 Ziqing Zhao
Ziqing Zhao Liping He3,4,5
Liping He3,4,5 Yuguo Liu
Yuguo Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 09 June 2023
Sec. Population, Community, and Ecosystem Dynamics
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1199874
This article is part of the Research Topic Recent Advances in Understanding the Relationship between Biodiversity and Ecosystem Functioning in Forest Ecosystem View all 5 articles
The formation and continued maintenance of beta diversity are essential factors contributing to the effective conservation of plant community biodiversity and sustainable development of ecosystem functions. While it is widely acknowledged that two different patterns, including turnover and nestedness, strongly influence plant community beta diversity, their relative importance in the unique dry and hot valleys of Southwest China is yet to be fully understood. For this study, we established a 100 km belt transect in the Nujiang dry and hot valley of Southwest China and conducted community investigations. The calculations of species and phylogenetic beta diversity composition patterns allowed us to explore the relative importance of environmental and dispersal limitations in terms of their contributions to beta diversity. The findings indicated that in the Nujiang dry and hot valley of Northwest Yunnan, species and phylogenetic beta diversity in the plant communities were mainly composed of the turnover component. The values of phylogenetic beta diversity and its turnover components were lower than those of species beta diversity. Both environmental and dispersal limitations played a role in partitioning beta diversity in the region, with environmental limitations being more dominant than dispersal limitations. The analysis of specific environmental factors revealed that climate factors, such as mean annual precipitation and Hargreaves reference evaporation, and soil factors, such as pH, were the primary drivers of species and phylogenetic beta diversity in the region. In addition, we observed that various limiting environmental factors related to the growth and development of different life forms existed within the dry and hot valley communities. The study highlighted the potential benefits of establishing conservation reserves featuring multiple dominant shrub species in the Nujiang dry and hot valley of Southwest China for the protection of the vegetation. The findings demonstrated that environmental factors, particularly hydrothermal conditions, played a significant role in constraining the maintenance of beta diversity in plant communities in the dry and hot valley region at a regional scale, given the geological history and environmental changes that have occurred over time.
The formation and maintenance of beta diversity have been key areas of focus within community ecology research, as they are crucial for the preservation of biodiversity and the stability of ecosystem functions (Mariano et al., 2020; He et al., 2022). Generalized beta diversity is used to characterize species composition differences in biological communities across various sites or environmental gradients within a region (Whittaker, 1960). The loss of beta diversity can lead to biological homogeneity and result in a decrease in ecosystem functions (Socolar et al., 2016). Plant community composition is influenced not only by biotic interactions occurring within the community structure and environmental factors, (such as climate, soil, and topography) but also by differences in species richness between locations, which in turn contribute to changes in species composition(Tilman, 1982). In recent years, the focus of research has shifted to studying plant community partitioning beta diversity as a means of investigating the effects on species composition differences (Asefa et al., 2019; Pereira et al., 2021; Li et al., 2023). To better understand the potential patterns underlying community differences, traditional beta diversity can be decomposed into two distinct components: turnover and nestedness. The two components of traditional beta diversity, reflect distinct factors underlying community differences. Turnover reflects the replacement of species between different locations, which can be attributed to competition, environmental factors, and geographical barriers. On the other hand, nestedness reflects the gain and loss of species, wherein sites with low species richness represent a subset of sites with high species richness, and this process is primarily affected by extinction and colonization (Baselga, 2010; Baselga, 2012). Meanwhile, incorporating phylogenetic beta diversity components can link evolutionary history and control for potential confounding effects of phylogenetic diversity differences on diversity assessments (Leprieur et al., 2012; Qian et al., 2020; Li et al., 2021). Current research has focused on integrating the two partitions and limiting effects to provide a quantitative explanation for beta diversity (Soininen et al., 2007; Myers et al., 2013).
Studies of Chinese angiosperms have revealed that environmental limitations based on climate tolerance are a major driver of beta diversity patterns (Qian et al., 2020). In the desert plant community of Northwest China, beta diversity was primarily governed by turnover and nestedness partitions, as well as elusive factors, with environmental factors playing a more prominent role than dispersal limitations (Yang et al., 2021). Research conducted in natural reserves located in Northeast China found that non-biological factors, such as the geographic topography of ecological limiting, were the primary driving force behind overall plant beta diversity (She et al., 2022). Yunnan, located in the Southwest region of China, is considered the most biologically diverse area in the country. The unique combinations of climatic and topographical features found in the dry and region have allowed for the growth of “semi-Savanna vegetation” (Jin, 1999; Jin and Ou, 2000). At present, studies on plant communities in hot and dry valleys of Yunnan primarily focus on classification and community characteristics (Zhang et al., 2010), vegetation flora and origin (Ma et al., 2020), spatial patterns of populations (Long et al., 2022; He et al., 2023), and diversity characteristics of established communities via internal ecological processes and community environmental factors (Zhu et al., 2020, Liu et al., 2021; Du et al., 2022). The perennial debate surrounding the role of ecological interactions, environmental limitations, and dispersal limitations in elucidating community partition beta diversity mechanisms within the dry and hot valley community continues to be the prominent focal points in ecological and evolutionary studies (Meng et al., 2017; Qian et al., 2020; Shi et al., 2021). Moreover, the mechanisms underlying beta diversity maintenance may vary depending on the geographic region and specific research subjects under investigation (Li et al., 2016; Zhang et al., 2020). In large-scale spatial analyses of the Jinsha River valley region located in Northwest Yunnan, the plant community structure was primarily influenced by geographic isolation (Liu et al., 2016). By incorporating phylogenetic biology, we gain insights into the evolutionary drivers behind species formation and adaptation, as well as the dispersal and colonization processes that occur. The findings suggest that the majority of communities are composed of both recent and historical elements. Throughout the historical process, the formation and adaptation of beta diversity have played a pivotal role in community assembly (Webb, 2000; Webb et al., 2002). To further examine the impact of historical factors on the construction of plant communities within the dry and hot valley region, studies have integrated phylogenetic information (Meng et al., 2017). In the dry and hot valley communities of Yuanjiang in Southeast Yunnan, geographical distances and environmental differences were found to significantly impact both species and phylogenetic beta diversity, with geographical limitations potentially exerting a greater effect (Li et al., 2016). It has been observed that the Northwest and Southeast dry and hot valley regions of Yunnan, located at opposite ends of the Yunnan plateau, share some common geological histories (Zhu and Yan, 2003; Meng et al., 2013). Incorporating phylogenetic principles, the dry and hot valley plant communities of Nujiang, located in Northwest Yunnan, present an ideal research site for exploring the maintenance and driving factors of beta diversity. Characterized by considerable spatial distribution and distinct environmental gradients, this region offers a unique opportunity to enrich the theoretical foundations underlying partitioning beta diversity mechanisms in the dry and hot valley of Southwest China (Yang et al., 2016). Additionally, such research can provide important insights into the current state of plant community assembly in the Nujiang region, especially in light of the unprecedented geological and environmental changes impacting this area.
Investigating and describing the patterns of species and phylogenetic composition across various spatial and environmental gradients is crucial for comprehensively understanding the underlying mechanisms of partitioning beta diversity and biodiversity maintenance in ecological systems. To analyze species and phylogenetic beta diversity patterns, a total of 22 plots were explored along a 100 km belt transect in the Nujiang dry and hot valley region of Southwest China. To quantify environmental limitations, we utilized the Euclidean distance between environmental factors (Qian et al., 2020), and to evaluate the influence of geographical limitation among the plots, we incorporated spatial distance analyses (Myers et al., 2013). The study aims to examine three primary questions: (1) Is the formation of species and phylogenetic beta diversity in the Nujiang dry and hot valley region of Southwest China driven by turnover or nestedness? (2) How do the environmental factors including climate and soil factors affect beta diversity? (3) What is the relative importance of environmental filtering and dispersal limitation on observed beta diversity patterns? Given the vulnerability of vegetation in the dry and hot valley region, we aim to identify the key factors driving environmental heterogeneity while examining their relative influence on partitioning beta diversity. Additionally, we propose measures for the protection of vegetation in this region to enhance the stability of ecosystem function.
The study area for this investigation is located in the Nujiang River region, stretching from 24°57′–25°38′N and 98°49′–98°52′E in northwest Yunnan, Southwest China, and situated along the middle reaches of the river basin. This region features a typical dry and hot valley landform, with altitudes ranging from 738 to 1,189 m. The mean annual temperature falls between 18.7 and 21.48°C, while the mean annual precipitation ranges from approximately 780 to 1051.08 mm, concentrated primarily between May and September. Additionally, the zonal soil type in this study area is dry red soil.
To conduct this study, a 100 km belt transect was established in the dry and hot valley region of the study area. Sample plots were established at 4 km intervals, and vegetation community surveys to conduct (Figure 1). The use of GPS technology facilitated the precise identification and location of each site and elevation (ELV). Specifically, each 10 × 10 m plot was further divided into four 5 × 5 m quadrats for accurate measurement and assessment purposes. As part of this study, thorough investigations of trees and shrubs were conducted within the 10 × 10 m plots, while the 2 × 2 m quadrats arranged at each of the four vertex corners within the same plot were used to document the herbaceous species. The situation and dominant species composition of each plot were shown in Supplementary Table 1.
