 Carmen Huerta1
*
†
Carmen Huerta1
*
†
 Magdalena Cruz-Rosales1
†
Magdalena Cruz-Rosales1
†
 Patricia González-Vainer2
†
Ivette Chamorro-Florescano3
†
Patricia González-Vainer2
†
Ivette Chamorro-Florescano3
†
 José D. Rivera1
,4
*
†
José D. Rivera1
,4
*
†
 Mario E. Favila1
†
Mario E. Favila1
†
- 1Red de Ecoetología, Instituto de Ecología, A.C., Xalapa, Mexico
- 2Sección Entomología, Facultad de Ciencias, Universidad de la República Iguá, Montevideo, Uruguay
- 3Facultad de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana, Tuxpan, Mexico
- 4Instituto de Ecología, Universidad Nacional Autónoma de México, Ciudad de México, Mexico
Dung beetles display complex reproductive behaviors involving sexual findings, sexual recognition, fighting for mates and food used for nesting, sperm competition, and parental care. Over the past 40 years, significant advances have been made regarding the knowledge of various aspects of the sexual and nesting behavior of Neotropical dung beetles. However, human activities modify the natural habitats of dung beetles at an alarming rate, affecting food availability and altering the ecological functions performed by the species in their different habitats. A deeper understanding of the reproductive behavior of dung beetles may contribute significantly in understanding the evolutionary diversification of these insects and their response to environmental changes. The present study reviews and analyzes studies regarding the sexual and reproductive behavior of Neotropical dung beetle species under field and laboratory conditions. We gathered 132 studies and 146 species; 42% of the available data were based on field observations, 23% on laboratory observations, 30% under both field and laboratory conditions, and 5% unspecified. Our review detected significant knowledge, geographic, and habitat gaps regarding the reproductive behavior of Neotropical dung beetles. Based on our findings, we propose future research goals and alternative methods to measure the behavioral responses of Neotropical dung beetles to the impacts of human activities.
Introduction
Dung beetles (Coleoptera: Scarabaeidae: Aphodiinae and Scarabaeinae, and Geotrupidae) are evolutionarily related species that use dung, carrion and decomposing fruits as organic sources for feeding and reproduction (Halffter and Matthews, 1966; Halffter and Edmonds, 1982; Hanski and Cambefort, 1991; Scholtz et al., 2009; Simmons and Ridsdill-Smith, 2011). According to how they handle the food resource and the structure of the nests they construct, Halffter and Matthews (1966) established four nesting groups (I-IV). Later, Halffter (1977) and Villalva et al. (2002) mentioned two evolutionary nesting lines closely related to the food resource they consume: burrower and roller beetles, also referred to as paracoprids and telecoprids, respectively. According to Halffter (1977) and Halffter and Edmonds (1982), there are seven nesting patterns in the Scarabaeinae (I to VII), whereas the Geotrupidae displays only the nesting pattern I, and most Aphodiinae are non-nesters.
Nesting patterns I, II, and III are observed in burrower beetles that construct underground nesting galleries, generally below or to one side of the food source, where they accumulate and manipulate food for their young. Simple underground nests contain only one brood mass or ball in each gallery, while compound nests contain two or more brood masses or brood balls per gallery. Typically, each brood mass or brood ball contains only one egg. In pattern I, the female leaves the nest after the egg is laid in the brood mass; a female can construct several brood masses. In the species showing pattern II, the female adds a layer of soil to the food after oviposition, which can be several millimeters thick, forming a brood ball, and then abandons the nest. The nest can contain up to two or three brood balls. Pattern III includes species in which the female builds nests with several brood balls and cares for them during the larval development of the progeny (Figure 1).
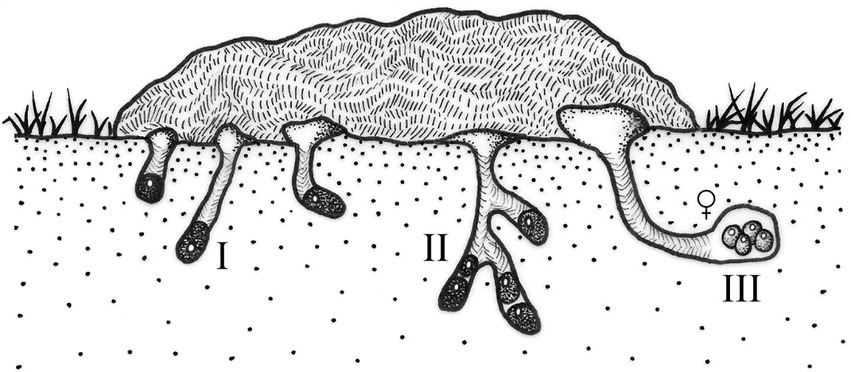
Figure 1. Nesting patterns I, II, and III (sensu Halffter and Edmonds, 1982).
Patterns IV and V (Halffter and Edmonds, 1982) group roller beetles in which the male makes a food ball of dung or carrion from the food source, attract a female and rolls the food ball, with the female transported on the ball some meters away from the food source where the nest will be established. While searching for food, mate, and rolling, males fight for food balls and females. The patterns of roller beetles are differentiated by the complexity of the nesting behavior of both males and females (Figure 2). Pattern IV groups species that build simple nests, formed by one brood ball that may or may not be covered by a layer of soil after oviposition and that are abandoned by the female. In nesting pattern V, the female builds several brood balls from a single ball. Each ball is covered with a layer of soil, and the egg is laid in a chamber built by the female at the apical section of the brood ball (Figure 2; Halffter and Edmonds, 1982; Cambefort and Hanski, 1991; Halffter, 1997; Simmons and Ridsdill-Smith, 2011; Halffter et al., 2013). After brood balls are built, the female takes care of the nest during larval development; the male also stays in the nest, protecting the brood ball and the female, avoiding other males from mating it (Favila et al., 2005). In pattern VI, the female makes brood balls built directly at the food source. Most of the species of the genus Eurysternus Dalman belong to this pattern, which usually displays maternal care (Huerta et al., 2003). Within the Scarabaeinae, the members of the Eucraniini tribe, which are endemic to the arid and semi-arid zones of Argentina, were considered rollers by Monteresino and Zunino (2003) (named telephagic and telecoprid by these authors). However, these dung beetles neither make nor roll balls, so they were not grouped in the nesting patterns established by Halffter and Edmonds (1982). Currently, little is known about their reproduction (Figure 3).
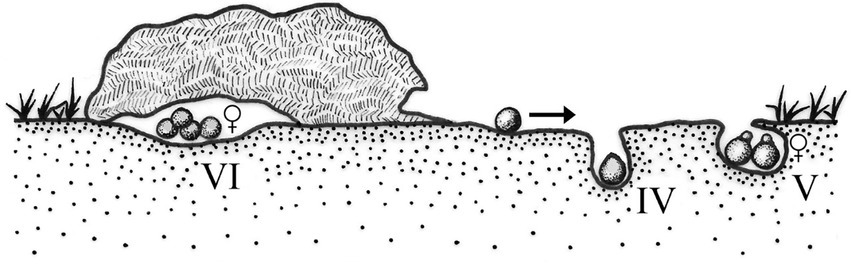
Figure 2. Nesting patterns IV, V, and VI (sensu Halffter and Edmonds, 1982).
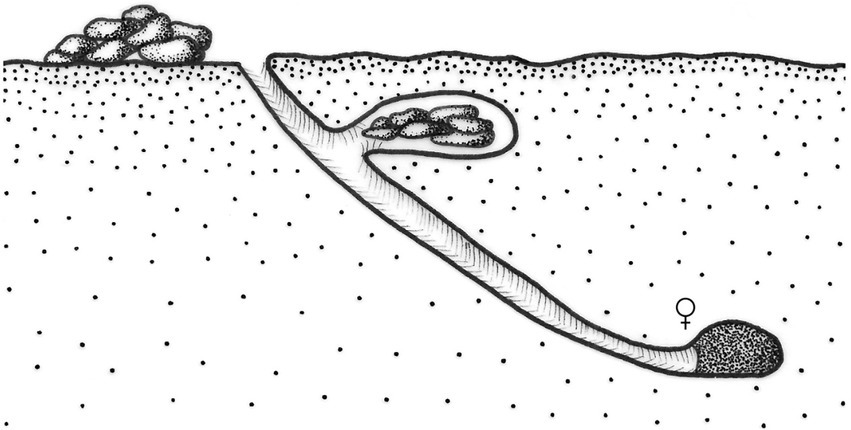
Figure 3. Nesting pattern for Eucranium Brullé (modified from Monteresino and Zunino, 2003).
After the hatching or emergence from the brood balls or nesting balls, adults are immature (tenerals), and the pre-reproductive period begins, during which adult beetles primarily search and fight for food and also feed. According to Zunino and Palestrini (1986) and Tonelli (2021), young adults feed in different ways: (a) directly above the food source (epiphagic behavior); (b) within the food source (endophagic behavior); (c) under the food source (mesophagic behavior); (d) burying the food in underground galleries to consume it subsequently (hypophagic behavior), and (e) making food balls that are first rolled some distance away from the food source, then buried, and later consumed (telephagic behavior). Throughout the pre-reproductive period, predominate feeding behavior, but there are various aggressive intra- and interspecific interactions, in addition to sexual interactions, including copulation at the end of this period. The pre-reproductive period, which varies according to the species, is followed by the reproductive period. In the subfamily Aphodiinae, some species do not build nests but lay their eggs directly in or under the food; larvae are free-living and directly consume the food source (Halffter and Edmonds, 1982; Hanski and Cambefort, 1991; Nichols et al., 2008; Huerta et al., 2013; Tonelli, 2021). Most Geotrupidae and Scarabaeinae species build nests during this stage. The post-reproductive stage has been little studied in coprophagous beetles, mostly under laboratory conditions; it is known that old individuals have limited activity and ultimately die.
According to Halffter and Matthews (1966), most nesting patterns of Neotropical dung beetle species (non-nesting, paracoprid and telecoprid) are well-represented and can be found in different habitats. However, while the reproductive behavior (including nesting patterns) of some dung beetle species has been studied in great detail, these are usually isolated efforts involving a set of model species (e.g., Canthon cyanellus, Eurysternus, Copris incertus; see Huerta and Halffter, 2000; Huerta et al., 2003, 2005; Favila et al., 2012). Thus, the current state of knowledge on the reproductive behavior of dung beetles in the Neotropics is still to be determined. We aim to review studies regarding the sexual and reproductive behavior of Neotropical dung beetle species under field and laboratory conditions. Our research will contribute to systematizing the information available to date, as well as to guide future research efforts, especially in those tribes or regions with significant information gaps.
Materials and methods
Literature search
The database was constructed through a systematized search of peer-reviewed papers on Web of Science (WoS) regarding the reproductive behavior of Neotropical dung beetles. The search was conducted in the second semester of 2022, covering articles published between 1980 and 2022. We employed the following search terms across the title, abstract, and keywords of each paper: [“dung beetle*” OR Scarabaeinae OR Aphodiinae OR Geotrupidae) AND (“reproductive behavior*” OR reproduct* OR etholog* OR behavior* OR “reproductive success” OR mating* OR “mating success” OR competition OR combat OR contest) AND (nest* OR nesting OR “nesting behavior” OR “resource reloca*” OR “food reloca*” OR “resource reloca*” behavior OR “resource reloca* behavior”].
Inclusion criteria and screening protocol
The WoS search retrieved 117 articles, of which we included only those meeting the following criteria: (1) the study should be based on species of the subfamily Scarabaeinae, Aphodiinae, or the family Geotrupidae; (2) the study should be conducted within the Neotropics, including also the North of Mexico, and the Andean region (sensu Morrone et al., 2022), with species native to the region; (3) the study should evaluate behaviors associated with at least one of the three main reproductive stages of beetles: pre-reproduction, reproduction, or post-reproduction. Behaviors associated with the pre-reproductive stage include rolling, construction, excavation of galleries, food relocation, and some reproductive behaviors such as courtship, competition/combat (intra- and intersexual competition), and copulation. Regarding the reproductive stage, we considered female–male behaviors associated with building nests and nesting galleries, as well as rolling and fighting behaviors that may emerge between individuals trying to obtain food resources for nesting, sperm competition, and female selection. The behaviors observed and recorded after reproduction were considered post-reproductive stages. After reviewing the abstracts and titles of the full WoS search results, we selected 35 articles for a full review. The database was supplemented with 97 articles from the authors’ library, as these journals lacked a digital repository or the papers were published before 1980 (Supplementary Table S1).
Data extraction
We extracted the following data from the selected literature: (a) the taxonomic identity of each dung beetles species studied (current names of species were used in case of synonyms or new combinations, Supplementary Table S2); (b) study location, including locality and country; the environment and known food resources of each species (Supplementary Table S3); (c) copulation data observed in different Scarabaeinae species, including duration, location relative to the food ball, and life stage; (d) average duration of nest care and preimaginal development (egg, larvae, pupa, and imago emergence) observed in different Scarabaeinae species; (e) fecundity data of different Scarabaeinae species, including the number of nests or galleries during lifetime, the average number of balls or masses per nest and the number of balls or masses during lifetime; (f) nest characteristics of Scarabaeinae and Geotrupidae (structure, complexity, and location relative to the surface and the food source); (g) male/female nesting behavior (food provisioning, excavation, brood construction, and care); (h) nesting pattern (Supplementary Table S4); (i) Aphodiinae reproductive behavior and preimaginal development data.
Results
We reviewed 132 publications and 146 species associated with at least one of the three reproductive stages: pre-reproductive, reproductive, or post-reproductive. Species were evaluated based on the different recording techniques used, of which 42% were field observations, 23% were laboratory observations, 30% were studies performed under field and laboratory conditions, and 5% did not specify the conditions under which the study was carried out.
Observations associated with the pre-reproductive behavior were mentioned in 100 studies containing 101 species records. We found 119 studies with 132 species focused on reproductive observations, such as those associated with nest building and behaviors that may arise from the defense of limited resources (e.g., food and females). Only 40 papers (30%) included observations related to the post-reproductive stage in 18 species.
Study locations
Studies on various aspects of the reproductive behavior of the Scarabaeidae (Aphodiinae and Scarabaeinae) and Geotrupidae were conducted in 15 countries, ranging from Mexico to Argentina (Figure 4). The recorded species were found mainly in three countries: Mexico (38%), Argentina (28%), and Brazil (21%). The remaining countries had less than three species analyzed (Table 1).
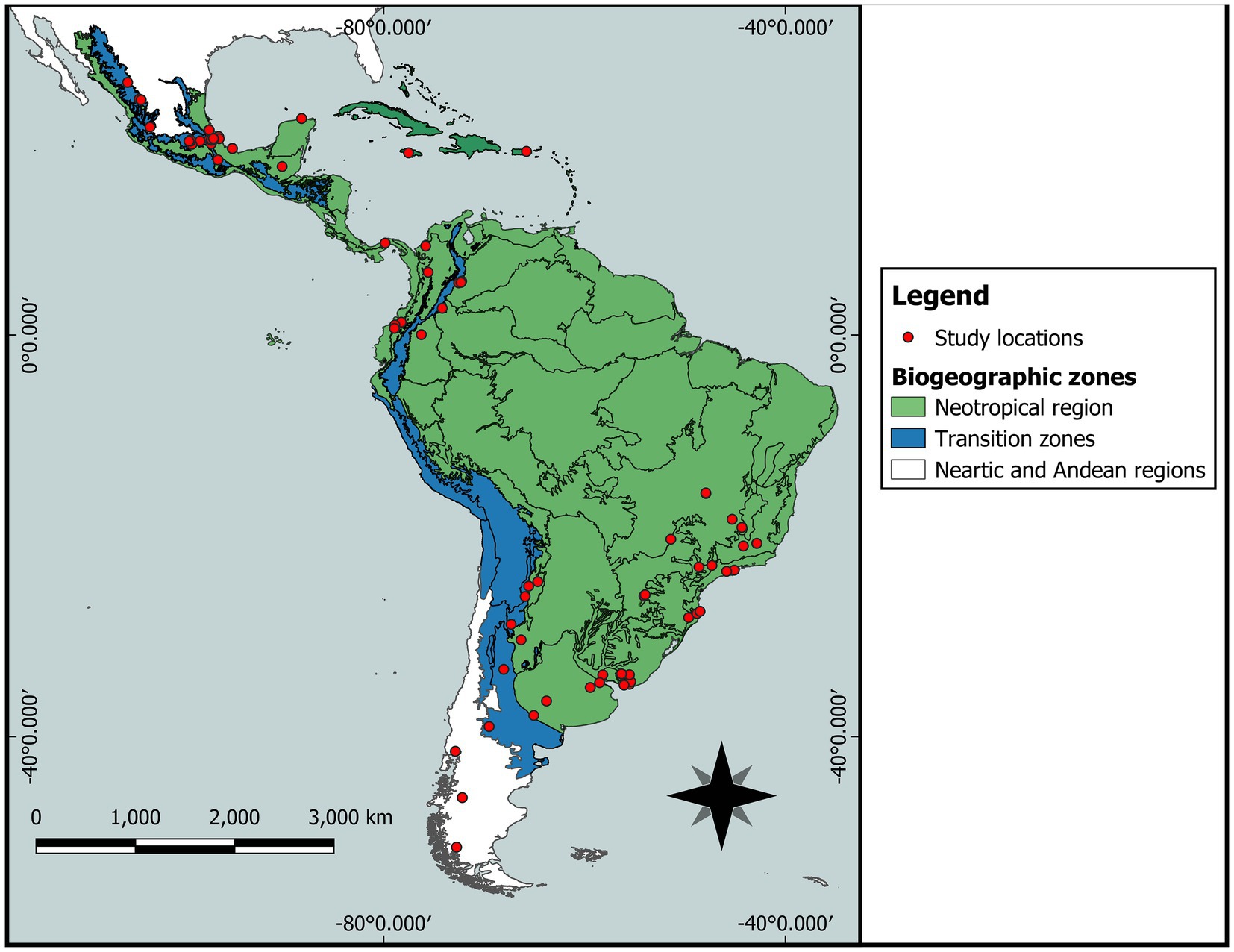
Figure 4. Study locations in countries of Latin America regarding the reproductive behavior of Aphodiinae, Scarabaeinae, and Geotrupidae.
