 Jun Zhao
Jun Zhao Yujing Li
Yujing Li Paul A. Selden
Paul A. Selden- 1Research Center of Paleobiology, Yuxi Normal University, Yuxi, Yunnan, China
- 2Yunnan Key Laboratory of Plateau Geographical Processes and Environmental Changes, Faculty of Geography, Yunnan Normal University, Kunming, Yunnan, China
- 3Department of Geology, University of Kansas, Lawrence, KS, United States
- 4Natural History Museum, London, United Kingdom
Sessile epibenthos were diverse and played important part in the process of energy flow in the Cambrian marine ecosystem. Based on new specimens from the Gaoloufang Section of the Wulongqing Formation, we describe two new representatives of the group that show character traits with cnidarians and bryozoans. If confirmed, the new material can help us understand the origin and early evolution of these two phyla. The discovery of more sessile epibenthos suggests that the benthic ecosystem of the Guanshan biota (Cambrian Series 2, Stage 4) is more diverse than previously thought.
Introduction
As one of the most significant Burgess Shale-type Lagerstätten, the Guanshan biota (Cambrian Series 2, Stage 4) has yielded nearly 100 species classified into more than 10 metazoan groups from over 10 fossil sites (Hu et al., 2013, 2023; Chen et al., 2019; Zhao et al., 2020). Chronologically, the Guanshan biota lies among some famous Cambrian biotas (e.g., the Chengjiang and Xiaoshiba from Stage 3 and, Kaili and the Burgess Shale from Wuliuan), thus forming an important link in the history of the early evolution of marine animals (Hu et al., 2013; Liu et al., 2016; Chen et al., 2019). In addition to the detailed systematic assessments of metazoan groups, recent studies have also been focused on the ecological complexities and sedimentology of the Guanshan biota (Ding et al., 2020; Chen et al., 2022), which further reveal a comprehensive appearance of the exceptionally preserved fossil deposit.
Characterized by a body of radial symmetry, cnidarians (anthozoans + medusozoans) are among the most basal metazoans (Hou et al., 2017; Han et al., 2020; Ou et al., 2022); polypoid cnidarian adults live anchored to the sea floor and by collecting food with circumoral tentacles (Hou et al., 2017). Han et al. (2020) discussed the evolution of early Cambrian cnidarians from the Yangtze Block, and concluded that during the “Cambrian explosion” the abundance and diversity of cnidarians peaked at the very beginning and then declined in Stages 2 and 3 of the Cambrian, with this change possibly contributing to the rise of bilateral animals. A few taxa from the Chengjiang biota (Cambrian Series 2, Stage 3) have been previously interpreted as potential cnidarians, including Xianguangia sinica Chen and Erdtmann, 1991. Recently, Xianguangia, alongside with Dinomischus venustus Chen et al., 1989, and Daihua sanqiong Zhao et al., 2019 (two epibenthic metazoans also from the Chengjiang biota), was included in the stem lineage of ctenophores (Zhao et al., 2019). However, the most recent phylogenetic analysis has proposed that Xianguangia, Dinomischus, Daihua, and Nailiana elegans Ou et al., 2022 (a polypoid form from the Chengjiang biota) were among the basal stem within Cnidaria (Ou et al., 2022). Only a tubular metazoan from the Guanshan biota was reported as a possible cnidarian (Hu et al., 2013), but its anatomy has not been documented in detail yet.
Bryozoa (also known as Ectoprocta) are a group of dominantly sessile, filter-feeding lophophorates that are abundant, diverse and widely distributed in marine, and freshwater environments (Taylor and Waeschenbach, 2015; Schwaha et al., 2020). They are colonial and characterized by iterated units (zooids) showing hierarchical levels of modularity (Lidgard et al., 2012; Zhang et al., 2021). Morphological data and molecular clock estimation strongly indicated a Cambrian origin for bryozoans (Taylor et al., 2015; Taylor and Waeschenbach, 2015; Hageman and Ernst, 2019; Ernst, 2020), and character traits expected in ancestral Cambrian representatives were also proposed (Taylor and Waeschenbach, 2015). No unequivocal fossil records of Cambrian bryozoans were accepted widely until Zhang et al. (2021) first recognized a bryozoan (Protomelission gatehousei Brock and Cooper, 1993) from Cambrian Stage 3 sections of Australia and South China, thus dating back the appearance of this phylum by approximately 35 million years. Recently, a potential bryozoan was described from the Cambrian Stage 4 Harkless Formation, USA, which could greatly push back the occurrence of mineralized skeletons in Bryozoa (Pruss et al., 2022).
