 Jun Zhao
Jun Zhao- 1Research Center of Paleobiology, Yuxi Normal University, Yuxi, China
- 2Yunnan Key Laboratory of Plateau Geographical Processes and Environmental Changes, Faculty of Geography, Yunnan Normal University, Kunming, China
- 3Department of Geology, University of Kansas, Lawrence, KS, United States
- 4Natural History Museum, London, United Kingdom
Annelids are abundant and speciose in the modern world but are comparatively few in the fossil record. Primitive annelids were expected to have developed eyes and nuchal organs, but until now definitive evidence is still lacking. Based on a new specimen from the Wulongqing Formation, we describe Gaoloufangchaeta bifurcus gen. et sp. nov. from the Guanshan biota (Cambrian Series 2, Stage 4) of Yunnan province, China. The overall profile of the body and the presence of tentacles and stout parapodia with simple chaetae establish it as a primitive polychaete. By bearing bicellular eyes and possible nuchal organs, the new form has developed relatively strong sensory abilities. Our material further confirms that polychaetes were already diverse by Cambrian Series 2, indicating a much earlier origin for the group.
Introduction
Annelida is a highly diverse phylum that includes approximately 17,000 described species (Nanglu and Caron, 2018). The records of fossil representatives are sparse because annelids are entirely soft-bodied and decay rapidly (Briggs and Kear, 1993; Parry et al., 2014). Whole-body fossil annelids have been recovered from early Cambrian to Carboniferous strata (Conway Morris, 1979; Schram, 1979; Thompson, 1979; Briggs et al., 1996; Sutton et al., 2001; Huang et al., 2004; Farrel and Briggs, 2007; Vinther et al., 2008; Högström et al., 2009; Briggs and Bartels, 2010; Liu et al., 2015; Han et al., 2019). In particular, the Cambrian annelids are predominantly polychaetes, which are strikingly various and widely distributed, including Burgessochaeta setigera (Walcott, 1911), Canadia spinosa (Walcott, 1911), Peronochaeta dubia (Walcott, 1911), Insolicorypha psygma (Conway Morris, 1979), Stephenscolex argutus (Conway Morris, 1979), and Kootenayscolex barbarensis (Nanglu and Caron, 2018) from the Burgess Shale, Phragmochaeta canicularis (Conway Morris and Peel, 2008), and Pygocirrus butyricampum (Vinther et al., 2011) from Sirius Passet, Ipoliknus avitus (Han et al., 2019), and Adelochaeta sinensis (Han et al., 2019) from Chengjiang, Dannychaeta tucolus (Chen et al., 2020) from the Cambrian Canglangpu Formation of Yunnan, and Guanshanchaeta felicia (Liu et al., 2015) from Guanshan. Nanglu and Caron (2018) proposed that primitive annelids could have developed eyes and nuchal organs. However, of all the polychaetes documented above, none has been demonstrated to possess eyes thus far.
Here, we describe a new primitive polychaete, Gaoloufangchaeta bifurcus gen. et sp. nov., from the Guanshan biota, which is characterized by the presence of a possibly tripartite body profile, a pair of tentacles, bicellular eyes, possible nuchal organs, and a bifid pygidium. These are the first taxon-bearing visual organs among Cambrian annelids. Our material provides a new epibenthic soft-bodied metazoan for the Burgess Shale-type Lagerstätte and further confirms the diversity of polychaetes by Cambrian Series 2, demonstrating that the group originated in a deeper time.
Geological setting
The Guanshan biota occurs in the Wulongqing Formation (Cambrian Series 2, Stage 4), which is distributed widely in eastern Yunnan, southwest China, including the Kunming-Wuding and Malong-Yiliang areas. The best-studied sections include the Gaoloufang and Gangtoucun sections of Kunming, the Shijiangjun section of Wuding, the Lihuazhuang section of Yiliang, and the Wulongqing section of Malong (Hu et al., 2013; Figure 1).

