
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Ecol. Evol., 26 April 2023
Sec. Phylogenetics, Phylogenomics, and Systematics
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1084009
This article is part of the Research TopicNeotropical Dung Beetle Diversity: Ecological, Historical, and Anthropogenic PerspectivesView all 11 articles
 Lucrecia Arellano1*†
Lucrecia Arellano1*† Jorge Ari Noriega2†
Jorge Ari Noriega2† Ilse J. Ortega-Martínez1†
Ilse J. Ortega-Martínez1† José D. Rivera1,3
José D. Rivera1,3 César M. A. Correa4
César M. A. Correa4 Andrés Gómez-Cifuentes5
Andrés Gómez-Cifuentes5 Alfredo Ramírez-Hernández6
Alfredo Ramírez-Hernández6 Felipe Barragán6
Felipe Barragán6Dung beetles are one of the most representative groups of insects associated with livestock, as they take advantage of the manure of livestock for food and reproduction. They have been widely used as a bio-indicator group to evaluate their responses to land-use change and other environmental disturbances by analyzing species diversity at different spatial and temporal scales. However, the impacts of livestock management practices, forms, and history on dung beetle diversity are still poorly understood. This paper is an exhaustive and systematic review of the existing peer-reviewed and indexed literature on the taxonomic diversity (species richness and composition), functional diversity, and ecological functions of dung beetles from different provinces and biogeographic domains in tropical grazing lands of the Neotropics. We analyzed the timeline of the studies conducted so far, and we detected increasing literature produced mainly in South America. We included the most frequent objectives, tendencies, software, and statistical analyses. Given the significant heterogeneity of livestock landscapes in the Neotropics, the reviewed studies have conceptualized broadly what a pasture is, and the authors have used different descriptions of other grazing lands. Additionally, management data and livestock practices vary widely among studies, and management history is poorly described. In future research, it is relevant to include as much information as possible and the consequences of different livestock management practices on additional ecological attributes of dung beetle assemblages at different landscape scales (spatial and temporal) to predict how ecological processes change in Neotropical landscapes. Considering the importance of the data to be included in future work, we propose a collection format to unify the information collected in the field when conducting ecological studies in grazing lands. Moreover, we define a classification system to homogenize the features that distinguish the multiple tropical grazing lands reported in the literature. Preserving dung beetle diversity and associated ecological functions is urgent for maintaining ecosystem services in grazing lands. The challenge for research institutions is to continue filling gaps in knowledge to help those who work in transferring knowledge, to help ranchers exercise better options for more sustainable livestock farming, and to publish results for conservation decision-making.
Livestock activities have transformed natural ecosystems into landscapes dominated by semi-open grazing areas associated with original vegetation for breeding several species of domesticated livestock (Guevara and Lira-Noriega, 2004). Projections for 2050 warn that the need for food will increase, such that land area destined for food production will increase (Herrero et al., 2015; FAO, 2017). Intensive and sustained land management over time has caused a cascading loss of native vegetation. There are, therefore, many overgrazed areas in the Neotropics that require special attention for biodiversity conservation purposes (Mellink and Riojas-López, 2020).
Biodiversity supports essential life-support services (Summers et al., 2012). Ecosystem services are the conditions and processes which provide ecosystem goods to sustain and fulfill human life and ecological continuity (e.g., food, medicine, firewood, freshwater; Daily, 1997; MEA [Millennium Ecosystem Assessment], 2005). Improving our knowledge of biodiversity conservation and its relationship with ecosystem services is critical in identifying humankind’s main ecological problems and finding solutions (Brand and Vadrot, 2013). According to Yu et al. (2017), there are several common indicators to monitor biodiversity loss and the implications for the sustainable provision of ecosystem services, and one of them is biodiversity per se because it has a substantial impact on ecosystem functions through the different functional traits presented among species in the community. An ecological indicator is a biological group sampled under various habitat conditions resulting from human activities (Calow, 1987; Moreno et al., 2007).
Dung beetles (Coleoptera: Scarabaeidae) are a valuable ecological indicator group for biodiversity assessment (Halffter and Favila, 1993; Spector, 2006; Nichols et al., 2007; Numa et al., 2009; Otavo et al., 2013), because they are well known taxonomically and ecologically, and susceptible to changes in habitat management (Favila and Halffter, 1997; Martínez et al., 2017; Barragán et al., 2021). Moreover, they exhibit wide variation in life history strategies and body sizes that are reflected in functional traits (Halffter and Edmonds, 1982; Hanski and Cambefort, 1991), as well as ecological functions that can be measured (e.g., dung removal, seed dispersion, soil bioturbation; see Andresen, 2002; Braga et al., 2013; Noriega et al., 2021a; Barragán et al., 2022). Therefore, dung beetles are adequate models for functional diversity and ecosystem functionality studies (Barragán et al., 2011; Braga et al., 2013), and they are essential for the maintenance of ecosystem functioning in grazing lands (Louzada and Carvalho e Silva, 2009). In this context, dung beetles are one of the most representative taxonomic groups of insects associated with grasslands and livestock. Most adult coprophagous beetles in grazing lands use fresh dung of mammalian herbivores (native, introduced, and domestic) for feeding and nesting; hence, dung removal has been one of the most measured ecological variables to evaluate dung beetle functionality (Holter, 2016). Consequently, dung beetle diversity is crucial for dung degradation; without them dung accumulates, leading to various health problems for human populations and ecological issues in ecosystems (Pecenka and Lundgren, 2018). Thus, the services performed by dung beetles reduces the use of financial resources to treat livestock health and soil fertilization (Lousey and Vaughan, 2006; Lopez-Collado et al., 2017).
We conducted an exhaustive and systematic evaluation of the peer-reviewed and indexed literature on the taxonomic (species richness and composition) and functional diversity, as well as the ecological functions of dung beetles in Neotropical grazing lands. We analyzed the timeline of the studies conducted to date and defined a classification system to homogenize the features that distinguish the multiple Neotropical grazing lands reported in the literature. Our review consists of several sections. First, we analyzed the regional and local changes in taxonomic dung beetle diversity (e.g., species richness, abundance, biomass), as well as the effect of grazing lands and livestock management on biodiversity from different provinces and biogeographic domains in tropical grazing lands of the Neotropics. Second, we discuss the meaning, importance, and vision of functional diversity studies on dung beetles. We considered functional groups and traits (both effect and response traits), including their selection, the most used, and the justification, in addition to analytical methods and relevant case studies. Third, we argued the relationship between biodiversity and ecosystem function (BEF). This relationship and its consequence for ecosystem services have predominantly been studied by controlled, short-term, and small-scale experiments under standardized environmental conditions and constant assemblage compositions. We discuss in the three sections the grazing land types and changes over time and we described these lands for understanding the dynamics of dung beetle assemblages. We describe livestock management practices and forms of management reported in the literature on dung beetle ecology and suggest some additional practices that we believe should be added to works on the subject. The history of land use is very important in this context. Finally, this article synthesizes the main findings, new research frontiers, and answers some open questions, current research gaps, potential developments in the field, and future challenges.
We used the Scopus and Web of Science databases to search for literature on the taxonomic and functional diversity and functions of dung beetles in pasture habitats, following the PRISMA methodology (Moher et al., 2009), which only considers indexed articles. The following search terms were used for the taxonomic diversity of dung beetles: ((“Dung beetle*” OR Scarabaeinae) AND disturb* AND (“Species richness” OR diversity OR Abundance) AND (Communit* OR Assemblage*) AND (“Tropical Forest*” OR Tropic*)). For functional diversity, we employed the following search terms: ((“Dung beetle*” OR Scarabaeinae) AND (disturb* OR “habitat disturb*” OR “land-use type*” OR “land-use change*” OR “land-use disturb*” OR anthropic* OR modification OR fragmentation OR “natural grassland*” OR Grassland* OR margeland* OR meadow* OR steppe OR “grazed rangeland*” OR Llanos OR Pampa OR savanna* OR Woodland* OR brushland* OR shrubland* OR Campos OR Cerrado OR Agroforestr* OR silvopastor* OR “forest grazing” OR “graz* forest*” OR Agriculture OR Pasture OR “livestoc* graz*” OR “cattle graz*” OR grazed OR “Open fores* “OR Temperature OR Microclim*) AND (“functional diversity” OR “functional trait*” OR “functional group*” OR Thermoregulation OR “ecological function*” OR “dung removal” OR “manure removal”) AND (Communit* OR Assemblage*) AND (“Tropical Forest” OR Tropic*)). Regarding dung beetle functions, we used the following search terms: ((“scarab*” OR “dung beetle*”) AND (remov* OR func* OR disper* OR biotur*) AND graz*). The search window of time covered articles published between January 1980 to February 2022.
Our search returned 272 taxonomic diversity articles, 109 functional diversity articles, and 81 articles regarding dung beetle functions. We complemented the taxonomic diversity search by including articles published in Spanish and Portuguese from the authors’ collection. We also surveyed among the three topics covering all articles obtained to add potential complementarity. The relevant articles for taxonomic and functional diversity and dung beetle functions were selected using the following criteria: i) the study includes species from Aphodiinae and/or Scarabaeinae subfamilies, ii) the study is based partly or entirely in grazing lands (pastures, cattle systems, agroforestry systems, and similar habitats), iii) the study evaluates taxonomical and/or functional diversity and/or dung beetle functions (e.g., dung burial, soil removal, fly control), and iv) the study is conducted in the Neotropics. Under these criteria, 76 taxonomic diversity articles, 26 functional diversity articles, and 18 dung beetle function articles were retained for data extraction (Figure 1; see Supplementary Table S1).
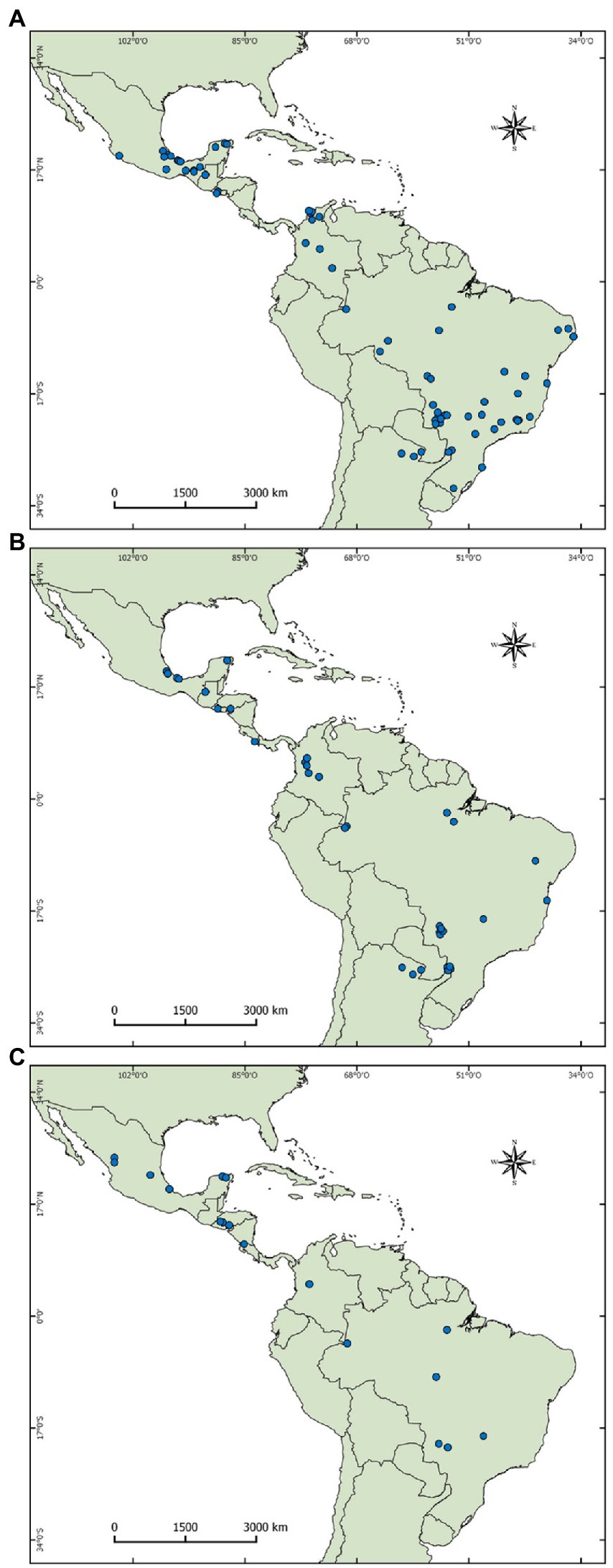
Figure 1. Maps with locations of dung beetle studies (Coleoptera: Scarabaeidae) in grazing environments included in the analytical review. (A) Taxonomic diversity studies, (B) functional diversity studies, (C) ecological functions studies.
We extracted the following information for all articles: (a) Neotropical biogeographic domains and provinces (sensu Morrone, 2017), including the year in which the study was published, the spatial and temporal scale, elevation (m a.s.l.) of the sampling sites, including sociopolitical divisions such as Town, County, State, and Country; (b) grazing land descriptions (e.g., grassland type), livestock management scheme and history, its surrounding matrix (at landscape scales) and whether cattle are present; (c) functional groups and traits of the dung beetle species; and (d) analytical methods and software used (Supplementary Table S2).
In this review, grazing land is any vegetated land that is grazed or that has the potential to be grazed by animals. Pasture lands are complex and globally diverse agricultural systems that vary in vegetation types (i.e., land cover) and management. Land cover in pasture systems can consist of (i) natural grasses alone, (ii) include naturally occurring shrubs and/or trees, or (iii) be entirely planted either in monocultures or as a component of mixed crop-live-stock systems. To analyze how authors have handled the concept of pasture and the description of grazing land and the cattle-grazed landscape, we have classified grazing lands (based on information from Butler et al., 2003; Allen et al., 2011; Holechek et al., 2011; Longland, 2013; Dixon et al., 2014; Costa et al., 2015; Hasanuzzaman, 2020; Oliveira et al., 2020; Jurado-Guerra et al., 2021). In Table 1, we show the grazing land definitions, descriptions, and categorizations (supported with pictures: Supplementary Figure S1) currently in use. We have classified grazing lands and proposed this classification for use in future studies by analyzing how different authors have handled the concept of pasture and the description of grazing lands and the cattle-grazed landscape.
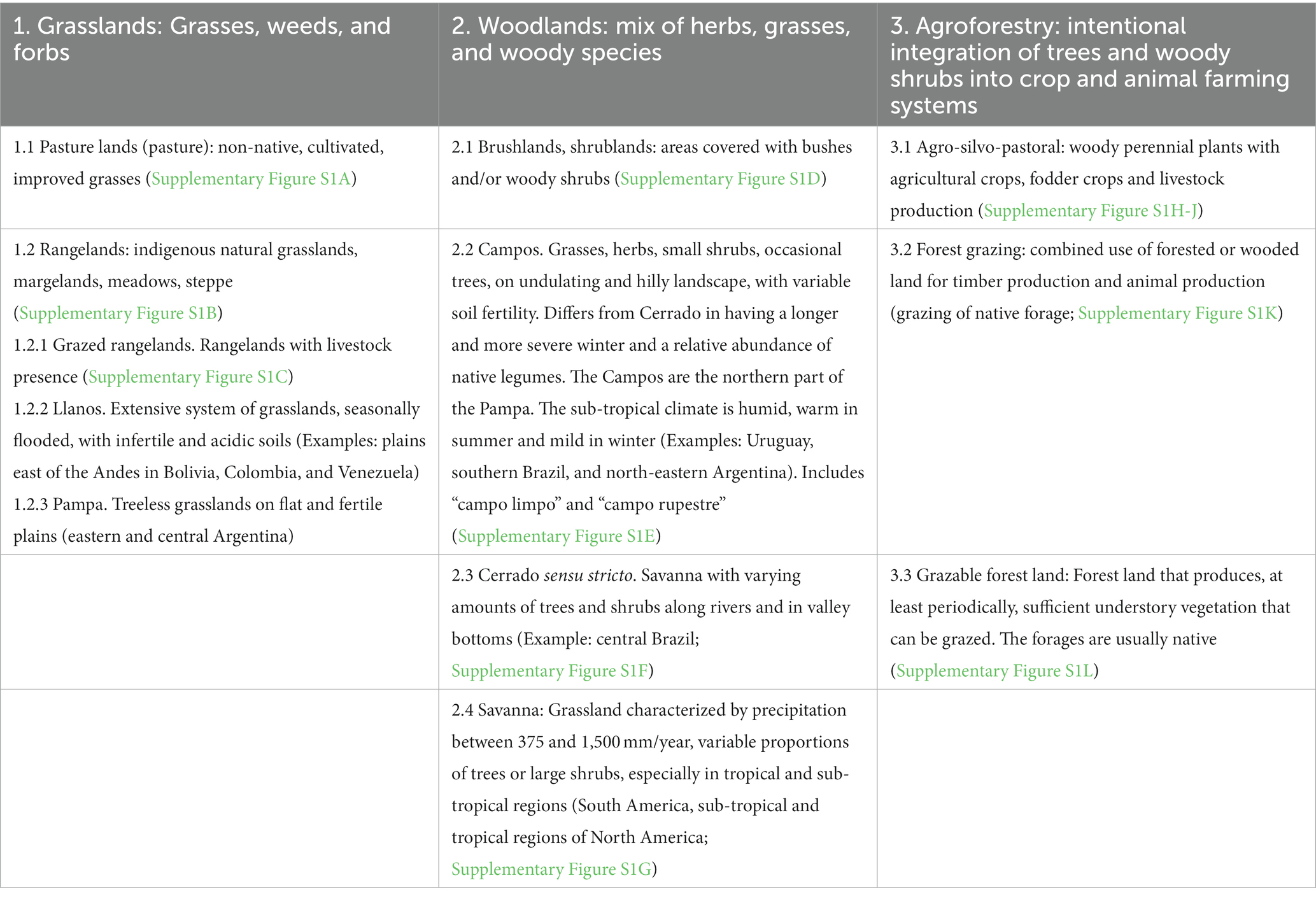
Table 1. Grazing land classification.
