
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 02 March 2023
Sec. Phylogenetics, Phylogenomics, and Systematics
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1096208
This article is part of the Research TopicNeotropical Dung Beetle Diversity: Ecological, Historical, and Anthropogenic PerspectivesView all 11 articles
 Eder F. Mora-Aguilar1‡
Eder F. Mora-Aguilar1‡ Alfonsina Arriaga-Jiménez2‡
Alfonsina Arriaga-Jiménez2‡ César M. A. Correa3‡
César M. A. Correa3‡ Pedro Giovâni da Silva4‡
Pedro Giovâni da Silva4‡ Vanesca Korasaki5‡
Vanesca Korasaki5‡ Pablo A. López-Bedoya6,7‡
Pablo A. López-Bedoya6,7‡ Malva Isabel Medina Hernández8‡
Malva Isabel Medina Hernández8‡ José D. Pablo-Cea9,10‡
José D. Pablo-Cea9,10‡ Renato Portela Salomão11,12‡Gorky Valencia13‡Kevina Vulinec14‡Felicity A. Edwards15,16‡David P. Edwards16‡Gonzalo Halffter17†‡
Renato Portela Salomão11,12‡Gorky Valencia13‡Kevina Vulinec14‡Felicity A. Edwards15,16‡David P. Edwards16‡Gonzalo Halffter17†‡ Jorge Ari Noriega18*‡
Jorge Ari Noriega18*‡Introduction: The standardization of sampling protocols is imperative for robustly studying any taxonomic group. Replicable methods allow the comparison of data between different spatial and temporal studies. In the case of dung beetles, one of the best-studied indicator groups in analyses of environmental disturbance, a wide range of collection methodologies are used, from basic pitfall traps to more complex or complementary methods such as mini-Winkler extractor. Also, different types of attractive baits, sampling effort, durations, and designs are used in dung beetle studies. Variations in methodological approaches are particularly noted in the Neotropics, which may be related to the vast number of biological strategies and behavior of dung beetles that inhabit this region. A lack of methodological unification for the Neotropical region makes a cross-sectional analysis of the information impossible.
Methods: We performed a compilation and analytical review of the existing literature for dung beetle sampling in the Neotropics, discussing the most used methodologies, their advantages and disadvantages, and specific cases in which particular models are more efficient.
Results: Pitfall traps baited with human excrement are the most common sampling method, but there is a wide range of models and variations in the structure of this trap. The complementary effect generated by flight interception traps, light traps, and direct collections, particularly within microhabitats, is exciting for the potential of finding new species. Some methodologies, such as mini-Winkler extractor, fogging, or very specific baits, are infrequently used.
Discussion: There was a lack of inclusion of spatial and temporal variation among studies. Therefore, it is necessary to consider broader sampling windows, which include different spatial scales, seasons, and years. Finally, we propose a standard protocol for sampling dung beetles in the Neotropics, depending on each objective, and including a basic methodology for obtaining complete local inventories.
Biodiversity, a somewhat loosely delineated concept in the ecological literature, needs defining for every case where it is used. In many situations, it is meant to describe all the species in an area; in others, it is restricted to one group of organisms. The latter is probably a more precise measure if one is trying to compare different habitats or locations. Defining biodiversity as a metric for a group of similar and related species allows a more accurate picture of the effect of geography, habitat, altitude, or other comparisons among species comprising these locations. It has been well-documented that there is declining species abundance and richness across the globe, which is why these studies are so critical. The diversity of birds (Fujisaki et al., 2008), amphibians (Becker et al., 2007), and mammals (Spooner et al., 2018), among many other groups, were documented as declining, with causes ranging from climate change, overhunting, and the introduction of exotic species. Nevertheless, fewer investigations have examined insect declines in the Neotropics (Scheffers et al., 2012; Cardoso and Leather, 2019; Cardoso et al., 2020). In order to clearly assess biodiversity within a biological group, robust and standardized surveying methodologies are required. However, many different methodologies have been employed to study each biological group, and this issue may pose challenges regarding the comparisons among studies.
The Neotropics comprise multiple ecosystems that are biodiversity hotspots for many taxa (Myers et al., 2000; Durães et al., 2013; Ríos-Touma and Ramírez, 2019). While controversy remains about the species extinction risk of tropical deforestation (Laurance, 2007), the current increase in habitat loss has become a critical concern for many vulnerable species and the environment as a whole (Barbosa et al., 2021). The Neotropics are particularly vulnerable because of uncontrolled logging, cattle ranching, mining, and farming (Gibbs et al., 2010; Ferreira et al., 2014). Numerous studies have documented the effect of anthropic activity, with declines in many species and extirpation of others (e.g., Noriega et al., 2021a). These works often examine species that are especially vulnerable to habitat change and are considered indicators. Such bioindicators are plants or animals that are thought of as “canaries in the coal mine,” meaning they may be the initial species to show declines or actual physiological changes with disturbance (Salomão et al., 2019a). These species are often considered bioindicators because they also have important roles in the local ecology (Vulinec, 2002; Valente-Neto et al., 2021). Different insect groups are often referred as bioindicators because of their relative ease of capture and standardized methodology, rapid response to environmental degradation, and cost efficiency ratio (Gardner T. et al., 2008). Recent studies have included dragonflies (Silva et al., 2021), ants (Pérez-Espona, 2021; da Silva W. B. et al., 2022), and butterflies (Doré et al., 2021) as bioindicators, among others.
Dung beetles (Coleoptera: Scarabaeinae) feature as one of the best-studied indicator groups in the analysis of environmental disturbance (e.g., Nichols et al., 2007; Gardner T. et al., 2008). Most of the studies have shown significant changes in dung beetle richness and abundance with even small changes in intact forests (Nichols et al., 2007). The presence of dung beetles also indicates that large mammals are present, providing the food source for dung beetles (Vulinec et al., 2006; Raine and Slade, 2019; Correa-Cuadros et al., 2022). There are, however, a wide range of sampling methodologies for dung beetles, from basic pitfall traps to more complex or complementary methods such as mini-Winkler extractor (Iannuzzi et al., 2021). These varied methodologies and sampling efforts make comparisons difficult among the many studies. In this scenario, the standardization of sampling protocols is an imperative need for studying any taxonomic group. Replicable methods allow for comparing data between different spatial and temporal studies. Nevertheless, no consensus allows a methodological unification, presenting works with different sampling designs that make a cross-sectional analysis of the information impossible. Among the biogeographic domains, Neotropical regions comprise one of the best-studied areas regarding dung beetle ecology, thus serving as a model region to depict the dynamics of ecological studies in dung beetles. In this paper, we thus focus on the Neotropical region to make a cross-sectional analysis of the importance of different types of traps, baits, sampling efforts, durations, and designs in sampling dung beetles. We also reviewed several studies to determine the most employed methodologies for the analysis of the effect of habitat alteration on dung beetle richness and abundance. Finally, after reviewing the existing literature, we propose a protocol for the group aiming for standardizations in dung beetle sampling in the Neotropics.
We performed a literature search following the PRISMA methodology (Page et al., 2021) to identify articles dealing with dung beetle trap sampling published from 1968 to 2021 (maximum time search window). Firstly, we conducted bibliographic queries in Web of Science (WoS) and Scientific Electronic Library Online (SciELO) databases using the keyword string: (“scarab*” OR “escarab*” OR “dung beetle*”) AND (“neotropic*” OR “tropic*”) AND (“trap*” OR “tramp*”), looking for matches in the title, abstract, and/or keywords. Therefore, from the initial search (updated on February 2022), we retrieved 4,799 records (WoS = 4,632 and SciELO = 167). We then eliminated duplicate records, studies out of the boundaries of the Neotropics (see Morrone et al., 2022), experimental, meta-analysis, revision, taxonomical or without richness data associated with trapping (i.e., articles that appeared more than once in the different search engines or the same platform due to typographical errors). All references not related to any dung beetle species of the subfamily Scarabaeinae were also excluded.
The following information was collected from each selected publication: year of publication, author(s), title, journal, language, country, biomes, ecosystems, if the study is about disturbances and what type of disturbance it is, geographic coordinates, elevation, trap type, the number of traps used, time active of each trap, bait, study approach (i.e., taxonomic or ecological), number of samples in space (spatial replicates), number of samplings in time (temporal replicates), seasonality, and any relevant additional observation. It is important to clarify that an article can represent more than one item for the analysis. For more detailed information from each of the analyzed studies, please see Supplementary Annex 1. To keep consistency with the literature, we used the biogeographical proposal by Morrone et al. (2022) to standardize and unify the biomes.
This literature search type has several limitations, which were carefully considered when analyzing the data and interpreting the results. First, the search may miss some relevant papers simply because either the title, abstract, or keywords did not contain the focal keywords. Other authors have previously identified these limitations using similar search approaches (see Prather et al., 2013). It is evident that the language, especially in the Neotropical region, is a limiting factor in the search and that articles in Spanish and Portuguese could have been left out. Finally, we may have failed to include some works that were not indexed by the platforms used here. Despite these limitations, we believe that the data retrieved gives us enough relevant information to examine general trends in dung beetle trapping research in the Neotropics. With our current literature revision, we may identify knowledge gaps that could help us to develop future research strategies to build more precise methodological approaches.
As a result of our bibliographic search, we found 241 articles studying dung beetles in the Neotropical region (Figure 1; Supplementary Annex 1). We recorded evidence of a marked trend toward an increase in publications on dung beetles in the last three decades (Figure 2A). We registered very few works in the 90s and none before 1990, which is related to the restrictions of the search method and the words used. Most of the registered articles are in English (n = 194, 80.5%), followed by Spanish and Portuguese (Figure 2B). We find a wide variety of registered journals, with more than 74 different journals. The journals with the highest number of articles included are Revista Brasileira de Entomologia, Journal of Insect Conservation, Revista Mexicana de Biodiversidad, Neotropical Entomology, and PLoS ONE (Figure 2C).
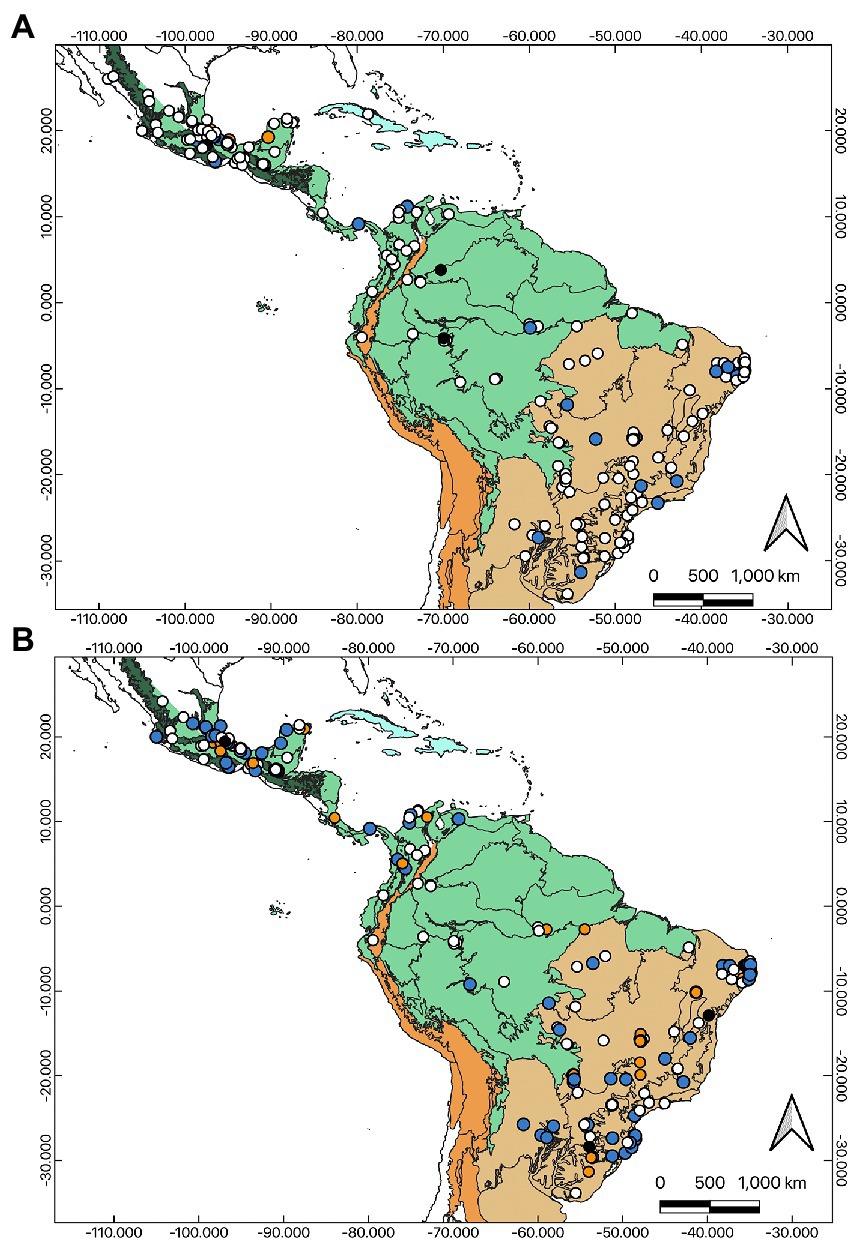
Figure 1. Distribution of published studies of dung beetles sampling compiled in our dataset in the biogeographical regions in the Neotropics. Each dot corresponds to one paper. (A) Studies using only pitfall trap (open diamonds), and other complementary traps. One trap (open dots), two traps (light gray dots), three traps (dark gray dots), and four traps (black dots). (B) Studies using different types of baits. One bait (open dots), two baits (light gray dots), three baits (dark gray dots), and four baits (black dots).