Figure 1. Paleogeographic and geographic maps, showing the occurrence of Gaoloufangchaeta bifurcus gen. et sp. nov. (A) paleogeographic reconstruction in early and middle Cambrian, modified from Torsvik and Cocks (2017); (B) the main fossil sites of the Guanshan biota in eastern Yunnan. 1, Shijiangjun; 2, Gangtoucun; 3, Gaoloufang; 4, Lihuazhang; 5, Wulongqing.
The specimen described in this study was collected from the Gaoloufang section (24.95916°N, 102.80539°E), which is ~ 15 km southeast of the downtown area of Kunming (Figure 1). The Cambrian succession in this section mainly consists of the Hongjingshao and Wulongqing Formations. The former is composed of dark brown sandstones and gray argillaceous siltstones, while the latter comprises grayish-yellow silty mudstones interbedded with dark brown siltstones.
Materials and methods
The single specimen (RCP-ZJ-0001), preserved in a gray-yellow mudstone, was prepared using a fine needle under a Nikon SMZ 800 N microscope, revealing parts covered by matrix. Digital photographs were taken using a Canon EOS 5D SR camera with a Canon MP-E 65 (1-5X) macro lens under cross-polarized light, with the brightness and contrast of the resulting images processed in Adobe Photoshop CS 5. The specimen was imaged using an FEI Quanta 650 FEG scanning electron microscope. Elemental mapping was undertaken using an EDAX Pegasus energy dispersive X-ray spectroscopy (EDX) system, 112 × magnification, 10.3 mm working distance, and an accelerating voltage of 20 kV. The specimen is housed in the Research Center of Paleobiology, Yuxi Normal University (RCP).
The specimen is of a bizarre appearance. Behind the second pair of uniramous parapodia, there is an apparent constriction, forming a neck-like structure, which is not known for any other Cambrian polychaetes (Figure 2). Based on the overall morphology of the body, we suspect that this is a taphonomic artifact. We tentatively divide the body into three regions as follows: anterior part, trunk, and pygidium, as shown in Figure 2.

Figure 2. Gaoloufangchaeta bifurcus gen. et sp. nov. from the early Cambrian Guanshan biota (Yunnan Province, China). AP, anterior part; Pg, pygidium; Tr, trunk. The scale bar is 2 mm.
The current publication and the new species are registered in ZooBank as urn:lsid:zoobank.org: act:54120A6E-2195-478E-A924-E686FD88D633 and urn:lsid:zoobank.org:act:D0F29632-9BDC-4CF6-9299-D84279B28A46, respectively.
Systematic paleontology
Phylum Annelida (Lamarck, 1809)
Class Polychaeta (Grube, 1850)
Gaoloufangchaeta gen. nov.
Derivation of name
Gaoloufang, referring to the location where the fossil was found; chaeta, a diagnostic characteristic of polychaetes.
Type species
Gaoloufangchaeta bifurcus sp. nov.
Diagnosis
Body tripartite (tentative) and elongated, bearing a pair of tentacles, bicellular eyes, and possible nuchal organs; the anterior part of the body being a reversed trapezoid in outline; at least seven pairs of uniramous parapodia with simple chaetae; pygidium large and bifid.
Gaoloufangchaeta bifurcus sp. nov.
Derivation of name
From the Latin bifurcus, referring to the bifid structure on the pygidium.
Holotype
RCP-ZJ-0001, a complete specimen.
Type locality
Gaoloufang section, Kunming, Yunnan, China.
Type horizon
Wulongqing Formation (Cambrian Series 2, Stage 4).
Diagnosis
As for genus.
Description
The body is bilaterally symmetrical and possibly tripartite (composed of the anterior part, trunk, and pygidium) and preserved dorsoventrally flattened in parallel aspect, measuring 9 mm long (excluding tentacles).
