 Johan T. du Toit1,2*
Johan T. du Toit1,2*- 1Department of Wildland Resources, Utah State University, Logan, UT, United States
- 2Mammal Research Institute, Department of Zoology and Entomology, University of Pretoria, Pretoria, South Africa
If transparent and inclusive stakeholder discussion delivers a consensus for active rewilding, then five steps are recommended for operationalizing that decision, focused initially on the large herbivore assemblage. Consideration of large predators could follow, contingent upon the establishment of prey populations. First, determine the potential biomass density (kg/km2) of large mammalian herbivores in the target landscape. Regression models based on rainfall or primary productivity are helpful if applicable, otherwise comparative studies are needed. Second, use empirical data from reference ecosystems to apportion biomass density among functional types, crudely defined by body size and feeding type (grazer, browser, mixed feeder). Third, identify specific functional traits (coarse grazing, endozoochory, etc.) of particular local importance. Fourth, identify species within each functional type that are already present, estimate their potential biomass densities, and thus identify the shortfall within each cell of the body size x feeding type matrix. A candidate set of native and non-native (surrogate) species is then identified to make up the shortfalls. This is followed by an iterative process of estimating equilibrium population sizes, stakeholder acceptance, and viability of each potential population. Fifth, stakeholders must be inclusively re-engaged to visualize the potential assemblage, its expected functional interactions, the ecosystem services to be delivered, and the long-term costs (including opportunity costs) and benefits. When a plan is supported, local stakeholders should be integrated as active participants in the implementation, monitoring, and championing of their rewilding project.
1. Introduction
One of the greatest challenges facing humanity in the Anthropocene is the diminishment of ecosystem services. Addressing the challenge involves two thrusts: first, the alleviation of anthropogenic forcing on natural systems; second, the repair of ecosystem function. The first is replete with wicked problems for the global populace and so my focus here is on the second, which for decades has been the domain of restoration ecology. However, it is increasingly apparent that, despite the best efforts of restoration practitioners, their successes are hard-won and then chronically vulnerable to back-sliding. This is because any benchmark state that existed prior to a transformational disturbance is subsequently elusive due to a set of quaintly-named effects in ecosystem dynamics. One is the Humpty-Dumpty effect, meaning that after a complex system has fragmented, it cannot be reassembled in the way it once was because the parts (predators, prey, habitats, etc.) have undergone quantitative and qualitative change (Pimm, 1991; Evans et al., 2022). Another is the hysteresis effect, which arises because feedbacks among processes within the system have changed, causing the “return” trajectory to be different from the “departure” (Beisner et al., 2003). Then, the Red Queen effect (Van Valen, 1973) can apply at the ecosystem level, meaning that because the environment is always changing the system must continually reorganize and adapt if it is to persist within state thresholds.
I suggest it is pragmatic to find ways of working with, rather than against, the above effects especially where stakeholder-desired outcomes are dependent on ecosystem function. Identify what ecosystem services are required from an ecosystem, which ecosystem processes generate them, and which functional types of organism maintain those processes, before choosing which species to invest conservation resources in. This is where the rewilding concept (Pettorelli et al., 2019) is making advances, with (depending on one’s definition of choice) its focus on functional type composition and acceptance of continual change and reorganization. To operationalize the concept for an ecologically degraded landscape, it is first vital to initiate a facilitated process of fully-inclusive stakeholder engagement. This is to reach consensus on which of three options is best aligned with the stakeholders’ priorities: (1) restoring historical species composition; (2) enabling some future state of undefined wildness to evolve; or (3) repairing ecosystem function. A decision tree can be used to move forward with each of those options (du Toit and Pettorelli, 2019) but here I focus on the case of ecosystem function being the priority for an area where the stakeholder consensus is for active rewilding (Option 3). Also, it is assumed that the project area is degraded to the extent that the large mammal community is diminished, with large predators being absent or rare. Then, recognizing the impossibility of identifying all the elements and interactions involved in an ecosystem’s function, a start can be made with the functional types represented by large herbivores (>5 kg body mass). A natural assemblage of syntopic large herbivores typically includes a relatively modest number of species (seldom >10) that can interactively impose strong top-down modifying effects on vegetation structure and composition, and on biogeochemical cycling (Vynne et al., 2022). Large herbivores are also visible and relatable agents of natural processes, thus facilitating a bond between stakeholders and their rewilding project. Developing an appropriate assemblage should happen within an adaptive co-management framework and with a social license to operate, so that practitioners and stakeholders may establish a trusting relationship (Butler et al., 2021). Concurrently, the technical team needs to follow a logical process to estimate how many of which large herbivore species would be required for the rewilding project, as suggested–with due recognition of knowledge gaps–in the steps that follow.