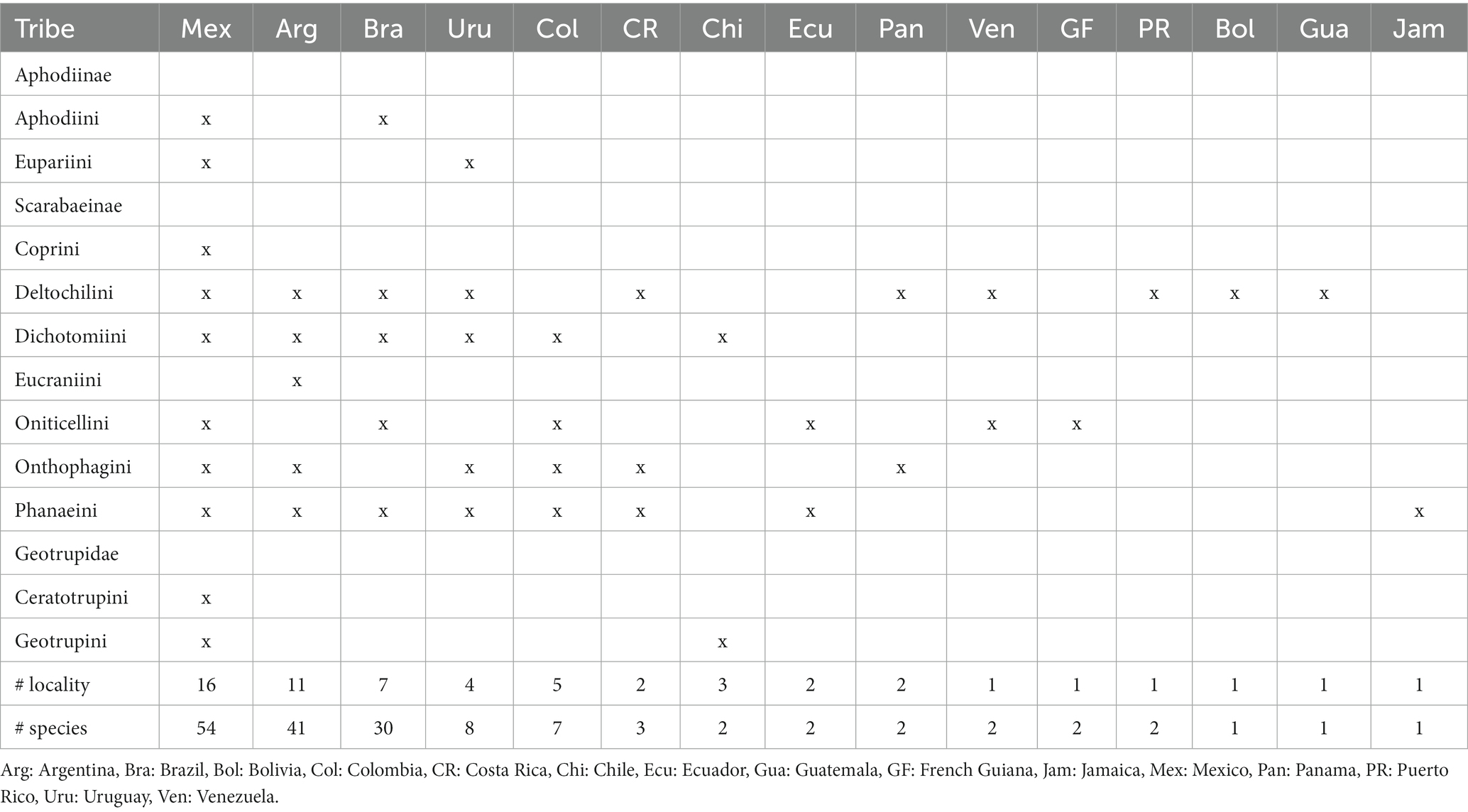
Table 1. Paper records on the reproductive behavior of Aphodiinae, Scarabaeinae, and Geotrupidae, according to country, locality, tribe, and study species.
The diversity of localities followed the same order mentioned by country. In Mexico, most of the localities referred to in the studies correspond to 16 states, mainly Veracruz, Chiapas, and the State of Mexico. In Argentina, we recorded 11 provinces, including La Rioja, Jujuy, and San Luis. Brazil included seven states, of which São Paulo and Minas Gerais were most frequently mentioned. Colombia and Uruguay had five and four departments, respectively. For each of the remaining countries, only one or two localities are cited (Figure 4).
Considering the family or subfamily of the beetles evaluated, Scarabaeinae was the most studied group in almost all countries, followed by Aphodiinae, studied only in Mexico and Uruguay, and Geotrupidae, studied only in Mexico and Chile. At the level of the Scarabaeinae tribes, Deltochilini was studied in nine countries, followed by Phanaeini in eight countries, and Dichotomiini, Oniticellini, and Onthophagini in six countries. The Eucraniini tribe was studied exclusively in Argentina, and Coprini in Mexico (Table 1).
Among the Scarabaeinae genera, Canthon Hoffmannseg was the most represented, with 22 species in five countries, followed by Onthophagus Latreille with 13 species distributed in six countries, Eurysternus Dalman with 10 species in six countries, Deltochilum Eschscholtz with nine species in five countries, and Dichotomius Hope with nine species in four countries (Table 2).
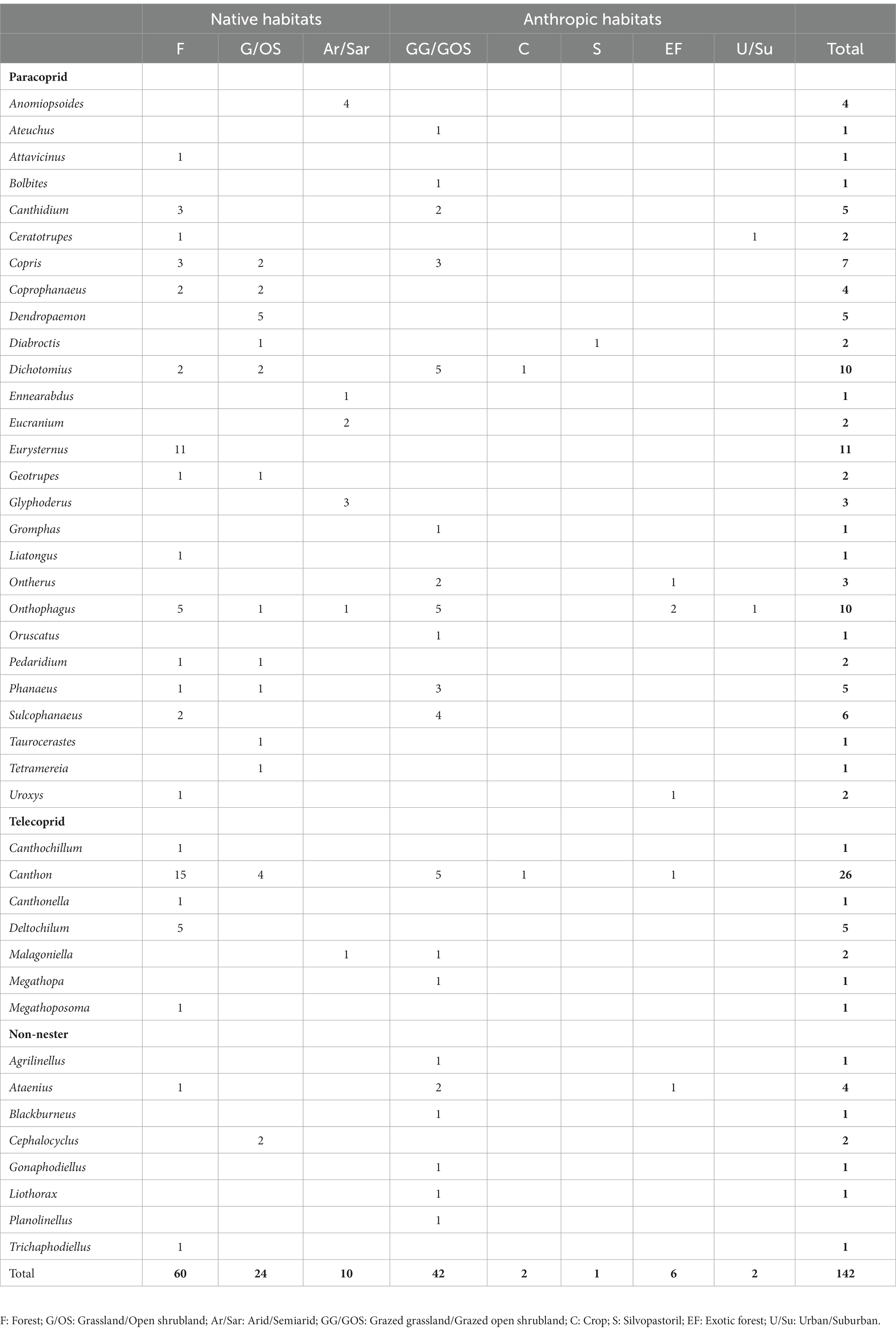
Table 2. Number of species by genus in the studied environments.
Habitats
The habitats where dung beetles display their feeding or reproductive behaviors were mentioned for approximately 97% of the recorded species (Table 2). About half of these species live in native or introduced open environments, such as grasslands in tropical and temperate zones of Mexico, the Pampean region and grasslands of Uruguay, the steppes of Argentina, and the savanna of Brazil. In multiple studies (64%), specimens were collected in fields or pastures dedicated to livestock raising. The species studied within grassland ecosystems were mainly burrower Scarabaeinae (36), with type I, II, or III nesting patterns. Studies on the behavior of roller and non-nesting Aphodiinae species in grasslands were lower (11 and 9, respectively; Table 2; Supplementary Table S3).
Behavioral studies on species collected in forest environments represent one-third of the total; these forest ecosystems include tropical rain forests, temperate mountain forests, or xeric forests, and fruit or forest plantations. Among the species of Scarabaeinae studied in forest ecosystems, 28 were burrower species, and 24 were roller species. A small proportion of all studied species (less than 10%) were recorded in a variety of habitats including native forests, grassland environments, and silvopastoral systems (i.e., Canthon virens (Mannerheim) and Diabroctis cadmus Harold (Forti et al., 2012; Halffter et al., 2013; Murillo-Ramos et al., 2016). Similarly, Canthon cyanellus cyanellus LeConte (Martínez, 1992a,b), and C. imitator Brown (Martínez et al., 2019) were collected in both tropical forests and neighboring pastures or crops. The burrower species Onthophagus curvicornis Latreille (Montes-Rodríguez, 2017), Ceratotrupes fronticornis (Erichson; Ramírez-Restrepo and Halffter, 2016), and Dichotomius satanas (Harold; Barretto et al., 2021a,b), which generally occurs in forest environments, also display their activities in pastures dedicated to livestock raising, crops, and even in urban or semi-urban areas. Finally, the tribe Eucraniini represented about 10% of the recorded species studied. These are exclusive burrowers of arid and semi-arid environments, endemic to the northwestern region of Argentina (Zunino et al., 1989; Monteresino and Zunino, 2003; Ocampo, 2005, 2010; Ocampo and Philips, 2005).
Feeding, gallery construction, and food relocation
Coprophagous burrower beetles may feed directly on food or build subterranean galleries to store and later consume the resource during the pre-reproductive stage (Halffter and Matthews, 1966; Halffter and Edmonds, 1982; Halffter et al., 1985; Hanski and Cambefort, 1991; Ocampo and Hawks, 2006; Scholtz, 2009). The pre-reproductive behavior has been studied in detail in eight species of Phanaeus (Halffter et al., 1974; Halffter and López, 1977; Price and May, 2009; Huerta et al., 2010). These Phanaeus species roll pieces of food some distance away from the source, pushing them with the head and forelegs (butting behavior), then burying them in a feeding gallery for subsequent consumption. According to Huerta and Halffter (2000), in three species of Copris Müller (C. armatus Harold, C. lugubris Boheman and C. incertus Say) during the feeding stage, males preferentially store food in sausage-shaped galleries, while females accumulate food masses or nest cakes in subterranean chambers, rather than galleries.
Coprophagous and necrophagous roller species cut and transport a food ball at different distances from the food source, which is used for feeding by sexually immature beetles or nesting by sexually mature beetles (Matthews, 1963; Halffter and Matthews, 1966; Halffter and Edmonds, 1982). During rolling, the beetle pushes the ball with the hind and middle legs; the forelegs are applied on the ground to move the ball. This rolling position is known as the pushing position (Halffter and Matthews, 1966). Most Neotropical species move food by the pushing position. During rolling, the food ball is covered by a layer of soil to avoid its drying out and the arrival of other competitors; in addition, the food ball is protected by chemicals from the exocrine glands of the abdominal and pygidia region of the beetles (Bellés and Favila, 1983; Pluot-Sigwalt, 1988a,b, 1991). The substances released by these glands play multiple functions: defense against competitors such as flies; fungicides or fungistatic; pheromones, and prevention of food decomposition, allowing its consumption by larvae in the brood ball (Favila et al., 2012). However, various species of the genus Canthon (e.g., C. septemmaculatus, Latterille, C. tristis, Harold, C. obliquus Horn, C. edentulus Harold) and Deltochilum orbignyi Blanchard form the food ball but do not roll it; instead, they bury it shallowly below the food source (Halffter and Matthews, 1966; Halffter and Halffter, 1989; Halffter et al., 2013). Eurysternus species are considered roller beetles; however, they feed and nest directly below the food source during the pre-reproductive and reproductive stages, respectively (Huerta et al., 2003).
Male competition for food and females
Studies on male competition for food or potential nesting partners are scarce in Neotropical coprophagous dung beetles. Studies on fights between females for food resources are even scarcer. Fight dynamics during the pre-reproductive and reproductive stages have been mentioned or addressed in field and laboratory studies of only six species: Phanaeus tridens, Laporte, Onthophagus acuminatus, Harold, Canthon quinquemaculatus, Laporte, C. lituratus, Germar, C. bispinus Germar, and C. cyanellus (Halffter and Edmonds, 1982; Favila, 1988; Favila and Díaz, 1996; Rodrigues and Flechtmann, 1997; Price and May, 2009; Halffter et al., 2013; Cantil et al., 2014b; González-Vainer, 2015; Salomão et al., 2019).
Pre-reproductive field records on the combat behavior between Phanaeus tridens males showed that the winner joins a female, pushing and burying the excrement fragment next to her. Male fights associated with food thievery, gallery invasion and fights between females have been documented in this species (Halffter and Edmonds, 1982; Price and May, 2009). Different fighting strategies for the gallery and the female have been observed in Onthophagus acuminatus, where the male owner guards the gallery entrance against intruders and periodically patrols along the tunnel (Emlen, 1997). The owner male fighting to defend the tunnels is horned and mates repeatedly with the occupant female. Intruder males can confront the owner and fight until one of them leaves the tunnel. However, hornless intruder males can also sneak inside, avoiding the guarding male. The intruder male digs side tunnels that intersect guarded tunnels and stealthily copulates with the female of the nest-owning male (Emlen, 1997). The guarding male can catch sneaking males before encountering the female. In this case, sneaking males return to their side tunnels or build new tunnels and remain inactive for several hours, but later they attempt to enter the primary tunnel again in search of the female (Emlen, 1997).
Regarding Canthon quinquemaculatus and C. lituratus, the theft or splitting of the food ball during rolling has been frequently observed. Specifically, in C. quinquemaculatus a case is mentioned where the male rolling the food ball with a female was displaced by the intruder and the female that was rolling with the owner male remained with the thief (Rodrigues and Flechtmann, 1997; Halffter et al., 2013; Cantil et al., 2014b). In C. bispinus, intrasexual fights for the food ball were recorded during the food ball rolling and burying by male–female pairs (González-Vainer, 2015).