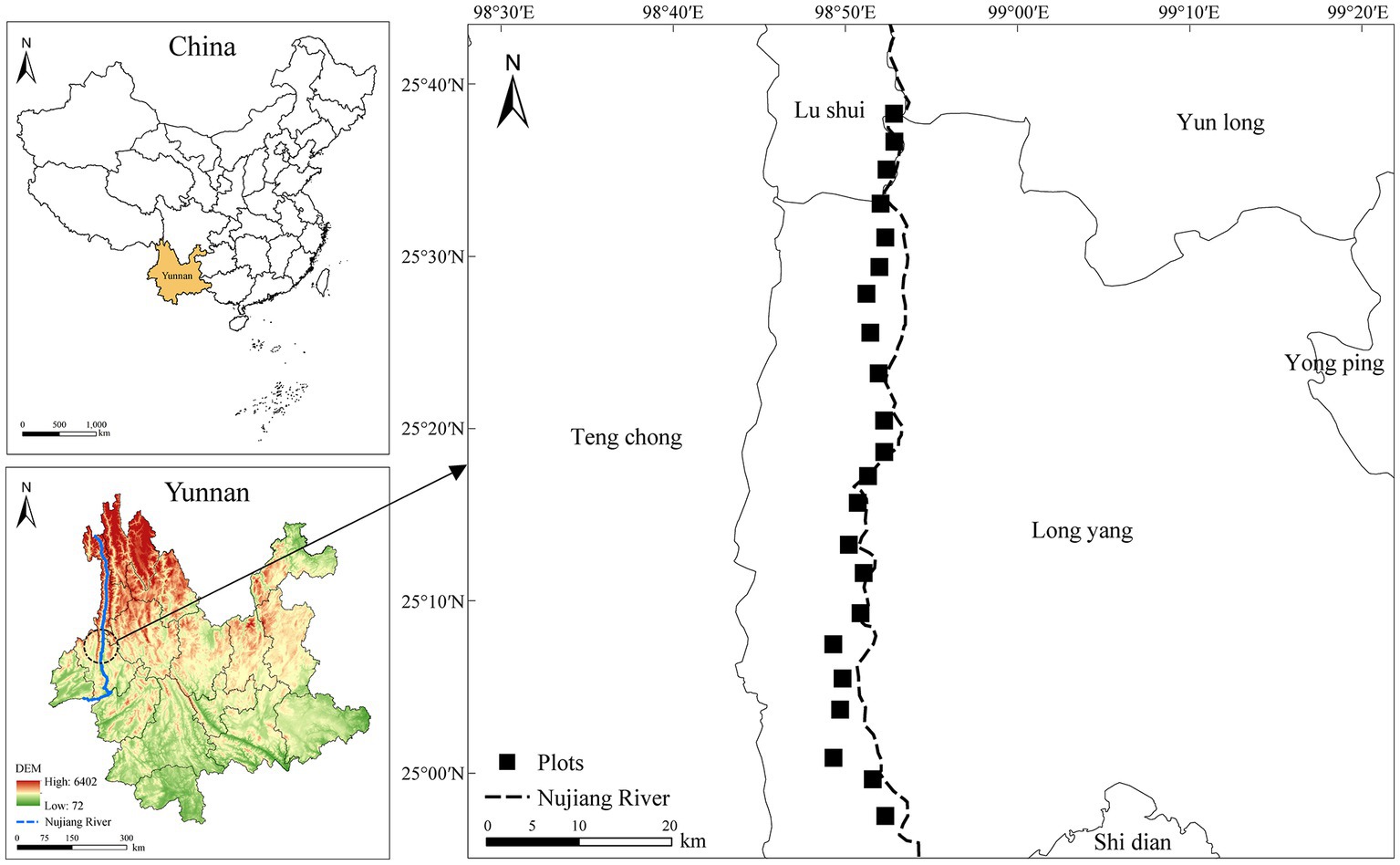
Figure 1. Location of quadrats setting in the Nujiang River.
The climate data utilized in this study, including the mean annual precipitation (MAP), mean annual temperature (MAT), Hargreaves reference evapotranspiration (EREF), and annual heat-moisture index (AHM), were obtained from the ClimateAP website (https://climateap.net/). Additionally, the total nitrogen (TN) was quantified using the selenium–cupric sulfate (CuSO4)–potassium sulfate (K2SO4)–heating digestion method. To evaluate available nitrogen (AN) content, the Kjeldahl nitrogen determination method was utilized, while the total phosphorus (TP) was determined via the alkali fusion-Mo-Sb colorimetric method. Soil organic matter (OM) was assessed using the potassium dichromate (K2Cr2O7) oxidation-heating method, and soil fast-acting potassium (AK) content was determined by ammonium acetate leaching and subsequent flame photometry. To quantify total potassium (TK), flame photometer analyses were performed, with soil dry-to-wet ratio (DW) measured utilizing the aluminum box method. Finally, the determination of soil pH was carried out using an electrode method.
To assess total beta diversity and its respective components of turnover and nestedness, the Sorensen dissimilarity coefficient was applied. Species beta diversity was computed based on the occurrences of species at each site, while phylogenetic beta diversity was determined using the phylogenetic tree generated by the “V.PhyloMaker 2” package (Jin and Qian, 2022), which abolished phylogenetic nest structure. Beta diversity calculations were performed using R version 4.1.0 software (R Core Team, 2021). Specifically, the “betapart” package was utilized, with functions such as “beta.pair” and “phylo.beta.pair” utilized to evaluate species and phylogenetic beta diversity, and “beta.multi” and “phylo.beta.multi” functions were implemented to calculate the patterns of species and phylogenetic beta diversity (Baselga, 2012).
Before conducting any analyses, we implemented a screening process to identify and eliminate any multicollinearity among the environmental factors. We calculated the Pearson correlations and variance inflation factors (VIF) for each environmental factor (Supplementary Table 2). Subsequently, we removed the predictor with the largest VIF to evaluate collinearity and recomputed the VIFs for the remaining predictors until all values were brought below the threshold of 5 (Dormann et al., 2013). We excluded AHM, MAT, ELV, and OM, and focused only on the remaining environmental factors. To investigate the effects of these environmental factors on the two dimensions of beta diversity, mantel correlation analyses were conducted to assess the relationships between nine environmental factors (pH, TN, TK, TP, AN, AK, DW, MAP, EREF) and both species and phylogenetic beta diversity. To calculate the environmental distance matrix, we standardized the data for soil factors (pH, OM, TN, TK, TP, AN, AK, DW), climate factors (MAP, MAT, EREF, AHM), and topographic factors (ELV) before eliminating any dimensionality effects by using the function “scale” in R.
To construct the geographical distance matrix, we relied on the latitude and longitude coordinates of the plots within the dry and hot valley (Zhao et al., 2017). The “rdacca.hp” package in R was utilized with the “rdacca.hp” function to perform distance-based redundancy analysis (db-RDA) and hierarchical partitioning (HP) using Sorensen and PhyloSor dissimilarity matrices in addition to both the environmental and geographical distance matrices (Lai et al., 2022). These approaches facilitated the quantification of the independent contributions of individual explanatory variables and multiple variables. The analyses aimed to assess the relative effects of environmental and geographical limitations on beta diversity. For those environmental factors that had negative explanatory contributions, our findings indicated limited variation within the environmental gradient and minor effects on beta diversity. As such, these factors were re-evaluated and removed, while the remaining factors were recalculated to determine their respective explanatory contributions. All statistical analyses were carried out in R version 4.1.0 (R Core Team, 2021) utilizing several key packages, including “picante” (Kembel et al., 2010), “vegan” (Oksanen et al., 2019), “V.PhyloMaker 2” (Jin and Qian, 2022), and “rdacca.hp” (Lai et al., 2022).
A total of 125 species of vascular plants were identified, representing 51 families and 104 genera. This included 41 species of trees (19 families, 33 genera), 50 species of shrubs (25 families, 41 genera), and 34 species of herbaceous plants (23 families, 37 genera). The analysis revealed that the species beta diversity within dry and hot valley plant communities was estimated to be 0.89, with the turnover and nestedness components accounting for 0.87 (97.04%) and 0.07 (2.96%), respectively (Figure 2). Furthermore, phylogenetic beta diversity was estimated to be 0.83, with turnover and nestedness components accounting for 0.79 (94.98%) and 0.04 (5.02%), respectively. Turnover was the predominant component of both species and phylogenetic beta diversity across all studied life forms. In pairwise beta diversity of plots in community and different life forms, species beta diversity was higher than phylogenetic beta diversity (Figure 3). Additionally, the turnover component consistently exceeded the nestedness component across life forms, with trees displaying the highest turnover component followed by shrubs and herbs (Figure 3, Table 1).
