K. A. (2002). Extra pair paternity in birds: a review of interspecific variation and adaptive function. Mol. Ecol. 11 (11), 2195–2212. doi: 10.1046/j.1365-294X.2002.01613.x
Grimm N. B., Faeth S. H., Golubiewski N. E., Redman C. L., Wu J., Bai X., et al. (2008a). Global change and the ecology of cities. science 319 (5864), 756–760. doi: 10.1126/science.1150195
Grimm N. B., Foster D., Groffman P., Grove J. M., Hopkinson C. S., Nadelhoffer K. J., et al. (2008b). The changing landscape: ecosystem responses to urbanization and pollution across climatic and societal gradients. Front. Ecol. Environ. 6 (5), 264–272. doi: 10.1890/070147
Gunnarsson T. G., Gill J. A., Newton J., Potts P. M., Sutherland W. J. (2005). Seasonal matching of habitat quality and fitness in a migratory bird. Proc. R Soc. B: Biol. Sci. 272, 2319–2323. doi: 10.1098/rspb.2005.3214
Hau M., Dominoni D., Casagrande S., Buck C. L., Wagner G., Hazlerigg D., et al. (2017). Timing as a sexually selected trait: the right mate at the right moment. Philos. Trans. R. Soc. B: Biol. Sci. 372 (1734), 20160249. doi: 10.1098/rstb.2016.0249
Heppner J. J., Ouyang J. Q. (2021). Incubation behavior differences in urban and rural house wrens, Troglodytes aedon. Frontiers in Ecology and Evolution. 9 89. doi: 10.3389/fevo.2021.590069
Holtmann B., Santos E. S., Lara C. E., Nakagawa S. (2017). Personality-matching habitat choice, rather than behavioural plasticity, is a likely driver of a phenotype–environment covariance. Proc. R Soc. B: Biol. Sci. 284, 284. doi: 10.1098/rspb.2017.0943
Hope S. F., Hopkins W. A., Angelier F. (2022). Parenting in the city: effects of urbanization on incubation behaviour and egg temperature in great tits, Parus major. Anim. Behav. 194, 1–11. doi: 10.1016/j.anbehav.2022.09.004
Hyman J., Hughes M., Nowicki S., Searcy W. (2004). Individual variation in the strength of territory defense in male song sparrows: correlates of age, territory tenure, and neighbor aggressiveness. Behaviour 141 (1), 15–27. doi: 10.1163/156853904772746574
Jehle G., Yackel Adams A. A., Savidge J. A., Skagen S. K. (2004). Nest survival estimation: a review of alternatives to the Mayfield estimator. Condor 106 (3), 472–484. doi: 10.1093/condor/106.3.472
Jonsomjit D., Jones S. L., Gardali T., Geupel G. R., Gouse P. J. (2007). A guide to nestling development and aging in altricial passerines. US Fish & Wildlife Publications 161, 1–66.
Juárez R., Chacón-Madrigal E., Sandoval L. (2020). Urbanization has opposite effects on the territory size of two passerine birds. Avian Res. 11, 1–9. doi: 10.1186/s40657-020-00198-6
Ketterson E. D., Nolan J. V. (1994). Male parental behavior in birds. Annu. Rev. Ecol. Systematics 25 (1), 601–628. doi: 10.1146/annurev.es.25.110194.003125
Ketterson E. D., Nolan C. (1999). Adaptation, exaptation, and constraint: a hormonal perspective. Am. Nat. 154 (S1), S4–S25. doi: 10.1086/303280
Klug P. E., Jackrel S. L. (2010). Linking snake habitat use to nest predation risk in grassland birds: the dangers of shrub cover. Oecologia 162 (3), 803–813. doi: 10.1007/s00442-009-1549-9
Kuznetsova A., Brockhoff P. B., Christensen R. H. (2017). lmerTest package: tests in linear mixed effects models. J. Stat. software 82 (1), 1–26. doi: 10.18637/jss.v082.i13
Lane S. J., Sewall K. B. (2022). What about females? Urban female song sparrows elevate aggressive signaling compared to rural. Integr. Comp. Biol. 62 (3), 487–495. doi: 10.1093/icb/icac106
Linden M., Møller A. P. (1989). Cost of reproduction and covariation of life history traits in birds. Trends Ecol. Evol. 4 (12), 367–371. doi: 10.1016/0169-5347(89)90101-8
Loss S. R., Will T., Marra P. P. (2013). The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 4, 1396. doi: 10.1038/ncomms2380
Lowry H., Lill A., Wong B. B. (2013). Behavioural responses of wildlife to urban environments. Biol. Rev. 88 (3), 537–549. doi: 10.1111/brv.12012