Fights in Canthon cyanellus have been extensively documented in field and laboratory studies. Fights occur mainly between males during the breeding season; intruder males attempt to steal the food ball and the female is transported by an owner male to the nesting site (Favila and Díaz, 1996; Chamorro-Florescano and Favila, 2008). Differences in body size and the reproductive status of females and males (virgins or with previous nesting) influence the fighting success between contestant males for the females transported in the food ball (Chamorro-Florescano and Favila, 2008; Chamorro-Florescano et al., 2011). In Copris laeviceps Harold, fights during nesting have also been documented but generated by experimentally placing an intruder female in a nest where brood balls are looked after by a resident female (Klemperer, 1986). Klemperer (1986) also reported cases of a female taking care of brood balls constructed by another female in a foreigner nest.
Mating behavior
Copulation is not easy to observe because it mainly occurs in underground galleries, making it difficult to determine when and how it happens. However, we found information on different aspects of the copulation behavior in 73 species of Neotropical Scarabaeinae beetles (Table 3). The copulation process has been described for only seven species: Megathoposoma candezei (Harold; Wille, 1973), Phanaeus daphnis, P. mexicanus (Halffter and López, 1977), Liatongus rhinocerulus (Bates; Halffter and Edmonds, 1982), Canthon cyanellus, C. indigaceus chevrolati (Martínez and Cruz, 1990), and Onthophagus acuminatus (Emlen, 1997).
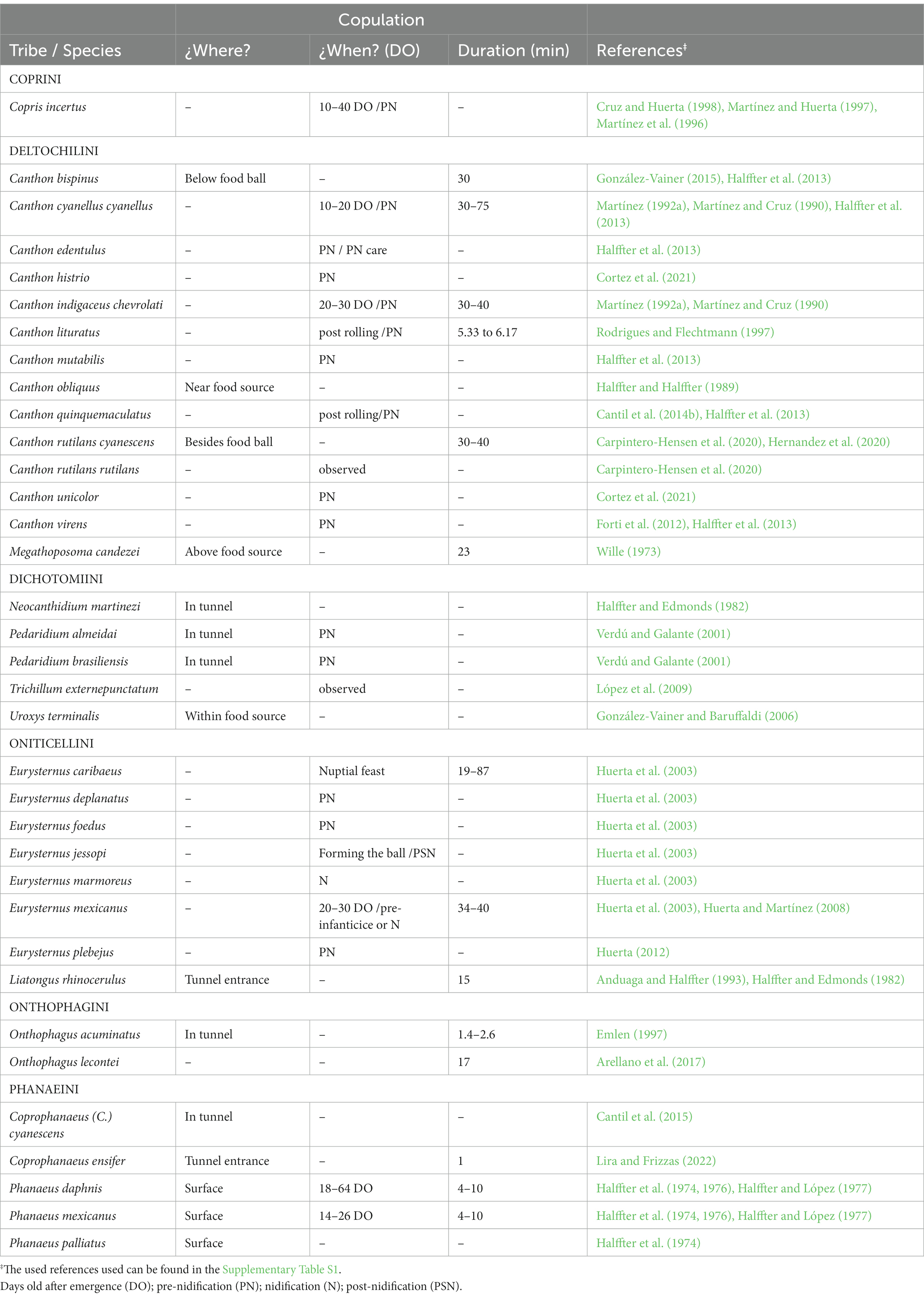
Table 3. Copulation data observed in different Scarabaeinae species.
Halffter and López (1977) defined four stages during copulation: (a) approach of the male to the female, generally near the food source as a meeting place; (b) detection of the female by the male; (c) positive response of the female and male mounting on her and attaching his parameres at the genital opening; (d) insertion of the male aedeagus. During this last stage, males can remain still or include touching or tapping movements with their front legs on the thorax, elytra, or sides of the female, supposedly intended to keep it calm. Although Halffter and Edmonds (1982) considered that there is no elaborate pre-copulatory or courtship behavior among Scarabaeinae, Emlen (1997) interpreted the tapping behavior observed in several species of insects as a copulatory courtship. Finally, at the end of copulation, the male separates and may remain near the female, guarding it against other males, or he may bury himself, searching for food or leaving the nest, depending on the species.
Details related to the site, time, or duration of copulation are only known for 34 species (Table 3), 14 of which are Canthon species because these are more easily observed during the rolling of food balls, and for seven Eurysternus species next to the food source. The remaining copulation observations correspond to species for which male–female pairs were found at their gallery entrance. A first copulation during the pre-nesting stage, which allows females to complete oocyte maturing and initiate the nesting stage, has only been observed in Phanaeus mexicanus, P. daphnis, Canthon cyanellus cyanellus, C. indigaceus chevrolati, Copris incertus, and Eurysternus mexicanus Harold (Halffter et al., 1976; Halffter and López, 1977; Martínez and Cruz, 1990; Martínez et al., 1996; Huerta et al., 2003). Most copulations observed on the surface at the gallery entrance occur before the nest ball or mass elaboration stage. The copulation duration varied from very short, lasting only one or 2 min in Coprophanaeus ensifer (Germar) and Onthophagus acuminatus (Emlen, 1997; Lira and Frizzas, 2022), up to maximum times of 75 min in Canthon cyanellus cynellus (Martínez and Cruz, 1990) and 84 min in Eurysternus caribaeus (Herbst; Huerta et al., 2003). The remaining observed cases ranged from 5 to 40 min (Table 3).
Sperm competition and female selection
Sperm competition in Neotropical dung beetle species has only been studied in Canthon cyanellus, observing that the male repeatedly copulates during nesting and before and during the elaboration of nest balls (Favila et al., 2005). It was experimentally demonstrated, with a cuticular genetic marker, that with this behavior, males significantly increase their paternity under a sperm competition scenario (Favila et al., 2005). In another study, Chamorro-Florescano and Favila (2009) found that males regulate copulation frequency during nesting; if the female has already copulated with other males, the latest male increases copulation frequency between the elaboration of nest balls; however, when a male is nesting with a virgin female, he does not mate several times. Besides, only males with previous reproductive experience increase their paternity, contrary to virgin males; that is, the reproductive experience of the female affects the frequency of mating, and the reproductive status of the male affects its reproductive success during the nesting stage (Chamorro-Florescano and Favila, 2009). In Canthon edentulus, copulations have also been observed during the nest care period (Halffter et al., 2013), suggesting that in this species, sperm competition also occurs and that the male can reduce it by repeated mating.
Cryptic choice by females can lead to direct or indirect benefits when choosing the male with whom they will mate (Andersson, 1994). The behaviors of males during combats may be signals allowing females to evaluate and select the most successful males; this, in turn, may be related to attributes indicative of male “quality” (Berglund et al., 1996; Briffa and Sneddon, 2007). Few studies of Neotropical beetles have recorded female behaviors that may reflect the choice or preference towards males with particular attributes. For example, Canthon cyanellus females that rolled together with a male were observed to actively participate during male–male combat over attempted ball-stealing by an intruder male, favoring and selecting males with reproductive experience, i.e., with previous mating and nesting (whether intruder or owner) over virgin males (Chamorro-Florescano et al., 2011). Another study found that the combat outcome can significantly affect the reproductive success of both combatants. Winning males increase their acquired paternity under conditions of sperm competition compared to loser males in previous combats (Chamorro-Florescano and Favila, 2016; Chamorro-Florescano et al., 2017).
Sperm competition and cryptic female choices play an essential role for dung beetles during and after copulation. Females copulating with different males may store sperm indefinitely until oocyte fertilization (Halffter and Edmonds, 1982; Eberhard, 1996). However, multiple factors can affect these dynamics. Sexual recognition in dung beetles occurs through cuticular hydrocarbons (Ortiz-Domínguez et al., 2006). These compounds act as short-distance contact pheromones and vary according to several factors, such as the changes at the gonad level in females and males during reproduction (Halffter and López, 1977; Huerta et al., 1981; Martínez and Cruz, 1990; Martínez, 1992b; Martínez and Huerta, 1997; López-Guerrero and Halffter, 2000; Howard and Blomquist, 2005; Huerta and Martínez, 2008; Ginzel, 2010; Favila et al., 2012).
The spermatheca plays a fundamental role in sperm competition and the cryptic choices of females. In Scarabeinae and Aphodiinae, the spermatheca is characterized by a C-shaped sclerotized receptacle with a striated muscle that joins the distal and basal end and a duct. After copulation, sperm migrates through this duct from the spermatophore or seminal fluid to the receptacle, where it is maintained and released until oocyte fertilization (López-Guerrero and Halffter, 2000; Martínez et al., 2001). In the family Geotrupidae, tribe Geotrupini, the spermatheca is pyriform with transverse striations at the base. In most species, it is elongated and has a short duct. Studies suggest that this type of spermatheca is primitive compared to those found in Scarabaeinae (Halffter and Edmonds, 1982; Martínez and Trotta-Moreu, 2010). Although both females and males obtain various benefits by mating with different partners, sperm competition may continue even after copulation under this scenario. However, the female may also perform sperm choice of specific phenotypes by shedding sperm and biasing sperm utilization by muscle contractions, among other possible mechanisms (Eberhard, 1996). These mechanisms have not been explored, but it has been reported that C. cyanellus females may benefit certain males having reproductive experience and a successful combat outcome (Chamorro-Florescano and Favila, 2016; Chamorro-Florescano et al., 2017).
Chemical communication at the reproductive stage: Semiochemicals
Chemical communication occurs through semiochemicals, either pheromone for intraspecific interactions or allelochemicals for interspecific interactions (Cortez, 2013). In dung beetles, both compounds are produced by exocrine glands distributed throughout the body, varying according to species and sex, as seen in Canthon cyanellus, C. indigaceus chevrolati and C. femoralis Chevrolat (Favila, 1988, 2001; Pluot-Sigwalt, 1988a,b). However, other attributes influence differences at the glandular level; for example, roller beetles have more exocrine glands than burrower beetles (Pluot-Sigwalt, 1991; Halffter et al., 2013). Glandular asymmetry has evolved in response to the different ecological pressures experienced by roller and burrower beetles, such as the relocation of the food resource competed by conspecific and heterospecific individuals (Hanski and Cambefort, 1991; Favila, 2001).
Since sexually active beetles can also find each other around food to form reproductive pairs, the presence of short- and long-distance sex pheromones during this stage has been suggested (Favila et al., 2012, 2016; Halffter et al., 2013). Recognition in sexually active males and females occurs when both individuals meet, extend their antennae, and touch each other. With this behavior, they can recognize each other through short-distance cuticular compounds. Immature and same-sex individuals will not display cooperative behavior. However, individuals of different sexes or sexually mature collaborate in elaborating and rolling the food ball for subsequent nesting, as demonstrated in C. cyanellus (Ortiz-Domínguez et al., 2006).
Long-distance pheromone emission by males has been described in different species of dung beetles, attributed as a strategy to attract females for nesting. However, it has also been suggested that the compounds emitted may repel other males or act as defensive chemicals against potential predators (Favila et al., 2016). During pheromone emission, the male holds the first and second pairs of legs upside down and rubs the abdominal sternites with the hind legs, where several exocrine glands are located. At synchronized time intervals, it rubs its legs and lifts them into the air (Tribe, 1975; Favila and Díaz, 1996; Favila, 2001). This behavior has been reported in different species of dung beetles next to the food source or a food ball, irrespective of whether a female is present, such as Canthon bispinus, C. chalybaeus, Blanchard, C. femoralis, C. cyanellus, C. lituratus, C. virens. However, in all these species, no female was observed arriving (Bellés and Favila, 1983; Favila and Díaz, 1996; Rodrigues and Flechtmann, 1997; Silveira et al., 2006; Vaz-de-Mello and Génier, 2009; Favila et al., 2012; Halffter et al., 2013; Cantil et al., 2014b; González-Vainer, 2015; Martín et al., 2021). In C. cyanellus and C. quiquemaculatus, the male exhibits this behavior in a nest where a female is already present (Bellés and Favila, 1983; Cantil et al., 2014b). However, two unidentified male species of Dendropaemon Perty were observed releasing pheromones, displaying the same behavior described at the chamber entrance. The arrival of a female touching the male’s abdomen during the emission behavior, has been described as a recognition behavior in the nesting site, and after this sexual recognition, they enter the gallery (Vaz-de-Mello and Génier, 2009). In C. femoralis, once the male buries the food ball, he performs the pheromone emission behavior until the arrival of the female (Favila, 2012).
Allomones are another multifunctional chemical found in dung beetles, related to defense, aggression, and protection of the food resource and the nest. This behavior has been explored in C. cyanellus, C. femoralis and Deltochilum furcatum Laporte (Halffter and Edmonds, 1982; Bellés and Favila, 1983; Favila, 2001; Cortez et al., 2012, 2015; Favila et al., 2012; Ix-Balam et al., 2018). An example of the above is the secretion of the pygidial glands of C. cyanellus, which produce chemicals that act as a defensive substance against Camponotus sericeiventris Guérin-Méneville (Cortez et al., 2012). When the ant attempts to attack C. cyanellus, it approaches the beetle and touches its body with its antennae. However, defensive compounds repel it, such as phenol in C. cyanellus and geraniol secreted by the pygidial glands of C. femoralis, or guaiacol produced by the pygidial glands of C. cyanellus (Favila, 2001; Cortez et al., 2012). In both species, the importance of the compounds released to impregnate and protect the brood balls from fungi has been proven. Canthon cyanellus displays nest care during almost all larval development and until the emergence of teneral adults, while C. femoralis does not look after the brood ball constructed by the female (Bellés and Favila, 1983; Favila, 2001; Favila et al., 2012). Deltochilum furcatum is another species that does not show nest care. It is suggested that pygidial secretions can reduce the oviposition of Lucilia cuprina Wiedemann flies on food balls rolled by beetles. However, whether this is due to the effect of secretions on microbial activity or an allomone effect is unknown (Ix-Balam et al., 2018).
The pygidial and abdominal secretions with which dung roller beetles impregnate nest balls during rolling are not only essential to avoid heterospecific competition. The chemicals secreted by the pygidial glands of C. cyanellus and C. femoralis, such as acetic acid and benzoic acid, have been found to prevent the decomposition of the food with which beetles elaborate nest balls and on which larvae feed until emergence. These compounds also have microbial activities (Favila, 1993; Cortez et al., 2012; Favila et al., 2016).
Nest care and preimaginal development
Subsocial behavior involving nest care by the female until the emergence of offspring is considered a relatively rare behavior within the Scarabaeinae (Halffter et al., 2013). Only 18 species of three genera have been identified in the Neotropical zone, mainly Copris and some Canthon and Eurysternus species, which take care of the nest until a particular stage of preimaginal development. Of special note is the contribution of males during the nest-care phase, albeit for a short time, in five Canthon species (Halffter et al., 2013; González-Vainer, 2015) (Table 4).
