 David García de León
David García de León José M. Rey Benayas
José M. Rey Benayas Pedro Villar-Salvador
Pedro Villar-Salvador
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 28 August 2023
Sec. Conservation and Restoration Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1206000
Conservation and restoration of hedgerows promote biodiversity and multi-functionality in agricultural landscapes. However, in Mediterranean environments, natural regeneration of hedgerows is often poor, and factors hindering the establishment and growth of hedgerow woody species remain unclear. This study aimed to address this gap by (1) quantifying natural regeneration, including seedling establishment and resprouting of planted hedgerow species and the establishment of woody species coming from outside the hedgerows; and (2) experimentally assessing the impact of drought, herbivory, and herb competition on seedling establishment. We selected 12 planted, 11–13-year-old hedgerows in four sites in Central Spain to sample their natural regeneration. In addition, we transplanted 1,356 seedlings of Colutea arborescens L., Crataegus monogyna Jacq. and Rhamnus alaternus L. to two field sites under three treatments that were applied factorially for two years: irrigation to alleviate summer drought, protection against herbivory, and herb clipping to reduce competition with transplanted seedlings. We found that only 66 juveniles of woody species, including 29 of species that were not planted in the hedgerows, developed from seed germination, and 316 sprouts recruited spontaneously along 10 years (14 individuals ha−1 year−1). Of the transplanted seedlings, only 22% survived after two years with protection against herbivory strongly enhancing seedling survival. To a lesser extent, irrigation and herb competition affected survival through interactions with herbivory and species identity. Irrigation and protection against herbivores accelerated seedling growth. Survival and growth of C. arborescens (14%; seedling height = 12 ± 9 cm) were lower than those of C. monogyna (41%; 21 ± 11 cm) and R. alaternus (17%; 13 ± 8 cm). We conclude that natural regeneration in Mediterranean planted hedgerows is low, herbivory is the main limiting factor for seedling survival, especially for deciduous species, and summer drought and herb competition mostly limit growth. These findings have important implications for the conservation and restoration of hedgerows in Mediterranean environments.
Agricultural production must be more sustainable to redress its negative impact on biodiversity and climate (IPBES, 2019; Pörtner et al., 2021). To achieve this goal, the EU Biodiversity Strategy for 2030 aims to convert at least 10% of farmland into high-rich landscape elements (European Commision, 2020). These landscape elements encompass several non-cultivated habitats of woody vegetation, such as hedgerows and forest patches (Rey Benayas and Bullock, 2012; Rey Benayas and Bullock, 2015).
Hedgerows are linear landscape green infrastructure commonly found around agricultural fields (Lajos et al., 2020). They can occur naturally or be planted. Hedgerows contribute to the multi-functionality of agricultural landscapes by promoting biodiversity, carbon sequestration, run-off control and nutrient regulation, and providing resources such as forage (García de León et al., 2021). Hence, the conservation and restoration of hedgerows should be prioritized in many agricultural landscapes worldwide. However, there are multiple socio-ecological factors that can limit the conservation and restoration (McCarthy, 1994; Footitt et al., 2018). In Mediterranean regions, natural regeneration of hedgerows is generally poor (Rey Benayas et al., 2021). The recruitment of plants in or near hedgerows in Mediterranean environments and the underlying demographic processes and their interaction with environmental factors and plant functional types are not well understood.
Drought stress is a major limitation for plant establishment in Mediterranean ecosystems (Rey Benayas et al., 2002; Granda et al., 2014). However, the impact of herbaceous competition on seedling survival of woody species in hedgerows remains controversial. While some studies indicate that herb competition significantly limits hedgerow regeneration (Kollmann and Reiner, 1996), others have found little to no effect (McCarthy, 1994). Herbs, however, often exacerbate the negative effects of summer drought stress on planted seedlings in Mediterranean cropland afforestation (Rey Benayas et al., 2005; Cuesta et al., 2010).
The presence of established vegetation in the landscape may facilitate natural regeneration of hedgerows through various mechanisms. Firstly, hedgerow plants produce seeds, and the effectiveness of colonization depends on distance. According to Janzen-Connell´s hypothesis, seedling recruitment increases with distance from mother plants up to a maximum value, after which it decreases due to dispersal limitations (Janzen, 1970; Deckers et al., 2005; Jacquemyn et al., 2010). However, several factors can distort this pattern at different scales. Conspecific adults can reduce the survival of the seedlings growing under their canopy through competition (Jacquemyn et al., 2010). Additionally, predators may forage under reproductive adults, attracted by the high abundance of seeds and seedlings, thereby reducing juvenile density near mother plants (Deckers et al., 2005). This effect is more pronounced in plants with fleshy fruits that are dispersed far from mother plants by birds, carnivore mammals, and herbivores (Suárez-Esteban et al., 2013; González-Varo et al., 2015). Secondly, although conspecific competition can hinder seedling establishment, facilitative interactions between plant species are common in Mediterranean environments and promote the establishment of juveniles, especially in late successional woody species (Cuesta et al., 2010; Hegland et al., 2021). Thirdly, hedgerows can influence herbivore activity. The structural complexity of hedgerows, which changes over time, can modify animal–plant interactions and impact the effect of herbivory on woody plant recruitment and performance (Endels et al., 2004; Suárez-Esteban et al., 2013; Dainese et al., 2015). For instance, more complex hedgerows can provide refugia and act as a reservoir for herbivores such as slugs (Marshall et al., 1999) and rabbits (Oryctolagus cuniculus).
Functional attributes drive the growth, competitiveness, and stress resistance of plant species in specific environments (Soliveres et al., 2014; Martínez-Vilalta et al. (2010); López-Iglesias et al., 2014). Evergreen and deciduous woody species often coexist in Mediterranean forest ecosystems (Fyllas et al., 2020). Deciduous Mediterranean woody species usually have higher drought mortality and are more susceptible to herbivory than their evergreen counterparts (López-Iglesias et al., 2014). The lower performance of deciduous woody plants can be attributed to their higher specific leaf area and growth rate (Paal et al., 2020), which are traits negatively associated with stress resistance (López-Iglesias et al., 2014; Andivia et al., 2021). However, their higher growth rate can enhance competitiveness when growing with weeds (Suárez-Esteban et al., 2013). Additionally, legumes have higher nitrogen concentration than other plant families, making them highly palatable to herbivores (Sánchez-Moreno et al., 2018). The interaction between functional attributes and environmental factors in determining plant establishment in hedgerows has been little explored.
This study aims to (1) quantify natural regeneration of planted species in hedgerows, including seed-based and resprout regeneration, as well as the establishment of species originating from outside the planted hedgerows; and (2) experimentally evaluate the interaction of limiting environmental factors that influence the seedling establishment for three functionally distinct woody seedlings in Mediterranean planted hedgerows. We hypothesized (H) and predicted (P) that ecological factors that shape the agricultural landscape determine plant regeneration in hedgerows (H1), and summer drought, herbivory, and herb competition synergistically limit recruitment of woody seedlings (P1); there is interspecific variation in species survival, growth, and recruitment (H2), and different species will respond differently to these limiting factors (P2), with context factors at the study site level influence early seedling establishment (P3). By providing insights into these aspects, the outcomes of this study will contribute to the design and establishment of hedgerows in Mediterranean environments.
The following four plots were studied with well-developed planted hedgerows located in Central Spain (Figure 1A):
● San Carlos del Valle (Ciudad Real), site “Vista Alegre”, olive orchard (38°48´N, 3°13´W; 837 m a.s.l.; 4.55 ha; Figure 1B).
● Valdepeñas (Ciudad Real), site “El Peral”, olive orchard (38°48´N, 3°21´W; 705 m a.s.l.; 1.76 ha; Figure 1C).
● Novés (Toledo), site “Fuente del Albañal”, olive orchard (40°3´N, 4°17´W; 538 m a.s.l.; 2.65 ha; Figure 1D).
● Novés (Toledo), site “Los Billares”, barley field (40°1´N, 4°14´W; 538 m a.s.l.; 12.66 ha; Figure 1E).
