 Yuelin Li1,2,3*†
Yuelin Li1,2,3*† Maina John Nyongesah4†
Maina John Nyongesah4† Libin Deng5†Fasih Ullah Haider1,3
Libin Deng5†Fasih Ullah Haider1,3 Shizhong Liu1,3
Shizhong Liu1,3 Brian Njoroge Mwangi1,2,3Qianmei Zhang1,3Guowei Chu1,3Deqiang Zhang1,3Juxiu Liu1,3Ze Meng1,3
Brian Njoroge Mwangi1,2,3Qianmei Zhang1,3Guowei Chu1,3Deqiang Zhang1,3Juxiu Liu1,3Ze Meng1,3- 1Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou, China
- 2South China National Botanical Garden, University of Chinese Academy of Sciences, Beijing, China
- 3South China National Botanical Garden, Guangzhou, China
- 4Department of Biological Sciences, Jaramogi Oginga Odinga University of Science and Technology, Bondo, Kenya
- 5Academy of Forestry Inventory and Planning, State Forestry Administration, Beijing, China
Over various terrain types, natural forests exhibit tree size and biomass variation. We started long- term research that consists of forest vegetation surveys in the Dinghushan National Nature Reserve to comprehensively investigate productivity based on the structure and species composition of China’s forest communities. We grouped the trees into three categories, i.e., as large or mature (DBH > 30 cm), medium-sized or growing (DBH 10–30 cm), and small-sized or regenerating seedlings (DBH < 10 cm). Forest data observations, i.e., species DBH, height, and biomass components (trunk, leaves, branches, above-ground dry weight, and below-ground dry weight) were recorded by use of standard protocols. All recorded observations were statistically analyzed by use of SPSS version 25. To comprehend the connections between the many elements of forest bio-productivity, linear regression analysis was utilized. Total above-ground biomass was 34.19 ± 5.75Kg/tree, but varied results were obtained when the forest was clustered based on DBH. Large-sized trees contributed an average of 2400.54 ± 510.4 kg/tree (93.24%), while medium-sized trees contributed 171.61 ± 25.06 kg/tree, and the least was regenerating seedlings which contributed 3.013 ± 0.07 kg/tree. There were positive linear relationships for all life forms between biomass and DHB, as well as DBH and height. The evergreen broadleaved shrubs were shorter in height (3.06 ± 0.99 m) than palm-leaved life forms (19.29 ± 5.39 m). Height influenced biomass accumulation and hence C gain, where life forms with tall stands had higher biomass. Generalized biomass estimation without clustering based on life forms or size class underestimates biomass components and hence lower C stocks for most forests worldwide. It’s also crucial to remember that trees with big DBH have tall, broad, well-lit crowns, which raise the primary productivity of forests and increase carbon storage. Besides natural disturbance (Typhoon) and climate change, it could be interesting to understand the relationship between soil resources such as nutrient and soil moisture content. These are factors that have direct impact on growth and biomass accumulation.
1. Introduction
Comparative productivity and large-scale patterns of plant species richness have been described in parts of Asia (Guo, 1999; Shen et al., 2013; Wu et al., 2015), but more attempts have yet to be made to identify the mechanisms of such patterns. Forests are constantly in flux as they grow and vary in size, composition, structure, and health, as well as in response to climate shifts and oscillations (Shen et al., 2013; Maina and Li, 2021). Within natural forests and plantations, tree size and stock diversity can be noticed at any moment, especially over varied terrain (Weiner and Thomas, 1986; Maina and Li, 2021). Because of this temporal and spatial heterogeneity, it can be challenging to derive accurate estimates of forest carbon stocks and biomass accumulation (Skovsgaard and Vanclay, 2013). Climate change driven increase in CO2 has been predicted to affect forest productivity by affecting biomass accumulation in different plant functional groups (Shaver et al., 2000) as well as their carbon storage (carbon stock) ability (Shaver et al., 2000; Gielen and Ceulemans, 2001). Therefore, understanding the factors that influence changes in forest carbon stocks and biomass accumulation is essential for sound management decisions and scientific researches.
Site index, or average height at a given age, is a popular indicator for forest site quality used by forest growth modelers to consider productivity differences between stands. The application of site index is not appropriate for trees that have experienced height suppression since it frequently relies on time- consuming increment core ring-count data to estimate stand age and needs to completely account for changes in volume growth (Skovsgaard and Vanclay, 2013). The ability to stratify sampling among more homogeneous subsets of the population of interest and to identify relatively homogeneous areas to serve as “blocks” in replicated field experiment installations are made possible by changes in forest growth (Husch et al., 2002). When links between forest production and its environment are taken into account or known and integrated into decision support systems, we can research or forecast changes in forest structure and productivity brought on by climate shifts, management actions, or other disturbances in ecosystems such as Dinhushan nature reserve. Aboveground biomass in the forest reflects the status of forest in terms of environmental conditions and health of the entire ecosystem (KoÈh et al., 2017; Lutz et al., 2018). Although this biomass is a key component in forest contribution to gas and water fux, available studies have focused on remotely sensed data (Skovsgaard and Vanclay, 2013), with scanty data available for in situ measurements especially for Dinghushan nature reserve. Available measurements have considered the entire ecosystem without clustering in trunk diameter classes. Field-based data from Dinghushan forest will provide relevant information on the forest characteristics and unveil determinants of forest carbon stocks, the structure and composition as well the history of disturbance.
For modelers of forest carbon stocks or biomass accumulation, different growth forms or habits lead to further problems, particularly in multi-aged stands (Murali et al., 2005). This study intends to overcome this challenge by estimating biomass, tree height, and DBH based on life forms and grouped DBH.
Several researchers have reported allometric connections between standing biomass and diameter at breast height (DBH) (Comley and McGuinness, 2005; Kirui et al., 2006) and linear equations between DBH and height for biomass estimation (Murali et al., 2005). These correlations have been calculated between DBH and above-ground biomass or its component elements (Kirui et al., 2006). However, limited information is available for allometric relationships or linear equations between above-ground biomass and DBH or height and DBH for different life forms of group datasets in a heterogenous forest like the Dinghushan forest. We explored different growth facets in the Dinghushan forest to examine relationships between biophysical gradients and productivity. Our goal was to determine which biophysical factors account for regional variations in forest productivity. We hypothesized the strongest correlations between DBH and height for various biophysical parameters for distinct life forms.
2. Materials and methods
2.1. Study site description
This study was carried out in Dinghushan National Nature Reserve (23°09′21″–23°11′ 30″N, 112°30′39″–112°33′41″E), which is located in Dinghu District, Zhaoqing City, Guangdong Province, covering a total area of 1,155 ha. The region experiences a south subtropical monsoon climate, with 1714 mm of annual rainfall and 76% annual humidity. The dry season lasts from October to March, and the wet season is from April to September. About 80% of the annual precipitation falls during the wet season. The coldest month is January, which has a mean temperature of 13.8°C, while the hottest month is July, which has a mean temperature of 28.8°C. The predominant topography comprises low mountains and hills ranging from 100 to 700 meters. Southern subtropical lateritic red soil comprises the zonal soil, which comprises Devonian sandstone, shale, sandstone, and quartz sandstone (Zou et al., 2018). Dinghushan’s plant community has a 95% canopy density and is year-round evergreen. The plant community in Dinghushan can be separated into three stages based on the succession of the forest ecosystem: Masson pine coniferous forest in the early stage, coniferous and broad-leaved mixed forest in the middle stage, and monsoon evergreen broad-leaved forest in the later stage. Aidia canthioides, Macarashell cnga sampsonii, Gironniera subaequalis, Cryptocarya chinensis, Cryptocarya concinna, Schima superba, Castanopsis chinensis, etc., are the dominant species in the tree layer of monsoon evergreen broad-leaved forests (Zou et al., 2018).
