 Ĺubomír Kováč1*
Ĺubomír Kováč1* Martina Žurovcová2Natália Raschmanová1Nikola Jureková1Vladimír Papáč3Andrea Parimuchová1
Martina Žurovcová2Natália Raschmanová1Nikola Jureková1Vladimír Papáč3Andrea Parimuchová1- 1Department of Zoology, Faculty of Science, Institute of Biology and Ecology, Pavol Jozef Šafárik University, Košice, Slovakia
- 2Institute of Entomology, Biology Centre AS CR v. v. i., České Budějovice, Czechia
- 3Slovak Caves Administration, State Nature Conservation SR, Rimavská Sobota, Slovakia
Introduction: Using an integrative taxonomic approach, we investigated the morphological and molecular characters to identify the species of the genus Pseudosinella occurring in caves of the Western Carpathians and to clarify their phylogenetic relationships. Based on morphological characters, we hypothesized that Pseudosinella aggtelekiensis (Stach, 1929) and Pseudosinella paclti (Rusek, 1961) originated from different phyletic lineages.
Methods: We used the barcoding fragment of the mtDNA COI gene from 87 individuals from 16 caves to reconstruct the phylogenetic history of the genus Pseudosinella.
Results: The molecular phylogenetic tree revealed two distinct species groups with allopatric distributions. The first group consisted of P. aggtelekiensis populations from the Slovak Karst and three undescribed species from fragmented and isolated karst areas in southeastern Slovakia. The second group, P. paclti populations in the Central Western Carpathians, was merged. This group included Pseudosinellamuranensis, a new species taxonomically described in this work. It is characterized by highly developed troglomorphic features and is restricted to a small karst area, the Muranska planina Plateau. The phylogeny of the genus Pseudosinella from the caves of the Western Carpathians revealed a Miocene diversification. According to molecular calibration, the two distinct Pseudosinella lineages were separated in the Middle Miocene, about 14.51 Mya, followed by further diversification in the P. paclti lineage 10.89 Mya and in the P. aggtelekiensis lineage 11.14 Mya.
Discussion: This phylogeny is consistent with the uplift of Triassic limestones during the early formation of the Western Carpathians in the Paratethys region and the initial development of caves during this period. The study provides further important evidence that the Western Carpathians played a significant role as an independent speciation center of the obligate cave fauna in Europe.
1. Introduction
Collembolans are “wingless” hexapods, one of the most abundant and diverse groups of soil mesofauna with nearly 9,000 species described to date (Bellinger et al., 1996–2022), probably accounting for only about 20% of the hypothetical diversity of this group (Potapov et al., 2020). Collembolans are also among the most diverse and abundant invertebrates in caves and other subterranean habitats (Juberthie, 2000), with over 400 troglobiotic (obligate cave) species known (Deharveng and Bedos, 2018).
The genus Pseudosinella (Schäffer, 1897) belongs to collembolan genera with high species diversity. Members of the genus inhabit a wide range of habitats, from xerophilous environments to wetlands and caves (Winkler et al., 2021). Currently, it includes 374 species (Bellinger et al., 1996–2022; Kováč, pers. database) with a predominantly Holarctic distribution, although recent discoveries in subterranean environments of Brazil (Cipola et al., 2020) and Australia (Guzik et al., 2021) suggest that the diversity of this genus in the Southern Hemisphere is far from well understood. More than 140 species (about 37%) are obligate cave dwellers (Lukić, 2019) with more than 100 species distributed in southern Europe (Fiera et al., 2021).
This genus is apparently a polyphyletic taxon, with phyletic lineages derived from various ancestors of the genus Lepidocyrtus (Bourlet, 1839) (Christiansen, 1961; Gama, 1984; Soto-Adames, 2002; Wang et al., 2004). Pseudosinella representatives can be distinguished from Lepidocyrtus relatives essentially by the reduced number of eyes (Christiansen, 1961), an artificial diagnostic morphological feature that does not account for phylogenetic relationships among these taxa. Species of the genus Pseudosinella generally have additional distinguishing characteristics, such as loss of pigmentation and modified unguis structure. These characters are related to their mode of life, which is restricted to deeper soil layers, shallow subterranean habitats, or caves.
Phylogenetic relationships among higher Collembola taxa have traditionally been based on morphology, but more recently, phylogenetic studies have benefited from molecular data. Internal relationships within Entomobryomorpha subfamilies have been investigated in several molecular-based studies (Leo et al., 2019; Sun et al., 2020; Cucini et al., 2021, Guzik et al., 2021). Within the Lepidocyrtinae, Gama (1984) made the first attempt to reconstruct the complex Pseudosinella phylogeny which was based on presumably non-adaptive morphological characters, namely the modifications of the basal chaetae on the labial triangle, and the presence/absence of specific chaetae on the abdominal segments II and IV. Morphological characters traditionally used for species diagnosis of Lepidocyrtinae taxa, especially head and tergal chetotaxy, are very conservative indicators of genetic divergence (Soto-Adames, 2002) and represent synapomorphies for the subfamily (Zhang et al., 2015). However, the polyphyly of Pseudosinella as indicated by the morphology has not been confirmed at the molecular level (Zhang et al., 2015; Godeiro et al., 2021, 2023).
In terms of geographic distribution, Pseudosinella lineages show obvious subterranean radiation in the Pyrenees (Deharveng and Bedos, 2018), the USA, Mexico (Christiansen and Culver, 1987), and Brazil (Cipola et al., 2020). In Southeast Asia, the diversity of troglobiotic Pseudosinella is concentrated in Sulawesi and Papua New Guinea (Deharveng, 1988; Deharveng and Bedos, 2012). Recently, Guzik et al. (2021) revealed extreme genetic diversity of this genus in the subterranean calcretes of arid Australia.
Based on a study carried out in caves in the eastern USA, Christiansen and Culver (1987) defined levels of Pseudosinella troglomorphy that are confined to specific morphological adaptations to the subterranean domain. Troglomorphic species are essentially recognized by a larger body size, an elongated unguis with reduced internal and lateral teeth, acuminate tenent hair, and an elongated apical mucronal tooth. In contrast to these forms, the highly troglomorphic species represent a more advanced stage of morphological adaptation to caves in which unguis is extremely elongated, the internal and lateral teeth are completely absent, the two basal teeth greatly reduced, tenent hairs acuminate and shortened, and the apical mucronal teeth are markedly elongated. In addition, these “advanced troglomorphs” often have distinctly elongated antennae, Pseudosinella christianseni (Salmon, 1965) being an extreme case. Identical characters of Pseudosinella in the advanced troglomorphy stage were specified by Deharveng (1988). Worldwide, there are about 70 troglomorphic species described mainly from Europe, of which 19 species show advanced troglomorphy. The distribution of these forms agrees well with the observations of Christiansen and Culver (1987), which showed a clear biogeographic pattern of decreasing geographic range with increasing troglomorphy. Moreover, these authors arrived at two important assumptions (implications) that increasing troglomorphy (1) decreases the ability to disperse and (2) points to increasingly earlier times of initial cave colonization.
The geologically complex mountain unit of the Western Carpathians in Central Europe is part of the Carpathian Mountains, one of the biodiversity hotspots in Europe (Mráz and Ronikier, 2016). The present study builds on systematic surveys of cave biota in the Western Carpathians in Slovakia carried out over the last two decades (Kováč et al., 2014). Slovakia is relatively rich in karst, covering an area of over 2,700 km2. In total, more than 7,200 karst and pseudokarst caves have been documented in the country to date (Bella et al., 2018).
So far, two troglomorphic Pseudosinella species are known from the Western-Carpathian caves, but Kováč and Rusek (2012) indicated the presence of several undescribed species. Furthermore, Kováč et al. (2016) pointed out the important role of the Western Carpathians as glacial refugia for subterranean Collembola. The authors recognized five obligate cave Pseudosinella species, three of them undescribed, based on morphological characters. They found that these species are allopatric in distribution and restricted to karst caves in the central and southern parts of the Western-Carpathian region.
The profound differences in morphological characters led us to hypothesize that Pseudosinella aggtelekiensis (Stach, 1929) and Pseudosinella paclti (Rusek, 1961) represent separate phylogenetic lineages. These lineages probably diverged during the pre-Quaternary period when significant palaeogeographic changes occurred in the former Paratethys area. We also considered species with highly evolved troglomorphic features to be evolutionary old taxa.
The aim of the present study was (1) to reconstruct the basic phylogeny of Pseudosinella populations occurring in the Western-Carpathian caves using an approach of integrative taxonomy, i.e., a combination of molecular and morphological characters; and (2) to estimate divergence times of individual Pseudosinella cave lineages based on molecular calibration.
2. Materials and methods
2.1. Morphological examination
For morphological examination, specimens were mounted individually on slides in Swann medium (Liquido de Swann), modified from Rusek (1975), and examined with a Carl Zeiss Axio 5 phase-contrast microscope and a Leica DM 2500 microscope equipped with DIC optics (differential interference contrast), a measuring micrometric eyepiece, and a drawing arm. Images were captured using a Carl Zeiss Axiocam 208 color camera and ZEN imaging software. The drawings were processed using Adobe Photoshop CS6.
Abbreviations used in text, tables, and figures are as follows: Abd., abdominal tergum; al, anterolateral sensillum; accp, accessorial p–sensillum; Ant., antennal segment; a.s.l., above sea level; cm, conical microchaeta; IBE FS UPJS, Institute of Biology and Ecology Faculty of Science, P. J. Šafárik University, Košice; mac, macrochaeta; mic, microchaeta; psp, pseudoporus; ms, microsensillum; Ti., tibiotarsus; Th., thoracic tergum.
2.2. Nomenclature of chaetae
The dorsal chaetotaxy of the head follows the two systems of Gisin (1967) and Soto-Adames (2010), with Gisin's original system retained for consistency with older taxonomic literature on the genus. Notation of labial palps is following Fjellberg (1999), labial chaetotaxy following Gisin (1964), and postlabial chaetotaxy following Chen and Christiansen (1993) and Cipola et al. (2020). Dorsal chaetotaxy of thoracic and abdominal segments follows both Gisin's (1967) and Szeptycki's (1979) systems, and the notation of sensilla (specialized S–chaetae) on terga follows Zhang and Deharveng (2015).
2.3. Pseudosinella material for the study
The specimens of Pseudosinella used in this study were collected from 16 caves in the Western Carpathians, Slovakia (Table 1). We used two main methods to collect specimens: (1) visual searching and hand collection from cave walls, sediment, rotten wood, and the surface of standing water (small sinter ponds and puddles in mud), and (2) pitfall trapping with 4% formaldehyde solution or 95.6% ethanol as a fixative liquid. Traps were exposed to cave sediment for no longer than 5 months to minimize their negative impact on local arthropod communities.
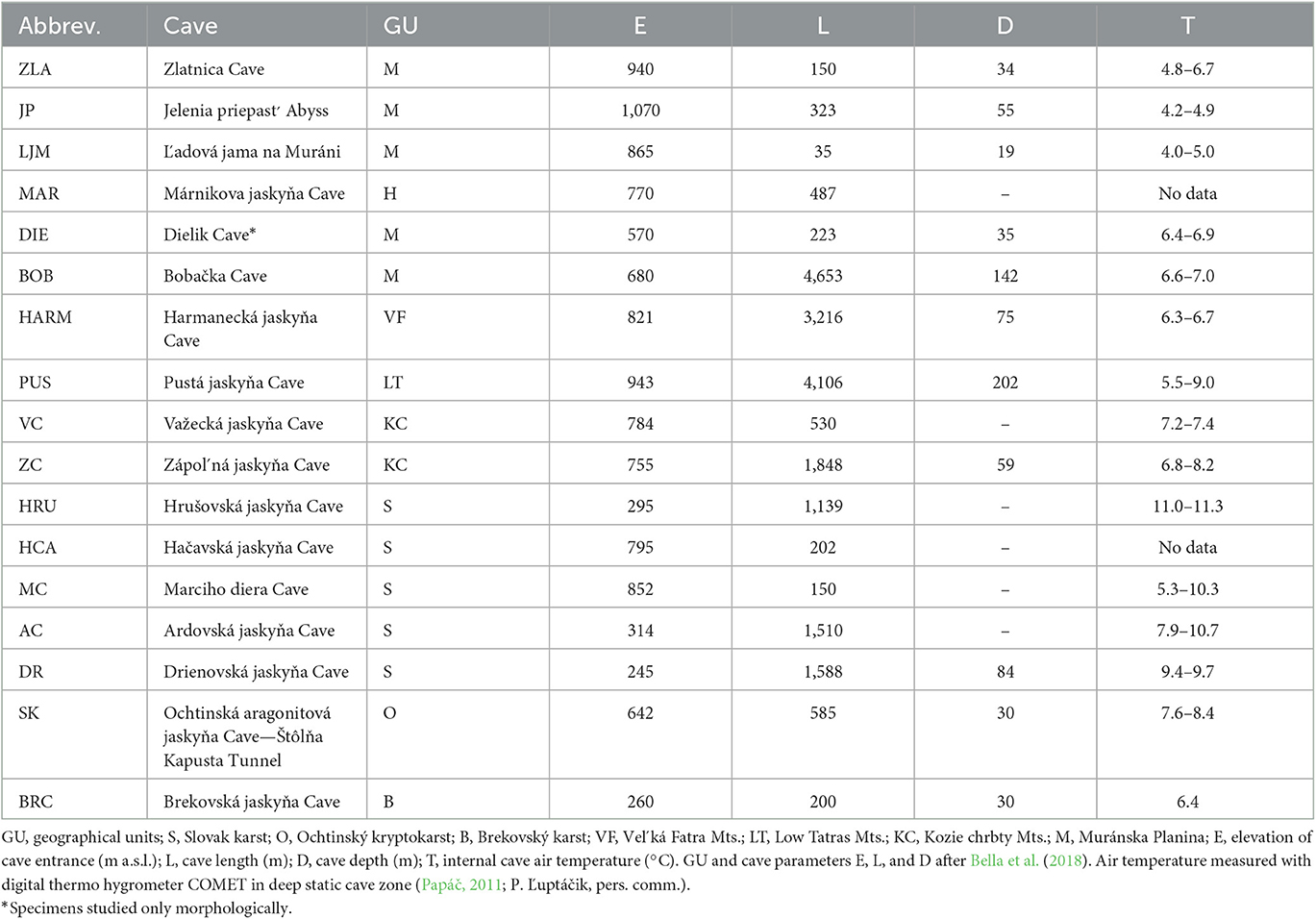
Table 1. List of caves with Pseudosinella populations used for morphological and molecular studies.
2.4. Molecular data analysis and species delimitation methods
A total of 87 Pseudosinella specimens from 16 caves in the Western Carpathians (see Supplementary Table S1) were analyzed in the molecular laboratories of the Institute of Entomology, Biology Center AS CR, Ceské Budějovice, Czech Republic, and the Department of Zoology, IBE FS UPJS, Košice, Slovakia.
To avoid contamination, all DNA laboratory work was performed under sterile conditions and using barrier tips. Total DNA was extracted using the Qiagen DNeasy Blood and Tissue Kit according to the manufacturer's modified protocol (see Parimuchová et al., 2017). A polymerase chain reaction (PCR) (Saiki et al., 1988) was performed with a reaction volume of 12.5 μl consisting of 1 μl template DNA (not quantified), 10 × PCR buffer (TopBio), 12.5 mM dNTP mix, 5 μM of each primer, and 0.125 units of Taq polymerase (TopBio) on a GenePro (Bioer Co., Ltd, China) thermal cycler. A fragment of the COI gene (660 bp) was amplified with the universal primers LCO1490 (5′-ggt caacaaatcataaagatattg g-3′) and HCO2198 (5′-taa act gggtgaccaaaaaat ca-3′; Folmer et al., 1994). Thermal cycling conditions were as follows: 94°C for 1 min followed by 37 cycles (94°C for 20 s, 47°C for 40 s, and 70°C for 50 s), followed by 1 min 30 s at 72°C. After verification on agarose electrophoresis, the reaction products were purified using Exo I/FastAP (Thermo Fisher Scientific). Sequencing of the purified products was performed using LCO1490 according to the Sanger method (SEQme s.r.o. in Dobris, Czech Republic). In cases where the primer did not produce a high-quality chromatogram, sequencing was performed with the reverse primer. Sequences were edited with Geneious Prime 2022.1.1 (Copyright © 2005–2022 Biomatters Ltd.) to remove unreadable short segments (~30 bp at the 5′ and 3′ ends). Because none of the sequences contained stop codons or indels in ORF, they were assumed to be true mitochondrial copies and not nuclear copies. All sequences were checked for their similarity to relatives of the family Entomobryidae using the GenBank BLASTn search (Mega Blast algorithm with the default setting). Sequences were aligned using Geneious Prime 2022.1.1 software (Copyright © 2005–2022 Biomatters Ltd.) according to the Muscle (Codons) algorithm using the Invertebrate Mitochondrial GeneCode and default parameters.
