 Alexander Averianov
Alexander Averianov Ekaterina Obraztsova2
Ekaterina Obraztsova2 Jian-Hua Jin
Jian-Hua Jin
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 27 February 2023
Sec. Paleontology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1076819
This article is part of the Research Topic Biotic Changes in Terrestrial Environments around the Eocene–Oligocene Transition View all 10 articles
A new hyaenodont Maocyon peregrinus, gen. et sp. nov., is described based on a partial skull and associated mandible from the upper Eocene Youganwo Formation at Maoming locality in Guangdong Province, China. It shows certain similarities with the Hyainailouroidea in the skull structure, including anteroposteriorly extended jugal/squamosal suture, presence of a preglenoid crest, a lateral expansion of the squamosal posterior to the zygomatic arch, a transversally expanded mastoid process, a nuchal crest that does not extend laterally to mastoid process, and large occipital condyles. The phylogenetic analysis clusters the new taxon with Orienspterodon dahkoensis from the late middle Eocene of China and Myanmar and places this clade within the Hyainailouridae in a polytomy with the Apternodontinae and the Hyainailourinae.
Hyaenodonta is an extinct group of placental carnivorous mammals that flourished mostly in the Paleogene of Africa, North America, Europe, and Asia. The oldest hyaenodonts are known from the Paleocene of Africa (Gheerbrant, 1995; Solé et al., 2009) and Asia (Meng et al., 1998). Hyaenodonts included a number of small-sized mesocarnivorous animals, as well as larger hypercarnivorous taxa with specialized sectorial dentition. According to the recent phylogenetic analyses, the hypercarnivory was evolved independently in two (Polly, 1996), or three lineages of hyaenodonts (Rana et al., 2015; Borths et al., 2016; Borths and Seiffert, 2017). These three lineages are Hyaenodontinae, Teratodontinae, and Hyainailourinae. The sister taxa Hyainailourinae and Apterodontinae form the clade Hyainailouridae and the latter with its sister taxon Teratodontinae form the clade Hyainailouroidea (Borths et al., 2016; Borths and Seiffert, 2017). The Hyainailourinae were most diverse in Africa where about a dozen of taxa are currently recognized (von Stromer, 1926; Savage, 1965, 1973; Holroyd, 1999; Morlo et al., 2007; Rasmussen and Gutiérrez, 2009; Morales and Pickford, 2017; Borths and Stevens, 2019). Four genera are known in Europe (Lange-Badré, 1979; Ginsburg, 1980; Solé et al., 2015). Two genera of the Hyainailourinae each known from North America and Asia, are Hemipsalodon and Orienspterodon, respectively (Mellett, 1969; Egi et al., 2007).