Grazing land management, in turn, varies in animal density, from animals moving freely over large areas (often called ‘extensive grazing’), to concentrated and rotated over small areas (termed ‘intensive rotational grazing’). Grazing land use also varies in terms of animal occupancy, with some systems primarily reliant on grazing and most others using pasture and feed lots (Oliveira et al., 2020) and livestock rotation. Other aspects in livestock management are livestock breed, production objective, types of grasses, water management for livestock, mating for livestock breeding, and infrastructure. Landowner management practices associated with grazing lands in this review are related to agrochemical use (herbicides, insecticides, dewormers), fertilizers, livestock feed supplements, fire, tillage, vitamins, and vaccines.
Seventy-six articles were analyzed in this section (Supplementary Table S1A). Publication numbers increased over time, where 8% (n = 6) were published in the 1990s, 24% (n = 18) in the 2000s, and 68% (n = 52) in the 2010s where the highest values were reached, and it seems that this trend will continue into the 2020s (due to the number of articles already available for publication in 2021 and 2022; Figure 2A). The most significant number of published papers came from Brazil (n = 45, 59.21%), followed by Mexico (n = 20, 26.31%), Colombia (n = 7, 9.21%), El Salvador-Nicaragua (n = 2, 2.63%) and Argentina (n = 2, 2.63%; Figure 2B). These tendencies are reflected in the biogeographic domains in which these studies were carried out; mainly in the Chacoan, Mesoamerican, Parana, and South Brazilian domains (Figure 2C). Brazil and Mexico have published papers on taxonomic diversity and grazing lands during the analyzed period, but Brazil increased its production of articles on this topic from 2010 onwards (n = 31).
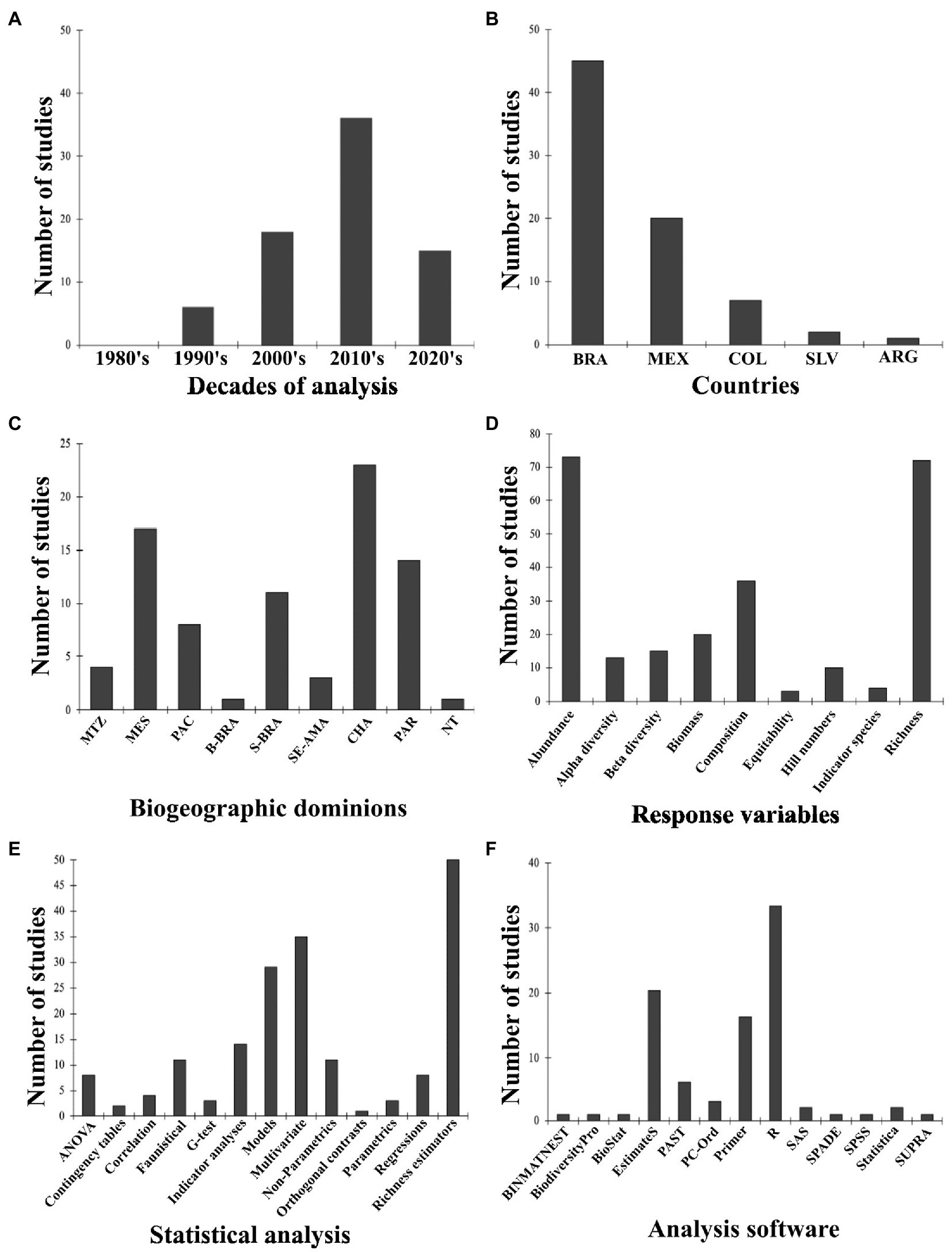
Figure 2. Number of taxonomic diversity studies. (A) Over time (decades). (B) Per country. Countries: Brazil = BRA, Mexico = MEX, Colombia = COL, El Salvador = SLV, Argentina = ARG. (C) Per biogeographic dominion. Mexican Tranzition Zone = MTZ, Mesoamerican = MES, Pacific = PAC, Boreal Brazilian = B-BRA, S-BRA = South Brazilian, South Eastern Amazonian = SE-AMA, Chacoan = CHA, Parana = PAR, Not tropical = NT. (D) Per response variable. (E) Per statistical analysis. (F) Per software protocol.
Most articles described grazing lands as grasslands (75.21% of the 91 sampling sites), woodlands in only 8.26% (n = 10), and agroforestry systems in 15.70% (n = 19). The proportions of each type of grassland, woodland and agroforestry system in the reviewed articles are shown in Supplementary Table S3. Pasture lands included exotic pastures and treeless pastures. The most common species of grasses (mainly African) are shown in Supplementary Table S4. The woodland systems essentially belonged to South America grazing lands like Cerrado sensu stricto, Pampa, Campos (“campo rupestre,” “campo limpo”), and Savanna (Supplementary Table S3). The principal agroforestry systems included silvopastoral systems with leguminous fodder plants (Vachellia spp., Guazuma ulmifolia Lam., Leucaena leucocephala (Lam.) de Wit, Brosimum alicastrum Sw.) and pastures, silvopastoral systems with native trees and fruit species, and silvopastoral systems with exotic species (e.g., Pinus spp.) and living fences (Supplementary Table S3).
Livestock management was included in the reviewed studies mainly for grazing description, animal information, production objective, water aspects, and infrastructure (Supplementary Table S5). Grazing information included animal rotation, type of grazing system, and fodder levels. Animal information included livestock stocking rate data and breed description. The main production objectives were meat and dual-purpose livestock (Supplementary Table S5). Information about water use only included irrigation and some aspects of water supplies. In terms of infrastructure, only the construction of stables is mentioned. We found that 43.63% of the reviewed manuscripts do not include livestock management aspects (Supplementary Table S5). Regarding the general objectives of the reviewed articles, the most common was the analysis of the effect of land use changes on taxonomic diversity (50% of all reviewed papers), followed by the impacts of surveys on distribution (26.32%), pasture management (13.16%), disturbance gradients (11.84%), fragmentation (9.21%), and silvopastoral systems (2.62%). The most used comparison type among the studies was diversity changes along management gradients (e.g., among forest, plantation, pasture, crops; 26.32% of the studies), followed by the forest vs. pasture diversity comparisons (19.74%), native vs. exotic pastures (13.16%), pastures (11.84%), exotic pasture diversity assessment (9.21%), vegetation gradients (e.g., primary forest, secondary forest, and pasture; 7.89%) and other comparisons representing the remainder (cattle presence or absence, restored gradients, living fences vs. pastures, natural fields, traps and bait functioning). The most important analyzed response variables were species richness and abundance, followed by the analysis of changes in species composition and biomass (Figure 2D). The more relevant tendencies were the decrease in species diversity from forest to pastures or along vegetation gradients from forest to pasture, or along management gradients. Species composition generally changed among conditions, mainly between wooded and open areas. In comparisons between native pastures or vegetation and exotic pastures, richness was higher in the native environments. In seasonal comparisons, the rainy season showed the highest richness.
Among the 76 articles evaluated, the most frequent analyses used were richness estimators (n = 50, 66.6%), multivariate analysis (n = 35, 46.6%), and models (n = 29, 38.6%). Richness estimator groups include Chao 1 and 2, Jackknife 1 and 2, Bootstrap, and Sample coverage. Multivariate analysis included Canonical Correlation Analysis (CCA), Principal Coordinate Analysis (PCO), Principal Component Analysis (PCA), Redundancy Analysis (RDA), and Permutational multivariate analysis of variance (PERMANOVA). Model group analyses consisted of Linear Models (LMs), Generalized Linear Models (GLMs), Generalized Linear Mixed Models (GLMMs), and Generalized Additive Models (GAMs; see Figure 2E, Supplementary Table S1). In relation to Software programs, R was the most used (n = 33, 44%), followed by EstimateS (n = 20, 26.6%) and Primer (n = 16, 21.3%). Finally, the R packages most used were iNext (n = 18, 24.0%), vegan (n = 17, 22.6%), and Betapart (n = 6, 8%; Figure 2F). Most of the studies that analyzed taxonomic diversity in grazing lands in the Neotropics have been carried out using a local spatial approach (~ 74%). It should be noted that 7.6% of the total articles reviewed had a scope at the local and landscape level; authors reflected on the variables affecting diversity at the local, landscape, and/or regional scales. Finally, very few studies (n = 7) make comparisons using a temporal approach (six were short-term and one was long-term).
The study of dung beetle functional diversity in Neotropical livestock systems began with Halffter et al. (2007) in Mexico. Moreover, Mexico is the only country that performed functional studies over the last three decades, with two studies per decade (Figure 3A). In the next decade, more than twice as many papers were published compared to the 2000s, with 60% of them conducted in Brazil. After 2020, the greatest number of functional studies have been published (n = 9, 34.62%; Figure 3A) in Brazil, the country with the most functional studies published to date, while El Salvador and Costa Rica showed opposite patterns, with one article published per country (Figure 3B). Additionally, the study of dung beetle functional diversity in livestock systems has been relatively new in countries such as Argentina (2019), Colombia (2020), and Costa Rica (2021; Figures 3A,B). Almost 40% of the reviewed papers were performed between two biogeographic provinces: 20% in Parana (between Brazil and Argentina) and 17% in Rondônia (Brazil). Moreover, 60% were divided among the remaining 13 provinces, most of them with only one published paper (Figure 3C). Approximately 57.70% of the articles were performed in the Parana (n = 8, 30.77%) and Chaco (n = 7, 26.92%) biogeographic domains. Mesoamerica (n = 6, 17.65%) and southern Brazil (n = 6, 17.65%) had equivalent numbers of studies, followed by the Mexican transition zone (n = 2, 5.88%), the Pacific (n = 2, 5.88%) and southeastern Amazon (n = 2, 5.88%), and the South American transition zone (n = 1, 2.94%; Figure 3C).
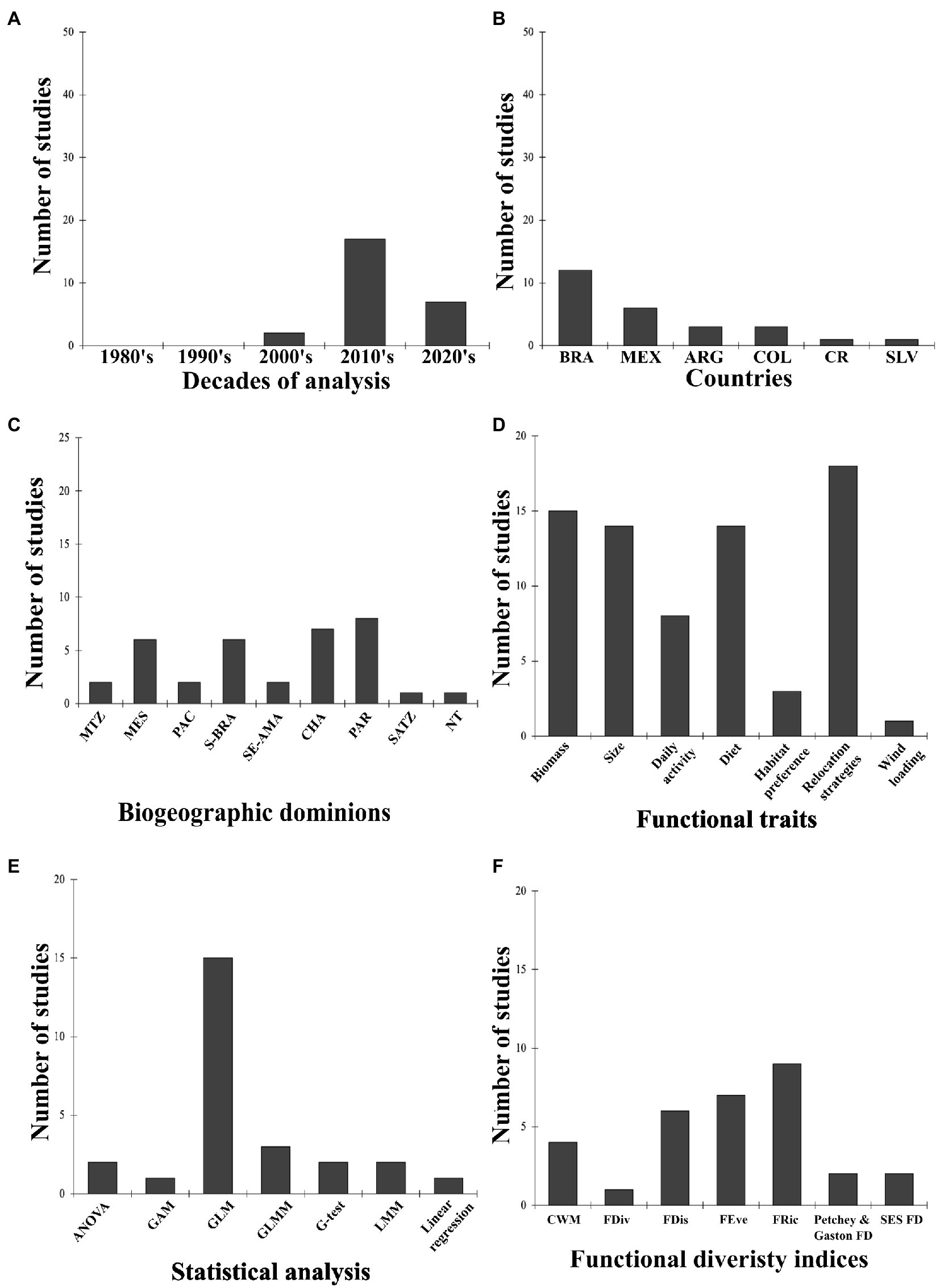
Figure 3. Number of functional diversity studies. (A) Over time (decades). (B) Per country. Brazil = BRA, Mexico = MEX, Colombia = COL, Costa Rica = CR, El Salvador = SLV, Argentina = ARG. (C) Per biogeographic dominion. Mexican Tranzition Zone = MTZ, Mesoamerican = MES, Pacific = PAC, Boreal Brazilian = B-BRA, SATZ = South American Transition Zone, South Eastern Amazonian = SE-AMA, Chacoan = CHA, Parana = PAR, Not tropical = NT. (D) Per functional trait. (E) Per statistical analysis. (F) Per functional diversity index.