Species delimitation was performed using both barcoding gap and evolutionary models. A total of 87 COI sequences (length 609 bp) were used for analyses. FaBox (Villesen, 2007) was used to assemble the final set of DNA sequences into unique haplotypes. The Assemble Species by Automatic Partitioning (ASAP) method (Puillandre et al., 2021) used genetic distances to propose hypotheses about the species. The Kimura (K2P) model with default parameters was used to group sequences together.
The Poisson tree processes (PTP) model, used to delineate species based on the number of substitutions, was performed using online software (Zhang et al., 2013). Unique haplotypes were only used for this purpose. A maximum likelihood (ML) tree was derived using the auto-substitution model and a 1,000-ultrafast bootstrap analysis (Hoang et al., 2018) in the software IQ-TREE (Nguyen et al., 2015).
The estimation of divergence time was performed according to the scheme proposed by Katz et al. (2018) and Katz (2020). Input data were processed in BEAUTI v 2.6.7, and further analyses were performed in BEAST 2 (Drummond and Rambaut, 2007; Suchard and Rambaut, 2009) via the CIPRES Science Gateway (www.phylo.org; Miller et al., 2012). For molecular clock calibration, we used COI = 3.54%/Mya as recommended by Papadopoulou et al. (2010), Katz et al. (2018), and Katz (2020). bModelTest (Bouckaert and Drummond, 2017) was used to estimate the site model. A relaxed log-normal clock model was applied, with the clock rate parameter set to 0.0168 and the Yule process for speciation priorities. Analyses were run twice with an MCMC chain of 100 million generations (sampling every 10,000 generations). The ESS values (>200) and convergence were checked in Tracer 1.7.1 (Rambaut et al., 2018) with 10% burn-in. The tree with maximum clade reliability was created using TreeAnnotator v.1.4.7 (Drummond et al., 2012) and visualized in FigTree v 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/).
All new sequences are publicly available in GeneBank, and the accession number for each specimen is provided in Supplementary Table S1.
The research complied with the conditions of licenses No. 3102/2009-2.1, No. 7905/2013-2.3, and No. 2661/2017-6.3 of the Ministry of Environment of the Slovak Republic, Certificate of Competence according to Law No. 543/2002.
3. Results
3.1. Molecular species delimitation and divergence time estimation
A 609 bp fragment of COI was aligned for 87 Pseudosinella specimens from 16 caves. A total of 28 haplotypes were identified, including nine singletons (haplotypes represented by only one individual). We used two delimitation methods to define molecular operational taxonomic units (MOTUs) and combined them with morphological traits and geographic distribution to evaluate MOTUs at the species level (Figure 1). The ASAP method delimited eight MOTUs, and the best partition had an ASAP score of 1.0 (p < 0.1). The distribution of K2P distances showed a clear barcode gap, with haplotypes that diverged at a distance >9% belonging to a different species. The bPTP method estimated nine MOTUs with support ranging from 0.519 to 1.0, overestimating the ASAP delimitation (Figure 1). Pseudosinella aggtelekiensis, the species endemic to the Slovak and Aggtelek Karst (identified as AC, HCA, MC, and HRU), was well defined by ASAP, morphology, and geography, while bPTP recognized two distinct MOTUs within these populations. One of them inhabits Ardovská jaskyna Cave (AC), located at the edge of this geomorphological unit. Another known species, P. paclti, is well-defined by its morphology and distributed in three karst areas in the central Western Carpathians (VF, LT, and KC). Three different MOTUs were detected within the populations using both molecular methods. The MOTUs designated as JP, LJM, MAR, BOB, and ZLA were confirmed by both molecular methods. It is a highly troglomorphic new species Pseudosinella muranensis endemic to the caves of the Muránska planina Plateau and the neighboring Horehronské podolie Valley. The species is described below in the taxonomic part of the Results section under the scientific name P. muranensis (Kováč and Parimuchová). The delimitation analyses ASAP and bPTP resulted in three new Pseudosinella MOTUs supported by diagnostic morphological characters and geographical distribution. Thus, they represent new species, designated as Pseudosinella sp. 1, 2, and 3 (Figure 1, Table 2). Pseudosinella sp. 1 inhabits a small cryptokarst area of Devonian origin in the southern Western Carpathians, Pseudosinella sp. 2 was detected in a small and well-isolated karst area in the eastern part of the Western Carpathians. And finally, Pseudosinella sp. 3 was found in the Drienovská jaskyna Cave (DR) in the Slovak Karst, i.e., within the distribution range of P. aggtelekiensis. The geographic range of the studied Pseudosinella cave populations and their assignment to species based on integrative taxonomy is shown in Figure 2.
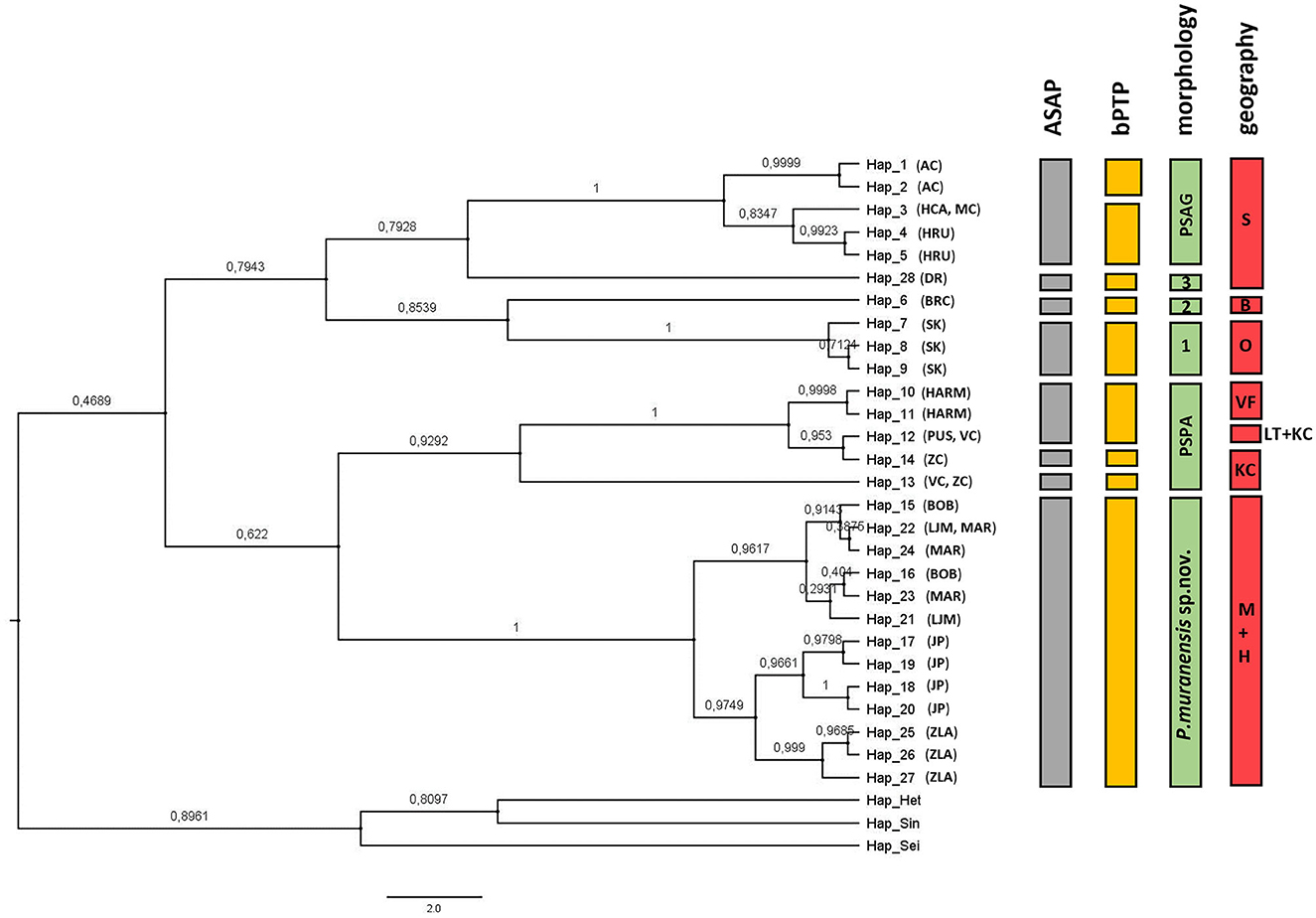
Figure 1. Species delimitation of cave Pseudosinella with ML tree based on COI marker. The values above the branches are Bayesian posterior probabilities. Heteromurus nitidus (MT611220.1), Sinella curviseta (NC 042755.1), and Seira dowlingi (MW419950.1) were used as outgroups. The graph is supplemented with a species delimitation (ASAP and bPTP) based on morphological characteristics and geographic distance. S, Slovak karst; O, Ochtinský kryptokarst; B, Brekovský karst; VF, Velká Fatra Mts.; LT, Low Tatras Mts.; KC, Kozie chrbty Mts.; M, Muránska Planina; H, Horehronské podolie. For abbreviations of caves in parentheses, see Table 1.
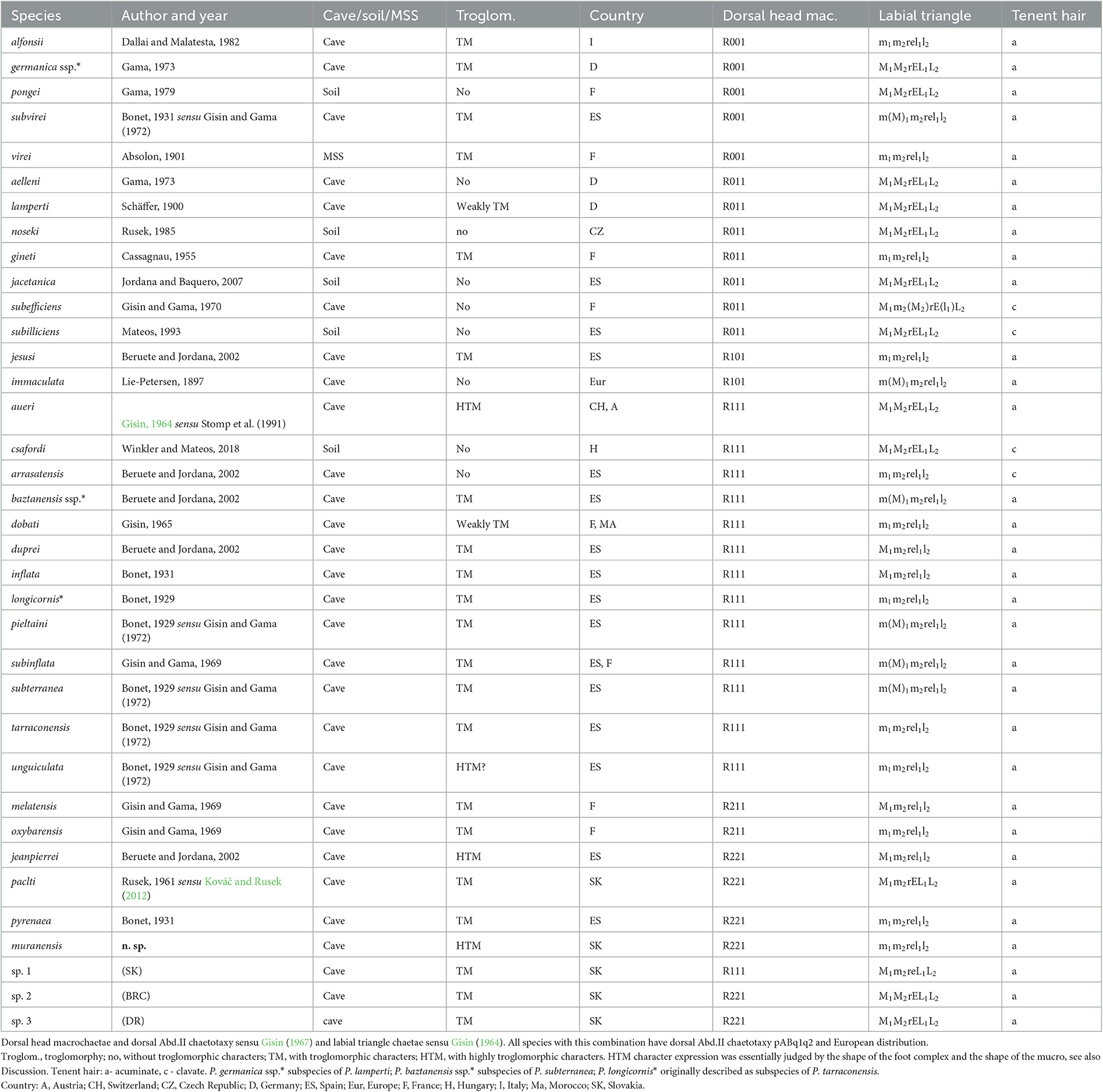
Table 2. List of eyeless Pseudosinella with 32/0201+2 medial tergal macrochaetae on Th.II–Abd.IV segments.
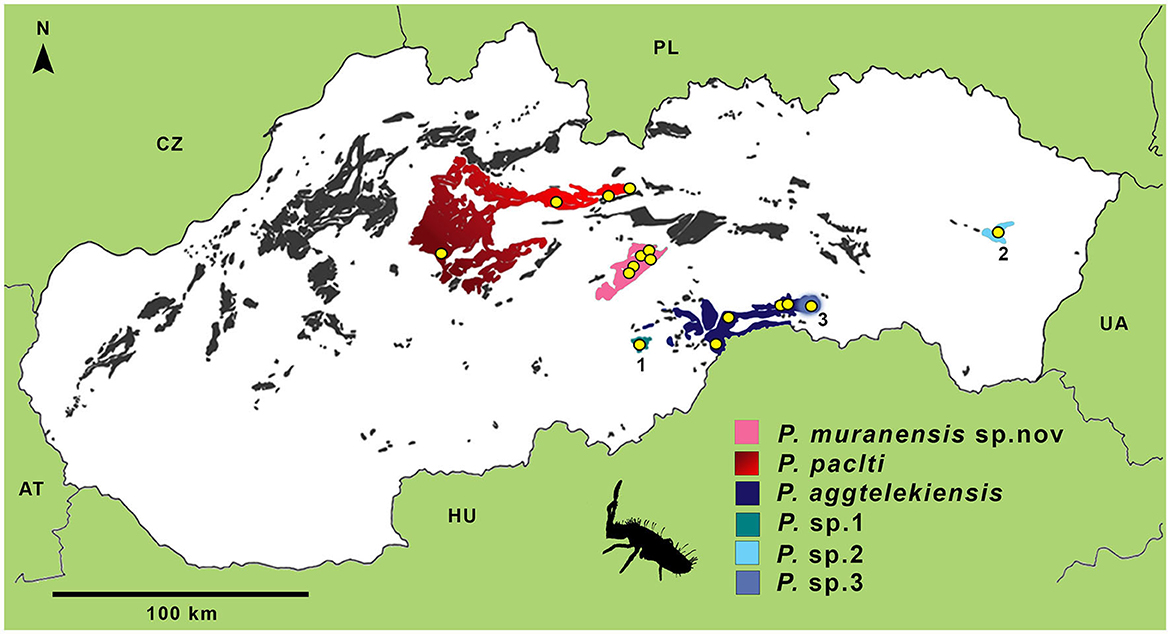
Figure 2. Distribution of Pseudosinella taxa in the caves of the Western Carpathians as a result of the integrative taxonomy study. The yellow circles stand for the sampling locations, the spots for the karst areas: black-red, Velká Fatra and Low Tatras; purple, Muránska planina Plateau; dark-blue, Slovak karst; 1, Ochtinský kryptokarst with P. sp.1; 2, Brekovský karst with P. sp. 2; 3, Drienovská Cave in the Slovak Karst with P. sp. 3. Karst areas were digitally processed by P. Gažík.