The oldest hyainailourid is Orienspterodon from the middle Eocene of South China and Myanmar, known from mandible and maxilla fragments and dentition (Chow, 1975; Egi et al., 2007). The genus Orienspterodon, originally referred to the Hyainailouridae (Egi et al., 2007), was once considered as a member of the Hyaenodontinae (Lewis and Morlo, 2010; Solé et al., 2014). Removal of Orienspterodon from the Hyainailouridae leaves no counterarguments about the African origin of that clade (Solé et al., 2014, 2015). However, the more recent phylogenetic analyses placed Orienspterodon as the most basal member of the Hyainailourinae (Borths et al., 2016; Borths and Seiffert, 2017; Borths and Stevens, 2017, 2019). Here we describe a new hyaenodont taxon, closely related to Orienspterodon, based on a partial skull from the late Eocene of South China (Figure 1).
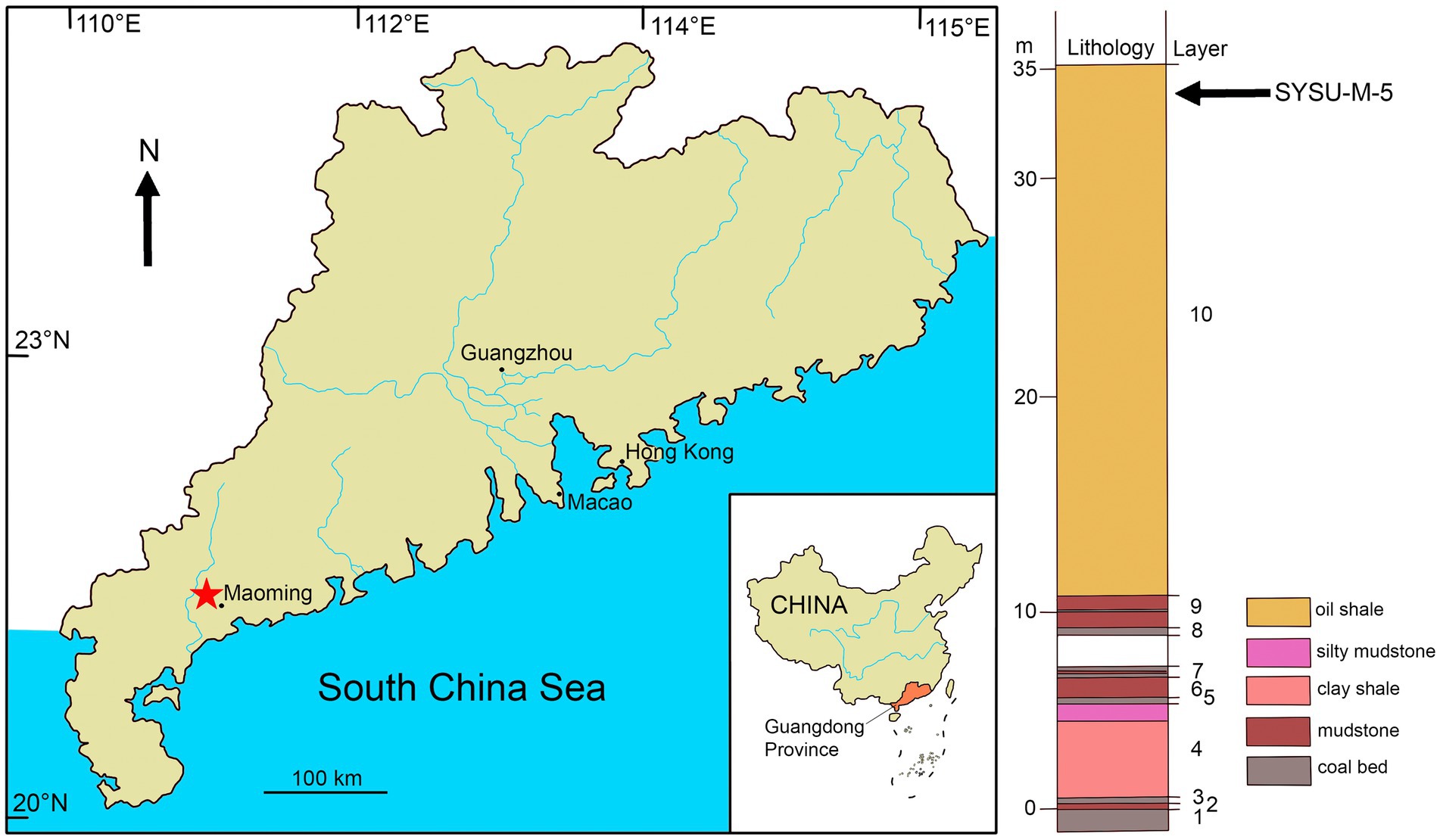
Figure 1. Geographic position of Maoming locality designated by star on the map of Guangdong Province, China (left) and stratigraphic position of SYSU-M-5, holotype of Maocyon peregrinus, gen. et sp. nov., indicated by arrow on the stratigraphic column of the Youganwo Formation (right).
Dental terminology and measurements follow Borths and Seiffert (2017).
We calculated the body mass of the new taxon as based on the equation Log10(P) = [3.5104 × Log10((ΣM)/3)] – 2.6469 provided by Solé et al. (2021), where P is the estimated body mass (in grams) and ΣM the sum of the length of the three lower molars (in mm). The length of the lower molars (m1-3) in SYSU-M-5 is equal to 48.5 mm (Table 1; the unknown m1 length was estimated as 11.7 mm based on its alveolus). The body mass of this specimen calculated using this equation is 39.43 kg. The body mass of Orienspterodon dahkoensis is estimated as 89.73 kg based on the same equation (m1-3 length is 61.3 mm; Egi et al., 2007).
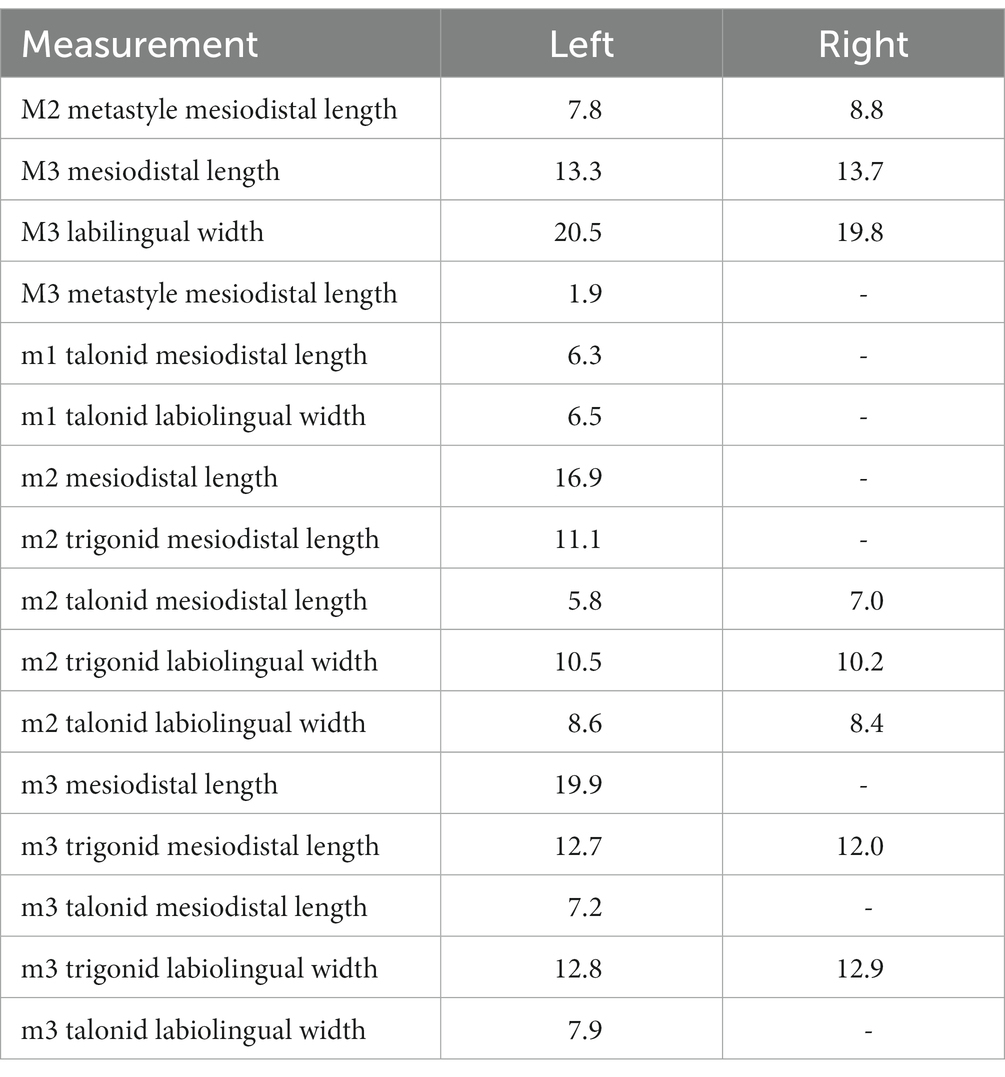
Table 1. Dental measurements of SYSU-M-5, the holotype of Maocyon peregrinus, gen. et sp. nov.