Grazing lands were described in most articles on functional diversity as grassland (71.79%, n = 28), woodland (5.13%, n = 2), and agroforestry system (23.08%, n = 9). The proportion of each type of grassland, woodland, and agroforestry system in the reviewed articles are shown in Supplementary Table S3. Pastureland included exotic pastures and treeless pastures. The most common species of grasses are shown in Supplementary Table S4. Rangelands comprised native grassland of Andropogon spp. and Axonopus spp. without cattle presence, and grazed rangelands having the same species, but with cattle presence (Supplementary Table S3). The woodland systems belonged to South America grazing lands like Cerrado sensu stricto, and Grota (Supplementary Table S3). The principal mentioned agroforestry systems included silvopastoral systems (SPS) with leguminous fodder plants [Vachellia pennatula (Schltdl. & Cham.) Seigler & Ebinger, Leucaena leucocephala (Lam.) de Wit] and pastures; SPS with commercial species and native trees, SPS combining fruit species (Psidium guajava L.) and native trees (Quercus insignis M. Martens & Galeotti), and SPS with exotic species (e.g., Pinus spp.; Supplementary Table S3). Livestock management information is included in the reviewed studies about functional diversity, mainly about grazing, but including animal information, water aspects, and total surface (Supplementary Table S5). Grazing information contains animal rotation aspects, food surveys, plant successional information, grass type and management, description of the silvopastoral design and/or silvopastoral grass cover, pasture age, and grazing system management (Supplementary Table S5). Animal information references mainly cattle density, the total number of cattle, and livestock breed description (Supplementary Table S5). Information about water use is general (Supplementary Table S5). Approximately 19.23% of the reviewed manuscripts do not include livestock management information. Livestock practice data were scarce (68.42% of the studies without information), and the only mentioned practice was agrochemical use (Supplementary Table S5).
Most papers evaluated in this section used functional effect traits to perform functional analyses because of their relationship with the ecosystem functions and dung beetle performance in livestock systems (Supplementary Table S1B). Food relocation strategies were commonly used functions in most studies (n = 18, 69.23%), while wing loading and habitat preference were the least used (n = 1, 3.85%; Figure 3D). In addition, a trend in the selection of four functional traits was observed: biomass, body size, diet, and relocation strategies (Figure 3D). According to the authors, biomass and body size were selected because those traits are related to the amount of organic matter manipulated and buried in the soil. Relocation behavior determined spatial distribution of resources (vertically and horizontally), and diet preference inferred the type of resource that dung beetles manipulated.
The reviewed studies used various statistical methods to model the observed functional diversity with environmental or biological parameters. These ranged from one-way ANOVAs to general linear mixed models. However, over half of the studies used general linear models (n = 11, 57.70%), followed by general linear mixed models (n = 2, 11.5%; Figure 3E). The remaining analytical methods were represented by one study each. Eleven studies (61%) used the FD statistical package to analyze functional diversity, and four (22.22%) used the dbFD function. The other functions from FD and the Picante package were used only by one or two studies. Functional diversity was evaluated using two approaches: functional groups and indices. Most reviewed studies grouped the selected traits into functional groups, with counts and proportions of these traits the most used approaches. In the second approach, three indices were the most used: functional richness (n = 9, 29.03%), evenness (n = 7, 22.58%), and dispersion (n = 6, 19.35%), respectively (Figure 3F).
Twenty-four of the reviewed functional diversity studies (92%) employed a local spatial approach, and the remaining studies used a regional system (n = 2, 7.7%). It should be noted, that of those 24 local-based articles, eight (33%) considered more than three habitats in addition to the grassland environment (with which they tried to understand the role of grasslands in the landscapes studied), and 16 studies (67%) only considered grasslands, generally comparing them with fragments of preserved vegetation. The two studies with a regional spatial approach only make a comparison between grasslands and preserved forests. Finally, two works (7.7%) made temporal comparisons.
The total number of papers in our literature search that evaluated at least one function provided by dung beetles was 18 (Supplementary Table S1C). There has been an increasing trend in the number of studies dealing with grazing lands and functions in the dung beetle literature over the last five decades (Figure 4A). This result matches the general taxonomic and functional diversity sections. Before the 1990’s we found no papers working on this subject. The pick of studies on this subject was in the 2010s with 10 papers, and the number of articles will probably be higher at the end of the 2020s. In terms of the countries supporting these papers, we found studies for six countries, with Mexico (n = 8, 44.44%) and Brazil (n = 6, 33.33%) as the main contributors, and several other countries with only one study (Figure 4B). This pattern is slightly different from the trend we found with the other topics, with Brazil being the dominant country. We did not find any article from several countries in Latin America (e.g., Bolivia, Chile, Ecuador, Guatemala, Honduras, Nicaragua, Panama, Venezuela, and all the Antilles) regarding work on grazing lands with a functional perspective. Biogeographically, we found studies in the principal dominions of Latin America, showing that there is no notable dominance of any dominion (Figure 4C). Mesoamerican (n = 6), Mexican Transition Zone (MTZ; n = 5), South Brazilian (n = 3), and Chacoan (n = 3) were the most studied regions. No papers were registered for functional studies in the Boreal or Parana dominions.
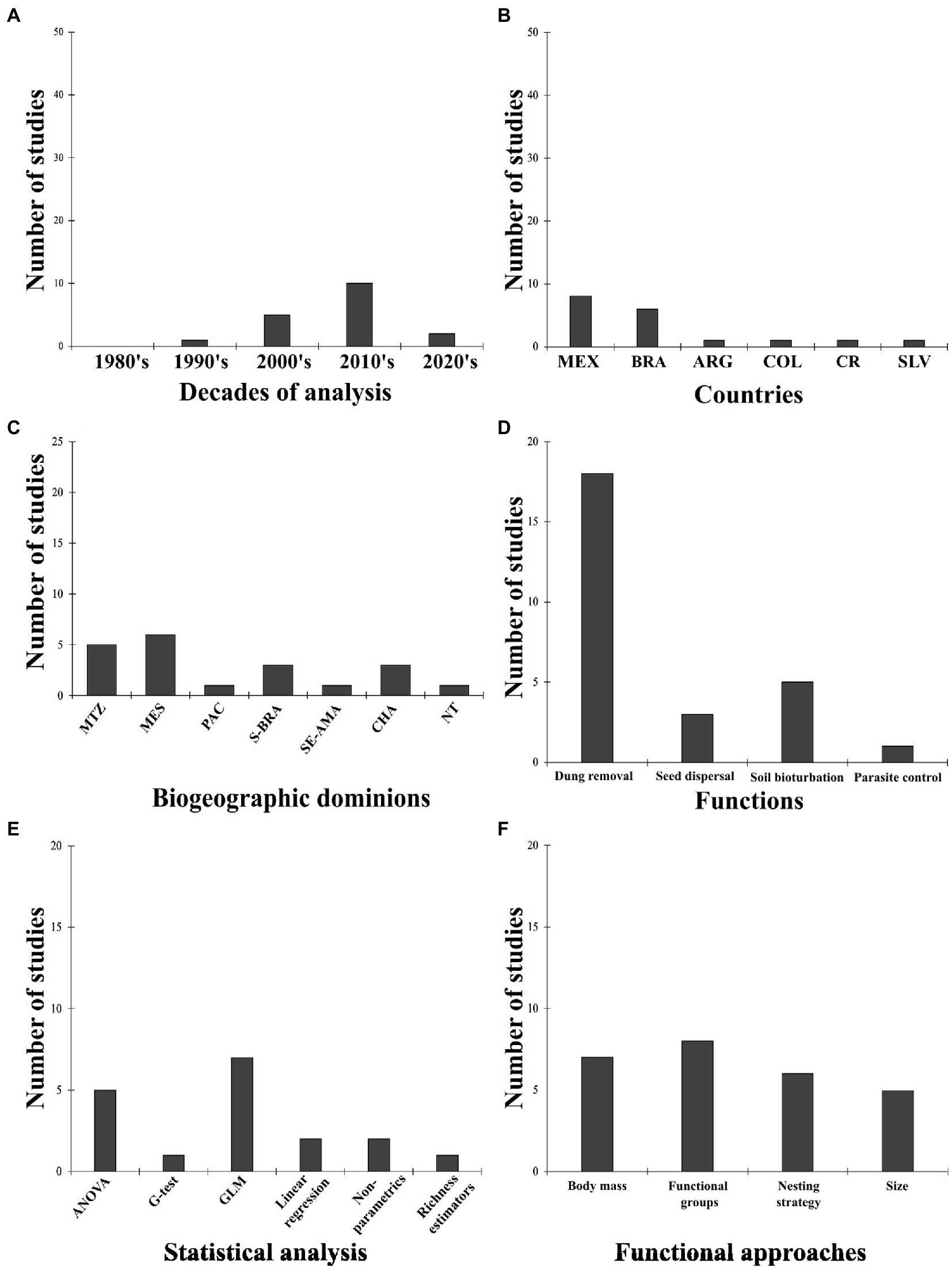
Figure 4. Number of ecological functions studies. (A) Over time (decades). (B) Per country. Brazil = BRA, Mexico = MEX, Colombia = COL, Costa Rica = CR, El Salvador = SLV, Argentina = ARG. (C) Per biogeographic dominion. Mexican Tranzition Zone = MTZ, Mesoamerican = MES, Pacific = PAC, Boreal Brazilian = B-BRA, SATZ = South American Transition Zone, South Eastern Amazonian = SE-AMA, Chacoan = CHA, Parana = PAR, Not tropical = NT. (D) Per function. (E) Per statistical analysis. (F) Per functional approach.
The trends observed in our review of articles on ecological functions and ecosystem services in grazing lands were like those we have found in our analysis of taxonomic and functional diversity. Most articles described grazing lands as grassland (76.92% of the 20 sampling sites), woodlands in only 3.85% (n = 1), and agroforestry systems described in 19.23% (n = 5). The proportions of each type of grassland, woodland, and agroforestry system in the reviewed articles are shown in Supplementary Table S4. Pasture lands included exotic pastures and treeless pastures. The most common species mentioned belonged to African grasses (Supplementary Table S4). The woodland systems belonged to South America grazing lands (Savanna; Supplementary Table S3). The principal mentioned agroforestry systems included a mix of pastures and banks of protein-rich legumes, such as Leucaena leucocephala (Lam.) de Wit (Fabaceae), and silvopastoral systems with commercial and native trees. Forest grazing included grazed primary and secondary vegetation and grazable forest land comprising small patches of grasses and ferns, predominantly covered by pine (Pinus oocarpa Schiede ex Schltdl.) with small numbers of Callitropsis lusitanica (Mill.) D.P. Little (formerly Cupressus lusitanica Mill.) and with little undergrowth vegetation (Supplementary Table S3). The information about livestock management in ecological functions and ecosystem services was similar to the authors’ topics (Supplementary Table S5). However, other elements are integrated into the management description, such as land ownership, type of livestock operation, and technological aspects (Supplementary Table S5). Approximately 22.22% of the revised manuscripts do not include livestock management aspects. In articles reviewed for ecological functions and ecosystem services, the information about livestock management practices was scarce; in 96.15% of the documents, there was no information. Only the use of veterinary medical products for cattle is mentioned in the few studies where information on livestock management practices is available (Supplementary Table S5).
Taking into consideration the wide range of functions that dung beetles provide in different ecosystems, in this literature review for pastures we only found studies evaluating four functions: (i) dung removal, (ii) seed dispersal, (iii) soil bioturbation, and (iv) parasite control (Figure 4D). The primary function was dung removal in 18 studies, followed by soil bioturbation (n = 5, 27.78%). Other functions like reducing greenhouse gas emissions, nutrient cycling, soil structure (e.g., aeration, permeability, porosity), pollination, or food supply, were not mentioned in any published papers in our literature review for grazing lands in Latin America. Thus, it seems easy to study and design experimental studies that quantify some of these functions instead of others. Regarding the statistical analyses used in the studies, we found that the most common methods were GLM, ANOVA, linear regressions, and non-parametric approaches (Figure 4E). Finally, in terms of function, we found four main variables: body mass, functional groups, nesting strategy, and size (Figure 4F). The most used were functional groups (n = 8, 44.44%), followed by body mass (n = 7, 38.89%). The conceptual separation between these approaches is not clear enough because some studies use size, body mass, or nesting strategy to build different functional groups.
The most common goal was evaluating the effects of “land use changes.” Indeed, land-use change has been one of the major causes of natural ecosystem transformation, causing global biodiversity declines (Newbold et al., 2015). Currently, livestock farming is the most significant land-use sector on Earth, occupying more than 30% of the planet’s continental surface as grazing lands (FAO, 2017). Therefore, it is understandable that dung beetle researchers have driven their studies to understand the impacts of the conversion of natural vegetation to grazing lands, because that theme is urgent to preserve biodiversity and associated ecosystem functions and services.
Among studies, we found two approaches explaining the changes in species diversity in grazing lands. First, in the articles conducted in tropical rain forest is observed a high number of total and exclusive species, and they become locally extinct with the loss of trees or canopy cover in humid tropical landscapes. A loss of species is observed as vegetation fragments become smaller due to changes in land use and increased distance among the remnants. These small fragments serve as refuges for some forest species (Favila, 2005), and the diversity depends on nearby species pools, possibly primary forests, and host a greater spatial heterogeneity in species composition. Forest cover is the best predictor of dung beetle assemblages, positively related to species diversity and biomass across multiple spatial scales (Alvarado et al., 2018). Yet, landscape homogenization resulting from increased extension of grazing lands leads to changes in resource food selection related to the preference in the attraction of dung beetles to exotic omnivores and livestock (Alvarado et al., 2021).
In articles conducted more frequently in fragmented landscapes of tropical dry forest we found a the second approach: the authors found there has been no net reduction in regional species richness, although local species richness in natural ecosystems has declined (Halffter and Arellano, 2002). A change in species composition is observed as grassland species invade vegetation fragments. However, forest remnants, wooded systems, living fences, and silvopastoral systems generate new assemblages (species with different habitat requirements) within species-rich landscapes with greater connectivity (Reyes-Novelo et al., 2007; Arellano et al., 2008, 2013; de Farias et al., 2015); therefore, changes in species composition and species turnover becomes important. There are clear signs that tree structure and microclimatic conditions like forests, as found in agroforestry, can help preserve biodiversity by creating a propitious habitat for native species (Righi et al., 2018). Silvopastoral systems can buffer the adverse effects of rapid expansion of open areas and the consequent reduction of tropical dry forest area generated by conventional technified systems (Arellano et al., 2013). Although richness remains relatively constant in landscapes with intermediate degrees of disturbance (such as those that have been partially modified for human use), richness at the local level changes notably over short lapses of time, and the assemblage composition is very fluid (Halffter et al., 2007).
Finally, because dung beetles were proposed as indicator groups in taxonomic diversity studies, the integration of functional groups such as habitat preferences (Favila, 2005; Díaz et al., 2010; Bourg et al., 2016), daily activity (Navarrete and Halffter, 2008); relocation strategies (Escobar and Chacón de Ulloa, 2000; Horgan, 2008; Basto-Estrella et al., 2012; Noriega et al., 2012; Gómez-Cifuentes et al., 2019; Correa et al., 2019a, 2020a; Cajaiba et al., 2020; Salomão et al., 2020); body size (Escobar and Chacón de Ulloa, 2000; Halffter and Arellano, 2002; Navarrete and Halffter, 2008; Korasaki et al., 2013), food preferences (Favila, 2005; da Silva et al., 2008; Horgan, 2008; da Silva and Audino, 2011; Correa et al., 2013; Cajaiba et al., 2020) and their representativeness under different conditions or land uses continue to be considered, as well as the use of biomass as an essential value (e.g., Noriega et al., 2012; Costa et al., 2017; Alvarado et al., 2021), forming the basis for subsequent studies of functional diversity with precise and statistically more robust methods.