The divergence time of the Pseudosinella phylogenetic tree was estimated using representatives of the family Entomobryidae as outgroups. Appropriate sequences of COI genes from mitogenomes deposited in GenBank were used for Heteromurus nitidus (MT611220.1), Sinella curviseta (NC 042755.1), and Seira dowlingi (MW419950.1; Figure 3). The graph (Figure 3) shows that two distinct Pseudosinella lineages—“paclti” and “aggtelekiensis”—diverged from the common ancestor during the Middle Miocene ca. 14.51 Mya, followed by subsequent diversification in the P. paclti lineage 10.89 Mya and P. aggtelekiensis lineage 11.14 Mya.
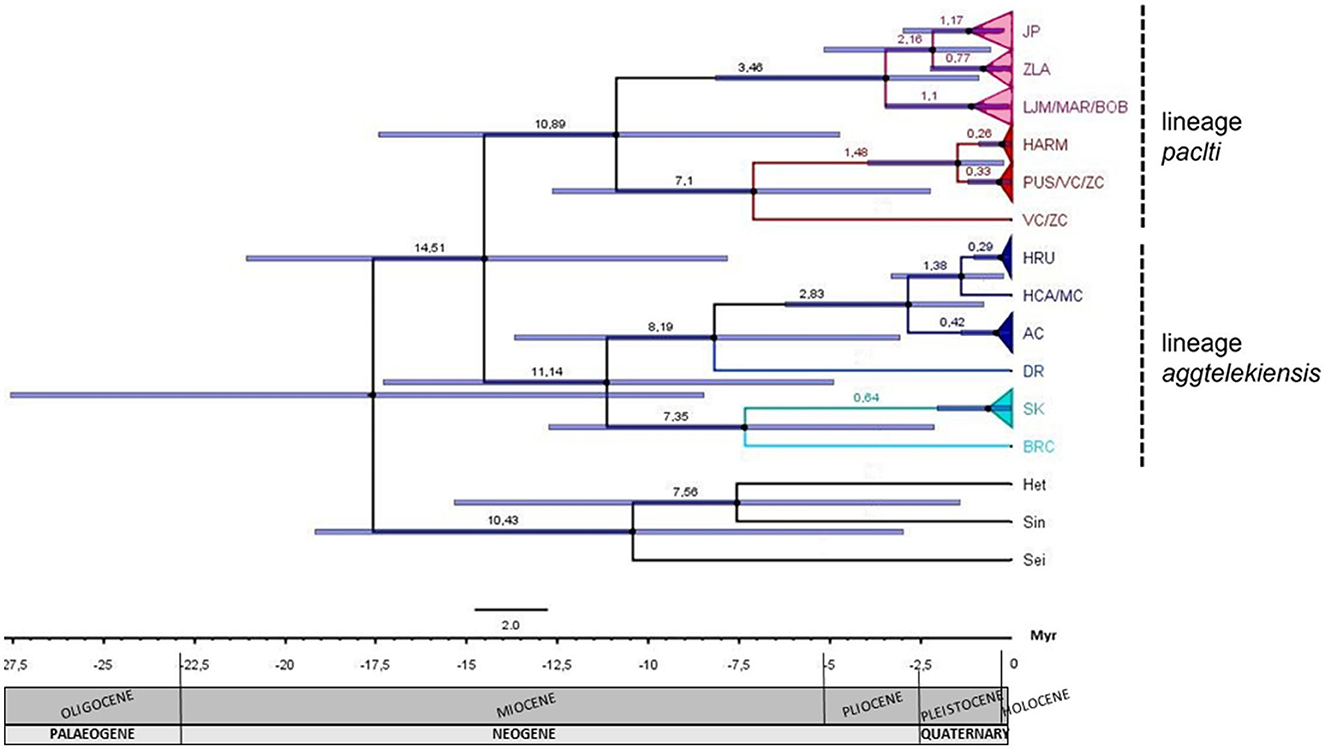
Figure 3. Bayesian tree in BEAST 2 for the Pseudosinella populations from the Western Carpathians caves based on the COI marker (visualized by FigTree). The purple boxes around each internode correspond to the 95% confidence intervals of the divergence time for each branch of the Pseudosinella phylogeny. For illustration, the geologic time scale is shown below the phylogeny. The different karst areas are represented by triangles of different colors: the Muránska planina Plateau in pink, the Low Tatras in red, the Slovak Karst in dark blue, the Ochtinský kryptokarst in green, and the Brekovský karst in turquoise. Heteromurus nitidus (Het), Sinella curviseta (Sin), and Seira dowlingi (Sei) are used as outgroups. The value 3.54%/Myr was used for the calibration of the molecular clock. For abbreviations of caves, see Table 1.
3.2. Taxonomy—Description of the new species
Class Collembola (Lubbock, 1873)
Order Entomobryomorpha (Börner, 1913)
Family Entomobryidae (Schäffer, 1896)
Subfamily Lepidocyrtinae (Wahlgren, 1906)
Genus Pseudosinella (Schäffer, 1897)
Pseudosinella muranensis (Kováč and Parimuchová), new species
Pseudosinella paclti (Rusek, 1961)—Kováč et al. (2002)
Pseudosinella cf. paclti (Rusek, 1961)—Kováč et al. (2014)
Pseudosinella sp. 3—Kováč et al. (2016)
Pseudosinella sp.—Parimuchová et al. (2020)
https://zoobank.org/NomenclaturalActs/12BE12B0-154A-4A05-B59F-9A49BB683F8C

Figure 4. Pseudosinella muranensis, general habitus of the new species. Jelenia priepast´, Muránska planina Plateau. Photo: L´. Kováč and A. Parimuchová.
3.2.1. Material
Type material. Holotype: Slovakia. ♀ on the slide (Nr. 204-00), Muránska planina Plateau, Bobačka Cave (680 m a.s.l.), Riečna chodba Passage, pitfall trap, 9 Nov. 2000, leg. L´. Kováč. Paratypes: Muránska planina Plateau, Bobačka Cave, 9 Nov. 2000, leg. L´. Kováč: eight ♀ and one ♂ on the slide (Nr. 204-00), one ♂ (Nr.: 205-00), three juveniles (Nr. 207-00), same data as holotype; one ♀ (Nr. 200-00) and one juvenile (Nr. 201-00), Koncový sifón Passage, pitfall trap, leg. L´. Kováč; one ♀ (Nr. 214-00), Vel´ryba Passage, pitfall trap, one ♀ (Nr. 200-00); six ♀ and three ♂ (Nr. 175-00), hind part, hand collecting, 5 Oct. 2000, leg. L´. Kováč and P. L´uptáčik; one ♀ and one ♂ (Nr. 422-11), Chodba tušenia Passage, 5 Sep. 2011, leg. L´. Kováč.
Other materials.
Muránska planina Plateau, Jelenia priepast´ Abyss (1,070 m a.s.l.): Biely dóm Hall, four ♀ and two specimens with undetermined sex (Nr. 561-11), central part, hand collecting on rotten wood, 2 Sep. 2011, leg. L´. Kováč; Sien pagody Hall, nine ♀ and two specimens with undetermined sex (Nr. 117-18), hand collecting on the surface of sinter pool, 18 May 2018, leg. A. Parimuchová;
Muránska planina Plateau, Zlatnica Cave (940 m a.s.l.): hind hall, 11 ♀ and two juveniles (Nr. 232-18), hand collecting on rotten wood, 14 June 2018, leg. L´. Kováč, V. Papáč, A. Mock, P. L´uptáčik, A. Parimuchová; second hall, two ♀ (Nr. 233-18), hand collecting of surface of the water pool, 14 June 2018, leg. A. Mock; entrance part, one ♀ and one ♂ (Nr.87-09), hand collecting on stony debris, 1 Oct 2009, leg. V. Papáč; Antifa Hall, four ♀ (Nr. 85-09), pitfall trap, four ♀ (Nr. 69-09), hand collecting on sediment, two ♀ and one ♂ (Nr. 70-09), hand collecting on rotten wood, Oct 2009, leg. V. Papáč; SNP Hall, eight ♀ (Nr. 23-09), hand collecting on rotten wood, 21 May 2009, leg. V. Papáč;
Muránska planina Plateau, L´adová jama na Muráni (Studna) Cave (1,165 m a.s.l.), three ♀ and one juvenile (Nr. 273-19), hand collecting on rotten wood, 3 June 2019, leg. A. Parimuchová; one ♀ (Nr. 61-08), hand collecting on rotten wood, 7 July 2008, leg. V. Papáč;
Muránska planina Plateau, Dielik Cave (570 m a.s.l.), Hall of fossils, three ♀ (Nr. 4-10), hand collecting on clay, one ♀ (Nr. 3-10), hand collecting on surface of water pool, 5 Jan. 2001, leg. V. Papáč;
Horehronské podolie, Márnikova jaskyňa Cave (770 m a.s.l.), six ♀ and 16 juveniles (Nr. 237-18), hand collecting on speleothems, 14 June 2018, leg. V. Papáč, A. Parimuchová; three ♀ (Nr. 240-18), pitfall trap, 13 March 2018, leg. J. Lakota.
All material is deposited in the IBE FS UPJS.
3.2.2. Description
Diagnosis: Eyes and pigmentation are absent. Diagnostic characters sensu Gisin (1967): (1) labium with basal chaetae as m1m2rel1l2, chaeta r minute, all labial chaetae smooth, (2) basic pattern of dorsal macrochaetae as R1R2221/32/0201+2, with number of cephalic macrochaetae M-S-Pa varying as 221 or 111, (3) chaetal pattern of Abd. II tergum pABq1q2, and (4) supplementary chaeta s anterior to trichobothrium on Abd. IV present; labral chaetae smooth; postlabial part of head and Abd. II–V terga with numerous additional ciliated mesochaetae; posterior macrochaetae of Abd. IV–V distinctly elongated; antennal segments I, II, and postlabial part of the head with conical microchaetae cm; unguis distinctly elongated, internal and lateral teeth absent, and basal teeth strongly reduced; external lamella of unguiculus is smooth; tibiotarsal tenent hair short and acuminate; trochanteral organ on leg III with 10 smooth microchaetae; and mucro with distinctly elongated apical teeth.
Habitus: General habitus as in Figure 4; Body length 2.1–2.4 mm (head 460 μm, thorax 550 μm, and abdomen 1,340 μm); White, without traces of pigmentation; Scales on antennae and legs absent, ventral side of manubrium with scales.
Head: Eyes absent; basic pattern of dorsal mac: A2, A3, M1, M2, S2, S3, and Pa5 that is R1R2221 sensu Gisin (1967), or R1R2111 (see Variability); An1–3 as mesochaetae and An4–8 as macrochaetae; Ocular area with mic p; Trichobothrium (Pa6) 50 μm long (Figure 5); Chaetotaxy of clypeus not seen well. Labrum with 4 prelabral and 5, 5, 4 labral chaetae, all smooth (Figure 6B); papillae on labral edge not differentiated; Pattern of basal chaetae on labium with m1m2rel1l2 (sensu Gisin, 1967; Figure 6A), chaeta r minute, all chaetae of labial triangle smooth; Outer maxillary palp with one basal chaeta and three smooth sublobal hairs; Labial papilla D (sensu Fjellberg, 1999) with three guard chaetae, papilla E with finger-shaped lateral process, barely reaching the top of papilla and four guard chaetae; Postlabial part of head plurichaetotic, chaetae mostly ciliated, except for four smooth chaetae in G-row and smooth chaeta J1; two ciliated chaetae in b.c. position; and three chaetae cm situated apically.
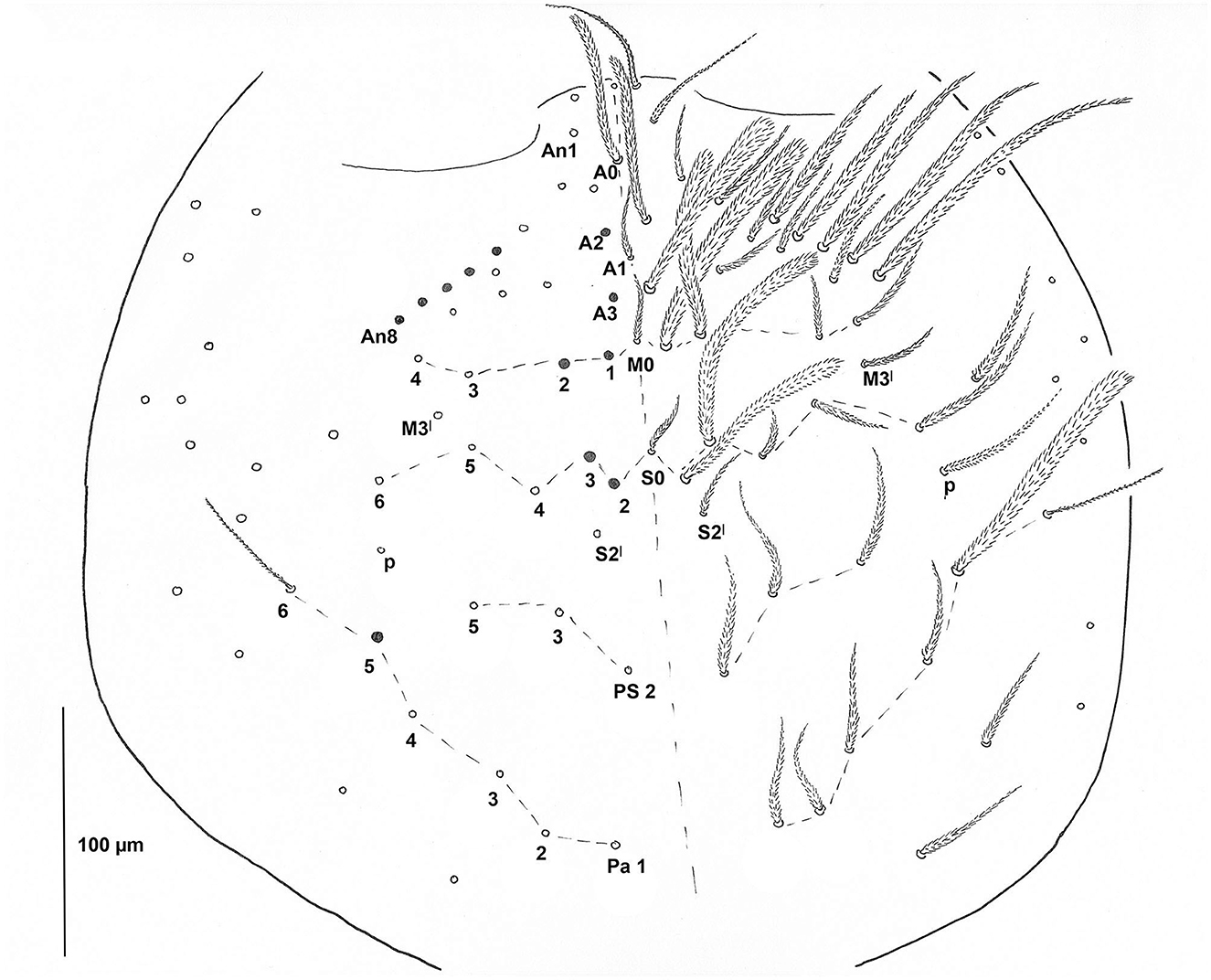
Figure 5. Pseudosinella muranensis, chaetotaxy of the head (sensu Soto-Adames, 2010). Trichobothria Pa6 drawn in full length.