Here, we describe two new epibenthos from the Guanshan biota (Cambrian Series 2, Stage 4). The first is soft-bodied and cnidarian-like; the second is preserved attached to trilobite fragments and lives in a skeletonized colony-like structure, with individuals (each with a suboval apical opening) being densely packed: some basic character traits of Bryozoa. If confirmed as representatives of Cnidaria and Bryozoa, the two new forms would provide new information about the early evolutionary history of these two phyla. Our findings will markedly increase the known taxonomic diversity and morphological disparity of the Guanshan biota. In addition, the new material also demonstrates that the epibenthic ecosystem of the Guanshan biota is rather sophisticated.
Materials and methods
One specimen of the cnidarian-like metazoan (RCP-ZJ-0002), with part and counterpart; two slabs of the bryozoan-like organism, with six colony-like assemblages (RCP-ZJ-0003-0008). All the fossils were collected from the Gaoloufang section (24.95916°N and 102.80539°E) in Kunming, Yunnan Province, southwest China. Digital photographs were taken using a Canon EOS 5D SR camera with a Canon MP-E 65 (1–5X) macro lens under cross-polarized light, with the brightness and contrast of the resulting images processed in Adobe Photoshop CS 5. The specimens are housed at the Research Center of Paleobiology, Yuxi Normal University (RCP).
The cnidarian-like metazoan (RCP-ZJ-0002)
Description
The body is 27 mm in length and has two distinct regions (upper portion and lower portion), which are delineated by a weak constriction in between (Figure 1).

Figure 1. The unnamed cnidarian-like metazoan (RCP-ZJ-0002). (A) Part; yellow arrow indicates the weak constriction between the upper portion and the lower portion of the body; (B) counterpart; yellow arrow indicates the weak constriction between the upper portion and the lower portion of the body. The scale bars are 5 mm. UP, upper portion; LP, lower portion.
The upper portion measures 12 mm long on the part of the specimen (Figure 1A) and is a cluster of tentacle-like structures that are thin, semi-transparent (Figures 2A, B), and preserved to be straight from base to top. “The supposed tentacles” are lamellar and overlapping (Figure 2B); although the exact number is hard to count due to the poor preservation, it is no fewer than eight.

Figure 2. The unnamed cnidarian-like metazoan (RCP-ZJ-0002). (A) Detail of the tentacle-like structures on the counterpart (position marked by frame b in Figure 1B); (B) detail of the tentacle-like structures [position marked by frame a in panel (A)]; yellow arrows indicate the thin and overlapped tentacle-like structures; (C) detail of the lower portion of the body on the part (position marked by frame a in Figure 1A); white and black arrows indicate a bump and a depression, respectively, yellow arrows indicate longitudinal ridges; blue arrows indicate the sheath-like structure. The scale bars are 2 mm for panels (A,C) and 1 mm for panel (B).
The lower portion (trunk) is column-like, measuring 15 mm in length, and 9 mm in maximum width (at the midheight). Bumps and depressions are evident on the surface (Figure 2C), and prominent sub-parallel longitudinal ridges are located at the lower right of the trunk (Figure 2C). The bottom is flat and measures 6 mm in width. Remains of some comparatively rigid structures are visible at the lower left side and the bottom (Figure 2C), suggesting the trunk might be enclosed by a sheath-like structure in life. Dark remains (Figure 1) are concentrated on the lower part of the trunk, indicating the presence of an internal cavity.
No openings can be seen on this laterally compressed specimen.