2. Steps for assembling a functionally diverse community of large herbivores
2.1. Estimate total potential biomass density
Macroecological meta-analyses have confirmed that in the absence of major anthropogenic distortions, the biomass density of an intact large herbivore community is generally expected to vary as a function of primary productivity, which in turn varies as a function of mean annual rainfall. That expectation currently holds in Africa, whereas on other continents the biomass of indigenous large herbivores is anthropogenically depleted (Fløjgaard et al., 2022). Fortunately, aerial survey data from African wildlife areas date back to the colonial era and enable a predictive understanding of the consumer-producer relationship. Coe et al. (1976) found a linear relationship by which variation in the logarithm of mean annual rainfall explained 77% of variation in the logarithm of large herbivore biomass density across wildlife and pastoral systems in the savannas of eastern and southern Africa. That relationship is, however, strongly influenced by soil nutrient status and 82%–85% of the variation can be explained by separate models for “low” and “medium to high” soil nutrient classes (Fritz and Duncan, 1994). Then, the diluting effect of rainfall means decreasing forage quality in wetter savannas, depressing herbivore biomass density from its peak of ~1,700 kg/km2 (for wildlife excluding elephants, Loxodonta africana) in areas receiving >700 mm of mean annual rainfall (Hempson et al., 2015). Also, on a global scale, potential stocking rates can be estimated from remotely sensed data on net primary production in grasslands (Piipponen et al., 2021), although that underestimates community biomass because browsing is not accounted for.
Overall, evidence confirms that large herbivore biomass is controlled from the bottom up by food availability and mechanistic models can explain the relationship to the extent permitted by available data. Where the predator guild is intact, populations of smaller ungulates (<150 kg) are regulated by predation (Sinclair et al., 2003) but this would not (yet) occur where rewilding projects are being planned. Across ecosystems that are anthropogenically distorted, there is a need for standardized open-access data-bases of equilibrium densities of large mammals in reference areas such as national parks. Such data would not necessarily provide a local benchmark for target conditions in a geographically similar area, but could be used to guide rewilding projects in any area where the state of the ecosystem is forecast to be similar to the current state in the reference area over a timeframe that is meaningful to stakeholders. Meanwhile, rewilding should progress through an adaptive management cycle (Figure 1) for which an estimate of potential biomass density is only initially useful as an upper “ball-park” value to begin planning within.

Figure 1. The process proposed to operationalize a rewilding project. Green boxes are actions performed by practitioners on the land. Yellow boxes are analyses performed in collaboration among practitioners, technical resources, and stakeholder groups, following steps described in the text. Blue diamonds are decisions on community assembly ultimately requiring ratification by stakeholders.