Functional diversity has been evaluated by its importance to ecology and biological conservation studies since it is a component of biodiversity that contributes to the understanding of the structure and functioning of ecosystems, including information on the identity of species through the description of their functional traits (Moore, 2001; Tilman, 2001). A first approach to functional diversity is the formation of functional groups, which are defined as a set of species that have a similar life history, such that they use a resource similarly in a given space and time (Moore, 2001; Steneck, 2001; Tilman, 2001), or that are related to some activity in ecosystems (Naeem et al., 2009). Species with similar functional traits are commonly assigned to functional groups (Moore, 2001; Steneck, 2001; Tilman, 2001). In Neotropic grazing lands, we found that in the reviewed articles, both analyses of functional groups and functional traits are carried out. However, most studies analyze each trait independently because counts and proportions are the most commonly used methods. Also, with this approach, it is necessary to consider the use of relevant terminology regarding dung beetle nesting behavior since it is one of the most widely used functional traits for categorizing functional groups (Tonelli, 2021).
In general, the studies reviewed on functional diversity showed how different traits, both ecological and morphological, have been considered over time (e.g., Halffter et al., 2007; Barretto et al., 2020; Davies et al., 2020, 2021; Souza et al., 2020; Correa et al., 2020b, 2021b; Whitworth et al., 2021; Guerra-Alonso et al., 2022). Beetle size (large, medium, and small species) has been the most frequently used qualitative functional trait (Barretto et al., 2020; Davies et al., 2020, 2021; Noriega et al., 2021b), although they also highlight the feeding pattern of relocation (telecoprid-rollers, paracoprid-tunnelers, and endocoprid-dwellers), daily activity (nocturnal, diurnal, and continuous or mixed activity), and diet (coprophagous, necrophagous, and generalist species). Biomass (dry weight) has been quantitatively estimated as an important trait (i.e., Correa et al., 2018; Gómez-Cifuentes et al., 2019, 2020), although it has also been used frequently in taxonomic diversity analyses, as a surrogate for abundance. A work that stands out in this review is Guerra-Alonso et al. (2022) because the authors measured different morphological response traits of dung beetles: (i) body area, (ii) biomass, (iii) total length, (iv) sphericity, (v) area of the head, (vi) width of the pronotum, (vii) length of the anterior tibia, (viii) area of the anterior tibia, (ix) area of the anterior femur, and (x) tooth width. Wing loading and color pattern (metallic, uniform, and patterned colors) also have recently been used as indicators of change in functional diversity (Whitworth et al., 2021).
Of the total number of articles reviewed, seven articles (27%) analyzed functional diversity as a complement to the taxonomic approach, applying these two methodological approaches to understand how grasslands modulate the function and diversity of beetles in the Neotropics (Correa et al., 2018, 2019b, 2020b, 2021a,b; Gómez-Cifuentes et al., 2019; Cajaiba et al., 2020). Likewise, we were able to visualize key conclusions revealing the context of the landscape and type of management needed to generate alterations in the role of beetles, for example: (i) the most disturbed ecosystems are related to significant reductions in functional redundancy, which can have detrimental effects on the future resilience of the landscape (Cajaiba et al., 2020); (ii) for cattle grazing rotation to be effective, in introduced Brazilian pastures it has been seen that an extended period of time (at least 1 month) is required for cattle removal; this strategy can be helpful to conserve the diversity of grasses, dung beetles and, consequently, ecological functions (Correa et al., 2021a); and (iii) some authors such as Carvalho et al. (2021) have indicated that the responses of dung beetle assemblages and their ecological functions to subtle changes within a type of land cover are uncoupled, idiosyncratic and depend on the context, making it difficult to make predictions and generalizations in grasslands. Likewise, Guerra-Alonso et al. (2022) have shown that native forests and forests with cattle maintain functional diversity in all regions. In the case of open pastures, the authors argue that the answer depends on the regional context because the substitution of the native forest for open pastures strongly affects functional diversity.
The low number of papers (n = 18) that include and evaluate at least one function provided by dung beetles in grazing lands, compared to the number of articles working with taxonomic (n = 76) or functional diversity (n = 26) in our review, is troubling. Even though there has been an increasing trend over the last five decades, it is indeed a new topic regarding grazing lands (e.g., Ortega-Martínez et al., 2016; Alvarado et al., 2019; Carvalho et al., 2021). There is a clear thematic dominance in terms of the countries that carry out this type of study (i.e., Mexico - Basto-Estrella et al., 2016; Ortega-Martínez et al., 2016; Huerta et al., 2018; Alvarado et al., 2019 and Brazil - Braga et al., 2013; Correa et al., 2019a; Carvalho et al., 2021). Most Latin American countries lack studies on ecosystem functions in grazing systems (see section 3.3). As well, there are few studies that analyze and study functions at the level of a single species (e.g., Miranda et al., 2000; Anduaga and Huerta, 2007; Martínez et al., 2018), and few studies that use mesocosms (e.g., Anduaga, 2004; Ortega-Martínez et al., 2016; Alvarado et al., 2019), or that carry out studies under laboratory conditions (e.g., Horgan, 2001; Mariategui et al., 2001; Ortega-Martínez et al., 2016), aspects to be explored in future studies.
Regarding ecosystem functions, there was a tendency to evaluate dung removal as the main activity of dung beetles (e.g., Miranda et al., 2000; Mariategui et al., 2001; Anduaga, 2004; Cruz et al., 2012; Alvarado et al., 2019). The main reason for this trend is the importance of this function as the basis for many ecosystem services and the methodological ease of evaluating it. However, it is important to mention that when comparing different studies, some experimental variations in the use of controls and units would need to be verified and standardized. The other ecosystem functions we reported were seed dispersal, soil bioturbation, and parasite control (see Giraldo et al., 2011; Braga et al., 2013; Martínez et al., 2018; Correa et al., 2019a; Carvalho et al., 2020, 2021). Other essential functions like greenhouse gas emissions control, nutrient cycling, soil aeration, permeability, porosity, pollination, or food supply (Nichols et al., 2008; Slade et al., 2016) were not registered in this review. It is possible that some services, such as seed dispersal, are more relevant in forest systems than in grasslands (e.g., Andresen, 2002), which would explain the absence of studies on this function. Likewise, the study of some services that may be important in livestock systems, such as the control of greenhouse gases, may be limited by the technical requirements and costs of this type of evaluation (Slade et al., 2016). This should undoubtedly be a priority for future studies in this type of system in the region.
Another critical aspect that needs to be standardized are the variables used in functional approaches. We found four main variables: (i) body mass (e.g., Anduaga, 2004; Braga et al., 2013; Alvarado et al., 2019), (ii) functional groups (e.g., Basto-Estrella et al., 2016; Correa et al., 2019b; Carvalho et al., 2021), (iii) nesting strategy (e.g., Braga et al., 2013; Alvarado et al., 2019; Correa et al., 2019b), and (iv) size (e.g., Mariategui et al., 2001; Anduaga, 2004; Carvalho et al., 2021). Most studies mix these concepts indistinctly, especially in the case of biomass, which is almost always assumed by utilizing the weight of individuals as a proxy. In many of these papers, the conceptual separation between these approaches, the way they are measured, or the categorization is not clear enough. The construction of functional groups using the combination between food resource relocation guilds (i.e., paracoprids, telecoprids, endocoprids, and kleptocoprids) and individual size (i.e., small, medium, large) seems to be a helpful strategy (see Noriega et al., 2021b). However, the construction and categorization of functional groups require an in-depth theoretical review and experimental studies that allow us to understand their separation at an operational level.
As well, we registered for taxonomic and functional diversity, a lack of information on livestock systems, type of management, use of anthelmintics, characteristics of pastures and soils, cattle species, grass species, and climatic conditions. The absence of this information in most of the papers is not only a severe methodological problem that constrains replicability, but also prevents large-scale comparisons or meta-analyses. This is one of the main reasons we propose a standard data collection format (Supplementary Box I) for work in these environments.
In articles regarding the effects of grazing lands on dung beetle diversity, functions, and ecosystem services, we must start by recognizing that there is no clear standardization or unification of the criteria for “grazing lands.” Over time, the approach and concept of pastures and the effect of the transformation of natural ecosystems to treeless and/or grazing lands on the taxonomic and functional diversity of dung beetles and their ecosystem functions and services have been evolving and transforming. Grazing land studies have long been focused on the presence/absence of tree cover (decreasing species diversity from forest to pastures: e.g., Navarrete and Halffter, 2008; da Silva and Hernández, 2014, 2016; Bourg et al., 2016; Silva et al., 2017; Salomão et al., 2020), and in most reviewed articles (using taxonomic and functional diversity and functions) they were described as open areas dominated by grasses and herbs (e.g., Halffter et al., 1992; Horgan, 2008; Navarrete and Halffter, 2008; Bourg et al., 2016; Salomão et al., 2020).
Currently, there are multiple studies describing declining species diversity over vegetation gradients (e.g., Halffter et al., 1992; Escobar and Chacón de Ulloa, 2000; Andresen, 2008; Almeida de and Louzada, 2009; Cajaiba et al., 2020; Rivera et al., 2020; Alvarado et al., 2021), or management gradients (e.g., Arellano et al., 2013; Korasaki et al., 2013; Montoya-Molina et al., 2016; Costa et al., 2017; de Farias and Hernández, 2017; Righi et al., 2018). Recent approximations compare species diversity between grazed rangelands and pasturelands (exotic pastures; e.g., Almeida et al., 2011; Correa et al., 2016a,b; Macedo et al., 2020). As the description of grazing lands becomes more precise, a better understanding of these spaces is permitted.
Approaches have been changed gradually, and other aspects have begun to be analyzed, such as the effect of boundaries between natural ecosystems and pastures (e.g., Silva et al., 2017; Martínez-Falcón et al., 2018), the relative importance of the presence of exotic food (cattle excrement), the loss of cover in managed secondary vegetation (Halffter and Arellano, 2002) and the spatial distribution of pastures in the landscape, their adjacent areas, and boundaries with other land uses (Arellano et al., 2008), or the species within pastures present in the areas. Since the 2010s, silvopastoral systems and managed secondary vegetation and their benefits for biodiversity conservation have begun to be incorporated into grazing lands. The variety, or the species of grass (e.g., Abot et al., 2012; Flota-Bañuelos et al., 2012; Correa et al., 2013, 2016a; Righi et al., 2018), the composition and density of tree species in pastures, the surrounding matrix, and local knowledge are being mentioned in dung beetle studies. Meanwhile, silvopastoral systems will increase in the coming decades due to the growing global population, with an estimated 30–70% increase in demand for timber, cellulose, oils, and food (Solorio et al., 2017). Further studies are needed to clearly understand how different silvopastoral systems (with different plant density, composition, and management) may affect dung beetle biodiversity in the Neotropics. There is little information regarding the impact of silvopastoral systems on Neotropical dung beetles (Colombia, see Montoya-Molina et al., 2016; Argentina, see Gómez-Cifuentes et al., 2019; Mexico, see Arellano et al., 2013, de Farias et al., 2015).
We need to understand the effect of vegetation structure of grazing lands (Agrosilvopastoral systems: ASPS, Voisin silvopastoral and pastoral grazing, biodiverse grasslands, monoculture pastures without trees) on Neotropical dung beetle assemblages. ASPS is a viable strategy for extensive livestock farming based on the principles of agroecology, agroforestry, and animal production. The objective of these systems is to optimize the positive interactions between agriculture, forestry, livestock, and the physical environment, and to maximize land productivity through spatial or sequential spatial arrangements or in temporal sequences (Murgueitio and Solorio, 2008; Nahed-Toral et al., 2013). However, afforestation often involves the creation of fast-growing tree plantations or SASP on non-forest lands. What will be the possible impacts of afforestation on the biodiversity of local species (e.g., Ueda et al., 2015)? Although grazing lands represent a large area in most terrestrial landscapes, management decisions within these systems that can affect the conservation of the dung beetle biodiversity are still poorly understood (Correa et al., 2021a).
Currently, livestock is the most prominent land-use sector on Earth, occupying more than 30% of the global continental land area (FAO, 2017). To understand the impact of livestock activities on dung beetle diversity over the last 20 years, analyses are increasingly investigating livestock practices and management. Some past studies have assessed the effects of shaded fields (Horgan, 2002, 2005), site complexity, soil quality, and management (de Farias et al., 2015); ivermectin use (Flota-Bañuelos et al., 2012; Tovar et al., 2016), fire use (Rangel-Acosta et al., 2020); cattle grazing abandonment time (Correa et al., 2019a, 2020b), animal consortium (e.g., horse and cattle, Louzada and Carvalho e Silva, 2009; sheep and cattle; Correa et al., 2020c), rotational pasture management (Correa et al., 2021a) and livestock intensification (Alvarado et al., 2018).
Research interest is growing in the Neotropics regarding the effect of agrochemicals, mainly anthelmintics (doramectin, eprinomectin, ivermectin, or moxidectin) and herbicides on dung beetles (Souza et al., 2018; Villada-Bedoya et al., 2019; Carvalho et al., 2020; Correa et al., 2022). The spatial and temporal impact of the macrocyclic lactone parasiticide treatments, which result in insecticide residues in cattle feces, will depend on a range of farm management factors, such as the frequency of anthelmintic use, the number of animals treated, and the choice of active ingredient, as well as a range of insect-related factors, such as abundance, population dynamics, and dispersal rates. More studies are needed to understand dung beetle assemblage responses to impacts from livestock activities on taxonomic and functional diversity, and in their ecological functions in the Neotropics. Equally important is how herd management, grazing intensity, area size, grazing time, and local history contribute to understanding how dung beetle species respond to pasture management in grazing lands and how this can help cattle farmers in decision-making to improve conservation strategies on their properties.
In the Neotropics, we also find differences in cattle raising protocols and their effects among countries and biogeographic regions. The greatest differences are found in the Pantanal, where the size of the herds is much larger than in other Neotropical areas, with no inputs used, and the animals are moved so that the land is left to rest for a sufficient time. The problems encountered have more to do with the introduction of exotic grasses. In the Brazilian Pantanal, the vast natural grassland plains, allied with a favorable climate, promoted extensive cattle ranching in this ecosystem in often pervasive areas (10,000 ha; Seidl et al., 2001; Eaton et al., 2011). Thus, in many Pantanal sites using cattle, there are private lands with livestock histories of at least 100 years using native grasses resistant to the seasonal flooding that occurs in this biome. These practices occur without intensive management (no use of fertilizers, herbicides, and veterinary drugs in cattle), with stocking rates between 0.5 and 1.0 animal unit ha−1. Therefore, cattle breeding in natural grasslands of the Brazilian Pantanal can integrate livestock production with the conservation of dung beetles and their ecological functions (Correa et al., 2019a).
In Mesoamerica and other regions of South America, such as Mexico and Colombia, areas previously occupied by tropical forests were deforested, and monoculture pastures were implemented, which were gradually dominated by introduced African grasses. In the Mexican High Plateau, where extensive cattle ranching is common, grazing lands include open areas for crops, semi-open areas for cattle, and closed areas with native shrubby vegetation, which during the dry season are used for cattle foraging. Those grazing practices have transformed the natural arid environment into landscapes dominated by semi-open grazing areas, maintaining several species of domestic livestock associated with the original vegetation (Mellink and Riojas-López, 2020). The responses of dung beetle assemblages are modulated by the ecological conditions resulting from the transformation of native vegetation into grazing areas and the history of the species inhabiting different biogeographical provinces. The impact of grazing on dung beetle diversity is dependent to a considerable degree on the local ecological conditions and the biogeographical context that has shaped the composition of assemblages over time (Barragán et al., 2014).