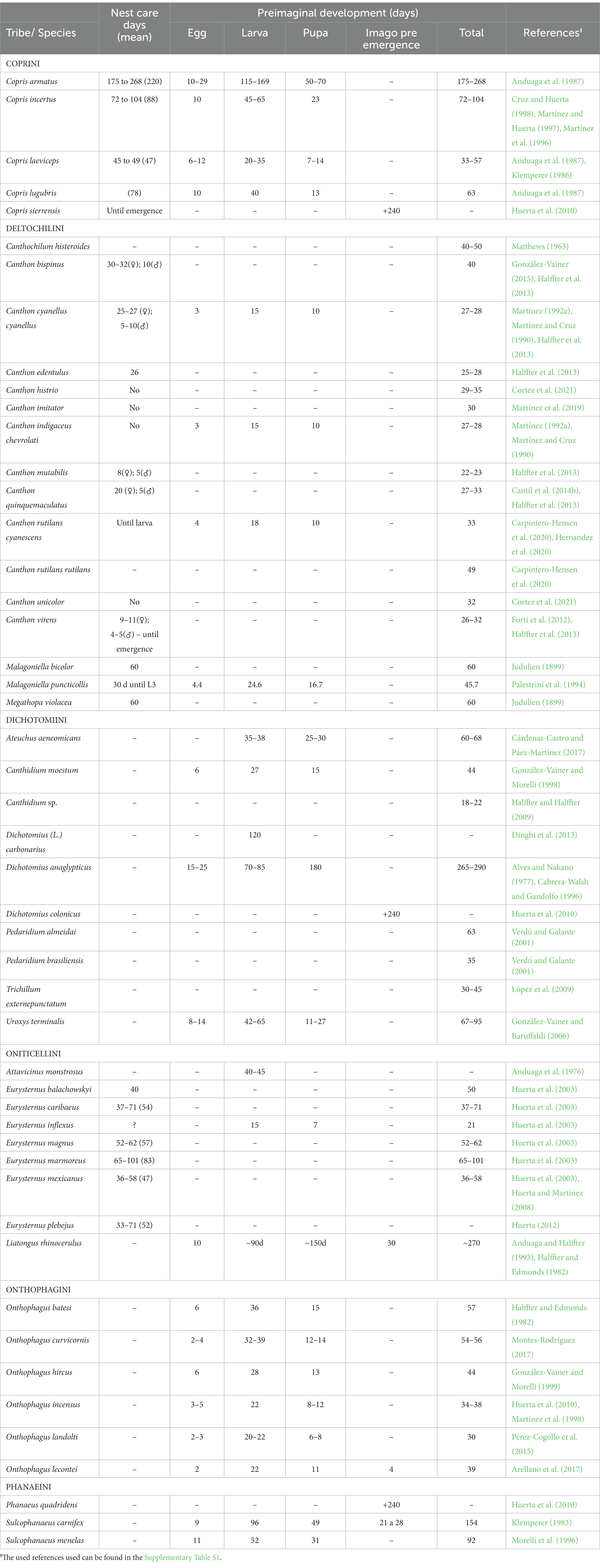
Table 4. Nest care and preimaginal development data observed from different Scarabaeinae species.
The nest care duration varies among species, particularly those that have annual cycles and live in temperate zones, such as Copris armatus and C. sierrensis Matthews, which look after the nest for more than seven and eight months, respectively, until the emergence of the imago, after the winter season ends and temperature improves (Anduaga et al., 1987; Huerta et al., 2010). The other Copris species studied are found in more tropical areas, allowing them to have shorter cycles. In these species, care time varies from 47 days on average in C. laeviceps to 78 and 88 days on average in C. lugubris and C. incertus, respectively (Anduaga et al., 1987; Martínez et al., 1996). Four Canthon species, Malagoniella bicolor (Guerin) and Megathopa violacea Blanchard have been identified among the roller dung beetles that perform nest care almost until imago emergence. Canthon rutilans cyanescens Harold and Malagoniella punticollis (Blanchard) only care for the nest until the larval stage. Canthon virens is a particular case, as several studies report that these perform nest-care only during the larval stage, requiring further confirmatory research (Forti et al., 2012; Halffter et al., 2013). In Canthon mutabilis Harold, females and males look after the nest only for eight and 5 days, respectively, of the 23 days of preimaginal development (Halffter et al., 2013; Table 4). In Canthon cyanellus, the female cares for the nest until the hatching of the offspring, while the male stays in the nest until the female constructs all the brood balls, taking care of both the brood balls and the female to avoid other males mating her (Favila, 1993; Favila et al., 2005).
Among the 11 species of Eurysternus studied, seven take care of the nest; in two species, it is undefined whether or not they perform nest caring, and only E. jessopi Martínez has been described as not taking care of the nest. The known duration of preimaginal development in this genus varies from 21 days in E. inflexus Germar to 83 days on average in E. marmoreus Laporte (Huerta et al., 2003, Table 4).
Of all the Neotropical Scarabaeinae species studied, the duration of preimaginal development until the emergence of the imago is known for 43 species. Many species complete their development until emergence within one to two months, as observed in some members of the tribes Deltochilini, Onthophagini, Coprini, Oniticellini, and Dichotomini (Table 4). Dichotomius (L.) carbonarius (Mannerheim) and Attavicinus monstrosus (Bates) are the only species for which the duration of the larval stage is known. In Copris sierrensis, Dichotomius colonicus (Say), and Phanaeus quadridens (Say), under field conditions, there were still pupae and imagos without emerging 8 months after the onset of preimaginal development, perhaps because they inhabit a temperate zone and were awaiting a more favorable climate to surface (Huerta et al., 2010; Table 4). Therefore, further field or laboratory studies are needed to broaden our understanding of the preimaginal development of multiple dung beetle species, of which little or nothing is currently known.
Fecundity
The fecundity — the number of eggs a female lays over its lifetime — can be considered high in the Aphodiinae (Halffter and Edmonds, 1982). Their nesting behavior is quite simple. The Geotrupidae show a moderate fecundity, with some effort invested in nest building. Finally, in Scarabaeinae, fecundity is relatively low but is compensated by greater reproductive effort or nesting complexity, which may include nest care (Table 5). In this case, some Copris species elaborate one to five nests per year or breeding season, with an average of three or four balls per nest (Anduaga et al., 1987; Martínez et al., 1996; Huerta et al., 2010). Also, Eurysternus species show a low fecundity, with two to three nests per season and three to five balls per nest, but their nesting behavior can be highly complex. For example, E. balachowskyi Halffter and Halffter, E. caribaeus, E. marmoreus, and E. mexicanus build temporary nests that are destroyed before the final nest, implying the likely loss of offspring and a higher fecundity (Huerta et al., 2003). The species with a simpler behavior and low fecundity include Liatongus rhinocerulus Bates, which makes only a single nest per season, consisting of a branched gallery with 6 or 7 nest masses (Anduaga and Halffter, 1993).
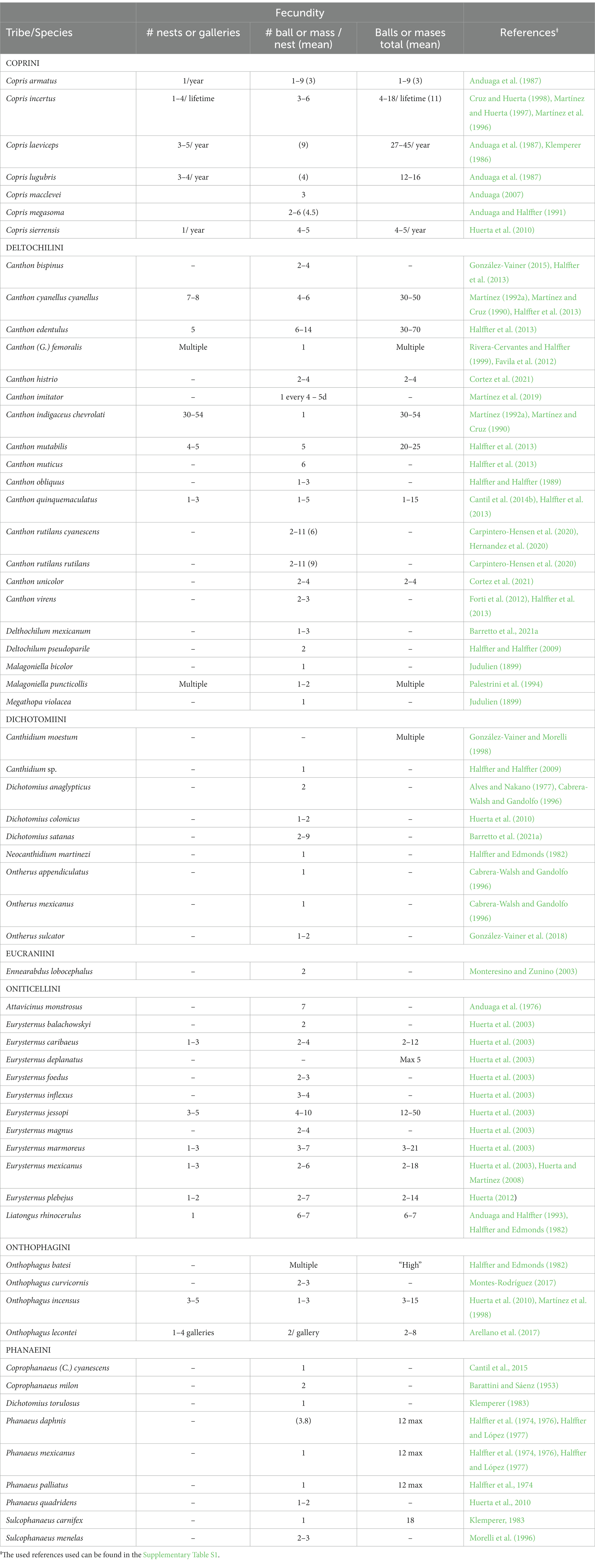
Table 5. Fecundity data of Scarabaeinae species.
Halffter and Edmonds (1982) considered that most Scarabaeinae species produce an average of 20 eggs per female over their lifetime, which still requires significant field and laboratory work to confirm. Although there is plenty of data regarding the nesting behavior of Neotropical species, only 18 species have information on the number of nests or brood balls, brood masses per nest, or galleries built by females over their lifetime or per breeding season. Moreover, 41 species have only partial information. Therefore, there are few species for which the relative fecundity per breeding season is known, obtained from the average number of balls or brood masses produced in each nest. The species known to produce an average of more than 20 eggs over their lifetime or per breeding season include four roller dung beetles: Canthon mutabilis (22.5), C. cyanellus (37.5), C. indigaceus chevrolati (42), and C. edentulus (50). On the other hand, those elaborating less than 10 balls or brood masses per breeding season are four species of Eurysternus, two of Copris, two of Onthophagus, and two of Canthon (Table 5). In most cases, only the number or type of nest balls or masses found in the field are reported, but further data are required to estimate their fecundity by season.
Nesting behavior patterns: Tunnellers and rollers
Pattern I
The Pattern-I nesting behavior comprises the largest number of records, having been observed in 28 Neotropical species to date, mainly of the genera Onthophagus (11 species), Dichotomius (6), and Canthidium Erichson (4). It has also been occasionally observed in species of Attavicinus Philips and Bell and Liatongus Reitter (Oniticellini); Ateuchus Weber, Neocanthidium, and Uroxys Westwood (Dichotomiini); Gromphas Brullé (Phanaeini) and Geotrupes (Geotrupidae; Supplementary Table S4).
The main characteristic of Pattern I is the so-called “brood mass,” which is packed in the blind bottom of a simple or branched gallery. The final shape of the brood mass is determined by the cavity, which may be cylindrical (i.e., “sausage-shaped”), oval, or spheroidal (Halffter and Edmonds, 1982). An exception is the nest mass of Dichotomius carbonarius, which has a peculiar structure and shape, composed of two connected parts, a lower spherical main structure, and an upper cylindrical protuberance (Dinghi et al., 2013). Also noteworthy is the brood mass of Neocanthidium martinezi, Edmonds and Halffter, which is pyriform, resembling a brood ball, resting loosely inside a subterranean cavity. However, it lacks the typical soil cover of brood balls produced by species with a Pattern II nesting behavior (Halffter and Edmonds, 1982).
Pattern-I nests are generally built below the food source. However, individuals of Canthidium megathopoides, Boucomont, may cut dung fragments and move them some distance, pushing them with their head and forelegs, before digging a gallery to bury them (Rodrigues and Flechtmann, 1997). This behavior has been extensively observed in several Phanaeini species with a Pattern-II nesting behavior, but not in Pattern-I species. Bisexual cooperation in food provisioning into the nest, whether simple or compound, has been observed in several species with a Pattern-I behavior (Supplementary Table S4). However, only females elaborate brood masses, with a single egg laid in each. Attavicinus monstrosus is an exception since the female can oviposit up to three eggs, 10 cm apart, along the sausage-shaped nest (Anduaga et al., 1976; Halffter and Edmonds, 1982).
Regarding the Neotropical Geotrupidae, the nest-building process has been studied in detail only in Geotrupes cavicollis Bates in Mexico. This species exhibits a Pattern-I nesting behavior with compound nests and bisexual cooperation, as described above (Halffter et al., 1980b, 1985). Moreover, Taurocerastes patagonicus Philippi records showed that they construct galleries up to 35 cm deep stocked with sausage-shaped droppings. These could be only for adult feeding; oviposition was not observed, and larvae of the species were found freely buried, not related to any food mass or gallery (Howden and Peck, 1987).
The majority of the species with Pattern-I behavior studied (72%) elaborate their nests with excrement, mainly bovine, and, to a lesser extent, with feces of other mammals (equines, goats, pigs, rodents, and humans). A considerably lower number of species, which inhabit tropical forests, use various material sources for building their nest masses: fruits, i.e., Onthophagus rhinolophus Harold, and Canthidium sp. (Halffter and Halffter, 2009; Sarges et al., 2012); decaying leaves, i.e., Dichotomius carbonarius (Dinghi et al., 2013); seeds, i.e., Canthidium laetum Harold, and Onthophagus orphnoides Bates (Halffter and Halffter, 2009); and carrion, i.e., Canthidium puncticolle Harold (Halffter and Edmonds, 1982). Other species that live associated with Atta Fabricius ant hills in anthropized subtropical and xerophytic scrub environments nest inside or under them, using their detritus as a food source for larvae (i.e., Ateuchus granigerum Harold Attavicinus monstrosus and Onthophagus rufescens Bates; Halffter and Matthews, 1966; Anduaga et al., 1976; Halffter and Halffter, 2009; See Supplementary Table S4). In addition, Onthophagus browni Howden and Carthwright, and Onthophagus coproides Horn construct their nests exclusively in burrows of the rodents Neotoma albigula Hartley and Thomomys umbrinus (Richardson), respectively, using their detritus and excrements (Anduaga and Halffter, 1991; Anduaga, 2007).
Pattern II
The Pattern-II nesting behavior has been described in 20 Neotropical species, of which 15 belong to the tribe Phanaeini, genera Bolbites Harold (1), Coprophanaeus Olsufieff (2), Diabroctis Gistel (1) Oxysternon Laporte (1), Phanaeus (6), and Sulcophanaeus Olsufieff (4). The remaining five species belong to Dichotomiini tribe, Ontherus Erichson (3) and Dichotomius (2) genera (Supplementary Table S4). Regarding the Phanaeini species with incomplete nest descriptions, particularly from the genera Coprophaneus, Phanaeus and Sulcophanaeus, we have considered that their nesting behavior corresponds to pattern II according to Halffter and Edmonds (1982).
In behavioral Pattern II, the excavation and subsequent food provisioning into the nesting gallery usually occur below or to one side of the food source. This characteristic allows classifying these nests as paracoprid. However, the transfer of large food fragments over the surface and a typical paracoprid nest construction has been observed in all the studied Phanaeus species. These fragments are pushed and rolled some distance with the head and prothorax or forelegs (“butting” behavior) before being buried. This can be done by the female alone or along with the male, depending on the species (Supplementary Table S4). Upon arrival at the appropriate site, the female starts excavating the gallery. Then the male and the female participate cooperatively in the food provisioning: the male cuts small fragments of food and deposits them at the entrance of the gallery, while the female introduces them into the nest, first by pulling and then pushing them inside (Halffter and López, 1977; Halffter and Edmonds, 1982; Price and May, 2009; Huerta et al., 2010). This butting behavior has also been recorded occasionally for Oxysternon conspicillatum Weber (Halffter and Matthews, 1966).
Bisexual cooperation in gallery construction and food provisioning occurs in 15 Phanaeini species of the genera Bolbites, Coprophanaeus, Phanaeus and Sulcophanaeus, but not in Oxysternon or Dichotomiini species with nesting pattern II. In Coprophanaeus ensifer, Sulcophanaeus leander (Waterhouse), and several species of Phanaeus, both sexes cooperate in both gallery excavation and food storage (Noriega, 2002; Price and May, 2009; Lira and Frizzas, 2022; Supplementary Table S4). In B. onitoides, Harold, S. carnifex (Linnaeus) and S. imperator (Chevrolat), the male introduces food fragments into the upper part of the gallery and the female carries them to the nest chamber to elaborate nest balls; males never enter the nest chamber (Klemperer, 1983; Cabrera-Walsh and Gandolfo, 1996). However, in several of these species, females can also nest alone. Nest care has not been observed in any species with a Pattern-II nesting behavior; the female leaves the nest after elaborating the last ball.