2.2. Allocate biomass to functional types
Having an idea of potential biomass density, even if a ball-park estimate, allows the functional composition of a (yet notional) large herbivore community to be sketched out. As stressed by Bell (1982) in his seminal work on community structure in African savannas, mean annual rainfall and soil nutrient availability interact in ways that govern both the total biomass of large herbivores and the structure of the community (see also East, 1984). Areas of relatively low-quality soil receiving relatively high rainfall have abundant vegetation of relatively low quality to herbivores, because of the increase in structural tissue (lignin, fiber) needed for mechanical support by large plants (trees, bunchgrasses). The larger the individual herbivore, however, the wider its dietary tolerance and the greater its ability to utilize abundant low-quality forage that smaller herbivores cannot survive on. Body size and feeding style thus provide the basis for a first-order classification of functional types: small (<100 kg), medium (100–1,000 kg), large (>1,000 kg); grazers, mixed feeders, browsers (Bell, 1984). Finer categorizations have been proposed (Hempson et al., 2015) but a body size x feeding-style matrix (Figure 2) allows a visualization of how the composition of a large herbivore community can differ substantially between one ecosystem and another.
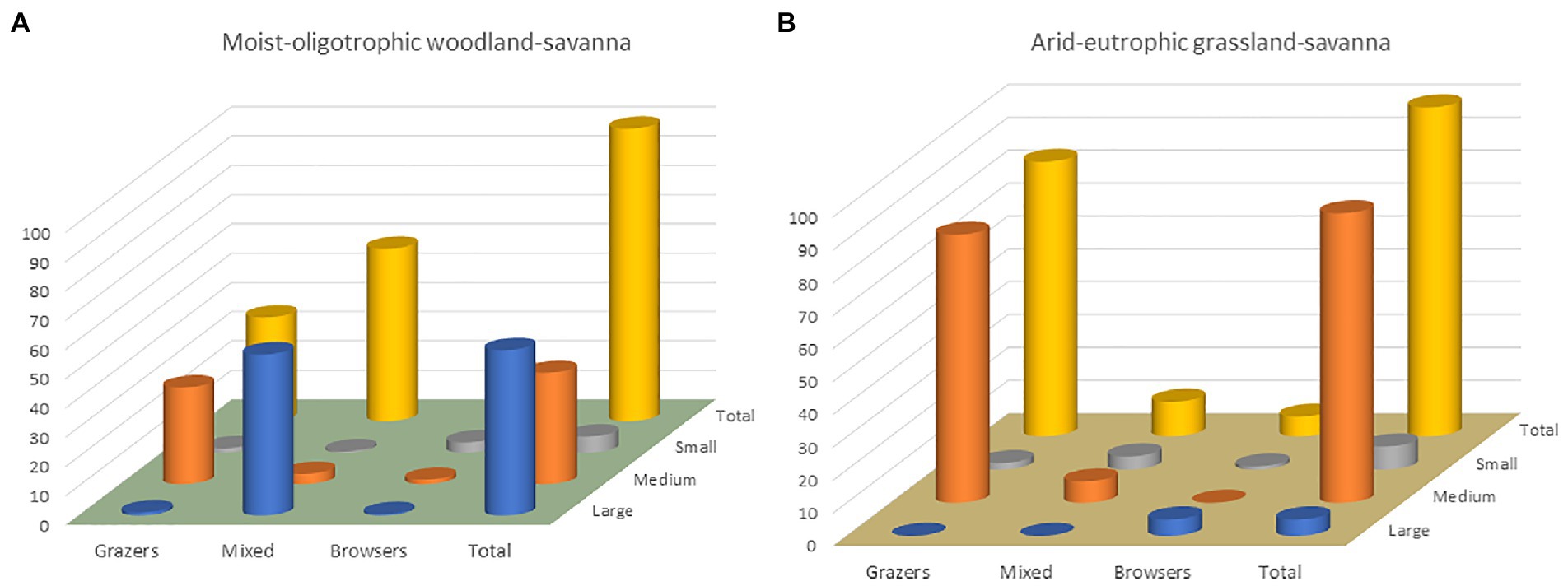
Figure 2. Distributions of large-herbivore community biomass (%) among functional types coarsely defined by feeding style (grazer, mixed feeder, browser) and body size (large, >1,000 kg; medium, 100–1,000 kg; small, <100 kg) in two African savanna ecosystems: (A) a moist-oligotrophic woodland and (B) an arid-eutrophic grassland (data from Bell, 1984).