Remarks
The overall profile and the presence of bumps and depressions demonstrate the new taxon is a sessile and soft-bodied epibenthos, with the bottom of the trunk attached to the seafloor in life. The lamellar structures of the upper portion are non-prehensile, reminiscent of the tentacle rods in Xianguangia. The relatively regular longitudinal ridges (Figure 2C) imply some internal supporting structures of the trunk, i.e., the presence of the mesenterial divisions. There are some similarities to the Xianguangia from the Chengjiang biota, which bears proximal tentacle rods, a preserving outline of gut in the trunk and the mesenteries inside. The bipartite body and the possession of possible tentacles portray a polypoid outline for the new taxon, suggesting some relationships with cnidarians.
Until now, the Chengjiang biota has produced the most diverse purported cnidarians from Cambrian Series 2, e.g., Dinomischus, Xianguangia, Daihua, and Nailiana. The new taxon resembles Xianguangia in having tentacular structures and a main body (figure 1 in Zhao et al., 2019), but tentacles of the latter are feather-like (figure 1 in Ou et al., 2017), a character not found in the former. It is similar to Xianguangia and Daihua in the overall profile of a bipartite body (figure 2 in Zhao et al., 2019), however, the presence/absence of circumoral domes and the lowermost part of the body being flat (figure 1) or tapering to a blunt tip (figure 2I in Zhao et al., 2019) clearly discriminate the two. The new taxon approximates Nailiana in the general shape of the body (composed of unbranched tentacles and a trunk), but the tentacles of Nailiana are prehensile (Ou et al., 2022), whereas those of the new taxon more likely function as sclerotized proximal tentacle rods. The absence of a stalk sets the new taxon apart from Dinomischus.
Based on the morphological similarities and discrepancies documented above with the purported cnidarians from the Chengjiang biota, we tentatively leave the new metazoan from the Guanshan biota in open nomenclature and describe it as a cnidarian-like metazoan. So far, no exact cnidarians have been documented from the Guanshan biota. If the new material can be confirmed as a representative of Cnidaria, it will provide us with very important evidence for the evolution of cnidarians from the Chengjiang to Guanshan biotas.