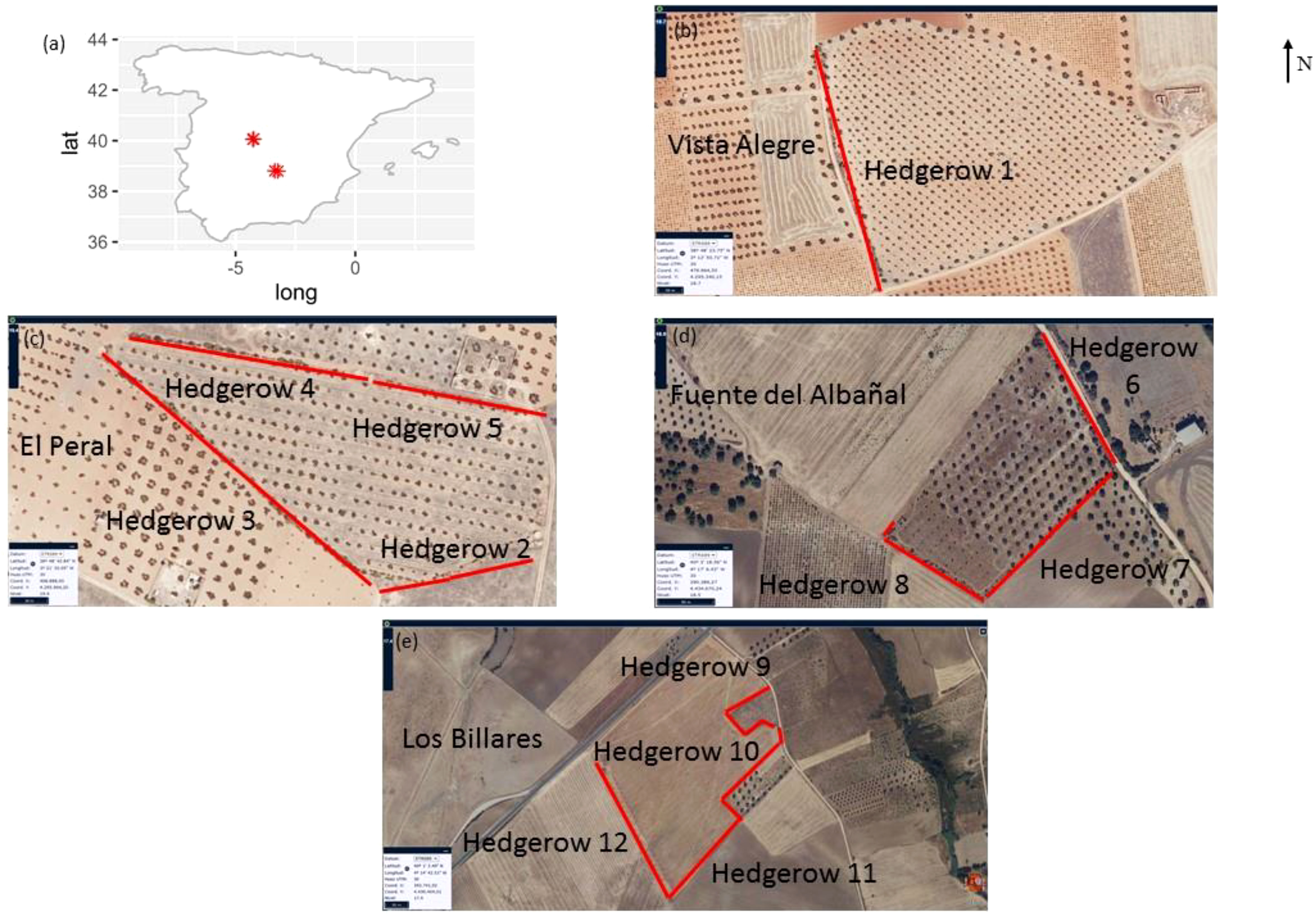
Figure 1 Location in peninsular Spain (A) and aerial photographs of the four studied fields taken from SIGPAC https://sigpac.mapa.es/fega/visor/. The fields correspond to sites “Vista Alegre” (B), “El Peral” (C), “Fuente del Albañal” (D), and “Los Billares” (E).
The four sites were managed as organic farms. The olive orchard sites included a green cover of spontaneous herbs and were mowed and fertilized only once every 4–5 years. “Los Billares” followed the rotation cereal–leguminous–fallow sequence. Adjacent to the hedgerows, there were a barley field (Los Billares), a grassland (El Peral), olive orchards (every site), a sunflower field (Vista Alegre), vineyards (Los Billares), and a winter wheat field (Los Billares). The annual crops were yearly ploughed, the olive orchards were extensive farming systems, and the grassland was not managed at all.
The studied sites have a continental Mediterranean climate characterized by cold winters and a long summer drought that imposes a severe water stress to the vegetation. In Valdepeñas and S. Carlos del Valle, total annual precipitation averaged 343 mm and mean annual air temperature averaged 15.3 °C during the period 2010–2021. In Novés, these values were 272 mm and 14.8 °C, respectively, during the same period. Soils of all sites were deep and fertile. The first two were a cambisol type and come from limestone, whereas the last two were a deep luvisol type and come from arkoses. Mean values of soil physical and chemical properties are reported in Table S1.
Twelve hedgerows distributed among the four sites were sampled. Total richness in the planted hedgerows was 22 woody plant species (17 planted and 5 non-planted species; details in the Results). Rosa canina L., Retama sphaerocarpa (L.) Boiss., Colutea arborescens, and Crataegus monogyna represented 15%, 10%, 9% and 8%, respectively, out of 748 adult individuals in the plots sampled in 2019. Site “Vista Alegre” harbored one hedgerow, “Fuente del Albañal” harbored three hedgerows, and sites “El Peral” and “Los Billares” four hedgerows each (Figures 1B–E). Each hedgerow was distinguished from others by natural discontinuities such as corners and stone piles. Hedgerows were planted in 2009 (sites “Vista Alegre” and “Fuente del Albañal”), 2010 (site “El Peral”) and 2011 (site “Los Billares”). Hedgerows were 85 to 273 m long and 2.5 m wide when they were sampled in 2019. Initial cover of woody plants, height (h), the perpendicular diameters of the crown, and the number of fruits of all planted individuals in the hedgerows were measured between 19th and 24th September 2019. The number of fruits were categorized as 0 (without fruits), 1 (1–10 fruits), 2 (11–100 fruits), and 3 (> 100 fruits) in September 2019. Fruit production data was scaled to the plant volume.
We first assessed natural regeneration in and around the planted hedgerows. A first in toto sampling was conducted in the 12 selected hedgerows and surrounding fields between the 21st of June and 30th of July 2019. Conspicuous new, non-planted individuals at the hedgerows were identified. In addition, a detailed second sampling was done in 44 transects of 10 × 2.5 m located every 50 m along the hedgerows. In this sampling event, all woody plants that were found were noted, distinguishing the plants derived from seeds (located at > 20 cm away between the crown edge of an adult of the same species and the crown edge of a recruit) and the resprouts of the planted individuals (< 20 cm away from an adult plant of the same species). To sample the vegetation around the planted hedgerows, the observers walked around for 20 minutes in each of the adjacent fields (1 ha) to each hedgerow and identified every woody plant individual. The ability of each planted species to resprout (Pausas et al., 2016) was used as an additional criterion to differentiate between the plants originating from seeds and those from resprouting. In every plot, mean woody vegetation cover, distance from recruited plants to the closest reproductive plant of the same species – regardless of transect – and the biometric data for both types of new established plants were registered.
A multifactorial field experiment was established to determine the interactive effects of limiting factors of the establishment of woody plant seedlings in the hedgerows. For this experiment, we selected “Los Billares” and “El Peral” sites. The treatments were applied on seedlings produced in nurseries and transplanted to the field of the species bladder-senna Colutea arborescens L., hawthorn Crataegus monogyna Jacq. and Mediterranean buckthorn Rhamnus alaternus L. These three species are shrubs and represent different functional types of species that were previously planted in the hedgerows. Specifically, they represented 9%, 8%, and 1%, respectively, of all hedgerow’s individuals. C. arborescens is a deciduous, fast-growing legume. C. monogyna is deciduous, grows more slowly (personal observations) than C. arborescens and produces fleshy fruits. Finally, R. alaternus is an evergreen and drought tolerant species with scleromorphic leaves that produces fleshy fruits. We tested three treatments: summer irrigation (I− and I+) to reduce summer drought; seedling protection (P− and P+) to exclude herbivory by rabbits, the most common herbivore in the studied sites; and herb clipping (C− and C+) in a 25-cm radius around each planted seedling to reduce competition (see details below). Herbivory pressure was estimated by counting rabbit latrines in September 2020, 2021, and 2022 and in February 2021 and 2022.
Some 1,356 seedlings were transplanted following the replication scheme in Table 1, namely 30 to 33 individuals of each species at Los Billares site and between 18 and 32 individuals at El Peral site. The unbalanced number of transplanted individuals at both sites was due to plant availability. Transplanting to the field was conducted between the 5th of October and the 20th of November 2020. Seedlings were 7- to 8-months-old and 5 to 15 cm in height. Seedlings were planted in gaps within the hedgerow plantation lines. All possible combinations of treatments were assured, and each transplanted seedling was at least 0.5 m far from any other transplanted seedling to avoid that treatment applied to one individual affected their transplanted neighbors.