The anterior part is a reversed trapezoid in the overall profile and measures 2 mm in length and 2.5 mm in maximum width (including parapodia, just above mid-length; Figures 2, 3A). A pair of tentacles stretches out anterolaterally from near the center of the anterior margin and tapers distally, with the best-preserved branch (left) measuring 1 mm long and 0.5 mm wide at the base (Figure 3). A pair of bicellular eyes is located behind the anterior margin and close to the bases of tentacles, with the diameter of each being 0.3 mm (Figure 3A). A total of two short longitudinal grooves, with each being 0.2 mm in length, are situated behind the eyes, which are interpreted here as possible nuchal organs (Figure 3A). A sub-ovoid dark patch lies behind the possible nuchal organs and may represent traces of the mouth or a partly preserved pharynx on the ventral side (Figure 3A). The posterior region of the anterior part is apparently constricted, forming a neck-like structure (Figure 2). Two pairs of uniramous parapodia are lined along both sides of the anterior part and are roughly triangular in outline (Figures 2, 3).

Figure 3. Gaoloufangchaeta bifurcus gen. et sp. nov. (A) detail of Figure 2, position marked by frame a. (B, C) SEM-EDX maps of (A), the specimen was rotated slightly to the left. Ey, eye; Mo?, possible mouth; NO?, possible nuchal organ; Pa, parapodium; Te, tentacle. The scale bars are 0.5 mm.
The trunk is elongated and tapers posteriorly, measuring 6 mm in length and approximately 3 mm in maximum width (at the first pair of parapodia). It comprises at least five homonomous segments, with each bearing a pair of uniramous parapodia that are broadly based and taper slightly distally, ending in blunt terminations (Figures 2, 4A, B). Chaetae are simple, thin, and situated at the distal ends of parapodia (Figures 4A, C). The posterior right side of the trunk is obscured, likely as a result of decay (Figure 4C). The boundaries of trunk segments are faint. The posterior part of the trunk is constricted slightly and associated with the pygidium and is devoid of any parapodia (Figure 4D). Column-like structures along the median trunk region are interpreted as traces of the gut (Figure 4A).
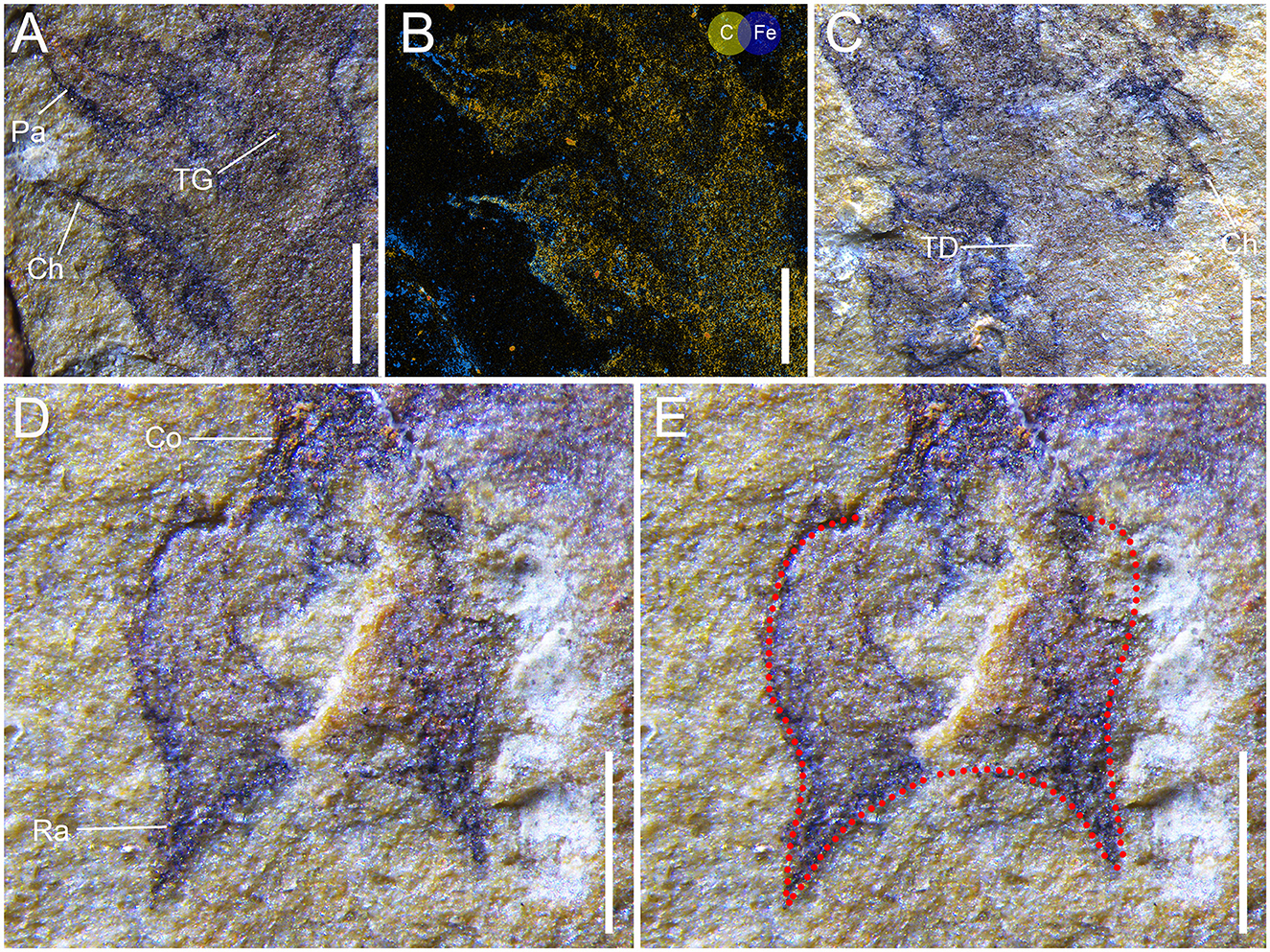