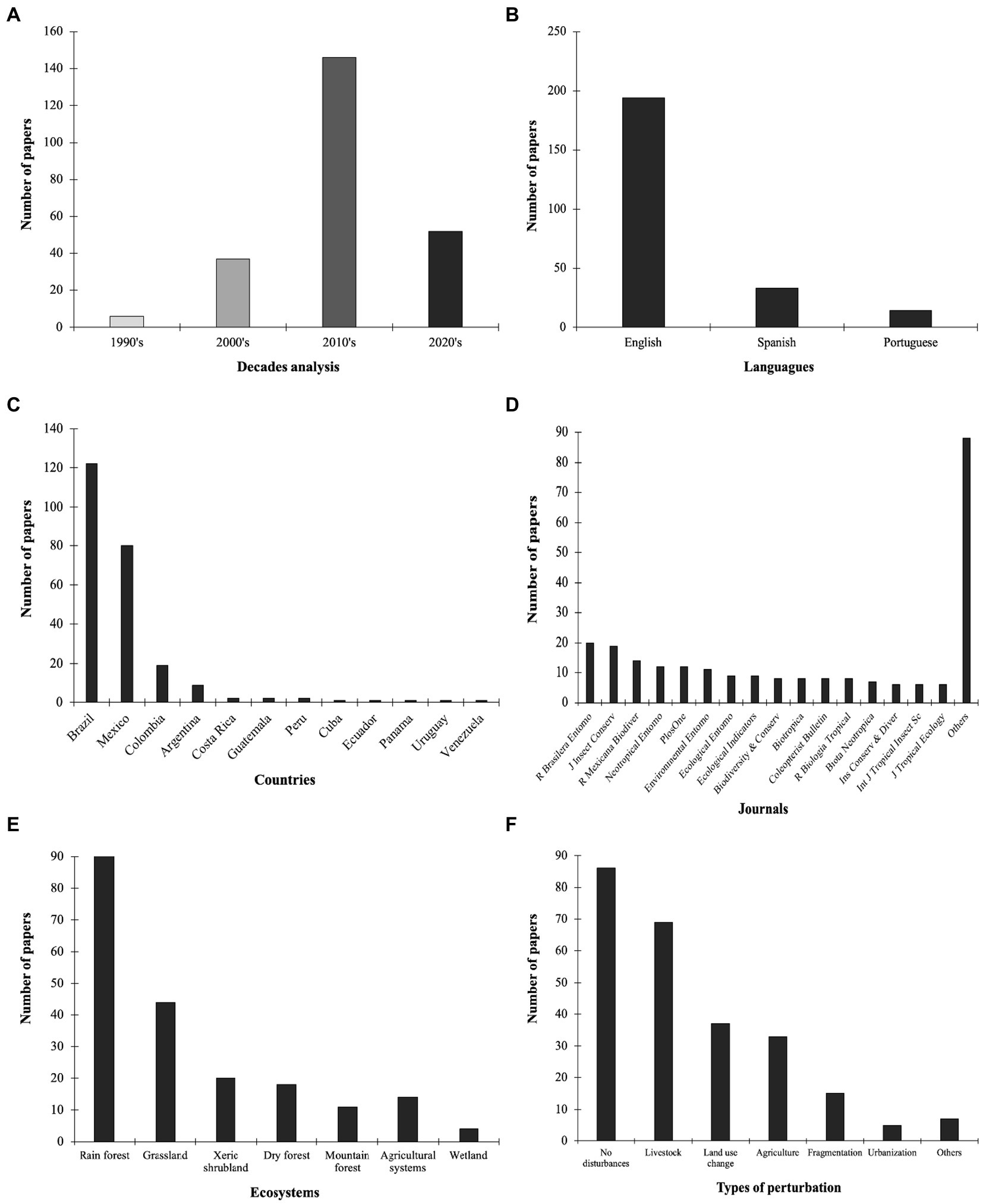
Figure 2. Number of published papers on dung beetle sampling compiled in the Neotropics according to different variables: (A) Decades analysis; (B) Languages; (C) Journals; (D) Countries; (E) Ecosystems; and (F) Types of perturbation.
Twelve countries are represented in our dataset, comprising almost the full range of the Neotropical region, with the southernmost study being carried out in Uruguay and the northernmost in Mexico (Figures 1, 2D). The studies included South, Central, North America, and the Caribbean. Brazil had the largest number of studies (n = 122, 50.6%), followed by Mexico (n = 80; 33.2%), and Colombia (n = 19, 7.9%; Figure 2D). Data from these three countries represented 91.7% of our dataset. Argentina was represented by nine papers (3.7%). Among the countries with the lowest number of publications in this review, Costa Rica, Guatemala, and Peru were represented by two papers each (0.8%); Cuba, Ecuador, Panama, Uruguay, and Venezuela were represented by only one study each (0.4%). Nine countries, within the Neotropical region had no studies recorded in this review. In Brazil, most studies took place in the Paraná dominion (Atlantic province, Araucaria Forest province, and Paraná Forest province) and Chacoan dominion (Caatinga province and Cerrado province). In Mexico, most studies were conducted in the Mexican Transition Zone (Trans-Mexican Volcanic Belt province) and the Mesoamerican dominion (Veracruzan province and Yucatán Peninsula province). In Colombia, most studies belonged to the Pacific dominion (Guajira province, Magdalena province). To cite some prolific researchers in Mexico: G. Halffter, M.E. Favila, A. Estrada, L. Arellano, and R.P. Salomão; in Brazil: J.N.C. Louzada, F.Z. Vaz-de-Mello, M.I.M. Hernández, P.G. da Silva, and C.M.A. Correa; and in Colombia: L.C. Pardo-Locarno, F. Escobar, and J.A. Noriega.
Natural forests were the most studied ecosystem, with 62.6% of the reviewed studies (n = 151; Figure 2E). Among them, tropical rainforests showed the highest number of papers (n = 122, 50.6%), followed by dry forests (n = 18, 7.5%), and mountain forest (n = 11, 4.5%). Other natural ecosystems, such as xeric shrubland and wetlands, represented together 9.9% of the total number of studies. Research on Neotropical dung beetles also showed a high number of papers in other non-forested ecosystems. Among these papers, some of the most studied include grassland (including both natural and anthropic, with 18.2%) and agricultural systems (5.8%; Figure 2E) which encompassed coffee, corn, or oil palm plantations. In addition, around two-thirds of the studies evaluated dung beetles under the effects of anthropic disturbance (Figure 2F). Among them, livestock was the most studied anthropic impact (n = 69, 28.6%), but other agriculture or fragmentation were also common.
Pitfall traps were used in the majority of studies (n = 206, 85.48%), while manual capture (n = 21, 8.71%), light traps (n = 13, 5.39%), flight interception traps (n = 11, 4.56%), and NTP-80 (permanent necro trap model 80, n = 7, 2.90%) were less frequently used. Carp traps, dung pats, Malaise traps, aerial traps, Shannon traps, and platform traps were used in very few studies (totaling n = 17, 7.05%; Figure 3A). None of the typical techniques were used in a small number of studies (n = 5; 2.07%); instead, other observations or experimental methodologies were used. Most studies (n = 208, 86.31%) surveyed dung beetles using only one trap type, two trap types were simultaneously used in 21 studies (8.71%), while three trap types were used in one study (0.41%) and four different types were used in four studies (1.66%; Figure 3B). Pitfall traps were used as the only surveying method in 182 (75.52%) studies; samplings using exclusively manual capture was performed in eight studies (3.32%), and light traps were only used in seven studies (2.90%). Studies using only flight interception traps to sample dung beetles comprised four studies (1.66%).
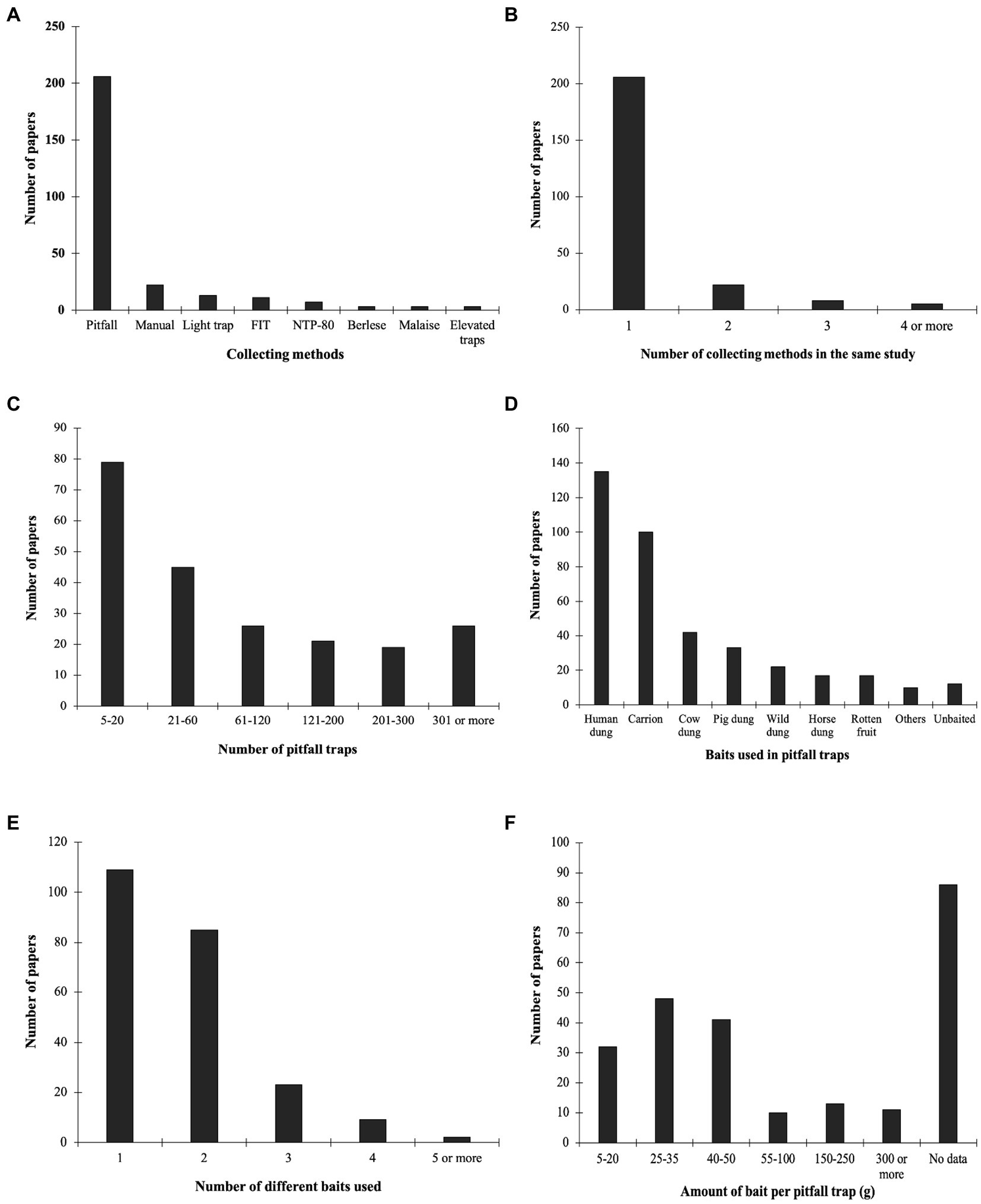
Figure 3. Number of published papers on dung beetle sampling compiled in the Neotropics according to different variables: (A) Collecting methods; (B) Number of collecting methods in the same study; (C) Number of pitfall traps; (D) Baits used in pitfall traps; (E) Number of different baits used; and (F) Amount of bait per pitfall trap.
Overall, studies that used more than one sampling method always used pitfall traps (e.g., pitfall and light traps, pitfall and NTP-80, pitfall and platform traps, pitfall and Shannon traps, pitfall and carp traps). In very few studies, pitfall traps were combined with flight interception traps (n = 6, 2.49%) and direct collection (n = 5, 2.07%). Among the studies that used pitfall traps, the number of traps ranged from four to more than 300 traps. Almost one-third of the studies (n = 62, 30.10%) had a sampling effort ranging from four to 20 traps. In 42 of the analyzed studies (20.39%), there was a sampling effort ranging from 21 to 60 traps; in 73 of the studies (35.43%) from 61 to 300 traps were used; 300 or more traps in 26 studies (12.62%; Figure 3C). In three studies (1.46%), the number of traps used was not reported.