Lüdecke D., Ben-Shachar M. S., Patil I., Waggoner P., Makowski D. (2021). performance: An R package for assessment, comparison and testing of statistical models. J. Open Source Software 6 (60). doi: 10.21105/joss.03139
Lyon B. E., Montgomerie R. D. (1987). Ecological correlates of incubation feeding: a comparative study of high arctic finches. Ecology 68 (3), 713–722. doi: 10.2307/1938477
Martin T. E. (1995). Avian life history evolution in relation to nest sites, nest predation, and food. Ecol. Monogr. 65 (1), 101–127. doi: 10.2307/2937160
Martin T. E., Briskie J. V. (2009). Predation on dependent offspring: a review of the consequences for mean expression and phenotypic plasticity in avian life history traits. Ann. New York Acad. Sci. 1168 (1), 201–217. doi: 10.1111/j.1749-6632.2009.04577.x
Martin T. E., Geupel G. R. (1993). Nest-Monitoring Plots: Methods for Locating Nests and Monitoring Success (Métodos para localizar nidos y monitorear el éxito de estos). J. Field Ornithology 64, 507–519. Available at: https://www.jstor.org/stable/4513862
Martin T. E., Ghalambor C. K. (1999). Males feeding females during incubation. I. Required by microclimate or constrained by nest predation? Am. Nat. 153 (1), 131–139. doi: 10.1086/303153
Marzluff J. M. (2001). “Worldwide urbanization and its effects on birds,” in Avian ecology and conservation in an urbanizing world, vol. 19-47. (Norwell, MA: Springer, Boston, MA).
McGlothlin J. W., Jawor J. M., Ketterson E. D. (2007). Natural variation in a testosterone-mediated trade-off between mating effort and parental effort. Am. Nat. 170 (6), 864–875. doi: 10.1086/522838
McKinney M. L. (2002). Urbanization, Biodiversity, and Conservation The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 52, 883–890. doi: 10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2
McKinney M. L. (2006). Urbanization as a major cause of biotic homogenization. Biol. Conserv. 127 (3), 247–260. doi: 10.1016/j.biocon.2005.09.005
McKinney M. L. (2008). Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst. 11 (2), 161–176. doi: 10.1007/s11252-007-0045-4
Møller A. P., Birkhead T. R. (1993). Certainty of paternity covaries with paternal care in birds. Behav. Ecol. Sociobiology 33 (4), 261–268. doi: 10.1007/BF02027123
Mouton J. C., Tobalske B. W., Wright N. A., Martin T. E. (2020). Risk of predation on offspring reduces parental provisioning, but not flight performance or survival across early life stages. Funct. Ecol. 34 (10), 2147–2157. doi: 10.1111/1365-2435.13650
Ortega C. R., Cruz A., Mermoz M. E. (2005). Issues and controversies of cowbird (Molothrus spp.) management. Ornithological Monogr. 2005, 6–15. doi: 10.2307/40166810
Otter K., Ratcliffe L. (1996). Female initiated divorce in a monogamous songbird: abandoning mates for males of higher quality. Proc. R. Soc. London. Ser. B: Biol. Sci. 263 (1368), 351–355. doi: 10.1098/rspb.1996.0054
Paredes R., Jones I. L., Boness D. J. (2005). Reduced parental care, compensatory behaviour and reproductive costs of thick-billed murres equipped with data loggers. Anim. Behav. 69 (1), 197–208. doi: 10.1016/j.anbehav.2003.12.029
Payne R. B. (1977). The ecology of brood parasitism in birds. Annu. Rev. Ecol. systematics 8, 1–28. doi: 10.1146/annurev.es.08.110177.000245
Peters S. S., Searcy W. A., Marler P. (1980). Species song discrimination in choice experiments with territorial male swamp and song sparrows. Anim. Behav. 28 (2), 393–404. doi: 10.1016/S0003-3472(80)80048-0
Pryke S. R., Griffith S. C. (2009). Socially mediated trade-offs between aggression and parental effort in competing color morphs. Am. Nat. 174 (4), 455–464. doi: 10.1086/605376
Pyle P. (1997). Identification guide to North American birds: a compendium of information on identifying, ageing, and sexing" near-passerines" and passerines in the hand (Point Reyes. Station, CA: Slate Creek Press).