Figure 4. Gaoloufangchaeta bifurcus gen. et sp. nov. (A) detail of Figure 2, position marked by frame b. (B) EDX map of (A), the specimen was rotated anticlockwise slightly. (C) Detail of Figure 2, position marked by frame c. (D) Detail of Figure 2, position marked by frame d; the specimen was rotated clockwise a little to show the pygidium in upright orientation. (E) Detail of Figure 2, position marked by frame d; the red dotted line shows the profile of the pygidium. Ch, chaeta; Co, constriction; Pa, parapodium; Ra, ramus; TD, traces of decay; TG, traces of gut. The scale bars are 0.5 mm.
The pygidium is sub-rectangular in outline and measures 0.5 mm in length and 1.2 mm in width. From its posterior–lateral corners, two triangular structures arise (0.5 mm long for each), creating a more or less U-shaped morphology of the posterior margin (Figures 4D, E).
Remarks
The general shape of the body and the presence of tentacles, eyes, parapodia with chaetae, and pygidium clearly demonstrate that Gaoloufangchaeta bifurcus is a polychaete annelid. Chaetae are hard parts and most easily to be preserved, but they are absent in most parapodia of the new form. Decay could result in the detachment of chaetae in Cambrian polychaetes, e.g., Canadia spinosa (Walcott, 1911; Briggs et al., 1994; Parry et al., 2016). Traces of decay are present at the posterior right side of the trunk of G. bifurcus (Figure 4C), indicating that the body indeed experienced a period of decay prior to burial.
Gaoloufangchaeta bifurcus resembles Guanshanchaeta felicia (Liu et al., 2015) in having an elongated body, a pair of tentacles, and a bifid pygidium. However, G. bifurcus differs from G. felicia in that the body is much shorter and the trunk segments fewer; parapodia are uniramous and the first two pairs are followed by a constriction, forming a reversed trapezoid for the anterior part of the body (although this could be a taphonomic artifact; Figure 2). In addition, in G. bifurcus, the pygidium is strongly differentiated from the trunk and large relative to the body size (Figures 2, 4D, E), whereas it is poorly defined and rather smaller in G. felicia (figure 1 in Liu et al., 2015). G. felicia tapers both anteriorly and posteriorly, a morphological characteristic not found in G. bifurcus. Lastly, G. bifurcus differs from G. felicia in the presence of eyes and possible nuchal organs and the absence of a buccal tube. The differences above discriminate G. bifurcus from being a juvenile of G. felicia.