The bryozoan-like metazoan (RCP-ZJ-0003-0008)
Description
RCP-ZJ-0003-0004 are preserved on a molted trilobite glabella (Figure 3) and RCP-ZJ-0005-0008 on a librigena (Figure 4). The specimens are dark-brown and in high relief (e.g., Figures 3B, C), with each bearing numerous, densely packed openings (e.g., Figure 3B).
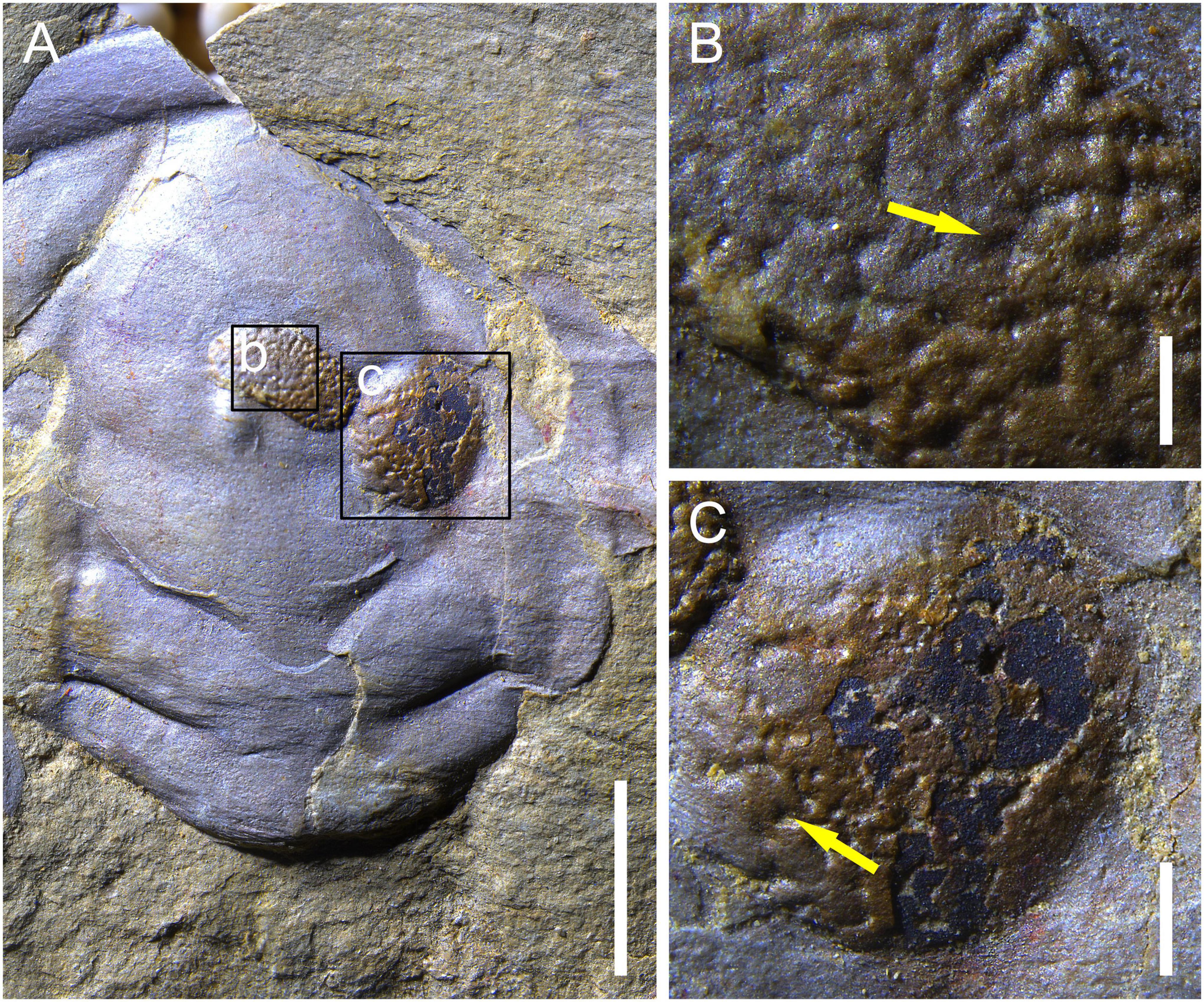
Figure 3. The unnamed bryozoan-like metazoan (RCP-ZJ-0003-0004). (A) RCP-ZJ-0003-0004 are preserved on a trilobite glabella; (B) detail of RCP-ZJ-0003 [position marked by frame b in panel (A)], yellow arrow indicates an opening of the specimen; (C) detail of RCP-ZJ-0004 [position marked by frame c in panel (A)], yellow arrow indicates an opening of the specimen. The scale bars are 5 mm for panel (A), 0.5 mm for panel (B), and 1 mm for panel (C).

Figure 4. The unnamed bryozoan-like metazoan (RCP-ZJ-0005-0008). (A) RCP-ZJ-0005-0008 are preserved on a trilobite librigenal; (B) detail of RCP-ZJ-0005 [position marked by frame b in panel (A)], yellow arrow indicates an opening of the specimen; (C) detail of RCP-ZJ-0006-0008 [position marked by frame c in panel (A)], white arrow indicates RCP-ZJ-0006; blue arrow indicates RCP-ZJ-0007 and yellow arrow indicates RCP-ZJ-0008. The scale bars are 5 mm for panel (A) and 1 mm for panels (B,C).
RCP-ZJ-0003 is oval in overall profile and, measures 4.5 × 3 mm (Figures 3A, B). The diameters of the openings range from 180 to 250 μm, with most being ca. 200 μm. RCP-ZJ-0004 is circular in outline and 4 mm in diameter (Figures 3A–C). The diameters of the openings measure 117 to 235 μm. RCP-ZJ-0005-0008 are counterparts (Figure 4); parts were collapsed into pieces during excavation and impossible to repair for photography. RCP-ZJ-0005 (Figure 4B) is sub-rectangular in general shape and measures 4.5 × 3.5 mm. The diameters of the openings vary from 114 to 286 μm. RCP-ZJ-0006-0008 are irregular in outline and, each comprises fewer openings (Figure 4C), with the diameters ranging from 91 to 152 μm.