A long-term monitoring experiment on the dynamics of the monsoon evergreen broadleaved forest was started in 1978 to examine the longer-term effects of climate change and human activity on ecological resilience, biodiversity, and ecosystem services. A 1-ha permanent monitoring site was also created in the monsoon evergreen broad-leaved forest. The core section of the reserve is made up of monsoon evergreen broadleaved forest that has not been altered and is located adjacent to a Buddhist monastery (Zhou et al., 2013). The landscape of the location before (A) and after (B) the typhoon is depicted in Figure 1 as the well-preserved monsoon evergreen broad-leaved forest suffered greatly from typhoon Mangkhut in 2018. Although previous studies have indicated Dinghushan nature reserve has not been heavily disturbed for more than 400 years (Zhou et al., 2006, 2013), the 2018 typhoon affected the number of tree stands as indicated in our previous study (Li et al., 2021). This study also illustrated that trees with <10 cm DBH and < 10 m heaigh were completelu destroyed by the storm as shown in Figure 1B while those with > 20 cm DBH were moderately destroyed. Table 1 provides a quick summary of the permanent site based on historical monitoring data (Zhang, 2011; Zhou et al., 2013, 2014).

Figure 1. Figure showing landscape of the study site location before (A) and after (B) the typhoon.

Table 1. Description of the permanent site.
3. Methods
3.1. Tree selection
Within the 100 m by 100 m 1-ha permanent plot, 10 sites were established and sub-divided into 10 m by 10 m, 100 subplots (quadrats) where inventories of trees was carried out. In the clearly defined plots and with the appearance of being in good health, trees of the four life types were marked for measurements. Following that, DBH measurements were taken for each tree, with each multi-stemmed tree’s stems being treated separately, as reported by Clough et al. (1997). Each stem’s DBH was measured at a height of around 1.3 meters above the substrate or above the highest buttress root. The life/growth forms includes: Broadleaf evergreen trees that had large leaves, that stay green all year long; Broadleaf evergreen shrub that are small tree-like multi-stemmed trunk plants with year round green leaves; deciduous evergreen broadleaved tree that shed their leaves during cold and dry seasons and palm leaved trees that were tall, slender with smooth trunks and occasionally with no branches but have large spreading leaves at the top.
3.2. Biomass
Based on the harvesting quadrats described by Luo et al. (2009), biomass for all the components was estimated in all 100 plots. All trees had a diameter at breast height larger than 3 cm; their height (H, m) and diameter (D, cm) were measured. We removed six to eight trees in each sub-plot, each with a different diameter, and measured the amounts of stem, branch, leaf, and root biomass. Linear equation as described by Murali et al. (2005) was adopted where height and basal area relations facilitated calculation of biomass. Additionally, the “clear-cutting approach” (all trees in a plot were chopped down) and the “mean tree method” (a number of “mean trees” in a plot were fallen) were used in a few plots to estimate the tree biomass.
Data analysis.
SPSS 25 was used for all statistical analysis (IBM, Armonk, NY, United States). To comprehend the correlations between the biomass of various components and predictor variables, linear regression analysis was utilized. All values were reported as mean and standard error for each parameter.
4. Results
4.1. Stand characteristics
In the study, 6,883 trees and shrubs stands were recorded in the 100 sub-plots from which four life forms were identified within which several species were investigated for their diameter at breast height (DBH), stand height and biomass of trunk, leaves, branches, total above ground and below ground dry weights. The plots were dominated by evergreen broadleaved trees (5,723) followed by evergreen broadleaved shrubs (1,145), palm type life form (7), and the least dominant being deciduous broadleaved trees (8). Total number of trees per hectare varied before and after the typhoon as described in our study site section and presented in Figure 1. A study carried out by Li et al. (2021) indicated a total of 5,682 trees in 2015 and 5,022 trees per hectare in 2018. This is indicated that the typhoon caused a decline in the number of tree stands. Our study indicated an increase in trees and shrubs by 1,861 stands accounting for 37% recovery in 2020.
4.2. Tree height (m)
The average tree height for the entire forest was 4.7 ± 3.22 m. This height varied depending on the life form where the tallest trees were from the palm leaved trees (19.29 ± 5.39 m) followed by deciduous broadleaved trees (8.05 ± 4.55 m), evergreen broadleaved trees (5.11 ± 3.35 m) and shortest being evergreen broadleaved shrubs (3.06 ± 0.99 m; Figure 2A). Tall trees from evergreen broadleaved trees and palm leaved life forms with over 30 cm DBH had heights above 22 m (Figure 3D).
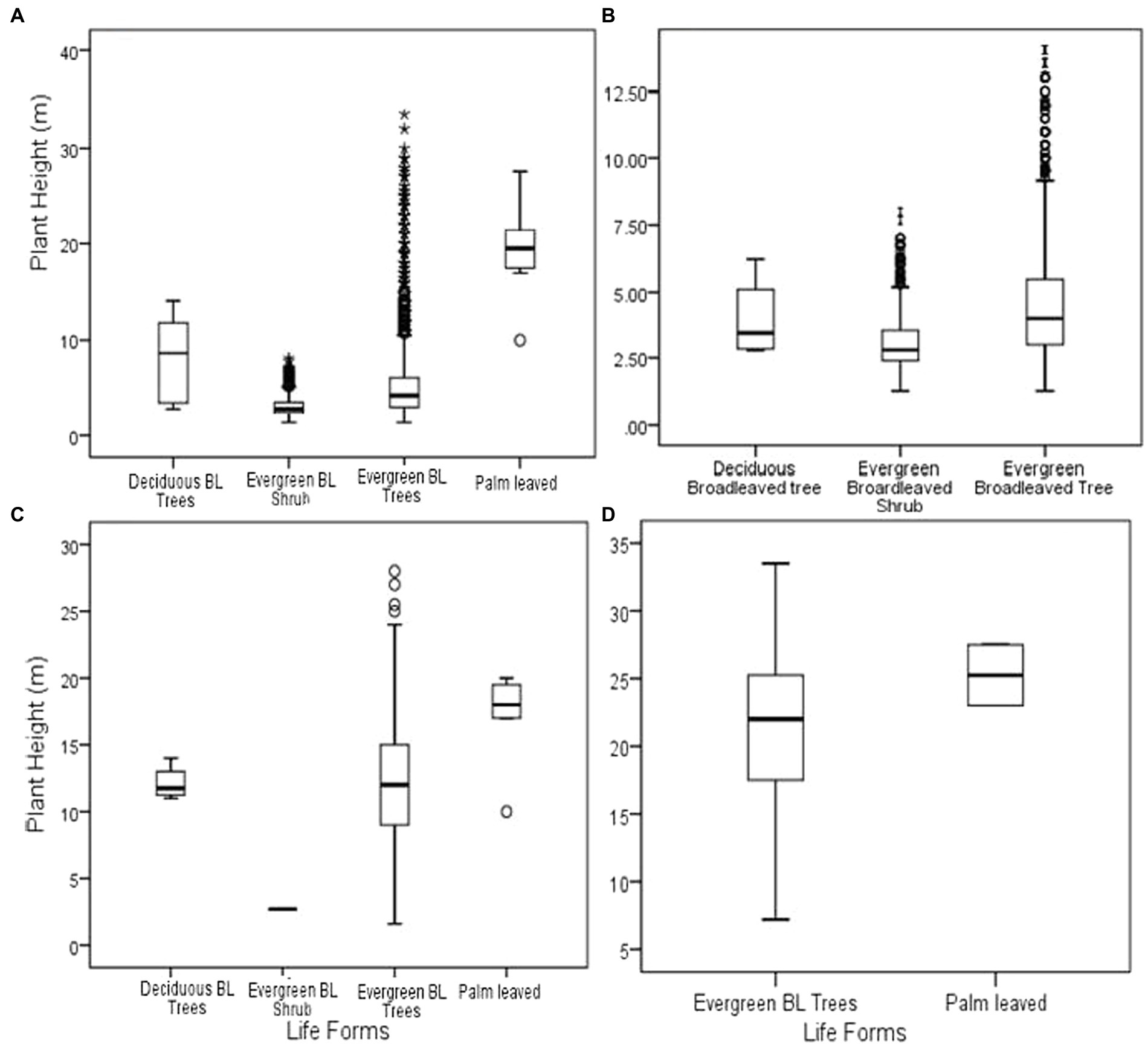
Figure 2. Tree height for the entire forest (A); less than 10 cm DBH (B), 10–30 cm DBH (C), and above 30 cm DBH (D). *BL (Broad leaved).