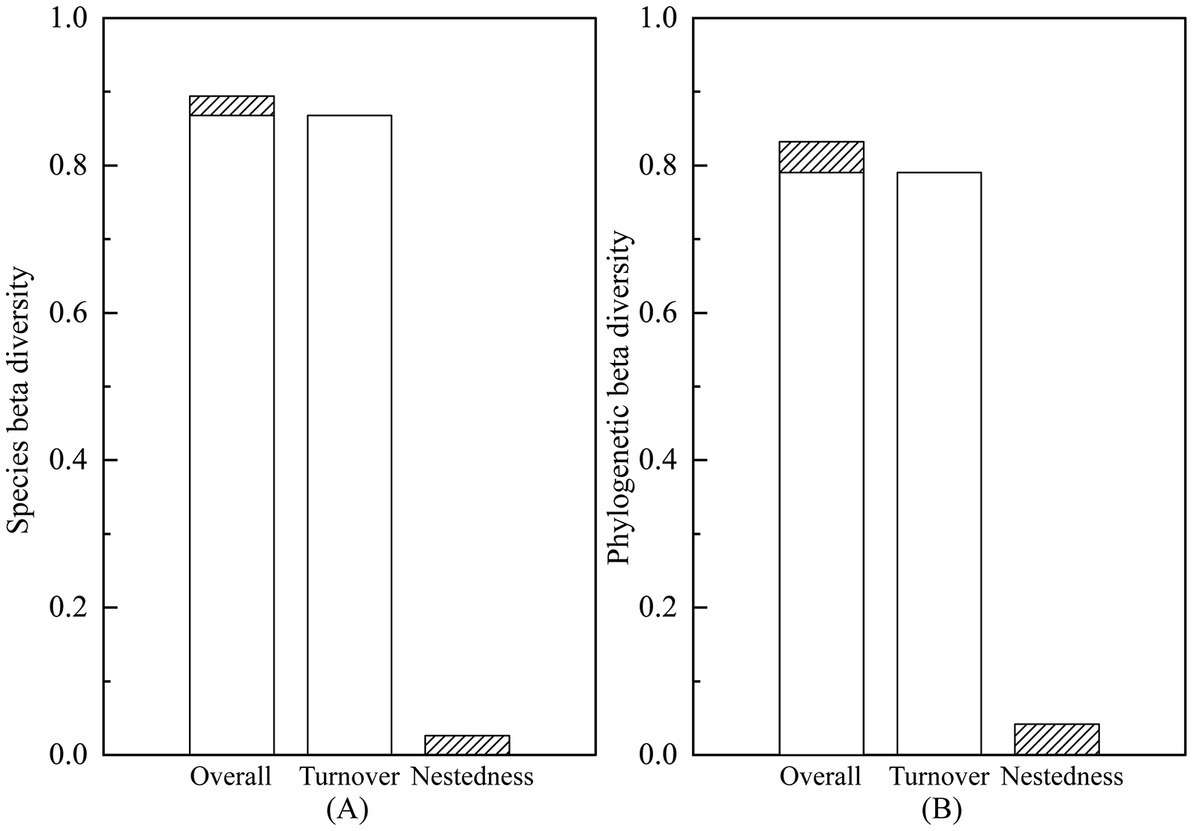
Figure 2. Beta diversity and its compositions in dry and hot valley plant communities: (A) species beta diversity; (B) phylogenetic beta diversity.
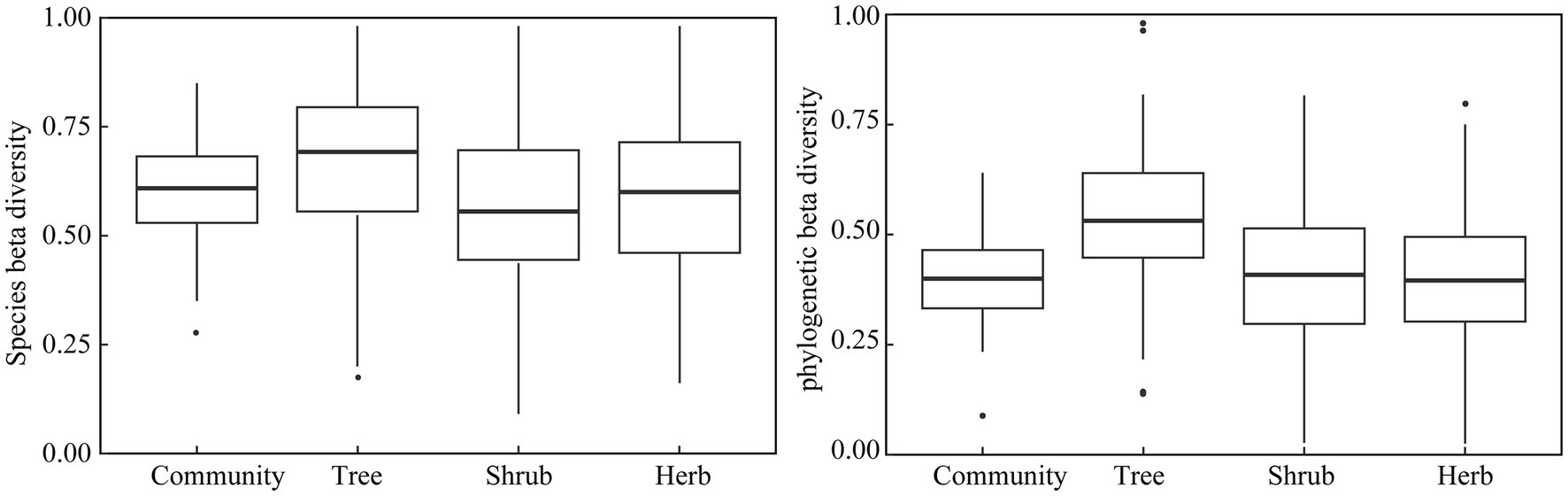
Figure 3. Pairwise beta diversity of plots in dry and hot valley plant communities.
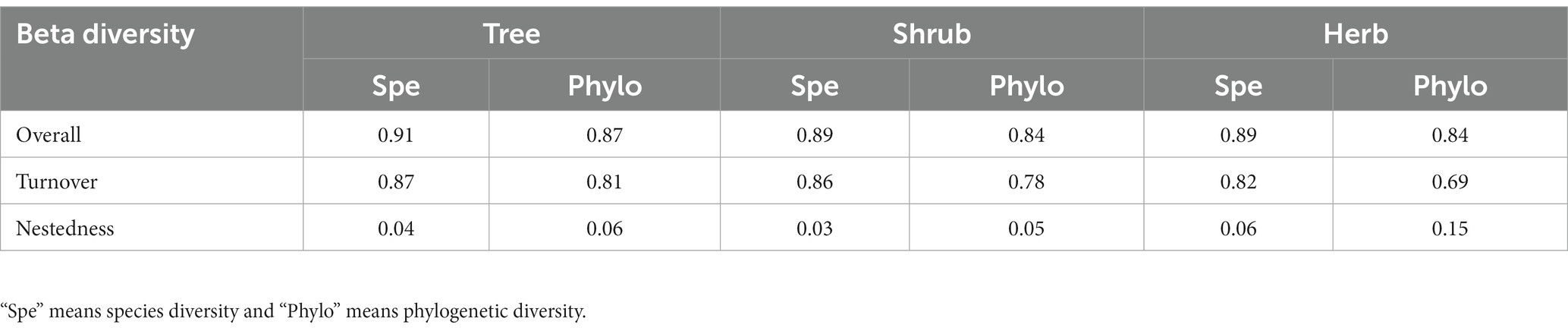
Table 1. Beta diversity and its compositions of the different life forms.
The analyses revealed that different life forms within the dry and hot valley plant communities displayed varying responses to environmental factors. In particular, the Mantel correlation test identified only three environmental factors (pH, MAP, EREF) that exhibited significantly positive correlations with species beta diversity (Figure 4). Specifically, the species beta diversity of trees demonstrated weak associations with the environmental factors assessed. It indicated that within specific life forms, the correlations between environmental factors and beta diversity varied. For instance, soil pH and MAP were significantly and positively correlated with the species beta diversity of shrubs, while only EREF factors demonstrated a significant positive correlation with the species beta diversity of herbs. Moreover, the associations observed for phylogenetic beta diversity were consistent with those observed for species beta diversity (Table 2).