Baited techniques to sample dung beetles were used in most of the studies analyzed, and only 2.9% used non-baited collecting methods. Human dung was the most used bait, corresponding to 56% of the papers evaluated (n = 135). Secondly, carrion was used in 41.5% of the studies (n = 100), followed by cattle dung (n = 42, 17.4%) and pig dung (n = 34, 14.1%; Figure 3D). In a smaller number of studies, other dung types were used as baits, including horse, wild vertebrate [native and exotic species, e.g., waterbuck – Kobus ellipsiprymnus (Ogilby, 1833) and jaguar – Panthera onca (Linnaeus, 1758)], and combinations of different types of dung, such as human and pig dung. There was no consensus concerning the number of baits used in the sampling protocols. Almost half of the papers (n = 110, 45.6%) used only one bait type, whereas 35.2% (n = 85) of them used two types of bait and 9.1% used three types of bait (n = 22; Figure 3E). The amount of bait used varied and ranged from 25 g to 35 g (n = 48, 19.9%), followed by 40–50 g (n = 41, 17.0%), and 5–25 g (n = 31, 12.8%; Figure 3F). Nonetheless, a significant number of analyzed papers (n = 86, 35.6%) did not include this information.
A considerable number of studies did not present a clear temporal (n = 48, 19%) and spatial (n = 52, 21.6%) distribution of traps. This included studies with unclear sampling techniques or studies with sampling techniques that did not comprise the use of traps per se (e.g., direct collection in dung pats). The time length during which traps were kept active in the experiments varied greatly, ranging from 24 h (1 day) to more than 480 h (20 days; Figure 4A). Among the studies that reported the time in which traps were kept active in the field, most of them had traps installed for 48 h (n = 114, 47.3%), followed by 24 h (n = 40, 16.6%), with fewer choosing 480 h or more (n = 28, 11.6%). Three studies (1.2%) let traps remain active for <24 h. Distances between traps varied widely, from 2 m to 1,000 m (Figure 4B). From the studies in which trap spacing was reported, most of them had traps spaced 50 m apart (n = 76, 40.2%). A considerable number of studies used the spacing intervals 2–20 m (n = 52, 27.5%) and 25–40 m (n = 25, 13.2%), while a few studies spaced traps more than 50 m (from 60 m to 100 m, n = 13, 6.8%; 150 m or more, n = 11, 5.8%). In terms of spatial sampling, of the 241 articles reviewed, the majority (>55%) used between 1 and 3 replicates per study, however a single replica was used by most of the authors (Figure 4C). Forty percent used more than four replicates, of these less than half (45%) used more than 10 replicates. A minority of studies (4%) did not give sufficient information on the sampling.
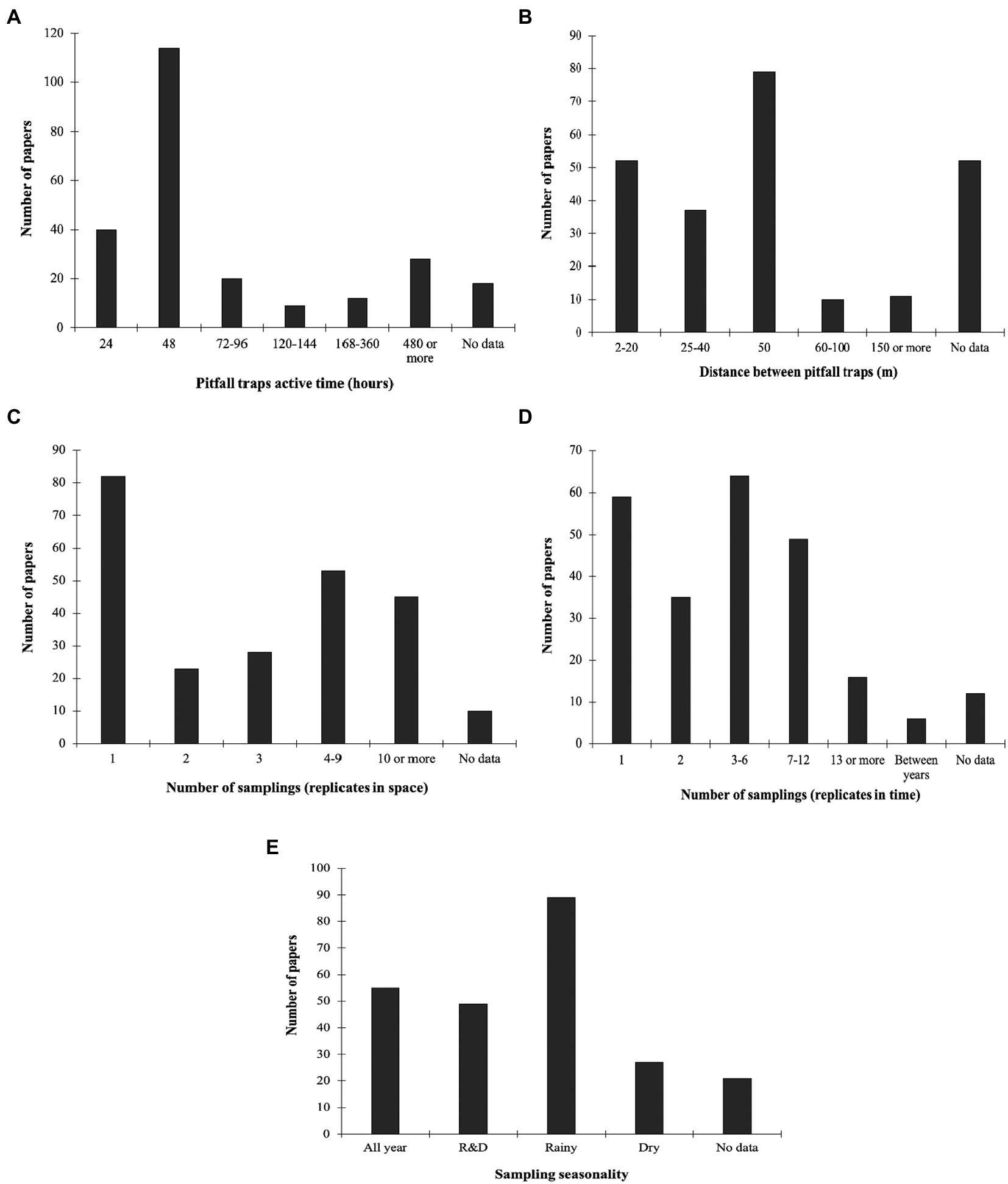
Figure 4. Number of published papers on dung beetle sampling compiled in the Neotropics according to different variables: (A) Pitfall traps active time; (B) Distance between pitfall traps; (C) Number of samplings in space; (D) Number of samplings in time; and (E) Sampling seasonality.
Concerning temporal variation, research conducted throughout the year corresponded to only 22.8% (n = 55) of the papers reviewed (Figure 4D). In general, research was carried out only in the rainy season (n = 89, 36.9%), while other works also include the dry season with 20.3% of the total (Figure 4E). There are very few studies carried out only in the dry season, as well as papers that do not present explicit information about the time of year in which the study took place.
The large number of studies found in Brazil, Mexico, and Colombia reflects the pioneering aspect and growth of research centers and researchers aimed at studying the biology, ecology, and taxonomy of Scarabaeinae in these countries. Most of the work carried out with dung beetles in the Neotropics is carried out in natural forest environments, likely due to the large coverage of Neotropical forest biomes, but also strongly influenced by the geographical location of researchers throughout history, initially focused on Mexico with the pioneering work carried out by Halffter and collaborators. After the proposal on using dung beetles as ecological indicators (Halffter and Favila, 1993), many works emerged comparing communities in pristine and anthropized environments (e.g., Gardner T. A. et al., 2008; López-Bedoya et al., 2021, 2022). When comparing multiple environments (i.e., natural vs. anthropic), it is necessary to carefully standardize the sampling methodology, to avoid collecting bias. For example, baits placed in environments with high direct solar incidence can quickly lose efficiency due to water loss (Lobo et al., 1998) representing a potential bias on capture rates between environments. Several classical studies of the 90’s or previous were not included; these papers are found in local or not indexed journals. These are located in Mexico and Brazil, but this does not alter the general pattern. Almost 14 provinces do not show any studies in this work, which suggest low or null effort, particularly in the Subregion called the South American Transition zone. In the following paragraphs, we will discuss the findings of this revision and suggest trends encompassing sampling protocols, aiming to improve and guide future ecological studies with dung beetles.
Pitfall traps consist of a container buried at ground-surface level filled with liquid (soapy water or ethanol), allowing crawling animals to fall in but preventing them from leaving (Southwood, 1978; Brown and Matthews, 2016). Our results show that pitfall traps are the dominant method for capturing dung beetles. However, there is a great diversity of models of this trap (Lobo et al., 1988; Veiga et al., 1989; Halffter and Favila, 1993) that have been implemented throughout history to capture a great diversity of taxa (e.g., Newton and Peck, 1975; Spence and Niemelä, 1994; Buchholz and Möller, 2018). The use, adaptation, and importance of pitfall traps for dung beetle capture were described by Lobo et al. (1988), Veiga et al. (1989), and Halffter and Favila (1993). Pitfall traps are popular because they are inexpensive and relatively simple to construct, install, collect, and are efficient in capturing beetles, especially when combined with bait suspended above the trap (Lobo et al., 1988; Halffter and Favila, 1993; Kočárek, 2000; Hohbein and Conway, 2018). The design of pitfall traps is not universal (Lobo et al., 1988), being contingent on the creativity of researchers (e.g., Porter, 2005; McKnight et al., 2013; Buchholz and Möller, 2018), availability of resources, and characteristics of the ecosystem where they are deployed (e.g., Spence and Niemelä, 1994; Porter, 2005; Noriega and Fagua, 2009). This is the main reason why a clear description of the trap adaptations used is essential, so methodologies are replicable, and results are comparable (Brown and Matthews, 2016; Hohbein and Conway, 2018); rather than simply stating that “pitfall traps were used to capture beetles” (e.g., Sarges et al., 2012; Trujillo-Miranda et al., 2016; Salomão et al., 2019a). On the other hand, we must also consider that the current trends that focus on publishing shorter and more precise publications often results in articles with limited methodological descriptions, removing details of trapping methods.
The number of traps set has not been contemplated in most studies evaluating different aspects of the methodological design of studies using pitfall traps (Boetzl et al., 2018). In the few studies that have considered it, the number of traps was identified as one of the most critical factors in the sampling design (Engel et al., 2017). However, there has been and continues to be a considerable variation in the number of traps used in studies (Brown and Matthews, 2016), and dung beetles are no exception (see Supplementary Annex 1). In most studies, 10–40 pitfall traps were placed per sampling event. However, there are studies in which the number of traps implemented exceeded one thousand (e.g., Estrada et al., 1999; Sarges et al., 2012; Bourg et al., 2016), and the most extreme case evaluated is 2,400 active traps per sampling event (Estrada and Coates-Estrada, 2002). However, some studies do not include the number of pitfall traps used (Martínez and Suárez, 2006; Morón-Ríos and Morón, 2016; Salomão et al., 2019a); providing this information is critical to calculate capture rate and ensure replicability. The spatial distribution and number of traps across habitat gradients should also aim to be standardized to ensure unbiased evaluation of the impact of anthropogenic activities.
In a recent study, Rivera and Favila (2022) demonstrated that ecological studies in the Neotropics often collect more dung beetle individuals than necessary to obtain a representative diversity sample. They suggest that we are currently oversampling the dung beetle community. In future studies, it is crucial to assess the optimal number of traps between effort and efficiency (richness, abundance, and diversity of captured dung beetles), which is the most widely used criterion for selecting the sampling methodology (Noriega and Fagua, 2009). Finding this optimal number is important since the potential impact on dung beetle populations has not been quantified, and it is possible that with fewer traps, the species asymptote will be reached, avoiding over-capture. Also, the optimal number of traps is important, especially considering that dung beetle sorting and identification can be time-demanding activities that limit the development and conclusion of ecological studies.
The use of direct collection and active searching for beetles in dung pats is based on how easy it is to find fresh excrement and insects (e.g., Morelli and Gonzalez-Vainer, 1997; Mendes and Linhares, 2006; Lopes et al., 2020). In our work, manual capture was the most used method for collecting dung beetles after pitfall traps. Its main limitation is that it is especially useful for capturing endocoprids (i.e., Eurysternus spp., beetles that nest inside the excrement) and not for collecting paracoprids or telecoprids. Light traps have been primarily used to capture phytophagous and saprophytophagous beetles that are photophilic (Ratcliffe and Cave, 2009). Nonetheless, their use to capture Scarabaeinae is based on the fact that light trap can catch species that do not fall into other types of traps (Hill, 1996; Abot et al., 2012), such as some species of Dichotomius Hope or Digitonthophagus Balthasar. Flight interception and Malaise traps are also used, intended to intercept insects randomly as they move through the air without avoiding or attracting into the trap (Southwood, 1978; Boiteau, 2000). The effectiveness of flight interception traps is limited because flying adults avoid them and may bounce off the trap without being picked up (Boiteau, 2000). We noted that these traps were among the most used after manual capture and light traps (e.g., da Costa et al., 2009; Rodrigues et al., 2010; Otavo et al., 2013; Puker et al., 2020; de Moura et al., 2021). The popularity of their use is based on the active flight displayed by dung beetles, which allows them to be intercepted if the traps are appropriately located (Puker et al., 2020). Flight intercept traps allow the capture of dung beetles not attracted by omnivore bait, as some species of Onthophagus Latreille, Deltochilum Eschscholtz, Phanaeus MacLeay, Canthidium Erichson, Cryptocanthon Balthasar, or Anomiopus Westwood, which have other food preferences (i.e., carrion, predatory, fugivory, mycetophagy).