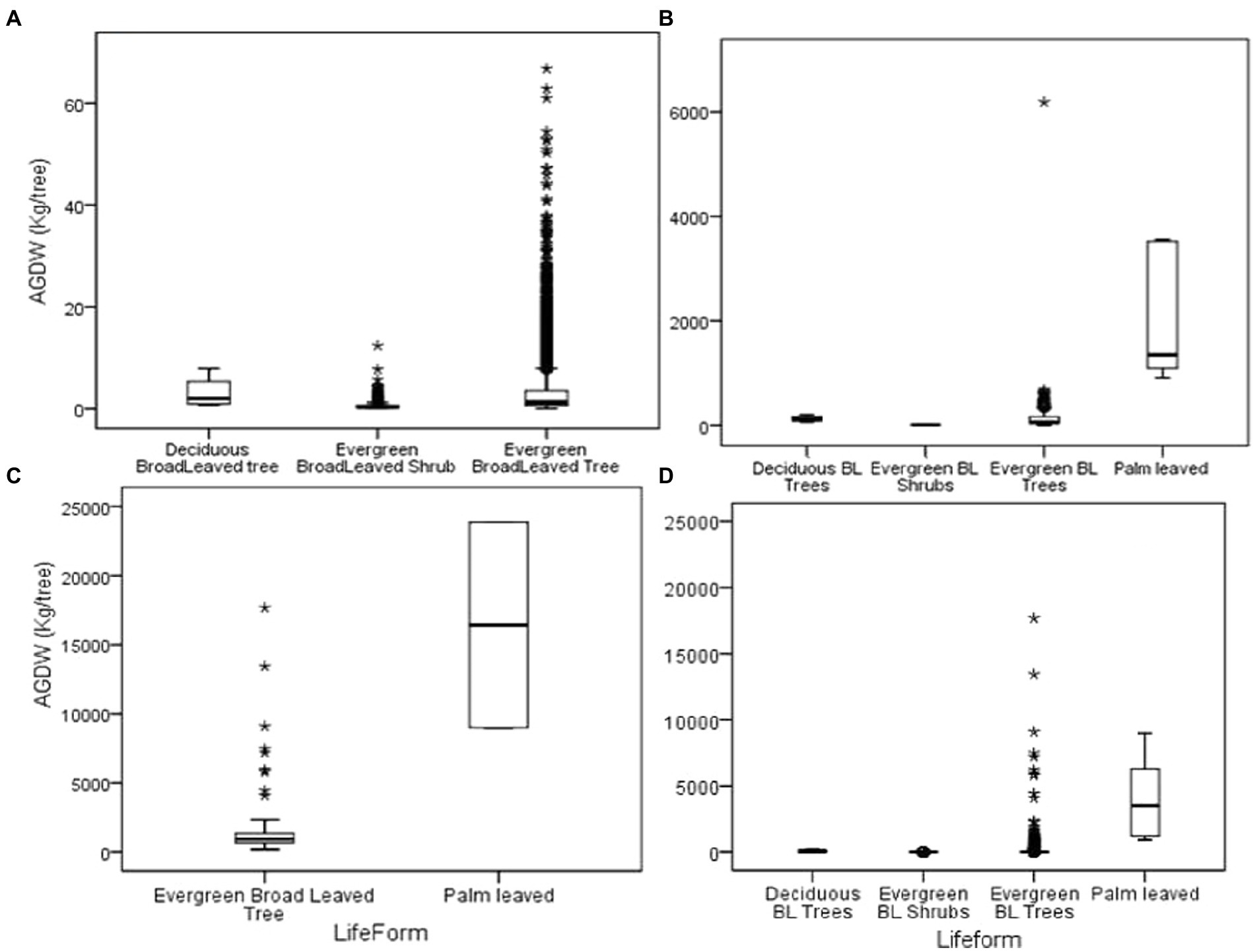
Figure 3. Above-ground dry weight (ABDW) for Dinghushan forest: (A) below 10 cm DBH; (B) 10– 30 cm DBH; (C) above 30 cm DBH and (D) the entire forest data.
Similar to stand height, the mean forest DBH was 3.76 ± 6.26 cm but this was contributed differently by different life forms. Palm-leaved trees had large trunks with DBH of 25.8 ± 8 cm followed by deciduous broadleaved trees (8.97 ± 6.88 cm), evergreen broadleaved trees (4.13 ± 6.73 cm), and the smallest trees being evergreen broadleaved shrubs (1.75 ± 0.73 cm) Figure 4A.