Han et al. (2019) described two polychaetes [Ipoliknus avitus (Han et al., 2019) and Adelochaeta sinensis (Han et al., 2019)] and three additional polychaete specimens from the earlier Chengjiang biota. Gaoloufangchaeta bifurcus differs from Ipoliknus avitus, in particular, in the absence of sclerites and evident segment boundaries. G. bifurcus is different from Adelochaeta sinensis in that the latter has a clearly defined head region and aciculae, which are absent from the former. The most obvious similarity between G. bifurcus and the three unidentified polychaetes is the presence of chaetae; further comparisons are hard to make owing to the incomplete preservation of the three individuals. The absence of characteristics such as palps, prostomium, thorax, and parapodia with lateral lamellae discriminates G. bifurcus from Dannychaeta tucolus (Chen et al., 2020), a tube-dwelling polychaete from the Canglangpu formation (Cambrian Series 2, Stage 3) of Yunnan province.
Hitherto, the Burgess Shale has yielded the most abundant polychaetes of various morphologies in Cambrian Lagerstätten. Gaoloufangchaeta bifurcus approximates Burgessochaeta setigera and Canadia spinosa in the elongated profile of the body and by having uniramous parapodia. However, the number of body segments and the presence or absence of branchia discriminate the three taxa. Peronochaeta dubia and Stephenscolex argutus most differ from G. bifurcus in that they bear papillae at the anterior end of the body, instead of tentacles. G. bifurcus shares the same body length with Insolicorypha psygma, whereas the latter has a bipartite head region and more trunk segments. Kootenayscolex barbarensis bears a pair of large palps and a median antenna, which are absent from G. bifurcus.
Different from Gaoloufangchaeta bifurcus, Phragmochaeta canicularis (Conway Morris and Peel, 2008), a polychaete from the Sirius Passet Lagerstätte, has numerous long and thin chaetae and more trunk segments. The body terminating in a bifid structure is the most apparent similarity between G. bifurcus and Pygocirrus butyricampum (Vinther et al., 2011), another polychaete from the Sirius Passet. However, the profile of the bifid structure is different between the two taxa: V-shaped for P. butyricampum and more or less U-shaped for G. bifurcus. In addition, P. butyricampum has more trunk segments.
Discussion
Gaoloufangchaeta bifurcus gen. et sp. nov. is described based on a single specimen with decay traces, and thus, some of its characteristics cannot be confirmed as original. The neck-like structure (Figure 2) may be an artifact formed during burial given the soft nature of annelid bodies, and hence, the tripartite outline of the body of the new form is tentative in this study; the parapodia lined along both sides of the trunk appear to be too stout, and whether each is composed of two lobes (i.e., notopodium and neuropodium) cannot be confirmed on this comparatively poorly preserved specimen; they are temporarily interpreted here as uniramous; the morphologies of the inferred nuchal organs and mouth are not clearly shown on the specimen, and their identities need to be confirmed by additional and better-preserved ones; decay traces obscure the morphology of the posterior part of the trunk (Figures 2, 4C).
Gaoloufangchaeta bifurcus from Cambrian Series 2 of southwest China is one of the oldest fossil annelids recovered so far. Structures such as eyes and nuchal organs have previously been expected but not found in ancestral annelids (Weigert et al., 2014; Nanglu and Caron, 2018). The specimen of G. bifurcus clearly shows evidence of eyes and possible nuchal organs (Figures 2, 3) for the first time in Cambrian polychaetes, demonstrating that the oldest annelids are not all devoid of these sensory organs. By having tentacles, eyes, possible nuchal organs, and stout parapodia, G. bifurcus clearly had comparatively strong sensory abilities and an active and epibenthic lifestyle. Although no evidence of a proboscis with jaws is found in the single specimen, the possibility of the new taxon being a predator or scavenger cannot be ruled out. Our material not only provides a new epibenthic representative for early Cambrian marine animals but also a new soft-bodied metazoan for the Guanshan biota.