For the phylogenetic analysis we used the data matrix presented by Solé et al. (2021). The matrix includes 107 taxa and 156 characters. Maocyon peregrinus gen. et sp. nov. can be coded by 53 of these characters (34.0%): 30(1); 31(2); 32(1); 33(2); 34(2); 35(1); 36(1); 37(1); 38(0); 39(1); 40(1); 41(2); 42(1); 43(0); 44(2); 45(2); 46(2); 48(2); 49(2); 50(0); 51(1); 52(1); 53(0); 54(1); 55(1); 56(1); 57(1); 58(1); 59(0); 60(1); 61(1); 62(0); 63(1); 64(1); 65(1); 86(0); 87(2); 88(0); 99(1); 101(2); 107(1); 109(0); 110(2); 114(1); 115(1); 117(0); 118(1); 125(0); 126(1); 127(1); 128(1); 129(0); and 130(1). The characters 41, 42, 46, 48, 49, 50, 51, 52, 53, 54, 55, 87, 101, and 110 were treated as ordered. Phylogenetic analysis was performed using the Bayesian “tip-dating” phylogenetic methods first applied to Hyaenodonta by Borths et al. (2016). We run the analysis in MrBayes 3.2 program (Ronquist et al., 2012) using the MrBayes formatted nexus files with all analytical parameters provided by Solé et al. (2021). This nexus file is included in the Supplementary materials.
Class Mammalia (Linnaeus, 1758)
Infraclass Eutheria (Gill, 1872)
Superoder Ferae (Linnaeus, 1758)
Order Hyaenodonta (Van Valen, 1967)
Superfamily Hyainailouroidea (Pilgrim, 1932)
Family Hyainailouridae (Pilgrim, 1932). Genus Maocyon, gen. Nov.
Maocyon peregrinus sp. nov.
urn:lsid:zoobank.org:act:66BEBEAA-87C6-49C4-92C2-41DA70748FB4.
As for the type and only species.
From Maoming City and Cyon (Noun, masculine), Latinized form of Greek κύων, a dog.
Maocyon peregrinus sp. nov.
SYSU-M-5, a partial skull with associated mandible.
SYSU-M, Mammal fossil collection from the Maoming Basin in the School of Life Sciences, Sun Yatsen University, Guangzhou, China.
The oil shale quarry (21°42’ N, 110°53′ E) located near Maoming City, Maoming Basin, Guangdong Province, China; Youganwo Formation, upper Eocene.
Referred to the Hyainailouroidea by anteroposteriorly extended suture of the jugal/squamosal, presence of a preglenoid crest, a lateral expansion of the squamosal posterior to the zygomatic arch, a transversally expanded mastoid process, a nuchal crest that does not extend laterally to mastoid process, and large occipital condyles (diagnostic characters are after Solé et al. (2015)). Differs from all Hyainailouroidea except Orienspterodon by retention of a rudimentary (ridge-like) metaconid on lower molars. Differs from Orienspterodon (Chow, 1975; Egi et al., 2007) by more rudimentary metaconid on m3 and dorsoventrally shallower mandibular body.
From Latin peregrīnus (Adjective, masculine), foreign, alien, or exotic, an allusion to the Asiatic provenance of this taxon belonging to the predominantly Afro-European clade.
SYSU-M-5 represents the posterior portion of the skull associated with posterior parts of the left and right dentaries (Figures 2–6). The ventral part of the orbit and zygoma are preserved on the right side (Figures 2, 3, 5) while only posterior part of the zygoma is present on the left side (Figures 2–4). The left dentary is missing anterior to m1 (Figure 4) and the right dentary is broken at the p4 (Figure 5). The palate and anterior part of the basicranium is unprepared and obscured by the attached dentaries (Figure 3). The skull is slightly distorted dorsoventrally. The specimen belongs to an aged individual with heavily worn dentition and obliterated sutures between the skull bones.