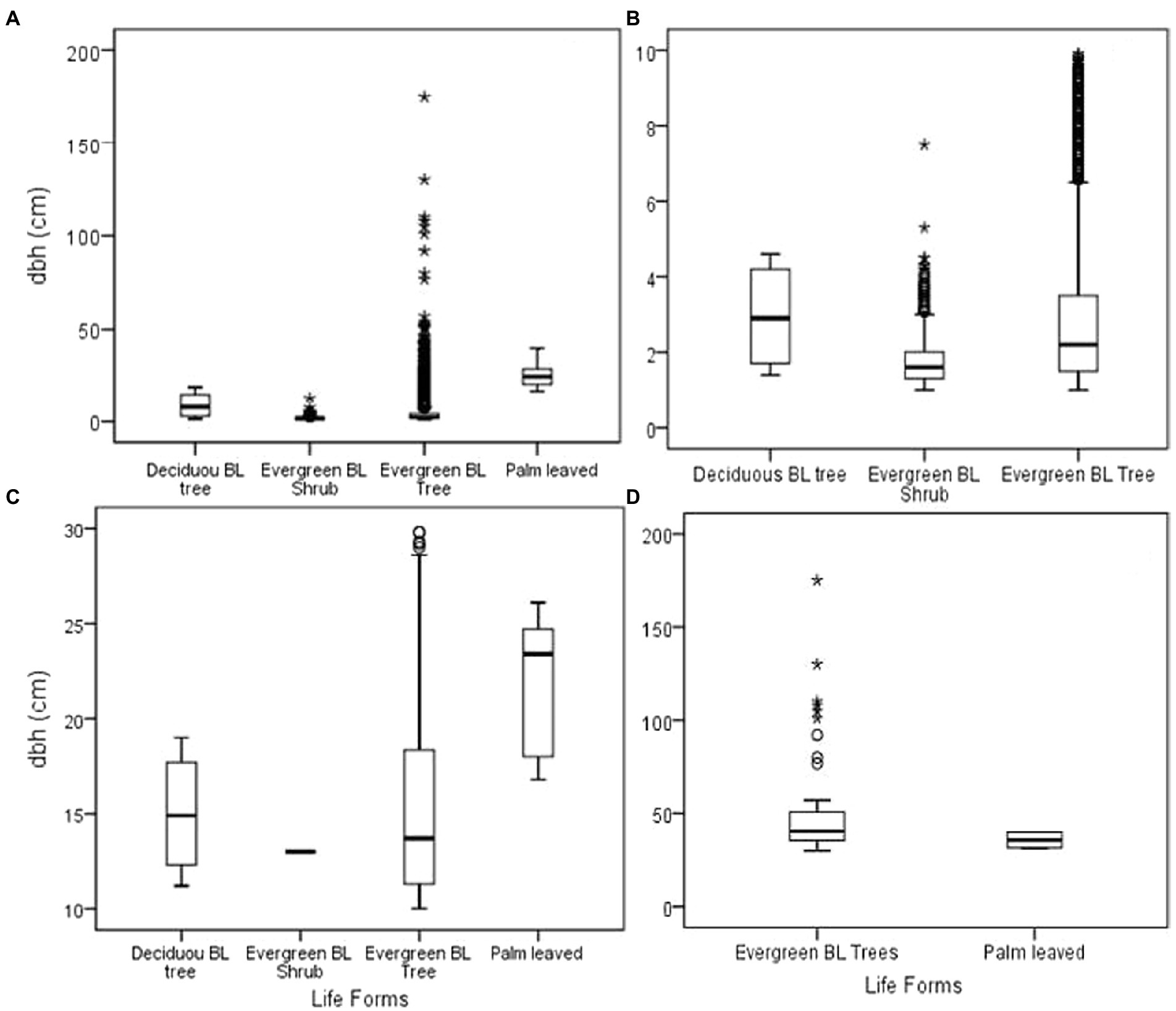
Figure 4. Diameter at breast height (cm) for the entire forest (A); less than 10 cm DBH (B), 10–30 cm DBH (C), and above 30 cm DBH (D). *BL (Broad leaved).
4.3. Forest biomass
Biomass varied depending on the life form whereas the highest biomass was recorded in broad- leaved evergreen forest and the least in deciduous broad-leaved evergreen trees. Looking at the entire above-ground dry weight for the forest, 63.91 kg/tree was recorded. However, forest life forms contributed differently to this biomass where palm-leaved trees recorded the highest above ground (6180.46 kg/tree) followed by deciduous broadleaved trees (63.91 ± 26.43 kg/tree), evergreen broadleaved trees (33.36 kg/tree) and evergreen broadleaved shrub had the least above ground biomass (0.56 kg/tree; Figure 3).
Below-ground dry weight for the Dinghushan forest was lower than the above-ground biomass. The palm-leaved type had the highest biomass of 1827.02 ± 24.82 kg/tree. Deciduous broad-leaved trees recoded 8.89 ± 3.52 kg/tree while evergreen broad-leaved shrubs had the lowest below- ground biomass of 0.11 ± 2.2 kg /tree. Grouping datasets based on DBH size class revealed that only broad- leaved evergreen and palm tree type had below-ground biomass of >30 cm DBH (Figure 5).
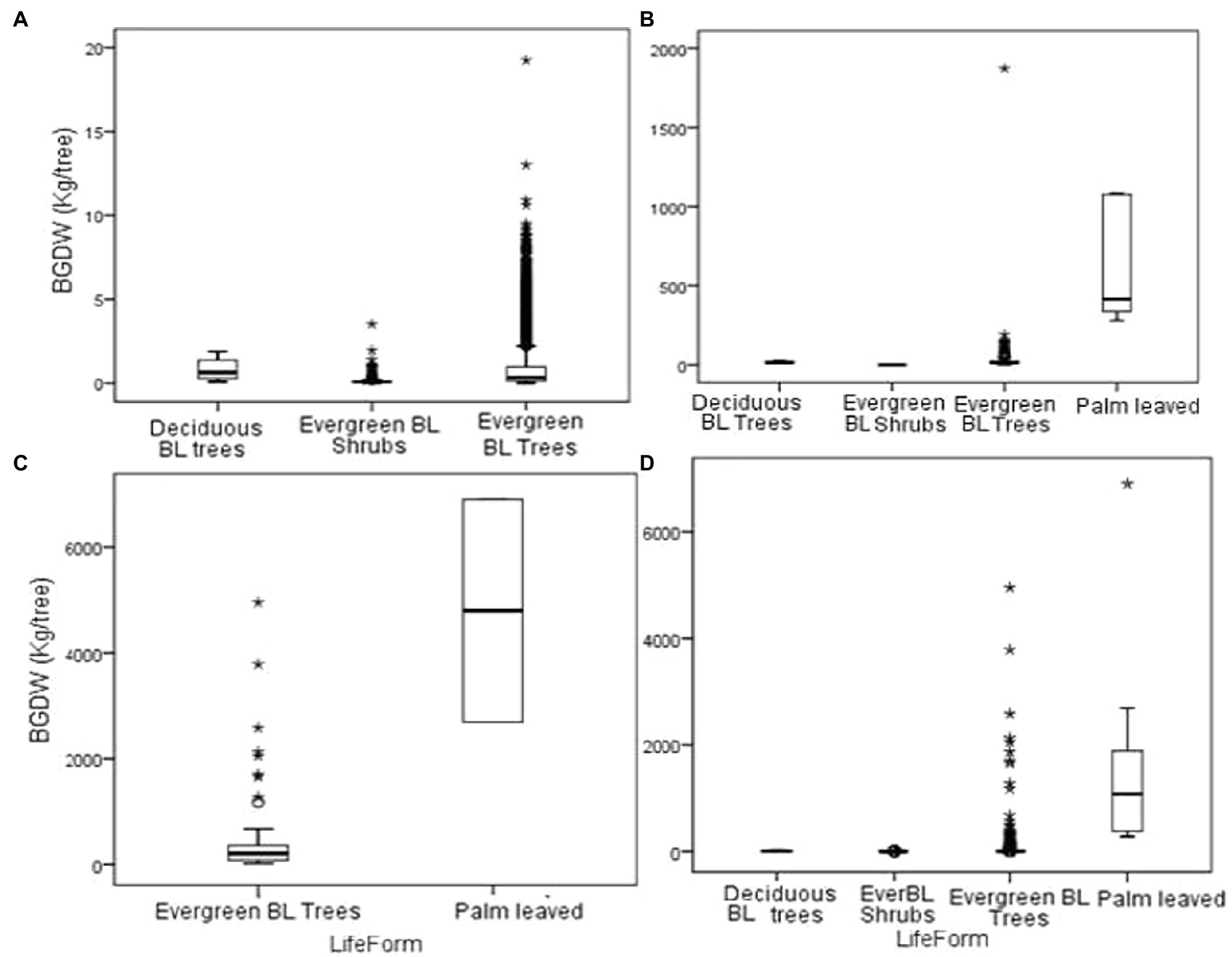
Figure 5. Below ground dry weight (ABDW) for Dinghushan forest: (A) below 10 cm DBH; (B) 10– 30 cm DBH; (C) above 30 cm DBH and (D) the entire forest data. *BL (Broad leaved).