Until now, fossil polychaetes are the most diversified and abundant in the Burgess Shale (Cambrian Miaolingian, Wuliuan); the earliest whole-body preserved ones date back to Cambrian Series 2, Stage 3, represented by Phragmochaeta canicularis and Pygocirrus butyricampum from Sirius Passet, and Ipoliknus avitus and Adelochaeta sinensis from Chengjiang. Together with Gaoloufangchaeta bifurcus, the slightly younger Guanshan biota (Figure 5) has yielded two types of polychaetes. These occurrences confirm that polychaetes were already diverse in morphology by Cambrian Series 2, indicating a much earlier origin for the group.
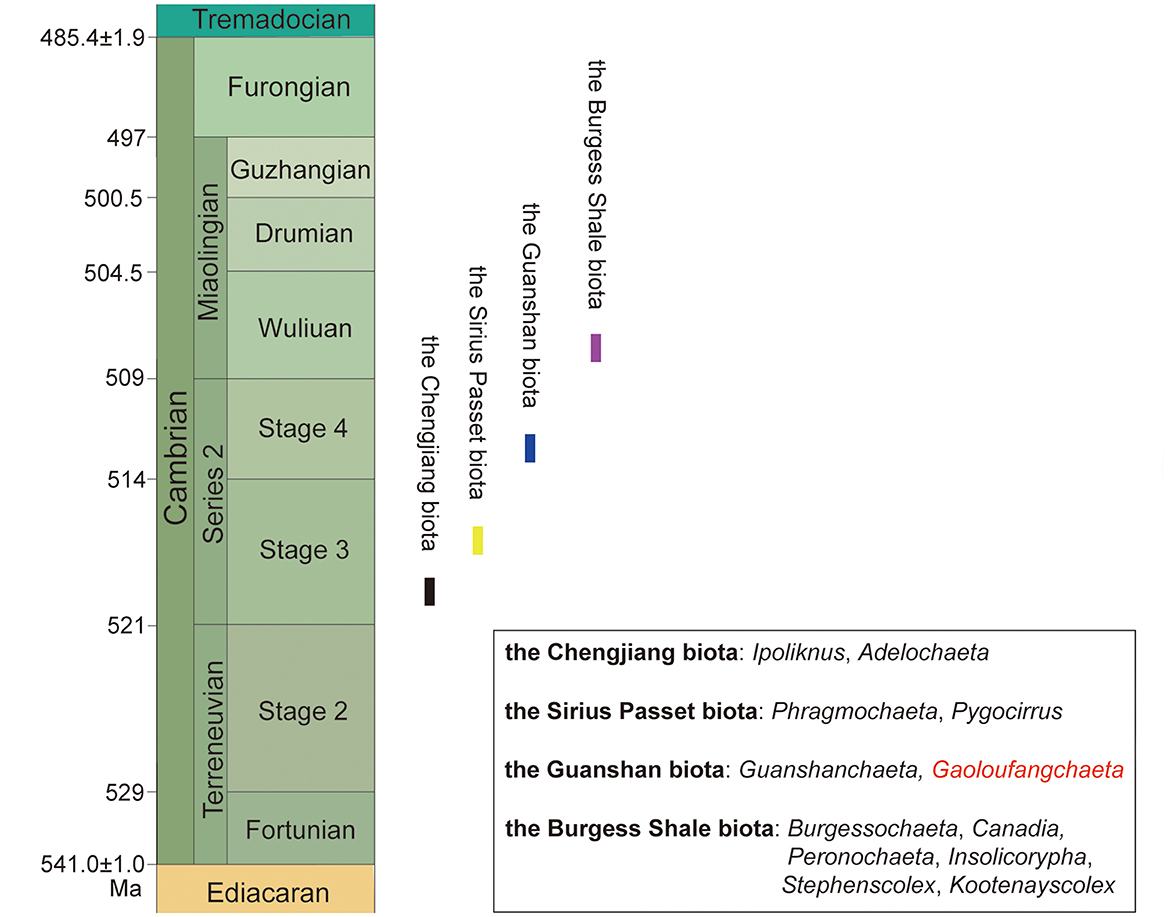
Figure 5. Main early and middle Cambrian fossil Lagerstätten yielding polychaetes.