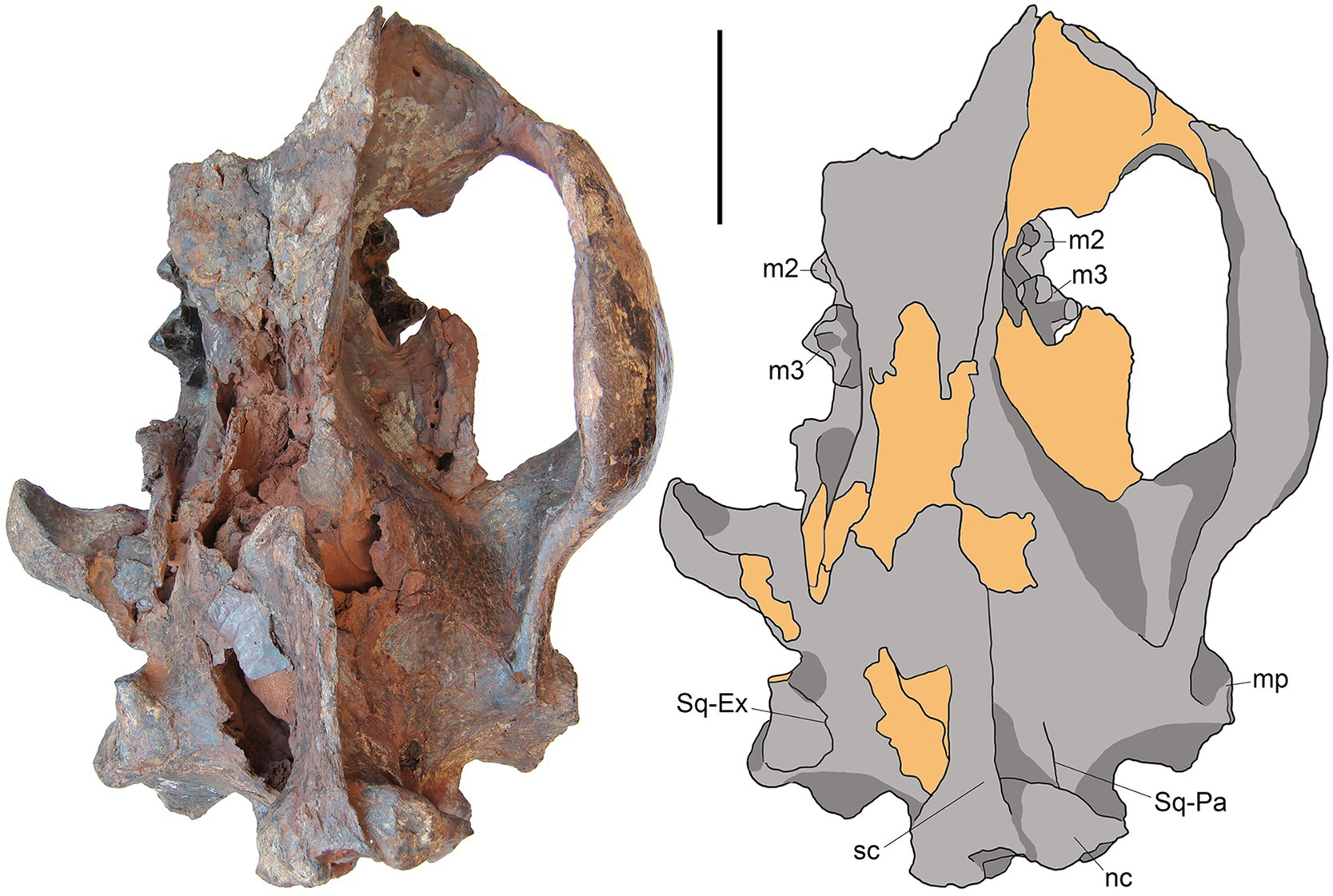
Figure 2. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, a partial skull with associated mandibles in dorsal view (photograph and explanatory drawing). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. mp, mastoid process; nc, nuchal crest; sc, sagittal crest; Sq-Ex, squamosal-exoccipital suture; Sq-Pa, squamosal-parietal suture. On this and following figures, the broken area and matrix are designated by the yellow color. Scale bar equals 5 cm.
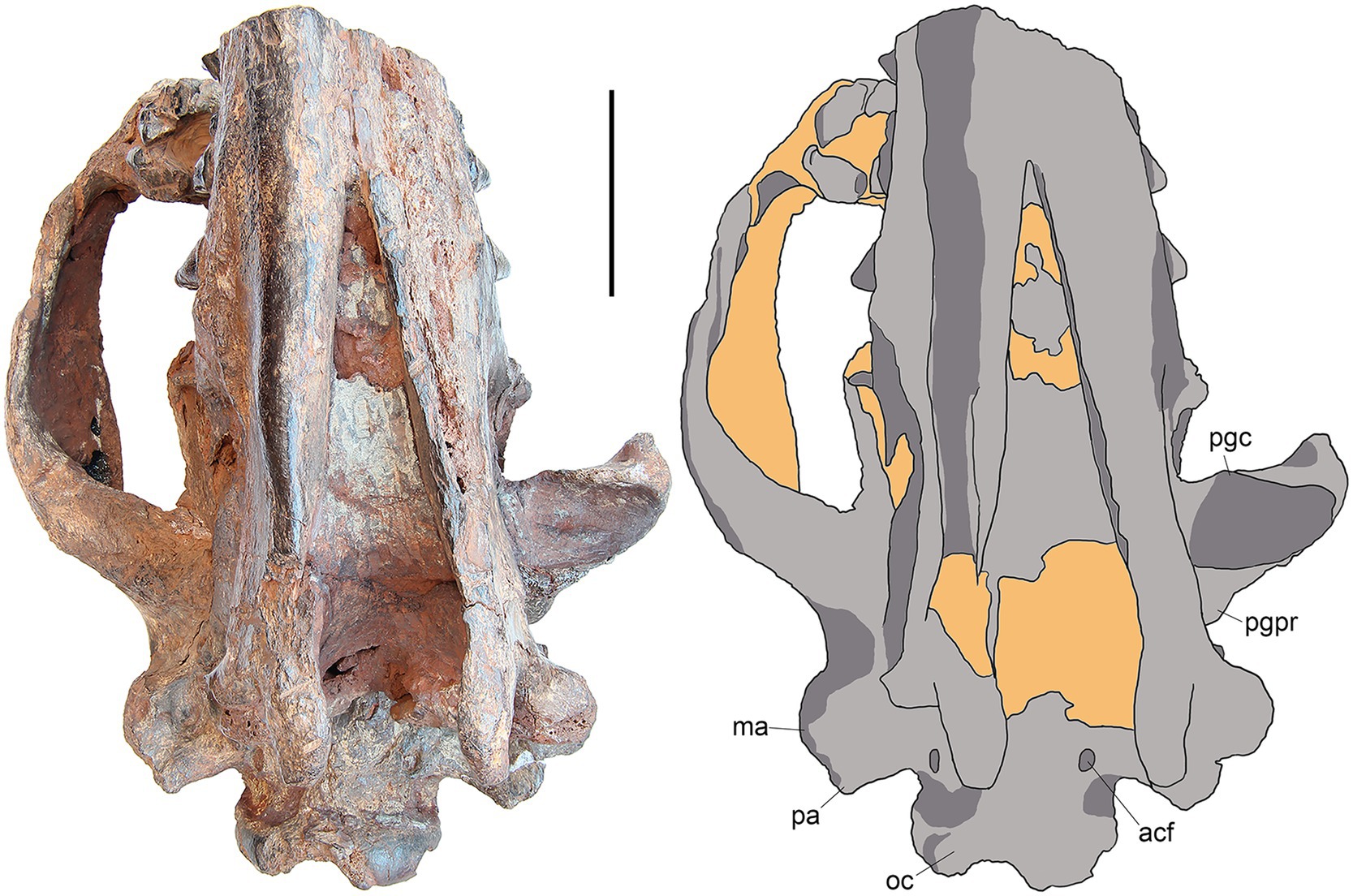
Figure 3. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, a partial skull with associated mandibles in ventral view (photograph and explanatory drawing). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. acf, anterior condyloid foramen; ma, mastoid apophysis; oc, occipital condyle; pa, paroccipital apophysis; pgc, preglenoid crest; pgpr, postglenoid process. Scale bar equals 5 cm.