The other most commonly used trap to capture dung beetles was NTP-80 (a model invented by Miguel A. Morón), a modification of the pitfall traps designed for the collection of insects with an affinity for decaying organic matter of animal origin, which can remain active for extended periods, and that has the main advantage of preventing looting by mammals attracted by the bait (Morón and Terrón, 1984). All the papers citing this trap were performed in Mexico (e.g., Trevilla-Rebollar et al., 2010; Deloya et al., 2013; González-Hernández et al., 2015), suggesting that it is a local modification that is not commonly used in other countries. Other types of complementary traps (e.g., aerial traps, light traps, flight interception traps, mini-Winkler extractor) for the capture of dung beetles are extremely limited because they usually incur extra expense and time, and generate discrete results in the effort and efficiency ratio. However, it has been mentioned by several authors that these traps can be used to capture rare species that do not usually fall into pitfall traps (e.g., Hill, 1996; Noriega, 2011; Abot et al., 2012; Touroult et al., 2017; Silva et al., 2020; Ong et al., 2022) and so helpful to taxonomical approaches to get rare and/or small species (e.g., Mora-Aguilar and Delgado, 2018, 2019). Therefore, studies with these traps are more relevant for taxonomic research and not for bioindicator studies. Thus, studies that evaluate these aspects in the future should be conducted.
It is virtually impossible to collect all species in a taxonomic group with only one sampling technique or bait type (e.g., Missa et al., 2009). However, a high sampling efficiency of the assemblage is vital to any research involving the biodiversity of dung beetles (e.g., Marsh et al., 2013; Noriega, 2015; Correa et al., 2018) since they are widely used as bioindicators of environmental changes (Halffter and Favila, 1993; Nichols et al., 2007). Human dung is the most used bait to sample dung beetles in the Neotropical region. Indeed, the feeding preference of dung beetles for omnivorous mammal dung usually attracts a more significant number of species and individuals relative to herbivore dung, carnivore dung, rotten fruits, or carrion (Filgueiras et al., 2009; Bogoni and Hernández, 2014; Correa et al., 2016, 2018; Salomão et al., 2018). Human dung is one of the most attractive baits for the dung beetle sample (Martín-Piera and Lobo, 1996) and is a resource available worldwide wherever the researcher travels (Marsh et al., 2013). For these reasons, human dung features as the bait type most used to sample a high abundance and species richness of dung beetle in ecological studies at the assemblage scale (Howden and Nealis, 1975; Gardner T. A. et al., 2008; Correa et al., 2016). Marsh et al. (2013) suggested using human-pig dung mixes in different proportions, with this mixed dung bait (human:pig) exhibiting efficiency comparable to human dung (see Marsh et al., 2013) and is used in more recent studies (e.g., Braga et al., 2013; França et al., 2020; Noriega et al., 2021a; see Supplementary Annex 1), demonstrating a possible tendency for future studies in the Neotropical region. In contrast, omnivorous dung can be an ineffective bait for species with a preference for open areas and/or herbivorous dung (Noriega personal observation).
Most of the studies used a single bait type in their sampling protocol, usually human dung which allows standardized comparisons among different habitats (see Howden and Nealis, 1975; Gardner T. A. et al., 2008; Correa et al., 2016). Using a single bait has clear logistical advantages, such as reduced time to set up traps in the field, reduced physical effort, and fewer financial resources (Gardner T. et al., 2008). Nevertheless, due to the trophic specialization of dung beetles (Halffter and Matthews, 1966), using multiple baits may attract a more diverse group of beetles and thus result in a better characterization of assemblages (e.g., Larsen et al., 2006; Noriega, 2015; Correa et al., 2016; Chamorro et al., 2019). Although, carrion was widely recorded in studies that used two or three bait types, mainly together with a dung type (e.g., human, cattle, or pig dung) but almost never used as the only bait in a study. The use of carrion is important due to the possibility of sampling generalists and necrophagous species (Halffter et al., 2007). Still, contrasting to dung there is no standardized carrion type to sample dung beetles, and studies use different carrion types, including fish, chicken, bovine, and pig (see Supplementary Table S1). Pivotally, the use of baits (individual or combined) will depend on the main objective of the research (see Correa et al., 2018).
The amount of bait used ranged from 5 g to 50 g. Indeed, there is no consensus in the literature on the amount of bait needed to sample dung beetles effectively, even though it has been reported that the amount (e.g., size and volume) of bait has a positive effect on the number of species and individuals captured (see Peck and Howden, 1984; Gill, 1991; Raine et al., 2020; Martínez-Hernández et al., 2022). The most commonly used dung type, human dung, can be in short supply, with a single person generating fresh dung for about 8–10 traps per day, based on a standard bait size of 20 g proposed by Marsh et al. (2013). This reduced the number of traps per day and severely limited sampling effort, and because of the high sampling effort employed in dung beetle ecological research (Gardner T. et al., 2008), larger amounts of human dung are required. This fact may drive the researchers to use lower amounts of bait per trap, aiming to increase the number of traps in their studies. To understand better how the collection method can affect the quantification of the community, further studies should assess the efficiency of different amounts of baits in sampling dung beetles in the Neotropical region (Martínez-Hernández et al., 2022). Furthermore, to our knowledge, it is relatively unknown how the amount of bait may affect the attractiveness of dung beetle assemblages in scenarios with distinct environmental and landscape conditions. In dry ecosystems, such as in tropical dry forests in the Neotropics, dung dries more quickly compared to wet ecosystems (e.g., tropical rainforests). Regarding ecosystem types, the amount of feces could be considered and modulated in order to maintain a similar attractiveness during the sampling period among different regions. Thus, this information may help researchers to use a standardized and/or ideal amount of bait per trap in future studies.
Most studies using pitfall traps ranged from 24 to 48 h of active trapping. Previous studies state that dung beetles have a high colonization rate on decaying material during the first 48 h of resource availability (Kessler and Balsbaugh, 1972; Sullivan et al., 2017; Wassmer, 2020). There are two important factors related to the time in which baited traps are active: (i) the decrease in the potential of the attractiveness of the resource with the advance of time (Hanski and Cambefort, 1991), and (ii) decaying organisms that fall in the pitfall produce odors that may attract or repel organisms other than those attracted to the bait used in the experiment (Schmitt et al., 2004; Fletchmann et al., 2009). In tropical rainforests, 48 h comprises the optimal time-lapse to obtain the most bait-attracted dung beetles. Nonetheless, in tropical dry forests, there is a high evapotranspiration dynamic (Sampaio, 1995; Velloso et al., 2002), which results in the rapid drying out of food resources. In dry-forest ecosystems, it is relatively common to install pitfall traps for 24 h (e.g., Barraza et al., 2010; Rangel-Acosta and Martínez-Hernández, 2010; Salomão et al., 2018). Decaying organisms in pitfall traps may attract insect-feeding vertebrates (e.g., Caracara plancus (Miller, 1777), Oliveira-Ribeiro personal observation; Young, 2015), resulting in the consumption of dung beetles within pitfall traps. Considering the decrease of attractiveness after 48 h and the biased attractiveness caused by decaying material, the time duration of 48 h is the most appropriate for dung beetle surveys at the assemblage level, although it is also possible to re-bait traps every 24 or 48 h, eliminating the problem of attractiveness decline. However, it would lead to an increase in the time spent on collecting.
Trap spacing had an astonishing range, from two to 1,000 m apart. Nonetheless, most studies (more than 80%) spaced traps up to 50 m. Standardized trap spacing guarantees accurate ecological comparisons among ecological studies (Larsen and Forsyth, 2005; Noriega and Fagua, 2009; da Silva and Hernández, 2015; but see Moctezuma, 2021). Dung beetle trap spacing relates to the study sampling unit: studies in which traps are treated as individual samples require spatial independence, while studies that consider a set of traps as a sample need spatial independence among samples. To determine the appropriate trap spacing that avoids pseudo replication issues (i.e., guaranteeing spatial independence among traps or set of traps), previous studies tried to assess the optimal distance among sampling units (Larsen and Forsyth, 2005; da Silva and Hernández, 2015). According to these studies, trap spacing from 50 to 150 m (depending on the mobility of the species and environmental conditions) would be an adequate distance to avoid interference between samples. In studies that evaluate the landscape process, it is most beneficial to distribute traps in a way that allows effective regional sampling, which is limited by the smallest study sites (e.g., islands, forest fragments, see Filgueiras et al., 2015; Storck-Tonon et al., 2020; Rodriguez-Garcia et al., 2021). Whenever a habitat is spatially limited, traps need to be clustered spatially, and thus trap spacing can be relatively small (e.g., Arellano et al., 2005; Costa et al., 2013). Traps can be installed close to each other to evaluate bait attractiveness or food preference (e.g., 2–3 m; see Louzada and Carvalho e Silva, 2009; Correa et al., 2018), while ecological studies that do not aim to sample the diversity of a region (e.g., studies of seed dispersal or to obtain a focal species), optimum trap spacing is not necessarily a rule.
The highest percentage of studies used only one sample (space-for-time replicates), due to several reasons. Some large-scale studies (i.e., comparing bioregions) use few samples, either to randomize a large number of sites avoiding pseudoreplication or to study biogeographical patterns (e.g., da Silva P. G. et al., 2022). Other studies focused more on behavior, natural history, or ecosystem services (e.g., Salomão et al., 2018; Noriega et al., 2021a), do not usually include gradients or a spatial analysis comparison. In addition, studies that are not necessarily large-scale will choose small sampling replicates to avoid spatial autocorrelation (Leather et al., 2014; Negrete-Yankelevich and Fox, 2015) or to study spatiotemporal diversity (e.g., Ferreira et al., 2018). Twenty percent of the reviewed studies used two or three samplings, most of which used spatial controls or replicas of the same habitat (Gómez-Cifuentes et al., 2019), while ~22% of studies used from four to nine replicates, including works with spatial replicability, studying beetles at the landscape level (e.g., Ramírez-Ponce et al., 2019; Correa et al., 2021). Studies with a larger number of samples (>10) are mainly due to studies with multi-year sampling (e.g., Salomão et al., 2020; Noriega et al., 2021b), studies in wider areas with multiple sites and replicates, or analyzing longer gradients (e.g., Vulinec, 2002; Correa et al., 2019). Lastly, the absence of detailed information on sampling or replication in some articles is a widespread pattern in other sub-themes, where the description of the methodological component is very incomplete, especially when the articles are concerned with details of natural history, food preferences, phenology, etc.
In terms of seasonality and temporal sampling, the rainy season may be ideal for collecting a higher abundance of adults that can be attracted to baited traps (Halffter and Matthews, 1966; Andresen, 2005; Correa et al., 2018). This is due to the behavior of dung beetles, which is strongly influenced by the rains (Halffter and Matthews, 1966; Doube, 1991; Hanski and Cambefort, 1991) and temperatures throughout the year (Verdú et al., 2006; Hernández and Vaz-de-Mello, 2009; da Silva et al., 2018). These activity peaks in rainy periods (mainly in environments with slight thermal variation throughout the year) are related to the physiological characteristics of insects, which must be able to survive by minimizing the loss of body water (Verdú et al., 2019), extracted from the trophic resources (e.g., excrement or other types of organic matter). In addition, the moisture of the resource, or the amount of water that the excrement can hold, is an important factor both in the spread of smell over long distances and the water availability provided by these beetles (Fletchmann et al., 2009; Dormont et al., 2010; Holter, 2016). Baits in traps for dung beetles suffer intense dehydration in dry periods, which produces a lower attractiveness (Lobo et al., 1998). Nonetheless, it is important to consider that seasonality among insects in the tropics is still uncertain (Kishimoto-Yamada and Itioka, 2015), mainly in ecosystems that are evergreen and that do not have a marked dry season. Such an argument is often used in ecological studies of dung beetles in the Neotropics that are performed during the dry season (e.g., Salomão et al., 2019b).
Ambient temperature is an excellent variable in predicting Neotropical dung beetle species richness (Lobo et al., 2018). Although some dung beetle species can slightly control their body temperature (e.g., Verdú and Lobo, 2008; Gallego et al., 2018); they are animals that depend on environmental temperature to perform their physiological functions, with an ideal temperature range (Chown, 2001; Sheldon et al., 2011). As humidity and temperature are strongly associated, spring or rainfall periods are suitable for these individuals to leave the nests for feeding or reproductive purposes. In this sense, it is important to take into account that the dispersion of individuals at these times can mask the dependence that many species have on their habitat since, during these favorable periods, it is possible to capture species in habitats where they would not survive during the dry season (Hernández et al., 2014). Another relevant issue is that unfavorable environmental conditions are less critical in burrowing species since they are less subject to seasonal climatic variations, remaining in the tunnels for long periods, where they have sufficient food for themselves and their offspring (Halffter and Matthews, 1966; Hanski and Cambefort, 1991; Scholtz et al., 2009).