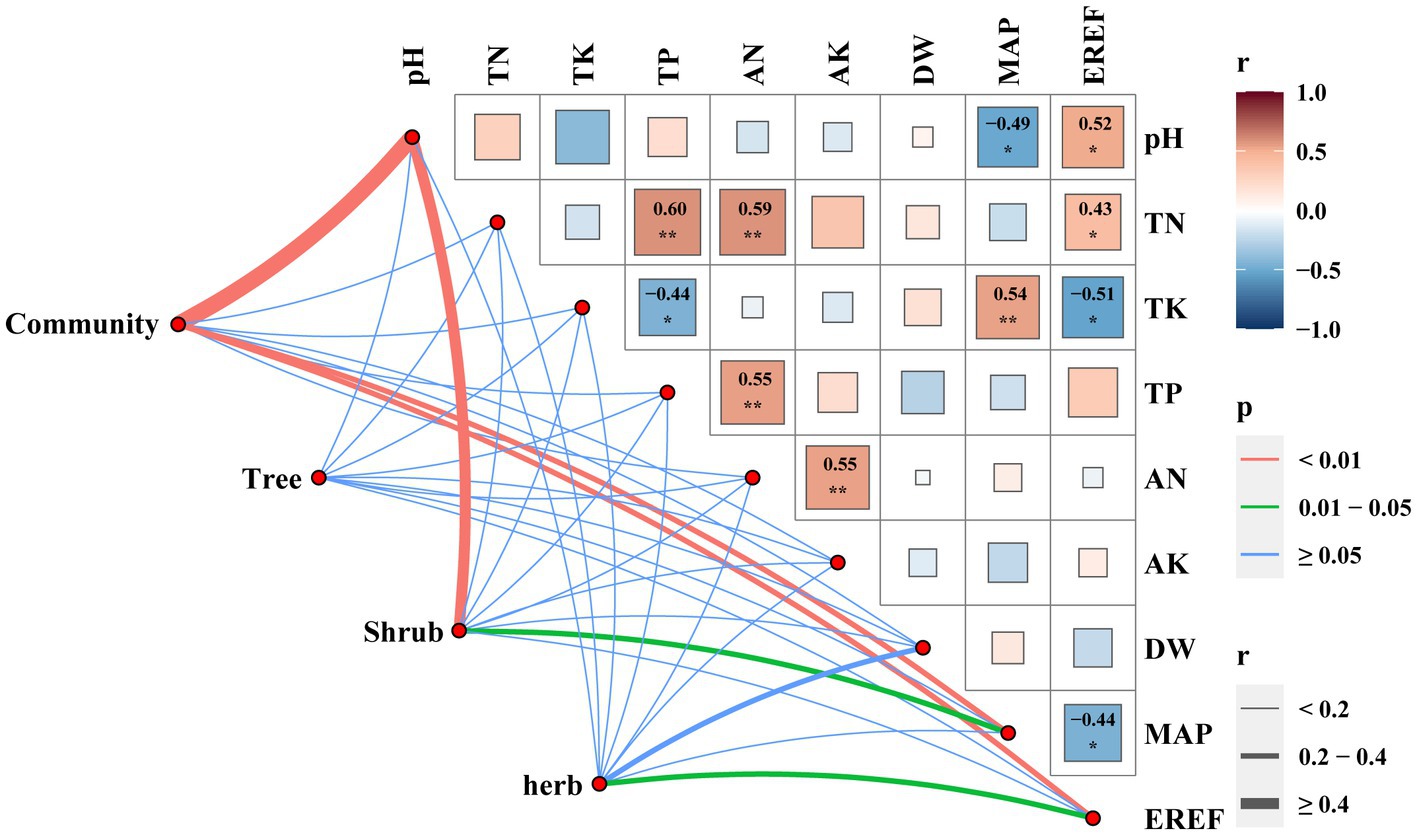
Figure 4. Mantel correlations between species beta diversity and environmental factors.
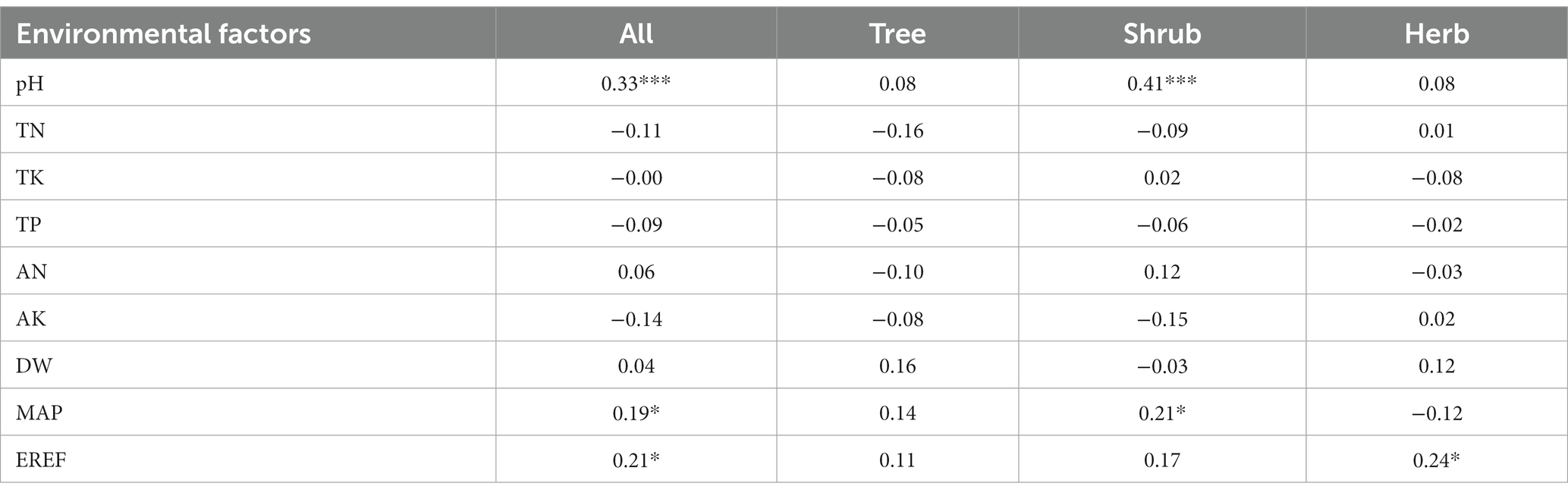
Table 2. Mantel correlations between phylogenetic beta diversity and environmental factors.
We utilized distance-based redundancy analysis (db-RDA) and hierarchical partitioning (HP) to evaluate the relative contribution of each environmental factor to beta diversity (Figure 5). The analysis revealed that the contribution of environmental factors to both species and phylogenetic beta diversity within the dry and hot valley plant community of Nujiang accounted for 34.40 and 29.20%, respectively, with climate factors and pH demonstrating the largest effects. The findings demonstrated that EREF, pH, MAT, and AHM were the most important environmental factors influencing both species and phylogenetic beta diversity. Moreover, the analyses revealed that the impact of environmental factors on beta diversity varied across different life forms. For instance, environmental factors accounted for 60.20% of the explanatory contribution for species beta diversity in herbs, whereas in trees, this figure was only 30.80%. Similarly, we observed that for phylogenetic beta diversity, shrubs displayed the greatest explanatory contribution from environmental factors (58.00%), while herbs exhibited the smallest contribution (15.60%). The analyses revealed that the central factors influencing both species and phylogenetic beta diversity within tree and shrub populations were EREF and TP, and EREF and pH, respectively. In contrast, we observed that ELV and AHM played the most prominent roles in influencing species beta diversity among herb populations, while AN and TK were identified as key factors influencing phylogenetic beta diversity among herbs.
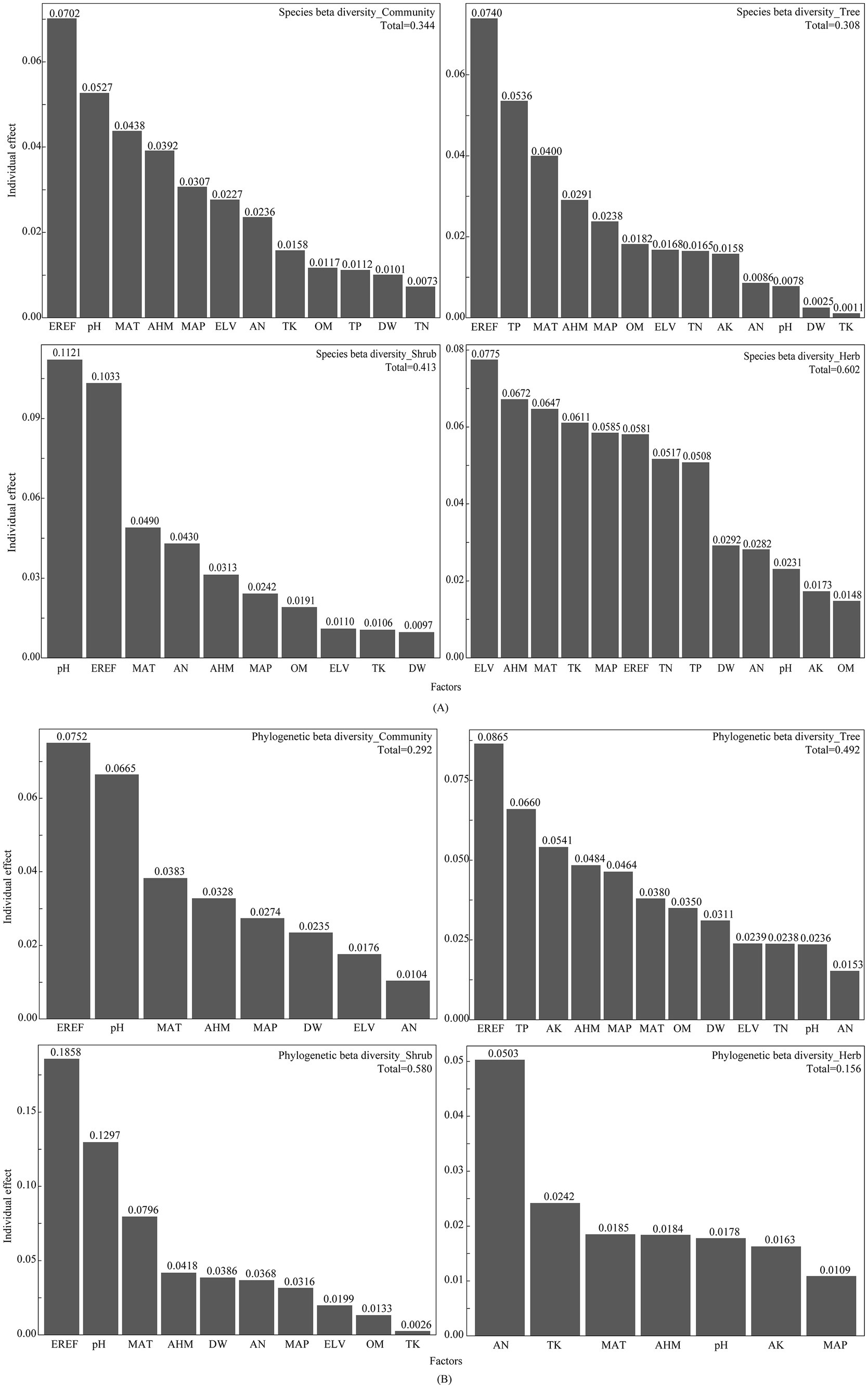
Figure 5. db-RDA and HP between beta diversity and environmental factors: (A) species beta diversity; (B) phylogenetic beta diversity.