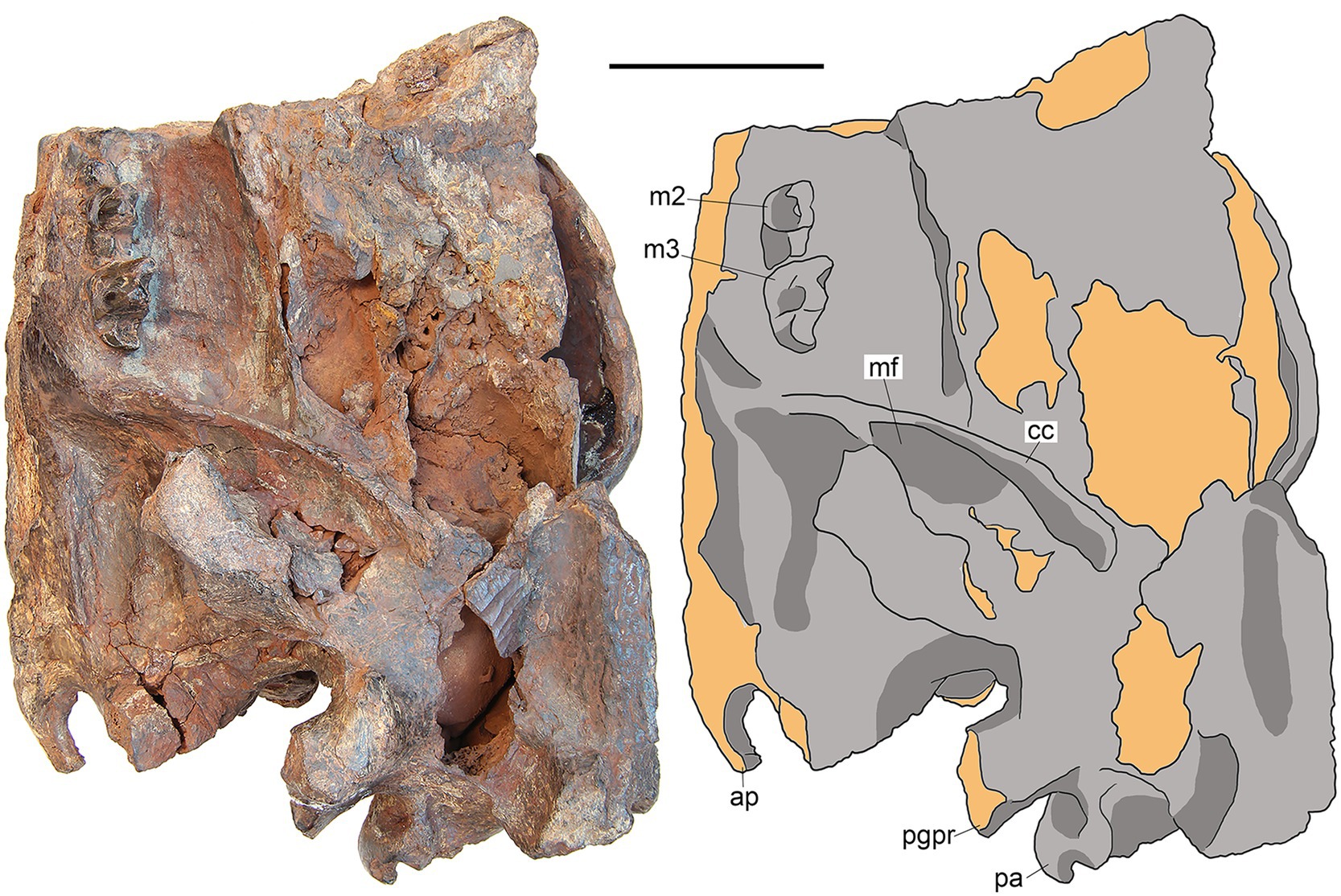
Figure 4. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, a partial skull with associated mandibles in left lateral view (photograph and explanatory drawing). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. ap, angular process; cc, coronoid crest; mf, masseteric fossa; pa, paroccipital apophysis; pgpr, postglenoid process. Scale bar equals 5 cm.