From historical wildlife survey data collected across African savannas, it is apparent that pure browsers (mainly giraffe, Giraffa spp. and black rhino, Diceros bicornis) generally contribute only ~10% or less to community biomass and across feeding styles the smallest body-size class (<10 kg) seldom represents >10% (Bell, 1984; Figure 2). Nevertheless, the most speciose size-class among African antelopes is 10–32 kg (du Toit and Cumming, 1999) and so the functional trait diversity of any one size class could be inversely related to its community biomass contribution. How generalizable these African patterns are at the global scale can be debated with the limited data on expected “natural” community structure for ecosystems on other continents. Paleoecological reconstructions for particular places at times prior to human disturbance are valuable for understanding species composition. Yet, for the total biomass of all syntopic species and the distribution of community biomass among functional types, the historical African data provide at least a guiding template formed by ecological principles (Bell, 1982) that are universal.
2.3. Prioritize ecosystem services and requisite functional traits
Rewilding projects aim to repair the functional properties of degraded ecosystems but the levels and types of degradation, and consequently the priorities for rewilding, differ between project areas. If a priority is the provision of herbaceous forage in an area encroached with woody vegetation, then requisite functional traits could be browsing, debarking, and seedling predation. If a priority is fire suppression then a requisite functional trait could be coarse grazing; if stream flow regulation then pond building; if revegetation then endozoochory; if soil amelioration then nutrient transport and tilling/rooting/churning; and so on. This step of prioritizing the functions needed from large herbivores sharpens the resolution of the body size x feeding style classification and sets up the next step of identifying species–with their requisite functional traits–to be prioritized by managers.
2.4. Prioritize native species but consider surrogates
After establishing a ballpark estimate of biomass density for the potential large herbivore assemblage in the area designated for rewilding, and then assigning a biomass distribution across the 3 × 3 matrix of body size classes and feeding styles (as in Figure 2), the rewilding planner now has nine “bins” of biomass to work with on paper. Native species can be allocated to each bin as appropriate, including species already in the project area and those that could be reintroduced. Then, species with required functional traits should be prioritized as above, ideally with redundancy because increased species richness in a large herbivore assemblage is associated with increased delivery of multiple ecosystem services (Maestre et al., 2022).
In most cases the megaherbivore (>1,000 kg) biomass bin is likely to remain empty due to extinctions, costs, and potential human-wildlife conflicts, but estimating the size of that bin is important for downsizing the overall biomass estimate. It might also be impossible or impractical to fill some other bins–while prioritizing required functional traits–with native species. In theory, non-native wildlife species could be considered as surrogates but in practice there are multiple legal, social, and economic barriers to that option. Nevertheless, various domestic species (horses, cattle, pigs, goats, sheep, etc.) can immediately be considered as surrogates and stocked together with native species at the densities required to fill those bins needing topping up.
Once all feasible biomass has been penciled in, and each species allocated a share of the biomass in its bin, the potential population of each species can be estimated simply by dividing its biomass by unit mass (⅔ adult female mass). Now, further adjustments can be made to account for population viability within the project area, with particular attention to social and economic viability. If a population is unlikely to be viable then it can be scratched and the population estimate for another species in that bin increased to take up the freed biomass.