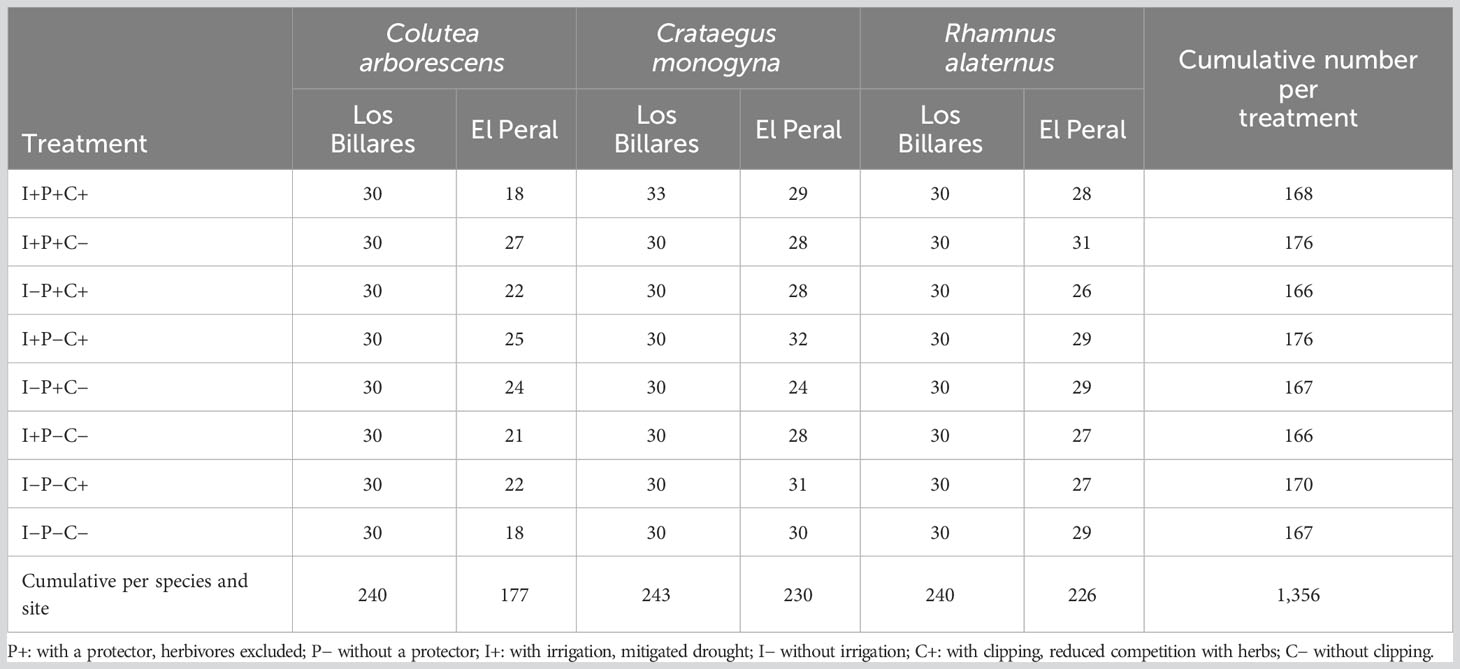
Table 1 Number of planted individuals per species, treatment, and site in the field multifactorial experiment.
Production of C. monogyna and R. alaternus seedlings was ordered to a forest nursery (Viveros Alborada; https://www.viverosalborada.com/) and plants were grown in containers of 375 mL volume. Seedlings of C. arborescens were cultivated by us in the nursery of the Real Jardín Botánico Juan Carlos I of Universidad de Alcalá due to its unavailability at commercial nurseries. The climate in this nursery is similar to that in the commercial nursery. To produce C. arborescens seedlings, 150 g of seeds from the Centro Nacional de Recursos Genéticos Forestales “El Serranillo” were used. Seeds were pretreated by immersion in H2SO4 (96–98%) for 20 min. Next, seeds were washed to eliminate burnt leftovers tissues and extended on blotting paper for 24 hours. Finally, the seeds that had not swollen were selected. Selected seeds were placed on germination trays, filled with a mix of peat and coconut fiber (Projar PS SEED PRO-5050 A0 pH 5.5–6, Madrid, Spain) and washed-river sand in proportion 3:2 at 1-cm depth in February 2020. The germination of C. arborescens was >80%. Once seeds germinated, they were transplanted into 20 rigid trays of 35 cells (Herku QP 35 T, 5 × 5 × 11.5 cm; volume 287 mL, Madrid, Spain) and were watered on demand until November 2020. Anti-snail poison was applied once to all trays to control a pest at the UAH nursery facility.
The treatments were as follows: (1) Irrigation was applied eleven times: five from the 8th of June to the 24th of August 2021 and six from the 23rd of May to the 29th of August 2022. Every year, the irrigation started with 8 L/m2. Two weeks later, irrigation dose was risen to 12 L/m2 due to the increase of temperature and hence potential evapotranspiration and water demand. Two weeks later, irrigation dose was increased to 16 L/m2 every 3 weeks until the end of each summer. The irrigation dose was determined to maximize watering in a logistically feasible manner. (2) The protection against herbivory was applied at the time of transplanting. We used cylindric nets with a 1 cm mesh. The diameter was 25 cm and a height of 30 cm above the ground. (3) The reduction of herb competition by clipping was applied five times, namely between the 12th and 21st of March 2021, 1st and the 18th of May 2021, 15th and 29th of November 2021, 5th and 19th March 2022, and 6th and 11th of May 2022. Clipped herb biomass was dried in the oven at 60°C for 1 week and weighed.
Herb biomass was similar in “Los Billares” and in “El Peral” (mean cumulative total biomass ± se was 0.25 ± 0.01 and 0.24 ± 0.01 kg m−2, respectively; Mann Whitney U, W = 5,760; p = 0.89). Overall, there were more herb biomass in the non-irrigated treatments than in the irrigated treatments (W = 6,662; p = 0.05). There were not significant differences in herb biomass that grew with and without protectors (W = 4,383; p = 0.91).
Pressure of herbivores was estimated by counting the number of rabbit latrines along the hedgerow perimeter. There were not available data on herbivory pressure at the beginning of this study; however, we estimated that there were more rabbits in “Fuente del Albañal” site than in the other three sites. In September 2020, we found seven rabbit latrines in “Los Billares”, whereas only one latrine was found in the olive orchard of “El Peral”. However, in February 2021, September 2021, February 2022, and September 2022, we observed 3, 5, 3, and 6 latrines in “Los Billares” and 10, 15, 6 and 19 latrines in “El Peral”, respectively (Repeated-measures ANOVA; Site: F1,4 = 2.69, p = 0.18; Date: F4,4 = 0.99, p = 0.51).
Survival counting took place in mid-February 2021, on the last week of April 2021, first week of September 2021, at mid-February 2022, mid-May 2022, and mid-September 2022. A seedling was considered dead when the stems were completely dry and it did not resprout in consecutive measurements.
The height of the established seedlings was measured in 4th to 5th September 2021, 12th to 13th February 2022, and 14th September 2022. Seedling growth was estimated as the height in September 2022 minus height in September 2021.
Seedling survival was assessed by a survival analysis that used a Cox’s proportional risk regression model as it is implemented in function coxph of survival R package (Therneau and Grambsch, 2000; Therneau, 2021). Model predictors were site (“Los Billares”, “El Peral”), hedgerow age (11 or 13 years), species (C. arborescens, C. monogyna, R. alaternus), and the three experimental treatments. To determine relevant main factors and their interaction terms, we used the null and the full starting models with forward and backward selection following function stepAIC in R package MASS (Venables and Ripley, 2002). We considered the use of mixed models, but we decided to use linear models because the possible random factor “site” had fewer than five levels (Zuur et al., 2009) and it accounted for the variability related to a specific prediction (i.e., context factors, P3).
All seedlings were of the same age and the initial (November 2020) difference in height was distributed at random among treatments and replicates. The effects of nearest neighbors established before 2019 were assumed negligible due to the low density of natural recruits. To estimate studied factors’ effects on seedling growth, a linear model was fitted with the species and treatment as predictors [i.e., other main effects or interactions terms could not be assessed due to a small sample size (n=8)] using function lm from R package stats (R Core Team, 2021). It was unfeasible to estimate growth in height for the dead seedlings and a substantial number of seedlings died along the experiment. Thus, the average growth per treatment on the alive seedling basis was quantified. Statistical significance of studied factors was tested with a multifactorial ANOVA using function Anova from R package car, with an error type-III structure to avoid relevance of the order of factors (Fox and Weisberg, 2019). Variation partitioning was applied to disentangle the proportion of variance explained by every predictor on survival and growth with function varpart from vegan R package (Oksanen et al., 2022).
It was expected that irrigation increased herb biomass, which competes against the transplanted seedlings. Thus, a Mann-Whitney’s U was used to test whether there were significant differences in herb biomass between treatments (I−/I+ and P−/P+). In addition, cumulative biomass was calculated as the sum of dry weight eliminated in every clipping event. Due to the random distribution of samples one can assume initial dry biomass was similar around clipped (C+) and non-clipped (C−) seedlings.
In summer 2019, woody-plant cover in the 44 sampled transects in the hedgerows averaged (± se) 61 ± 1%. Average plant height, major diameter (D1), and perpendicular diameter to the major diameter (d2) of adult plants were 96 ± 2 cm, 88 ± 4 cm, and 66 ± 63 cm, respectively. Such measures led to an average crown volume [height × D1 × d2] of 1.99 ± 0.20 m3 (Table 2).