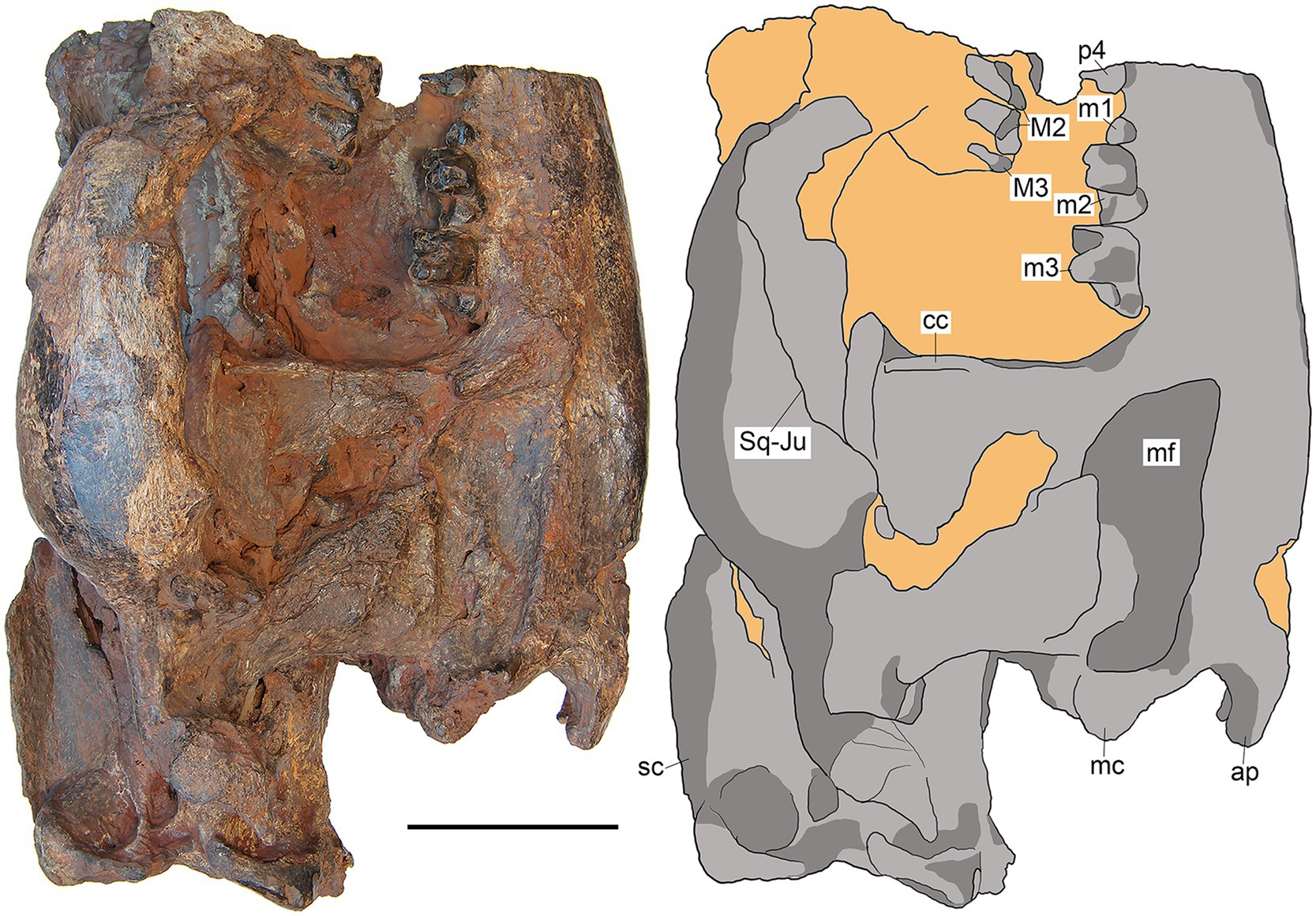
Figure 5. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, a partial skull with associated mandibles in right lateral view (photograph and explanatory drawing). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. ap, angular process; cc, coronoid crest; mc, mandibular condyle; mf, masseteric fossa; sc, sagittal crest; Sq-Ju, squamosal-jugal suture. Scale bar equals 5 cm.
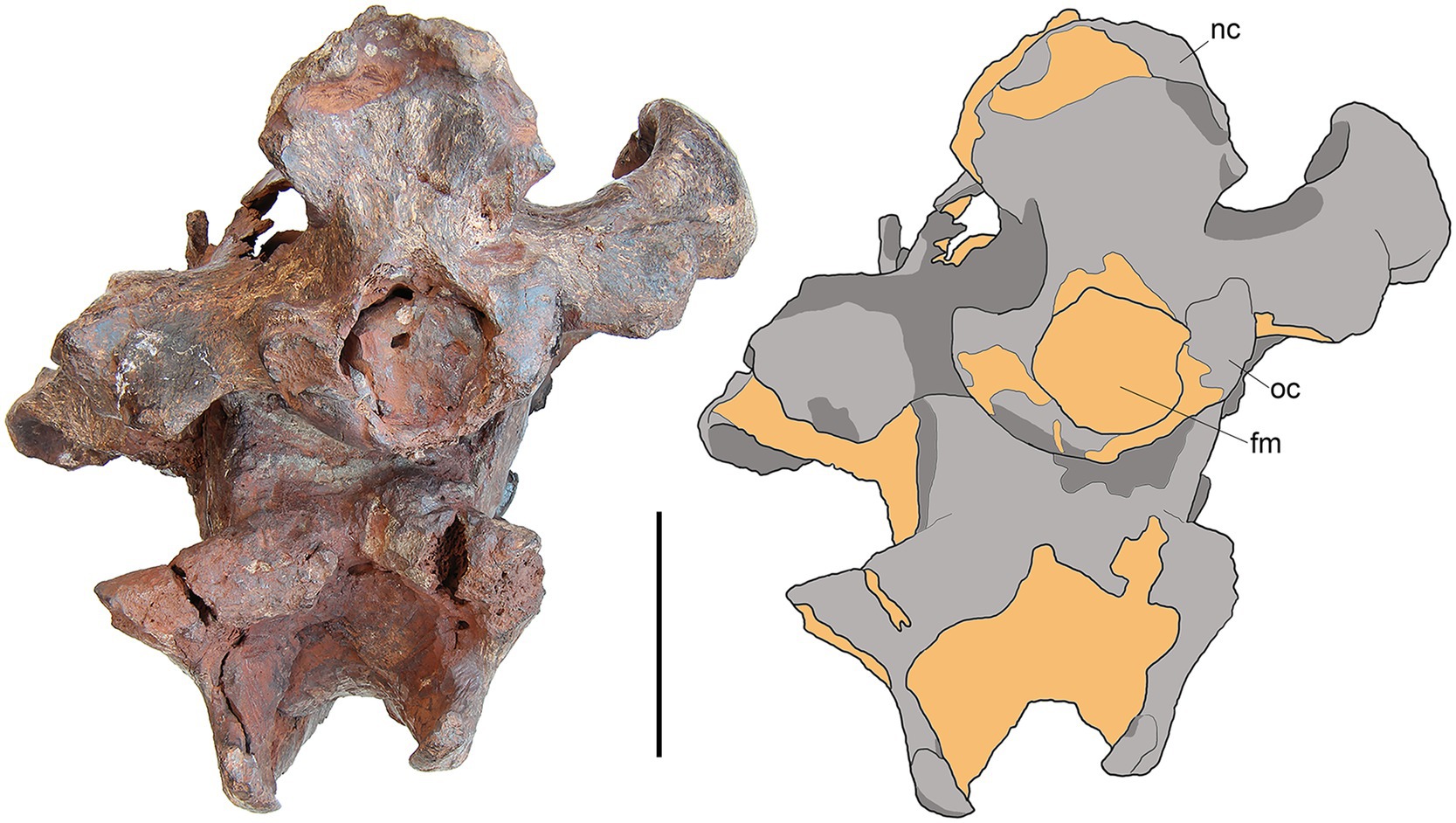
Figure 6. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, a partial skull with associated mandibles in posterior view (photograph and explanatory drawing). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. fm, foramen magnum; oc, occipital condyle; nc, nuchal crest. Scale bar equals 5 cm.