Gaoloufangchaeta shows the characteristics proposed for primitive annelids, i.e., errant polychaete body form, parapodia with simple chaetae, and prostomial sensory organs (Parry et al., 2014; Weigert et al., 2014), and thus, it is among the primitive representatives of Annelida. Of all the early and middle Cambrian polychaetes, only the bodies of Pygocirrus, Guanshanchaeta, and Gaoloufangchaeta terminate in a bifid structure, suggesting a similar phylogenetic grade in between. Gaoloufangchaeta bears eyes and possible nuchal organs that are absent from Pygocirrus and Guanshanchaeta, which implies that Gaoloufangchaeta might be comparatively more derived and/or even a representative of the last common ancestor of annelids.
Conclusion
We describe a new primitive polychaete, Gaoloufangchaeta bifurcus gen. et sp. nov., from the Cambrian Series 2, Stage 4 of southwest China. This is the second polychaete from the Guanshan biota and among the oldest records of fossil annelids globally. Our material not only demonstrates that relatively strong sensory organs had developed in Cambrian annelids but also further confirms that polychaetes were already diverse in morphology by Cambrian Series 2.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
JZ and YL designed the study, collected and prepared the fossil, and wrote the first draft of the manuscript with substantial input from PS. JZ photographed the specimen and prepared the figures. All authors described and interpreted the specimen. All authors contributed to the article and approved the submitted version.
Acknowledgments
We are grateful to Fei Li and Zhongwei Li for their assistance during fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Briggs, D. E. G., and Bartels, C. (2010). Annelids from the Lower Devonian Hunsrück Slate (Lower Emsian, Rhenish Massif, Germany). Palaeontology 53, 215–232. doi: 10.1111/j.1475-4983.2009.00927.x
Briggs, D. E. G., Erwin, D. H., and Collier, F. J. (1994). The Fossils of the Burgess Shale. Washington, DC; London: Smithsonian Institution Press.
Briggs, D. E. G., and Kear, A. J. (1993). Decay and preservation of polychaetes: taphonomic thresholds in soft-bodied organisms. Paleobiology 19, 107–135. doi: 10.1017/S0094837300012343
Briggs, D. E. G., Siveter, D. J., and Siveter, D. J. (1996). Soft-bodied fossils from a Silurian volcaniclastic deposit. Nature 382, 248–250. doi: 10.1038/382248a0
Chen, H., Parry, L. A., Vinther, J., Zhai, D. Y., Hou, X. G., and Ma, X. Y. (2020). A Cambrian crown annelid reconciles phylogenomics and the fossil record. Nature 583, 248–250. doi: 10.1038/s41586-020-2384-8
Conway Morris, S. (1979). Middle Cambrian polychaetes from the Burgess Shale of British Columbia. Philos. Trans. R. Soc. of Lond. B. 285, 227–274. doi: 10.1098/rstb.1979.0006
Conway Morris, S., and Peel, J. S. (2008). The earliest annelids: lower Cambrian polychaetes from the Sirius Passet Lagerstätte, Peary Land, north Greenland. Acta Palaeontol. Pol. 53, 137–148. doi: 10.4202/app.2008.0110
Farrel, Ú., and Briggs, D. E. G. (2007). A pyritized polychaete from the Devonian of Ontario. Proc. R. Soc. B 274, 499–504. doi: 10.1098/rspb.2006.0063
Han, J., Conway Morris, S., Hoyal Cuthill, J. F., and Shu, D. (2019). Sclerite-bearing annelids from the lower Cambrian of South China. Sci. Rep. 9, 1–11. doi: 10.1038/s41598-019-40841-x
Högström, A. E. S., Briggs, D. E. G., and Bartels, C. (2009). A pyritized lepidocoleid machaeridian from the Lower Devonian Hunsrück Slate, Germany. Proc. R. Soc. B 276, 1981–1986. doi: 10.1098/rspb.2008.1875
Hu, S. X., Zhu, M. Y., Luo, H. L., Steiner, M., Zhao, F. C., Li, G. X., et al. (2013). The Guanshan Biota. Kunming: Yunnan Science and Technology Press [in Chinese with English summary].