2.5. Re-engage stakeholders
By this stage the project manager has an on-paper design of a large-herbivore assemblage that would be ecologically compatible with the area designated for rewilding, and would include the suite of functional traits required for the prioritized ecosystem services. Next, the stakeholders should be re-engaged for a decision-into-practice exercise (Butler et al., 2021). Using the on-paper design, the project manager can present a vision of the rewilding project with estimations on the likelihood of the project achieving its goals, at what costs (implementation, running, and opportunity costs), and over what timeframe. That presentation should include alternative scenarios for dealing with biomass bins that cannot be filled. For example, the missing megaherbivores could be functionally substituted to some extent by machinery (Bocherens, 2018), such as the mulching machines used for mastication of woodlands and shrublands in the western United States (Brennan and Keeley, 2017) where woody encroachment is a multi-billion-dollar problem (Morford et al., 2022). This would clearly infringe on the minimal-intervention ideals of rewilding but pragmatism might have to prevail for intermittent operations. Then, the implications of population regulation must be worked through for scenarios with and without an intact or partially intact predator community. Offtake through regulated hunting can control populations and earn revenue if stakeholders accept the ethical implications. Alternatively, populations can be allowed to self-regulate through density dependence, with die-offs in harsh years (Fløjgaard et al., 2022), requiring full recognition of the implications for animal welfare (Capozzelli et al., 2020).
Through a fully inclusive and equitable process, all stakeholders should be facilitated in their decision-making to reach consensus on whether to support or reject rewilding as a means toward meeting their needs and aspirations for the project area. If supported, then implementation should proceed with local participation in reintroduction operations, citizen-science surveys, field educational exercises, etc., to keep stakeholders engaged in their rewilding project.
3. Discussion
The stepwise, functionally-focused approach proposed here does not describe the ways in which most self-described rewilding projects (reviewed in Pettorelli et al., 2019) come into being. In Europe, the process most commonly involves one keystone species (Segar et al., 2022) after which the project progresses (or not) through learning-by-doing. That was a completely reasonable approach under past circumstances but now, with the rate of global change, the scale of the problem of faltering ecosystem services, and the growing interest in rewilding, there is a mounting need to equip the concept with a comprehensive plan for operationalization. Each case has its idiosyncrasies but the approach suggested here is aimed at the primary goal of establishing the range of ecological functions performed by a large herbivore assemblage in a terrestrial ecosystem. Thereafter, small carnivores will mostly take care of themselves whereas large carnivores, whether already present or not, require special considerations for rewilding (Linnell and Jackson, 2019).
Increasingly, conservation practitioners are having to abandon the ideal of restoring species composition to benchmark conditions and accept the existence of novel ecosystems (Hobbs et al., 2006). Such acceptance comes with the challenge of recognizing the value of the services that novel ecosystems can provide under present and forecast conditions. The rewilding concept holds promise for achieving the best–or perhaps least bad–alternative state that a self-organizing system could stabilize in and sustain the delivery of ecosystem services to its stakeholders. For land managers to visualize that state and articulate it to stakeholders requires tangible operational elements, and large herbivores are both compelling symbols of wildness and effective agents of key ecological processes. Furthermore, ecologically appropriate large-herbivore communities can be designed, assembled, and left to self-organize with minimal ongoing management. The ecological principles were revealed by foundational work in African savannas that took advantage of historical data on intact wildlife communities (Coe et al., 1976; Bell, 1982, 1984). Real-world application can be found in the extensive conversion of cattle ranches to private wildlife reserves and conservancies in southern and eastern Africa, where a rewilding movement of its own has been underway for decades now (Carruthers, 2008; Bothma and du Toit, 2016). There is no reason why the same ecological principles should not apply outside of African savannas where, in general, we can expect large-herbivore equilibrium biomass to vary predictably in response to primary productivity and soil nutrient availability. We can also expect the distribution of total biomass among functional types to differ predictably between ecosystem types.
In closing, a suggestion to facilitate the operationalization of the rewilding concept on a global scale is to develop typologies of functional types of animals and plants within bioregions of trait space (sensu Harris et al., 2022). These could be mapped in space and time by correspondence with latitude, climatic envelope, and soil nutrients. Each bioregion would have a generic (non-taxonomic) “parts list,” which could be assembled (or allowed to self-assemble) in a project area from native species and surrogates as needed. The assembled community would optimally deliver ecosystem services and could be reorganized (or allowed to self-organize) as bioregions shift across the project area due to climate change. While this suggestion is likely heretical to conservation traditionalists, the dire and compounding ecological challenges of the Anthropocene demand non-traditional solutions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
The author confirms being the sole contributor to this work and has approved it for publication.