Finally, most species displaying Pattern-II build their nests with herbivorous mammal droppings. Exceptions are Phanaeus halffterorum Edmonds, which uses fungi and carrion in pine and oak forests in Mexico (Price and May, 2009), and Coprophanaeus species that use omnivorous and carnivorous mammal droppings or carrion in tropical forests and grasslands in South America (Barattini and Sáenz, 1953; Cantil et al., 2015; Lira and Frizzas, 2022).
Cases worth highlighting are the species of the tribe Eucraniini, endemic to the arid and semi-arid region of northwestern Argentina (South American Transition Zone). These burrowing species display a unique behavior: they carry dung pellets with their front legs and move forward with their middle and hind legs to bury them in a deep gallery excavated in advance; this operation is repeated several times. It has been suggested that this telephagic behavior is a strategy to exploit the scattered dry dung, mainly from rodents and camelids, by gathering and rehydrating it in the wetter bottom of tunnels (Zunino et al., 1989). The previous gallery excavation is a typical behavior of burrowing species (paracoprids); therefore, they have been considered paracoprids in this work (Supplementary Table S4).
Behavioral studies within the tribe Eucraniini have been conducted in 11 species of the genera Anomiopsoides Blackwelder, Ennearabdus Lansberge, Eucranium Brullé, and Glyphoderus Weswood (Zunino et al., 1989; Monteresino and Zunino, 2003; Ocampo, 2004; Ocampo and Philips, 2005; Ocampo, 2005, 2010; Supplementary Table S4). Bisexual cooperation in gallery construction and food provisioning has been recorded in eight species; brood-ball elaboration, only in two: Anomiopsoides heteroclyta (Blanchard; unpublished observation by A. Martínez, cited by Monteresino and Zunino, 2003; Figure 5) and Ennearabdus lobocephalus (Harold) (Monteresino and Zunino, 2003; Ocampo, 2010). The final nest structure for the other species is unknown. In E. lobocephalus, a composite paracoprid nest was built below bovine excrement under experimental conditions. The tunnel was filled like a sausage, and it bifurcated at the distal end, communicating with a lateral gallery containing stored food and a chamber containing two brood balls (Monteresino and Zunino, 2003). The observed characteristics of this paracoprid nest may correspond to a Pattern-II behavior. However, brood balls are not housed in separate chambers, as is typical in the nests corresponding to this pattern. On the other hand, Ocampo and Philips (2005) proposed that Eucranium arachnoides Brullé and E. planicolle, Burmeister, as well as others of the related genus Anomiopsoides, do not elaborate masses or brood-balls, but that larvae develop by feeding freely on fragmented and fermented pellets stored at the bottom of tunnels. These authors observed pupae of Anomiopsoides (species not indicated) under laboratory conditions, enclosed in cells made of organic material and sand. Undoubtedly, it is necessary to advance in the knowledge of the nesting behavior of Eucraniini species to define their behavioral patterns in further detail, considering that there may be significant differences in behavior between genera and species of this group.
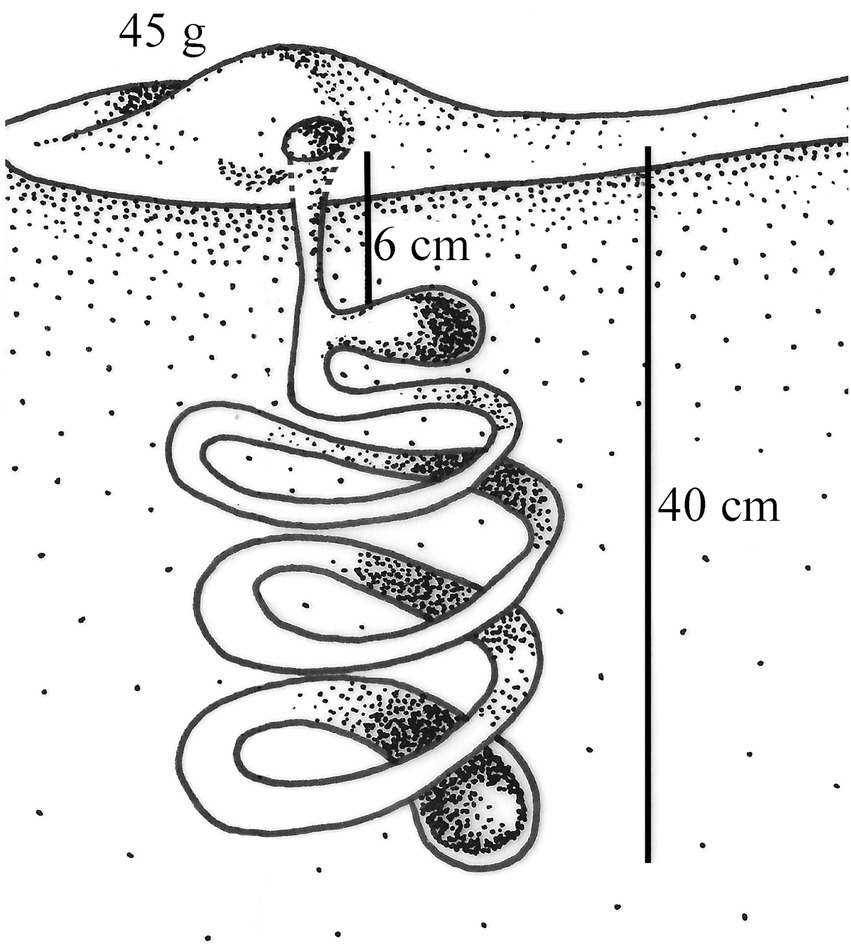
Figure 5. Anomiopsoides heteroclyta (Blanchard) nest (modified from Monteresino and Zunino, 2003).
Pattern III
The Pattern-III nesting behavior has been described for eight species of Neotropical dung beetles of the genus Copris (Tribe Coprini) and one species of the genus Eurysternus (Oniticellini; Supplementary Table S4) in studies conducted in Mexico. All Copris species are paracoprid and subsocial, and their behavior is highly uniform. Females make a nesting cake placed in a spacious chamber built at the end of a gallery. From the cake, brood balls are cut to build compound nests, which are taken care of until the emergence of the offspring. During the nest-care stage, the female adds an outer layer of soil to each brood balls; this, together with the care, protects the progeny from desiccation and fungal attack. There are slight variations between species, particularly regarding nest depth (Anduaga et al., 1987; Huerta and Halffter, 2000). Copris megasoma Matthews and Halffter and Copris macclevei Warner breed exclusively on gourd excrement (Anduaga and Halffter, 1991; Anduaga, 2007).
The nesting behavior of Eurysternus foedus Guérin-Méneville was classified as Pattern III by Huerta et al. (2005). This species builds underground nests where the female store food in a brood mass in a chamber. This mass is not divided into balls, but the female oviposits two to six eggs, where larvae develop. The female takes care of the brood mass until the progeny emerges. All studies on this species have been conducted in Mexico.
Patterns IV and V
Nesting Patterns IV and V correspond to roller species, which belong exclusively to the tribe Deltochilini in the Neotropics. Nesting Pattern IV has been recorded for 15 species, mainly in the genera Canthon (8) and Deltochilum (4) and occasionally in Canthochillum Chapin, Malagoniella Martínez, and Megathopa Eschscholtz. Nesting Pattern V has been recorded in nine species of Canthon and one of Malagoniella (Supplementary Table S4).
In most roller species, during the reproductive season, males and females meet at the food source, where they cut and elaborate balls which are rolled together; sometimes, the male initiates the rolling of a ball alone and attracts a female, completing the rolling and burial together. In Pattern IV, the resulting nest is simple, made up of a single brood ball shaped by the female, which she abandons after oviposition. In contrast, Pattern-V nests are compound, cared for by the female for some time, occasionally almost to the emergence of the offspring. Bisexual cooperation in nest care has been observed in some species. The male may remain with the female for nearly the entire duration of preimaginal development (i.e., Canthon cyanellus cyanellus, C. edentulus and Malagoniella puncticollis), while in other species, the male stays for a shorter time (i.e., Canthon bispinus, C. rutilans rutilans Laporte, C. rutilans cyanescens Harold and C. virens virens). During nest care, the beetle pair does not feed.
Behavior Patterns IV and V have recorded exceptions concerning nest care. Judulien (1899) described the nests of Malagoniella bicolor (cited as Megathopa bicolor) and Megathopa violacea (cited as Malagoniella intermedia) in detail, pointing out that these nests are simple and cared for by the female over approximately 2 months. It is necessary to confirm the simple condition of the nests of these species through further studies since this may vary. Malagoniella puncticollis normally builds simple nests but occasionally builds compound nests; in all cases, the female cares for them over up to 30 days. Canthon rutilans cyanescens builds a simple nest that is cared for by the couple until the larva hatches (Hernandez et al., 2020). Canthon bispinus, which makes characteristic Pattern-V nests, also cares for simple nests under laboratory conditions (González-Vainer, 2015). In contrast, in Canthon mutabilis, the pair separates and leaves the compound nest after the last oviposition. Males and females can build up to five nests in the same reproductive period under laboratory conditions (Halffter et al., 2013).
Parental care of the nest dramatically increases the survival of the offspring; in C. cyanellus cyanellus, brood balls that are left with no parental care are attacked readily by fungi (Favila, 1993). In C. bispinus, the female guards the nest for 30 days, resulting in a high progeny survival rate (92%; González-Vainer, 2015). Male C. c. cyanellus secretes chemicals in their abdominal glands that prevent the development of fly larvae, saprophagous and entomopathogenic fungi, and phytopathogenic bacteria in brood balls where the female lays the eggs (Favila et al., 2012). Chemical protection of the brood in roller beetles does not necessarily require the presence of the female or both parents in the nest. Canthon femoralis femoralis builds simple nests and leaves them on the surface; however, it probably has a defense and protection mechanism for brood balls by depositing more stable chemicals than those produced by C. c. cyanellus (Favila et al., 2012). In C. rutilans cyanescens, it was observed that the female defecates on the chamber walls before placing the egg and that the couple often walks on the ball, probably applying protective chemical secretions, until the larva hatches (Hernandez et al., 2020).
Brood balls of roller species are typically pear- or teardrop-shaped, usually covered by thick layers of soil, except for C. edentulus, which lacks a cover (Halffter et al., 2013). However, several studies do not indicate this aspect of the brood ball structure. The nests of most species are housed either in surface chambers covered with soil or leaves or underground, reaching 60 cm depth, as in the case of Deltochilum orbignyi (Halffter and Matthews, 1966).
Some species of roller dung beetles do not roll the brood balls but make them, whether simple or compound, in underground or surface chambers below the food source or at the soil-excrement interface. These species are Canthochilum histeroides, (Harold) Canthon edentulus, C. mutabilis, C. obliquus and D. orbignyi (Matthews, 1963; Halffter and Matthews, 1966; Halffter and Halffter, 1989; Halffter et al., 2013). In the case of C. mutabilis, this condition is facultative and depends on the size of the food source: if it is large, beetles make paracoprid nests; if it is small, beetles build telecoprid nests (Halffter et al., 2013).
Approximately 70% of the nests described for roller species are made exclusively with dung, mostly from herbivorous mammals. Nests built preferably with feces of omnivorous mammals have been recorded to a much lesser extent, as are the cases of C. femoralis femoralis, which displays a clear preference for monkey feces (Rivera-Cervantes and Halffter, 1999) and C. rutilans cyanescens which prefers Cerdocyon thous (crab-eating fox) dung to make the brood balls (Hernandez et al., 2020). Another species, Canthon rutilans rutilans build nests exclusively with dung from omnivorous and carnivorous mammals (Carpintero-Hensen et al., 2020). Only four species were found to build their nests with carrion: Canthon bispinus, C. cyanellus cyanellus, C. virens virens and C. virens aff. paraguayanus Balthasar (Halffter and Edmonds, 1982; Favila and Díaz, 1996; Halffter et al., 2013; Cantil et al., 2014a; González-Vainer, 2015). Canthon mutabilis has been the only recorded species capable of successfully building its nests with carrion and dung (Halffter et al., 2013), while Deltochilum pseudoparile Paulian builds nests with both feces and decomposed Brosimum alicastrum Swartz fruits (Halffter and Halffter, 2009; Halffter et al., 2013; Supplementary Table S3).
Concerning necrophagous species, most experimental studies have used beef or fish as food resources with which the species studied have built their nests. However, in the field, C. c. cyanellus has occasionally been observed feeding on carcasses of crickets and other dung beetles of the genera Phanaeus and Copris (Villalobos et al., 1998). In laboratory conditions, C. c. cyanellus can make food balls with the abdominal contents of crickets; even more, it can successfully build nests with the body of the diplopod Orthoporus ornatus (Girard) (Villalobos et al., 1998).
Another necrophagous species, Canthon virens virens, has been observed in the field feeding on the bodies of different arthropods such as Diplopods, Heteroptera, Coleoptera, and Lepidoptera. Under laboratory conditions, it can build nests with the bodies of crickets and cockroaches, both alive and dead, being an opportunistic predator (Halffter et al., 2013). In addition, several field studies have recorded that C. v. virens is an active predator of female ants of the genus Atta Fabricius during the short period of their nuptial flight after the ants clip their wings (Hertel and Colli, 1998; Vaz-De-Mello et al., 1998; Silveira et al., 2006; Forti et al., 2012). The female of C. v. virens catches an ant, decapitates it, rolls its body, buries it superficially, and makes two or three brood ball with it (Silveira et al., 2006; Forti et al., 2012).
Another necrophagous roller species that is an opportunistic predator is Canthon chalybeus, which preys on the snail Bulimulus apodemetes d’Orbigny (Martín et al., 2021). Males and females can kill a healthy snail, cut its shell, and roll its body into a ball. This beetle can also use snail carcasses. Canthon morsei has also been observed feeding on a wounded, still-living specimen of the diplopod Rhysodesmus dasypus (Gervais) (Villalobos et al., 1998), and it has been caught in pitfall traps baited with carcasses of the millipedes Amplinus bitumidus and Anadenobolus putealis (Bedoussac et al., 2007). Necrophagous species likely use the bodies of dead or dying invertebrates found in their habitat to build their nests. The capability of feeding and nesting with the flesh of wounded invertebrates could have been an evolutionary step before the development of the predation behavior of some Deltochilini (Villalobos et al., 1998; Martín et al., 2021).
Pattern VI
Pattern VI groups most species of the genus Eurysternus (Halffter, 1977; Halffter and Edmonds, 1982) with known reproductive behavior. This Neotropical genus includes a total of 53 species (Génier, 2009); the feeding and nesting behavior has been studied only in 10 of these, mainly in Mexico (Génier, 2009; Supplementary Table S3). Except for E. foedus, whose female builds brood masses, the known behavior of this genus consists of building balls solely for nesting. Males of the species studied have never been observed performing this behavior (Huerta et al., 2003). Adults feed directly above or below the food source. No straightforward relocation of food has been observed during nesting; the female builds the balls by getting under the food and moving within it, using her middle legs as oars (see Halffter et al., 1980a).
The Eurysternus species studied show several differences (Supplementary Table S4; Huerta et al., 2003). Some supposed subsocial species do not care for their offspring. The morphological characteristics of this genus are directly related to their nesting behavior and cannot be related to any of the evolutionary lines of the Scarabaeinae (paracoprids and telecoprids), nor can they be considered endocoprids because they do not nest within the dung mass but at the dung-soil interface (Figure 6; Halffter et al., 1980a).
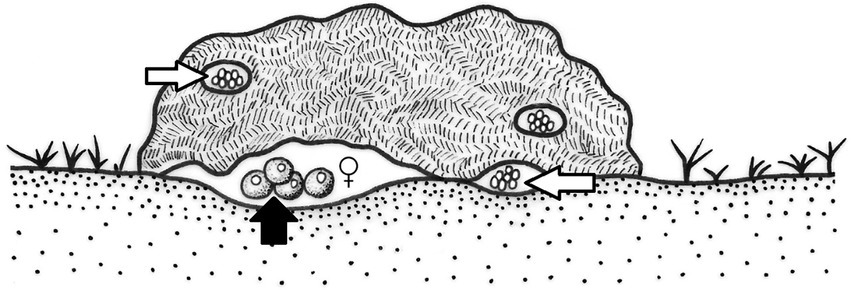
Figure 6. Oviposition sites of the Aphodiinae within the dung mass or at the dung-soil interface (white arrows), as also observed in some Eurysternus species (black arrow).
Aphodiinae reproductive behavior
Studies on the reproductive behavior of the subfamily Aphodiinae are few, consisting of scarce data on eight species of Aphodiini and three of Eupariini, mainly from Mexico. They are coprophagous dwellers that do not build nests but lay their eggs in the dung or at the dung-soil interface, depending on the species (Figure 6). The knowledge state of gonad maturity in both sexes helped to distinguish young from already mature individuals, as in Gonaphodiellus opisthius (Bates), Cephalocyclus hogei, (Bates) Planolinellus vittatus (Say), and the three Ataenius Harold species studied (Cruz et al., 2002; Martínez and Suárez, 2012; González-Vainer et al., 2018). The other species practically emerge mature and ready to reproduce. Most of the lifetime of Aphodiinae beetles is spent in the preimaginal stages or diapause; when adults emerge, they have a short period of activity lasting two or 3 months to reproduce, and then they die (Table 6).