In this review, we examined the various methods used to depict dung beetle assemblages, diversity, and abundance in 241 papers published in peer-reviewed journals. We limited our search to the Neotropics, and those papers focused on dung beetle biodiversity. We analyzed several variables related to trapping design. Based on our analysis, we made a series of recommendations for the optimal procedures to examine dung beetle diversity and abundance, and we propose some minimum requirements for a standard protocol (see Box 1). In the interest of staying within our stated scope in this paper, we did not delve into other issues of importance for dung beetle diversity studies. We did not examine the methods for collecting beetles once they are in the trap, such as what kill solution is preferred (for example, the old technique of using ethylene glycol is no longer recommended because of its toxic effect on mammals). We also did not discuss live-trapping versus kill-trapping, labeling, storage, or identification (still problematic due to the many beetle species and the low number of taxonomists). In addition, several concerns in dung beetle studies were not discussed here, including the definition of diversity, the best metrics to use in describing dung beetle assemblages, and what statistical methods should be employed in comparing two or more habitats, to name a few. These are more complex and controversial subjects and need to be examined further.

One of the critical aspects of scientific studies is the possibility of replicating them. In ecological studies, the sampling design (i.e., number of samples, spatio-temporal distribution of the samples, detailed sampling technique) configure as a primordial aspect that will allow replicability in further studies. Under such rationale, it is essential to note that a considerable number of ecological studies in this review did not correctly detail their sampling methodology. For example, some studies do not include the number of pitfall traps used, which is a determinant in calculating capture rate and ensuring replicability. Besides, the absence of detailed information on sampling or replication in some articles is a widespread pattern in other sub-themes. It is often expected that the characterization of methodological components is very poor, especially when the articles are concerned with details of natural history, food preferences, phenology, etc. Such practices may come together with the requirements of ecological journals for manuscripts to present a concise description of methods. With the data presented in this study, we reinforce the importance of adequately detailing sampling methodology in ecological studies encompassing the dung beetles.
In this context, we found different information gaps in both geographical and methodological contexts. This investigation shows how multiple countries of the Neotropical region present limited knowledge on dung beetles (evidenced by the few researches found). For this reason, an increased research effort is recommended on dung beetles in countries with high biodiversity potential, such as Ecuador, Peru, and Venezuela. In this context, dung beetles have been used in different meta-analyses (e.g., Nichols et al., 2007; López-Bedoya et al., 2022). However, a substantial limitation of interpretation of results when evaluating global patterns is the standardization of methodologies between investigations included in a meta-analysis. To provide better conclusions on global patterns, it is necessary to start with methodological standardization on dung beetles. The idea of standardizing a sampling methodology is to allow comparisons on a large scale (continental, neotropics, etc.), allowing for studying global changes. However, we understand that we are still some ways away from standardization, as the more crucial things while developing a methodology are the question we want to answer and the material and financial resources available to answer it. In saying that, we acknowledge that much of the research carried out in Latin America has limited funding, so the methodology followed is subject to the financial and human resources available to researchers.
Standardization in models and size of other types of traps (including pitfall) is poorly evaluated. In some cases, the experience in the right location, orientation, selection of corridors or gaps, and the number of this type of traps and samples (e.g., mini-Winkler, interception) could be an essential factor in efficiency in obtaining favorable results in rare species of scarabs. This inexperience could be derived from the fact that this type of trap is more commonly used by taxonomists o to other groups of beetles or insects. Collaboration between experts in ecology and taxonomy should be mandatory for the correct determination of species, better analysis of data, and a deep study of the numerous specimens collected and frequently stored. This way, several of these specimens or a synoptic sample should be preserved, pinned, and deposited in public collections. The standardized methodology for sampling dung beetles suggested in this study (see Box 1) may be helpful for studies directed toward the knowledge of dung beetle fauna that can support data on the species distribution in the Neotropical region. Here, we offer a methodological guideline that can be replicated in studies at local scales in the Neotropics or other regions of the world with a high diversity of dung beetles (e.g., Afrotropical, Oriental).
Considering the time, financial and logistical resources, which in many cases are scarce, we suggest a standardization for the dung beetles sampling. The use of transect for dung beetle sampling is widely used (e.g., Gardner T. et al., 2008; Gardner T. A. et al., 2008; Macedo et al., 2020; Noriega et al., 2021a), being suitable because it presents a greater amplitude in the dispersion of the odor plume of the baits when compared with other denser configurations of traps. Considering that most forest areas in the Neotropics are small (e.g., Ribeiro et al., 2009), very long transects are not possible due to the edge effect (see Laurance and Bierregaard, 1997); therefore, the distance of 100 between traps within the same site is more feasible, considering the configuration of the areas, especially forest areas in the Neotropics. However, if it is not possible to use 100 m, at least a minimum distance of 50 m is recommended. We suggest five or six traps per site (see Rivera and Favila, 2022), therefore considering a transect of 400 m, with five traps spaced at 100 m by type of bait, considered as a replica, and at least three replicas, regarding the distance between sites.
We suggest that the distance between sites should not be <1.5 km since large species can disperse, covering a distance of 1 km in 2 days (Peck and Forsyth, 1982). This distance between areas ensures that traps installed for 48 h do not receive dung beetles from other areas studied. Small traps can limit the capture of beetles, especially in tropical forests, which within 48 h, can attract large numbers of insects. Therefore, we suggest traps with a capacity of at least 0.5–1 L, and around 1/3 of their liquid capacity can be added (water + salt + neutral detergent). The short period (48 h) makes the salt efficient for preserving the dung beetles and does not interfere with the attraction of the beetles. The detergent helps to break the surface tension of the water. For bait, plastic coffee cups (50 mL) are easy to find in any country, so we suggest their use, with the bait supply in at least 1/3 of its capacity, with a mix between pig-human dung bait, due to the ease of use, wherever the researcher is (Marsh et al., 2013). As a suggestion for the pitfall traps, plastic plates can be used to avoid the bait’s desiccation and prevent its contents from leaking due to rain. We also suggest installing the traps in places with little solar radiation; for this, the researcher can find more suitable places as far as possible. These suggestions aim at the bait’s attractiveness during the entire period of trap activity.
In terms of potential research areas, there are several future studies. One topic that has not been fully explored is the attraction of dung beetles to specific chemicals. In order to know and determine the volatile organic compounds (VOCs) to which beetles are most attracted may evaluate the best baits to use in the field. This multi-disciplinary research area requires chemical analysis of the VOCs, followed by lab and field behavioral trials. Another aspect to explore is experimental approaches focused on diversity complementarity by comparing different types of traps, types of baits, and proportions of bait mixes need to be explored. It is essential to include and evaluate several types of baits and species or groups of mushrooms, fruits, carrion, dung, and mesofauna, as myriapods and gastropods are necessary. In addition, sample coverage estimators may be considered in future meta-analysis studies aiming to propose the most appropriate sampling effort in dung beetle studies in different Neotropical ecosystems.
In terms of potential aspects to improve is the omission of information on the richness and abundance captured in each trap in ecological studies related to dung beetles. Providing this information as Supplementary material in future studies would not mean any additional effort because it is an essential step in organizing and analyzing the information in this type of study. Henceforth, we recommend providing this information as the best way to improve our understanding of the efficiency of the methodologies implemented (e.g., type of trap, the number of traps, the type of bait, the distance between traps) in research related to dung beetles. Analyzing this information will let us propose standardized, efficient sampling protocols for dung beetles in the upcoming years.
Due to the decrease in biodiversity due to anthropic causes, we believe it is crucial to think about pitfall traps that can keep the specimens alive after their identification in the field. Many species that fall into the traps are easily identifiable, and after being counted (and perhaps marked to avoid counting them in the following capture), these specimens could be released and returned to nature, thinking that they can continue to perform their ecosystem functions. Those species that are not easily identified or of which there are few specimens in scientific collections can be captured and deposited, as well as vouchers for each species. The need for intensive work on the biodiversity of many taxa is becoming more essential as climate change and habitat destruction increase, and surveys of organisms over time will enhance our understanding of the detrimental effects of these factors on our biodiversity (Brodie et al., 2012). Whatever way one defines biodiversity, it is still well known that the wealth of species once seen on this planet is rapidly declining. Documenting this decline may give us clues to ameliorate it if we can do this quickly. Dung beetles are a suitable ecological indicator species, have essential roles in the ecosystem, and are a proxy for surveying large mammalian fauna, especially in the Neotropics. This is why it is critical to standardize our survey methodology and promote solid techniques across the landscapes where we collect these valuable insects.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
EM-A and JN conceived the idea, designed the research, and structured the manuscript. EM-A, AA-J, CC, PS, VK, PL-B, MH, JP-C, RS, GV, and JN gathered and analyzed the data. All authors interpreted the results, contributed to the writing of the paper, and approved the last version of the document.
Funding was provided to DE and FE by the Natural Environment Research Council (grant no. NE/R017441/1). MH thanks CNPq (Science and Technology Ministry of Brazil), for a Productivity Grant (proc. 304713/2021-0). PS thanks the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) for post-doctoral grant (Process 88882.316025/2019-01, Code 001). RS was supported by Programa Nacional de Pós-doutorado/Capes (Government funds PNPD/CAPES, Brazil). This is publication #36 of the Biodiversity, Agriculture, and Conservation in Colombia [Biodiversidad, Agricultura, y Conservación en Colombia (BACC)] project. PALB was supported by a Master’s scholarship from Minas Gerais State Agency for Research and Development (FAPEMIG).
We thank Roberta Moura for his valuable participation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1096208/full#supplementary-material
Abot, A. R., Puker, A., Taira, T. L., Rodrigues, S. R., Korasaki, V., and Oliveira, H. N. (2012). Abundance and diversity of coprophagous beetles (Coleoptera: Scarabaeidae) caught with a light trap in a pasture area of the Brazilian Cerrado. Stud. Neotrop. Fauna Environ. 47, 53–60. doi: 10.1080/01650521.2012.662846
Andresen, E. (2005). Effects of season and vegetation type on community organization of dung beetles in a tropical dry forest. Biotropica 37, 291–300. doi: 10.1111/j.1744-7429.2005.00039.x
Arellano, L., Favila, M. E., and Huerta, C. (2005). Diversity of dung and carrion beetles in a disturbed Mexican tropical montane cloud forest and on shade coffee plantations. Biodivers. Conserv. 14, 601–615. doi: 10.1007/s10531-004-3918-3
Barbosa, L. G., Santos Alves, M. A., and Viveiros Grelle, C. E. (2021). Actions against sustainability: dismantling of the environmental policies in Brazil. Land Use Policy 104:105384. doi: 10.1016/j.landusepol.2021.105384
Barraza, J. M., Montes, J. F., Martínez, N. H., and Deloya, C. (2010). Ensamblaje de escarabajos coprófagos (Scarabaeidae: Scarabaeinae) del Bosque Tropical Seco, Bahía Concha, Santa Marta (Colombia). Rev. Col. Entomol. 36, 285–291. doi: 10.25100/socolen.v36i2.9160
Becker, C. G., Fonseca, C. R., Haddad, C. F. B., Batista, R. F., and Prado, P. I. (2007). Habitat split and the global decline of amphibians. Science 318, 1775–1777. doi: 10.1126/science.1149374
Boetzl, F. A., Ries, E., Schneider, G., and Krauss, J. (2018). It’s a matter of design—how pitfall trap design affects trap samples and possible predictions. PeerJ 6:e5078. doi: 10.7717/peerj.5078
Bogoni, J. A., and Hernández, M. I. M. (2014). Attractiveness of native mammal’s feces of different trophic guilds to dung beetles (Coleoptera: Scarabaeinae). J. Insect Sci. 14:299. doi: 10.1093/jisesa/ieu161
Boiteau, G. (2000). Efficiency of flight interception traps for adult Colorado potato beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 93, 630–635. doi: 10.1603/0022-0493-93.3.630
Bourg, A., Escobar, F., MacGregor-Fors, I., and Moreno, C. E. (2016). Got dung? Resource selection by dung beetles in Neotropical forest fragments and cattle pastures. Neotrop. Entomol. 45, 490–498. doi: 10.1007/s13744-016-0397-7
Braga, R. F., Korasaki, V., Andresen, E., and Louzada, J. (2013). Dung beetle community and functions along a habitat-disturbance gradient in the Amazon: a rapid assessment of ecological functions associated to biodiversity. PLoS One 8:e57786. doi: 10.1371/journal.pone.0057786
Brodie, J. F., Post, E., and Laurance, W. (2012). Climate change and tropical biodiversity: a new focus. Trends in Ecology & Evolution 27, 145–150.