Funding
The ideas presented in this publication were developed while the author was funded by Utah State University and, previously, the University of Pretoria.
Acknowledgments
This work benefitted from discussions with participants in the Restoration and Rewilding session within Theme 3 of the INTECOL 2022 congress at Geneva, Switzerland.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Beisner, B. E., Haydon, D. T., and Cuddington, K. (2003). Alternative stable states in ecology. Front. Ecol. Environ. 1, 376–382. doi: 10.1890/1540-9295(2003)001[0376:ASSIE]2.0.CO;2
Bell, R. H. V. (1982). “The effect of soil nutrient availability on community structure in African ecosystems” in The ecology of tropical savannas. eds. B. J. Huntley and B. H. Walker (Berlin: Springer-Verlag), 193–216.
Bell, R. H. V. (1984). “Carrying capacity and off-take quotas” in Conservation and wildlife Management in Africa. eds. R. H. V. Bell and E. McShane-Caluzi (Washington, DC: US Peace Corps), 146–181.
Bocherens, H. (2018). The rise of the Anthroposphere since 50,000 years: an ecological replacement of megaherbivores by humans in terrestrial ecosystems? Front. Ecol. Evol. 6:3. doi: 10.3389/fevo.2018.00003
Bothma, J. D. P., and du Toit, J. G., eds. (2016). Game Ranch Management 6th ed. Pretoria: Van Schaik Publishers.
Brennan, T. J., and Keeley, J. E. (2017). Impacts of mastication fuel treatments on California, USA, chaparral vegetation structure and composition. Fire Ecol. 13, 120–138. doi: 10.4996/fireecology.130312013
Butler, J. R. A., Marzano, M., Pettorelli, N., Durant, S. M., du Toit, J. T., and Young, J. C. (2021). Decision-making for rewilding: an adaptive governance framework for social-ecological complexity. Front. Conserv. Sci. 2:681545. doi: 10.3389/fcosc.2021.681545
Capozzelli, J. F., Hecht, L., and Halsey, S. J. (2020). What is the value of wild animal welfare for restoration ecology? Restor. Ecol. 28, 267–270. doi: 10.1111/rec.13114
Carruthers, J. (2008). “Wilding the farm or farming the wild”? The evolution of scientific game ranching in South Africa from the 1960s to the present. Trans. R. Soc. S. Afr. 63, 160–181. doi: 10.1080/00359190809519220
Coe, M. J., Cumming, D. H. M., and Phillipson, J. (1976). Biomass and production of large African herbivores in relation to rainfall and primary production. Oecologia 22, 341–354. doi: 10.1007/BF00345312
du Toit, J. T., and Cumming, D. H. M. (1999). Functional significance of ungulate diversity in African savannas and the ecological implications of the spread of pastoralism. Biodivers. Conserv. 8, 1643–1661. doi: 10.1023/A:1008959721342
du Toit, J. T., and Pettorelli, N. (2019). The differences between rewilding and restoring an ecologically degraded landscape. J. Appl. Ecol. 56, 2467–2471. doi: 10.1111/1365-2664.13487
East, R. (1984). Rainfall, soil nutrient status and biomass of large African savanna mammals. Afr. J. Ecol. 22, 245–270. doi: 10.1111/j.1365-2028.1984.tb00700.x
Evans, A. E., Zimova, M., Giery, S. T., Golden, H. E., Pastore, A. L., Nadeau, C. P., et al. (2022). An eco-evolutionary perspective on the humpty-dumpty effect and community restoration. Oikos 2022:e08978. doi: 10.1111/oik.08978