Agricultural production systems are dynamic and complex because they have non-linear properties and experience imbalances over time by new interactions among their unique components. Therefore, the current status and response potential of livestock systems results from a complex historical process of interactions between physical, biological, and socioeconomic factors, adaptive tensions between society and nature, and production and ecosystem services. The changes in local grazing land conditions are driven by human management practices (Hutton and Giller, 2003; Alvarado et al., 2019; Gómez-Cifuentes et al., 2022). The knowledge of the historical trajectory of livestock farming in the Neotropics facilitates identification of the essential stages of the process to recognize the causes of biodiversity loss (Rodríguez-Moreno et al., 2020). Livestock management history in the Neotropics is a theme touched upon by some, not as a variable for analysis, but as a theme to involve the reader in the context of land use changes. How livestock management affects dung beetles in Neotropical grazing lands remains to be investigated in different biogeographical and social contexts.
As presented, a high proportion of reports on changes in dung beetle species diversity come from Brazil, Mexico, and Colombia. The latest research trends reported by UNESCO (2021), show that Brazil, Mexico, and Argentina have the highest rates of public investment in overall research and scientific production, as well as Ph.D. students in Latin America. Moreover, such productivity should also be reflected in environmental research with dung beetles. However, lack of investment creates a critical information gap, resulting in poor understanding of each country’s identity, diversity, ecology, and species distribution. In addition, considering the publication trend over the last three decades, a geographical transition of publications from Mexico to Brazil can be seen due to a generational change in researchers.
This review shows that the number of articles on the temporal changes in species diversity has been declining (e.g., Reyes-Novelo et al., 2007; Silva et al., 2010; Lopes et al., 2011; Flota-Bañuelos et al., 2012; Correa et al., 2016a, 2018, 2021b; Salomão et al., 2020). Although it is important to know monthly or seasonal variations in species diversity, long-term studies on the impact of land management on dung beetle assemblage dynamics are needed to understand the processes involved and suggest better management strategies. Overall, we found a lack of studies that include extensive time windows (more than 1 year of sampling) or analyses at broad spatial scales (landscape or between countries or regions). This produces an explicit spatio-temporal limitation of the patterns described and makes the results extremely local and punctual. Absence of this form of study is due to the non-existence of projects and funding sources covering more than 1 year, and the reduced interaction between scientists throughout Latin America. In this sense, it is necessary to expand these windows of time and space to compare studies among years and decades, as well as among countries and biogeographical regions, and create a more robust network of researchers in Latin America.
As we mentioned, there is a growing interest among researchers in the Neotropics regarding the effect of agrochemicals on dung beetle species. Most studies have been carried out using small, short-cycle species [e.g., Aphodiinae, Euoniticellus intermedius (Reiche)], which perform efficiently in the laboratory. However, it also is necessary to know the effect of these agrochemicals on large burrowing species (e.g., Iwasa et al., 2007), which are more sensitive to land use and management changes. Large bodies are often associated with lower fecundity and longer generation times (Halffter and Edmonds, 1982; Chown and Gaston, 2010), so a reduction in the ability of populations to compensate for increased mortality under anthropogenic pressures would be expected (Chown and Klok, 2011; Nichols et al., 2013). Species’ abundances or the occurrence of specific sensitive species should be investigated concerning habitat and landscape factors before management and conservation plans of semi-natural pastures are made (Söderström et al., 2001). Performing more studies on the effect of herbicides, insecticides (e.g., Kryger et al., 2006; Sands and Wall, 2018), hormones, and vaccines on dung beetles would be highly beneficial. Thus, many field studies are currently being initiated (Villada-Bedoya et al., 2019).
Grazing management strategies are not carried out exclusively in space or time, so we must analyze the effect of grazing synergistically with other management practices (fire, agrochemical use, artificial fertilization, soil management practice, tillage, type of livestock) on dung beetle assemblages. This will facilitate the design of conservation and management strategies that favor the diversity of dung beetle species. Also, the importance of landscape composition for mobile organisms entails that management activities should focus on broader scales. Finally, studies evaluating dung beetle economic contributions to cattle production in tropical grazing lands are required (see Lopez-Collado et al., 2017). Information on the monetary value of dung beetle services may stimulate and encourage farmers to develop management plans for livestock production to conserve dung beetle diversity and their ecological benefits.
According to our review, the impact of animal livestock grazing and management on the environment has been of increasing interest over time, although the approach in each paper provides contrasts between the Old World and the New World due to the different contexts and histories of animal livestock maintenance. Livestock farming is an ancient human activity, which began with the domestication of cattle (sheep, goats) in the Neolithic (e.g., hieroglyphics in the Middle East, in Egypt dating from 4,500 years BC). In Europe, there are traditional management systems such as grazing of small herds of small livestock and the “dehesa” ecosystem, which, according to the legal context of the European Union (Habitat Directive), are considered Cultural Landscapes or semi-natural ecosystems and serve as model ecosystems in ecological restoration (Gann et al., 2019). The perpetuation of this type of landscape (since prehistoric times) is an example showing anthropic intervention in ecosystems can be sustainable, ensuring productive benefits while conserving biodiversity and climate regulation (Ferraz-de-Oliveira et al., 2016; Garrido et al., 2017).
To understand the reasons for the grazing and management practices used according to a particular context and management history also is important in order to suggest possible management strategies (e.g., Europe: Bokdam and Gleichman, 2000; Verdú et al., 2000; Lobo et al., 2006; Macagno and Palestrini, 2009; Rosa-García et al., 2013; Tocco et al., 2013). For example, we must consider that the tree species used in livestock systems, management practices, and the objectives in silvopastoral systems (SPS) are not the same. In Central America and Mexico, the tree species most used in SPS are forage and fruit trees, and in South America, timber species. The impact of the use of introduced grasses in pastures concerning native grasses is very different in Mesoamerica and South America.
Within agricultural management regimes, management history is important because of its implications for soil conditions, seed deposition and seedling propagation, existing diversity, and ecosystem resilience. Yet, in the articles we reviewed, less than 50% of the studies considered taxonomic diversity (46%), functional diversity (46.15%), or ecological functions (38.88%), nor included the history of land use (only as a descriptive explanation in the methods section or the introduction) to facilitate the understanding of the context in which the work takes place, mainly regarding the changes in land use over time, or the age of establishment of land uses. In the remaining studies, there is no information about land use history. This element is often not considered a variable in diversity analyses.
Dung removal and burial is a crucial ecological function of dung beetles because it generates ecosystem services such as soil improvement, pasture cleaning and increased grazing area, control of cattle flies, greenhouse gas reduction, and secondary seed dispersal (Nichols et al., 2008). There have been multiple studies related to soil improvement, seed dispersal, and the identification of dung beetle species efficient in the control of cattle flies. However, there are no studies in the Neotropics related to the activity of Neotropical beetles and the emission of greenhouse gasses or fecal helminth transmission. More studies are needed that involve multiple functions and ecosystem services in grazing lands to have a more comprehensive assessment of the individual functional contributions of particular species and/or the mixture of species that drive ecosystem functioning (Manning et al., 2016; Slade et al., 2017; Piccini et al., 2018). We must identify changes in the function of manure burial and removal in response to traces of agrochemicals (e.g., Manning et al., 2017), hormones, vitamins, and other substances used in livestock management of manure, which change its quality. More economic valuations at local or regional scales on the ecosystem services of dung beetles are needed to communicate the importance of this group of insects and their roles to decision-makers. In terms of laboratory studies with one or several species or experimental designs in the field, we also found very few works showing alternative empty niches of possible analyses in livestock systems where physiological, behavioral, and ecological aspects of the assemblage could be evaluated.
Studies using functional trait approaches increase the understanding of ecological processes and inform conservation and restoration of ecosystems since traits are morphological, biochemical, physiological, structural, phenological, or behavioral characteristics that influence species’ fitness (Nock et al., 2016). In future work, it is essential to include a variety of traits that represent species ecology and physiology to have a complete overview of functional diversity and the consequences of human disturbance on ecosystem functioning (Giménez-Gómez et al., 2022). In addition, it is necessary to assess the relationship between dung beetle traits and ecosystem functions delivered by them (deCastro-Arrazola et al., 2020), and distinguish dung beetle trait clusters that reflect assemblage adaptations to land use changes since they may better allow for generalizations of adaptive responses in ecosystems (Bui et al., 2020).
Most studies on dung beetle ecology are related to individual anthropogenic drivers of decline in richness, abundance, and biomass, changes in composition, increases of generalist species, and homogenization of assemblages. Further studies are required to analyze the synergic effects of the identified drivers (fragmentation, deforestation, agricultural management, defaunation, hunting, fire, invasive species, urbanization, global environmental change). Especially in Neotropical livestock systems, the presence of the invasive species Digitonthophagus gazella (Fabricius; see Noriega et al., 2020) can have a negative effect on the structure, diversity, and function of assemblages, which has not been experimentally quantified in the field.
There is a need for Neotropical research, mainly in population dynamics and natural history of dung beetle species and in monitoring any recent declines in population size or geographical range of some species in response to livestock activities. Comparisons between past and present population and range sizes over long periods are few. Most of these studies are carried out in Europe, Asia, and Africa. Adding information on geographic areas of dung beetles, where there are often significant knowledge gaps, contributes to the international conservation of species by deepening our understanding of their distribution, spatial niches, and phylogeographic barriers. The dung beetle species most vulnerable to isolation and local extinction are those with naturally low population levels, restricted niches, high biomass, and are forest specialists, which may require forest tracts greater than 200 ha to maintain the evolutionary variability of their communities (Larsen et al., 2008; Rivera et al., 2022). The main threat affecting these species is the destruction of their natural habitats due to anthropogenic activities. Many forest-dependent species, such as those native to the Atlantic Forest of Brazil, are at high risk due to accelerated deforestation resulting from expanding agricultural and livestock activities (Rezende et al., 2018). Species depending on highly specialized habitats, such as the burrows of small mammals (e.g., rats, gophers), caves, snail droppings, or species that have monetary value because of their size or beauty should be considered candidates for conservation and/or protection using legal coverage. For some Red List species, their scarcity may not signal danger, as they have not been assessed adequately (low collection effort, insufficient biological study), which is very likely to occur for some dung beetle species.
In the other hand, using a meta-analytical approach with dung beetles, Rivera et al. (2023) evaluated how anthropogenic habitat disturbances influence taxonomic, functional, and phylogenetic diversity to conserved forest sites in the Neotropics, and they found that heavy disturbances erode and homogenized all diversity dimensions of dung beetles with close dependence on forest habitats species. Moreover, they promote the protection of disturbed off-reserve forests (e.g., second-growth forests and agroforestry systems) in management schemes, since favoring the coexistence between functional and phylogenetically distant species and maintaining assemblages similar to those in conserved forests.
Therefore, safeguarding ecosystem functions and services that insects perform in the Neotropics required to add efforts in new public policies in protected areas including Indigenous and Community Ecological Reserves as well as species-specific action policies to prevent further declines and ensure their continuity in the ecosystems. In addition, it is required to potentiate the study of insect distribution modeling to identify areas of priority conservation habitat (Duffus et al., 2023), considering an integral vision of the study of biodiversity (i.e., taxonomic, and functional diversity) in the tropics (Moreno et al., 2018); since global warming is a growing threat, and its synergies are potentially far-reaching with other causes of anthropogenic origin (Laurance et al., 2011).
Most of the studies included in this analysis more commonly comprise the Scarabaeinae subfamily, with few investigations incorporating the subfamily Aphodiinae. If this subfamily is included, their taxonomic identification is often deficient. This shows a lack of taxonomic tools (i.e., keys and academic proficiency) to identify this group, but at the same time, a lack of ecological, physiological, and behavioral information on this group, which is extremely relevant and essential in grassland areas. This should be a priority for future studies in this region.
We recommend following a framework trait-based, since it is recognized the multi-functionality of traits of the dung beetle, considering characteristics of their morphology, feeding, reproduction, physiology, activity, and movement which involve traits with response to the environment and affect ecosystem processes in different spatial, temporal, and biological scales (for more details deCastro-Arrazola et al., 2023). Also, is therefore required to address the trophic complexity of ecosystems, since traits also influence interactions within trophic levels. For example, via competitive interactions between dung beetle species with similar ecological niches (Schleuning et al., 2023).
Forest cover is the best predictor of dung beetle assemblages, being positively related to species diversity and biomass across multiple spatial scales (Alvarado et al., 2018). Landscape homogenization resulting from increases in the extension of open grasslands reduces species diversity and composition. The most disturbed ecosystems are related to significant reductions in functional redundancy, which can have detrimental effects on the future resilience of a landscape (Cajaiba et al., 2020). In the case of open pastures, the answer depends on the regional context because the substitution of native forest for open pastures strongly affects functional diversity (Guerra-Alonso et al., 2022).
Native forests and forests with cattle maintain functional diversity in all regions. Biodiverse grazing lands with tree species are an option for not reducing regional species richness in fragmented landscapes. There are clear signs that tree structure and microclimatic conditions provided by forests, as found in agroforestry, can help preserve biodiversity by creating productive habitats for native species (Righi et al., 2018). Silvopastoral systems can buffer the adverse effects of rapid expansion of open areas and the consequent reduction of tropical dry forest area generated by technified conventional systems (Arellano et al., 2013). Forest remnants, wooded systems, living fences, and silvopastoral systems favor new assemblages (species with different habitat requirements) within species-rich landscapes with greater connectivity (Reyes-Novelo et al., 2007; Arellano et al., 2008, 2013; de Farias et al., 2015).
Species with similar functional traits are commonly assigned to functional groups (Moore, 2001; Steneck, 2001; Tilman, 2001). However, the construction and categorization of functional groups requires an in-depth theoretical review and experimental studies that permit a better understanding of their separation at an operational level. As well, the variables used in functional approaches must be standardized.
Most Latin American countries lack studies on ecosystem functions in grazing systems. In our review, there are few papers that include and evaluate at least one function provided by dung beetles in grazing lands (mainly dung removal) compared to the number of articles working with taxonomic or functional diversity. The responses of dung beetle assemblages and their ecological functions to subtle changes within a type of land cover are uncoupled, idiosyncratic and depend on the context, making it difficult to make predictions and generalizations in grasslands (Carvalho et al., 2021). Considering the importance of the data to be included in future work, we propose a collection format to unify the information collected in the field when conducting ecological studies in grazing lands. We recorded a lack of information on livestock systems, type of management, use of anthelmintics, characteristics of pastures and soils, cattle species, grass species, and climatic conditions. The absence of this information in most of the papers is not only a severe methodological problem that constrains replicability, but also prevents large-scale comparisons or meta-analyses. This is one of the main reasons we propose a standard data collection format (Supplementary Box I) for work in these environments. Moreover, we define a classification system to homogenize the features that distinguish the multiple tropical grazing lands found in the literature. Preserving dung beetle diversity and associated ecological functions is urgent for maintaining ecosystem services in grazing lands.
LA, JN, and IO-M contributed to the conception and approach of the text and coordinated the progress of each of the main sections of the document. JN, JR, CC, AG-C, FB, and AR-H were responsible for the design of the final figures and methods. All authors participated with content of the manuscript, review of papers, synthesis of ideas, results, Supplementary material, and general redaction and writing of the document.
Some of the ideas in our article, mainly those referring to the classification of grazing systems came from the 80-132 TAMU-Conacyt project, but the project does not finance publication of results. We leave it to your consideration whether to leave it or replace it with None.
We thank the Instituto de Ecología, A. C., Universidad de Los Andes, Universidade Federal de Mato Grosso do Sul, Instituto de Biología Subtropical (IBS), Universidad Nacional de Misiones and Instituto Potosino de Investigación Científica y Tecnológica (IPICYT) for the facilities offered to develop this contribution. We thank Juliana Kuchenbecker (picture B and E), J. León and A. Armida: México (picture D), Pedro H. Ribeiro: Brasil (picture G) and S. López: México (picture J) for image contributions to Supplementary Figure S1. W. Bruce Campbell reviewed English of the final version of the article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1084009/full#supplementary-material
Supplementary Table S1 | Sources for analysis of taxonomic diversity, functional diversity, and dung beetle ecological functions in grazing lands.
Supplementary Table S2 | Taxonomic, Functional and Ecological Functions and ecosystem services synthesis of Neotropical dung beetles from pasture habitats.At the end of the Table, please insert: In most of the revised publications, the authors considered more than one category of this table, therefore, the percentage described in results vary and include more of 26 studies in count of functional synthesis
Supplementary Table S3 | Information about grazing lands in the reviewed articles.At the end of the Table, please insert: In most of the reviewed publications the authors considered more than one type of grazing land for sampling sites. Then, in the row type of grazing land, we included the percentage of sites by each type of grazing land and in parentheses the number of sites for each main category (grassland, woodland, agroforestry and others) included in all articles according to the type of analysis. In the row Particular grazing lands are mentioned grazing lands in particular. See levels of grazing lands in Methods. SPS= silvopastoral systems. TDF= Tropical Deciduous Forest.