Brown, G. R., and Matthews, I. M. (2016). A review of extensive variation in the design of pitfall traps and a proposal for a standard pitfall trap design for monitoring ground-active arthropod biodiversity. Ecol. Evol. 6, 3953–3964. doi: 10.1002/ece3.2176
Buchholz, S., and Möller, M. (2018). Assessing spider diversity in grasslands – does pitfall trap color matter? J. Arachnol. 46, 376–379. doi: 10.1636/JoA-S-16-062.1
Cardoso, P., Barton, P. S., Birkhofer, K., Chichorro, F., Deacon, C., Fartmann, T., et al. (2020). Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 242:108426. doi: 10.1016/j.biocon.2020.108426
Cardoso, P., and Leather, S. R. (2019). Predicting a global insect apocalypse. Insect Conserv. Divers. 12, 263–267. doi: 10.1111/icad.12367
Chamorro, W. R., Gallo, F. O., Delgado, S., Enríquez, S. I., Guasumba, V., and López-Iborra, G. (2019). Los escarabajos estercoleros (Coleoptera: Scarabaeidae: Scarabaeinae) del Bosque Protector Oglán Alto, Pastaza. Ecuador. Biota Colomb. 20, 34–49. doi: 10.21068/c2019.v20n01a03
Chown, S. L. (2001). Physiological variation in insects: hierarchical levels and implications. J. Insect Physiol. 47, 649–660. doi: 10.1016/S0022-1910(00)00163-3
Correa, C. M. A., Braga, R. F., Puker, A., Abot, A. R., and Korasaki, V. (2018). Optimising methods for dung beetle (Coleoptera: Scarabaeidae) sampling in Brazilian pastures. Environ. Entomol. 47, 48–54. doi: 10.1093/ee/nvx191
Correa, C., Braga, R. F., Puker, A., and Korasaki, V. (2019). Patterns of taxonomic and functional diversity of dung beetles in a human-modified variegated landscape in Brazilian Cerrado. J. Insect Conserv. 23, 89–99. doi: 10.1007/s10841-018-00118-6
Correa, C. M. A., da Silva, P. G., Puker, A., and Abot, A. R. (2021). Spatiotemporal patterns of taxonomic and functional β-diversity of dung beetles in native and introduced pastures in the Brazilian Pantanal. Austral Ecol. 46, 98–110. doi: 10.1111/aec.12963
Correa, C. M. A., Puker, A., Korasaki, V., Ferreira, K. R., and Abot, A. R. (2016). The attractiveness of baits to dung beetles in Brazilian Cerrado and exotic pasturelands. Entomol. Sci. 19, 112–123. doi: 10.1111/ens.12169
Correa-Cuadros, J. P., Gómez-Cifuentes, A., and Noriega, J. A. (2022). Comparative effect of forest cutting and mammal hunting on dung beetle assemblages in Chocó biogeographic forests in Colombia. Int. J. Trop. Insect Sci. 42, 3045–3055. doi: 10.1007/s42690-022-00839-x
Costa, F. C., Pessoa, K. K. T., Liberal, C. N., Filgueiras, B. C. K., Salomão, R. P., and Iannuzzi, L. (2013). What is the importance of open habitat in a predominantly closed forest area to the dung beetle (Coleoptera, Scarabaeinae) assemblage? Rev. Bras. Entomol. 57, 329–334. doi: 10.1590/S0085-56262013000300012
da Costa, C. M. Q., Silva, F. A. B., Farias, A. I., and Moura, R. C. (2009). Diversity of Scarabaeinae (Coleoptera, Scarabaeidae) collected with flight intercept trap in the Charles Darwin ecologic refuge, Igarassu-PE. Brazil. Rev. Bras. Entomol. 53, 88–94. doi: 10.1590/S0085-56262009000100021
da Silva, W. B., Cajaiba, R. L., Santos, M., and Périco, E. (2022). Effectiveness of ant communities to detect anthropogenic disturbance in Neotropical forest landscapes: a case study in the Brazilian Amazon. Biodivers. Conserv. 31, 211–226. doi: 10.1007/s10531-021-02329-x
da Silva, P. G., and Hernández, M. I. M. (2015). Spatial patterns of movement of dung beetle species in a tropical forest suggest a new trap spacing for dung beetle biodiversity studies. PLoS One 10:e0126112. doi: 10.1371/journal.pone.0126112
da Silva, P. G., Lobo, J. M., Hensen, M. C., Vaz-de-Mello, F. Z., and Hernández, M. I. M. (2018). Turnover and nestedness in subtropical dung beetle assemblages along an elevational gradient. Divers. Distrib. 24, 1277–1290. doi: 10.1111/ddi.12763
da Silva, P. G., Souza, J. G., and Neves, F. D. S. (2022). Dung beetle B-diversity across Brazilian tropical dry forests does not support the Pleistocene arc hypothesis. Austral Ecol. 47, 54–67. doi: 10.1111/aec.13080
de Moura, R. S., Noriega, J. A., Serpa, R. A., Vaz-de-Mello, F. Z., and Klemann, L. Jr. (2021). Dung beetles in a tight-spot, but not so much: quick recovery of dung beetles assemblages after low-impact selective logging in central Brazilian Amazon. For. Ecol. Manag. 494:119301. doi: 10.1016/j.foreco.2021.119301
Deloya, C., Madora, A. M., and Covarrubias, M. D. (2013). Scarabaeidae y Trogidae (Coleoptera) necrófilos de Acahuizotla, Guerrero, México. Rev. Colomb. Entomol. 39, 88–94.
Doré, M., Willmott, K., Leroy, B., Chazot, N., Mallet, J., Freitas, A. V. L., et al. (2021). Anthropogenic pressures coincide with Neotropical biodiversity hotspots in a flagship butterfly group. Divers. Distrib. 28, 2912–2930. doi: 10.1111/ddi.13455
Dormont, L., Jay-Robert, P., Bessière, J. M., Rapior, S., and Lumaret, J. P. (2010). Innate olfactory preferences in dung beetles. J. Exp. Biol. 213, 3177–3186. doi: 10.1242/jeb.040964
Doube, B. M. (1991). “Dung beetles of southern Africa” in Dung beetle ecology. eds. I. Hanski and Y. Cambefort (Princeton, NJ: Princeton University Press), 133–155.
Durães, R., Carrasco, L., Smith, T. B., and Karubian, J. (2013). Effects of forest disturbance and habitat loss on avian communities in a Neotropical biodiversity hotspot. Biol. Conserv. 166, 203–211. doi: 10.1016/j.biocon.2013.07.007
Engel, J., Hertzog, L., Tiede, J., Wagg, C., Ebeling, A., Briesen, H., et al. (2017). Pitfall trap sampling bias depends on bodymass, temperature, and trap number: insights from an individual-based model. Ecosphere 8:e01790. doi: 10.1002/ecs2.1790
Estrada, A., Anzures, D. A., and Coates-Estrada, R. (1999). Tropical rain forest fragmentation, howler monkeys (Alouatta palliata), and dung beetles at Los Tuxtlas, Mexico. Am. J. Primatol. 48, 253–262. doi: 10.1002/(SICI)1098-2345(1999)48:4<253::AID-AJP1>3.0.CO;2-D
Estrada, A., and Coates-Estrada, R. (2002). Dung beetles in continuous forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodivers. Conserv. 11, 1903–1918. doi: 10.1023/A:1020896928578
Ferreira, J., Aragão, L. E. O. C., Barlow, J., Barreto, P., Berenguer, E., Bustamante, M., et al. (2014). Brazil’s environmental leadership at risk. Science 346, 706–707. doi: 10.1126/science.1260194
Ferreira, S. C., da Silva, P. G., Paladin, A., and Di Mare, R. A. (2018). Climatic variables drive temporal patterns of α and β diversities of dung beetles. Bull. Entomol. Res. 109, 390–397. doi: 10.1017/S0007485318000676
Filgueiras, B. K. C., Liberal, C. N., Aguiar, C. D. M., Hernández, M. I. M., and Iannuzzi, L. (2009). Attractivity of omnivore, carnivore and herbivore mammalian dung to Scarabaeinae (Coleoptera, Scarabaeidae) in a tropical Atlantic rainforest remnant. Rev. Bras. Entomol. 53, 422–427. doi: 10.1590/S0085-56262009000300017
Filgueiras, B. C. K., Tabarelli, M., Leal, I. R., Vaz-de-Mello, F. Z., and Iannuzzi, L. (2015). Dung beetle persistence in human-modified landscapes: combining indicator species with anthropogenic land use and fragmentation-related effects. Ecol. Indic. 55, 65–73. doi: 10.1016/j.ecolind.2015.02.032
Fletchmann, C. A. H., Tabet, V. G., and Quintero, I. (2009). Influence of carrion smell and rebaiting time on the efficiency of pitfall traps to dung beetle sampling. Entomol. Exp. Applic. 132, 211–217. doi: 10.1111/j.1570-7458.2009.00885.x
França, F. M., Ferreira, J., Vaz-de-Mello, F. Z., Maia, L. F., Berenguer, E., Ferraz Palmeira, A., et al. (2020). El Niño impacts on human-modified tropical forests: consequences for dung beetle diversity and associated ecological processes. Biotropica 52, 252–262. doi: 10.1111/btp.12756
Fujisaki, I., Pearlstine, E. V., and Miller, M. (2008). Detecting population decline of birds using long-term monitoring data. Popul. Ecol. 50, 275–284. doi: 10.1007/s10144-008-0083-7
Gallego, B., Verdú, J. R., and Lobo, J. M. (2018). Comparative thermoregulation between different species of dung beetles (Coleoptera: Geotrupinae). J. Therm. Biol. 74, 84–91. doi: 10.1016/j.jtherbio.2018.03.009
Gardner, T., Barlow, J., Araujo, I. S., Ávila-Pires, T. C., Bonaldo, A. B., Costa, J. E., et al. (2008). The cost-effectiveness of biodiversity surveys in tropical forests. Ecol. Lett. 11, 139–150. doi: 10.1111/j.1461-0248.2007.01133.x
Gardner, T. A., Hernández, M. I. M., Barlow, J., and Peres, C. A. (2008). Understanding biodiversity consequences of habitat change: the value of secondary and plantations forests for Neotropical dung beetles. J. Appl. Ecol. 45, 883–893. doi: 10.1111/j.1365-2664.2008.01454.x
Gibbs, H. K., Ruesch, A. S., Achard, F., Clayton, M. K., Holmgren, P., Ramankutty, N., et al. (2010). Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Proc. Natl. Acad. Sci. U. S. A. 107, 16732–16737. doi: 10.1073/pnas.0910275107
Gill, B. (1991). “Dung beetles in tropical American forests” in Dung beetle ecology. eds. I. Hanski and Y. Cambefort (Princeton, NJ: Princeton University Press), 211–229.
Gómez-Cifuentes, A., Giménez, V. C., Moreno, C. E., and Zurita, G. A. (2019). Tree retention in cattle ranching systems partially preserves dung beetle diversity and functional groups in the semideciduous Atlantic Forest: the role of microclimate and soil conditions. Basic Appl. Ecol. 34, 64–74. doi: 10.1016/j.baae.2018.10.002
González-Hernández, A. L., Navarrete-Heredia, J. L., Quiroz-Rocha, J. A., and Deloya, C. (2015). Coleópteros necrócolos (Scarabaeidae: Scarabaeinae, Silphidae y Trogidae) del Bosque Los Colomos, Guadalajara, Jalisco, México. Rev. Mex. Biodivers. 86, 764–770. doi: 10.1016/j.rmb.2015.07.006
Halffter, G., and Favila, M. E. (1993). The Scarabaeinae (Insecta: Coleoptera): an animal group for analysing, inventorying and monitoring biodiversity in tropical rainforest and modified landscapes. Biol. Int. 27, 15–23.
Halffter, G., and Matthews, E. G. (1966). The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomol. Mex. 12-14, 1–312.
Halffter, G., Pineda, E., Arellano, L., and Escobar, F. (2007). Instability of copronecrophagous beetle assemblages (Coleoptera: Scarabaeinae) in a mountainous tropical landscape of Mexico. Environ. Entomol. 36, 1397–1407. doi: 10.1603/0046-225X(2007)36[1397:IOCBAC]2.0.CO;2
Hanski, I., and Cambefort, Y. (1991). Dung beetle ecology. Princeton University Press, New Jersey, USA. 520.
Hernández, M. I. M., Barreto, P. S. C. S., Costa, V. H., Creão-Duarte, A. J., and Favila, M. E. (2014). Response of a dung beetle assemblage along a reforestation gradient in a Restinga forest. J. Insect Conserv. 18, 539–546. doi: 10.1007/s10841-014-9645-5
Hernández, M. I. M., and Vaz-de-Mello, F. (2009). Seasonal and spatial species richness variation of dung beetle (Coleoptera, Scarabaeidae s. str.) in the Atlantic Forest of southeastern Brazil. Rev. Bras. Entomol. 53, 607–613. doi: 10.1590/S0085-56262009000400010
Hill, C. J. (1996). Habitat specificity and food preference of an assemblage of tropical Australian dung beetles. J. Trop. Ecol. 12, 449–460. doi: 10.1017/S026646740000969X
Hohbein, R. R., and Conway, C. J. (2018). Pitfall traps: a review of methods for estimating arthropod abundance. Wildl. Soc. Bull. 42, 597–606. doi: 10.1002/wsb.928
Holter, P. (2016). Herbivore dung as food for dung beetles: elementary coprology for entomologists. Ecol. Entomol. 41, 367–377. doi: 10.1111/een.12316
Howden, H. F., and Nealis, V. G. (1975). Effects of deforestation clearing in a tropical rain forest on the composition of the coprophagous scarab beetle fauna (Coleoptera). Biotropica 7, 77–83. doi: 10.2307/2989750
Iannuzzi, L., Liberal, C. N., de Souza, T. B., Pellegrini, T. G., da Cunha, J. C. S., Koroiva, R., et al. (2021). “Sampling methods for beetles (Coleoptera)” in Measuring arthropod biodiversity. eds. J. C. Santos and G. W. Fernandes (Cham: Springer), 125–185.