Huang, D. Y., Chen, J. Y., Vannier, J., and Saiz Salinas, J. (2004). Early Cambrian sipunculan worms from southwest China. Proc. R. Soc. Lond. B 271, 1671–1676. doi: 10.1098/rspb.2004.2774
Liu, J., Ou, Q., Han, J., Li, J., Wu, Y., Jiao, G., et al. (2015). Lower Cambrian polychaete from China sheds light on early annelid evolution. Sci. Nat. 102, 1–7. doi: 10.1007/s00114-015-1285-4
Nanglu, K., and Caron, J. B. (2018). A new Burgess Shale polychaete and the origin of the annelid head revisted. Curr. Biol. 28, 319–326. doi: 10.1016/j.cub.2017.12.019
Parry, L., Tanner, A., and Vinther, J. (2014). The origin of annelids. Palaeontology 57, 1091–1103. doi: 10.1111/pala.12129
Parry, L. A., Edgecombe, G. D., Eibye-Jacobsen, D., and Vinther, J. (2016). The impact of fossil data on annelid phylogeny inferred from discrete morphological characters. Proc. R. Soc. B 283, 20161378. doi: 10.1098/rspb.2016.1378
Schram, F. R. (1979). Worms of the Mississippian Bear Gulch Limestone of central Montana, USA. San Diego, CA: Transactions of the San Diego Society of Natural History.
Sutton, M. D., Briggs, D. E. G., and Siveter, D. J. (2001). A three-dimensionally preserved fossil polychaete worm from the Silurian of Herefordshire England. Proc. R. Soc. B 268, 2355–2363. doi: 10.1098/rspb.2001.1788
Thompson, I. (1979). Errant polychaetes (Annelida) from the Pennsylvanian Essex fauna of northern Illinois. Palaeontogr. A. 163, 169–199.
Torsvik, T. H., and Cocks, L. R. M., (eds.). (2017). “Cambrian,” in Earth History and Palaeogeography (Cambridge: Cambridge University Press), 85–100. doi: 10.1017/9781316225523.006
Vinther, J., Eibye-Jacobsen, D., and Harper, D. A. T. (2011). An early Cambrian stem polychaete with pygidial cirri. Bio. Lett. 7, 929–932. doi: 10.1098/rsbl.2011.0592
Vinther, J., Roy, P. V., and Briggs, D. E. G. (2008). Machaeridians are palaeozoic armoured annelids. Nature 451, 185–188. doi: 10.1038/nature06474
Walcott, C. D. (1911). “Middle Cambrian annelids,” in Cambrian Geology and Paleontology II (Washington, DC: Smithsonian Institution), 109–144.
Keywords: Annelida, Burgess Shale-type Lagerstätte, South China, Cambrian explosion, soft-bodied organism
Citation: Zhao J, Li Y and Selden PA (2023) A new primitive polychaete with eyes from the lower Cambrian Guanshan biota of Yunnan Province, China. Front. Ecol. Evol. 11:1128070. doi: 10.3389/fevo.2023.1128070
Received: 20 December 2022; Accepted: 28 March 2023;
Published: 26 April 2023.
Edited by:
Farid Saleh, Yunnan University, ChinaReviewed by:
Danny Eibye-Jacobsen, University of Copenhagen, DenmarkJianni Liu, Northwest University, China
Copyright © 2023 Zhao, Li and Selden. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yujing Li, yujingli@ynnu.edu.cn