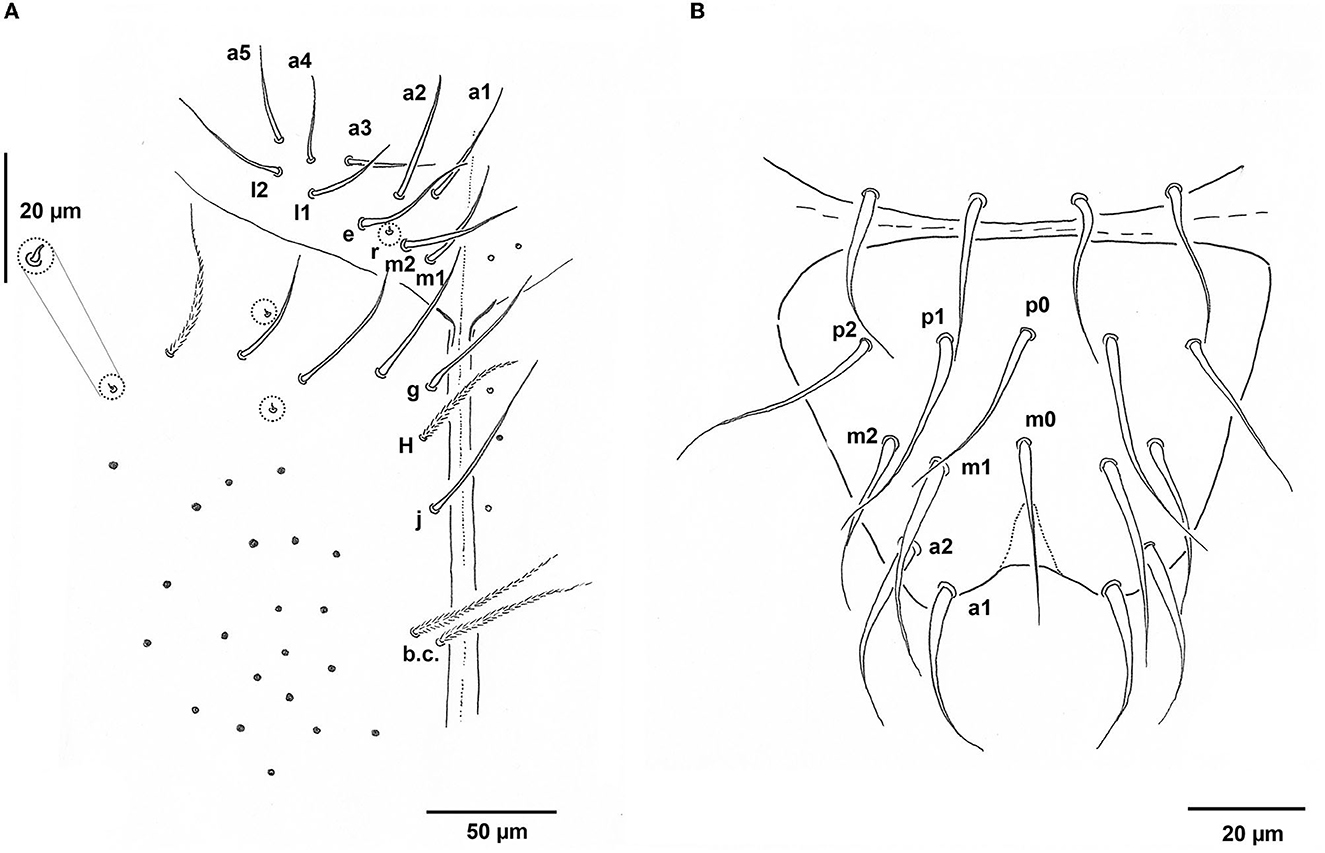
Figure 6. Pseudosinella muranensis. (A) Labium with postlabial chaetae (enlargement of microchaeta on the left); (B) Labrum.
Antennae: Length ratio of antennae and head diagonal (measured from apex of the labrum to posterior edge of head) 20.9; Relation of antennal segments I–IV as 1:2.2:2:3.5 (holotype); Ant. I with a ventral group of up to 20 slightly curved microsensilla and an external group of about 5 conical microchaetae (Figure 7C); External side of Ant.II with numerous curved sensilla, slightly curved microsensilla, and several conical microchaetae; distally with a dorso-external group of 5 leaf-shaped sensilla (Figure 7A); Antennal segments I and II with conical microchaetae cm; Ant. III organ with two thick leaf-shaped sensilla partly behind a cuticular fold, two thin guard sensilla, and 1 microsensillum (Figure 7B); Ant. III segment with numerous thin and curved sensilla, numerous straight microsensilla, and several cm on its dorsal side (Figure 7D); Ant. IV dorsally with seven types of chaetae: numerous long and thin blunt-tip sensilla, thick and band sensilla, smooth thin acuminate sensilla, six leaf-shaped sensilla at external side of segment, short spine-like chaetae, many ciliated mesochaetae, and apically one dorso-external modified chaeta (type a according to Cipola et al., 2020) apical bulb absent (Figure 7E).
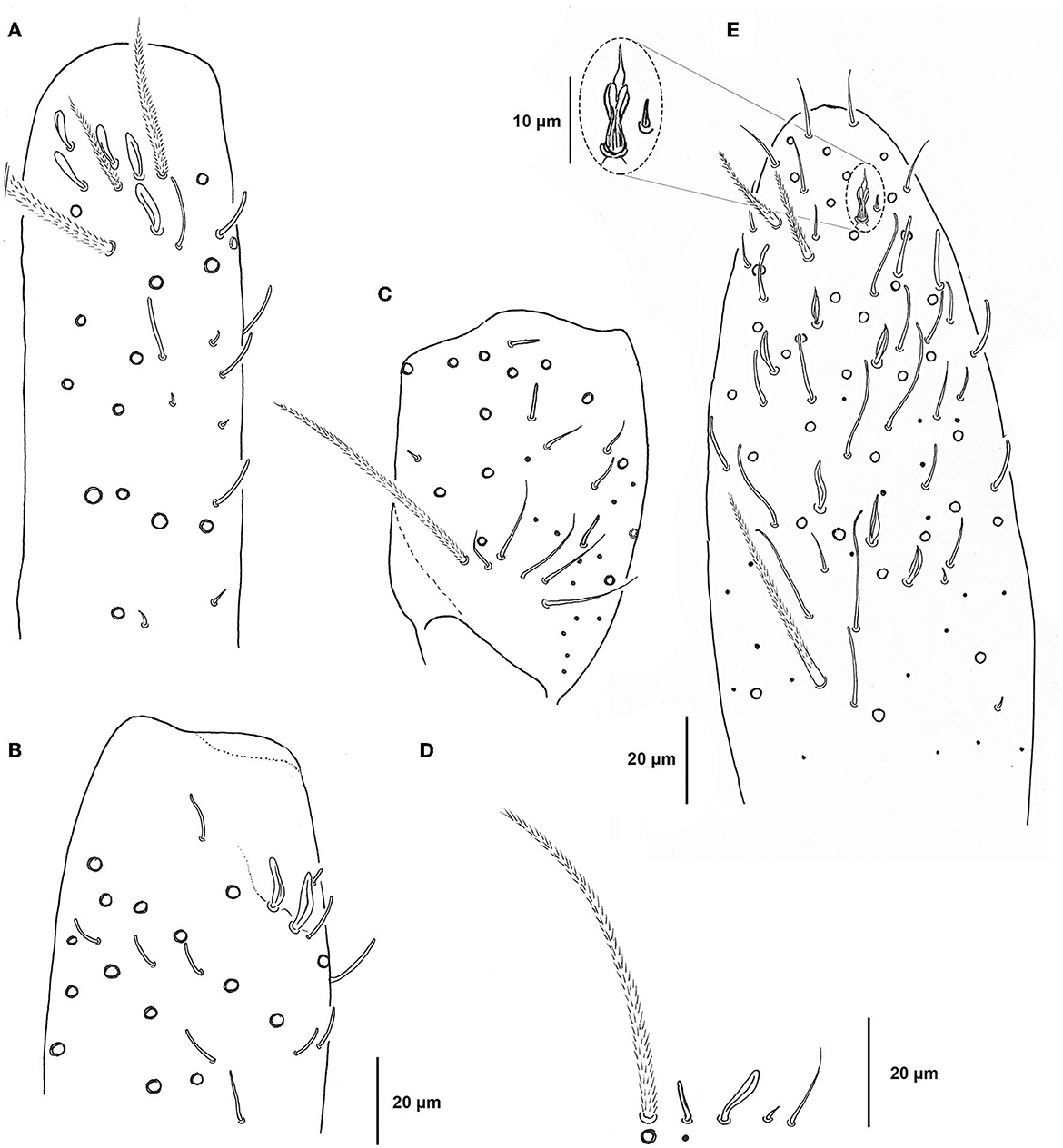
Figure 7. Pseudosinella muranensis, antennal segments I–III. (A) Apical part of right Ant.II (lateral view, all five sensilla shown); (B) Apical part of right Ant.III with AOIII (lateral view); (C) Right Ant.I (ventro-lateral view); (D) type of chaetae on Ant.I–III; (E) apical part of right Ant.IV with enlargement of subapical chaeta (empty circles—ciliated Mac, full dots—long thin sensilla).
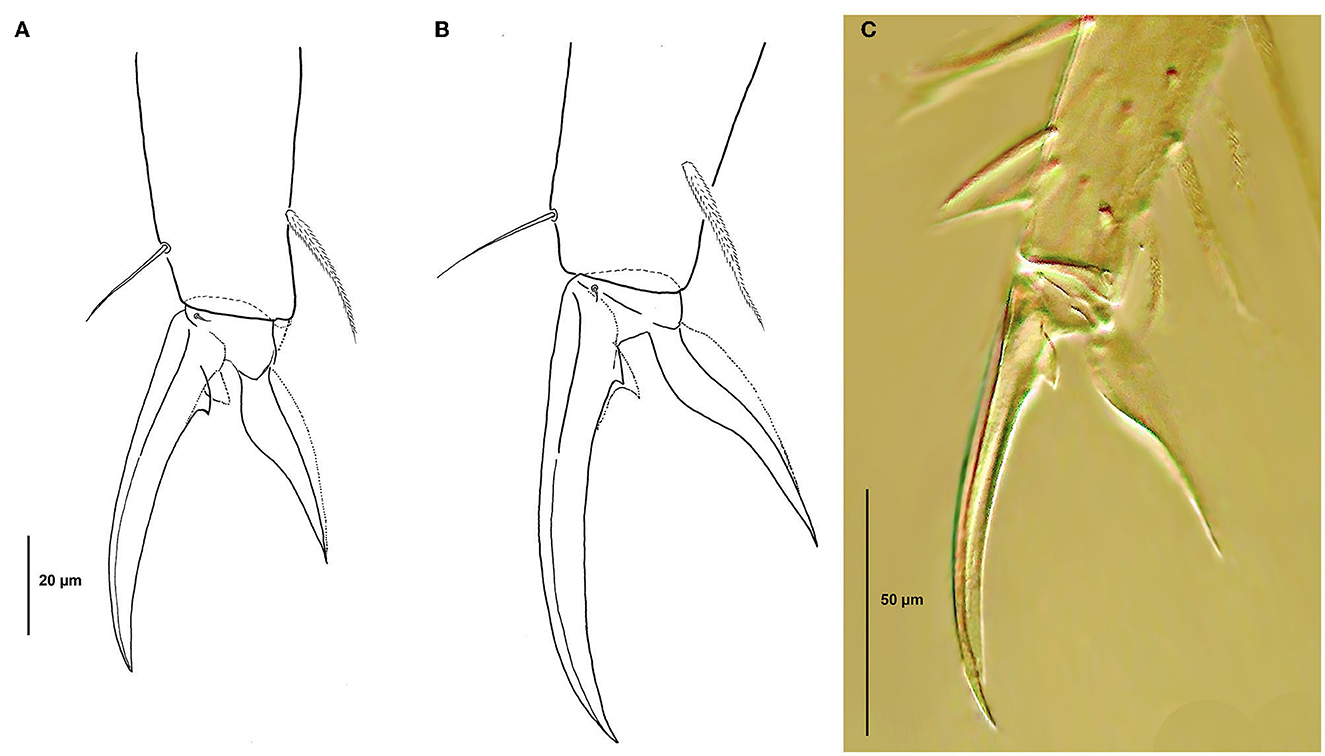
Figure 8. Pseudosinella muranensis, foot complex of leg I. (A) Shorter type from Bobačka Cave (1:2.3); (B) Longer type from Jelenia priepast´ Abyss (1:3); (C) DIC contrast image.
Thorax and abdomen: Dorsal mac formula from Th. II to Abd. IV sensu Gisin (1967) 32/0201+2; Th. II with 3 mac, 1 sensillum al, and 1 microsensillum ms in anterolateral position; Th. III with 2 mac and S-chaeta al, S-chaeta ms absent.
Abd. I with lateral S-chaeta ms; Chaetotaxy of Abd II–III as in Figure 9A; Chaetal pattern of Abd. II between 2 dorso-medial trichobothria (m2, a5) pABq1q2 sensu Gisin (1967); sensu Szeptycki (1979) as p = a2p, A = a2, B = m3, q1 = m3e, and q2 = p4; Chaeta d3 on Abd III absent; Chaetotaxy and trichobothrial complex of Abd IV in 1st instar, juvenile, and adult developmental stages as in Figures 10, 11. Mac B5, B6, C1, D3, E2, E3, E4, and F1 are thick and long with broad sockets, and B5, B6, and D3 are extremely elongated (350 μm); Numerous thinner mac with smaller sockets: A1, A1', A4, B4, C4, T5, T7, Te7, D2, De1, De3, E1, E4p2, F2, F2p, F3, F3p, Fe4, and 3 chaetae in r row; Abd IV with 4 fan-shaped supplementary chaetae anteriorly to anterior trichobothrium T2: D1, s, a, m; Numerous ciliated mesochaetae associated with longitudinal T-row of chaetae with two trichobothria (T2, T4); Two as and 1 ps S-chaeta, long dorsal S-chaetae on Abd. IV absent. Dorsal chaetotaxy of Abd V as in Figure 9B. Three S-chaetae present, as, accp4, and accp5; seven thick mac on each side, m2, m3, and m4 extremely elongated (237.5 μm), and 1 unpaired mac p0.