Based on Table 2, generalizing biomass for the entire forest underestimates the contribution of each life form in the Dinghushan forest. Total above-ground biomass was 34.19 ± 5.75Kg/tree but varied results are obtained when the forest is stratified based on DBH. Trees with DBH above 30 cm contributed an average of 2400.54 ± 510.4 kg/tree while trees with DBH 20–30 contributed 171.61 ± 25.06 kg/tree and the least was below 10 cm DBH which contributed 3.013 ± 0.07 kg/tree. More than 50% of the total above-ground mass was mainly contributed by trunk dry weight regardless of the size of the tree. The leaves contributed less than 10.3% to the total above- ground biomass whereas large-diameter trees had only a 7.3% contribution from leaves. Similarly, below-ground biomass was highest in trees with more than 30 cm DBH (632.23 ± 148.68 kg/tree). The overall forest below-ground biomass was 8.66 ± 1.64 kg/tree which was higher than trees with less than 10 cm DBH (0.77 ± 0.016 kg/tree).
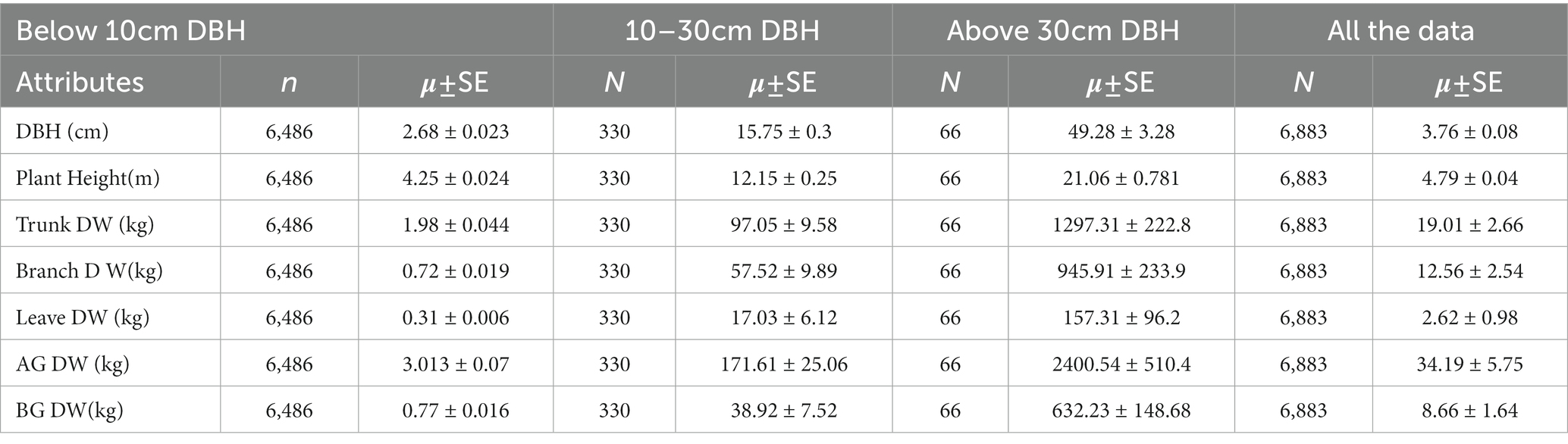
Table 2. Forest attributes for the entire data stratified based on DBH.
4.4. Relationships between biophysical characteristics and biomass
This study found variability in the relationship between DBH and tree height depending on class size clustering (Figure 6). Regardless of tree size (DBH), DBH was positively correlated with tree heights whereas below 10 cm DBH was highly correlated (R2 = 0.63) compared to large tree size which had an R2 of 0.102. Pooled data without clustering showed a highly positive relationship (R2 = 0.68).
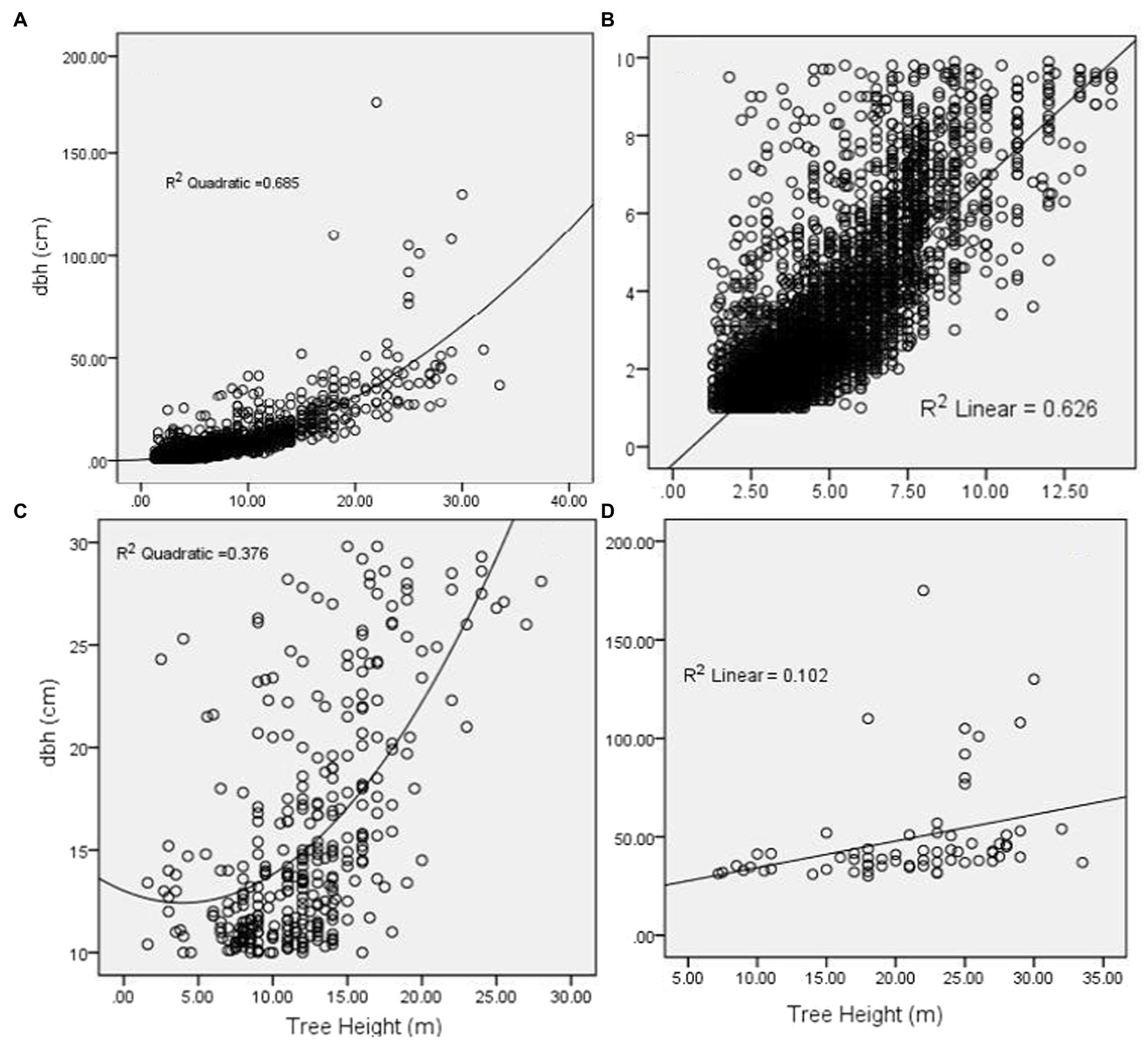
Figure 6. A quadratic and linear relationship between DBH and height for pooled data (A), DBH below 10 cm (B), DBH between 10 and 30 cm (C), and DBH above 30 cm (D).