Table 6. Pre-reproductive and reproductive behavior data, and preimaginal development time (days) for Neotropical Aphodiinae species.
Although there are data available on the phenology and reproductive cycles of several species of Mexican Aphodiini (see Martínez et al., 2022), information on their reproductive behavior is scarce, including mating and oviposition, since they are not easy to observe given their small size and because they spend most of their time inside the dung pats. Martínez et al. (2022) synthesize the available information on the reproductive cycles of some Aphodiini: Agrilinellus ornatus (Schmidt), G. opisthius, and Liothorax levatus (Schmidt), by characterizing mature, maturing, or immature individuals by ovary size variations, and the presence or absence of spermatozoa in the female spermatheca once they have already copulated. While in males, it is mainly due to the size of the glandular reservoir. In Eupariini, besides the anatomical variations mentioned to characterize the maturity state of individuals, in Ataenius perforatus Harold was possibly observed copulating inside the dung mass, at the end of which a spermatophore is formed in the vagina of females. This structure was also observed in the other two Ataenius species studied and only in a single Aphodiini species (Cephalocyclus hogei; Cruz et al., 2002). The number of eggs laid is known for some species, such as G. opisthius, which lays 15 to 18 agglutinated eggs in an underground chamber under the manure, and Blackburneus saylorea (Robinson), which lays eight eggs per nesting chamber in the two to three chambers it builds over its lifetime (Cruz et al., 2002). In Planolinellus vittatus, each egg is deposited in a laying chamber built within the manure still wet from the manure crust, although the total number of eggs laid per female is not reported (Martínez, 2008). On the other hand, in the three studied Ataenius species, eggs may be laid either under the manure or at the manure-soil interface; the number of eggs laid varies from 10 to 18 eggs per clutch. Finally, the time elapsed from preimaginal development to imago formation is known for only three Aphodiini species and all three Eupariini species. This time varies from 19 days to 66 days. The development time of each preimaginal stage is known for only four species; most of this time is spent in the larval stage (Table 6).
Geotrupidae breeding behavior
Studies on the reproductive behavior of Geotrupidae species are also scarce in the Neotropics. There are data on only four species belonging to two tribes — Ceratotrupini and Geotrupini —of which three are Mexican, and one is Chilean (Supplementary Table S4). All these species are coprophagous and paracoprid; the exception is Taurocerastes patagonicus, considered roller or telecoprid for moving food with its front legs; it is also a rare species that does not fly (Howden and Peck, 1987).
The species with the best-known reproductive behavior is Geotrupes cavicollis, observed under laboratory conditions (Halffter et al., 1980b, 1985); there is less information for the other species (Supplementary Table S4). This species also undergo maturation after emerging, and immature individuals build simple feeding galleries near or under the manure at a maximum depth of 10 cm. This period lasts 35 to 40 days after emergence; the reproductive period begins when the male and the female copulate at the gallery entrance for 10 min. Food is stored in galleries with the couple’s collaboration during this period. The nest comprises a maximum of five galleries, each with one egg in the distal end. In rare cases, two eggs can be laid per gallery, with soil in between the two eggs; the second was laid 24 h after the first. The subsequent oviposition took place 10 to 15 days later. Geotrupidae nest range between eight to 21 cm in depth, depending on the galleries and their radial or parallel distribution, with an average length of 21.5 cm. According to Halffter et al. (1980b, 1985), for the female to continue laying more than one or two eggs, the male must copulate and help with nest provisioning. Within each brood mass, preimaginal development starts with the embryo, and it takes 10 days for the larva to hatch; the larval stage lasts 210 days, followed by pupae, which take 44 days, and then the adult or imago remains in the nest for 30 to 40 days before emerging to the surface. Females die after 80 to 90 days (maximum 140 days) and only lay five eggs over their lifetime. However, females of G. cavicollis still have large oocytes not laid, differing from Scarabaeinae and Aphodiinae species, in which females show the reduction of their oocytes before dying (Martínez, 1992a,b; Martínez et al., 1996; Martínez, 2008; Martínez and Suárez, 2012).
Regarding the other species, Ceratotrupes fronticornis (Erichson) buries dog feces to build nests, but the type of the nest is not mentioned (Ramírez-Restrepo and Halffter, 2016). In Geotrupes (Halffterius) rufoclavatus Jekel, females oviposit in February because they have matured or maturing ovaries; however, no information is available on their oviposition behavior or the type of nest (Trotta-Moreu et al., 2007). Finally, in Taurocerastes patagonicus, one study describes that food is moved as pellets, then buried in simple, winding galleries inclined 70° and measuring 10 to 35 cm long (Howden and Peck, 1987). At the bottom, they accumulate food, forming a mass 5 to 7 cm long by 2 to 3 cm high. Nevertheless, it is not mentioned whether this food is only for adults or also for the young. Although male–female pairs were observed inside the gallery with food, no eggs were found. In another site, larvae were observed buried at 35 cm depth, although not associated with any food reserve or mass (Howden and Peck, 1987). Further studies are required to clarify this peculiar behavior.
Discussion
This review found that behavioral studies on dung beetles have been carried out on a low number of species (146), considering that the estimated richness of Scarabaeinae alone in the Neotropical region is approximately 1,250 species (Escobar, 2000). If the Aphodiinae and Geotrupidae are also included, behavioral studies have been conducted on less than 10% of the total dung beetle species. These studies are more abundant for Scarabaeinae (85%) compared to Aphodiinae (9%) and Geotrupidae (6%). The above is partly due to the number of Scarabaeinae species recorded in the Neotropics; more specifically, because only in the subfamily Scarabaeinae food relocation and nest construction are key features of the adult behavior in the vast majority of their species (Halffter and Edmonds, 1982); therefore, they constitute interesting model for behavioral studies. The pre-nesting stage has been studied in 101 Scarabaeinae species, the reproductive stage in 128 species, and the post-reproductive stage in only 18 species. Forty-two percent of the behavioral studies were based on field observations, 23% on laboratory observations, 30% under both field and laboratory conditions, and 5% were unspecified.
Behavioral studies on Neotropical Scarabaeinae species have been performed in 15 countries, mainly Mexico, Brazil, and Argentina. For the subfamily Aphodiinae, behavioral studies have been carried out only in Mexico and Uruguay, whereas some species of Geotrupidae have been studied only in Mexico and Chile. This review highlights the need to conduct further behavioral studies in several Central-South American and Caribbean countries, including Guatemala, El Salvador, Honduras, Nicaragua, Cuba, Haiti, Paraguay, Peru, Guyana, and Suriname. Isolated studies have been carried out in Venezuela, Bolivia, Ecuador, Chile, and Jamaica. The lack of behavioral studies for the Amazon rainforest has also become evident, particularly in Brazil, where such studies have addressed the southern area of this country. Although it is necessary to continue behavioral studies on Neotropical species from a biogeographical point of view, studies should also address the species inhabiting the Caribbean and Amazonian subregions (sensu Morrone et al., 2022). The above is increasingly urgent, given the accelerated loss of tropical rainforests. It is also necessary to fill the information gap on species inhabiting some provinces within the Chacoan subregion (e.g., Caatinga and Chaco), as well as on species living in the South American Transition Zone and the Andean region.
Another critical aspect is that at least one-half of the studied species were collected in open habitats (i.e., grasslands of temperate zones of Mexico, the Pampean region, and steppes of Argentina, or grasslands of Uruguay and the savanna in Brazil). These species are related to livestock and are burrowing species, with nesting Patterns I, II, and III. Only one-third of the studies have been conducted on both tropical and temperate forest species, mostly on burrowing species. Human activities profoundly modify native Neotropical environments; records of the dung beetle species studied indicate that only a small proportion of them (less than 10%) are habitat generalists that live in native forests, grasslands, and silvopastoral systems. Therefore, few dung beetle species may adapt to the continuous changes occurring in this region.
Feeding is crucial for dung beetles because it allows them to perform all their functions, including sexual maturation and reproduction. Intraspecific and interspecific male competition for food and females, which occurs during the pre-reproductive and reproductive stages, has been studied in a few burrowing and roller species. Although most of these studies have focused on fights between males, few have evaluated fights for these resources between females. Mate selection by males and females is fundamental for the reproductive success of dung beetle species. Fights have been analyzed in detail only for Canthon cyanellus. Evaluating the factors involved in the ability to win or lose a fight for limited resources and once this competition is transferred to the sperm level or the cryptic selection by females is still limited. Comparative studies will be highly relevant to explore the behavioral patterns within the context of evolutionary and ecological pressures the species have been exposed to during the pre-reproductive, reproductive, and post-reproductive stages.
Copulation has been described only in seven species. The duration of copulation has been recorded only in roller species because it generally occurs during the ball-rolling phase or in shallow nests. Few species have records regarding courtship before and during copulation and its incidence on reproductive success. Likewise, chemical communication has been investigated only in roller species; however, the effect of pheromones on intraspecific interactions and allelochemicals on interspecific interactions has been studied in a few roller species.
Although studies on nesting behavior in Scarabaeinae have been carried out on few species, most are representative of all the behavioral patterns proposed by Halffter and Edmonds (1982) and belong to almost all Neotropical tribes (e.g., Deltochilini, Coprini, Phanaeini, Dichotomiini, Oniticellini, and Onthophagini). Over the past 40 years, these studies have deepened our knowledge of behavioral patterns in dung beetles and revealed their plasticity. Subsocial behavior, the degree of development that varies among species, has also been studied in detail. This behavior has been analyzed mainly in Copris and some Canthon and Eurysternus species, which take care of the nest until a particular stage of preimaginal development. Conversely, the incipient knowledge about the nesting behavior of the Eucraniini has evidenced that it does not conform to any known patterns, highlighting it as a group of great interest for study.
This work has identified a few species for which there is a considerable accumulation of knowledge, ideal for answering questions or testing particular hypotheses. Canthon cyanellus is a good model for behavioral studies (Favila, 2001), also known as a model system (Dugatkin, 2001). Undoubtedly, studying the reproductive behavior of dung beetles in their environment, where their behavior is displayed naturally, is fundamental for understanding the ecological processes surrounding the variation in the reproductive behavior of each species. Transferring these studies to the laboratory allows us to have controlled conditions and test hypothesis that can be quantified regarding the effect of previously identified variables that affect or regulate behavioral patterns. Both field and laboratory data are essential for understanding the ecological processes and evolutionary contexts of Neotropical dung beetles. However, laboratory studies can only be conducted for some species, whereas only some are sound model systems for behavioral studies. The above depends on several conditions that must be controlled to maintain the reproductive behavior of the studied dung beetle species, which is hard to achieve. This is one of the main reasons why conclusive laboratory results have yet to be obtained for some species of dung beetles.
Finally, it is necessary to continue studying the behavioral biology and ecology of dung beetles in their original habitats in Neotropical forests since these habitats are being heavily anthropized, and information on many species is still lacking (about 88% of the richness estimated of Scarabaeinae). We risk losing species or altering behavioral patterns due to the effects of these macro and micro environmental changes on the Neotropical region, which are likely to affect species’ survival and the environmental services provided by this group of beetles.
Author contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
JR thanks the Consejo Nacional de Ciencia y Tecnología (CONACYT) for PhD grant # 70982.
Acknowledgments
Thanks to Gonzalo Halffter, pioneer in dung beetle behavior studies. We also thank Sara Lariza Rivera Gasperín for the elaboration of the figures. María Elena Sánchez-Salazar edited the manuscript in English.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1102477/full#supplementary-material
References
Alves, S. B., and Nakano, O. (1977). Biologia e importancia economica do Dichotomius anaglypticus (Mannerheim, 1829) (Coleoptera, Scarabaeidae). [dissertation/thesis]. [São Paulo]: Universidade de São Paulo, Piracicaba, Brazil. Available at: https://repositorio.usp.br/item/000722614
Anduaga, S. (2007). Nuevos registros de escarabajos coprófagos (Coleoptera: Scarabaeidae) en detritus de madrigueras de Neotoma albigula Hartley (Rodentia: Muridae). Acta Zool. Mex. 23, 143–144. doi: 10.21829/azm.2007.233600
Anduaga, S., and Halffter, G. (1991). Escarabajos asociados a madrigueras de roedores (Coleoptera: Scarabaeidae, Scarabaeinae). Folia Entomol. Mex. 81, 185–197.
Anduaga, S., and Halffter, G. (1993). Nidificación y alimentación en Liatongus rhinocerulus (Bates) (Coleoptera: Scarabaeidae: Scarabaeinae). Acta Zool. Mex. 57, 1–14. doi: 10.21829/azm.1993.55571677
Anduaga, S., Halffter, V., and Halffter, G. (1976). Etograma de Liatongus monstrosus (Bates) (Coleoptera: Scarabaeidae). Folia Entomol. Mex. 36, 48–49.
Anduaga, S., Halffter, G., and Huerta, C. (1987). Adaptaciones ecológicas de la reproducción en Copris. Boll. Mus. Reg. Sci. Nat. Torino. 5, 45–65.
Arellano, L., Castillo-Guevara, C., Huerta, C., Germán-García, A., and Lara, C. (2017). Nesting biology and life history of the dung Onthophagus lecontei (Coleoptera: Scarabaeinae) Anim. Biol. 67, 41–52. doi: 10.1163/15707563-00002520
Barattini, L. P., and Sáenz, A. C. (1953). Nuevos aportes para el conocimiento del desarrollo de Phanaeus milon (Blanch.) (Col.: Scarab.). Rev. Soc. Entomol. Argen. 16, 25–30.