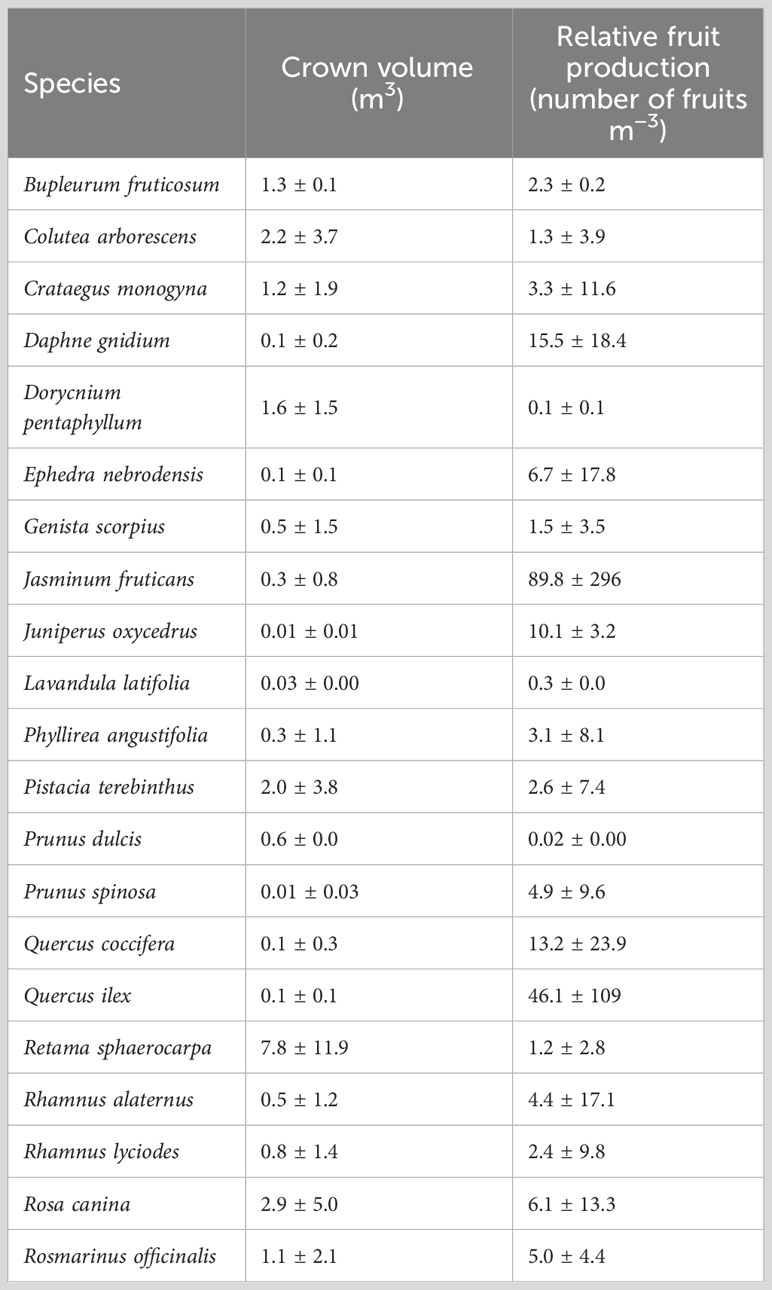
Table 2 Projected crown volume (in September 2019) per species and hedgerow and number of produced fruits normalized by plant size (estimated by a semi-quantitative number from 0 to 3; see Material and methods) measured in adult plants sampled in 10 × 2.5 m transects.
The volume of adult plants and the relative amount of fruit production in the hedgerows depended on species; it was highest for Jasminum fruticans and lowest for Prunus dulcis (Table 2). The plants with a larger volume produced more fruits than small plants (rho = 0.55, p < 0.01, n = 1,669). However, there were not significant relations between hedgerow age and plant volume (t = −0.03, p = 0.19, n = 1,699) or relative fruit production (t = −0.03, p = 0.20, n = 1,699). Relative fruit production (number of fruits/m3) differed among sites (Kruskal-Wallis, F3,662 = 5.45, p < 0.01) and averaged (± se) 0.57 ± 0.04 at “Vista Alegre”, 0.58 ± 0.04 at “El Peral”, 0.68 ± 0.05 at “Fuente del Albañal”, and 0.71 ± 0.04 at “Los Billares”.
We counted 66 and 316 individuals established from seeds and resprouts, respectively, of the planted species across hedgerows (Table 3). In addition, we counted 29 plants of species that were not planted in the hedgerows, namely 15 almond trees (P. dulcis), 10 grapevines (Vitis vinifera L.), two pistachio trees (Pistacia vera L.), a fig tree (Ficus carica), and an asparagus (Asparagus acutifolius L.). The average recruitment rate from seed of both planted and non-planted species was 14 individuals ha−1 year−1.
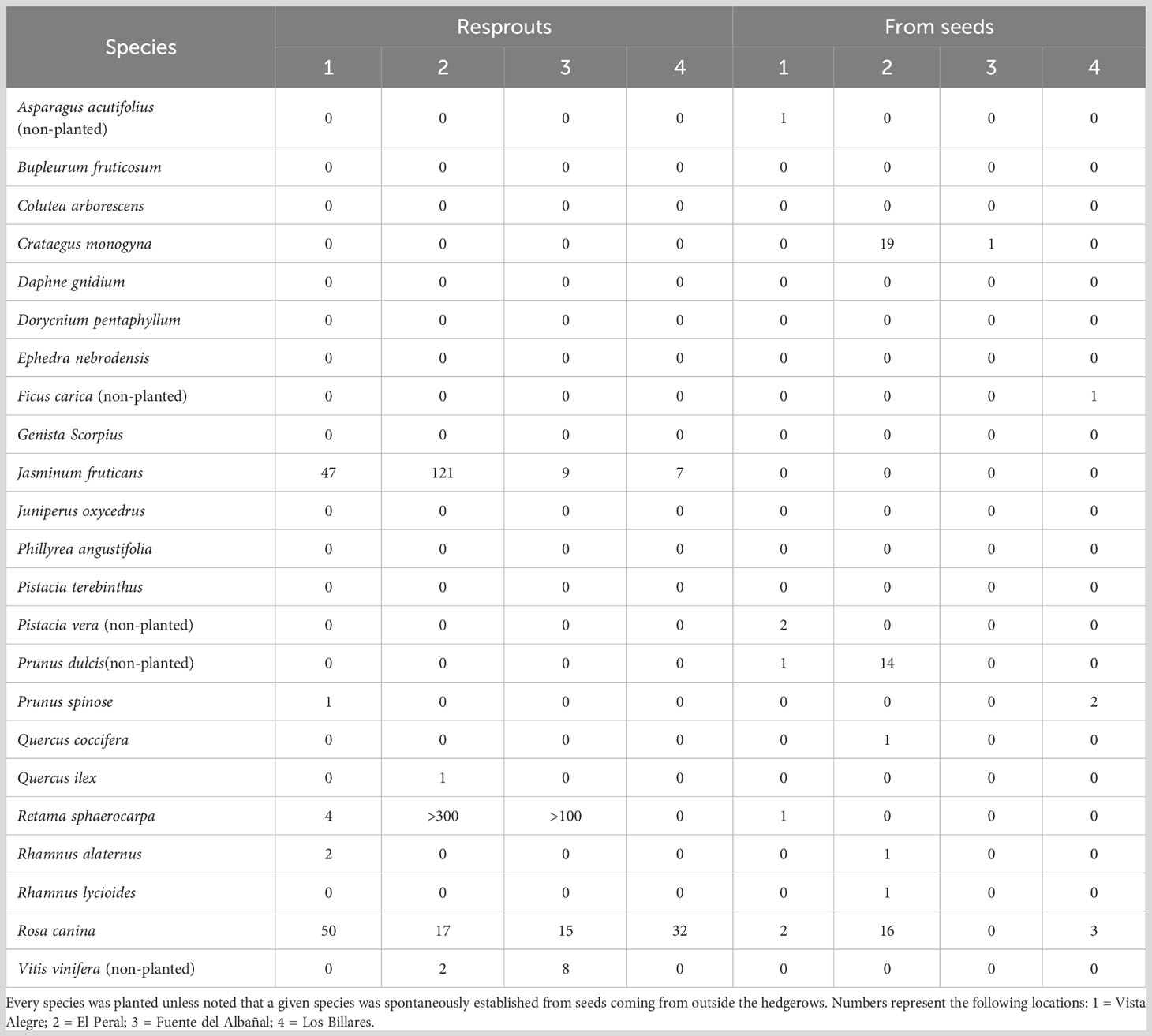
Table 3 Natural regeneration at the studied planted hedgerows, distinguishing between the number of juveniles from seed germination and resprouts.
We only identified one recruited individual of blackthorn (Rhamnus lycioides L.) in the abandoned agricultural fields adjacent to the hedgerows, which was located at 3 m from the hedgerow, and a colonization front of several hundreds of R. sphaerocarpa seedlings, which were apparently of the same age and whose mother plants were planted ten years before.