Diameter at breast height (DBH) showed a significant relationship with above-ground biomass (R2 = 0.7) for small-sized trees which reduced with the increase in tree size (Figure 7). 10–30 cm tree sizes exhibited the weakest relationship (R2 = 0.18) between DBH and biomass. Considering the overall relationship without size class clustering could overestimate the relationship for the 10–30 cm diameter trees by 45% and underestimate for the small-sized trees (less than 10 cm DBH) by 57%.
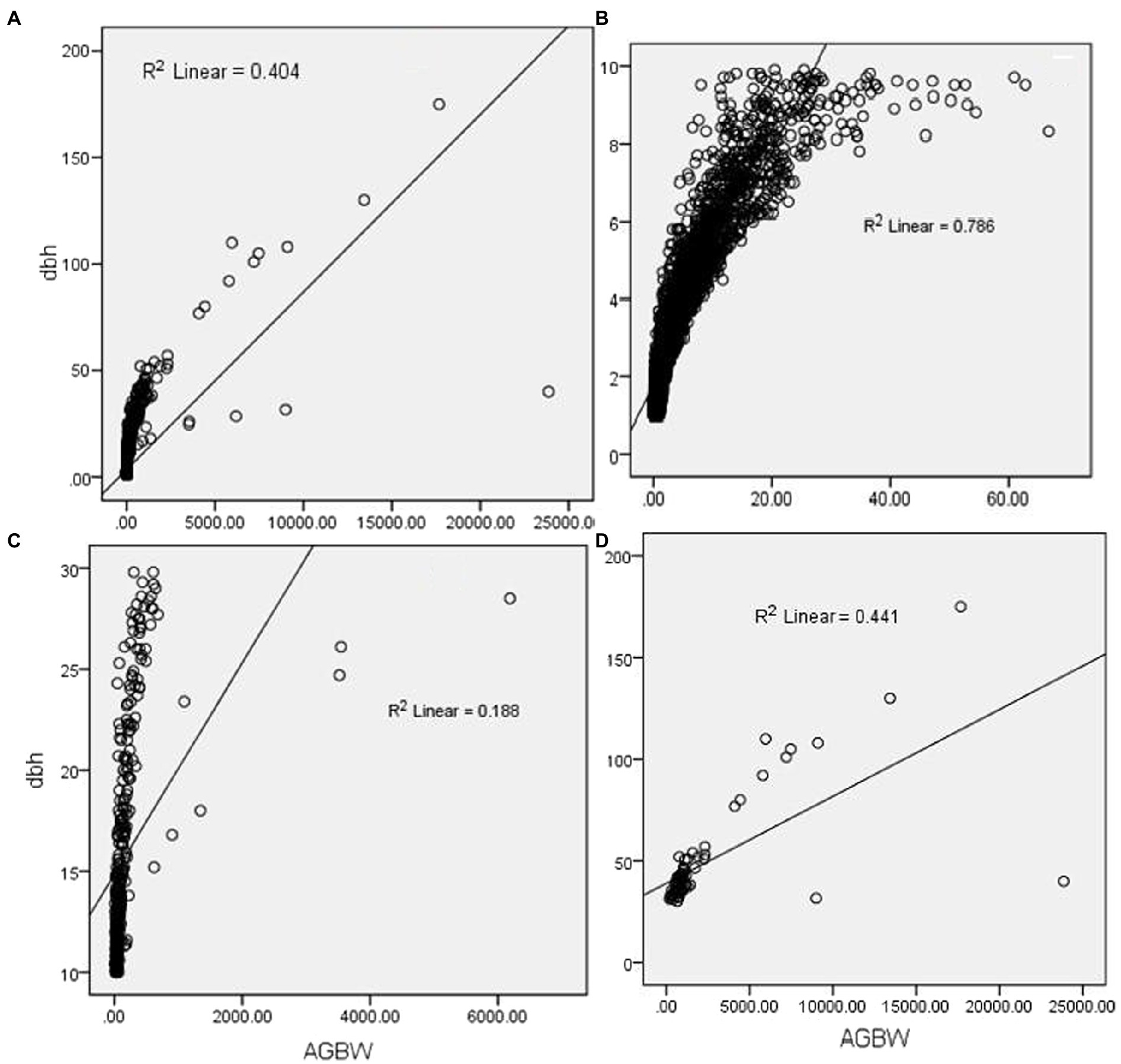
Figure 7. Relationship between DBH and above-ground dry weight; (A) (overall dataset); (B) (less than 10 cm DBH dataset); (C) (10–30 cm DBH dataset) and (D) (above 30 cm DBH dataset).
5. Discussion
Forests are always changing as they adapt to variations in climatic fluctuations by changing their structure and composition (Weiner and Thomas, 1986; Skovsgaard and Vanclay, 2013). In the current study, large trees (with DBH >30 cm) stored large amounts of biomass as observed from below-ground and above-ground biomass. They may have significant controls over the Dinghushan forest carbon store due to the dynamics and sensitivity of biomass to environmental change. Previous studies have shown that large-diameter trees constitute roughly half of the mature forest biomass worldwide (Lutz et al., 2018). According to previous research, big trees may be more susceptible to climate change, ultimately diminishing forest biomass storage (KoÈh et al., 2017; Lutz et al., 2018). In natural forests, tree size and stocking variation could be seen at any time, especially over varied terrain (Weiner and Thomas, 1986), similar to our study. This was true for the Dinghushan forest, where variability in tree height and diameter at breast height (DBH) produced varied biomass for different life forms. This study also showed that due to this temporal and spatial variability, when various life forms occur in a forest and individual species are of different size classes, it might be challenging to acquire reliable estimations of forest carbon stocks and biomass accumulation (Skovsgaard and Vanclay, 2013).
Previous studies have indicated that large-diameter forest trees (≥ 60 cm DBH) can account for up to 41% of above-ground biomass (AGB) globally (Lutz et al., 2018). In the current study site, big- sized trees (≥30 cm DBH) contributed 93.24% of the total AGB if we considered contribution based on DBH size class. Large-sized DBH trees had tall, broad, well-lit crowns that increased the primary productivity of forests and allowed them to store more carbon. This was different when we compared the contribution of evergreen broadleaved shrubs. The evergreen broadleaved shrubs were shorter in height (3.06 ± 0.99 m) than palm-leaved life forms (19.29 ± 5.39 m). Studies have demonstrated that the growth and biomass accumulation of trees are subject to year-to-year and long-term fluctuations (KoÈh et al., 2017), with the majority of the carbon being accumulated at older ages, even though we did not take the age of the trees into account. Therefore, it is assumed that the large-sized trees in our study accumulated more carbon and recorded higher biomass.