Kessler, H., and Balsbaugh, E. U. (1972). Succession of adult Coleoptera in bovine manure in east Central South Dakota. Ann. Entomol. Soc. Am. 65, 1333–1336. doi: 10.1093/aesa/65.6.1333
Kishimoto-Yamada, K., and Itioka, T. (2015). How much have we learned about seasonality in tropical insect abundance since Wolda (1988)? Entomol. Sci. 18, 407–419. doi: 10.1111/ens.12134
Larsen, T. H., and Forsyth, A. (2005). Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 37, 322–325. doi: 10.1111/j.1744-7429.2005.00042.x
Larsen, T. H., Lopera, A., and Forsyth, A. (2006). Extreme trophic and habitat specialization by Peruvian dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt. Bull. 60, 315–324. doi: 10.1649/0010-065X(2006)60[315:ETAHSB]2.0.CO;2
Laurance, W. F. (2007). Have we overstated the tropical biodiversity crisis? Trends Ecol. Evol. 22, 65–70. doi: 10.1016/j.tree.2006.09.014
Laurance, W. F., and Bierregaard, R. O. (1997). Tropical forests remnants: Ecology, management, and conservation of fragmented communities. Chicago: University of Chicago Press, 592.
Leather, S. R., Basset, Y., and Didham, R. K. (2014). How to avoid the top ten pitfalls in insect conservation and diversity research and minimise your chances of manuscript rejection. Insect Conserv. Divers. 7, 1–3. doi: 10.1111/icad.12066
Lobo, J. M., da Silva, P. G., Hensen, M. C., Amore, V., and Hernández, M. I. M. (2018). Exploring the predictive performance of several temperature measurements on Neotropical dung beetle assemblages: methodological implications. Entomol. Sci. 22, 56–63. doi: 10.1111/ens.12340
Lobo, J. M., Lumaret, J. P., and Jay-Robert, P. (1998). Sampling dung beetles in the French Mediterranean area: effects of abiotic factors and farm practices. Pedobiologia 42, 252–266.
Lobo, J. M., Martín-Piera, F., and Veiga, C. M. (1988). Las trampas pitfall con cebo, sus posibilidades en el estudio de las comunidades coprófagas de Scarabaeoidea (Col.). I. Características determinantes de su capacidad de captura. Rev. Ecol. Biol. Sol 25, 77–100.
Lopes, L. B., Pitta, R. M., Eckstein, C., Carneiroe Pedreira, B., Coelho, P., Sindeaux, E., et al. (2020). Diversity of coleopterans associated with cattle dung in open pastures and silvopastoral systems in the Brazilian amazon. Agrofor. Syst. 94, 2277–2287. doi: 10.1007/s10457-020-00549-8
López-Bedoya, P. A., Bohada-Murillo, M., Angel-Vallejo, M. C., Audino, L. D., Davis, A. L. V., Gurr, G., et al. (2022). Primary forest loss and degradation reduces biodiversity and ecosystem functioning: a global meta-analysis using dung beetles as an indicator taxon. J. Appl. Ecol. 59, 1572–1585. doi: 10.1111/1365-2664.14167
López-Bedoya, P. A., Magura, T., Edwards, F. A., Edwards, D. P., Rey-Benayas, J. M., Lövei, G. L., et al. (2021). What level of native beetle diversity can be supported by forestry plantations? A global synthesis. Insect Conserv. Divers. 14, 736–747. doi: 10.1111/icad.12518
Louzada, J. N. C., and Carvalho e Silva, P. R. (2009). Utilisation of introduced Brazilian pastures ecosystems by native dung beetles: diversity patterns and resource use. Insect Conserv. Divers. 2, 45–52. doi: 10.1111/j.1752-4598.2008.00038.x
Macedo, R., Audino, L. D., Korasaki, B. V., and Louzada, J. (2020). Conversion of Cerrado savannas into exotic pastures: the relative importance of vegetation and food resources for dung beetle assemblages. Agric. Ecosyst. Environ. 288:106709. doi: 10.1016/j.agee.2019.106709
Marsh, C. J., Louzada, J., Beiroz, W., and Ewers, R. M. (2013). Optimising bait for pitfall trapping of Amazonian dung beetles (Coleoptera: Scarabaeinae). PLoS One 8:e73147. doi: 10.1371/journal.pone.0073147
Martínez, M. I., and Suárez, M. T. (2006). Phenology, trophic preferences, and reproductive activity in some dung-inhabiting beetles (Coleoptera: Scarabaeoidea) in El Ilano de Las Flores, Oaxaca, Mexico. Proc. Entomol. Soc. Wash. 108, 774–784.
Martínez-Hernández, N. J., Rangel-Acosta, J. L., Beltrán-Díaz, H. J., and Daza-Guerra, C. A. (2022). ¡El tamaño sí importa! Incidencia del tamaño del cebo en la captura de escarabajos coprófagos en el bosque seco tropical. Rev. Biol. Trop. 70, 1–19. doi: 10.15517/rev.biol.trop.v70i1.46712
Martín-Piera, F., and Lobo, J. M. (1996). A comparative discussion of trophic preferences in dung beetle communities. Misc. Zool. 19, 13–31.
McKnight, D. T., Dean, T. L., and Ligon, D. B. (2013). An effective method for increasing the catch-rate of pitfall traps. Southwest. Nat. 58, 446–449. doi: 10.1894/0038-4909-58.4.446
Mendes, J., and Linhares, A. X. (2006). Coleoptera associated with undisturbed cow pats in pastures in southeastern Brazil. Neotrop. Entomol. 35, 715–723. doi: 10.1590/S1519-566X2006000600001
Missa, O., Basset, Y., Alonso, A., Miller, S. E., Curletti, G., De Meyer, M., et al. (2009). Monitoring arthropods in a tropical landscape: relative effects of sampling methods and habitat types on trap catches. J. Insect Conserv. 13, 103–118. doi: 10.1007/s10841-007-9130-5
Moctezuma, V. (2021). Spatial autocorrelation in a Mexican dung beetle ensemble: implications for biodiversity assessment and monitoring. Ecol. Indic. 125:107548. doi: 10.1016/j.ecolind.2021.107548
Mora-Aguilar, E. F., and Delgado, L. (2018). New species of Cryptocanthon Balthasar (Coleoptera: Scarabaeidae: Scarabaeinae) from the region of Chimalapas, Oaxaca, Mexico. Coleopt. Bull. 72, 792–796. doi: 10.1649/0010-065X-72.4.792
Mora-Aguilar, E. F., and Delgado, L. (2019). A new Mexican species of Rhyparus Westwood (Coleoptera: Scarabaeidae: Aphodiinae), with new records and a key to the Mexican and Guatemalan species. Zootaxa 4609, 196–200. doi: 10.11646/zootaxa.4609.1.13
Morelli, E., and Gonzalez-Vainer, P. (1997). Dung beetles (Coleoptera: Scarabaeidae) inhabiting bovine and ovine droppings in Uruguayan prairies. Coleopt. Bull. 51, 1–5.
Morón, M. A., and Terrón, R. (1984). Distribución altitudinal y estacional de los insectos necrófilos de la Sierra Norte de Hidalgo, México. Acta Zool. Mex. 3, 1–47. doi: 10.21829/azm.1984.132052
Morón-Ríos, A., and Morón, M. A. (2016). Evaluación de la fauna de Coleoptera Scarabaeoidea en la Reserva de la Biósfera de Calakmul, Campeche, México. Southwest. Entomol. 41, 469–484. doi: 10.3958/059.041.0217
Morrone, J. J., Escalante, T., Rodríguez-Tapia, G., Carmona, A., Arana, M., and Mercado-Gómez, J. D. (2022). Biogeographic regionalization of the Neotropical region: new map and shapefile. An. Acad. Bras. Cienc. 94:e20211167. doi: 10.1590/0001-3765202220211167
Myers, N., Mittermeier, R., Mittermeier, C., da Fonseca, G. A., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Negrete-Yankelevich, S., and Fox, G. A. (2015). “Spatial variation and linear modeling of ecological data” in Ecological statistics: Contemporany theory and application. eds. G. A. Fox, S. Negrete-Yankelevich, and V. J. Sosa (Oxford: Oxford Academic), 228–260.
Nichols, E., Larsen, T., Spector, S., Davis, A. L., Escobar, F., Favila, M., et al. (2007). Global dung beetle response to tropical forest modification and fragmentation: a quantitative literature review and meta-analysis. Biol. Conserv. 137, 1–19. doi: 10.1016/j.biocon.2007.01.023
Noriega, J. A. (2015). “How a locality can have so many species? A case study with dung beetles (Coleoptera: Scarabaeinae) in a tropical rain forest in Colombia” in Beetles: Biodiversity, ecology and role in the environment. ed. C. Stack (New York: Nova Science Publisher), 175–204.
Noriega, J. A., and Fagua, G. (2009). “Monitoreo de escarabajos coprófagos (Coleoptera: Scarabaeidae) en la Región Neotropical” in Técnicas de campo en ambientes tropicales: Manual para el monitoreo en ecosistemas acuáticos y artrópodos terrestres. eds. A. Acosta, G. Fagua, and A. M. Zapata (Bogotá: Editorial Pontificia Universidad Javeriana), 165–188.
Noriega, J. A., March-Salas, M., Castillo, S., Garcia-Q, H., Hortal, J., and Santos, A. M. C. (2021a). Human perturbations reduce dung beetle diversity and dung removal ecosystem function. Biotropica 53, 753–766. doi: 10.1111/btp.12953
Noriega, J. A., Santos, A. M. C., Calatayud, J., Chozas, S., and Hortal, J. (2021b). Short- and long-term temporal changes in the assemblage structure of Amazonian dung beetles. Oecologia 195, 719–736. doi: 10.1007/s00442-020-04831-5
Ong, X. R., Hemprich-Bennett, D., Gray, C. L., Kemp, V., Chung, A. Y. C., and Slade, E. M. (2022). Trap type affects dung beetle taxonomic and functional diversity in Bornean tropical forests. Austral Ecol. 47, 68–78. doi: 10.1111/aec.13124
Otavo, S. E., Parrado-Roselli, A., and Noriega, J. A. (2013). Superfamilia Scarabaeoidea (Insecta: Coleoptera) como elemento bioindicador de perturbación antropogénica en un parque nacional amazónico. Rev. Biol. Trop. 61, 735–752.
Page, M. J., Moher, D., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., et al. (2021). PRISMA 2020 explanation and elaboration: updated guidance and exemplars for reporting systematic reviews. BMJ 372:n160. doi: 10.1136/bmj.n160
Peck, S. B., and Forsyth, A. (1982). Composition, structure, and competitive behaviour in a guild of Ecuadorian rain forest dung beetles (Coleoptera; Scarabaeidae). Canadian J. Zool. 60, 1624–1634. doi: 10.1139/z82-213
Peck, S. B., and Howden, H. F. (1984). Response of a dung beetle guild to different sizes of dung bait in a Panamanian rainforest. Biotropica 16:235. doi: 10.2307/2388057
Pérez-Espona, S. (2021). Eciton Army ants—umbrella species for conservation in Neotropical forests. Diversity 13:136. doi: 10.3390/d13030136
Porter, S. D. (2005). A simple design for a rain-resistant pitfall trap. Insect. Soc. 52, 201–203. doi: 10.1007/s00040-004-0783-8
Prather, C. M., Pelini, S. L., Laws, A., Rivest, E., Woltz, M., Bloch, C. P., et al. (2013). Invertebrates, ecosystem services and climate change. Biol. Rev. 88, 327–348. doi: 10.1111/brv.12002
Puker, A., da Silva, K. K. G., dos Santos, D. C., Correa, C. M. A., and Vaz-de-Mello, F. Z. (2020). Dung beetles collected using flight intercept traps in an Amazon rainforest fragment and adjacent agroecosystems. Int. J. Trop. Insect Sci. 40, 1085–1092. doi: 10.1007/s42690-020-00132-9
Raine, E. H., Mikich, S. B., Lewis, O. T., and Slade, E. M. (2020). Linking dung beetle-mediated functions to interactions in the Atlantic Forest: sampling design matters. Biotropica 52, 215–220. doi: 10.1111/btp.12722
Raine, E. H., and Slade, E. M. (2019). Dung beetle–mammal associations: methods, research trends and future directions. Proc. R. Soc. Lond. B Biol. Sci. 286:20182002. doi: 10.1098/rspb.2018.2002
Ramírez-Ponce, A., Calderón-Patrón, J. M., Guzman, H. M., and Moreno, C. E. (2019). Biotic heterogeneity among scarab beetle communities in an anthropized landscape in the central valleys of Oaxaca, Mexico. J. Insect Conserv. 23, 765–776. doi: 10.1007/s10841-019-00169-3
Rangel-Acosta, J. L., and Martínez-Hernández, N. J. (2010). Comparison of copro-necrophagous beetle assemblages (Scarabaeidae: Scarabaeinae) among tropical dry forest fragments and the adjacent matrix in the Atlántico Department of Colombia. Rev. Mex. Biodiv. 88, 389–401. doi: 10.1016/j.rmb.2017.03.012
Ratcliffe, B. C., and Cave, R. D. (2009). The Dynastine scarab beetles of Mexico, Guatemala, and Belize (Coleoptera: Scarabaeidae: Dynastinae). Lincoln: University of Nebraska State Museum.