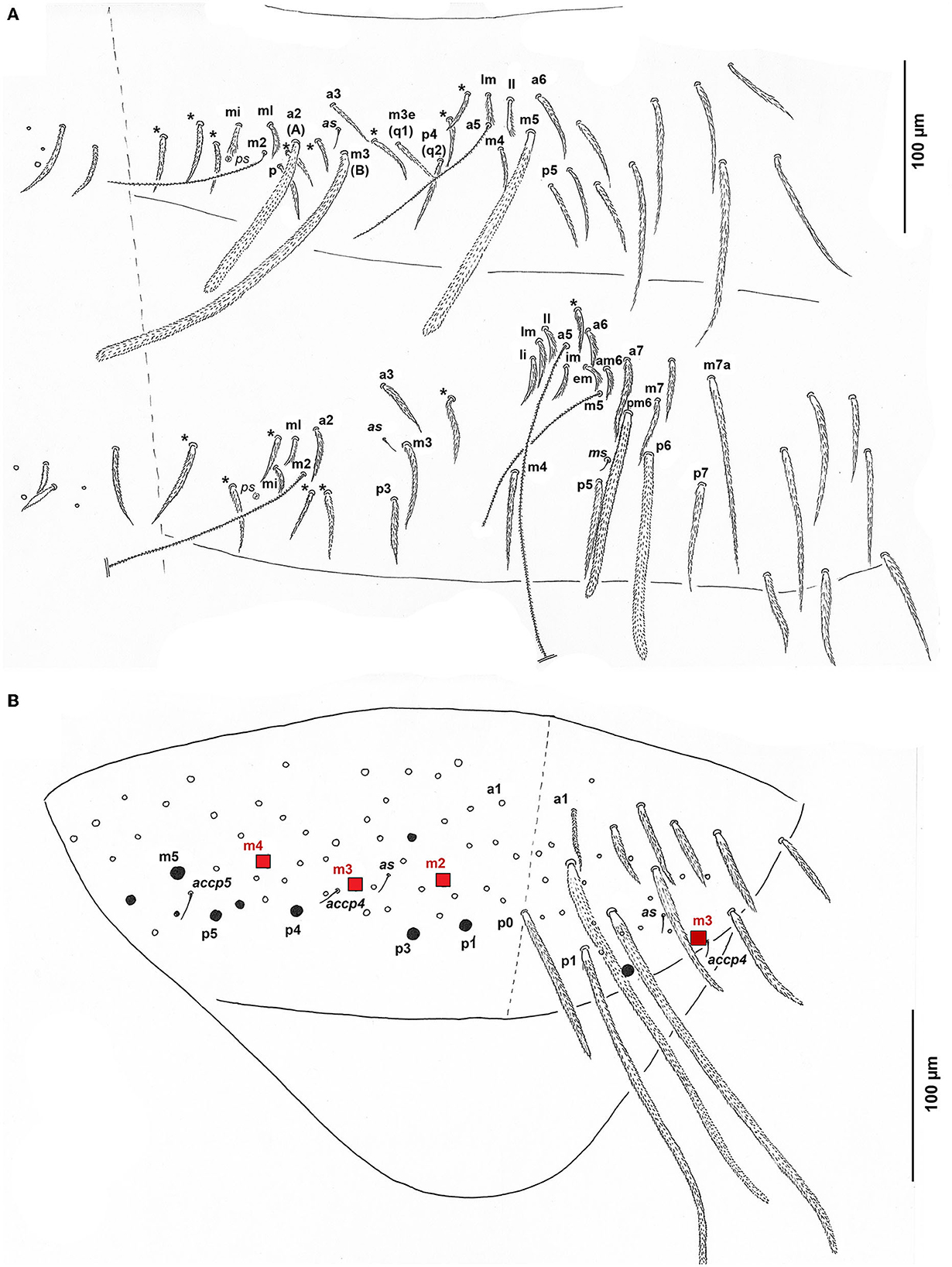
Figure 9. Pseudosinella muranensis, chaetotaxy of Abd.II–III and Abd V. (A) Abd.II–III terga sensu Soto-Adames (2010), in parentheses sensu Gisin (1967). The additional chaetae are marked with an asterisk; (B) Abd.V tergum. Red squares, long Mac (for symbols see legend in Figure 11).
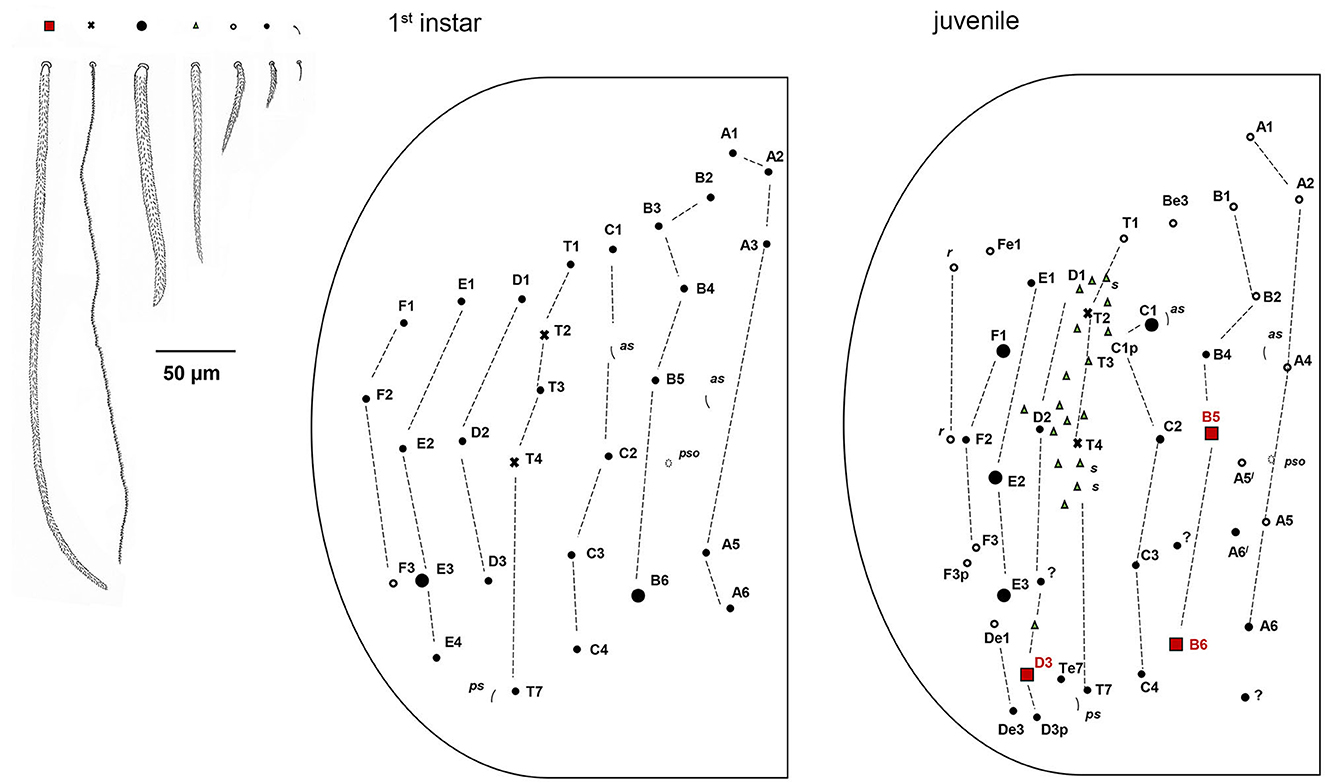
Figure 10. Pseudosinella muranensis, chaetotaxy of Abd.IV tergum in 1st instar and juvenile. Upper left—legend to the symbols used in the schemes.
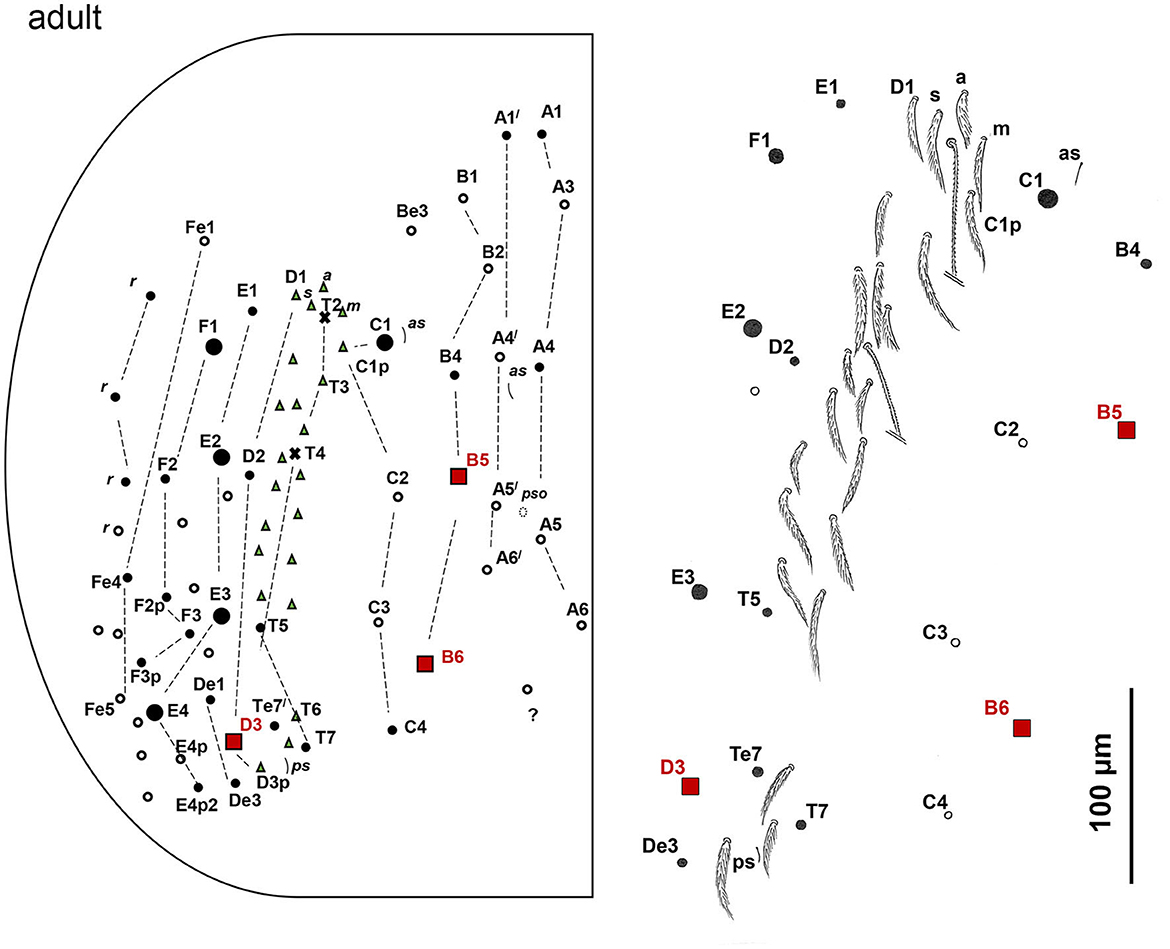
Figure 11. Pseudosinella muranensis, chaetotaxy of Abd.IV terga in adults, enlargement of the area with accessory chaetae (right).
Length of trichobothria: Head: p6–50 μm; Abd. II: m2–100 μm, a5–115 μm; Abd. III: m2–275 μm, a5–300 μm, m5–115 μm; Abd. IV: T2–350 μm, T4–125 μm.
Tubus ventralis with scales, with 5–7 subequal ciliated chaetae on the anterior side and 10–12 subequal ciliated chaetae on the posterior side; lateral side with around 10 ciliated and 2 smooth chaetae apically per half (Figure 12B).
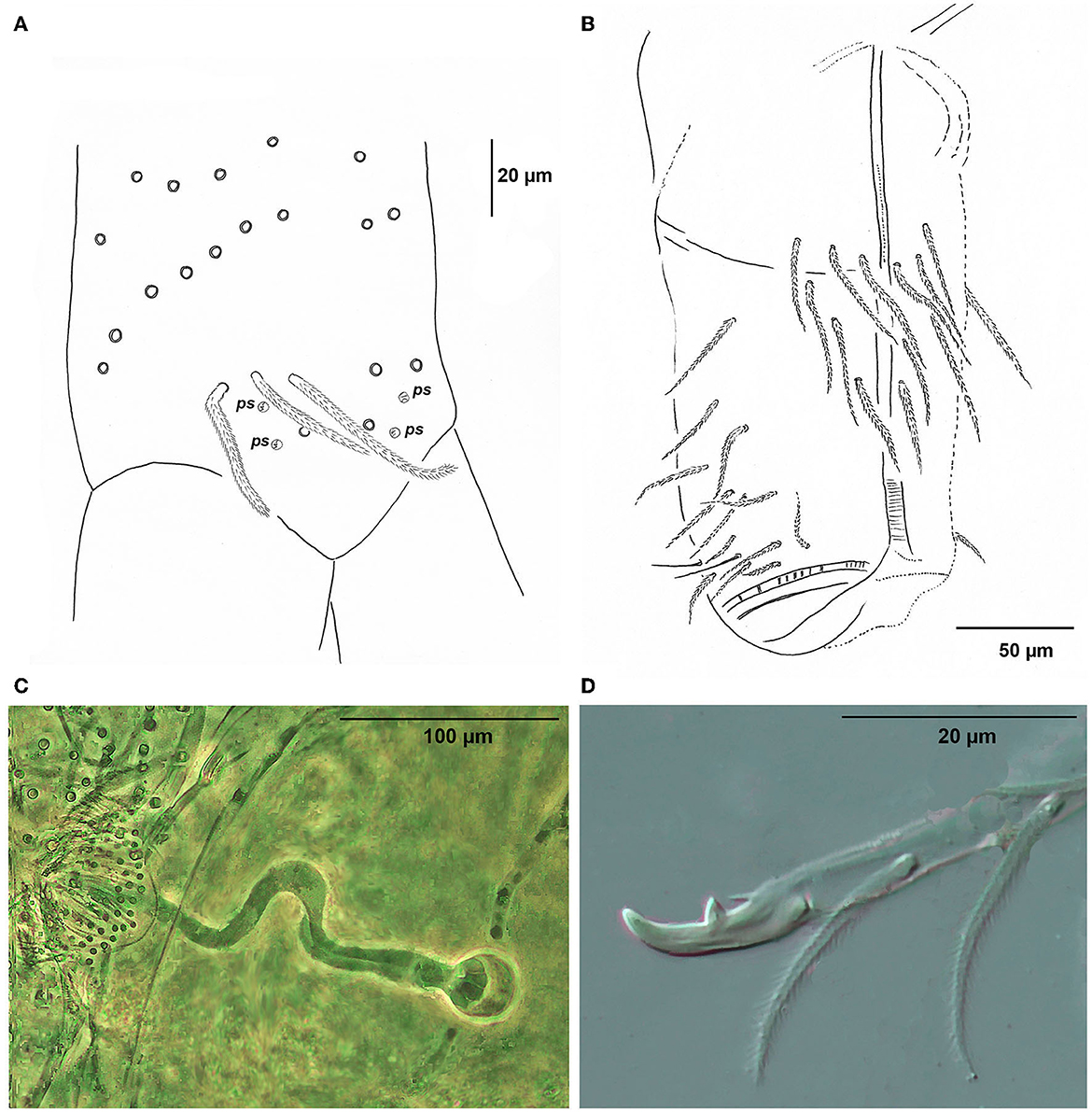
Figure 12. Pseudosinella muranensis, (A) Manubrium, dorsal chaetotaxy of apical part; ps, pseudopores; (B) Chaetotaxy of ventral tube (antero-lateral view); (C) Male genital plate with testiculus and ductus ejaculatorius (phase–contrast image); (D) Mucro and apical part of dens, lateral view (DIC contrast image).
Legs: Legs with scales; Subcoxa I with 3–4 chaetae and 2 psp; subcoxa II with a-row of 5–6 chaetae, p-row with 4 chaetae, and 2 psp; subcoxa III with a row of 6 chaetae and 2 posterior psp; Trochanteral organ on leg III with up to 10 thin and smooth spine-like chaetae distributed in V-shaped pattern; Internal edge of Ti. I–III with numerous thick acuminate ciliated chaetae approximately as long as claw, external edge with 0,1,1 long mac (more than three times the Ti. width) with a blunt apex; Tibiotarsal tenent hair short and acuminate; Supraempodial chaeta on Ti. III smooth and acuminate, ratio of supraempodial chaeta/unguiculus around 0.9; Unguis and unguiculus as in Figure 8; Unguis narrow and distinctly elongated; Internal and lateral teeth absent, basal teeth strongly reduced: external tooth strong and internal minute; Ratio pretarsus width/unguis length from 1:2.3 to 1:3.1; Unguiculus lanceolate, broad, tapering strongly toward apex; lamellae smooth and without teeth.
Furca: Manubrium ventrally with scales and 2 + 2 terminal ciliated chaetae; Dorsal manubrial plate without scales, with 2 larger inner chaetae and 2 chaetae external to 2 psp, all ciliated (Figure 12A); Length of the not ringed apical part of dens about 1.5 times the length of mucro; Mucro with distinctly elongated anterior tooth; mucronal basal spine short, not reaching the tip of anteapical tooth (Figure 12D); Ratio manubrium/dens/mucro as 7:9:1.
Both sexes are known. Male genital plate, testiculus, and ductus ejaculatorius are shown in Figure 12C.
Etymology: The species is named after the Murán region in central Slovakia, where the karst area of the Muránska planina Plateau is located.