Fløjgaard, C., Pedersen, P. B. M., Sandom, C. J., Svenning, J.-C., and Ejrnæs, R. (2022). Exploring a natural baseline for large-herbivore biomass in ecological restoration. J. Appl. Ecol. 59, 18–24. doi: 10.1111/1365-2664.14047
Fritz, H., and Duncan, P. (1994). On the carrying capacity for large ungulates of African savanna ecosystems. Proc. R. Soc. B 256, 77–82. doi: 10.1098/rspb.1994.0052
Harris, T., Ottaviani, G., Mulligan, M., and Brummitt, N. (2022). Trait hypervolumes based on natural history collections can detect ecological strategies that are distinct to biogeographic regions. J. Ecol. 111, 314–326. doi: 10.1111/1365-2745.14005
Hempson, G. P., Archibald, S., and Bond, W. J. (2015). A continent-wide assessment of the form and intensity of large mammal herbivory in Africa. Science 350, 1056–1061. doi: 10.1126/science.aac7978
Hobbs, R. J., Arico, S., Aronson, J., Baron, J. S., Bridgewater, P., Cramer, V. A., et al. (2006). Novel ecosystems: theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 15, 1–7. doi: 10.1111/j.1466-822x.2006.00212.x
Linnell, J. D. C., and Jackson, C. R. (2019). “Bringing back large carnivores to rewild landscapes” in Rewilding. Ecological reviews. eds. N. Pettorelli, S. M. Durant, and J. T. du Toit (Cambridge, UK: British Ecological Society and Cambridge University Press), 248–279.
Maestre, F. T., Le Bagousse-Pinguet, Y., Delgado-Baquerizo, M., Eldridge, D. J., Saiz, H., Berdugo, M., et al. (2022). Grazing and ecosystem service delivery in global drylands. Science 378, 915–920. doi: 10.1126/science.abq4062
Morford, S. L., Allred, B. W., Twidwell, D., Jones, M. O., Maestas, J. D., Roberts, C. P., et al. (2022). Herbaceous production lost to tree encroachment in United States rangelands. J. Appl. Ecol. 59, 2971–2982. doi: 10.1111/1365-2664.14288
Pettorelli, N., Durant, S. M., and du Toit, J. T., eds. (2019). Rewilding. Ecological reviews. Cambridge: British Ecological Society and Cambridge University Press.
Piipponen, J., Jalava, M., de Leeuw, J., Rizayeva, A., Godde, C., Cramer, G., et al. (2021). Global trends in grassland carrying capacity and relative stocking density of livestock. Glob. Change Biol. 28, 3902–3919. doi: 10.1111/gcb.16174
Pimm, S. L. (1991). The balance of nature? Ecological issues in the conservation of species and communities. Chicago: University of Chicago Press.
Segar, J., Pereira, H. M., Filgueiras, R., Karamanlidis, A. A., Saavedra, D., and Fernández, N. (2022). Expert-based assessment of rewilding indicates progress at site-level, yet challenges for upscaling. Ecography 2022:e05836. doi: 10.1111/ecog.05836
Sinclair, A. R. E., Mduma, S., and Brashares, J. S. (2003). Patterns of predation in a diverse predator-prey system. Nature 425, 288–290. doi: 10.1038/nature01934
Keywords: large herbivores, surrogate species, herbivore biomass, herbivory traits, ecosystem function, stakeholder engagement
Citation: du Toit JT (2023) Steps to operationalize a rewilding decision: Focus on functional types. Front. Ecol. Evol. 11:1114856. doi: 10.3389/fevo.2023.1114856
Edited by:
Anthony Lehmann, University of Geneva, SwitzerlandReviewed by:
Mario Santiago Di Bitetti, CONICET Institute of Subtropical Biology (IBS), ArgentinaCopyright © 2023 du Toit. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johan T. du Toit, am9oYW4uZHV0b2l0QHVzdS5lZHU=