Remarks
Morphological details cannot be observed clearly on these heavily mineralized specimens. In terms of the overall profile, each specimen resembles an assemblage of zooid tubes, i.e., a colony of bryozoans. Although it cannot be fully confirmed, the densely packed openings may represent the apical orifices of bryozoan zooids, which are used as the passage of the lophophore during life (e.g., figure 2 in Zhang et al., 2021; figure 3A in Pruss et al., 2022). Furthermore, the diameters of the openings also lie within the size range (ca. 50 μm–ca. 1 mm) of the orifices of zooids expected in ancestral Cambrian bryozoans (Taylor and Waeschenbach, 2015; Zhang et al., 2021). Attached to trilobite fragments, these fossils are definitively epibenthic metazoans, which add new epibenthos to the Guanshan biota. If confirmed as bryozoans, these specimens will further fill in the gap between the first fossil record of Cambrian Stage 3 to the diversity dynamics of Ordovician radiation of Bryozoa. The new material will represent the earliest known bryozoans from the Burgess Shale-type Lagerstätten and indicate that some representatives of Bryozoa were adapted for attaching to hard substrates in siliciclastic environments during Cambrian Stage 4.
Discussion
The “Cambrian explosion” (Brasier, 1979), beginning ca. 540 million years ago, was one of the most significant radiations of animal life in Earth’s history, and all major animal phyla started appearing in the fossil record during this period (Maloof et al., 2010; Hu et al., 2013; Tihelka et al., 2022). Studies have shown that this important evolutionary event resulted in the formation of metazoan-dominated ecosystems in the Phanerozoic (Erwin and Tweedt, 2012; Zhang and Shu, 2014; Zhang et al., 2021). The Burgess Shale-type Lagerstätten from Cambrian Series 2 and Miaolingian worldwide (e.g., the Chengjiang and Guanshan biotas from China and, the Burgess Shale from Canada) witnessed the construction of diversified marine ecosystems in this time interval. In terms of ecological niches, animals from these Lagerstätten can be classified into five categories: pelagic, nekton, vagile epibenthos, sessile epibenthos, and infauna (e.g., Hou et al., 2017). In any ecosystem, the routes of energy flow and matter cycling are determined by trophic relationships (Zhang et al., 2021). In this respect, biotic constituents from these deposits can be listed as producers (e.g., algae), consumers (e.g., euarthropods, priapulids, and radiodonts), and decomposers (which remain to be recognized).
Sessile epibenthos are indispensable in any marine ecosystem and play important part in the process of energy flow. Representatives of this group are abundant in the Guanshan biota, including sponges, chancelloriids, echinoderms, and some enigmatic taxa, e.g., Phlogites guangxiensis Hu et al., 2010 and a kleptoparasitic tube-dwelling organism (Zhang et al., 2020). The two new metazoans documented above show character traits with cnidarians and bryozoans; confirmed or not, both can be included in sessile epibenthos. The discovery of more epibenthic organisms suggests that the benthic ecosystem of the Guanshan biota is much more diversified than previously thought, and hence implies a complex trophic structure and rather sophisticated epibenthic ecological interactions for the Lagerstätte.
Conclusion
We describe two new sessile epibenthos and, their general morphology indicates them to be possible cnidarians and bryozoans. Our new material demonstrates that the Guanshan biota has developed an epibenthic ecosystem much more complex than previously expected.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
JZ and YL designed the study and collected and prepared the fossil specimens. JZ photographed the specimens and prepared the figures. JZ and YL wrote the first draft of the manuscript with substantial input from PS. All authors described and interpreted the specimens, contributed to the article, and approved the submitted version.
Acknowledgments
We are grateful to Fei Li and Zhongwei Li for the assistance during field work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Brasier, M. D. (1979). “The Cambrian radiation event,” in The origin of major invertebrate groups, ed. M. R. House (New York, NY: Academic Press), 103–159.