The mean distance (average ± se) between the naturally emerged seedlings and the nearest reproductive plant of the same species (i.e. potential mother plant) was: C. monogyna (0.63 ± 0.26 m), P. spinosa (0.80 ± 0.06 m), R. sphaerocarpa (2.04 ± 0.05 m), R. canina (3.10 ± 0.16 m), R. alaternus (3.80 m; only one individual), R. lycioides (7.00 m; only one individual), P. dulcis (25 ± 13 m), P. vera (40 ± 7 m), and Quercus coccifera L. (64 m; only one individual).
Survival of transplanted seedlings progressively decreased to 22% by the end of counting (approximately two years). Mortality was faster in “El Peral” than in “Los Billares” (LR χ2 = 33.71, p < 0.01; Figure S1A) and differed among species (LR χ2 = 22.94, p < 0.01; C. arborescens survived the least and C. monogyna survived the most; Figure S1B). Protection against herbivores had a positive effect on seedling survival (LR χ2 = 47.17, p < 0.01; Figure S1C) that differed among species (Species × Herbivory LR χ2 = 14.47, p < 0.01); R. alaternus and C. arborescens were the most and the least, respectively, benefited species by seedling shelters.
There were not significant main effects of irrigation (LR χ2 < 0.01, p = 0.99) or herb clipping (LR χ2 = 1.45, p = 0.23) on seedling survival. The interaction of these two treatments with species and protection against herbivores was significant: Species × Protection × Clipping (LR χ2 = 13.71, p < 0.01), and Species × Herbivory × Clipping × Irrigation (LR χ2 = 6.00, p = 0.05; Figure 2). Alleviating drought stress and competition had little to no effect on the unprotected plants. In contrast clipping increased survival for protected seedlings. R. alaternus was the species that benefited the most when protected against herbivores (higher survival increase; 28%), while C. arborescens was the least benefited (lowest survival increase; 18%). In general, irrigation only increased survival if herb competition was suppressed, and this effect was stronger for C. arborescens and R. alaternus. In summary, herbivory was the most important factor for seedling survival and that herb competition was more important than summer drought for survival.
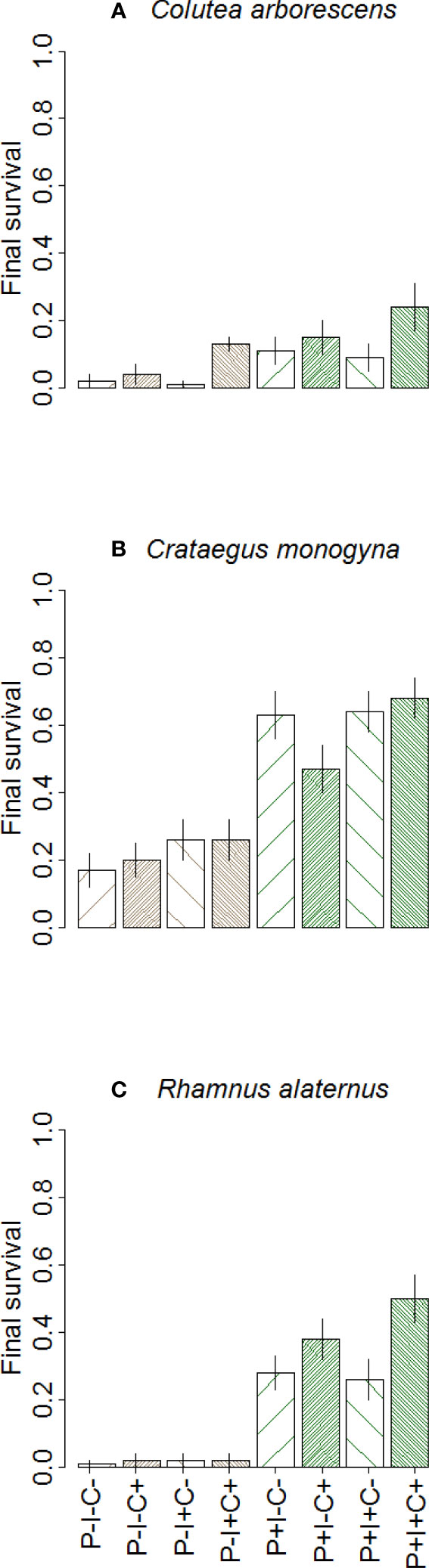
Figure 2 Effects of interactions among protection against herbivories (unprotected P−, brown bars; protected P+, green bars), irrigation (I−, without irrigation, line pattern rising right “/”, I+ with irrigation, line pattern rising left “”), and herb clipping (C− without clipping, low line density pattern; C+ with clipping, high line density pattern) on the survival of C. arborescens (A), C. monogyna (B), and R. alaternus (C) transplanted seedlings at the end of the experimental in September 2022. Error bars represent standard errors.
Other significant interaction terms were Site × Species × Clipping (LR χ2 = 6.93, p = 0.03), Site × Species × Irrigation (LR χ2 = 11.84, p < 0.01), and Site × Protection × Clipping × Irrigation (LR χ2 = 4.92, p = 0.03). The variation partition indicated that species identity and protection against herbivores explained each 5% of observed mortality, whereas both irrigation and herb clipping explained <1%.
Height growth of transplanted seedlings depended on species (F = 18.77, p < 0.01) and treatment (F = 20.47, p = 0.01; Figure 3). R. alaternus grew more than the other two species (t = 3.33, p < 0.01). Clipped (t = 2.38, p = 0.04) and protected (t = −2.04, p = 0.04) seedlings also grew more than non-clipped or unprotected seedlings. Irrigation did not affect seedling growth. According to the variation partition analysis, species and treatment explained 32% and 20% of seedling growth, respectively.
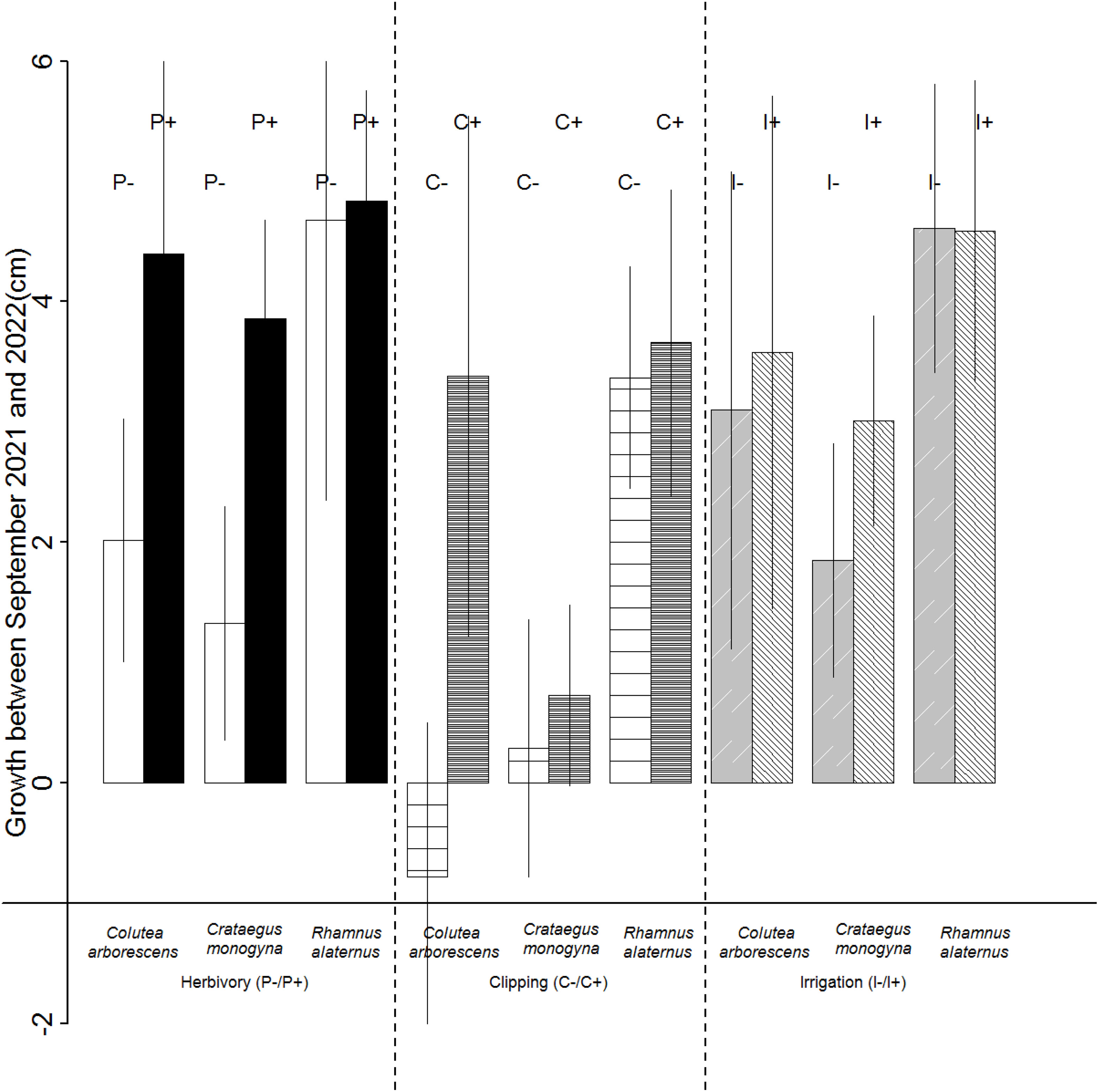
Figure 3 Difference in seedling height growth per species and treatment. P+ protection against herbivores; P− without protection against herbivores; C+ herb clipping, reduced competition; C− no clipping; I+ irrigation, reduced drought; I− no irrigation. Error bars represent standard errors.