Barretto, J., Baena, M. L., Huesca Domínguez, I., and Escobar, F. (2021a). Spatiotemporal variation in the adult sex ratio, male aggregation, and movement of two tropical cloud forest dung beetles. Curr. Zool. 2021:zoab101. doi: 10.1093/cz/zoab101
Barretto, J., Cruz, M., and Escobar, F. (2021b). Annual reproductive phenology of the coprophagous beetle Dichotomius satanas (Coleoptera: Scarabaeidae, Scarabaeinae) of the cloud forest in eastern Mexico. Can. Entomol. 153, 157–171. doi: 10.4039/tce.2020.75
Bedoussac, L., Favila, M. E., and López, R. M. (2007). Defensive volatile secretions of two diplopod species attract the carrion ball roller scarab Canthon morsei (Coleoptera: Scarabaeidae). Chemoecology 17, 163–167. doi: 10.1007/s00049-007-0375-y
Bellés, X., and Favila, M. E. (1983). Pretection chimique du nid chez Canthon cyanellus cyanellus LeCorte (Col. Scarabaeidae). Bull. Soc. Ent. Fr. 88, 602–607. doi: 10.3406/bsef.1983.18093
Berglund, A., Bisazza, A., and Pilastro, A. (1996). Armaments and ornaments: an evolutionary explanation of traits of dual utility. Biol. J. Linn. Soc. 58, 385–399. doi: 10.1111/j.1095-8312.1996.tb01442.x
Briffa, M., and Sneddon, L. U. (2007). Physiological constraints on contest behaviour. Funct. Ecol. 21, 627–637. doi: 10.1111/j.1365-2435.2006.01188.x
Cabrera-Walsh, G., and Gandolfo, G. (1996). Nidification of thirteen common argentine dung beetles (Scarabaeidae: Scarabaeinae). Ann. Entomol. Soc. Am. 89, 581–588. doi: 10.1093/aesa/89.4.581
Cambefort, Y., and Hanski, I. (1991). “Dung beetles population biology” in Dung beetle ecology. eds. I. Hamski and Y. Cambefort (Princeton, NJ: Princeton University Press), 36–50. doi: 10.1515/9781400862092.36
Cantil, L. F., Sánchez, M. V., Dinghi, P. A., and Genise, J. F. (2014b). Food relocation behavior, nests, and brood balls of Canthon quinquemaculatus Laporte de Castelnau (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 68, 199–208. doi: 10.1649/0010-065X-68.2.199
Cantil, L. F., Sánchez, M. V., and Genise, J. F. (2014a). The nest and brood ball of Canthon (Canthon) virens aff. paraguayanus Balthasar (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 68, 384–386. doi: 10.1649/072.068.0306
Cantil, L. F., Sánchez, M. V., Sarzetti, L., Molina, A., and Genise, J. F. (2015). Nest and brood balls of Coprophanaeus (Coprophanaeus) cyanescens (Olsoufieff, 1924) (Coleptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 69, 153–158. doi: 10.1649/0010-065X-69.1.153
Cárdenas-Castro, E., and Páez-Martínez, A. (2017). Comportamiento reproductivo de coleópteros coprófagos (Coleoptera: Scarabaeidae) en condiciones de laboratorio. Rev. Cien. Agr. 34, 74–83. doi: 10.22267/rcia.173401.64
Carpintero-Hensen, M., Lobo, J. M., and Hernandez, M. I. M. (2020). Differences in the reproductive behaviour and larval development of two Canthon rutilans subspecies reinforce their thermal regional segregation. Ent. Exp. Appl. 168, 827–835. doi: 10.1111/eea.12984
Chamorro-Florescano, I. A., and Favila, M. E. (2008). Male reproductive status affects contest outcome during nidification in Canthon cyanellus cyanellus LeConte (Coleoptera: Scarabaeidae). Behaviour 145, 1811–1821. doi: 10.1163/156853908786279637
Chamorro-Florescano, I. A., and Favila, M. E. (2009). The reproductive status of both sexes affects the frequency of mating and the reproductive success of males in the ball roller beetles Canthon cyanellus cyanellus (Coleoptera: Scarabaeidae). Behaviour 146, 1499–1512. doi: 10.1163/156853909X445560
Chamorro-Florescano, I. A., and Favila, M. E. (2016). Male success in intrasexual contests extends to sperm competition level: the case of the dung roller beetle. Ethology 122, 53–60. doi: 10.1111/eth.12443
Chamorro-Florescano, I. A., Favila, M. E., and Macías-Ordóñez, R. (2011). Ownership, size and reproductive status affect the outcome of food ball contests in a dung roller beetle: when do enemies share? Evol. Ecol. 25, 277–289. doi: 10.1007/s10682-010-9428-8
Chamorro-Florescano, I. A., Favila, M. E., and Macías-Ordoñez, R. (2017). Contests over reproductive resources in female roller beetles: outcome predictors and sharing as an option. PLoS One 12, 1–14. doi: 10.1371/journal.pone.0182931
Cortez, V. (2013). Ecología química y perspectivas de su aplicación en la conservación de la biodiversidad. Cuad. Biodiver. Univ. Alicante. 41, 16–21. http://dx.doi.org/10.14198/cdbio.2013.41.03
Cortez, V., Favila, M. E., Verdú, J. R., and Ortiz, A. J. (2012). Behavioral and antennal electrophysiological responses of a predator ant to the pygidial gland secretions of two species of Neotropical dung roller beetles. Chemoecology 22, 29–38. doi: 10.1007/s00049-011-0095-1
Cortez, V., Verdú, J. R., Ciares, W., and Halffter, G. (2021). Nesting behaviour of Canthon unicolor and C. histrio: a new subsocial nesting variation in dung beetles (Coleoptera: Scarabaeidae: Deltochilini). J. Nat. Hist. 55, 2187–2197. doi: 10.1080/00222933.2021.1989072
Cortez, V., Verdú, J. R., Ortiz, A. J., Trigos, Á. R., and Favila, M. E. (2015). Chemical diversity and potential biological functions of the pygidial gland secretions in two species of Neotropical dung roller beetles. Chemoecology 25, 201–213. doi: 10.1007/s00049-015-0189-2
Cruz, R. M., and Huerta, C. C. (1998). Comportamiento y actividad reproductora de los machos de Copris incertus Say (Coleoptera Scarabaeidae: Scarabaeinae). Acta Zool. Mex. (n.s.) 74, 163–173. doi: 10.21829/azm.1998.74741723
Cruz, R. M., and Martínez, M. I. (2002). Data on nesting and preimaginal development in two Mexican species of Ataenius Harold, 1867 (Coleoptera, Scarabaeoidea, Aphodiidae; Eupariinae). Folia Ent. Mex. 41, 1–5.
Cruz, R. M., Martínez, M. I., and Alvarado, O. M. (2002). Population and reproductive features of Aphodius (Trichaphodius) opisthius and Cephalocyclys hogei Bates (Coleoptera, Aphodiidae: Aphodiinae). Coleopt. Bull. 56, 21–235. doi: 10.1649/0010-065X(2002)056%5b0221:PARFOA%5d2.0.CO;2
Dinghi, P. A., Sánchez, M. V., Cantil, L. F., Sarzetti, L. C., and Genise, J. F. (2013). Leaf-litter brood chambers in Dichotomius (Luederwaldtinia) carbonarius (Mannerheim, 1829) (Coleoptera: Scarabaeidae): a novel behavior for dung beetles. Coleopt. Bull. 67, 388–396. doi: 10.1649/0010-065X-67.3.388
Dugatkin, L. A. (2001). Model systems in behavioral ecology: integrating conceptual, theoretical, and empirical approaches (Vol. 70). Princeton, NJ: Princeton University Press.
Eberhard, W. (1996). Female control: sexual selection by cryptic female choice (Vol. 17). Princeton NJ: Princeton University Press. doi: 10.1515/9780691207209
Emlen, D. J. (1997). Alternative reproductive tactics and male-dimorphism in the horned beetle Onthophagus acuminatus (Coleoptera: Scarabaeidae). Behav. Ecol. Sociobiol. 41, 335–341. doi: 10.1007/s002650050393
Escobar, F. (2000). “Diversidad y distribución de los escarabajos del estiércol (Coleoptera. Scarabaeidae: Scarabaeinae) de Colombia” in Hacia un Proyecto CYTED para el inventario y estimación de la diversidad entomológica en Iberoamérica: PiIBES-2000. eds. F. Martín-Piera, J. J. Morrone, and A. Melic (Zaragoza: m3m Monografías Tercer Milenio), 197–210.
Favila, M. E. (1988). Comportamiento durante el periodo de maduración gonádica en un escarabajo rodador (Coleoptera: Scarabaeidae, Scarabaeinae). Folia Entomol. Mex. 76, 55–64.
Favila, M. E. (1993). Some ecological factors affecting the life-style of Canthon cyanellus cyanellus (Coleoptera Scarabaeidae): an experimental approach. Ethol. Ecol. Evol. 5, 319–328. doi: 10.1080/08927014.1993.9523019
Favila, M. E. (2001). Historia de vida y comportamiento de un escarabajo necrófago: Canthon cyanellus cyanellus LeConte (Coleoptera: Scarabaeinae). Folia Entomol. Mex. 40, 245–278.
Favila, M. E. (2012). Historical, biogegraphical and ecological factors explain the success of some native dung beetles after the introduction of cattle in Mexico. Pastos 42, 161–181.
Favila, M., Cortez, V., Verdú, J. R., Ortíz, A. J., and Trigos, A. (2016). “Diversidad y función de los semioquímicos en los escarabajos rodadores del estiércol de la subfamilia Scarabaeinae” in Ecología Química y Alelopatía: Avances y Perspectivas. eds. A. L. Anaya-Lang, F. J. Espinosa-García, and M. J. Reigosa-Roger (México: Instituto de Ecología, UNAM y Plaza y Valdés SA de CV), 641–677.
Favila, M. E., and Díaz, A. (1996). Canthon cyanellus cyanellus LeConte (Coleoptera: Scarabaeidae) makes a nest in the field with several brood balls. Coleopt. Bull. 50, 52–60.
Favila, M. E., Nolasco, J., Chamorro-Florescano, I., and Equihua, M. (2005). Sperm competition and evidence of sperm fertilization patterns in the carrion ball-roller beetle Canthon cyanellus cyanellus LeConte (Scarabaeidae: Scarabaeinae). Behav. Ecol. Sociobiol. 59, 38–43. doi: 10.1007/s00265-005-0006-y
Favila, M. E., Ortiz-Domínguez, M., Chamorro-Florescano, I., and Cortez-Gallardo, V. (2012). “Comunicación química y comportamiento reproductor de los escarabajos rodadores de estiércol (Scarabaeinae: Scarabaeini): aspectos ecológicos y evolutivos, y sus posibles aplicaciones” in Temas Selectos en Ecología Química de Insectos, eds. Rojas, J. C., and Malo, E. A. (México: El Colegio de la Frontera Sur), 141–164.
Forti, L., Rinaldi, I., Camargo, R., and Fujihara, R. (2012). Predatory behavior of Canthon virens (Coleoptera: Scarabaeidae): a predator of leafcutter ants. Psyche 2012, 1–5. doi: 10.1155/2012/921465
Génier, F. (2009). Le genre Eurysternus Dalman, 1824 (Scarabaeidae: Scarabaeinae: Oniticellini): Révision taxonomique et clés de détermination illustrées. Bulgaria: Pensoft Publishers.
Ginzel, M. D. (2010). “Hydrocarbons as contact pheromones of longhorned beetles (Coleoptera: Cerambycidae)” in Insect hydrocarbons: Biology, biochemistry and chemical ecology. eds. G. J. Blomquist and A. G. Bagnères (New York: Cambridge University Press), 375–389.
González-Vainer, P. (2015). Feeding, reproductive, and nesting behavior of Canthon bispinus Germar (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 69, 61–72. doi: 10.1649/0010-065X-69.1.61
González-Vainer, P., and Baruffaldi, L. (2006). Nesting behavior and life cycle of Uroxys terminalis Waterhouse 1891 (Coleoptera: Scarabaeidae) under laboratory conditions. Coleopt. Bull. 60, 307–311.
González-Vainer, P., and Morelli, E. (1998). Estados preimaginales, nidificación y fenología de Canthidium (E) moestum Harold, 1867 (Coleoptera: Scarabaeidae: Coprini). Acta Zool. Mex. (n.s.) 73, 155–165. doi: 10.21829/azm.1998.73731733
González-Vainer, P., and Morelli, E. (1999). Phenology and biology of the dung beetle Onthophagus hirculus Mannerheim (Coleoptera: Scarabaeidae). Coleopt. Bull. 53, 303–309.
González-Vainer, P., Cantil, L., and Genise, J. (2018). Plasticity in nesting behavior of Ontherus sulcator (Scarabaeidae). Coleopt. Bull. 72, 203–208. doi: 10.1649/0010-065X-72.1.203
Halffter, G. (1977). Evolution of nidification in the Scarabaeinae (Coleoptera, Scarabaeidae). Quaest. Ent. 13, 231–253.
Halffter, G. (1997). “Subsocial behavior in Scarabainae beetles” in The evolution of social behavior in insects and arachnids. eds. J. C. Choe and B. J. Crespi (Cambridge: Cambridge University Press), 237–259.
Halffter, G., Cortez, V., Gómez, E. J., Rueda, C. M., Ciares, W., and Verdú, J. R. (2013). A review of subsocial behavior in Scarabaeinae rollers (Insecta: Coleoptera): an evolutionary approach. Zaragoza: m3m Monografías Tercer Milenio.
Halffter, G., and Edmonds, D. W. (1982). The nesting behavior of dung beetles (Scarabaeinae) an ecological and evolutive approach. México: Instituto de Ecología.
Halffter, G., and Halffter, V. (1989). Behavioral evolution of the non-rolling roller beetles (Coleoptera: Scarabaeidae; Scarabaeinae). Acta Zool. Mex. 32, 1–53. doi: 10.21829/azm.1989.31321937
Halffter, G., and Halffter, V. (2009). Why and where coprophagous beetles (Coleoptera, Scarabaeinae) eat seeds, fruits or vegetable detritus. Bol. Soc. Entomol. Arag. 45, 1–22.
Halffter, G., Halffter, V., and Huerta, C. (1980a). Mating and nesting behavior of Eurysternus (Coleoptera: Scarabaeinae). Quaest. Ent. 16, 599–620.
Halffter, G., Halffter, V., and Lopez, Y. (1974). Phanaeus behaviour: food transportation and bisexual cooperation. Environ. Entomol. 3, 341–345. doi: 10.1093/ee/3.2.341
Halffter, G., and López, G. Y. (1977). Development of the ovary and mating behavior in Phanaeus. Ann. Entomol. Soc. Am. 70, 203–213. doi: 10.1093/aesa/70.2.203
Halffter, G., López, G. I., and Halffter, V. (1976). Comportamiento de Phanaeus (Coleoptera, Scarabaeidae): cópula. Folia Entomol. Mex. 36, 44–45.
Halffter, V., López-Guerrero, Y., and Halffter, G. (1980b). Comportamiento sexual de Geotrupes (Coleoptera: Scarabaeidae: Geotrupinae). Folia Entomol. Mex. 45, 63–65.
Halffter, V., López-Guerrero, Y., and Halffter, G. (1985). Nesting and ovarian development in Geotrupes cavicollis Bates (Coleoptera: Scarabaeidae). Acta Zool. Mex. 7, 1–28. doi: 10.21829/azm.1985.271643
Halffter, G., and Matthews, E. G. (1966). The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera, Scarabaeidae). Folia Entomol. Mex. 12-14, 1–312.
Hanski, I., and Cambefort, Y. (1991). Dung beetle ecology. Princeton NJ: Princeton University Press.
Hernandez, M. I. M., Niero, M. M., Schumacher, F., and Wuerges, M. (2020). Feeding and reproductive behavior of the dung beetle Canthon rutilans cyanescens (Coleoptera: Scarabaeinae). Rev. Bras. Entomol. 64:e20190007. doi: 10.1590/1806-9665-RBENT-2019-0007
Hertel, F., and Colli, G. R. (1998). The use of leaf-cutter ants, Atta laevigata (Smith) (Hymenoptera: Formicidae), as a substrate for oviposition by the dung beetle Canthon virens Mannerheim (Coleoptera: Scarabaeidae) in Central Brazil. Coleopt. Bull. 52, 105–108. https://www.jstor.org/stable/4009370
Howard, R. W., and Blomquist, G. J. (2005). Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 50, 371–393. doi: 10.1146/annurev.ento.50.071803.130359
Howden, H. F., and Peck, S. B. (1987). Adult habits, larval morphology, and phylogenetic placement of Taurocerastes patagonicus Philippi (Scarabaeidae: Geotrupinae). Can. J. Zool. 65, 329–332. doi: 10.1139/z87-051
Huerta, C. (2012). Nesting behavior of Eurysternus plebejus Harold (Coleoptera: Scarabaeidae) under laboratory conditions. Coleopt. Bull. 66, 51–54. doi: 10.1649/072.066.0113
Huerta, C., Anduaga, S., and Halffter, G. (1981). Relaciones entre nidificación y ovario en Copris (Coleoptera, Scarabaeidae, Scarabaeinae). Folia Entomol. Mex. 47, 141–170.
Huerta, C., Anduaga, S., López-Portillo, J., and Halffter, G. (2010). Use of food and space by tunneler dung beetles (Coleoptera; Scarabaeinae) during reproduction. Environ. Entomol. 39, 1165–1169. doi: 10.1603/EN09321
Huerta, C., and Halffter, G. (2000). Factores involucrados en el comportamiento subsocial de Copris (Coleoptera: Scarabaeidae: Scarabaeinae). Folia Entomol. Mex. 108, 95–120.
Huerta, C., Halffter, G., and Halffter, V. (2005). Nidifiton in Eurysternus foedus Guérin-Méneville: its relationship to other dung beettle nesting paterns (Coleoptera: Scarabaeidae: Scarabaeinae). Folia Entomol. Mex. 44, 74–84.
Huerta, C., Halffter, G., Halffter, V., and López, R. (2003). Comparative analysis of reproductive and nesting behavior in several species of Eurysternus Dalman (Coleoptera, Scarabaeinae, Eurysternini). Acta Zool. Mex. 88, 1–41. doi: 10.21829/azm.2003.88881788
Huerta, C., and Martínez, M. I. (2008). Morphological changes in reproductive organs and neuroendocrine centers related to nesting, mating, and larvicide behavior in Eurysternus mexicanus Harold (Scarabaeinae: Eurysternini). Coleopt. Bull. 62, 123–132. doi: 10.1649/1024.1
Huerta, C., Martínez, M. I., Montes de Oca, E., Cruz-Rosales, M., and Favila, M. E. (2013). “The role of dung beetles in the sustainability of pasture and grasslands” in Ecological dimensions for sustainable socio economic development. eds. A. R. Yáñez-Arancibia, R. Dávalos Sotelo, J. W. Day, and E. Reyes (Boston: WIT Press Southampton), 441–463.
Ix-Balam, M. A., Oliveira, M. G. A., Louzada, J., McNeil, J. N., and Lima, E. (2018). The rolling of food by a dung beetles affects the oviposition of competing flies. Insects 9:92. doi: 10.3390/insects9030092
Judulien, F. (1899). Quelques notes sur plusieurs coprophages de Buenos Aires. Rev. Mus. Plata Argent. 9, 371–380.
Klemperer, H. G. (1983). Brood ball construction by the non-brooding Coprini Sulcophanaeus carnifex and Dichotomius torulosus (Coleoptera, Scarabaeidae). Ecol. Entomol. 8, 61–68. doi: 10.1111/j.1365-2311.1983.tb00483.x
Klemperer, H. G. (1986). Life history and parental behavior of a dung beetle from neotropical rainforest, Copris laeviceps (Coleoptera, Scarabaeidae). J. Zool. Lond. 209, 319–326. doi: 10.1111/j.1469-7998.1986.tb03594.x
Lira, L. A., and Frizzas, M. R. (2022). Behavioral aspects of Coprophanaeus ensifer (Germar) (Coleoptera: Scarabaeidae: Scarabaeinae) associated with pig carcasses in Central Brazil. Neotrop. Entomol. 51, 65–72. doi: 10.1007/s13744-021-00922-x
López-Guerrero, Y., and Halffter, G. (2000). Evolution of the spermatheca in the Scarabaeoidea (Coleoptera). Frag. Entomol. 32, 225–285.