This study revealed that the Dinghushan forest is constantly changing in size and composition over time as the trees develop and react to climatic fluctuations. When growing in a dense forest, small- diameter trees have been found to take advantage of canopy gaps and concentrate biomass production through height growth rather than diameter expansion until they reach the high crown layers (McGregor et al., 2021). Tree DBH and height relationship are important elements describing attributes of forest stands. Although stand height has been applied as a variable in volume and biomass models (Comley and McGuinness, 2005) and DBH with height relationships used to determine volume and biomass estimates (Gobakken et al., 2008), such relationships may not provide accurate information regarding the entire forest if datasets are not grouped based on life forms or size class for the heterogeneous forest like Dinghushan forest. Furthermore, generalizing the entire dataset without grouping based on DBH, as observed in our study, may underestimate the forest biomass for less than 30 cm DBH trees and overestimate for trees with more than 30 cm DBH. It is worth noting that large-diameter trees represented fewer species in the entire forest (low stem density). However, they exhibited tall trees with more biomass in the trunk, leaves, branches, and below and above-ground dry weights. The dry weight of the Dinghushan forest is a complex trait that combines a variety of functional processes and structural characteristics. These attributes are linked to individual tree height, forest bio-productivity, and stem density (Shen et al., 2013). Since long-term forest biomass is driven by a balance between the rates at which wood growth and mortality rates, the observed differences in biomass between different life forms in this study emphasize the importance of understanding forest carbon budgets separately based on growth forms. Furthermore, biomass estimates should be based on carbon allocation ratios to foliage, branches, roots, and woody tissues, as described in (Sitch et al. (2008).
The results were more life-form specific than the others, and as a result, the weights recorded underestimated the true below-ground biomass of the larger trees. This resulted in a poor relationship between DBH and below-ground biomass. Trees with larger DBH had the most expansive root system for the life forms studied. The observed linear relationship between above- ground and DBH, and height and DBH indicated a close linkage between stand-level productivity and biomass across this forest complex. The differences in biomass between different DBH classes for different life forms reflect the differences in species composition, tree density, and basal area. Although the tree density was higher in species with DBH less than 10 cm, the biomass was lower than in species with larger DBH >10 cm. This is due to lower height and hence less accumulation of biomass. This was evident where large-sized trees recorded higher biomass accumulation and hence C gain.
It’s possible that environmental and biological heterogeneity within the forest will make it difficult to generalize findings from one location to another. The results of this study occasionally differed significantly from those of Ong et al. (2004), where high tree density (10 cm DBH) contributed the least to below-ground and above-ground biomass. As a result of the lifetime of the main tree species, which provides outstanding contributions to their spectacular heights and vast biomass, studies have demonstrated that a few temperate rainforest locations have excessive above-ground biomass compared to their yearly net productivity (KoÈh et al., 2017). Dinghushan forest complex is a subtropical monsoon forest; trees have a more open canopy than rain forests, creating a dense, closed forest floor or a sub- tropical forest. Therefore, the high biomass accumulation in large-sized trees may be driven by their higher carbon use efficiency; hence large fractions of their photosynthetic products are used for accumulating biomass. Though tropical forests are often more productive, this may conflict with the fact that they utilize less carbon (DeLucia et al., 2007). The typical biomass turnover and mortality rates in tropical forests can be rather high, which prevents the buildup of living biomass (Maina and Li, 2021). Previous studies have indicated an above-ground biomass in the inland and coastal tropical dry evergreen forests of peninsular India to be estimated at 39.69 to 170.02 Mg/ha (Mani and Parthasarathy, 2017). This values were way too low if we compared our data where aboveground biomass ranged between 3.013 ± 0.07 kg/Tree and 2400.54 ± 510.4 kg/Tree depending on DBH class. We must emphasize that the natural disturbance in this forest such as the 2018 typhoon destroyed the forest and affected its structure and functioning based on comparative data by Li et al. (2021). Although other studies have revealed the commercial role of biomass (Mani and Parthasarathy, 2017), the obtained biomass estimates from Dinghushan nature reserve is scientifically relevant for understanding the entire of ecosystem health and productivity, nutrient flow and energy balance. Our findings are important for assessing the contribution of each life form to the entire forest role in global carbon cycle. With combined effects of climate change, different life forms may respond differently to disturbances and this is an area that should be researched across succession stages in the nature reserve.
Furthermore, generalized forest biomass underestimates the contribution of each life form to the total forest carbon stock. Nevertheless, apart from the life forms with less than 30 cm DBH, our study site still has on average the greatest biomass stocks. Most earlier research (Comley and McGuinness, 2005) concentrated only on the above-ground biomass C stocks in tree species, neglecting to take into account the contributions made by C stocks of other life forms like shrubs, herbs, or even roots (below ground), as well as small diameter < 10 cm DBH tree individuals in tropical forest ecosystems. Our focus on assessing the C stocks and their allocation in different structural components between life forms provides a new front in understanding the general components and their contribution to the overall biomass accumulation for the entire ecosystem.
6. Conclusion
This is the first study to cluster the Dinghushan forest based on tree size class. Biomass (C stocks) is influenced by structural attributes (DBH, height) and diversity attributes (life forms). The dynamics of individual tree collectives can be used to explain why the observed forest bio- productivity dynamics occur over such vast areas. High biomass carbon stocks cause size distributions to shift in favor of trees with bigger DBH. The Dinghushan forest complex’s C storage will be improved by maintaining big-sized trees and stand structures and continuing to preserve biodiversity. Although this study found a positive relationship between DBH and stand height (h), physical measurements of these parameters are time-consuming. Due to optical interference from nearby trees or rounded crown forms, it might be challenging to acquire a clear view of the treetop. Thus, measurement costs can be decreased while maintaining an acceptable degree of H accuracy by creating predictive models of H from DBH. The results help to highlight the regions in action plans that require additional attention for conservation and C stock improvement. We suggest managing forests to preserve existing big-diameter trees or those that may soon achieve large diameters as a straightforward method to preserve and possibly improve ecosystem services. Our study on biomass in four life forms in a forest plot ws based on data obtained after the typhoon destruction. These possess a limitation on wider application of our findings. Additionally, we did not consider biomass accumulation in relation to environmental variables within the nature reserve. Therefore, future studies could consider spatio-temporal variation in biomass in relation to environmental conditions.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
YL and MN analyzed data and writing. LD and FH data process and writing. SL data collection and vegetation investigation. BM data process and writing. QZ data process. GC data collection. DZ experimental design. JL data process and writing support. ZM site maintenance. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the National Natural Science Foundation of China (Grant No. 31961143023); the National Key Research and Development Program of China (Grant No. 2021YFF0703905); the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant No. XDA23080302); the National Science and Technology Basic Work Project (Grant No. 2015FY1103002); the Dinghushan Forest Ecosystem Positioning Research Station of the National Science and Technology Infrastructure Platform, the Chinese Ecosystem Research Network (CERN); and the Operation Service Project of the National Scientific Observation and Research Field Station of the Dinghushan Forest Ecosystem of Guangdong, the Ministry of Science and Technology of the People’s Republic of China.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1118175/full#supplementary-material
References
Comley, B. W. T., and McGuinness, K. A. (2005). Above- and below-ground biomass, and allometry, of four common northern Australian mangroves. Aust. J. Bot. 53, 431–436. doi: 10.1071/BT04162
Clough, B. F., Dixon, P., and Dalhaus, O. (1997). Allometric relationships for estimating biomass in multi-stemmed mangrove trees. Australian Journal of Botany 45, 1023–1031. doi: 10.1071/BT96075
DeLucia, E. H., Drake, J., Thomas, R. B., and Gonzalez-Meler, M. A. (2007). Forest carbon use efficiency: is respiration a constant fraction of gross primary production?. Global Change Biology 13, 1157–1167.