Mantel test results between beta diversity with spatial and environmental distance showed that species beta diversity and phylogenetic beta diversity were significantly correlated with environmental and dispersal limitations (Figure 6). The hierarchical partitioning (HP) of the db-RDA analyses indicated that both environmental and geographical limitations had a significant impact on both species and phylogenetic beta diversity (Figure 7). Collectively, these two factors accounted for 32.8 and 31.70% of the total contribution to species and phylogenetic beta diversity, respectively. Regarding species beta diversity, the analysis indicated that dispersal limitations accounted for only 4.46% of the total variation, while environmental limitations explained 28.23%. Furthermore, the findings suggested that soil factors demonstrated a larger explanatory contribution than both climate and topographic factors. Regarding phylogenetic beta diversity, the results showed that dispersal limitation accounted for only 5.66% of the total variation, while the remaining variation was explained by environmental limitations (26.01%). The findings also suggested that climate factors exhibited a larger explanatory contribution compared to both soil and topographic factors.
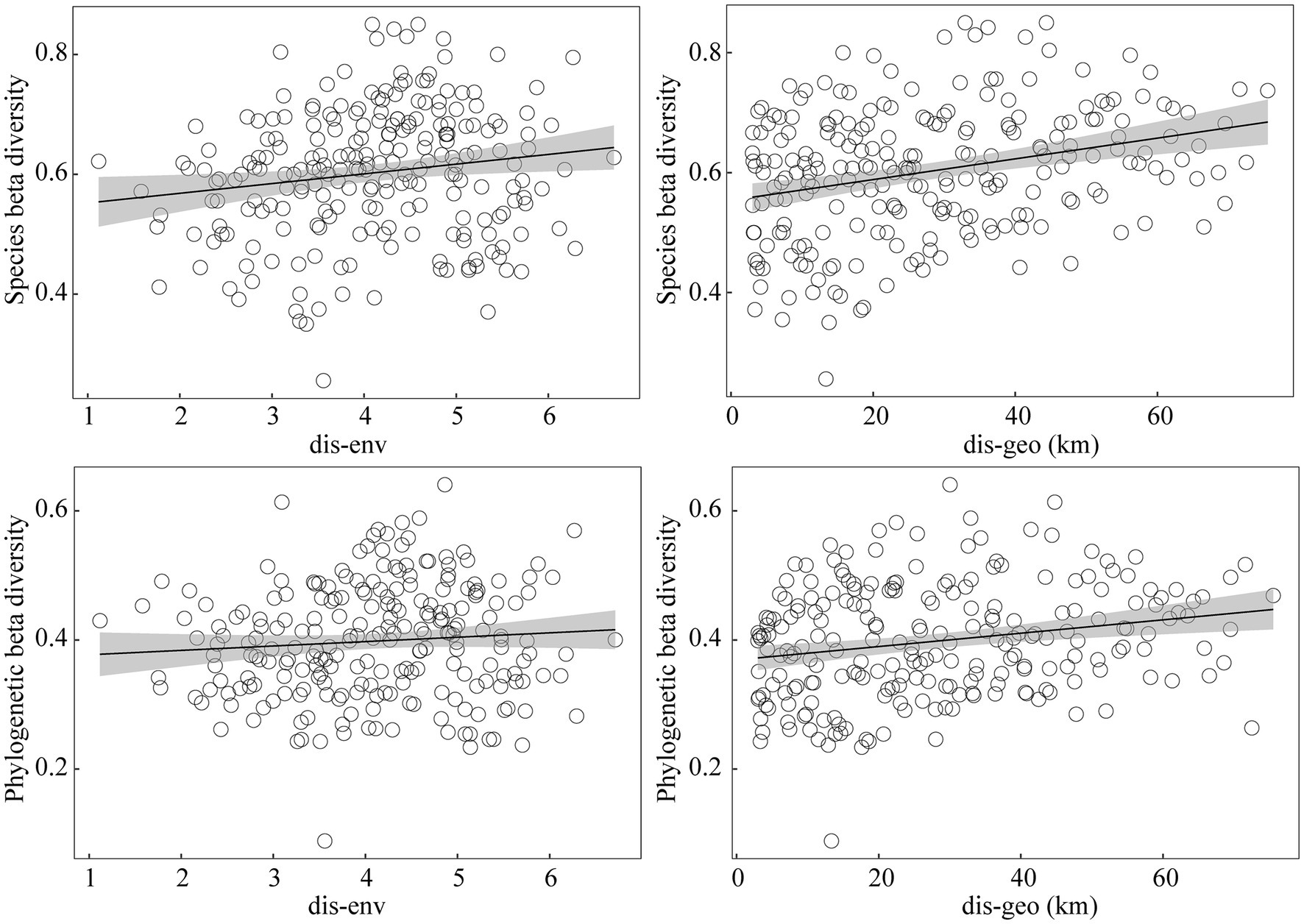
Figure 6. Mantel correlations between beta diversity with spatial and environmental distance.
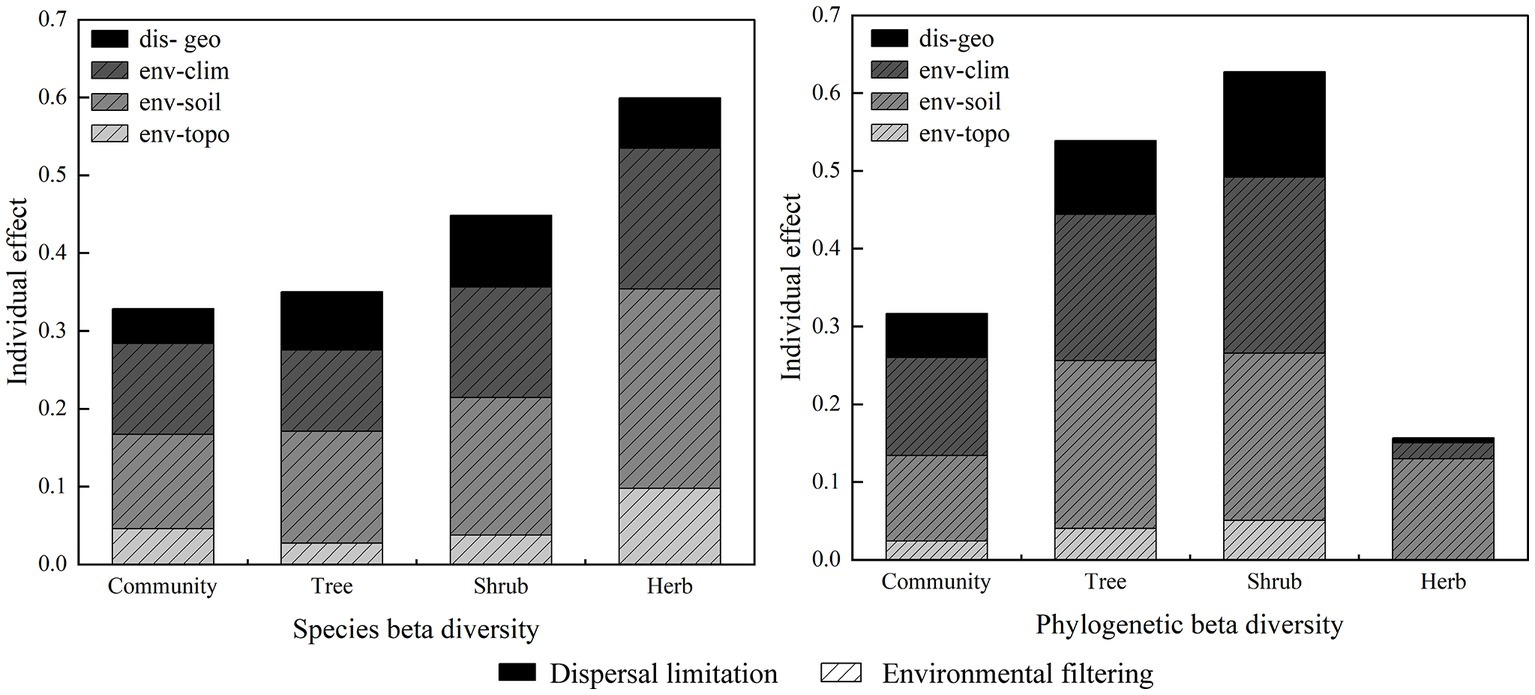
Figure 7. HP of db-RDA between beta diversity and environmental filtering and dispersal limitation.