Variability: A total of 16 specimens were examined for the pattern of the dorsal mac on the head. The pattern R1R2221 (M1, M2, S2, S3, Pa5) was observed in 13 specimens, R1R2111 (M1, S2, Pa5) in one specimen, and a left-right asymmetry 111 vs. 011 in mac numbers was observed in two specimens.
Habitat characteristics: The species inhabits a deep zone of caves of the Muránska planina Plateau with stable microclimatic conditions, characterized by absolute humidity and relatively low air temperature of 5.5–6.5°C. It was observed in numerous specimens on rotten wood, scattered bat guano, and on the water surface of small sinter pools or puddles in cave mud. It was also frequently sighted on sinter decorations with percolating water.
3.2.3. Taxonomic remarks
Pseudosinella muranensis n. sp. exhibits a combination of morphological characters that represent a high degree of adaptation to deep subterranean habitats:
(1) Relatively large body size (up to 2.4 mm),
(2) Absence of eyes and pigmentation,
(3) Smooth labral and labial chaetae,
(4) Number of medial macrochaetae on terga from head to Abd.IV essentially as R1R2221/32/0201+2 sensu Gisin (1967),
(5) Plurichaetotic postlabial part of head and Abd. II–V terga, with numerous additional ciliated mesochaetae, associated with longitudinal T-row of chaetae on Abd. IV,
(6) Extremely elongated macrochaetae on Abd. IV-V,
(7) Extremely elongated trichobothria m2 and a5 on Abd. III and T2 and T4 on Abd. IV,
(8) Ant. I–II and postlabial part of the head with conical microchaetae cm,
(9) Foot complex with distinctly elongated unguis, internal and lateral teeth absent, basal teeth much reduced, unguicular lamellae smooth,
(10) Short and acuminate tibiotarsal tenent hair,
(11) Distinctly elongated apical teeth on mucro.
The diagnostic characters of the eyeless Pseudosinella representatives are similar to P. muranensis n. sp. with the common formula of the medial macrochaetae on Th.II–Abd.IV as 32/0201+2 sensu Gisin (1967) are listed in Table 2. It is noteworthy that all these congeners share two other common features: (1) European distribution and (2) the chaetotaxy formula of Abd.II sensu to Gisin (1967) as pABq1q2. Furthermore, most of them are cave dwellers with endemic distributions restricted to caves/cave systems confined to small geographic areas. Combining R221 macrochaetae on the head and the labial triangle formula as m1m2rel1l2, the new species is most similar to Pseudosinella pyrenaea (Bonet, 1931) sensu Beruete et al. (2002). Pseudosinella pyrenaea occurs in five caves in the northern part of the Navarra region, Spain, suggesting the convergent evolution of the subterranean taxa. The new species also resembles P. paclti (Rusek, 1961), but this species differs, among other characters, in the formula of the labial triangle M1m2rEL1L2 and the shape of the foot complex. The differences in important characters between Western-Carpathian relatives are provided in Table 2. Another apparently troglomorphic species, P. aggtelekiensis, differs from these congeners mainly in the macrochaetotaxy of the Abd.IV tergum. With respect to Gisin's nomenclatural system, this species is characteristic of the macrochaetae formula R111/32/0201+3 and the labial triangle setae M1M2reL1L2. Among other features, it also has a characteristic shape of the foot complex (Kováč and Rusek, 2012).
3.2.4. Geographic distribution
Pseudosinella muranensis n. sp. is a short-range endemic species, whose distribution is restricted to the Muránska planina Plateau in the central Western Carpathians. This small karst has an area of 420 km2 and hosts ~500 caves (Bella et al., 2018).
Figure 13 shows the distribution of the highly troglomorphic Pseudosinella representatives in Europe. Their placement in this category was judged essentially by the shape of the foot complex and the shape of the mucro, and the selection follows Deharveng (1988), Deharveng and Thibaud (1989), and Christian (2002).
(1) Picos de Europa
Pseudosinella chapmani Gama, 1979; Santander, Spain;
(2) Pyrenees
Pseudosinella bessoni Deharveng, 1988; Grotte du Pic d'Orby, Atlantic Pyrenees, France;
Pseudosinella cabidochei Deharveng, 1986; Gouffre de la Pierre Saint-Martin, Atlantic Pyrenees, France;
Pseudosinella unguilonginea Jordana and Beruete, 1983; Navarra, Atlantic Pyrenees, Spain;
(3) Massif Central
Pseudosinella kersmaekersi Stomp, 1974; Grotte du Peintre, Grands Causses, Massif Central, France;
(4) Alps
Pseudosinella carthusiana Gisin, 1963; Grotte du Guiers-vif, Grande-Chartreuse, France;
Pseudosinella salisburgiana Gisin and Gama, 1969; Eisriesenwelt, Totengebirge and Schenkofen cave, Hagengebirge, Austria;
Pseudosinella aueri Gisin, 1964; Almberghöhle, Styria, Austria.
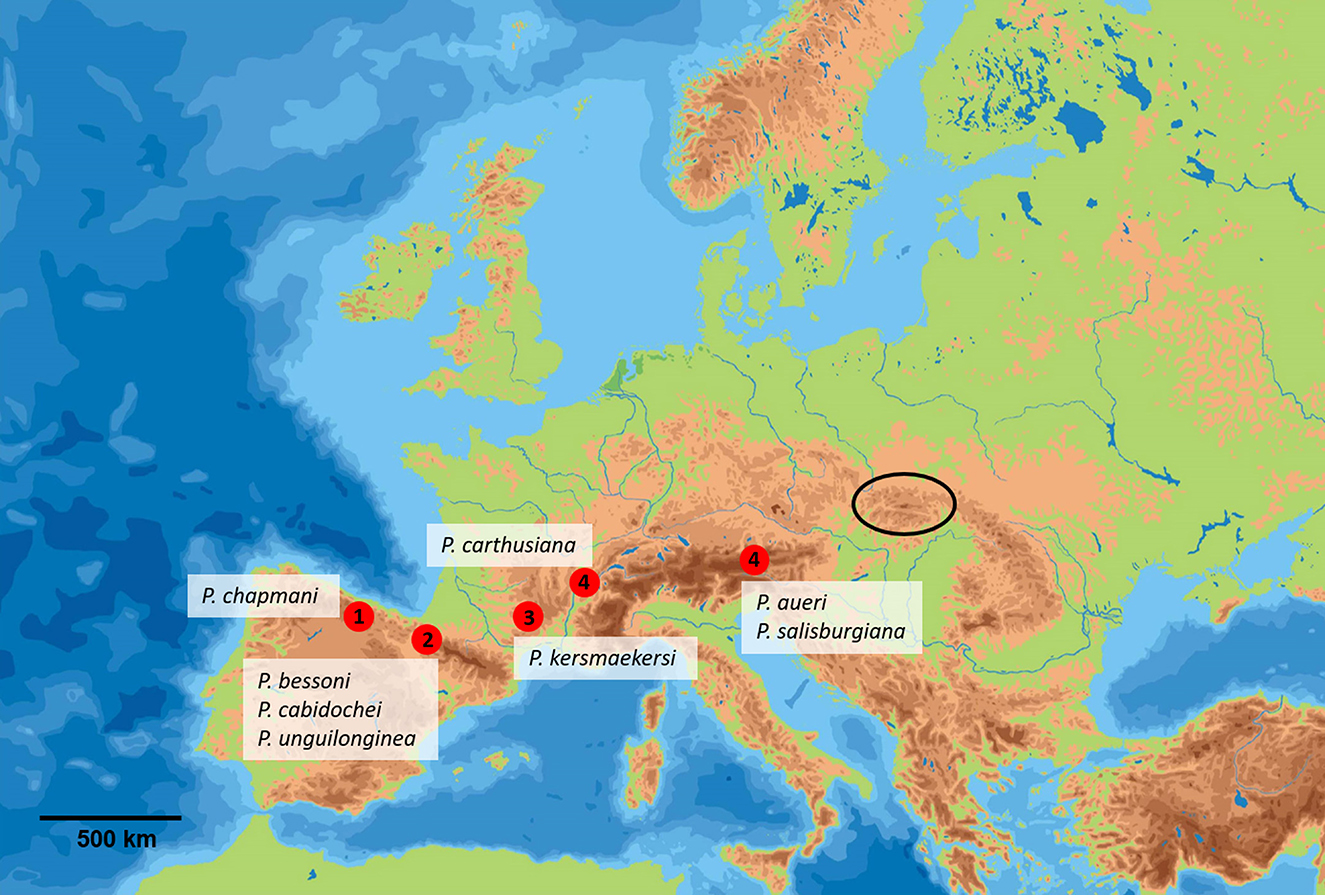
Figure 13. Distribution of the highly troglomorphic Pseudosinella representatives in Europe. Red dots represent localities, the black ellipse represents the Western Carpathians. (1) Picos de Europa; (2) Pyrenees; (3) Massif Central; (4) Alps (background map from gifex.com).
It is evident that the distribution of these forms correlates with the “mid-latitude biodiversity ridge in terrestrial cave fauna” defined by Culver et al. (2006), which includes the main mountain ridges from the Pyrenees to the Alps. The discovery of P. muranensis highlights the biogeographic importance of the Western Carpathians for the evolution of invertebrates highly adapted to a subterranean environment.
4. Discussion
4.1. Integrative taxonomy and phylogeny of the cave Pseudosinella populations
Cave-dwelling springtails provide excellent material for the study of adaptation (Christiansen and Culver, 1968). Our study of the Pseudosinella populations in the caves of the Western Carpathians, in which we took an integrative taxonomy approach, was primarily aimed at answering the question of whether this geologically complex area in Central Europe harbors ancient cave lineages, whose origin may predate the Quaternary. We also wanted to understand the importance of this area for the evolution of the highly adapted subterranean fauna.
As a first step, we examined the morphological characters of local Pseudosinella populations. Some characters are obviously unstable, such as the relative length of the antennae, and the relative length and shape of unguis that could be an expression of phenotypic plasticity. In contrast, macrochaetotaxy of the terga and labial chaetotaxy are considered complex, stable characters traditionally used for species diagnosis in Pseudosinella (Gisin, 1964, 1967; Soto-Adames, 2010). Recently, Zhang and Deharveng (2015) drew attention to the chaetotaxy of tergal sensilla (pattern of S-chaetae) as a powerful tool for revealing the phylogeny and systematics of the family Entomobryidae, which also seems to be very useful for the taxonomy of Pseudosinella. In the description of P. muranensis, we adopted the approach of Szeptycki (1979) for a detailed study of postembryonic development of tergal chaetotaxy in abdominal segments IV and V from the first instar to the adult stage. This approach helped us characterize the chaetotaxy complexity and assess the degree of adaptation to the cave environment. In this highly troglomorphic species, we observed the multiplication of ordinary ciliated chaetae on the last abdominal terga instead of the simplification of chaetotaxy seen in edaphic Pseudosinella counterparts such as Pseudosinella alba (Packard, 1873) (Szeptycki, 1979). The enormous elongation of anterior trichobothria on abdominal segments III–IV and posterior macrochaetae on abdominal segments IV–V is very characteristic of this newly described species.
In the molecular section, we identified eight to nine Pseudosinella MOTUs using barcoding-gap (ASAP) or evolutionary (bPTP) delimitation analyses, corresponding to observed differences in morphological characters among cave populations and their degree of geographic isolation. Populations of P. aggtelekiensis geographically confined to the Slovak-Aggtelek karst unit were also supported by ASAP and morphology. However, the bPTP method suggested two distinct MOTUs within this clade, one of which specifically inhabits Ardovská Cave (AC). This is likely a result of the isolation of the local population from the other four populations due to distance, specifically the 18.4 km (11.4 miles) to the nearest Hrušovská Cave (HRU) population in the same clade. To get a better idea, the five P. aggtelekiensis populations were located 7.7–37.7 km (4.8–23.6 miles) from the type locality, the Domica-Baradla Cave System. Based on morphology, the populations from the central Western Carpathians (Velká Fatra, Low Tatras, and Kozie chrbty Mts.) belonged to P. paclti, originally described from the Demänová Cave System in the Low Tatras. The four P. palcti populations were located 1.7–45.2 km (1.1–28.3 miles) from the type locality. However, both molecular delimitation methods signaled the presence of two other MOTUs in two caves of the Kozie chrbty Mts. At the moment, we are not able to distinguish these taxa morphologically, nor can we explain this sympatric pattern of MOTUs. The populations of P. muranensis, morphologically strongly adapted to caves, described in this work formed a compact clade, well-supported by molecular and morphological criteria, as well as by the geographic location of the caves in the Muránska planina Plateau in the central Western Carpathians. We also found three undescribed Pseudosinella species. The first is restricted to the Ochtinská aragonite Cave (SK) in a small area of isolated cryptokarst of Devonian origin in southern Slovakia. The populations of the second species, on the other hand, are endemic to the Brekovský kras Karst, a small and well-isolated karst area in eastern Slovakia. The third species was found in the Drienovská jaskyňa Cave (DR) in the eastern part of the Slovak-Aggtelek Karst, but not in sympatry with P. aggtelekienesis, which is characteristic of caves in this karst unit. All three species are well supported by both molecular and morphological delimitation criteria. The subterranean Pseudosinella from the Western-Carpathian caves were allocated into two main lineages: (1) the “paclti” lineage, restricted to the central Western Carpathian, and (2) the “aggtelekiensis” lineage, confined to the southern karst areas of the Western Carpathians. Thus, our study supports the main conclusion of Soto-Adames (2002) that several morphological characters traditionally used for species diagnosis in the genera Pseudosinella and Lepidocyrtus are very conservative indicators of genetic divergence and have a strong phylogenetic signal.
4.2. Troglomorphy in the Western-Carpathian Pseudosinella
Troglomorphisms represent a set of characteristic morphological modifications that are among the most peculiar adaptations of animals to a deep subterranean environment and occur in many phylogenetically unrelated groups of organisms (Christiansen, 1961, 1992; Deharveng and Bedos, 2018; Ribera et al., 2018). Therefore, morphological convergences may have evolved independently in different Pseudosinella lineages (Christiansen, 1992).
In terrestrial cave invertebrates, these modifications are divided into (1) regressive traits, such as reduction of eyes and body pigment, and (2) constructive traits, such as enlargement of the body, elongation of appendages, and claws on the legs. Collembola exhibits a great diversity of regressive troglomorphic traits, which results in high taxonomic diversity of troglobiotic forms in this group (Deharveng and Bedos, 2018). In addition, Collembola shows a number of specific constructive troglomorphic modifications such as hypertrophy and multiplication of antennal sensilla, and enlargement of paired sensilla in the Ant. III organ, elongation of unguis, reduction and basal shift of the two internal unguicular teeth, regression of the tenent hairs on the tibiotarsi, and elongation of the distal mucronal tooth (Christiansen and Culver, 1987; Deharveng, 1988; Christiansen, 2012).
It is important to note that each troglomorphic trait evolves independently within the cave lineages (Deharveng and Bedos, 2018). For example, anophthalmy without elongated appendages is common in the cave and edaphic species, whereas some cave species are oculate and weakly pigmented but have elongated appendages, e.g., Pseudosinella theodoridesi (Gisin and Gama, 1969), which inhabits caves in Haute-Garonne and Ariége in France. As Deharveng and Bedos (2018) pointed out, extreme appendage elongation is not necessarily accompanied by extreme elongation of unguis, and these differences in evolutionary changes result in complex patterns of troglomorphy. Indeed, patterns of troglomorphy are determined by the characteristics of the microhabitat that species inhabit (Deharveng, 1988; Lukić et al., 2018). The elongation of the unguis is clearly an advantage for locomotion over moist substrates or open water; this trait is also typical of aboveground Collembola inhabiting aquatic biotopes (Christiansen, 1961). However, the shape of the foot complex can vary within populations of the same Pseudosinella species (Soto-Adames, 2010). Christiansen and Culver (1987) defined three stages of troglomorphy seen in cave Entomobryinae. The newly described P. muranensis apparently shows a high level of troglomorphy with distinct morphological adaptations to a cave environment.