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Remeŝ V. (2006). Growth strategies of passerine birds are related to brood parasitism by the brown-headed cowbird (Molothrus ater). Evolution 60 (8), 1692–1700. doi: 10.1111/j.0014-3820.2006.tb00513.x
Renthlei Z., Borah B. K., Trivedi A. K. (2017). Effect of urbanization on daily behavior and seasonal functions in vertebrates. Biol. Rhythm. Res. 48 (5), 789–804. doi: 10.1080/09291016.2017.1345462
Reynolds J. S., Ibáñez-Álamo J. D., Sumasgutner P., Mainwaring M. C. (2019). Urbanisation and nest building in birds: a review of threats and opportunities. J. Ornithology 160 (3), 841–860. doi: 10.1007/s10336-019-01657-8
Reznick D., Nunney L., Tessier A. (2000). Big houses, big cars, superfleas and the costs of reproduction. Trends Ecol. Evol. 15 (10), 421–425. doi: 10.1016/S0169-5347(00)01941-8
Robinson S. K., Thompson F. R., Donovan T. M., Whitehead D. R., Faaborg J. (1995). Regional forest fragmentation and the nesting success of migratory birds. Science 267 (5206), 1987–1990. doi: 10.1126/science.267.5206.1987
Rodewald A. D. (2009). Urban-associated habitat alteration promotes brood parasitism of Acadian Flycatchers. J. Field Ornithology 80 (3), 234–241. doi: 10.1111/j.1557-9263.2009.00226.x
Roff D. A., Fairbairn D. J. (2007). The evolution of trade-offs: where are we? J. evolutionary Biol. 20 (2), 433–447. doi: 10.1111/j.1420-9101.2006.01255.x
Roff D. A., Remeš V., Martin T. E. (2005). The evolution of fledging age in songbirds. J. Evolutionary Biol. 18 (6), 1425–1433. doi: 10.1111/j.1420-9101.2005.00958.x
Rosenberg K. V., Dokter A. M., Blancher P. J., Sauer J. R., Smith A. C., Smith P. A., et al. (2019). Decline of the North American avifauna. Science 366 (6461), 120–124. doi: 10.1126/science.aaw1313
Saino N., Møller A. P. (1995). Testosterone-induced depression of male parental behavior in the barn swallow: female compensation and effects on seasonal fitness. Behav. Ecol. Sociobiology 36 (3), 151–157. doi: 10.1007/BF00177791
Saunders D. A., Hobbs R. J., Margules C. R. (1991). Biological consequences of ecosystem fragmentation: a review. Conserv. Biol. 5 (1), 18–32. doi: 10.1111/j.1523-1739.1991.tb00384.x
Scales J., Hyman J., Hughes M. (2011). Behavioral syndromes break down in urban song sparrow populations. Ethology 117 (10), 887–895. doi: 10.1111/j.1439-0310.2011.01943.x
Scales J., Hyman J., Hughes M. (2013). Fortune favours the aggressive: territory quality and behavioural syndromes in song sparrows, Melospiza melodia. Anim. Behav. 85 (2), 441–451. doi: 10.1016/j.anbehav.2012.12.004
Scolozzi R., Geneletti D. (2012). A multi-scale qualitative approach to assess the impact of urbanization on natural habitats and their connectivity. Environ. Impact Assess. Rev. 36, 9–22. doi: 10.1016/j.eiar.2012.03.001
Seress G., Liker A. (2015). Habitat urbanization and its effects on birds. Acta Zoologica Academiae Scientiarum Hungaricae 61 (4), 373–408. doi: 10.17109/AZH.61.4.373.2015
Seress G., Lipovits Á., Bókony V., Czúni L. (2014). Quantifying the urban gradient: a practical method for broad measurements. Landscape Urban Plann. 131, 42–50. doi: 10.1016/j.landurbplan.2014.07.010
Shochat E., Lerman S., Fernández-Juricic E. (2010). Birds in urban ecosystems: population dynamics, community structure, biodiversity, and conservation. Urban ecosystem Ecol. 55, 75–86. doi: 10.2134/agronmonogr55.c4
Shochat E., Warren P. S., Faeth S. H., McIntyre N. E., Hope D. (2006). From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21 (4), 186–191. doi: 10.1016/j.tree.2005.11.019
Smith J. N., Arcese P. (1994). Brown-headed cowbirds and an island population of song sparrows: a 16-year study. Condor 96 (4), 916–934. doi: 10.2307/1369102
Smith J. N., Roff D. A. (1980). Temporal spacing of broods, brood size, and parental care in Song Sparrows (Melospiza melodia). Can. J. Zoology 58 (6), 1007–1015. doi: 10.1139/z80-141
Sogge M. K., Van Riper C. (1988). Breeding biology and population dynamics of the San Miguel Island Song Sparrow (Melospiza melodia micronyx) (Davis, CA; Cooperative National Park Resources Studies Unit, University of California, Institute of Ecology).