The findings indicated that the total contribution of environmental and dispersal limitations to species beta diversity varied across different life forms (Figure 7). Specifically, herbs exhibited the highest total contribution (60.0%), followed by shrubs (48.8%) and trees (35.0%). Moreover, it observed that the total contribution of environmental and dispersal limitations to phylogenetic beta diversity was highest among shrubs (62.7%), followed by trees (53.9%) and herbs (15.6%). Environmental limitations exhibited a greater contribution than geographical limitations to both species and phylogenetic beta diversity across different life forms. Among the various environmental factors evaluated, the findings demonstrated that soil factors exerted the greatest impact on both species and phylogenetic beta diversity across different life forms. In contrast, we observed that for the phylogenetic beta diversity of shrubs, climate factors exhibited the most pronounced impact.
The study of beta diversity and the distribution patterns of its components holds significant implications for both ecology and biodiversity conservation (Baselga, 2010). In the investigation of the Nujiang dry and hot valley, we observed that plant community composition variations were largely attributed to turnover components, with the effect of richness being relatively minor. After the Miocene, the region of northwest Yunnan experienced significant geological changes, with the west end of the right-lateral axis moving northward due to plate collision and Himalayan uplift (Shi et al., 1999; Mehrotra et al., 2005). These events have resulted in a shift from tropical to subtropical climate in the area. The distinct environmental characteristics of the dry and hot valley have provided unique pathways and shelters for several species and allowed for the preservation of ancient and tropical-origin species during periods of climate change (Li et al., 2000). The dry and hot valley is characterized by natural barriers on either side that serve to limit long-distance species dispersal. Additionally, the complex topography of the region has resulted in heightened habitat heterogeneity, resulting in reduced gene exchange and an increased rate of species turnover. However, Zhang et al. (2015) found in the flora of plants in the Nujiang River that the wide channel downstream of Liuku has facilitated the migration of tropical Asian plants from south to north, making it easier for plant species to colonize the region. A previous research in the Himalayas and Hengduan Mountains in southwest China (Qian et al., 2020) has revealed that small variations in community beta diversity and high turnover components are often influenced by differences in altitude and environmental conditions. The results of this study were consistent. Moreover, given that various factors influence species turnover and nestedness, the development of diverse conservation strategies is critical (Baselga, 2010). Selecting communities with high species richness for single-point protection can better maintain vegetation beta diversity when nestedness components are dominated by species richness (Bergamin et al., 2017). Conversely, in cases where species turnover is the dominant component, establishing small nature reserves focused on the conservation of multiple dominant plant species simultaneously will be a more fruitful approach (Li et al., 2021). In the case of plant communities in the dry and hot valleys where turnover components dominate, it has been suggested the latter method of protection. In this regard, species such as Castanopsis delavayi and Pistacia weinmanniifolia shrubs have been recommended as ideal choices for conservation efforts in the Nujiang dry and hot valley. The analysis revealed that both phylogenetic beta diversity and its turnover components were lower compared to species diversity and its components, suggesting that species turnover with closely related phylogenetic relationships predominantly influenced beta diversity patterns (Qian et al., 2013). Indeed, research has demonstrated that species belonging to the same taxonomic unit, family, or genus share similar evolutionary histories, resulting in phylogenetic clustering (Webb et al., 2002). We observed that in the Nujiang dry and hot valley, several dominant species belonging to different life forms shared the same family. For instance, genera such as Toxicodendron, Lannea, and Pistacia were all members of the family Anacardiaceae. The woody dominant species such as Toxicodendron grandiflorum, Lannea coromandelica and Pistacia weinmanniifolia, despite being different genera and species, share the same family Anacardiaceae. Similarly, herb-dominant species such as Eragrostis ferruginea, Arthraxon prionodes, and Heteropogon contortus all belong to the relatively recent evolutionary family Poaceae (Jiang et al., 2019; Li et al., 2021).
The analysis of influencing factors has revealed that both environmental and geographical limitations significantly constrained the maintenance of beta diversity in plant communities in the Nujiang dry and hot valley. We observed that the impact of environmental limitations was notably higher, being 4–6 times greater than that of dispersal limitations, with climate and soil factors playing a critical role in environmental limitations (Andino et al., 2021). In contrast to the findings observed in the Dulongjiang and Yuanjiang valleys of Southwest China, the results indicated that geographic distance played a relatively minor role at the local scale in the Nujiang dry and hot valley (Li et al., 2016; Meng et al., 2017). As compared to Southeast Yunnan, the climate and plant floras of Northwest Yunnan have undergone significant changes following the plate movement, with species turnover being an adaptation strategy for species survival (Blundo et al., 2016). The vegetation in Northwest Yunnan has undergone significant changes over time, with the original tropical vegetation being gradually replaced by subtropical evergreen broadleaf forests. The examination of beta diversity in tropical and temperate forests revealed that environmental limitations played a more significant role in temperate forests, whereas dispersal limitations was more pronounced in tropical forests (Myers et al., 2013). At higher elevations, adjacent sites exhibited a wider ecological range, resulting in more similar species compositions (Tang and Fang, 2004). The plant communities in Northwest Yunnan have been shaped by a complex interplay of factors, including the Himalayan-Tibetan plateau and the Glacial–Interglacial cycle. The analysis suggests that the limitations imposed by hydrothermal conditions are particularly pronounced in the region. Given the historical and climactic changes observed in the area, it is evident that species with close phylogenetic relationships and similar adaptation abilities have been selectively coexisting in Northwest Yunnan as opposed to the dry and stable conditions observed in Southeast Yunnan. The analysis of different life forms among abundant woody plants in the Nujiang dry and hot valley has revealed that environmental factors, particularly hydrothermal conditions such as EREF, have a greater impact on the phylogenetic structure of trees and shrubs than on species beta diversity. Furthermore, the influencing factors of beta diversity are subject to constraints imposed by the study scale, with the environmental effect becoming more pronounced with increased scale, and dispersal limitation being more important in smaller scales (Chase, 2014; Zhao et al., 2017). In the valley regions, human activities and frequent species exchange have had a significant impact on plant communities, accelerating species propagation and weakening the influence of geographic distance on community composition (Cao and Jin, 1993; Li et al., 2016).
The analysis of specific environmental factors highlights that, in addition to hydrothermal conditions, different life forms in the valley exhibit varying responses to environmental factors. The dry and hot climate in the region has made it challenging for slope vegetation to recover, resulting in the widespread distribution of sparse shrubs and herbs with long lifespans (Cao and Jin, 1993). The investigation of Castanopsis delavayi in the Nujiang dry and hot valley revealed a significant absolute dominance cluster distribution pattern, with its companion species exhibiting more constrained distribution patterns that result in a fragmented distribution of woody plants, favoring the entry of herbs (Li et al., 2021). Such differences in the beta composition patterns of different life forms may contribute to variations in plant community composition. The analysis has shown that climate factors are stronger predictive factors at wider spatial scales (50 km2), whereas soil and topographic factors become relatively more important at medium (5 km2) and finer (0.5 km2) scales (Andino et al., 2021). Soil factors, in particular, were identified as important limiting factors in this study. Soil N, P, and K were found to play a critical role in maintaining plant productivity, with pH regulating nutrient availability and controlling the chemical elements of P and K, thereby influencing species nutrient uptake and impacting community beta diversity (John et al., 2007; Kong et al., 2018).
In addition, our study has identified environmental and dispersal limitations as the primary limiting factors that impact beta diversity and species turnover. However, despite these findings, there remains a significant amount of unexplained variation. The limited number of measured local environmental factors suggests that other non-biological factors (such as paleoclimate and additional soil factors), biological factors (including community structure), and species pools may also play a role in constraining the maintenance of beta diversity in communities (Myers et al., 2013; Zheng et al., 2019). As such, further analysis of the impact of additional environmental parameters will be necessary to more fully elucidate the complex mechanisms that govern community dynamics in this region.