Gielen, B., and Ceulemans, R. (2001). The likely impact of rising atmospheric CO2 on natural and managed Populus: a literature review. Environ. Pollut. 115, 335–358. doi: 10.1016/S0269-7491(01)00226-3
Gobakken, T., Lexerød, N. L., and Eid, T. (2008). A forest simulator for bioeconomic analyses based on models for individual trees. Scand. J. For. Res. 23, 250–265.
Guo, Q. (1999). Ecological comparisons between eastern Asia and North America: historical and geographical perspectives. J. Biogeogr. 26, 199–206. doi: 10.1046/j.1365-2699.1999.00290.x
Husch, B., Beers, T. W., and Kershaw Jr., J. A. (2002). Forest Mensuration. 4th edn. USA: John Wiley & Sons.
Kirui, B., Kairo, J. G., and Karachi, M. (2006). Allometric equations for estimating above ground biomassof Rhizophora mucronata Lamk. (Rhizophoraceae) mangroves at Gazi Bay, Kenya. Western Indian Ocean. J. Mar. Sci. 5, 27–34. doi: 10.4314/wiojms.v5i1.28496
KoÈh, M., Neupane, P. R., and Lotfiomran, N. (2017). The impact of tree age on biomass growth and carbon accumulation capacity: a retrospective analysis using tree ring data of three tropical tree species grown in natural forests of Suriname. PLoS One 12:e0181187. doi: 10.1371/journal.pone.0181187
Li, Y., Mwangi, B., Zhou, S., Liu, S., Zhang, Q., Liu, J., et al. (2021). Effects of typhoon Mangkhut on a monsoon Evergreen broad-leaved Forest Community in Dinghushan nature reserve, lower subtropical China. Front. Ecol. Evol. 9:692155. doi: 10.3389/fevo.2021.692155
Luo, T., Zhang, L., Zhu, H., Daly, C., Li, M., and Luo, J. (2009). Correlations between net primary productivity and foliar carbon isotope ratio across a Tibetan ecosystem transect. Ecography 32, 526–538. doi: 10.1111/j.1600-0587.2008.05735.x
Lutz, J. A., Furniss, T. J., Johnson, D. J., Davies, S. J., Allen, D., Alonso, A., et al. (2018). Global importance of large-diameter trees. Glob. Ecol. Biogeogr. 27, 849–864. doi: 10.1111/geb.12747
Maina, J. N., and Li, Y. (2021). Spatio-temporal variation in species diversity between plantation and secondary Forest of Kakamega tropical rain Forest in Kenya. Ecol. Eng. Environ. Technol. 5, 1–11. doi: 10.12912/27197050/139410
Mani, S., and Parthasarathy, N. (2017). Above-ground biomass estimation in ten tropical dry evergreen forest sites of peninsular India. Biomass Bioenergy 31, 284–290. doi: 10.1016/j.biombioe.2006.08.006
McGregor, I. R., Helcoski, R., Kunert, N., Tepley, A. J., Gonzalez-Akre, E. B., Herrmann, V., et al. (2021). Tree height and leaf drought tolerance traits shape growth responses across droughts in a temperate broadleaf forest. New Phytol. 231, 601–616. doi: 10.1111/nph.16996
Murali, K. S., Bhat, D. M., and Ravindranath, N. H. (2005). Biomass estimation equation for tropical deciduous and evergreen forests. Int. J. Agric. Resour. Gov. Ecol. 4, 81–92. doi: 10.1504/IJARGE.2005.006440
Ong, J. E., Gong, W. K., and Wong, C. H. (2004). Allometry and partitioning of mangrove, Rhizophora apiculata. For. Ecol. Manag. 188, 395–408. doi: 10.1016/j.foreco.2003.08.002
Shaver, G. R., Canadell, J., Chapin, F. S. III, Gurevitch, J., Harte, J., Henry, G., et al. (2000). Global warming and terrestrial ecosystems: a conceptual framework for analysis. Bioscience 50, 871–882. doi: 10.1641/0006-3568(2000)050[0871:GWATEA]2.0.CO;2
Shen, Y., Santiago, L. S., Ma, L., Guo-Jun, L., Ju-Yu, L., Hong-Lin, C., et al. (2013). Forest dynamics of a subtropical monsoon forest in Dinghushan, China: recruitment, mortality and the pace of community change. J. Trop. Ecol. 29, 131–145. doi: 10.1017/S0266467413000059
Sitch, S., Huntingford, C., Gedney, N., Levy, P. E., Lomas, M., Piao, S. L., et al. (2008). Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Global change biology. 14, 2015–2039. doi: 10.1111/j.1365-2486.2008.01626.x
Skovsgaard, J. P., and Vanclay, J. K. (2013). Forest site productivity: a review of spatial and temporal variability in natural site conditions. Forestry 86, 305–315. doi: 10.1093/forestry/cpt010
Weiner, J., and Thomas, S. C. (1986). Size variability and competition in plant monocultures. Oikos 47, 211–222. doi: 10.2307/3566048
Wu, X., Wang, X., Tang, Z., Shen, Z., Zheng, C., Xia, X., et al. (2015). The relationship between species richness and biomass changes from boreal to subtropical forests in China. Ecography 38:2015. doi: 10.1111/ecog.00940
Zhang, Q. (2011). China Ecosystem Positioning Observation and Research Data set Forest Ecosystem Volume Guangdong Dinghushan Station: 1998–2008. Beijing: China Agricultural Publishing House.
Zhou, G., Houlton, B. Z., Wang, W., Huang, W., Xiao, Y., Zhang, Q., et al. (2014). Substantial reorganization of China's tropical and subtropical forests: based on the permanent plots. Glob. Chang. Biol. 20, 240–250. doi: 10.1111/gcb.12385
Zhou, G. Y., Liu, S. G., Li, Z., Zhang, D. Q., Tang, X. L., Zhou, C. Y., et al. (2006). Old- growth forests can accumulate carbon in soils. Science 314:1417. doi: 10.1126/science.1130168
Zhou, G., Peng, C., Li, Y., Liu, S., Zhang, Q., Tang, X., et al. (2013). A climate change- induced threat to the ecological resilience of a subtropical monsoon evergreen broad- leaved forest in southern China. Glob. Chang. Biol. 19, 1197–1210. doi: 10.1111/gcb.12128
Keywords: bio-productivity, forest biomass, life forms, diameter at breast height, broadleaved
Citation: Li Y, Nyongesah MJ, Deng L, Haider FU, Liu S, Mwangi BN, Zhang Q, Chu G, Zhang D, Liu J and Meng Z (2023) Clustered tree size analysis of bio-productivity of Dinghushan National Nature Reserve in China. Front. Ecol. Evol. 11:1118175. doi: 10.3389/fevo.2023.1118175
Edited by:
Hui Zhang, Hainan University, ChinaCopyright © 2023 Li, Nyongesah, Deng, Haider, Liu, Mwangi, Zhang, Chu, Zhang, Liu and Meng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuelin Li, ✉ eXVlbGluQHNjYmcuYWMuY24=
†These authors have contributed equally to this work