Although the effects of interactions among treatments on growth could not be statistically tested owing to small sample size, some relationships can be guessed (Figure 3). Lack of protection and herb clipping reduced growth in all species but R. alaternus (interactions Protection × Species and Clipping × Species). Irrigation increased C. monogyna growth but had little or no effect for the two other species (interaction Irrigation × Species).
This study has (1) quantified the natural regeneration of species planted in 11- to 13-yr-old Mediterranean hedgerows and the colonization of species dispersed into the hedgerows and (2) experimentally assessed the limiting factors of woody seedlings establishment. Its novelty lies in the long-term estimation of natural recruitment and the assessment of the relative importance of three major limiting factors on the recruitment in Mediterranean hedgerows. Overall and in agreement with our predictions, summer drought, herbivory, and herb competition limited the early establishment of woody seedlings (P1), functionally different woody species exhibited contrasting responses to these limiting factors (P2), and limiting factors for plant establishment depended on the local context (P3).
Hedgerow plantings resulted on ca. one third of ground covered by woody vegetation, the amount of fruit production averaged up to ten fruits per m3 of hedgerow vegetation, and recruitment rate was 14 seedlings ha−1 yr−1. These values are consistent with the values reported in restoration trials in Mediterranean environments. For instance, Martínez-Baroja et al. (2022) found that 4.1 oaks (Q. ilex) ha−1 yr−1 were recruited from 30-yr-old woodland. Similarly, Vieco-Martínez et al. (2023) found 9.7 individuals ha−1 yr−1 that were spontaneously recruited under mixed 26-yr-old Pinus halepensis and Q. ilex plantations close to our study sites.
Nearly half of the newly established seed recruits in the studied hedgerows originated from non-planted species that successfully colonized them, indicating that hedgerows performed as effective applied nucleation elements (Corbin and Holl, 2012). However, we hardly found established individuals in the vicinity of the planted hedgerows, which we attribute to ploughing and several abiotic and biotic factors that limit plant establishment in exposed Mediterranean environments (Streit et al., 2002; García de León et al., 2014). Notably, all established seedlings were of vertebrate-dispersed species (e.g., C. monogyna, J. fruticans, P. dulcis, and Rosa canina), highlighting the importance of zoochory for hedgerow regeneration.
Our findings hint that natural regeneration assisted by hedgerows in Mediterranean fields is a slow process, despite the increasing propagule pressure. In addition to the limiting factors and processes we experimentally assessed here and discuss below, competition from established vegetation at hedgerows may hinder natural regeneration aligning with the Janzen-Connel’s hypothesis. Janzen (1970) suggested that maximum seedling establishment occurs at intermediate distances from mother plants; close to mother plants, adults outcompete seedlings, which eventually die, and few propagules are able to reach and establish at long distance sites. In our experiment, ploughed crops occupied the intermediate distances from mother plants.
Our transplanting experiment ascertained the factors limiting the early (two years) establishment of seedlings in the hedgerows. Around 80% of the transplanted seedlings died in the first two years with rabbit herbivory emerging as the main factor hindering natural regeneration. The high vulnerability of young seedlings to herbivores can be attributed to the limited capacity of seedlings to produce secondary defensive compounds and toughness (Barton and Hanley, 2013). Herbivory pressure has also been described as a key limitation to woody species recruitment in hedgerows (Endels et al., 2004; Suárez-Esteban et al., 2013). For instance, Suárez-Esteban et al. (2013) found that the low density of hackberry (Celtis australis) in Mediterranean hedgerows was due to a larger density of rabbits compared to other habitats. In agreement with our results, Endels et al. (2004) estimated that 80% of planted individuals in hedgerows were affected by herbivory.
The influence of abiotic factors, such as temperature and precipitation, on the regeneration of woody plants in hedgerows has also been highlighted in previous studies (McCarthy, 1994; Jacquemyn et al., 2010; Footitt et al., 2018). Notably, we did not find any significant effects of irrigation on survival and growth, suggesting that competition for water was severe even in the irrigated scenario. Hand irrigation may have been insufficient to compensate the fast soil drying (McCarthy, 1994). Rey Benayas et al. (2002) even found a marginal negative effect of irrigation on seedling survival, attributing it to increased competition by weeds in irrigated plots rather than the direct negative effects of water itself. This explanation aligns with our experiment, where we also found a negative effect of irrigation on protected C. monogyna seedlings (Figure 2B).
Contrary to our P1, herb clipping alone did not enhance the survival of woody seedlings in hedgerows. However, when combined with herbivory, herb clipping showed a positive effect (Figure 2). It is possible that the frequency of herb clipping was not sufficient to fully counteract the predicted effect of herb competition on survival. Previous studies support the idea that herb competition reduces woody seedling growth (Kollmann and Reiner, 1996; Rey Benayas et al., 2002; Rey Benayas et al., 2003; Rey Benayas et al., 2005; Rey Benayas et al., 2007). Kollmann and Reiner (1996) suggested that herbs compete for light with woody seedlings. Conversely, Rey Benayas et al. (2003) pointed out that the competition between woody species seedlings and herbs in Mediterranean abandoned cropland was mainly driven by water availability rather than light. The negative effects of herbs on Quercus coccifera seedlings resulted from competition for both water and light, while water competition was more important in the case of more mesic Quercus species (Rey Benayas et al., 2007). Some authors have also emphasized that herbs can limit the growth of woody seedlings through competition for nutrients (Tourte and Buchanan 2003; Cuesta et al., 2010; Jacquemyn et al., 2010).
We observed that seedlings of C. arborescens and R. alaternus were shorter when grown surrounded by a dense herb cover. This indicates that competition with herbs was relevant, although it did not directly contribute to the observed high seedling mortality (Figure 3). Our experimental design did not allow us to determine whether seedlings primarily competed for water (Rey Benayas et al., 2003), light (Kollmann and Reiner, 1996) or soil nutrients (Jacquemyn et al., 2010). Nevertheless, herb removal provided the surviving seedlings with more resources for their development. In our study, competition for water and light seems more plausible than competition for nutrients, because agricultural fields surrounding the hedgerows were fertilized, and the composition of planted hedgerows include legume shrubs, which serve as natural nitrogen fertilizers (Dendy et al., 2015).
Species responded differently to the treatments (P2). Species differences in survival could be attributed to their varying tolerance to drought and herbivory. Consistently, the deciduous legume C. arborescens had the lowest survival. Papachristou and Papanastasis (1994) found that C. arborescens and other legumes have high concentration of protein and low concentration of lignin, which make these species particularly attractive to herbivores. Surprisingly and contrary to our hypothesis, the evergreen species with scleromorphic leaves, R. alaternus, did not show the highest tolerance to drought and herbivory, while the deciduous C. monogyna was the least vulnerable species to the stress factors. Additionally, we cannot discard that differences in the quality of planted seedlings could have affected their field performance (Cortina et al., 2006; Mola et al., 2018).
Unlike survival, R. alaternus showed the highest growth rates, likely due to higher benefits from shade provided by adult plants in the hedgerows. Some individuals of this species reached a height growth of 30 cm in the first autumn after planting, particularly when located close to well-developed brooms (R. sphaerocarpa). Andivia et al. (2017) found that broom presence was a major determinant in the colonization of mid and late successional woody species like R. alaternus.
The studied species had contrasting values for most regeneration-related functional traits, with seedling height standing out (Shannon et al., 1996; Endels et al., 2004; Suárez-Esteban et al., 2013; Paal et al., 2020). Plant height is an easy measurable trait that directly relates to the maximum distance of propagule dispersal (Tamme et al., 2014). Our interpretation of contrasting responses of different species based on functional trait complementarity is consistent with that of Rey Benayas et al. (2005) for Quercus species planted in Mediterranean farmland, who found that shading increased growth only for the mesic Q. faginea, but not for the xerophytic Q. coccifera.