Ribeiro, M. C., Metzger, J. P., Martensen, A. C., Ponzoni, F. J., and Hirota, M. M. (2009). The Brazilian Atlantic forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 142, 1141–1153. doi: 10.1016/j.biocon.2009.02.021
Ríos-Touma, B., and Ramírez, A. (2019). “Chapter 12 – multiple stressors in the Neotropical region: environmental impacts in biodiversity hotspots” in Multiple stressors in river ecosystems. eds. S. Sabater, A. Elosegi, and R. Ludwig (Amsterdam: Elsevier), 205–220.
Rivera, J. D., and Favila, M. E. (2022). Good news! Sampling intensity needed for accurate assessments of dung beetle diversity may be lower in the Neotropics. Front. Ecol. Evol. 10:999488. doi: 10.3389/fevo.2022.999488
Rodrigues, S. R., Barros, A. T. M., Puker, A., and Ledesma, T. (2010). Diversidade de besouros coprófagos (Coleoptera, Scarabaeidae) coletados com armadilha de interceptação de voo no Pantanal Sul-Mato-Grossense, Brasil. Biota Neotrop. 10, 123–127. doi: 10.1590/S1676-06032010000200015
Rodriguez-Garcia, S., Carretero-Pinzon, X., Monroy, D., Medellin, F., Garcia, M., and Noriega, J. A. (2021). Influencia de la fragmentación de bosques de galeria en la Orinoquia Colombiana sobre la diversidad y estructura funcional del ensamblaje de escarabajos coprófagos. Bol. Cient. Mus. Hist. Nat. Univ. Caldas 25, 119–137. doi: 10.17151/bccm.2021.25.2.8
Salomão, R. P., Alvarado, F., Baena-Díaz, F., Favila, M. E., Iannuzzi, L., Liberal, C. N., et al. (2019b). Urbanization effects on dung beetle assemblages in a tropical city. Ecol. Indic. 103, 665–675. doi: 10.1016/j.ecolind.2019.04.045
Salomão, R. P., Favila, M. E., and González-Tokman, D. (2020). Spatial and temporal changes in the dung beetle diversity of a protected, but fragmented, landscape of the northernmost Neotropical rainforest. Ecol. Indic. 111:105968. doi: 10.1016/j.ecolind.2019.105968
Salomão, R. P., Favila, M. E., González-Tokman, D., and Chamorro-Florescano, I. A. (2019a). Contest dynamics for food and reproductive resources are defined by health condition in a dung beetle. Ethology 125, 343–350. doi: 10.1111/eth.12858
Salomão, R. P., Maia, A. C. D., Bezerra, B. M., and Iannuzzi, L. (2018). Attractiveness of different food resources to dung beetles (Coleoptera: Scarabaeidae) of a dry tropical area. Neotrop. Entomol. 47, 69–78. doi: 10.1007/s13744-017-0515-1
Sampaio, E. V. S. B. (1995). “Overview of the Brazilian Caatinga” in Seasonally dry tropical forests. eds. S. H. Bullock, H. A. Mooney, and E. Medina (Cambridge: Cambridge University Press), 35–63.
Sarges, R., Halffter, G., and Rojas, A. D. (2012). The importance of frugivory to the survival of the dung beetle Onthophagus rhinolophus Harold (Coleoptera: Scarabaeidae: Scarabaeinae) under changing ecological conditions. Coleopt. Bull. 66, 166–168. doi: 10.1649/072.066.0216
Scheffers, B. R., Joppa, L. N., Pimm, S. L., and Laurance, W. F. (2012). What we know and don’t know about Earth's missing biodiversity. Trends Ecol. Evol. 27, 501–510. doi: 10.1016/j.tree.2012.05.008
Schmitt, T., Krell, F., and Linsenmair, E. (2004). Quinone mixture as attractant for necrophagous dung beetles specialized on dead millipedes. J. Chemic. Ecol. 30, 731–740. doi: 10.1023/B:JOEC.0000028428.53797.cb
Scholtz, C. H., Davis, A. L. V., and Kryger, U. (2009). Evolutionary biology and conservation of dung beetles. Sofia: Pensoft.
Sheldon, K. S., Yang, S., and Tewksbury, J. J. (2011). Climate change and community disassembly: impacts of warming on tropical and temperate montane community structure. Ecol. Lett. 14, 1191–1200. doi: 10.1111/j.1461-0248.2011.01689.x
Silva, L. F. R., Castro, D. M. P., Juen, L., Callisto, M., Hughes, R. M., and Hermes, M. G. (2021). Functional responses of Odonata larvae to human disturbances in neotropical savanna headwater streams. Ecol. Indic. 133:108367. doi: 10.1016/j.ecolind.2021.108367
Silva, J. L., Silva, R. J., Fernandes, I. M., Sousa, W. O., and Vaz-de-Mello, F. Z. (2020). Species composition and community structure of dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) compared among savanna and forest formations in the southwestern Brazilian Cerrado. Fortschr. Zool. 37, 1–12. doi: 10.3897/zoologia.37.e58960
Southwood, T. R. E. (1978). Ecological methods: with particular reference to the study of insect populations. New York: Chapman and Hall.
Spence, J. R., and Niemelä, J. K. (1994). Sampling carabid assemblages with pitfall traps: the madness and the method. Can. Entomol. 126, 881–894. doi: 10.4039/Ent126881-3
Spooner, F. E. B., Pearson, R. G., and Freeman, R. (2018). Rapid warming is associated with population decline among terrestrial birds and mammals globally. Glob. Change Biol. 24, 4521–4531. doi: 10.1111/gcb.14361
Storck-Tonon, D., da Silva, R. J., Sawaris, L., Vaz-de-Mello, F. Z., da Silva, D. J., and Peres, C. A. (2020). Habitat patch size and isolation drive the near-complete collapse of Amazonian dung beetle assemblages in a 30-year-old forest archipelago. Biodivers. Conserv. 29, 2419–2438. doi: 10.1007/s10531-020-01982-y
Sullivan, G. T., Ozman-Sullivan, S. K., Lumaret, J. P., Bourne, A., Zeybekoglu, U., Zalucki, M. P., et al. (2017). How guilds build success; aspects of temporal resource partitioning in a warm, temperate climate assemblage of dung beetles (Coleoptera: Scarabaeidae). Environ. Entomol. 46, 1060–1069. doi: 10.1093/ee/nvx117
Touroult, J., Dalens, P.-H., Giuglaris, J.-L., Lapèze, J., and Boilly, O. (2017). Structure des communautés de Phanaeini (Coleoptera: Scarabaeidae) de Guyane: étude par échantillonnage massif au piège d’interception. Ann. Soc. Entomol. Fr. 53, 143–161. doi: 10.1080/00379271.2017.1319294
Trevilla-Rebollar, A., Deloya, C., and Padilla-Ramírez, J. (2010). Coleópteros necrófilos (Scarabaeidae, Silphidae y Trogidae) de Malinalco, Estado de México, México. Neotrop. Entomol. 39, 486–495. doi: 10.1590/S1519-566X2010000400005
Trujillo-Miranda, A. L., Carrillo-Ruiz, H., Rivas-Arancibia, S. P., and Andrés-Hernández, A. R. (2016). Structure and composition of the community of beetles (Coleoptera: Scarabaeoidea) from Chacateca's Hill, Zapotitlan, Puebla, Mexico. Rev. Mex. Biodivers. 87, 109–122. doi: 10.1016/j.rmb.2015.08.008
Valente-Neto, F. B., Téllez Martínez, R. M., Hughes, A., Ferreira, F., Severo-Neto, F., Leandro de Souza, R., et al. (2021). Incorporating costs, thresholds and spatial extents for selecting stream bioindicators in an ecotone between two Brazilian biodiversity hotspots. Ecol. Indic. 127:107761. doi: 10.1016/j.ecolind.2021.107761
Veiga, C. M., Lobo, J. M., and Martín-Piera, F. (1989). Las trampas pitfall con cebo, sus posibilidades en el estudio de las comunidades coprófagas de Scarabaeoidea (Col.). II. Análisis de efectividad. Rev. Ecol. Biol. Sol 26, 91–109.
Velloso, A. L., Sampaio, E. V. S. B., and Pareyn, F. G. C. (2002). Ecorregiões Propostas para o Bioma Caatinga. Recife: Instituto de Conservação Ambiental The Nature Conservancy do Brasil.
Verdú, J. R., Arellano, L., and Numa, C. (2006). Thermoregulation in endothermic dung beetles (Coleoptera: Scarabaeidae): effect of body size and ecophysiological constraints in flight. J. Insect Physiol. 52, 854–860. doi: 10.1016/j.jinsphys.2006.05.005
Verdú, J. R., Cortez, V., Oliva, D., and Giménez-Gómez, V. (2019). Thermoregulatory syndromes of two sympatric dung beetles with low energy costs. J. Insect Physiol. 118:103945. doi: 10.1016/j.jinsphys.2019.103945
Verdú, J. R., and Lobo, J. M. (2008). Ecophysiology of thermoregulation in endothermic dung beetles: ecological and geographical implications. Insect Ecol. Conserv. 9, 30–28. doi: 10.2307/2390087
Vulinec, K. (2002). Dung beetle communities and seed dispersal in primary forest and disturbed land in Amazonia. Biotropica 34, 297–309. doi: 10.1111/j.1744-7429.2002.tb00541.x
Vulinec, K., Lambert, J., and Mellow, D. J. (2006). Primate and dung beetle communities in secondary growth rainforests: implications for conservation of seed dispersal systems. Int. J. Primatol. 27, 855–879. doi: 10.1007/s10764-006-9027-2
Wassmer, T. (2020). Attractiveness of cattle dung to coprophilous beetles (Coleoptera: Scarabaeoidea and Sphaeridiinae) and their segregation during the initial stages of the heterotrophic succession on a pasture in Southeast Michigan. J. Insect Sci. 20, 1–15. doi: 10.1093/jisesa/ieaa040
Keywords: collecting methodology, flight intercept traps, monitoring protocol, tropical America, Scarabaeidae, survey
Citation: Mora-Aguilar EF, Arriaga-Jiménez A, Correa CMA, da Silva PG, Korasaki V, López-Bedoya PA, Hernández MIM, Pablo-Cea JD, Salomão RP, Valencia G, Vulinec K, Edwards FA, Edwards DP, Halffter G and Noriega JA (2023) Toward a standardized methodology for sampling dung beetles (Coleoptera: Scarabaeinae) in the Neotropics: A critical review. Front. Ecol. Evol. 11:1096208. doi: 10.3389/fevo.2023.1096208
Edited by:
William Wyatt Hoback, Oklahoma State University, United StatesReviewed by:
Andrey Frolov, Zoological Institute (RAS), RussiaCopyright © 2023 Mora-Aguilar, Arriaga-Jiménez, Correa, da Silva, Korasaki, López-Bedoya, Hernández, Pablo-Cea, Salomão, Valencia, Vulinec, Edwards, Edwards, Halffter and Noriega. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jorge Ari Noriega, am5vcmllZ0Bob3RtYWlsLmNvbQ==
†The author passed away prior to the submission of this paper
‡ORCID: Eder F. Mora-Aguilar https://orcid.org/0000-0002-6993-7243
Alfonsina Arriaga-Jiménez https://orcid.org/0000-0002-1242-7496
César M. A. Correa https://orcid.org/0000-0002-2804-7527
Pedro Giovâni da Silva https://orcid.org/0000-0002-5525-1291
Vanesca Korasaki https://orcid.org/0000-0001-6239-2976
Pablo A. López-Bedoya https://orcid.org/0000-0002-5525-1291
Malva Isabel Medina Hernández https://orcid.org/0000-0002-6820-5980
José D. Pablo-Cea https://orcid.org/0000-0001-5627-835X
Renato Portela Salomão https://orcid.org/0000-0001-9826-7472
Gorky Valencia https://orcid.org/0000-0003-3545-9818
Kevina Vulinec https://orcid.org/0000-0002-9068-9061
Felicity A. Edwards https://orcid.org/0000-0003-1742-2763
David P. Edwards https://orcid.org/0000-0001-8562-3853
Gonzalo Halffter https://orcid.org/0000-0003-1430-7049
Jorge Ari Noriega https://orcid.org/0000-0003-1760-7020
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.