Supplementary Table S4 | Species of grasses mentioned as a part of grazing lands in the reviewed articles according to the topic analyzed.
Supplementary Table S5 | (A) Percentage of studies with information on livestock management. (B) Percentage of studies with information on management practices.
Supplementary Box I | Data collection format for ecological pasture studies.
Abot, A. R., Puker, A., Taira, T. L., Rodrigues, S. R., Korasaki, V., and de Oliveira, H. N. (2012). Abundance and diversity of coprophagous beetles (Coleoptera: Scarabaeidae) caught with a light trap in a pasture area of the Brazilian Cerrado. Stud. Neotropical Fauna Environ. 47, 53–60. doi: 10.1080/01650521.2012.662846
Allen, V. G., Batello, C., Berretta, E. J., Hodgson, J., Kothmann, M., Li, X., et al. (2011). An international terminology for grazing lands and grazing animals. Grass Forage Sci. 66:2. doi: 10.1111/j.1365-2494.2010.00780.x
Almeida de, S. D. S., and Louzada, J. N. (2009). Estrutura da comunidade de Scarabaeinae (Scarabaeidae: Coleoptera) em fitofisionomias do Cerrado e sua importância para a conservação. Neotrop. Entomol. 38, 32–43. doi: 10.1590/S1519-566X2009000100003
Almeida, S., Louzada, J., Sperber, C., and Barlow, J. (2011). Subtle land-use change and tropical biodiversity: dung beetle communities in Cerrado grasslands and exotic pastures. Biotropica 43, 704–710. doi: 10.1111/j.1744-7429.2011.00751.x
Alvarado, F., Andrade, E. R., Santos, B. A., Prescott, G., Souza, G., and Escobar, F. (2018). Forest cover is more important than farmland heterogeneity and livestock intensification for the retention of dung beetle phylogenetic diversity. Ecol. Indic. 93, 524–532. doi: 10.1016/j.ecolind.2018.05.041
Alvarado, F., Dáttilo, W., and Escobar, F. (2019). Linking dung beetle diversity and its ecological function in a gradient of livestock intensification management in the Neotropical region. Appl. Soil Ecol. 143, 173–180. doi: 10.1016/j.apsoil.2019.06.016
Alvarado, F., Liberal, C. N., Dantas, T. A., Bérgamo, D. B., and Menezes, R. F. (2021). Diversity and resource selection of dung beetles in a relictual mountain forest in Brazil. Int. J. Trop. Insect Sci. 41, 1343–1353. doi: 10.1007/s42690-020-00327-0
Andresen, E. (2002). Dung beetles in a central Amazonian rainforest and their ecological role as secondary seed dispersers. Ecol. Entomol. 27, 257–270. doi: 10.1046/j.1365-2311.2002.00408.x
Andresen, E. (2008). Dung beetle assemblages in primary forest and disturbed habitats in a tropical dry forest landscape in western Mexico. J. Insect Conserv. 12, 639–650. doi: 10.1007/s10841-007-9100-y
Anduaga, S. (2004). Impact of the activity of dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) inhabiting pasture land in Durango, Mexico. Environ. Entomol. 33, 1306–1312. doi: 10.1603/0046-225X-33.5.1306
Anduaga, S., and Huerta, C. (2007). Importance of dung incorporation activity by three species of coprophagous beetle (Coleoptera: Scarabaeidae: Scarabaeinae) macrofauna in pastureland on “La Michilía” biosphere reserve in Durango, Mexico. Environ. Entomol. 36, 555–559.
Arellano, L., León-Cortés, J. L., and Halffter, G. (2008). Response of dung beetle assemblages and their conservation in remnant natural and modified habitats in southern Mexico. Insect Conserv. Divers. 1, 253–262. doi: 10.1111/j.1752-4598.2008.00033.x
Arellano, L., León-Cortés, J. L., Halffter, G., and Montero, J. (2013). Acacia woodlots, cattle and dung beetles (Coleoptera: Scarabaeinae) in a Mexican silvopastoral landscape. Rev. Mex. Biodivers. 84, 650–660. doi: 10.7550/rmb.32911
Barragán, F., Douterlungne, D., Ramírez-Hernández, A., Gelviz-Gelvez, S. M., Miranda, A. V. G., and Rodas-Ortíz, J. P. (2022). The rolling dung master: an ecosystem engineer beetle mobilizing soil nutrients to enhance plant growth across a grassland management intensity gradient in drylands. J. Arid Environ. 197:104673. doi: 10.1016/j.jaridenv.2021.104673
Barragán, F., Labastida, J., and Ramírez-Hernández, A. (2021). Response of dung beetle diversity to three livestock management systems in drylands of Central Mexico. J. Arid Environ. 193:104598. doi: 10.1016/j.jaridenv.2021.104598
Barragán, F., Moreno, C. E., Escobar, F., Bueno-Villegas, J., and Halffter, G. (2014). The impact of grazing on dung beetle diversity depends on both biogeographical and ecological context. J. Biogeogr. 41, 1991–2002. doi: 10.1111/jbi.12351
Barragán, F., Moreno, C. E., Escobar, F., Halffter, G., and Navarrete, D. (2011). Negative impacts of human land use on dung beetle functional diversity. PLoS One 6:e17976. doi: 10.1371/journal.pone.0017976
Barretto, J., Salomão, R. P., and Iannuzzi, L. (2020). Diversity of dung beetles in three vegetation physiognomies of the Caatinga dry forest. Int. J. Trop. Insect Sci. 40, 385–392. doi: 10.1007/s42690-019-00089-4
Basto-Estrella, G. S., Rodriguez-Vivas, R. I., Delfin-Gonzalez, H., Navarro-Alberto, J. A., Favila, M. A., and Reyes-Novelo, E. (2016). Dung removal by dung beetles (Coleoptera: Scarabaeidae) and macrocyclic lactone use on cattle ranches of Yucatan, Mexico. Rev. Biol. Trop. 64, 945–954. doi: 10.22201/ib.20078706e.2012.2.943
Basto-Estrella, G. S., Rodríguez-Vivas, R. I., Delfín-González, H., and Reyes-Novelo, E. (2012). Escarabajos estercoleros (Coleoptera: Scarabaeidae: Scarabaeinae) de ranchos ganaderos de Yucatán, México. Rev. Mex. Biodivers. 83, 380–386. doi: 10.22201/ib.20078706e.2012.2.943
Bokdam, J., and Gleichman, J. M. (2000). Effects of grazing by free-ranging cattle on vegetation dynamics in a continental north-west European heathland. J. Appl. Ecol. 37, 415–431. cc
Bourg, A., Escobar, F., MacGregor-Fors, I., and Moreno, C. (2016). Got dung? Resource selection by dung beetles in Neotropical forest fragments and cattle pastures. Neotrop. Entomol. 45, 490–498. doi: 10.1007/s13744-016-0397-7
Braga, R. F., Korasaki, V., Andreses, E., and Louzada, J. (2013). Dung beetle community and functions along a habitat-disturbance gradient in the Amazon: a rapid assessment of ecological functions associated to biodiversity. PLoS One 8:e57786. doi: 10.1371/journal.pone.0057786
Brand, U., and Vadrot, A. (2013). Epistemic selectivities and the valorisation of nature: the cases of the Nagoya protocol and the intergovernmental science-policy platform for biodiversity and ecosystem services (IPBES). Law Env't & Dev. J. 9:202.
Bui, V. B., Ziegler, T., and Bonkowski, M. (2020). Morphological traits reflect dung beetle response to land use changes in tropical karst ecosystems of Vietnam. Ecol. Indic. 108:105697. doi: 10.1016/j.ecolind.2019.105697
Butler, L., Cropper, J., Johnson, R., Norman, A., Peacock, G., Shaver, P., et al. (2003). National range and pasture handbook. USDA National Resources Conservation Service, Washington, DC, USA, 214.
Cajaiba, R. L., Périco, E., da Silva, W. B., Caron, E., Buss, B. C., Dalzochio, M., et al. (2020). Are primary forests irreplaceable for sustaining Neotropical landscapes biodiversity and functioning? Contributions for restoration using ecological indicators. Land Degrad. Dev. 31, 508–517. doi: 10.1002/ldr.3467
Carvalho, R. L., Andersen, A. N., Anjos, D. V., Pacheco, R., Chagas, L., and Vasconcelos, H. L. (2020). Understanding what bioindicators are actually indicating: linking disturbance responses to ecological traits of dung beetles and ants. Ecol. Indic. 108:105764. doi: 10.1016/j.ecolind.2019.105764
Carvalho, R. L., Andresen, E., Anjos, D. V., Franca, F., and Vasconcelos, H. L. (2021). Dung beetle functions in tropical planted pastures were barely explained by management variables and not at all by community metrics. Ecol. Indic. 125:107598. doi: 10.1016/j.ecolind.2021.107598
Chown, S. L., and Gaston, K. J. (2010). Body size variation in insects: a macroecological perspective. Biol. Rev. 85, 139–169. doi: 10.1111/j.1469-185X.2009.00097.x
Chown, S.L., and Klok, C.J. (2011) “The ecological implications of physiological diversity in dung beetles”, in Ecology and evolution of dung beetles, ed. L.W. Simmons, and T.J. Ridsdill-Smith (HNJ: Blackwell Publishing Ltd), 200–219.
Correa, C. M. A., Audino, L. D., Holdbrook, R., Braga, R. F., Menéndez, R., and Louzada, J. (2020b). Successional trajectory of dung beetle communities in a tropical grassy ecosystem after livestock grazing removal. Biodivers. Conserv. 29, 2311–2328. doi: 10.1007/s10531-020-01975-x
Correa, C. M. A., Braga, R. F., Louzada, J., and Menéndez, R. (2019a). Dung beetle diversity and functions suggest no major impacts of cattle grazing in the Brazilian Pantanal wetlands. Ecol. Entomol. 44, 524–533. doi: 10.1111/een.12729
Correa, C. M. A., Braga, R. F., Puker, A., Abot, A. R., and Korasaki, V. (2018). Optimizing methods for dung beetle (Coleoptera: Scarabaeidae) sampling in Brazilian pastures. Environ. Entomol. 47, 48–54. doi: 10.1093/ee/nvx191
Correa, C. M. A., Braga, R. F., Puker, A., and Korasaki, V. (2019b). Patterns of taxonomic and functional diversity of dung beetles in a human-modified landscape in Brazilian Cerrado. J. Insect Conserv. 23, 89–99. doi: 10.1007/s10841-018-00118-6
Correa, C. M. A., da Silva, P. G., Puker, A., and Abot, A. R. (2021b). Spatiotemporal patterns of taxonomic and functional β-diversity of dung beetles in native and introduced pastures in the Brazilian Pantanal. Austral. Ecol. 46, 98–110. doi: 10.1111/aec.12963
Correa, C. M. A., Ferreira, K. R., Abot, A. R., Louzada, J., and Vaz-de-Mello, F. Z. (2022). Ivermectin impacts on dung beetle diversity and their ecological functions in two distinct Brazilian ecosystems. Ecol. Entomol. 47, 736–748. doi: 10.1111/een.13158
Correa, C. M. A., Lara, M. A., Puker, A., Noriega, J. A., and Korasaki, V. (2021a). Quantifying responses of dung beetle assemblages to cattle grazing removal over a short-term in introduced Brazilian pastures. Acta Oecol. 110:103681. doi: 10.1016/j.actao.2020.103681
Correa, C. M. A., Peres, N. D., and Holdbrook, R. (2020c). Patterns of alimentary resource use by dung beetles in introduced Brazilian pastures: cattle versus sheep dung. Entomol. Sci. 23, 271–279. doi: 10.1111/ens.12425
Correa, C. M. A., Puker, A., and Abot, A. R. (2020a). Impacts of exotic pasture establishment on dung beetle assemblages (Coleoptera: Scarabaeidae: Scarabaeinae) in the Brazilian Cerrado. Environ. Entomol. 49, 1335–1344. doi: 10.1093/ee/nvaa132
Correa, C. M. A., Puker, A., Ferreira, K. R., Cristaldo, C. M., Ferreira, F. N. F., Abot, A. R., et al. (2016a). Using dung beetles to evaluate the conversion effects from native to introduced pasture in the Brazilian Pantanal. J. Insect Conserv. 20, 447–456. doi: 10.1007/s10841-016-9877-7
Correa, C. M. A., Puker, A., Korasaki, V., Ferreira, K. R., and Abot, A. R. (2016b). Attractiveness of baits to dung beetles in Brazilian savanna and exotic pasturelands. Entomol. Sci. 19, 112–123. doi: 10.1111/ens.12169
Correa, C. M. A., Puker, A., Korasaki, V., and Oliveira, N. G. D. (2013). Dung beetles (Coleoptera, Scarabaeinae) attracted to sheep dung in exotic pastures. Rev. Bras. Entomol. 57, 113–116. doi: 10.1590/S0085-56262013000100017
Costa, W., Fonseca, L., and Körting, T. (2015). “Classifying grasslands and cultivated pastures in the brazilian cerrado using support vector machines, multilayer perceptrons and autoencoders” in International workshop on machine learning and data Mining in Pattern Recognition (Cham: Springer), 187–198.
Costa, C., Oliveira, V. H. F., Maciel, R., Beiroz, W., Korasaki, V., and Louzada, J. (2017). Variegated tropical landscapes conserve diverse dung beetle communities. PeerJ 5:e3125. doi: 10.7717/peerj.3125
Cruz, M., Martínez, I., Lopez-Collado, J., Vargas-Mendoza, M., Gonzalez-Hernandez, H., and Platas-Rosado, D. E. (2012). Degradación del estiercol vacuno por escarabajos estercoleros en un pastizal tropical de Veracruz, Mexico. Rev. Colom. Entom. 38, 148–155.
da Silva, P. G., and Audino, L. D. (2011). Escarabeíneos (Coleoptera: Scarabaeidae: Scarabaeinae) atraídos a diferentes iscas em campo nativo de Bagé, Rio Grande do Sul, Brasil. Rev. Brasileira de Zoociências 13, 241–247.
da Silva, P. G., da Rosa Garcia, M. A., and Vidal, M. B. (2008). Espécies de besouros copro-necrófagos (Coleoptera: Scarabaeidae) da Região da Campanha do Rio Grande do Sul. Revista de Ciências Agroveterinárias. 7, 143–149.
da Silva, P. G., and Hernández, M. I. M. (2014). Local and regional effects on community structure of dung beetles in a mainland-island scenario. PLoS One 9:e111883. doi: 10.1371/journal.pone.0111883
da Silva, P. G., and Hernández, M. I. M. (2016). Spatial variation of dung beetle assemblages associated with forest structure in remnants of southern Brazilian Atlantic Forest. Rev. Bras. Entomol. 60, 73–81. doi: 10.1016/j.rbe.2015.11.001
Davies, R. W., Edwards, D. P., and Edwards, F. A. (2020). Secondary tropical forests recover dung beetle functional diversity and trait composition. Anim. Conserv. 23, 617–627. doi: 10.1111/acv.12584
Davies, R. W., Edwards, D. P., Medina-Uribe, C. A., Cárdenas-Bautista, J. S., Haugaasen, T., Gilroy, J. J., et al. (2021). Replacing low-intensity cattle pasture with oil palm conserves dung beetle functional diversity when paired with forest protection. J. Environ. Manag. 283:112009. doi: 10.1016/j.jenvman.2021.112009
de Farias, P. M., Arellano, L., Hernández, M. I. M., and López, S. (2015). Response of the copronecrophagous beetle (Coleoptera: Scarabaeinae) assemblage to a range of soil characteristics and livestock management in a Mexican tropical landscape. J. Insect Conserv. 19, 94–960. doi: 10.1007/s10841-015-9812-3
de Farias, P. M., and Hernández, M. I. M. (2017). Dung beetles associated with agroecosystems of southern Brazil: relationship with soil properties. Rev. Bras. Cienc. Solo. 41. doi: 10.1590/18069657rbcs20160248
deCastro-Arrazola, I., Andrew, N. R., Berg, M. P., Curtsdotter, A., Lumaret, J. P., Menéndez, R., et al. (2023). A trait-based framework for dung beetle functional ecology. J. Anim. Ecol. 92, 44–65. doi: 10.1111/1365-2656.13829
deCastro-Arrazola, I., Hortal, J., Noriega, J. A., and Sánchez-Piñero, F. (2020). Assessing the functional relationship between dung beetle traits and dung removal, burial, and seedling emergence. Ecology 101:e03138. doi: 10.1002/ecy.3138
Díaz, A., Galante, E., and Favila, M. E. (2010). The effect of the landscape matrix on the distribution of dung and carrion beetles in a fragmented tropical rain forest. J. Insect Sci. 10:81. doi: 10.1673/031.010.8101
Dixon, A. P., Faber-Langendoen, D., Josse, C., Morrison, J., and Loucks, C. J. (2014). Distribution mapping of world grassland types. J. Biogeogr. 41, 2003–2019. doi: 10.1111/jbi.12381
Duffus, N.E., Echeverri, A., Dempewolf, L., Noriega, J. A., Furumo, P. R., and Morimoto, J. (2023). The present and future of insect biodiversity conservation in the Neotropics: Policy gaps and recommendations. Neotrop. Entomol. doi:doi: 10.1007/s13744-023-01031-7 [Epub ahead of print].