Fruit type is an important functional trait that can affect the colonization of hedgerows by certain plant species such as the almond or fig trees. The type of fruit can also explain why species differently respond to limiting factors (Figure 2). Plants with fleshy fruits, which are dispersed by vertebrates, can colonize farther compared to those with non-fleshy fruits (Suárez-Esteban et al., 2013). Nevertheless, in our study we have only detected one specimen of fig tree as a representative of fleshy fruit species established in the hedgerows without being planted. Conversely, non-fleshy fruit species such as the almond tree and the pistachio were able to establish to a larger extent by spontaneously colonizing from outside the hedgerows. Approximately half of planted species in the studied hedgerows are dispersed by vertebrates (e.g., C. monogyna is animal dispersed, while Retama sphaerocarpa is wind dispersed; Table 2). We attribute this effect to the presence of abundant individuals of vertebrate-dispersed plant species recruited in the hedgerows and to the fact that they can be effectively dispersed by magpie (Pica pica). Magpie is also abundant in the studied areas (Sánchez-Moreno et al., 2018; Martínez-Baroja et al., 2019; Martínez-Baroja et al., 2021). However, there cannot be an optimum combination of factors that maximize the regeneration of all species simultaneously because species vary in their response to herbivory, irrigation, and clipping (Figure 2). Instead, managers might need to provide a set of micro-environments and species to enhance the regeneration of hedgerows (Paal et al., 2020).
Finally, hedgerow regeneration differed among study sites (P3). The abundance of rabbits (Suárez-Esteban et al., 2013) and propagule pressure of species near the hedgerows can explain in part differences among sites. Dainese et al. (2015) found that the largest regeneration can be observed in hedgerows with intermediate levels of herbivore and propagule pressures. Climate and soil characteristics might have also influenced the different establishment of seedlings across sites.
We found that natural regeneration in planted Mediterranean hedgerows is slow despite the presence of planted sources of propagules. Species dispersed by vertebrates were favored. Herbivory was the main limiting factor for seedling survival, especially for the deciduous C. arborescens. Summer drought and herb competition primarily restricted growth and were relevant when interacted with other limiting factors. Our study suggests that costs of replacing dead plants in planted hedgerows can be minimized conducting censuses to identify newly recruited seedlings and implementing protection measures against herbivores such as rabbits. Reducing spontaneous herbaceous cover by mowing or herbicides also contribute to the establishment of woody plants in hedgerows. We recommend planting species-rich hedgerows that incorporate a variety of functional traits to develop these linear green infrastructure elements in agricultural Mediterranean landscapes.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
JR and PV-S conceived the study and designed the experiment. DG established and maintained the experiment, collected the data, performed statistical analyses, and wrote the first draft. All co-authors contributed to revisions. All authors contributed to the article and approved the submitted version
This study was funded by the Regional Government of Madrid [REMEDINAL, ref. TE-CM S2018/EMT-4 338] and the University of Alcalá [CM/JIN/2019-023]. DG thanks to the Regional Government of Madrid and The University of Alcalá for the talent research grant 2018-T2/BIO-10995, and project TED2021-130908BC44, funded by MCIN/AEI/10.13039/501100011033 and the European Union NextGenerationEU/ PRTR.
Baiba Ozolina and Jekaterina Bikova helped in field transplantation. Maite Artero Morales and Pedro García de León Hernández participated in survival censuses and biometrical measures. Guillermo González Fradejas, Maite Artero Morales, Gloria Martínez Sagarra, Mario Vasquez Loy, Andrea Collado Gómez, and Aruca Pedrero Redondo participated in the experimental irrigation and clipping.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1206000/full#supplementary-material
Andivia E., Villar-Salvador P., Tovar L., Rabasa S., Rey Benayas J. M. (2017). Multiscale assessment of woody species recruitment in Mediterranean shrublands: facilitation and beyond. J. Veg. Sci. 28, 639–648. doi: 10.1111/jvs.12520
Andivia E., Villar‐Salvador P., Oliet J. A., Puértolas J., Dumroese R. K., Ivetić V., et al. (2021). Climate and species stress resistance modulate the higher survival of large seedlings in forest restoration worldwide. Ecol. Appl. e02394. doi: 10.1002/eap.2394
Barton K. E., Hanley M. E. (2013). Seedling-herbivore interactions: Insights into plant defence and regeneration patterns. Ann. Bot. 112, 643–650. doi: 10.1093/aob/mct139
Corbin J. D., Holl K. D. (2012). Applied nucleation as a forest restoration strategy. For. Ecol. Manage. 265, 37–46. doi: 10.1016/j.foreco.2011.10.013
Cortina J., Peñuelas J. L., Puértolas J., Savé R., Vilagrosa A. (2006). Calidad de planta forestal para la restauración en ambientes mediterráneos Ministerio de Medio Ambiente ISBN-10: 84-8014-670-2. Available at: https://www.miteco.gob.es/content/dam/miteco/es/biodiversidad/publicaciones/calidad_planta_forestal_tcm30-100334.pdf.
Cuesta B., Villar-Salvador P., Puértolas J., Rey Benayas J. M., Michalet R. (2010). Facilitation of Quercus ilex in Mediterranean shrubland is explained by both direct and indirect interactions mediated by herbs. J. Ecol. 98 (3), 687–696. doi: 10.1111/j.1365-2745.2010.01655.x
Dainese M., Luna D. I., Sitzia T., Marini L. (2015). Testing scale-dependent effects of seminatural habitats on farmland biodiversity. Ecol. Appl. 25, 1681–1690. doi: 10.1890/14-1321.1
Deckers B., Verheyen K., Hermy M., Muys B. (2005). Effects of landscape structure on the invasive spread of black cherry Prunus serotina in an agricultural landscape in Flanders, Belgium. Ecography 28, 99–109. doi: 10.1111/j.0906-7590.2005.04054.x
Dendy J., Cordell S., Giardina C. P., Hwang B., Polloi E., Rengulbai K. (2015). The role of remnant forest patches for habitat restoration in degraded areas of Palau. Restor. Ecol. 23, 872–881. doi: 10.1111/rec.12268
Endels P., Adriaens D., Verheyen K., Hermy M. (2004). Population structure and adult plant performance of forest herbs in three contrasting habitats. Ecography 27, 225–241. doi: 10.1111/j.0906-7590.2004.03731.x
European Commision. (2020). EU Biodiversity Strategy for 2030 Bringing nature back into our lives. Communication from the commission to the European parliament, the council, the European economic and social committee and the committee of the regions COM(2020) 380 final Brussels, 20.5.2020. Available at: https://environment.ec.europa.eu/strategy/biodiversity-strategy-2030_en.
Footitt S., Huang Z., Ölcer-Footitt H., Clay H., Finch-Savage W. E. (2018). The impact of global warming on germination and seedling emergence in Alliaria petiolata, a woodland species with dormancy loss dependent on low temperature. Plant Biol. 20, 682–690. doi: 10.1111/plb.12720
Fox J., Weisberg S. (2019). An R companion to applied regression. 3rd ed. (Thousand Oaks, CA, USA: Sage).
Fyllas N. M., Michelaki C., Galanidis A., Evangelou E., Zaragoza-Castells J., Dimitrakopoulos P. G., et al. (2020). Functional trait variation among and within species and plant functional types in mountainous Mediterranean forests front. Plant Sci. 11. doi: 10.3389/fpls.2020.00212
García de León D., Freckleton R. P., Lima M., Navarrete L., Castellanos E., González-Andújar J. L. (2014). Identifying the effect of density dependence, agricultural practices and climate variables on the long-term dynamics of weed populations. Weed. Res. 54, 556–564. doi: 10.1111/wre.12113
García de León D., Rey Benayas J., Andivia E. (2021). Contributions of hedgerows to people - a global meta-analysis. Front. Cons Sc 2, 789612.
González-Varo J. P., Fedriani J. M., López-Bao J. V., Guitián J., Suárez-Esteban A. (2015). Frugivoría y dispersión de semillas por mamíferos carnívoros: rasgos funcionales Ecosistemas. 24 (3), 43-50. doi: 10.7818/ECOS.2015.24-3.07
Granda E., Escudero A., Valladares F. (2014). More than just drought: complexity of recruitment patterns in Mediterranean forests. Oecologia 176 (4), 997–1007. doi: 10.1007/s00442-014-3064-x
Hegland S. J., Rydgren K., Lilleeng M. S., Moe S. R., Gillespie M. A. K. (2021). Junipers enable heavily browsed rowan saplings to escape ungulates in boreal forest. For. Ecol. Manage. 500, 119651. doi: 10.1016/j.foreco.2021.119651
IPBES (2019). Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Eds. Brondizio E. S., Settele J., Díaz S., Ngo H. T. (Bonn, Germany: IPBES secretariat), 1148. doi: 10.5281/zenodo.3831673
Jacquemyn H., Endels P., Honnay O., Wiegand T. (2010). Evaluating management interventions in small populations of a perennial herb Primula vulgaris using spatio-temporal analyses of point patterns. J. Appl. Ecol. 47, 431–440. doi: 10.1111/j.1365-2664.2010.01778.x
Janzen D. H. (1970). Herbivores and the number of tree species in tropical forests. Am. Nat. 104, 501–528. doi: 10.1086/282687
Kollmann J., Reiner S. A. (1996). Light demands of shrub seedlings and their establishment within scrublands. Flora 191, 191–200. doi: 10.1016/S0367-2530(17)30713-2
Lajos K., Császár O., Sárospataki M., Samu F., Tóth F. (2020). Linear woody landscape elements may help to mitigate leaf surface loss caused by the cereal leaf beetle. Landsc. Ecol. 35, 2225–2238. doi: 10.1007/s10980-020-01097-3
López-Iglesias B., Villar R., Poorter L. (2014). Functional traits predict drought performance and distribution of mediterranean woody species. Acta Oecol. 56, 10–18. doi: 10.1016/j.actao.2014.01.003
Marshall E. J. P., Grant A. J., Fairbairn S. (1999). “Spatial patterns of gastropod occurrence and herbivory in a field-scale habitat mosaic in winter,” in Heterogeneity in Landscape Ecology: pattern and scale. Eds. Maudsleyand M. J., Marshall E. J. P. (London, UK: IALE (UK), 239–244.