López, A. D., Halffter, G., and Vaz-de-Mello, F. (2009). Nesting behavior in Trichillum Harold, 1868 and related genera (Coleoptera: Scarabaeidae: Scarabaeinae: Ateuchini: Scatimina): a primitive process or a loss of nidification? Coleopt. Bull. 63, 289–297. doi: 10.1649/1132.1
Martín, C. M., Guanuco, A. V., Cortez, V., and Verdú, J. R. (2021). First observation on the predation of a non-arthropod species by a dung beetle species: the case of Canthon chalybaeus and the snail Bulimulus apodemetes. PLoS One 16:e0258396. doi: 10.1371/journal.pone.0258396
Martínez, M. I. (1992a). Données comparatives sur l’activité reproductrice de Canthon indigaceus chevrolati Harold et Canthon cyanellus cyanellus LeConte (Coleoptera, Scarabaeidae). Ann. Soc. Entomol. Fr. 28, 397–408.
Martínez, M. I. (1992b). L'activité ovarienne pendant la vie imaginale chez deux especies de Canthon (Coleoptera, Scarabaidae). Boll. Mus. Reg. Sci. Nat. Torino. 10, 367–386.
Martínez, M. I. (2005). Abundancias poblacionales y ciclos reproductivos de tres especies de escarabajos estercoleros (Coleoptera: Aphodiinae: Aphodiini) del volcán Cofre de Perote, Veracruz, México. Folia Ent. Mex. 44, 27–36.
Martínez, M. I. (2008). Bionomía del escarabajo estercolero Planolinellus vittatus (Say, 1825) (Coleoptera: Aphodiinae) en el Volcán Cofre de Perote, Veracruz. México. Dugesiana 15, 131–140. doi: 10.32870/dugesiana.v15i2.3893
Martínez, M. I., and Alvarado, O. M. (2001). Comportamiento de ovisposición en Aphodius opisthius Bates 1887 (Coleoptera: Scarabaeoidea: Aphodiidae). Elytron 15, 73–78.
Martínez, M. I., and Cruz, R. M. (1990). Cópula, función ovárica y nidificación en Canthon Hoffmannsegg (Coleoptera: Scarabaeidae). Elytron 4, 161–169.
Martínez, M. I., Dellacasa, M., Lumaret, J. P., and Dellacasa, G. (2022). Phenology and reproductive cycles in Mexican aphodiine dung beetles (Coleoptera: Scarabaeidae: Aphodiinae: Aphodiini). Ann. Soc. Entomol. Fr. 58, 173–185. doi: 10.1080/00379271.2022.2060859
Martínez, I., Deloya, C., and Dellacasa, M. (2001). Anatomical and functional data on female and male reproductive systems of some dung beetle species of Aphodiinae and Eupariinae of Mexico (Coleoptera: Scarabaeoidea: Aphodiidae). Proc. Entomol. Soc. Wash. 103, 227–248.
Martínez, M. I., Escobar, F., Almendarez, S., Martínez, A. K., González, G. L., and González-Tokman, D. (2019). Nuevos datos sobre la distribución y la biología reproductiva de Canthon imitator Brown, 1946 (Coleoptera: Scarabaeidae. Scarabaeinae). Acta Zool. 35, 1–7. doi: 10.21829/azm.2019.3502096
Martínez, M. I., and Huerta, C. (1997). Coordinated activity of the ovary, pars intercerebralis and corpus allatum during the prenesting and nesting cycles of Copris incertus Say (Coleoptera Scarabaeidae: Scarabaeinae). Coleopt. Bull. 51, 351–363.
Martínez, M. I., Huerta, C., and Cruz, R. M. (1996). Comportamiento reproductor en hembras de Copris incertus Say (Coleoptera, Scarabaeidae). Bull. Soc. Ent. Fr. 101, 121–130. doi: 10.3406/bsef.1996.17225
Martínez, M. I., Montes de Oca, T. E., and Cruz, R. M. (1998). Contribución al conocimiento de la biología del escarabajo coprófago Onthophagus incensus Say (Coleoptera: Scarabaeidae: Scarabaeinae): datos ecológicos y reproductivos en relación a su fenología. Folia Ent. Mex. 103, 1–13.
Martínez, M. I., and Suárez, M. T. (2012). Ciclo reproductivo de Blackburneus saylorea (Robinson, 1940) (Coleoptera: Aphodiinae), escarabajo coprófago de la ladera oriental del Volcán Cofre de Perote, Veracruz. México. Dugesiana 18, 181–187. doi: 10.32870/dugesiana.v18i2.4025
Martínez, M. I., and Trotta-Moreu, N. (2010). Comparative study of Mexican Geotrupini (Coleoptera: Scarabaeoidea: Geotrupidae) reproductive systems, with taxonomic commentaries. Coleopt. Bull. 64, 129–140. doi: 10.1649/072.064.0209
Matthews, E. G. (1963). Description of the larva and pupa of Canthochilum histeroides (Harold) with notes on its biology (Coleoptera: Scarabaeidae). Coleopt. Bull. 17, 110–116.
Monteresino, E., and Zunino, M. (2003). “Sobre el comportamiento de alimentación y nidificación de Eucraniini (Coleoptera Scarabaeidae: Scarabaeinae)” in Escarabeidos de Latinoamérica: Estado del conocimiento, comp. eds. G. Onore, P. Reyes-Castillo, and M. Zunino (Zaragoza: m3m Monografías Tercer Milenio), 75–80.
Montes-Rodríguez, J. M. (2017). Nidificación del escarabajo coprófago Onthophagus curvicornis Latreille, 1811 (Coleoptera: Scarabaeidae) en condiciones de laboratorio. Rev. Fac. Cien. 6, 20–28. doi: 10.15446/rev.fac.cienc.v6n2.64004
Morelli, E., González-Vainer, P., and Canziani, C. (1996). Nidificación, ciclo de vida y estados preimaginales de Sulcophanaeus menelas (Laporte, 1840) (Coleoptera: Scarabaeidae). Elytron 10, 11–22.
Morrone, J. J., Escalante, T., Rodríquez-Tapia, G., Carmona, A., Arana, M., and Mercado-Gómez, J. D. (2022). Biogeographic regionalization of the Neotropical region: new map and shapefile. An. Acad. Bras. Cienc. 94:e20211167. doi: 10.1590/0001-3765202220211167
Murillo-Ramos, L. C., Ayazo, B. R., and Linares, A. J. C. (2016). Notes on nesting tunnels of Diabroctis cadmus Harold, 1868 (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 70, 198–199. doi: 10.1649/072.070.0134
Nichols, E., Spector, S., Louzada, J., Larsen, T., Amezquita, S., and Favila, M. E. (2008). Ecological funtions and ecosytem services provided by Scarabaeinae dung beetles. Biol. Conserv. 141, 1461–1474. doi: 10.1016/j.biocon.2008.04.011
Noriega, J. A. (2002). Aportes a la biología del escarabajo sudamericano Sulcophanaeus leander (Waterhouse, 1891) (Coleoptera: Scarabaeidae). Acta Zool. Mex. 87, 67–82. doi: 10.21829/azm.2002.87871801
Ocampo, F. C. (2004). Food relocation behavior and synopsis of the southern south American genus Glyphoderus Westwood (Scarabeaidae: Scarabaeinae: Eucraniini). Coleopt. Bull. 58, 295–305. doi: 10.1649/685
Ocampo, F. C. (2005). Revision of the south American endemic genus Anomiopsoides Blackwelder, 1944 (Coleoptera: Scarabaeidae: Scarabaeinae: Eucraniini) and description of its food relocation behavior. J. Nat. Hist. 39, 2537–2557. doi: 10.1080/00222930502005928
Ocampo, F. C. (2010). The south American dung beetle genus Ennearabdus Lansberge (Coleoptera: Scarabaeidae: Scarabaeinae, Eucraniini). J. Insect Sci. 10, 1–12. doi: 10.1673/031.010.9301
Ocampo, F. C., and Hawks, D. C. (2006). Molecular phylogenetics and evolution of the food relocation behaviour of the dung beetle tribe Eucraniini (Coleoptera: Scarabaeidae: Scarabaeinae). Invertebr. Syst. 20, 557–570. doi: 10.1071/IS05031
Ocampo, F. C., and Philips, T. K. (2005). Comportamiento de reubicación del alimento de Eucranium, escarabajo estercolero endémico de la Argentina y comparación con Scarabaeus (Pachysoma) del sudoeste de África (Coleoptera: Scarabaeidae: Scarabaeinae). Rev. Soc. Entomol. Argent. 64, 53–59.
Ortiz-Domínguez, M., Favila, M. E., Mendoza-López, M. R., García-Barradas, O., and Cruz-Sánchez, J. S. (2006). Epicuticular compounds and sexual recognition in the ball-roller scarab, Canthon cyanellus cyanellus. Ent. Exp. Appl. 119, 23–27. doi: 10.1111/j.1570-7458.2006.00388.x
Palestrini, C., Barbero, E., Zunino, M., and Luzzatto, M. (1994). Reproductive biology in Malagoniella puncticollis (Blanchard) (Coleoptera Scarabaeidae Canthonina): first evidence. Ethol. Ecol. Evol. 6, 437–438.
Pérez-Cogollo, L. C., Rodríguez-Vivas, R. I., Delfin-González, H., Reyes-Novelo, E., and Morón, M. A. (2015). Life history of Onthophagus landolti Harold, 1880 (Coleoptera: Scarabaidae), with descriptions of the preimaginal stages. Coleopt. Bull. 69, 255–263. doi: 10.1649/0010-065X-69.2.255
Pluot-Sigwalt, D. (1988a). Le système des glandes tégumentaires des Scarabaeidae rouleurs, particulièrement chez deux espèces de Canthon (Coleoptera). Folia Entomol. Mex. 74, 79–108.
Pluot-Sigwalt, D. (1988b). Données sur l’activité et le role de quelques glandes tégumentaires, sternales, pygidiales et autres, chez deux espèces de Canthon (Coleoptera, Scarabaeidae). Bull. Soc. Ent. Fr. 93, 89–98. doi: 10.3406/bsef.1988.17532
Pluot-Sigwalt, D. (1991). Le système glandulaire abdominal des coléoptères coprophages Scarabaeidae: ses tendances évolutives et ses relations avec la nidification. Ann. Soc. Entomol. Fr. 27, 205–229.
Price, D. L., and May, M. L. (2009). Behavioral ecology of Phanaeus dung beetles (Coleoptera: Scarabaeidae): review and new observations. Acta Zool. Mex. 25, 211–238. doi: 10.21829/azm.2009.251621
Ramírez-Restrepo, L., and Halffter, G. (2016). A dung beetle in Mexico City: the case of Ceratotrupes fronticornis (Erichson) (Coleoptera: Geotrupidae). Acta Zool. Mex. 32, 213–214. doi: 10.21829/azm.2016.322954
Rivera-Cervantes, L. E., and Halffter, G. (1999). Monografía de las especies mexicanas de Canthon del subgénero glaphyrocanthon (Coleoptera: Scarabaeidae: Scarabaeinae). Acta. Zool. Mex. 77, 1–12. doi: 10.21829/azm.1997.70701749
Rodrigues, S. R., and Flechtmann, C. A. H. (1997). Aspectos biológicos de Canthon lituratus (Germar, 1813) e Canthidium (Canthidium) megathopoides Boucomont, 1928 (Coleoptera, Scarabaeidae). Acta Zool. Mex. 70, 1–12. doi: 10.21829/azm.1997.70701749
Salomão, R. P., Favila, M. E., González-Tokman, D., and Chamorro-Florescano, I. A. (2019). Contest dynamics for food and reproductive resources are defined by health condition in a dung beetle. Ethology 125, 343–350. doi: 10.1111/eth.12858
Sarges, R., Halffter, G., and Díaz, R. A. (2012). The importance of frugivory to the survival of the dung beetle Onthophagus rhinolophus Harold (Coleoptera: Scarabaeidae: Scarabaeinae) under changing ecological conditions. Coleopt. Bull. 66, 166–168. doi: 10.1649/072.066.0216
Scholtz, C. H. (2009). “Evolution of feeding, competition, and life history strategies” in Evolutionary biology and conservation of dung beetles. eds. H. Clarke, C. H. Scholtz, A. L. V. Davis, and U. Kryger (Bulgaria: Pensoft Publishers), 40–61.
Scholtz, C. H., Davis, A. L. V., and Kryger, U. (2009). Evolutionary biology and conservation of dung beetles. Bulgaria: Pensoft Publishes.
Silveira, F. A. O., Santos, J. C., Viana, L. R., Falqueto, S. A., Vaz-De-Mello, F. Z., and Fernandes, G. W. (2006). Predation on Atta laevigata (Smith, 1858) (Formicidae Attini) by Canthon virens (Mannerheim, 1829) (Coleoptera, Scarabaeidae). Trop. Zool. 19, 1–7.
Simmons, L. W., and Ridsdill-Smith, T. J. (2011). Ecology and evolution of dung beetles. Oxford: Wiley-Blackwell Publishing Ltd. doi: 10.1002/9781444342000
Tonelli, M. (2021). Some considerations on the terminology applied to dung beetle functional groups. Ecol. Entomol. 46, 772–776. doi: 10.1111/een.13017
Tribe, G. D. (1975). Pheromone release by dung beetles (Coleoptera: Scarabaeidae). South Afr. J. Sci. 71, 277–278.
Trotta-Moreu, N., Montes de Oca, E., and Martínez, M. I. (2007). Ecological and reproductive characteristics of Geotrupes (Halffterius) rufoclavatus Jekel 1865 (Coleoptera: Geotrupidae: Geotrupinae) on the Cofre de Perote volcano (Veracruz, Mexico). Coleopt. Bull. 61, 435–446. doi: 10.1649/0010-065X(2007)61[435:EARCOG]2.0.CO;2
Vaz-de-Mello, F. Z., and Génier, F. (2009). Notes on the behavior of “Dendropaemon” Perty and “Tetramereia” Klages (Scarabaeidae: Scarabaeinae: Phanaeini). Coleopt. Bull. 63, 364–366. doi: 10.1649/1191.1
Vaz-De-Mello, F. Z., Louzada, J. N. C., and Schoereder, J. H. (1998). New data and comments on Scarabaeidae (Coleoptera, Scarabaeoidea) associated with Attini (Hymenoptera, Formicidae). Coleopt. Bull. 52, 209–216.
Verdú, J. R., and Galante, E. (1997). Aphodius brasiliensis Castelnau (Coleoptera: Aphodiidae): larval morphology and notes on biology. Coleopt. Bull. 51, 378–383.
Verdú, J. R., and Galante, E. (2001). Larval morphology and breeding behavior of the genus Pedaridium Harold (Coleoptera: Scarabaeidae). Ann. Ent. Soc. Amer. 94, 596–604. doi: 10.1603/0013-8746(2001)094[0513296:LMABBO]2133.0.CO;2
Villalobos, F. J., Díaz, A., and Favila, M. E. (1998). Two species of Canthon Hoffmannsegg (Coleoptera: Scarabaeidae) feed on dead and live invertebrates. Coleopt. Bull. 52, 101–104.
Villalva, S., Lobo, J. M., Martín-Piera, F., and Zardoya, R. (2002). Phylogenetic relationships of Iberian dung beetles (Coleoptera. Scarabaeinae): insights on the evolution of nesting behavior. J. Mol. Evol. 55, 116–126. doi: 10.1007/s00239-002-2314-4
Wille, A. (1973). Observations on the behavior of a tropical rain forest dung beetle, Megathoposoma candezei (Harold) (Coleoptera: Scarabaeidae). Rev. Biol. Trop. 21, 45–57.
Zunino, M., Barbero, E., and Luzzatto, M. (1989). Food relocation behaviour in Eucraniina beetles (Scarabaeidae) and the constraints of xeric environment. Trop. Zool. 2, 235–240. doi: 10.1080/03946975.1989.10539440
Keywords: sexual behavior, parental care, Neotropical, foraging competition, Scarabaeinae
Citation: Huerta C, Cruz-Rosales M, González-Vainer P, Chamorro-Florescano I, Rivera JD and Favila ME (2023) The reproductive behavior of Neotropical dung beetles. Front. Ecol. Evol. 11:1102477. doi: 10.3389/fevo.2023.1102477
Edited by:
José F. Fontanari, University of São Paulo, BrazilReviewed by:
Malva Isabel Medina Hernández, Federal University of Santa Catarina, BrazilEduardo Galante, University of Alicante, Spain
Copyright © 2023 Huerta, Cruz-Rosales, González-Vainer, Chamorro-Florescano, Rivera and Favila. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carmen Huerta, ✉ Y2FybWVuLmh1ZXJ0YUBpbmVjb2wubXg=; José D. Rivera, ✉ amRyNDk1QGhvdG1haWwuY29t
†These authors have contributed equally to this work