Eaton, D. P., Santos, S. A., Santos, M. C. A., Lima, J. V. B., and Keuroghlian, A. (2011). Rotational grazing of native pasturelands in the Pantanal: an effective conservation tool. Trop. Conserv. Sci. 4, 39–52. doi:doi: 10.1177/194008291100400105
Escobar, F., and Chacón de Ulloa, P. (2000). Distribución espacial y temporal en un gradiente de sucesión de la fauna de coleópteros coprófagos (Scarabaeinae, Aphodiinae) en un bosque tropical montano, Nariño-Colombia. Rev. Biol. Trop. 48, 961–975.
Favila, M. E. (2005). “Diversidad alfa y beta de los escarabajos del estiércol (Scarabaeinae) en Los Tuxtlas, México” in Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma. eds. G. Halffter, J. Soberón, P. Koleff, and A. Melic (Monografías 3ercer Milenio. SEA / CONABIO / DIVERSITAS / CONACYT) (España: Zaragoza), 209–219.
Favila, M. E., and Halffter, G. (1997). The use of indicator groups for measuring biodiversity as related to community structure and function. Acta Zool. Mex. 72, 1–25.
Ferraz-de-Oliveira, M. I., Azeda, C., and Pinto-Correia, T. (2016). Management of Montados and Dehesas for high nature value: an interdisciplinary pathway. Agrofor. Syst. 90, 1–6. doi: 10.1007/s10457-016-9900-8
Flota-Bañuelos, C., López-Collado, J., Vargas-Mendoza, M., Fajersson, P., González-Hernández, H., and Martínez-Morales, I. (2012). Efecto de la ivermectina en la dinámica espacio-temporal de escarabajos estercoleros en Veracruz, México. Trop. Subtrop. Agroecosyst. 15, 227–239.
Gann, G. D., McDonald, T., Walder, B., Aronson, J., Nelson, C. R., Jonson, J., et al. (2019). International principles and standards for the practice of ecological restoration. Restor. Ecol. 27, S1–S46. doi: 10.1111/rec.13035
Garrido, P., Elbakidze, M., Angelstam, P., Plieninger, T., Pulido, F., and Moreno, G. (2017). Stakeholder perspectives of wood-pasture ecosystem services: a case study from Iberian dehesas. Land Use Policy 60, 324–333. doi: 10.1016/j.landusepol.2016.10.022
Giménez-Gómez, V. C., Verdú, J. R., Casanoves, F., and Zurita, G. A. (2022). Functional responses to anthropogenic disturbance and the importance of selected traits: a study case using dung beetles. Ecol. Entomol. 47, 503–514. doi: 10.1111/een.13135
Giraldo, C., Escobar, F., Chara, J. D., and Calle, Z. (2011). The adoption of silvopastoral systems promotes the recovery of ecological processes regulated by dung beetles in the Colombian Andes. Ins. Conserv. Div. 4, 115–122. doi: 10.1111/j.1752-4598.2010.00112.x
Gómez-Cifuentes, A., Giménez, G. V., and C., Moreno, C. E., and Zurita, G. A., (2019). Tree retention in cattle ranching systems partially preserves dung beetle diversity and functional groups in the semi-deciduous Atlantic forest: the role of microclimate and soil conditions. Basic Appl. Ecol. 34, 64–74. doi: 10.1016/j.baae.2018.10.002
Gómez-Cifuentes, A., Huerta, C., Zurita, G. A., and Arellano, L. (2022). The influence of biodiversity-friendly ranching practices on dung beetle diversity in a Mexican mountainous tropical landscape. J. Insect Conserv. 26, 721–734. doi: 10.1007/s10841-022-00414-2
Gómez-Cifuentes, A., Vespa, N., Semmartín, M., and Zurita, G. (2020). Canopy cover is a key factor to preserve the ecological functions of dung beetles in the southern Atlantic Forest. Appl. Soil Ecol. 154:103652. doi: 10.1016/j.apsoil.2020.103652
Guerra-Alonso, C. B., Zurita, G. A., and Bellocq, M. I. (2022). Livestock grazing impact differently on the functional diversity of dung beetles depending on the regional context in subtropical forests. Sci. Rep. 12, 1–11. doi: 10.1038/s41598-022-05616-x
Guevara, S., and Lira-Noriega, A. (2004) De los pastos de la selva a la selva de los pastos: la introducción de la ganadería en México. Pastos. 34, 109–150. Available at: http://polired.upm.es/index.php/pastos/article/view/1331 [Accessed August 16, 2022].
Halffter, G., and Arellano, L. (2002). Response of dung beetle diversity to human–induced changes in a tropical landscape. Biotropica 34, 144–154. doi: 10.1111/j.1744-7429.2002.tb00250.x
Halffter, G., and Edmonds, W.D. (1982). The nesting behavior of dung beetles (Scarabaeinae). An ecological and evolutive approach. México, DF: Instituto de Ecología.
Halffter, G., and Favila, M. E. (1993). The Scarabaeinae (Insecta: Coleoptera), an animal group for analyzing, inventorying and monitoring biodiversity in tropical rainforest and modified landscapes. Biol. Int. 27, 15–21.
Halffter, G., Favila, M. E., and Halffter, V. (1992). A comparative study of the structure of the scarab guild in Mexican tropical rainforest and derived ecosystems. Folia Entomol. Mex. 84, 131–156.
Halffter, G., Pineda, E., Arellano, L., and Escobar, F. (2007). Instability of copronecrophagous beetle assemblages (Coleoptera: Scarabaeinae) in a mountainous tropical landscape of Mexico. Environ. Entomol. 36, 1397–1407. doi: 10.1603/0046-225X(2007)36[1397:IOCBAC]2.0.CO;2
Hanski, I., and Cambefort, Y. (1991). Dung beetle ecology. Princeton, NJ: Princeton University Press. doi:doi: 10.1515/9781400862092
Hasanuzzaman, Md. (2020). Pasture management. In fodder production and management. AGRO 120. 4 pp. Available at: https://hasanuzzaman.webs.com/publications.htm [Accessed April 18, 2022].
Herrero, M., Wirsenius, S., Henderson, B., Rigolot, C., Thornton, P., Havlík, P., et al. (2015). Livestock and the environment: what have we learned in the past decade? Annu. Rev. Environ. Resour. 40, 177–202. doi:doi: 10.1146/annurev-environ-031113-093503
Holechek, J., Pieper, R. D., and Herbel, C. H. (2011). Range management: Principles and practices. Englewood Cliffs, NJ: Prentice Hall, 444.
Holter, P. (2016). Herbivore dung as food for dung beetles: elementary coprology for entomologists. Ecol. Entomol. 41, 367–377. doi: 10.1111/een.12316
Horgan, F. G. (2001). Burial of bovine dung by coprophagous beetles (Coleoptera: Scarabaeidae) from horse and cow grazing sites in El Salvador. Eur. J. Soil Biol. 37, 103–111. doi: 10.1016/S1164-5563(01)01073-1
Horgan, F. G. (2002). Shady field boundaries and the colonization of dung by coprophagous beetles in central American pastures. Agric. Ecosyst. Environ. 91, 25–36. doi: 10.1016/S0167-8809(01)00261-4
Horgan, F. G. (2005). Effects of deforestation on diversity, biomass and function of dung beetles on the eastern slopes of the Peruvian Andes. For. Ecol. Manag. 216, 117–133. doi: 10.1016/j.foreco.2005.05.049
Horgan, F. G. (2008). Dung beetle assemblages in forests and pastures of El Salvador: a functional comparison. Biodivers. Conserv. 17, 2961–2978. doi: 10.1007/s10531-008-9408-2
Huerta, C., Arellano, L., and Cruz-Rosales, M. (2018). Dung beetles (Coleoptera: Scarabaeidae, Scarabaeinae) and dung removal in Mexican livestock pastures: the relationship with functional groups. Rev. Mex. Biodivers. 89, 1280–1292. doi: 10.22201/ib.20078706e.2018.4.2495
Hutton, S. A., and Giller, P. S. (2003). The effects of the intensification of agriculture on northern temperate dung beetle communities. J. Appl. Ecol. 40, 994–1007. doi: 10.1111/j.1365-2664.2003.00863.x
Iwasa, M., Maruo, T., Ueda, M., and Yamashita, N. (2007). Adverse effects of ivermectin on the dung beetles, Caccobius jessoensis Harold, and rare species, Copris ochus Motschulsky and Copris acutidens Motschulsky (Coleoptera: Scarabaeidae), in Japan. Bull. Entomol. Res. 97, 619–625. doi: 10.1017/S0007485307005329
Jurado-Guerra, P., Velázquez-Martínez, M., Sánchez-Gutiérrez, R. A., Álvarez-Holguín, A., Domínguez-Martínez, P. A., Gutiérrez-Luna, R., et al. (2021). Los pastizales y matorrales de zonas áridas y semiáridas de México: Estatus actual, retos y perspectivas. Rev. Mex. Cienc. Pecu. 12, 261–285. doi: 10.22319/rmcp.v12s3.5875
Korasaki, V., Braga, R. F., Zanetti, R., Moreira, F., Vaz-de-Mello, F. Z., and Louzada, J. (2013). Conservation value of alternative land-use systems for dung beetles in Amazon: valuing traditional farming practices. Biodivers. Conserv. 22, 1485–1499. doi: 10.1007/s10531-013-0487-3
Kryger, U., Deschodt, C., Davis, A. L., and Scholtz, C. H. (2006). Effects of cattle treatment with a cypermethrin/cymiazol spray on survival and reproduction of the dung beetle species Euoniticellus intermedius (Coleoptera: Scarabaeidae). Bull. Entomol. Res. 96, 597–603. doi: 10.1079/BER2006463
Laurance, W. F., Useche, D. C., Shoo, L. P., Herzog, S. K., Kessler, M., Escobar, F., et al. (2011). Global warming, elevational ranges and the vulnerability of tropical biota. Biol. Conserv. 144, 548–557. doi: 10.1016/j.biocon.2010.10.010
Larsen, T. H., Lopera, A., and Forsyth, A. (2008). Understanding trait‐dependent community disassembly: dung beetles, density functions, and forest fragmentation. Conserv. Biol. 22, 1288–1298.
Lobo, J. M., Hortal, J., and Cabrero-Sañudo, F. J. (2006). Regional and local influence of grazing activity on the diversity of a semi-arid dung beetle community. Divers. Distrib. 12, 111–123. doi: 10.1111/j.1366-9516.2006.00194.x
Longland, A. C. (2013). “Pasture and pasture management” in Equine Applied and Clinical Nutrition. eds. J. Geor, A. Raymond, H. Patricia, and C. Manfred. Elsevier Ltd. 332–350. doi: 10.1016/C2009-0-39370-8
Lopes, J., Korasaki, V., Catelli, L. L., Marçal, V. V., and Nunes, M. P. B. (2011). A comparison of dung beetle assemblage structure (Coleoptera: Scarabaeidae: Scarabaeinae) between an Atlantic forest fragment and adjacent abandoned pasture in Paraná, Brazil. Zoologia 28, 72–79. doi: 10.1590/S1984-46702011000100011
Lopez-Collado, J., Cruz-Rosales, M., Vilaboa-Arroniza, J., Martínez-Morales, I., and Gonzalez-Hernandez, H. (2017). Contribution of dung beetles to cattle productivity in the tropics: a stochastic-dynamic modeling approach. Agric. Syst. 155, 78–87. doi: 10.1016/j.agsy.2017.05.001
Lousey, J. E., and Vaughan, M. (2006). The economic value of ecological services provided by insects. Bioscience 56, 311–323. doi: 10.1641/0006-3568(2006)56[311:TEVOES]2.0.CO;2
Louzada, J. N., and Carvalho e Silva, P. R. (2009). Utilization of introduced Brazilian pasture ecosystems by native dung beetles: diversity patterns and resource use. Insect Conserv. Divers. 2, 45–52. doi: 10.1111/j.1752-4598.2008.00038.x
Macagno, A. L. M., and Palestrini, C. (2009). The maintenance of extensively exploited pastures within the alpine mountain belt: implications for dung beetle conservation (Coleoptera: Scarabaeoidea). Biodivers. Conserv. 18, 3309–3323. doi: 10.1007/s10531-009-9643-1
Macedo, R., Audino, L. D., Korasaki, V., and Louzada, J. (2020). Conversion of Cerrado savannas into exotic pastures: the relative importance of vegetation and food resources for dung beetle assemblages. Agric. Ecosyst. Environ. 288:106709. doi: 10.1016/j.agee.2019.106709
Manning, P., Slade, E. M., Beynon, S. A., and Lewis, O. T. (2016). Functionally rich dung beetle assemblages are required to provide multiple ecosystem services. Agric. Ecosyst. Environ. 218, 87–94. doi: 10.1016/j.agee.2015.11.007
Manning, P., Slade, E. M., Beynon, S. A., and Lewis, O. T. (2017). Effect of dung beetle species richness and chemical perturbation on multiple ecosystem functions. Ecol. Entomol. 42, 577–586. doi: 10.1111/een.12421
Mariategui, P., Speicys, C., Urretabizkaya, N., and Fernández, E. (2001). Efecto de Ontherus sulcator F. (Coleoptera: Scarabaeidae) en la incorporacion de estiercol al suelo. Zootec. Trop. 19, 131–138.
Martínez, I., Flota-Bañuelos, C., Tokman, D. G., and Ortiz, M. R. (2018). Dung and gastrointestinal nematode eggs buried by Euoniticellus intermedius (Reiche) (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopterist Bull. 72, 188–194. doi: 10.1649/0010-065X-72.1.188
Martínez, I., Ramírez-Hernández, A., and Lumaret, J.-P. (2017). Medicinas veterinarias, pesticidas y los escarabajos del estiércol de la zona tropical de Palma Sola, Veracruz, México. Southwest. Entomol 42, 563–574. doi: 10.3958/059.042.0225
Martínez-Falcón, A. P., Zurita, G. A., Ortega-Martínez, I. J., and Moreno, C. E. (2018). Populations and assemblages living on the edge: dung beetles responses to forests-pasture ecotones. PeerJ 6:e6148. doi: 10.7717/peerj.6148
MEA [Millennium Ecosystem Assessment] (2005). Ecosystems and human well-being: Synthesis. Washington, DC: Island Press.
Mellink, E., and Riojas-López, M. E. (2020). Livestock and grassland interrelationship along five centuries of ranching the semiarid grasslands on the southern highlands of the Mexican plateau. Elem. Sci. Anth. 8:20. doi: 10.1525/elementa.416
Miranda, C. H. B., dos Santos, J. C., and Bianchin, I. (2000). The role of Digitonthophagus gazella in pasture cleaning and production as a result of burial of cattle dung. Pasturas Tropicales 22, 14–18.
Moher, D., Liberati, A., Tetzlaff, J., and Altman, D. G., Prisma Group (2009). Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Ann. Intern. Med. 151, 264–269. doi: 10.7326/0003-4819-151-4-200908180-00135
Montoya-Molina, S., Giraldo-Echeverri, C., Montoya-Lerma, J., Chará, J., Escobar, F., and Calle, Z. (2016). Land sharing vs. land sparing in the dry Caribbean lowlands: a dung beetles’ perspective. Appl. Soil Ecol. 98, 204–212. doi: 10.1016/j.apsoil.2015.10.017
Moore, J. C. (2001). “Diversity, taxonomic versus functional” in Encyclopedia of biodiversity. ed. S. A. Levin, vol. 2 (USA: Elsevier Science), 205–215.