The vegetation of the dry and hot valley is a rare and endangered ecosystem in Southwest China, and the study of its beta diversity components provides important insights into the biological conservation strategies and the limiting effects of environmental factors. Specifically, the investigation focused on the composition patterns and drivers of species and phylogenetic beta diversity in the Nujiang dry and hot valley in Northwest Yunnan, Southwest China. The investigation revealed three important findings. Firstly, both species and phylogenetic beta diversity were dominated by turnover components, underscoring the importance of establishing multiple nature reserves to protect multiple dominant species (such as Castanopsis delavayi and Pistacia weinmanniifolia shrubs) to maintain the unique vegetation found in the dry and hot valley. Phylogenetic beta diversity and its turnover components were lower than those of species diversity, indicating that closely related phylogenetic relationships dominated the beta diversity patterns. Secondly, the combined influence of environmental and dispersal limitations on beta diversity in the dry and hot valleys of the Nujiang region, with environmental limitations being the more dominant factor. Against the backdrop of geological history and environmental changes, climate and soil factors play a critical role in shaping the unique ecosystem of the hot and dry valley of Nujiang, with hydrothermal conditions such as EREF being particularly important. Thirdly, climate factors (including EREF and MAP) and soil factors (such as pH) exhibit a significant positive correlation with both species and phylogenetic beta diversity. These factors contribute significantly to the variation observed in specific environmental factors, accounting for the largest proportion of the variance. The investigation suggests that the growth and development of different life forms in the community are restricted by a diverse range of environmental factors, highlighting the complex ecological interactions and processes that govern community dynamics in this region. We recommend that future research expand beyond the study of beta diversity components to include other aspects of biodiversity, such as functional diversity. In addition, further investigation into the role of other limiting factors that may impact biodiversity maintenance in the region is warranted. By enriching the understanding of the various mechanisms that underlie biodiversity maintenance, we can provide a more comprehensive theoretical basis for driving ecosystem functions in the region.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
ZZ, YL, and YC: conception and design, manuscript writing. LH, GL, and SM: data collection, data analysis, and visualization. MC: supervision and writing – review and editing. All authors contributed to the article and approved the submitted version.
This work is supported by the National Key Research and Development Program of China (No. 2022YFF13024), the National Natural Science Foundation of China (No. 31860045) and Candidates of the Young and Middle-Aged Academic Leaders of Yunnan Province of China (No. 202205 AC160041).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer XH declared a shared affiliation with the authors ZZ, MC, and YL to the handling editor at the time of review.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1199874/full#supplementary-material
Andino, J. E. G., Pitman, N. C. A., ter Steege, H., Peralvo, M., Ceron, C., and Fine, P. V. A. (2021). The contribution of environmental and dispersal filters on phylogenetic and taxonomic beta diversity patterns in Amazonian tree communities. Oecologia 196, 1119–1137. doi: 10.1007/s00442-021-04981-0
Asefa, M., Brown, C., Cao, M., Zhang, G., Ci, X., Sha, L., et al. (2019). Contrasting effects of space and environment on functional and phylogenetic dissimilarity in a tropical forest. J. Plant Ecol. 12, 314–326. doi: 10.1093/jpe/rty026
Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 19, 134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Baselga, A. (2012). The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 21, 1223–1232. doi: 10.1111/j.1466-8238.2011.00756.x
Bergamin, R. S., Bastazini, V. A. G., Vélez-Martin, E., Debastiani, V., Zanini, K. J., Loyola, R., et al. (2017). Linking beta diversity patterns to protected areas: lessons from the Brazilian Atlantic rainforest. Biodivers. Conserv. 26, 1557–1568. doi: 10.1007/s10531-017-1315-y
Blundo, C., Gonzalez-Espinosa, M., and Malizia, L. R. (2016). Relative contribution of niche and neutral processes on tree species turnover across scales in seasonal forests of NW Argentina. Plant Ecol. 217, 359–368. doi: 10.1007/s11258-016-0577-x
Cao, Y. H., and Jin, Z. Z. (1993). A research on the vegetation of Nujiang dry-hot river valley in Lijiangba of Yunnan. Cuihaia. 2, 132–138.
Chase, J. M. (2014). Spatial scale resolves the niche versus neutral theory debate. J. Veg. Sci. 25, 319–322. doi: 10.1111/jvs.12159
Dormann, C. F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al. (2013). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Du, S. K., Tang, G. Y., Liu, Y. G., Lei, C. Y., Xu, Y. J., Ruan, C. M., et al. (2022). Plant diversity in various sections of Jinsha River dry-hot valley under different site environments. J. Zhejiang A&F Univ. 39, 742–749. doi: 10.11833/j.issn.2095-0756.20210572
He, G. X., Shi, Z. T., Yan, B. G., Yang, H. Z., Sun, Y., Wang, Y. D., et al. (2023). Effects of fencing enclosure on interspecific associations in a savanna grassland community in China’s arid-hot valley region. Acta Pratac. Si. 32, 1–14. doi: 10.11686/cyxb2022080
He, Y., Liang, S., Liu, R., and Jiang, Y. (2022). Beta diversity patterns unlock the community assembly of woody plant communities in the riparian zone. Forests 13:673. doi: 10.3390/f13050673
Kong, J. J., Yang, J., and Bai, E. (2018). Long-term effects of wildfire on available soil nutrient composition and stoichiometry in a Chinese boreal forest. Sci. Total Environ. 642, 1353–1361. doi: 10.1016/j.scitotenv.2018.06.154
Jiang, X. M., Ding, C., Brosse, S., Pan, B., Lu, Y., Xie, Z. C., et al. (2019). Local rise of phylogenetic diversity due to invasions and extirpations leads to a regional phylogenetic homogenization of fish fauna from Chinese isolated plateau lakes. Ecol. Indic. 101, 388–398. doi: 10.1016/J.ECOLIND.2019.01.041
Jin, Y., and Qian, H. (2022). V.PhyloMaker2: an updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Divers. 44, 335–339. doi: 10.1016/j.pld.2022.05.005
Jin, Z. Z. (1999). A phytosociological study on the semi-savanna vegetation in the dry-hot valleys of Yuanjiang River. Yunnan. Guihaia. 19, 289–302. doi: 10.3969/j.issn.1000-3142.1999.04.001
Jin, Z. Z., and Ou, X. K. (2000). Yuanjiang, Nujiang, Jinshajiang, Lancangjiang vegetation of dry-hot valley. Kunming: Yunnan University Press, Yunnan Science and Technoligy Press.