The skull has an anterioposteriorly elongated basicranial and ethmoidal regions, as in oldest hyainailourines such as Pterodon and Kerberos (Solé et al., 2015). The whole length of the skull could be around 30 cm based on proportions of Kerberos langebadreae (Solé et al., 2015). The narrowest part of the braincase is distinctly posterior to the missing postorbital process (Figure 2), as is typical for the Hyainailouridae (Solé et al., 2015).
Only posterior part of maxilla is preserved. The left maxilla fragment with M2-3 was separated from the skull during preparation (Figure 7). On the right maxilla, also preserving M2-3, there is the zygomatic process of the maxilla connecting with the jugal (Figure 5), but the maxillary-jugal suture is not discernable. The ventral border of orbit is horizontal.
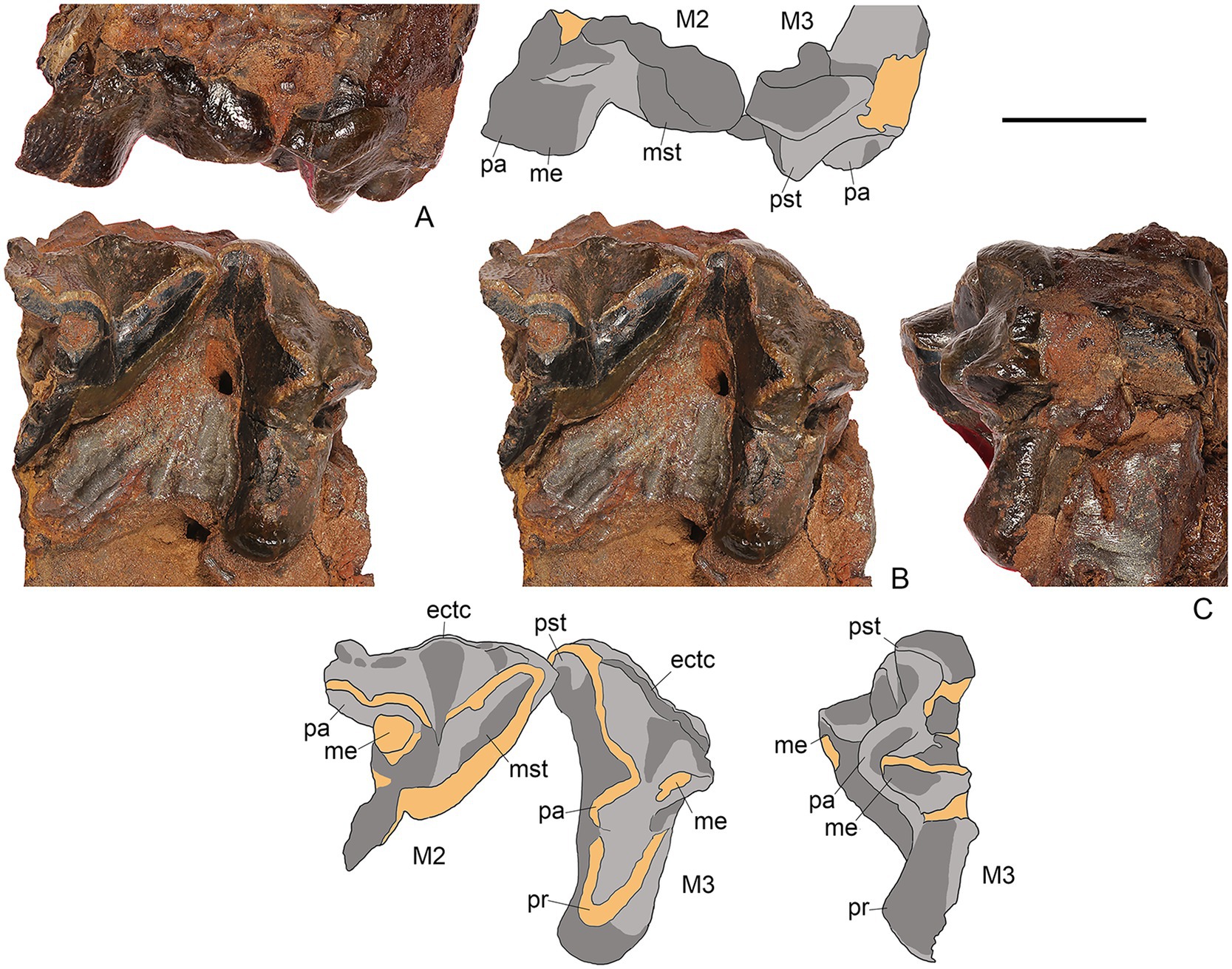
Figure 7. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, left maxilla with M2-3, in labial (A), occlusal [(B) stereopair], and posterior (C) views (photographs and explanatory drawings). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. ectc, ectocingulum; me, metacone; mst, metastyle; pa, paracone; pr, protocone; pst, parastyle. Scale bar equals 1 cm.