Soulsbury C. D., White P. C. (2016). Human–wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildlife Res. 42 (7), 541–553. doi: 10.1071/WR14229
Stamps J. A., Krishnan V. V. (1997). Functions of fights in territory establishment. Am. Nat. 150 (3), 393–405. doi: 10.1086/286071
Stracey C. M., Robinson S. K. (2012). Are urban habitats ecological traps for a native songbird? Season-long productivity, apparent survival, and site fidelity in urban and rural habitats. J. Avian Biol. 43 (1), 50–60. doi: 10.1111/j.1600-048X.2011.05520.x
van Heezik Y., Smyth A., Adams A., Gordon J. (2010). Do domestic cats impose an unsustainable harvest on urban bird populations? Biol. Conserv. 143 (1), 121–130. doi: 10.1016/j.biocon.2009.09.013
Venables W. N., Ripley B. D. (2002). Modern Applied Statistics with S, Fourth edition (New York: Springer). Available at: https://www.stats.ox.ac.uk/pub/MASS4/, ISBN: ISBN 0-387-95457-0.
Westneat D. F. (1987). Extra-pair fertilizations in a predominantly monogamous bird: genetic evidence. Anim. Behav. 35 (3), 877–886. doi: 10.1016/S0003-3472(87)80123-9
Westneat D. F., Hatch M. I., Wetzel D. P., Ensminger A. L. (2011). Individual variation in parental care reaction norms: integration of personality and plasticity. Am. Nat. 178 (5), 652–667. doi: 10.1086/662173
Whittingham L. A., Dunn P. O., Robertson R. J. (1994). Female response to reduced male parental care in birds: an experiment in tree swallows. Ethology 96 (3), 260–269. doi: 10.1111/j.1439-0310.1994.tb01014.x
Winkler D. W., Hallinger K. K., Pegan T. M., Taff C. C., Verhoeven M. A., Chang van Oordt D., et al. (2020). Full lifetime perspectives on the costs and benefits of lay-date variation in tree swallows. Ecology 101 (9), e03109. doi: 10.1002/ecy.3109
Keywords: fitness, parental care, territorial aggression, trade-offs, urbanization
Citation: Lane SJ, VanDiest IJ, Brewer VN, Linkous CR, Fossett TE, Goodchild CG and Sewall KB (2023) Indirect effects of urbanization: consequences of increased aggression in an urban male songbird for mates and offspring. Front. Ecol. Evol. 11:1234562. doi: 10.3389/fevo.2023.1234562
Received: 04 June 2023; Accepted: 18 July 2023;
Published: 22 August 2023.
Edited by:
Zoltan Elek, University of Szeged, HungaryReviewed by:
Attila Marton, Babeș-Bolyai University, RomaniaZoltán Németh, University of Debrecen, Hungary
Copyright © 2023 Lane, VanDiest, Brewer, Linkous, Fossett, Goodchild and Sewall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Samuel J. Lane, c2FtdWVsbGFuZTQ5NkBnbWFpbC5jb20=
†Present address: Samuel J. Lane, Department of Biological Sciences, North Dakota State University, Fargo, ND, United States
Valerie N. Brewer, Department of Integrative Biology, Oregon State University, Corvallis, OR, United States
Christopher G. Goodchild, Department of Biology, University of Central Oklahoma, Edmond, OK, United States