Moreno, C. E., Calderón-Patrón, J. M., Martín-Regalado, N., Martínez-Falcón, A. P., Ortega-Martínez, I. J., Rios-Díaz, C. L., et al. (2018). Measuring species diversity in the tropics: a review of methodological approaches and framework for future studies. Biotropica 50, 929–941. doi: 10.1111/btp.12607
Moreno, C. E., Sánchez-Rojas, G., Pineda, E., and Escobar, F. (2007). Shortcuts for biodiversity evaluation: a review of terminology and recommendations for the use of target groups, bioindicators and surrogates. Int. J. Environ. Health 1, 71–86. doi: 10.1504/IJENVH.2007.012225
Morrone, J.J. (2017). Neotropical biogeography. Regionalization and evolution. Boca Raton, FL, USA: CRC Press, Taylor & Francis Group.
Murgueitio, E., and Solorio, B. (2008). El Sistema Silvopastoril Intensivo, un modelo exitoso para la competitividad ganadera en Colombia y México. In V Congreso Latinoamericano de Agroforestería para la Producción Pecuaria Sostenible (proceedings). Universidad Rómulo Gallegos, Universidad Central de Venezuela, Universidad de Zulia. Maracay, Venezuela (electronic publication).
Naeem, S., Bunker, D. E., Hector, A., Loreau, M., and Perrings, C. (2009). Biodiversity, ecosystem functioning, and human wellbeing: An ecological and economic perspective. Oxford: University Press.
Nahed-Toral, J., Valdivieso-Pérez, A., Aguilar-Jiménez, R., Cámara-Cordova, J., and Grande-Cano, D. (2013). Silvopastoral systems with traditional management in southeastern Mexico: a prototype of livestock agroforestry for cleaner production. J. Clean. Prod. 57, 266–279. doi: 10.1016/j.jclepro.2013.06.020
Navarrete, D., and Halffter, G. (2008). Dung beetle (Coleoptera: Scarabaeidae: Scarabaeinae) diversity in continuous forest, forest fragments and cattle pastures in a landscape of Chiapas, Mexico: the effects of anthropogenic changes. Biodivers. Conserv. 17, 2869–2898. doi: 10.1007/s10531-008-9402-8
Newbold, T., Hudson, L. N., Hill, S., Contu, Sara, Lysenko, Igor, Senior, Rebecca A, et al. (2015). Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50. doi: 10.1038/nature14324
Nichols, E., Larsen, T., Spector, S., Davis, A. L., Escobar, F., Favila, M., et al. (2007). Global dung beetle response to tropical forest modification and fragmentation: a quantitative literature review and meta-analysis. Biol. Conserv. 137, 1–19. doi: 10.1016/j.biocon.2007.01.023
Nichols, E., Spector, S., Louzada, J., Larsen, T. H., Amezquita, S., Favila, M., et al. (2008). Ecological functions and ecosystem services of Scarabaeinae dung beetles: a review. Biol. Conserv. 141, 1461–1474. doi: 10.1016/j.biocon.2008.04.011
Nichols, E., Uriarte, M., Bunker, D. E., Favila, M. E., Slade, E. M., Vulinec, K., et al. (2013). Trait-dependent response of dung beetle populations to tropical forest conversion at local and regional scales. Ecology 94, 180–189. doi: 10.1890/12-0251.1
Nock, C.A., Vogt, R.J., and Beisner, B.E. (2016). Functional traits. eLS. John Wiley & Sons, Ltd: Chichester, 1–8.
Noriega, J. A., Floate, K. D., Génier, F., Reid, C. A. M., Kohlmann, B., Horgan, F. G., et al. (2020). Global distribution patterns provide evidence of niche shift by the introduced African dung beetle Digitonthophagus gazella. Entomol. Exp. Appl. 168, 766–782. doi: 10.1111/eea.12961
Noriega, J. A., March-Salas, M., Castillo, S., Garcia, Q. H., Hortal, J., and Santos, A. M. C. (2021a). Human perturbations reduce dung beetle diversity and dung removal ecosystem function. Biotropica 53, 753–766. doi: 10.1111/btp.12953
Noriega, J. A., Palacio, J. M., Monroy, J. D., and Valencia, E. (2012). Estructura de un ensamblaje de escarabajos coprófagos (Coleoptera: Scarabaeinae) en tres sitios con diferente uso del suelo en Antioquia, Colombia. Actu. Biol. 34, 43–54.
Noriega, J. A., Santos, A. M. C., Calatayud, J., Chozas, S., and Hortal, J. (2021b). Short- and long-term temporal changes in the assemblage structure of Amazonian dung beetles. Oecologia 195, 719–736. doi: 10.1007/s00442-020-04831-5
Numa, C., Verdú, J. R., Sánchez, A., and Galante, E. (2009). Effect of landscape structure on the spatial distribution of Mediterranean dung beetle diversity. Divers. Distrib. 15, 489–501. doi: 10.1111/j.1472-4642.2009.00559.x
Oliveira, J., Campbell, E. E., Lamparelli, R. A., Figueiredo, G. K., Soares, J. R., Jaiswal, D., et al. (2020). Choosing pasture maps: an assessment of pasture land classification definitions and a case study of Brazil. Int. J. Appl. Earth Obs. Geoinf. 93:102205. doi: 10.1016/j.jag.2020.102205
Ortega-Martínez, I. J., Moreno, C. E., and Escobar, F. (2016). A dirty job: manure removal by dung beetles in both a cattle ranch and laboratory setting. Entomol. Exp. Appl. 161, 70–78. doi: 10.1111/eea.12488
Otavo, S. E., Parrado-Rosselli, A., and Noriega, J. A. (2013). Scarabaeoidea superfamily (Insecta: Coleoptera) as a bioindicator element of anthropogenic disturbance in an amazon national park. Rev. Biol. Trop. 61, 735–752. doi: 10.15517/rbt.v61i2.11219
Pecenka, J. R., and Lundgren, J. G. (2018). The importance of dung beetles and arthropod communities on degradation of cattle dung pats in eastern South Dakota. PeerJ 6:e5220. doi: 10.7717/peerj.5220
Piccini, I., Nervo, B., Forshage, M., Celi, L., Palestrini, C., Rolando, A., et al. (2018). Dung beetles as drivers of ecosystem multifunctionality: are response and effect traits interwoven? Sci. Total Environ. 616, 1440–1448. doi: 10.1016/j.scitotenv.2017.10.171
Rangel-Acosta, J. L., Martínez-Hernández, N. J., and Yonoff-Zapata, R. (2020). Respuesta de los escarabajos coprófagos (Scarabaeidae: Scarabaeinae) a la modificación del hábitat causada por un incendio forestal en la Reserva Bijibana, Atlántico-Colombia. Rev. Mex. Biodivers. 91:e912879. doi: 10.22201/ib.20078706e.2020.91.2879
Reyes-Novelo, E., Delfín-González, H., and Morón, M. A. (2007). Copro-necrophagous beetle (Coleoptera: Scarabaeidae) diversity in an agroecosystem in Yucatan, Mexico. Rev. Biol. Trop. 55, 83–99. doi: 10.15517/rbt.v55i1.6059
Rezende, C. L., Scarano, F. R., Assad, E. D., Joly, C. A., Metzger, J. P., Strassburg, B. B. N., et al. (2018). From hotspot to hopespot: An opportunity for the Brazilian Atlantic Forest. Perspect. Ecol. conserv. 16, 208–214.
Righi, C. A., Sandoval Rodríguez, C., Ferreira, E. N., Godoy, W. A., and Cognato, A. I. (2018). Microclimatic conditions for dung beetle (Coleoptera: Scarabaeidae) occurrence: land use system as a determining factor. Environ. Entomol. 47, 1420–1430. doi: 10.1093/ee/nvy163
Rivera, J. D., de los Monteros, A. E., Saldaña-Vázquez, R. A., and Favila, M. E. (2023). Beyond species loss: how anthropogenic disturbances drive functional and phylogenetic homogenization of Neotropical dung beetles. Sci. Total Environ. 869:161663. doi: 10.1016/j.scitotenv.2023.161663
Rivera, J. D., Gómez, B., Navarrete-Gutiérrez, D. A., Ruíz-Montoya, L., Delgado, L., and Favila, M. E. (2020). Mechanisms of diversity maintenance in dung beetle assemblages in a heterogeneous tropical landscape. PeerJ 8:e9860. doi: 10.7717/peerj.9860
Rivera, J. D., Espinosa de los Monteros, A., da Silva, P. G., and Favila, M. E. (2022). Dung beetles maintain phylogenetic divergence but functional convergence across a highly fragmented tropical landscape. J. App. Ecol. 59:1781–1791.
Rodríguez-Moreno, O. G., Nahed-Toral, J., Guevara-Hernández, F., Alayón-Gamboa, J. A., and Grande-Cano, J. D. (2020). Historia y caracterización técnica y socioeconómica de la ganadería bovina en la Costa de Chiapas, México [history and technical and socioeconomic characterization of cattle on the coast of Chiapas, Mexico]. Trop. Subtrop. Agroecosyst. 23:55.
Rosa-García, R., Fraser, M. D., Celaya, R., Ferreira, L. M. M., García, U., and Osoro, K. (2013). Grazing land management and biodiversity in the Atlantic European heathlands: a review. Agrofor. Syst. 87, 19–43. doi: 10.1007/s10457-012-9519-3
Salomão, R. P., Favila, M. E., and González-Tokman, D. (2020). Spatial and temporal changes in the dung beetle diversity of a protected, but fragmented, landscape of the northernmost Neotropical rainforest. Ecol. Indic. 111:105968. doi: 10.1016/j.ecolind.2019.105968
Sands, B., and Wall, R. (2018). Sustained parasiticide use in cattle farming affects dung beetle functional assemblages. Agric. Ecosyst. Environ. 265, 226–235. doi: 10.1016/j.agee.2018.06.012
Schleuning, M., García, D., and Tobias, J. A. (2023). Animal functional traits: towards a trait-based ecology for whole ecosystems. Funct. Ecol. 37, 4–12. doi: 10.1111/1365-2435.14246
Seidl, A. F., Silva, J. F. V., and Moraes, A. S. (2001). Cattle ranching and deforestation in the Brazilian Pantanal. Ecol. Econ. 36, 413–425. doi: 10.1016/S0921-8009(00)00238-X
Silva, F. A. B., Costa, C. M. Q., Moura, R. C., and Farias, A. I. (2010). Study of the dung beetle (Coleoptera: Scarabaeidae) community at two sites: Atlantic forest and clear-cut, Pernambuco, Brazil. Environ. Entomol. 39, 359–367. doi: 10.1603/EN09180
Silva, R. J., Pelissari, T. D., Krinski, D., Canale, G., and Vaz-de-Mello, F. Z. (2017). Abrupt species loss of the Amazonian dung beetle in pastures adjacent to species-rich forests. J. Insect Conserv. 21, 487–494. doi: 10.1007/s10841-017-9988-9
Slade, E. M., Kirwan, L., Bell, T., Philipson, C. D., Lewis, O. T., and Roslin, T. (2017). The importance of species identity and interactions for multifunctionality depends on how ecosystem functions are valued. Ecology 98, 2626–2639. doi: 10.1002/ecy.1954
Slade, E. M., Riutta, T., Roslin, T., and Tuomisto, H. L. (2016). The role of dung beetles in reducing greenhouse gas emissions from cattle farming. Sci. Rep. 6:18140. doi: 10.1038/srep18140
Söderström, B. O., Svensson, B., Vessby, K., and Glimskär, A. (2001). Plants, insects and birds in semi-natural pastures in relation to local habitat and landscape factors. Biodivers. Conserv. 10, 1839–1863. doi: 10.1023/A:1013153427422
Souza, A., Ferreira-Châline, R., Châline, N., Korasaki, V., Beiroz, W., and Louzada, J. (2018). Does Neem trigger the same response as Ivermectin? Dung beetle behaviour and physiology. PeerJ 6:e26770v1. doi: 10.7287/peerj.preprints.26770v1
Souza, T. B., França, F. M., Barlow, J., Dodonov, P., Santos, J. S., Faria, D., et al. (2020). The relative influence of different landscape attributes on dung beetle communities in the Brazilian Atlantic forest. Ecol. Indic. 117:106534. doi: 10.1016/j.ecolind.2020.106534
Solorio, S. F. J., Wright, J., Franco, M. J. A., Basu, S. K., Sarabia, S. L., Ramírez, L., et al. (2017). Silvopastoral systems: best agroecological practice for resilient production systems under dryland and drought conditions. Quantification of climate variability, adaptation and mitigation for agricultural sustainability, 233–250.
Spector, S. (2006). Scarabaeinae dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae): an invertebrate focal taxon for biodiversity research and conservation. Coleopt. Bull. 5, 71–83. doi: 10.1649/0010-065X(2006)60[71:SDBCSS]2.0.CO;2
Steneck, R. S. (2001). “Functional groups” in Encyclopedia of biodiversity. ed. S. A. Levin, vol. 3 (USA: Elsevier Science), 121–139.
Summers, J. K., Smith, L. M., Case, J. L., and Linthurst, R. A. (2012). A review of the elements of human well-being with an emphasis on the contribution of ecosystem services. Ambio 41, 327–340. doi: 10.1007/s13280-012-0256-7
Tilman, D. (2001). “Functional diversity” in Encyclopedia of biodiversity. ed. S. A. Levin, vol. 3 (USA: Elsevier Science), 109–120.
Tocco, C., Probo, M., Lonati, M., Lombardi, G., Negro, M., Nervo, B., et al. (2013). Pastoral practices to reverse shrub encroachment of sub-alpine grasslands: dung beetles (Coleoptera, Scarabaeoidea) respond more quickly than vegetation. PLoS One 8:e83344. doi: 10.1371/journal.pone.0083344
Tonelli, M. (2021). Some considerations on the terminology applied to dung beetle functional groups. Ecol. Entomol. 46, 772–776. doi: 10.1111/een.13017
Tovar, H. L., Noriega, J. A., and Caraballo, P. (2016). Ivermectin effect on the structure of the ensembles of dung beetles (Coleoptera: Scarabaeidae: Aphodiinae-Scarabaeinae) in the Colombian savannahs of the Caribbean region. Actual. Biol. 38, 157–166. doi: 10.17533/udea.acbi.v37n105a03
Ueda, A., Dwibadra, D., Noerdjito, W. A., Kon, M., Ochi, T., Takahashi, M., et al. (2015). Effect of habitat transformation from grassland to Acacia mangium plantation on dung beetle assemblage in East Kalimantan, Indonesia. J. Insect Conserv. 19, 765–780. doi: 10.1007/s10841-015-9798-x
UNESCO (2021). Latin America. In: UNESCO science report: The race against time for smarter development. United Nations Educational, Scientific and Cultural Organization, Paris, France, p 200–233.
Verdú, J. R., Crespo, M. B., and Galante, E. (2000). Conservation strategy of a nature reserve in Mediterranean ecosystems: the effects of protection from grazing on biodiversity. Biodivers. Conserv. 9, 1707–1721. doi: 10.1023/A:1026506725251
Villada-Bedoya, S., Córdoba-Aguilar, A., Escobar, F., Martínez-Morales, I., and González-Tokman, D. (2019). Dung beetle body condition: a tool for disturbance evaluation in contaminated pastures. Environ. Toxicol. Chem. 38, 2392–2404. doi: 10.1002/etc.4548
Whitworth, A., Beirne, C., Flatt, E., Froese, G., Nuñez, C., and Forsyth, A. (2021). Recovery of dung beetle biodiversity and traits in a regenerating rainforest: a case study from Costa Rica's Osa Peninsula. Insect Conserv. Divers. 14, 439–454. doi: 10.1111/icad.12470
Keywords: cattle ranching, ecosystem functions and services, functional groups, management history, traits, Scarabaeinae
Citation: Arellano L, Noriega JA, Ortega-Martínez IJ, Rivera JD, Correa CMA, Gómez-Cifuentes A, Ramírez-Hernández A and Barragán F (2023) Dung beetles (Coleoptera: Scarabaeidae) in grazing lands of the Neotropics: A review of patterns and research trends of taxonomic and functional diversity, and functions. Front. Ecol. Evol. 11:1084009. doi: 10.3389/fevo.2023.1084009
Edited by:
Cristina Grande, Autonomous University of Madrid, SpainReviewed by:
Estefany Karen López Estrada, National Museum of Natural Sciences (CSIC), SpainCopyright © 2023 Arellano, Noriega, Ortega-Martínez, Rivera, Correa, Gómez-Cifuentes, Ramírez-Hernández and Barragán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucrecia Arellano, bHVjcmVjaWEuYXJlbGxhbm9AaW5lY29sLm14
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.