John, R., Dalling, J. W., Harms, K. E., Yavitt, J. B., Stallard, R. F., Mirabello, M., et al. (2007). Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. 104, 864–869. doi: 10.1073/pnas.0604666104
Kembel, S. W., Cowan, P. D., Helmus, M. R., Cornwell, W. K., Morlon, H., Ackerly, D. D., et al. (2010). Picante: r tools for integrating phylogenies and ecology. Bioinformatics 26, 1463–1464. doi: 10.1093/bioinformatics/btq166
Lai, J. S., Zou, Y., Zhang, J. L., and Peres-Neto, P. (2022). Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.Hp R package. Methods Ecol. Evol. 13, 782–788. doi: 10.1111/2041-210X.13800
Leprieur, F., Albouy, C., De Bortoli, J., Cowman, P. F., Bellwood, D. R., and Mouillot, D. (2012). Quantifying phylogenetic beta diversity: distinguishing between 'true' turnover of lineages and phylogenetic diversity gradients. PLoS One 7:e42760. doi: 10.1371/journal.pone.0042760
Li, D. D., Zhang, F., Luo, G. Y., Zhang, T., Lv, J. Q., Wang, W. C., et al. (2023). Taxon-dependent effects of dispersal limitation versus environmental filters on bryophyte assemblages-multiple perspective studies in land-bridge islands. Ecol. Evol. 13:e9844. doi: 10.1002/ece3.9844
Li, F., Yan, Y., Zhang, J., Zhang, Q., and Niu, J. (2021). Taxonomic, functional, and phylogenetic beta diversity in the Inner Mongolia grassland. Glob. Ecol. Conserv. 28:e01634. doi: 10.1016/j.gecco.2021.e01634
Li, X. H., Liu, Y. H., Liu, Y., Xu, Y., Yang, Y., and Shen, Z. H. (2016). Impacts of geographical distances and environmental differences on the beta diversity of plant communities in the dry-hot valley of the Yuanjiang River. Biodivers. Sci. 24, 399–406. doi: 10.17520/biods.2015245
Liu, Y., Xu, Y., Shi, S. L., Peng, P. H., and Shen, Z. H. (2016). Quantitative classification and environmental interpretations for the structural differentiation of the plant communities in the dry valley of Jinshajiang River. Biodivers. Sci. 24, 407–420. doi: 10.17520/biods.2015353
Liu, F. Y., Gao, C. J., Feng, D. F., and L, K., (2021). Effects of local species pool on plant diversity in dry and hot valley of Yunnan Province. J. W. China Forest. Sci. 50, 64–72. doi: 10.16473/j.cnki.xblykx1972.2021.05.010
Long, C., Yu, Z. X., Yang, Y. Q., and Shui, M. M. (2022). Research on spatial pattern and interspecific association of dominant populations in secondary savanna shrub forest in dry-hot valley. J. Sichuan Forest. Sci. Technol. 43, 44–52. doi: 10.12172/202109020002
Ma, H. C., Wu, J. R., Zheng, C. L., and Tang, J. R. (2020). Discussion on the formation characteristics of dry-hot valley and related problems of revegetation. J. S. W. Forest. Univ. 40, 1–8. doi: 10.11929/j.swfu.202003080
Mariano, R. F., Rezende, V. L., Mendes, C. N., dos Santos, R. M., de Souza, C. R., de Moura, A. S., et al. (2020). Phylogenetic beta diversity in an upper montane Atlantic Forest along an altitudinal gradient. Plant Ecol. 221, 671–682. doi: 10.1007/s11258-020-01041-0
Mehrotra, R., Liu, X. Q., Li, S. C., Wang, Y. F., and Chauhan, M. S. (2005). Comparison of the tertiary flora of Southwest China and Northeast India and its significance in the antiquity of the modern Himalayan flora. Rev Palaeobot Palno. 135, 145–163. doi: 10.1016/j.revpalbo.2005.03.004
Meng, G. T., Chai, Y., Yuan, C. M., Ai, H., Li, G. X., Wang, Q., et al. (2013). Community characteristics of the mid-montane humid ever-green broad-leaved forest in gaoligong mountains. Yunnan. Scientia Silvae Sinicae. 49, 144–151. doi: 10.11707/j.1001-7488.20130320
Meng, Y., Qiu, Y., Zhang, L., Wang, C., Zang, Z., Zhang, X., et al. (2017). Effects of geographical distance and differences in climate and altitude on species dissimilarity of vascular plant communities in the Dulongjiang River watershed area. Biodivers. Sci. 25, 1313–1320. doi: 10.17520/biods.2017076
Myers, J. A., Chase, J. M., Jiménez, I., Jørgensen, P. M., Araujo-Murakami, A., Paniagua-Zambrana, N., et al. (2013). Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly. Ecol. Lett. 16, 151–157. doi: 10.1111/ele.12021
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2019). Vegan: Community ecology package. Available online at: https://CRAN.R-project.org/package=vegan (Accessed October 11, 2022).
Pereira, I. S., Rezende, V. L., Meira-Neto, J. A. A., Clappe, S., and Eisenlohr, P. V. (2021). Phylogenetic structure as a predictive component of beta diversity: lessons from a comprehensive Neotropical biogeographic transition. Perspect. Plant Ecol. Evol. Syst. 49:125602. doi: 10.1016/j.ppees.2021.125602
Qian, H., Swenson, N. G., and Zhang, J. L. (2013). Phylogenetic beta diversity of angiosperms in North America. Glob. Ecol. Biogeogr. 22, 1152–1161. doi: 10.1111/geb.12076
Qian, H., Jin, Y., Leprieur, F., Wang, X. L., and Deng, T. (2020). Geographic patterns and environmental correlates of taxonomic and phylogenetic beta diversity for large-scale angiosperm assemblages in China. Ecography 43, 1706–1716. doi: 10.1111/ecog.05190
R Core Team. (2021). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
She, D. Q., Zhang, X. T., Xiao, L., Zhong, Z. L., Wang, H. M., and Wang, W. J. (2022). Plant beta diversity and its influence factors in the Liangshui National Nature Reserve in the central region of the Xiaoxing’an mountains. Biodivers. Sci. 30:21274. doi: 10.17520/biods.2021274
Shi, Y. F., Li, J. Y., Li, B. Y., Yao, T. D., and Zhang, Q. S. (1999). Uplift of the Qinghai Xizang (Tibetan) plateau and East Asia environmental change during late Cenozoic. Acta Geograph. Sin. 54, 10–21. doi: 10.11821/xb199901002
Shi, W., Wang, Y. Q., Xiang, W. S., Li, X. K., and Cao, K. F. (2021). Environmental filtering and dispersal limitation jointly shaped the taxonomic and phylogenetic beta diversity of natural forests in southern China. Ecol. Evol. 11, 8783–8794. doi: 10.1002/ece3.7711
Socolar, J. B., Gilroy, Z. J. J., Kunin, W. E., and Edwards, D. P. (2016). How should beta-diversity inform biodiversity conservation? Trends Ecol. Evolution 31, 67–80. doi: 10.1016/j.tree.2015.11.005
Soininen, J., McDonald, R., and Hillebrand, H. (2007). The distance decay of similarity in ecological communities. Ecography 30, 3–12. doi: 10.1111/j.2006.0906-7590.04817.x
Tang, Z. R., and Fang, J. Y. (2004). A review on the elevational patterns of plant species diversity. Chinese Biodiversity. 12, 20–28. doi: 10.3321/j.issn:1005-0094.2004.01.004
Tilman, D. (1982). Resource competition and community structure. Monogr. Popul. Biol. 17, 1–296. doi: 10.1515/9780691209654
Yang, H., Wang, Y., Wang, J. M., Xia, Y. G., Li, J. W., Jia, X. H., et al. (2021). Effects of environmental filtering and dispersal limitation on the β-diversity of plant communities in the south fringe of Kumtag Desert. J. Desert Res. 41, 147–154. doi: 10.7522/j.issn.1000-694X.2020.00129
Yang, Y., Han, J., Liu, Y., Zhongyong, C., Shi, S. L., Sina, C., et al. (2016). A comparison of the altitudinal patterns in plant species diversity within the dry valleys of the three parallel Rivers region, northwestern Yunnan. Biodivers. Sci. 24, 440–452. doi: 10.17520/biods.2015361
Webb, C. O. (2000). Exploring the phylogenetic structure of ecological communities: an example for rain forest trees. Am. Nat. 156, 145–155. doi: 10.1086/303378
Webb, C. O., Ackerly, D. D., McPeek, M. A., and Donoghue, M. J. (2002). Phylogenies and community ecology. Annu. Rev. Ecol. Evol. Syst. 33, 475–505. doi: 10.1146/annurev.ecolsys.33.010802.150448
Whittaker, R. H. (1960). Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 30, 279–338. doi: 10.2307/1943563
ZHAO Mingfei, 赵., XING Kaixiong, 邢., WANG Yuhang, 王., WANG Guoyi, 王., XUE Feng, 薛., ZUO Wanyi, 左., et al. (2017). Patterns and determinants of beta diversity in the understory vegetation layers of montane boreal conifer forest in Luya Mountain. Acta Ecol. Sin. 37, 3327–3334. doi: 10.5846/stxb201603050390
Zhang, J. L., Liu, X. K., Shen, R., Shi, W., Zhang, Z. M., Zhou, R., et al. (2010). The structure and similarity characteristic of the grassland community in hot-dry valley upper middle and lower of Jinsha River. Energy Environ. Sci. 19, 1272–1277. doi: 10.16258/j.cnki.1674-5906.2010.06.002
Zhang, X. K., Du, F., Chen, J. S., and Zhou, D. (2015). Endemism in the flora of seed plants in Nu River valley of Yunnan Province. Plant J. 33, 302–310. doi: 10.11913/PSJ.2095-0837.2015.30302
Zhang, Y. B., Wu, H. D., Yang, J., Song, X. Y., Yang, D., and He, F. L. (2020). Environmental filtering and spatial processes shape the beta diversity of liana communities in a valley savanna in Southwest China. Appl. Veg. Sci. 23, 482–494. doi: 10.1111/avsc.12514
Zheng, Y., Dong, L., Li, Z., Zhang, J., Li, Z., Miao, B., et al. (2019). Phylogenetic structure and formation mechanism of shrub communities in arid and semiarid areas of the Mongolian plateau. Ecol. Evol. 9, 13320–13331. doi: 10.1002/ece3.5787
Zhu, H., and Yan, L. C. (2003). Notes on the realities and significances of the “Tanaka link” and the “Ecogeographical diagonal line” in Yunnan. Advance in Earth Sciences. 18, 870–876.
Keywords: beta diversity, dry and hot valley, environmental filtering, dispersal limitation, Nujiang River
Citation: Zhao Z, He L, Li G, Ma S, Cui M, Liu Y and Chai Y (2023) Partitioning beta diversity of dry and hot valley vegetation in the Nujiang River in Southwest China. Front. Ecol. Evol. 11:1199874. doi: 10.3389/fevo.2023.1199874
Edited by:
Tongli Wang, University of British Columbia, CanadaReviewed by:
Xiaobo Huang, Chinese Academy of Forestry, ChinaCopyright © 2023 Zhao, He, Li, Ma, Cui, Liu and Chai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuguo Liu, bGl1eXVndW9AY2FmLmFjLmNu; Yong Chai, Y2hhaXlvbmdAeWFmZy5hYy5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.