Martínez-Baroja L., Pérez-Camacho L., Villar-Salvador P., Rebollo S., Quiles P., Gómez-Sánchez D., et al. (2019). Massive and effective acorn dispersal into agroforestry systems by an overlooked vector, the Eurasian magpie (Pica pica). Ecosphere (12), e02989. doi: 10.1002/ecs2.2989.
Martínez-Baroja L., Pérez-Camacho L., Villar-Salvador P., Rebollo S., Leverkus A. B., Pesendorfer M. B., et al. (2021). Caching territoriality and site preferences by a scatter-hoarder drive the spatial pattern of seed dispersal and affect seedling emergence. J. Ecol. 109 (6), 2342–2353. doi: 10.1111/1365-2745.13642.
Martínez-Baroja L., Rey-Benayas J. M., Pérez-Camacho L., Villar-Salvador P. (2022). Drivers of oak establishment in Mediterranean old fields from 25-year-old woodland islets planted to assist natural regeneration. Eur. J. For. Res. 141, 17–30. doi: 10.1007/s10342-021-01423-7
Martínez-Vilalta J., Mencuccini M., Vayreda J., Retana J. (2010). Interspecific variation in functional traits, not climatic differences among species ranges, determines demographic rates across 44 temperate and Mediterranean tree species. J. Ecol. 98 (6), 1462–1475. doi: 10.1111/j.1365-2745.2010.01718.x
McCarthy B. C. (1994). Experimental studies of hickory recruitment in a wooded hedgerow and forest. Bull. Tor. Bot. Club 121, 240–250.
Mola I., Sopeña A., de Torre R. (2018). Guía práctica de restauración ecológica Vol. 77 (Madrid, Spain: Fundación Biodiversidad. Ministerio para la Transición Ecológica).
Oksanen J., Simpson G., Blanchet F., Kindt R., Legendre P., Minchin P., et al. (2022). Vegan: community ecology package. Package. R package version 2.6-4. Available at: https://CRAN.R-project.org/package=vegan.
Paal T., Zobel K., Liira J. (2020). Standardized response signatures of functional traits pinpoint limiting ecological filters during the migration of forest plant species into wooded corridors. Ecol. Indic. 108, 105688. doi: 10.1016/j.ecolind.2019.105688
Papachristou T. G., Papanastasis V. P. (1994). Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo-pastoral systems for goats. Agr. Sys. 27, 269–282. doi: 10.1007/BF00705061
Pausas J. G., Pratt R. B., Keeley J. E., Jacobsen A. L., Ramirez A. R., Vilagrosa A., et al. (2016). Towards understanding resprouting at the global scale. New Phytologist 209, 945–954. doi: 10.1111/nph.13644
Pörtner H. O., Scholes R. J., Agard J., Archer E., Arneth A., Bai X., et al. (2021). IPBES-IPCC co-sponsored workshop report on biodiversity and climate change (New York, USA: IPBES and IPCC). doi: 10.5281/zenodo.4782538
R Core Team. (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/.
Rey Benayas J. M., Bullock J. M. (2012). Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 15, 883–899. doi: 10.1007/s10021-012-9552-0
Rey Benayas J. M., Bullock J. M. (2015). “Vegetation restoration and other actions to enhance wildlife in european agricultural landscapes,” in Rewilding european landscapes. Eds. Pereira H. M., Navarro L. M. (Cham: Springer), 127–142. doi: 10.1007/978-3-319-12039-3_7
Rey Benayas J. M., Espigares T., Castro-Díez P. (2003). Simulated effects of herb competition on planted Quercus faginea seedlings. Appl. Veg. Sc. 6 (2), 213–222. doi: 10.1111/j.1654-109X.2003.tb00582.x
Rey Benayas J. M., Fernández A., Aubenau A. (2007). Clipping herbaceous vegetation improves early performance of planted seedlings of the Mediterranean shrub Quercus coccifera. Web. Ecol. 7, 120–131. doi: 10.5194/we-7-120-2007
Rey Benayas J. M., López-Pintor A., García C., de la Cámara N., Strasser R., Sal A. G. (2002). Early establishment of planted Retama sphaerocarpa seedlings under different levels of light, water and weed competition. Plant Ecol. 159, 201–209. doi: 10.1023/A:1015562623751
Rey Benayas J., Martínez de Baroja L., García de León D., Crespo Cepas G., Pajares Guerra M., Pérez Camacho L., et al. (2021). “Caso práctico XVI Plantación de islotes forestales y setos como infraestructura verde para ayudar la regeneración natural en paisajes agrícolas mediterráneos,” in Bases técnicas y ecológicas de la repoblación forestal. Ministerio para la Transición Ecológica y el Reto Demográfico. Eds. Pemán J., Navarro R. M., Serrada R.(Madrid, Spain: Ministerio para la Transición Ecológica y el Reto Demográfico), 523–532.
Rey Benayas J. M., Navarro J., Espigares T., Nicolau J. M., Zavala M. A. (2005). Effects of artificial shading and weed mowing in reforestation of Mediterranean abandoned cropland with contrasting Quercus species. For. Ecol. Manage. 212, 302–314. doi: 10.1016/j.foreco.2005.03.032
Sánchez-Moreno S., Cano M., López-Pérez A., Rey Benayas J. M. (2018). Microfaunal soil food webs in Mediterranean semi-arid agroecosystems. Does organic management improve soil health? Applied Soil Ecology 125, 138-147.
Shannon D., Isaac L., Brockmann F . (1996). Assessment of hedgerow species for seed size, stand establishment and seedling height Agroforestry Systems. 35 (1), 95-110.
Soliveres S., Maestre F. T., Bowker M. A., Torices R., Quero J. L., García-Gómez M., et al. (2014). Functional traits determine plant co-occurrence more than environment or evolutionary relatedness in global drylands. Persp. Plant Ecol. Evol. Syst. 16 (4), 164–173. doi: 10.1016/j.ppees.2014.05.001
Streit B., Rieger S.B., Stamp P., Richner W. (2002). The effect of tillage intensity and time of herbicide application on weed communities and populations in maize in central Europe Agriculture, Ecosystem and Environment 92(2-3), 211-224T.
Suárez-Esteban A., Delibes M., Fedriani J. M. (2013). Unpaved road verges as hotspots of fleshy-fruited shrub recruitment and establishment. Biol. Conserv. 167, 50–56. doi: 10.1016/j.biocon.2013.07.022
Tamme R., Götzenberger L., Zobel M., Bullock J. M., Hooftman D. A. P., Kaasik A., et al. (2014). Predicting species’ maximum dispersal distances from simple plant traits. Ecology 95, 505–513. doi: 10.1890/13-1000.1
Therneau T. M., Grambsch P. M. (2000). Modeling Survival Data: Extending the Cox Model. Springer, Berlin. doi: 10.1007/978-1-4757-3294-8.
Therneau T. M., Grambsch P. (2000). Modeling survival data: extending the cox model (New York: Springer).
Tourte, Buchanan. (2003). Estimated costs and potential for a perennial hedgerow planting UC Cooperative Extension. Available at: https://caff.org/wp-content/uploads/2010/07/HedgerowPlanting.pdf.
Vieco-Martínez A., Rey-Benayas J. M., Oliet J. A., Villar-Salvador P., Martínez-Baroja L. (2023). Efectos de la forestación de tierras agrícolas mediterráneas y de su manejo en el establecimiento de árboles y arbustos. Ecosistemas 32, 2460.
Keywords: germination, herb competition, herbivory, resprouts, summer drought
Citation: García de León D, Rey Benayas JM and Villar-Salvador P (2023) Assessing the limiting factors of natural regeneration in Mediterranean planted hedgerows. Front. Ecol. Evol. 11:1206000. doi: 10.3389/fevo.2023.1206000
Received: 14 April 2023; Accepted: 11 August 2023;
Published: 28 August 2023.
Edited by:
Meredith Root-Bernstein, CNRS UMR 7204, FranceReviewed by:
Mariano Ordano, Fundación Miguel Lillo, ArgentinaCopyright © 2023 García de León, Rey Benayas and Villar-Salvador. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David García de León, ZGF2aWRnbGhAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.