4.3. Distribution pattern of cave Pseudosinella
The distribution pattern of cave Pseudosinella in the Western Carpathians follows one of the basic scenarios whereby aboveground species that colonize subterranean habitats can undergo adaptive radiation and become endemic to a cave system (Christiansen and Culver, 1968). Christiansen and Culver (1987) demonstrated a clear biogeographic pattern of decreasing geographic range with increasing troglomorphy in Pseudosinella species in caves of eastern North America, which can be explained by the assumption that increasing troglomorphy decreases dispersal ability. Apparently, parallel speciation occurred in the Western Carpathians, with morphologically similar species evolving independently in different caves, in response to a common selective pressure according to the evolutionary scenario described by Christiansen (1961, 1992). Patterns of genetic differentiation in troglobionts are likely driven primarily by isolation due to physical barriers and reflect vicariant speciation (Katz et al., 2018). This is because obligate cave invertebrates have very limited dispersal abilities and thus have limited ability to migrate between caves; as a result, they may display extreme genetic divergence even across small areas (e.g., Balogh et al., 2020). We observed sympatry of two troglobiotic Pseudosinella in only two caves in the Kozie Chrbty Mts., which are among the northernmost sites with troglomorphic invertebrates in Europe (Kováč et al., 2014). This essentially allopatric distribution pattern of Pseudosinella in the Western Carpathians suggests that isolation was the main driving force for the evolution of local lineages. Such a distribution indicates the absence of recent and historical gene flow and supports the idea of speciation by vicariance at the time of habitat colonization (Guzik et al., 2021). On the other hand, the co-ocurrence of multiple troglobiotic species as a result of subterranean adaptive radiation would support the “cave as island” hypothesis (Culver and Pipan, 2009).
In our study, a highly troglomorphic species P. muranensis belonging to the “paclti” phyletic lineage was discovered. It inhabits caves in the Muránska planina Plateau, a small karst area in the central Western Carpathians. Pseudosinella aggtelekiensis (Stach, 1929), a species with advanced troglomorphy, is also a short-range endemic, confined to most caves of the Slovak and Aggtelek Karst in the southern part of the Western Carpathians. In contrast, P. paclti and three undescribed species of the P. aggtelekiensis–group (sp. 1, 2, and 3) occur in more or less “peripheral” karst areas. Thus, this pattern is basically consistent with the concept of adaptive novelty, according to which more troglomorphic Pseudosinella forms in caves in the eastern USA tend to be distributed in the center of the area and less troglomorphic ones in the periphery (Christiansen and Culver, 1968, 1987). This pattern is also confirmed by the distribution of the highly troglomorphic collembolan Deuteraphorura muranensis (Parimuchová and Kováč, 2020) (fam. Onychiuridae), which is also endemic to the small karst area of the Muránska planina Plateau in the central part of the Western Carpathians Mts. It has the most pronounced troglomorphic traits of all known subterranean Deuteraphorura congeners (Parimuchová et al., 2020). The local caves studied are situated at elevations above 700 m and have relatively low internal air temperatures (Table 1), conditions comparable to other mountain caves in the surrounding regions. The long-term historical isolation of the local karst may be a clue to this unusual distribution pattern of highly troglomorphic taxa, despite its partial connection with the neighboring karst areas of Tisovský kras in the southwest and Slovak Paradise in the northeast, both of which are composed of the same Triassic limestone formations.
The distribution of three undescribed Pseudosinella species in southern karst areas contradicts the observation of Christman et al. (2005) that single-cave endemism is not high in isolated cave areas on the periphery. To obtain an overall picture, it is important to note that troglomorphic Pseudosinella representatives do not occur in the karst of the Tatras Mts. that form the northern ridges of the Western Carpathians (Kováč et al., 2016). The general pattern of a few terrestrial troglobionts in the northern karst areas of Western Carpathians is usually explained by the proximity to continental glaciers during the Pleistocene glacial events.
The distribution of troglomorphic Pseudosinella in Europe, their low diversity, and the absence of highly troglomorphic forms in the rest of the Carpathians (Ukraine, Romania) suggest a possible colonization of the Western Carpathians by an ancestral lineage from the Eastern Alps. However, a thorough integrative study of Pseudosinella populations from these areas is needed to clarify this assumption. The diversity of cave representatives of this genus in the Balkan Peninsula is still unclear, and so far no highly troglomorphic Pseudosinella species is known from the Dinaric caves (Lukić and Deharveng, 2008).
4.4. Divergence time of the subterranean Pseudosinella lineages
In general, many subterranean taxa apparently represent old phylogenetic lineages that have persisted for long periods in subterranean refugia (Gibert and Deharveng, 2002; Holsinger, 2012). The distribution range size of a subterranean animal is a good indicator of the age of its lineage, and increasing troglomorphy suggests increasingly earlier times of initial cave colonization (Christiansen and Culver, 1987; Christian, 2002). However, as Trajano (2007) noted, there are limitations of using troglomorphy to estimate phylogenetic age because of the complexity of the mechanisms underlying morphological differentiation, i.e., a complex balance among various genetic, ecological, and biological factors that produce the actual divergence rates. The evolution rate of a troglomorphism may depend on the environment or way of life of a species and its ancestors. For example, old species may be less troglomorphic than younger ones if they have a slower rate of phenotypic evolution (Pipan and Culver, 2012; Ribera et al., 2018).
It is also considered that a putative ancestral species is no longer found near the distributional ranges of troglomorphic species (Christiansen, 2012). However, this assumption still needs to be supported by a molecular study of the cave and edaphic populations of Pseudosinella (and edaphic populations of Lepidocyrtus) in terms of their phyletic relationships to reveal the evolutionary scenario of these lineages in the Western Carpathian, which is indeed a challenging task.
Estimating the divergence time of the phyletic lineages of Pseudosinella populations was one of the main objectives of the present study. Soto-Adames (2002) applied 2.3% sequence divergence per million years as the most commonly used calibration for the COI molecular clock in the Lepidocyrtus/Pseudosinella complex. In this study, we calibrated the divergence time according to Katz (2020), who estimated relative substitution rates in the COI unit inferred by Bayesian analysis over a topologically constrained phylogeny of Hexapoda using a relaxed molecular clock model. Rates for Collembola were not significantly different from the average rate or from rates estimated for most other groups compared. This suggests that clocks calibrated using “universal” insect rates may be appropriate for estimating evolutionary time scales in this group.
The troglomorphic Pseudosinella populations of the Western Carpathians occur exclusively in karst caves developed in Triassic limestone formations of marine origin, suggesting that the geological history of the area had a crucial role in the formation of their present distribution pattern. The phylogeny study of the genus Pseudosinella (Hexapoda, Collembola) in the Western Carpathian caves revealed an initial split of an ancient lineage in the Middle Miocene into lineages “paclti” and “aggtelekiensis,” with the common ancestor colonizing the area about 14.5 Mya. This finding thus supports our assumption that the local troglomorphic Pseudosinella populations are descendants of ancient lineages originating before the Quaternary (Kováč et al., 2016). Indeed, the history of a site has a major influence on the history of a lineage, as the characteristics of past environments (e.g., geological setting, contemporary global climate, and biotic composition) have significantly influenced the survival, dispersal, and diversification of phylogenetic lineages that occurred at these sites (Lomolino et al., 2010).
Undoubtedly, the initial diversification within the collembolan genus Pseudosinella in the Miocene was associated with complex geological (tectonic) events in the Paratethys region. This is consistent with the uplift of Triassic limestones during the early formation of the Western Carpathians (Popov et al., 2004) and most likely also with the initial development of caves during this time. Continentalization of Europe during the Neogene, triggered by the Alpidic orogeny and the simultaneous retreat of the Paratethys Sea, led to isolated evolution and an increase in the endemicity of the entire local biota (Neubauer et al., 2015). The central Western Carpathians formed an island in the Paratethys during the Burdigalian and Langhian stages of the Miocene, which ranged from 20.4 to 13.6 Mya (Kováč et al., 2017). This long-term isolation undoubtedly led to the independent evolution of the local biota after previous colonization of the territory by lineages from southern European orogenic units (e.g., Kováč, et al. 2014; Haase et al., 2021). The onset of cooling of the contemporary global climate, known as the Middle Miocene Climatic Transition—MMCT (Methner et al., 2020), could promote the colonization of deep subterranean spaces by edaphic invertebrates. Similarly, the large number of Pseudosinella subterranean species recorded in the subterranean calcretes of Western Australia is also explained by more rapid climatic changes in history (Guzik et al., 2021). The overall phylogenetic history of the genus Pseudosinella is likely longer than shown in our study and extends back to the Oligocene, indicating independent diversification of certain lineages from more than one ancestor and presumed polyphyly of the entire genus, similar to that observed by Guzik et al. (2021). Indeed, the phylogeny of Pseudosinella reflects the paleogeographic context. For example, all eyeless relatives with an identical number of dorsal macrochaetae from thorax to abdomen, 32/0201+2 sensu Gisin (1967), are European representatives (Table 2).
In their synthesis of the richness of European troglobionts, Culver et al. (2006) assessed the part of Central Europe north of the Dinaric Mountains as an area with very few or no troglobionts, and the reduction in the number of these forms was largely considered to be the result of Pleistocene glaciation events (e.g., Deharveng et al., 2012). However, our study highlights the biogeographic importance of the Western Carpathians as a major speciation center of troglomorphic troglobionts, located at their northern distribution limit in Europe. Not only do highly troglomorphic taxa occur here but the diversity of troglomorphic obligate cave invertebrates includes more than 20 terrestrial species with endemic distribution (Kováč et al., 2014). The palpigrade Eukoenenia spelaea (Peyerimhoff, 1902), the pseudoscorpion Neobisium slovacum (Gulička, 1977), the rhagidiid mite Foveacheles troglodyta (Zacharda, 1988), the millipede Hylaebainosoma gulickai (Tajovský et al., 2014), and the collembolans Megalothorax tatrensis (Papáč and Kováč, 2013), D. muranensis (Parimuchová and Kováč, 2020), P. aggtelekiensis (Stach, 1929), Neelus koseli (Kováč and Papáč, 2010), and a newly described Pseudosinella species are among the local taxa that are very well adapted to the cave environment.
We are aware of certain limitations of the present study. Further studies with larger population datasets and incorporating of additional molecular markers may shed more light on the questions of subterranean biodiversity and phylogeny of the genus Pseudosinella and the complexity of troglomorphic adaptations in relation to cave microhabitat structure. Subterranean collembolans of the genus Pseudosinella are indeed suitable organisms for studies of evolutionary processes and population genetics, and the same approach can be extended to other subterranean taxa distributed in the Western Carpathians, on the northern European frontier of troglomorphy.
5. Conclusion
The present integrative taxonomic study focused on the subterranean populations of the genus Pseudosinella (Hexapoda, Collembola) inhabiting caves in the Western Carpathians in Central Europe. Eight to nine molecular operational taxonomic units (MOTUs) were identified, three of which are undescribed species.
In addition to the typical troglomorphic characters, the newly described subterranean species P. muranensis is characterized in the adult stage by a multiplication of the common chaetae on the last abdominal terga, which is probably a common feature of the highly troglomorphic conspecifics. The species is endemic to a small karst area in the central Western Carpathians documenting the short-range distribution pattern of highly evolved troglomorphisms.
Age calibration of the Pseudosinella phylogeny in the Western Carpathian caves indicates diversification of the ancient Lepidocyrtus/Pseudosinella lineage in the Middle Miocene, supporting our hypothesis that local troglomorphic Pseudosinella populations are of ancient, pre-Quaternary origin.
Moreover, our study supports the assumption that morphological characters traditionally used in species diagnosis in the genus Pseudosinella are conservative indicators of genetic divergence.
Finally, this study highlights the role of the Western Carpathians as an important speciation center of the troglomorphic subterranean fauna at the northern distributional limit in Europe.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical review and approval was not required for the study on animals in accordance with the local legislation and institutional requirements.
Author contributions
L´K and AP designed the study. L´K, AP, and VP collected the data and analyzed morphological traits. AP, NR, and NJ extracted mtDNA and prepared the molecular data. MŽ and AP analyzed the molecular data. L´K wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was funded by the Slovak Research and Development Agency (grant APVV-21-0379) and by the Slovak Scientific Grant Agency (grant VEGA 1/0438/22).
Acknowledgments
The authors thank colleagues A. Mock and P. L´uptáčik (P.J. Šafárik University, Košice, Slovakia) for their assistance during the fieldwork in the caves, especially for the help in collecting Pseudosinella specimens; P. L´uptáčik provided the authors with most data on the internal temperature of caves. Prof. Dr. R. Jordana (Universidad de Navarra, Pamplona, Spain) kindly provided the authors with two paratypes of P. unguilonginea for morphological study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1169911/full#supplementary-material
References
Balogh, A., Ngo, L., Zigler, K. S., and Dixon, G. (2020). Population genomics in two cave-obligate invertebrates confirms extremely limited dispersal between caves. Sci. Rep. 10, 17554. doi: 10.1038/s41598-020-74508-9
Bella, P., Hlaváčová, I., and Holúbek, P. (2018). Zoznam Jaskýn Slovenskej Republiky. Liptovský Mikuláš: SMOPaJ-SSS-ŠOP SR.
Bellinger, P. F., Christiansen, K. A., and Janssens, F. (1996–2022). Checklist of the Collembola of the World. Available online at: http://www.collembola.org (accessed February 08, 2023).
Beruete, E., Baquero, E., and Jordana, R. (2002). New species of Pseudosinella (Collembola: Entomobryidae) from karst caves of the Basque bio-speleologic district. Ann. Soc. Entomol. Fr. 38, 385–398. doi: 10.1080/00379271.2002.10697351
Bouckaert, R., and Drummond, A. (2017). bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 17, 42. doi: 10.1186/s12862-017-0890-6
Chen, J.-X., and Christiansen, K. A. (1993). The genus Sinella with special reference to Sinella s. s. (Collembola: Entomobryidae) of China. Orient. Insects 27, 1–54. doi: 10.1080/00305316.1993.10432236
Christian, E. (2002). Distribution patterns of cavernicolous Collembola in Austria. Proc. Xth Int. Colloquium on Apterygota, Ceské Budějovice 2000: apterygota at the beginning of the third millennium. Pedobiologia 46, 261–266. doi: 10.1016/S0031-4056(04)70141-9
Christiansen, K. A. (1961). Convergence and parallelism in cave Entomobryinae. Evolution 15, 288–301. doi: 10.2307/2406229
Christiansen, K. A. (1992). “Biological processes in space and time: cave life in the light of modern evolutionary theory,” in The Natural History of Biospeleology, ed A. Camacho (Madrid: Monografias del Museo Nacional de Ciencias Naturales), 453–478.
Christiansen, K. A. (2012). “Morphological adaptations,” in Encyclopedia of Caves, eds W. B. White, and D. C. Culver (Amsterdam: Elsevier/Academic Press), 517–528. doi: 10.1016/B978-0-12-383832-2.00075-X
Christiansen, K. A., and Culver, D. (1968). Geographical variation and evolution in Pseudosinella hirsuta. Evolution 22, 237–255. doi: 10.2307/2406522
Christiansen, K. A., and Culver, D. (1987). Biogeography and the distribution of cave Collembola. J. Biogeogr. 14, 459–477. doi: 10.2307/2844976
Christman, M. C., Culver, D. C., Madden, M. K., and White, D. (2005). Patterns of endemism of the eastern North American cave fauna. J. Biogeogr. 32, 1441–1452. doi: 10.1111/j.1365-2699.2005.01263.x
Cipola, N. G., Oliveira, J. V. L. C., Bellini, B. C., Ferreira, A. S., Lima, E. C. A., Brito, R. A., et al. (2020). Review of eyeless Pseudosinella Schäffer (Collembola, Entomobryidae, Lepidocyrtinae) from Brazilian caves. Insects 11, 194. doi: 10.3390/insects11030194
Cucini, C., Fanciulli, P. P., Frati, F., Convey, P., Nardi, F., Carapelli, A., et al. (2021). Re-evaluating the internal phylogenetic relationships of Collembola by means of mitogenome data. Genes 12, 44. doi: 10.3390/genes12010044
Culver, D. C., Deharveng, L., Bedos, A., Lewis, J. J., Madden, M., Reddell, J. R., et al. (2006). The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 29, 120–128. doi: 10.1111/j.2005.0906-7590.04435.x
Culver, D. C., and Pipan, T. (2009). The Biology of Caves and Other Subterranean Habitats. New York, NY: Oxford University Press. ISBN 978– 0–19–921992–6.