Brock, G. A., and Cooper, B. J. (1993). Shelly fossils from the early Cambrian (Toyonian) Wirrealpa, Aroona Creek, and Ramsay Limestones of South Australia. J. Paleontol. 67, 758–787. doi: 10.1017/S0022336000037045
Chen, F. Y., Topper, T. P., Skovsted, C. B., Strotz, L. C., Shen, J., and Zhang, Z. F. (2022). Cambrian ecological complexities: Perspectives from the earlies brachiopod-supported benthic communities in the early Cambrian Guanshan Lagerstätte. Gondwana Res. 107, 30–41. doi: 10.1016/j.gr.2022.02.008
Chen, F. Y., Zhang, Z. F., Betts, M. J., Zhang, Z. L., and Liu, F. (2019). First report on Guanshan biota (Cambrian Stage 4) at the stratotype area of Wulongqing formation in Malong County, Eastern Yunnan. Geosci. Front. 10, 1459–1476. doi: 10.1016/j.gsf.2018.09.010
Chen, J. Y., and Erdtmann, B. D. (1991). “Lower Cambrian lagerstätte from Chengjiang, Yunnan, China: Insights for reconstructing early metazoan life,” in The early evolution of metazoa and the significance of problematic taxa, eds A. M. Simonetta and S. Conway Morris (Cambridge: Cambridge University Press), 57–76.
Chen, J. Y., Hou, X. G., and Lu, H. Z. (1989). Early Cambrian hock glass-like rare sea animal Dinomischus (Entoprocta) and its ecological features. Acta Pal. Sin. 28, 58–71.
Ding, Y., Liu, J. N., and Chen, F. F. (2020). Ichnology, palaeoenvironment, and ecosystem dynamics of the early Cambrian (stage 4, series 2) Guanshan biota, South China. Geol. J. 55, 77–94. doi: 10.1002/gj.3360
Ernst, A. (2020). “Fossil record and evolution of Bryozoa,” in Bryozoa, eds J. G. Helmcke, D. Starck, and H. Wermuth (Berlin: De Gruyter), 11–55. doi: 10.1515/9783110586312-002
Erwin, D. H., and Tweedt, S. (2012). Ecological drivers of the Ediacaran-Cambrian diversification of metazoa. Evol. Ecol. 26, 417–433. doi: 10.1098/rspb.2014.0038
Hageman, S. J., and Ernst, A. (2019). The last phylum: Occupation of bryozoa morpho-ecospace (colony growth habits) during the early phase of the Great Ordovician Biodiversification Event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 534:109270. doi: 10.1016/j.palaeo.2019.109270
Han, J., Guo, J. F., Ou, Q., Song, Z. C., Liu, P., Hao, W. J., et al. (2020). Evolutionary framework of early Cambrian cnidarians from South China. Earth Sci. Front. 27, 067–078. doi: 10.13745/j.esf.sf.2020.6.3
Hou, X. G., Siveter, D. J., Siveter, D. J., Aldridge, R. J., Cong, P. Y., Gabbott, S. E., et al. (2017). The Cambrian fossils of Chengjiang, China: The flowering of early animal life. Hoboken, NJ: John Wiley & Sons, 307. doi: 10.1002/9781118896372
Hu, S. X., Li, X. K., Tan, X. H., Zhao, D. Q., and Luo, H. L. (2010). Phlogites from the early Cambrian Guanshan fauna. Acta Pal. Sin. 49, 360–364.
Hu, S. X., Zhao, F. C., Liu, A. G., and Zhu, M. Y. (2023). A new Cambrian frondose organism: “Ediacaran survivor” or convergent evolution? J. Geol. Soc. 180. doi: 10.1144/jgs2022-088
Hu, S. X., Zhu, M. Y., Luo, H. L., Steiner, M., Zhao, F. C., Li, G. X., et al. (2013). The Guanshan biota. Kunming: Yunnan Science and Technology Press, 204.