Deharveng, L. (1988). Collemboles cavernicoles VII. Pseudosinella bessoni n, sp. et note sur l'évolution morphologique de la griffe chez les Pseudosinella. Rev. Suisse Zool. 95, 203–208. doi: 10.5962/bhl.part.79648
Deharveng, L., and Bedos, A. (2012). “Diversity patterns in the tropics,” in Encyclopedia of Caves, eds W. B. White, and D. C. Culver (Amsterdam: Elsevier Academic Press), 238–250. doi: 10.1016/B978-0-12-383832-2.00032-3
Deharveng, L., and Bedos, A. (2018). “Chapter 7. Diversity of terrestrial invertebrates in subterranean habitats,” in Cave Ecology, eds O. T. Moldovan, L. Kováč, and S. Halse (Berlin: Springer), 107–172. doi: 10.1007/978-3-319-98852-8_7
Deharveng, L., Gibert, J., and Culver, D. C. (2012). “Diversity patterns in Europe,” in Encyclopedia of Caves. eds W. B. White, and D. C. Culver (Amsterdam: Elsevier Academic Press), 219–228. doi: 10.1016/B978-0-12-383832-2.00030-X
Deharveng, L., and Thibaud, J. M. (1989). Acquisitions recentes sur les insectes collemboles cavernicoles d'Europe. Mem. Biospeol. 16, 145–151.
Drummond, A. J., and Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 7, 214. doi: 10.1186/1471-2148-7-214
Drummond, A. J., Suchard, M. A., Xie, D., and Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. E29, 1969–1973. doi: 10.1093/molbev/mss075
Fiera, C., Arbea, J. I., Vargovitsh, R. S., and Barjadze, S. (2021). A synthesis on troglobitic springtails in Europe. J. Zoolog. Syst. E59, 1874–1890. doi: 10.1111/jzs.12560
Folmer, O., Black, M., Hoeh, W., Lutz, R., and Vrijenhoek, R. (1994). DNA primers for the amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Marine Biol. Biotechnol. 3, 294–299.
Gama, M. M. D. (1984). Phylogénie des espèces européennes de Pseudosinella (Collembola: Entomobryidae). Ann. Soc. R. Zool. Belg. 114, 59–70.
Gibert, J., and Deharveng, L. (2002). Subterranean ecosystems: a truncated functional biodiversity. BioScience 52, 473–481. doi: 10.1641/0006-3568(2002)0520473:SEATFB2.0.CO
Gisin, H. (1964). Collemboles d'Europe VII. Rev. Suisse Zool. 71, 649–678. doi: 10.5962/bhl.part.75615
Gisin, H. (1967). Espéces nouvelles et lignées évolutives de Pseudosinella endogés. Mem. Estud. Mus. Zool. Univ. Coimbra 301, 1–25.
Godeiro, N. N., Bellini, B. C., Ding, N., Xu, C., Ding, Y., Zhang, F., et al. (2021). A mitogenomic phylogeny of the Entomobryoidea (Collembola): a comparative perspective. Zool. Scr. 50, 658–666. doi: 10.1111/zsc.12487
Godeiro, N. N., Ding, Y., Cipola, N. G., Jantarit, S., Bellini, B. C., Zhang, F., et al. (2023). Phylogenomics and systematics of Entomobryoidea (Collembola): marker design, phylogeny and classification. Cladistics 39, 101–115. doi: 10.1111/cla.12521
Guzik, M. T., Stevens, M. I., Cooper, S. J. B., Humphreys, W. F., and Austin, A. D. (2021). Extreme genetic diversity among springtails (Collembola) in subterranean calcretes of arid Australia. Genome 64, 181–195. doi: 10.1139/gen-2019-0199
Haase, M., Grego, J., Eross, Z. P., Farkas, R., and Fehér, Z. (2021). On the origin and diversification of the stygobiotic freshwater snail genus Hauffenia (Caenogastropoda: Hydrobiidae) with special focus on the northern species and the description of two new species. Eur. J. Taxon. 775, 143–184. doi: 10.5852/ejt.2021.775.1555
Hoang, D. T., Chernomor, O., von Haeseler, A., Minh, B. Q., and Vinh, L. S. (2018). UFBoot2: improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 35, 518–522. doi: 10.1093/molbev/msx281
Holsinger, J. R. (2012). “Vicariance and dispersalist biogeography,” in Encyclopedia of Caves, eds W. B. White, and D. C. Culver (Amsterdam: Elsevier Academic Press), 849–858. doi: 10.1016/B978-0-12-383832-2.00123-7
Juberthie, C. (2000). “The diversity of the karstic and pseudokarstic hypogean habitats in the world,” in Subterranean Ecosystems. Ecosystems of the World 30, eds H. Wilkens, D. C. Culver, and W. F. Humphreys (Amsterdam: Elsevier Academic Press), 17–39.
Katz, A. D. (2020). Inferring evolutionary timescales without independent timing information: an assessment of “universal” insect rates to calibrate a Collembola (Hexapoda) molecular clock. Genes 11, 1172. doi: 10.3390/genes11101172
Katz, A. D., Taylor, S. J., and Davis, M. A. (2018). At the confluence of vicariance and dispersal: phylogeography of cavernicolous springtails (Collembola: Arrhopalitidae, Tomoceridae) codistributed across a geologically complex karst landscape in Illinois and Missouri. Ecol. Evol. 8, 10306–10325. doi: 10.1002/ece3.4507
Kováč, L´., Elhottová, D., Mock, A., Nováková, A., Krištufek, V., Chronáková, A., et al. (2014). The Cave Biota of Slovakia. Speleologia Slovaca 5. Liptovský Mikuláš: State Nature Conservancy SR, Slovak Caves Administration.
Kováč, L´., L´uptáčik, P., and Mock, A. (2002). “Článkonožce (Arthropoda) jaskyne Bobačka (Muránska planina),” in: Výskum a Ochrana Prírody Muránskej Planiny 3, ed M. Uhrin (Revúca: OP – Správa Národného parku Muránska planina), 141–145.
Kováč, L´., Parimuchová, A., and Miklisová, D. (2016). Distributional patterns of cave Collembola (Hexapoda) in association with habitat conditions, geography and subterranean refugia in the Western Carpathians. Biol. J. Linn. Soc. 119, 571–592. doi: 10.1111/bij.12555
Kováč, L´., and Rusek, J. (2012). Redescription of two troglobiotic species of the genus Pseudosinella Schäffer, 1897 (Collembola, Entomobryidae) from the Western Carpathians. Zootaxa 3341, 32–45. doi: 10.11646/zootaxa.3341.1.2
Kováč, M., Márton, E., Oszczypko, N., Vojtko, R., Hók, J., Králiková, S., et al. (2017). Neogene palaeogeography and basin evolution of the Western Carpathians, Northern Pannonian domain and adjoining areas. Glob. Planet Change 155, 133–154. doi: 10.1016/j.gloplacha.2017.07.004
Leo, C., Carapelli, A., Cicconardi, F., Frati, F., and Nardi, F. (2019). Mitochondrial genome diversity in Collembola: phylogeny, dating and gene order. Diversity 11, 169. doi: 10.3390/d11090169
Lomolino, M. V., Riddle, B. R., Whittaker, R. J., and Brown, J. H. (2010). Biogeography, 4th ed. Sunderland, MA: Sinauer Associates.
Lukić, M. (2019). “Chapter 35 – Collembola,” in Encyclopedia of Caves, eds W. B. White, D. C. Culver, and T. Pipan (Amsterdam: Elsevier), 308–319. doi: 10.1016/B978-0-12-814124-3.00034-0
Lukić, M., and Deharveng, L. (2008). “Biodiversity and distribution of cave Collembola (Hexapoda) on Biokovo Mt., Croatia,” in Advances in the Studies of the Fauna of the Balkan Peninsula - Papers Dedicated to the Memory of Guido Nonveiller, eds D. Pavičević, and M. Perreau (Belgrade: Institute of Nature Conservation of Serbia), 119–136.
Lukić, M., Delić, T., Zagmajster, M., and Deharveng, L. (2018). Setting a morphological framework for the genus Verhoeffiella (Collembola, Entomobryidae) for describing new troglomorphic species from the Dinaric karst (Western Balkans). Invertebr. Syst. 32, 1118–1170. doi: 10.1071/IS17088
Methner, K., Campani, M., Fiebig, J., Löffler, N., Kempf, O., Mulch, A., et al. (2020). Middle Miocene long-term continental temperature change in and out of pace with marine climate records. Sci. Rep. 10, 7989. doi: 10.1038/s41598-020-64743-5
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2012). “The CIPRES science gateway: enabling high-impact science for phylogenetics researchers with limited resources,” in Proceedings, 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the eXtreme to the Campus and Beyond (Chicago: ACM), 1–8. doi: 10.1145/2335755.2335836
Mráz, P., and Ronikier, M. (2016). Biogeography of the Carpathians: evolutionary and spatial facets of biodiversity. Biol. J. Linn. Soc. 119, 528–559. doi: 10.1111/bij.12918
Neubauer, T. A., Harzhauser, M., Kroh, A., Georgopoulou, E., and Mandic, O. (2015). A gastropod-based biogeographic scheme for the European Neogene freshwater systems. Earth Sci. Rev. 143, 98–116. doi: 10.1016/j.earscirev.2015.01.010
Nguyen, L.-T., Schmidt, H. A., von Haeseler, A., and Minh, B. Q. (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. E32, 268–274. doi: 10.1093/molbev/msu300
Papáč, V. (2011). Spoločenstvá chvostoskokov (Hexapoda, Collembola) v jaskyniach Muránskej planiny, Drienčanského krasu a Cerovej vrchoviny [Dissertation thesis]. Bratislava, SR: Comenius University, Bratislava.
Papadopoulou, A., Anastasiou, I., and Vogler, A. P. (2010). Revisiting the insect mitochondrial molecular clock: the Mid-Aegean trench calibration. Mol. Biol. E27, 1659–1672. doi: 10.1093/molbev/msq051
Parimuchová, A., Kováč, L´., Žurovcová, M., Miklisová, D., and Paučulová, L. (2017). A glacial relict in the Carpathian caves – population variability or a species complex? Arthr. Syst. Phyl. 75, 351–362. doi: 10.3897/asp.75.e31911
Parimuchová, A., Žurovcová, M., Papáč, V., and Kováč, L´. (2020). Subterranean Deuteraphorura Absolon, 1901, (Hexapoda, Collembola) of the Western Carpathians—troglomorphy at the northern distributional limit in Europe. PLoS ONE 15, e0226966. doi: 10.1371/journal.pone.0226966
Pipan, T., and Culver, D. C. (2012). Convergence and divergence in the subterranean realm: a reassessment. Biol. J. Linn. Soc. 107, 1–14. doi: 10.1111/j.1095-8312.2012.01964.x
Popov, S. V., Rogl, F., Rozanov, A. Y., Steininger, F. F., Shcherba, I. G., Kováč, M., et al. (2004). Lithological-palaeogeographic maps of paratethys. 10 maps late eocene to pliocene. Cour. Forsch. Inst. Senckenberg 250, 1–46.
Potapov, A., Bellini, B., Chown, S., Deharveng, L., Janssens, F., Kováč, L., et al. (2020). Towards a global synthesis of Collembola knowledge – challenges and potential solutions. Soil Org. 92, 161–188. doi: 10.25674/so92iss3pp161
Puillandre, N., Brouillet, S., and Achaz, G. (2021). ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour. 21, 609–620. doi: 10.1111/1755-0998.13281
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., and Suchard, M. A. (2018). Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
Ribera, I., Cieslak, A., Faille, A., and Fresneda, J. (2018). “Chapter 10. Historical and ecological factors determining cave diversity,” in Cave Ecology. Ecological Studies, eds O. Moldovan, L. Kováč, and S. Halse (Cham: Springer), 229–252. doi: 10.1007/978-3-319-98852-8_10
Rusek, J. (1975). Eine Präparationstechnik für Sprungschwänze und ähnliche Gliederfüsser. Mikrokosmos 12, 376–381.
Saiki, R. K., Gelfand, D. H., and Stoffel, S. (1988). Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239, 150–160. doi: 10.1126/science.2448875
Soto-Adames, F. N. (2002). Molecular phylogeny of the Puerto Rican Lepidocyrtus and Pseudosinella (Hexapoda: Collembola), a validation of Yoshii's “color pattern species”. Mol. Phylogenet. Evol. 25, 27–42. doi: 10.1016/S1055-7903(02)00250-6
Soto-Adames, F. N. (2010). Two new species and descriptive notes for five Pseudosinella species (Hexapoda: Collembola: Entomobryidae) from West Viginian (USA) caves. Zootaxa 2331, 1–34.
Suchard, M. A., and Rambaut, A. (2009). Many-core algorithms for statistical phylogenetics. Bioinformatics 25, 1370–1376. doi: 10.1093/bioinformatics/btp244
Sun, X., Yu, D., Xie, Z., Dong, J., Ding, Y., Yao, H., et al. (2020). Phylomitogenomic analyses on collembolan higher taxa with enhanced taxon sampling and discussion on method selection. PLoS ONE 15, e0230827. doi: 10.1371/journal.pone.0230827
Szeptycki, A. (1979). Chaetotaxy of the Entomobryidae and its Phylogenetical Significance. Morpho-Systematic Studies on Collembola. IV. Krakow: Polska Akademia Nauk, Instytut Zoologiczny.
Trajano, E. (2007). The challenge of rstimating the age of subterranean lineages: examples from Brazil. Acta Carsol. 36, 191–198. doi: 10.3986/ac.v36i1.221
Villesen, P. (2007). FaBox: an online toolbox for fasta sequences. Mol. Ecol. Notes 7, 965–968. doi: 10.1111/j.1471-8286.2007.01821.x
Wang, F., Chen, J. X., and Christiansen, K. (2004). A survey of the genus Pseudosinella (Collembola: Entomobryidae) from East Asia. Ann. Entomol. Soc. Am. 97, 364–385. doi: 10.1603/0013-8746(2004)0970364:ASOTGP2.0.CO
Winkler, D., Németh, M. T., and Fiera, C. (2021). A new species of Pseudosinella Schäffer, 1897 (Collembola, Entomobryidae) from Hungary and Romania, with redescription of related species Pseudosinella huetheri Stomp, 1971. ZooKeys 1063, 121–137. doi: 10.3897/zookeys.1063.73094
Zhang, F., and Deharveng, L. (2015). Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 44, 298–311. doi: 10.1111/zsc.12100
Zhang, F., Sun, D., Yu, D., and Wang, B. (2015). Molecular phylogeny supports S-chaetae as a key character better than jumping organs and body scales in classification of Entomobryoidea (Collembola). Sci. Rep. 5, 12471. doi: 10.1038/srep12471
Keywords: biospeleology, short-range endemism, cryptic diversity, divergence, molecular calibration
Citation: Kováč Ĺ, Žurovcová M, Raschmanová N, Jureková N, Papáč V and Parimuchová A (2023) Troglomorphic adaptations on the northern European frontier: the phylogeny of the cave Pseudosinella (Hexapoda, Collembola) in the Western Carpathians. Front. Ecol. Evol. 11:1169911. doi: 10.3389/fevo.2023.1169911
Received: 20 February 2023; Accepted: 31 May 2023;
Published: 22 June 2023.
Edited by:
Matthew Niemiller, University of Alabama in Huntsville, United StatesReviewed by:
Jose Aurelio Castro, University of the Balearic Islands, SpainGergely Balázs, Eötvös Loránd University, Hungary
Copyright © 2023 Kováč, Žurovcová, Raschmanová, Jureková, Papáč and Parimuchová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ĺubomír Kováč, bHVib21pci5rb3ZhY0B1cGpzLnNr