Lidgard, S., Carter, M. C., Dick, M. H., Gordon, D. P., and Ostrovsky, A. N. (2012). Division of labor and recurrent evolution of polymorphisms in a group of colonial animals. Evol. Ecol. 26, 233–257. doi: 10.1007/s10682-011-9513-7
Liu, J. N., Han, J., Li, J. S., Wu, Y. C., Peng, J., Qi, N., et al. (2016). New localities and Palaeoscolecid worms from the Cambrian (Stage 4, Series 2) Guanshan biota in Kunming, Yunnan, South China. Acta Geol. Sin. 90, 1939–1945. doi: 10.1111/1755-6724.13013
Maloof, A. C., Porter, S. M., Moore, J. L., Dudas, F. Q., Bowring, S. A., Higgins, J. A., et al. (2010). The earliest Cambrian record of animals and ocean geochemical change. Geol. Soc. Am. Bull. 122, 1731–1774. doi: 10.1130/B30346.1
Ou, Q., Han, J., Zhang, Z. F., Shu, D. G., Su, G., and Mayer, G. (2017). Three Cambrian fossils assembled into an extinct body plan of cnidarian affinity. Proc. Natl. Acad. Sci. U.S.A. 114, 8835–8840. doi: 10.1073/pnas.1701650114
Ou, Q., Shu, D. G., Zhang, Z. F., Han, J., Iten, H. V., Cheng, M. R., et al. (2022). Dawn of complex animal food webs: A new predatory anthozoan (Cnidaria) from Cambrian. Innovation 3:100195. doi: 10.1016/j.xinn.2021.100195
Pruss, S. B., Leeser, L., Smith, E. F., Zhuravlev, A. Y., and Taylor, P. D. (2022). The oldest mineralized bryozoan? A possible palaeostomate in the lower Cambrian of Nevada, USA. Sci. Adv. 8:eabm8465. doi: 10.1126/sciadv.abm8465
Schwaha, T. F., Ostrovsky, A. N., and Wanninger, A. (2020). Key novelties in the evolution of the aquatic colonial phylum bryozoa: Evidence from soft body morphology. Biol. Rev. 95, 696–729. doi: 10.1111/brv.12583
Taylor, P. D., and Waeschenbach, A. (2015). Phylogeny and diversification of bryozoans. Palaeontology 58, 585–599. doi: 10.1111/pala.12170
Taylor, P. D., Lombardi, C., and Cocito, S. (2015). Biomineralization in bryozoans: Present, past and future: Bryozoan biomineralization. Biol. Rev. 90, 1118–1150. doi: 10.1111/brv.12148
Tihelka, E., Howard, R. J., Cai, C. Y., and Lozano-Fernandez, J. (2022). Was there a Cambrian explosion on land? The case of arthropod terrestrialization. Biology 11:1516. doi: 10.3390/biology11101516
Zhang, X. L., and Shu, D. G. (2014). Causes and consequences of the Cambrian explosion. Sci. China Earth Sci. 57, 930–942. doi: 10.1007/s11430-013-4751-x
Zhang, Z. F., Strotz, L. C., Topper, T. P., Chen, F. Y., Chen, Y. L., Liang, Y., et al. (2020). An encrusting kleptoparasite-host interaction from the early Cambrian. Nat. Commun. 11, 1–7. doi: 10.1038/s41467-020-16332-3
Zhang, Z. L., Zhang, Z. F., Ma, J. Y., Taylor, P. D., Strotz, L. C., Jacquet, S. M., et al. (2021). Fossil evidence unveils an early Cambrian origin for bryozoa. Nature 599, 251–255. doi: 10.1038/s41586-021-04033-w
Zhao, J., Li, Y. J., Selden, P. A., and Cong, P. Y. (2020). New occurrence of the Guanshan Lagerstätte (Cambrian Series 2, Stage 4) in the Kunming area, Yunnan, southwest China, with records of new taxa. Alcheringa 44, 343–355. doi: 10.1080/03115518.2020.1781257
Keywords: Cnidaria, Bryozoa, Burgess Shale-type Lagerstätten, South China, Cambrian Series 2, Cambrian explosion
Citation: Zhao J, Li Y and Selden PA (2023) Two new metazoans from the Cambrian Guanshan biota of China. Front. Ecol. Evol. 11:1160530. doi: 10.3389/fevo.2023.1160530
Received: 07 February 2023; Accepted: 14 March 2023;
Published: 31 March 2023.
Edited by:
Bertrand Lefebvre, Université Claude Bernard Lyon 1, FranceCopyright © 2023 Zhao, Li and Selden. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yujing Li, eXVqaW5nbGlAeW5udS5lZHUuY24=