The zygomatic arch is dorsoventrally high, increasing in height posteriorly (Figure 5). A minute postorbital process is present along the dorsal margin of the zygoma at the jugal-squamosal junction. Thus the orbit is largely open at least ventrally (the dorsal border of the orbit is not preserved). The jugal-squamosal suture is anteroposteriorly extended between the postorbital process and the preglenoid process (Figure 5), as in other hyainailourides (Solé et al., 2015).
On the lateral side of the braincase the fronto-parietal suture is not discernable. The early obliteration of the fronto-parietal suture is a typical feature for the Hyainailouridae (Solé et al., 2015). On the parietal, the sagittal crest is high, approximately 20 mm in its maximum height, which is about one third of the skull height in this part (Figures 2, 5). The maximum height of the sagittal crest is opposite the paroccipital apophysis where the parietals are ventrally depressed. The dorsal profile of the sagittal crest is somewhat convex.
The squamosal forms the mandibular glenoid with pre- and postglenoid processes (Figures 2, 3). The preglenoid process is in the form of a distinct crest along the anterior border of the mandibular glenoid (Figure 3), as in other Hyainailouridae (Solé et al., 2015). The mediolateral width of the glenoid fossa is more than twice greater than its anteroposterior length. The postglenoid process, better preserved on the left side (Figures 3, 4), deepens ventrally in medial direction. Its distalmost part is missing and the preserved portion shows no curvature in anterior direction. As preserved, the preglenoid process is dorsoventrally higher than the postglenoid process. The lateral margin of the squamosal between the postglenoid and mastoid processes is everted dorsally and medially. Posterior to the zygomatic arch the squamosal is mediolaterally expanded, reaching close to the sagittal crest (Figure 2). The latter condition is characteristic for the Hyainailouridae (Solé et al., 2015). The mastoid process is extensive, with the squamosal-exoccipital suture clearly visible on the dorsal side (Figure 2). The knob-like mastoid apophysis is projecting laterally (Figure 3), a typical character of the Hyainailouridae (Solé et al., 2015). A thin plate-like paroccipital apophysis is posterolaterally directed (Figures 3, 4).
On the occiput, the nuchal crests are short and do not reach the mastoid processes (Figures 2, 6), as typical for the Hyainailouridae (Solé et al., 2015). The nuchal crests converge ventrally and terminate above the occipital condyles. The supraoccipital forming the posterior surface of the nuchal crest is depressed dorsal to the foramen magnum. The large occipital condyles are lateral and ventral to the foramen magnum (Figure 6). This condition is characteristic for the Hyainailouridae (Solé et al., 2015). The ventral margin of the foramen magnum is indented anteriorly.
On the ventral side of the basicranium, the anterior condyloid foramen (=anterior opening of the hypoglossal canal) is placed at the base of the paroccipital apophysis (Figure 3).
Only M2-3 are preserved from the upper dentition; they are better preserved on the left side, where, however, M2 is anteriorly incomplete (Figure 7). On M2, the paracone and metacone are fused into an amphicone. Both cusps are separated by a shallow groove on the labial side. The metacone is ovoid in cross-section, slightly compressed labiolingually. The metastyle is long and blade-like, directed distolabially. It is distinctly longer than the postmetacrista and separated from the latter by the carnassial notch. The ectoflexus of M2 is shallow. The occlusal surface of the paracone and the metacone are heavily worn. The M3 has a long parastylar region and the preparacrista. The metacone is large, only slightly smaller than the paracone. The trigon is transversely long and mesiodistally short. The protocone apex is worn. There are faint lingual and distal cingula. All mesial side of M3 is heavily worn, with a distinct groove in the parastylar region. Both M2 and M3 have a rather strong ectocingulum.
The preserved part of the mandibular body tapers anteriorly, with the minimal height at p4 (Figure 5). The ventral margin of the mandibular body is straight. The coronoid process is an extensive thin plate with the anterior margin nearly perpendicular to the alveolar plane (Figure 5). The posterior margin of the coronoid process is distinctly convex. The mandibular condyle is massive, with the cylindrical and mediolaterally elongate articular surface. It is positioned above the alveolar plane (Figure 4). The angular process is small and hook-like, with the distal end directed posterodorsally (Figures 4, 5). There is no concavity along the ventral margin of the mandibular ramus anterior to the angular process, as in other Hyainailouridae (Solé et al., 2015). The masseteric fossa is deep, delimited anteriorly by a sharp coronoid crest and ventrally by the condyloid crest (Figures 4, 5). The anterior angle of the masseteric fossa is well posterior to m3.
Only posterior part of the right p4 is preserved (Figures 8C,D). This tooth was distinctly larger than m1. The talonid is damaged and it is not clear how large was its hypoconid. Only talonids of m1 are preserved from both sides (Figure 8). However, judging from it and the mesial alveolus size, it is clear that m1 was the smallest lower molar. m3 is the largest lower molar. The paraconid is about twice lower than the protoconid. The metaconid is ridge-like on m2-3. The talonid is simple, single-cusped, but distinctly basined, at least on m3. The talonid of m3 is proportionally shorter mesiodistally compared with m2. At least on m3 there is a distinct vertical keel along the mesiolabial side of the paraconid. There is a distinct labial cingulid on m2-3.
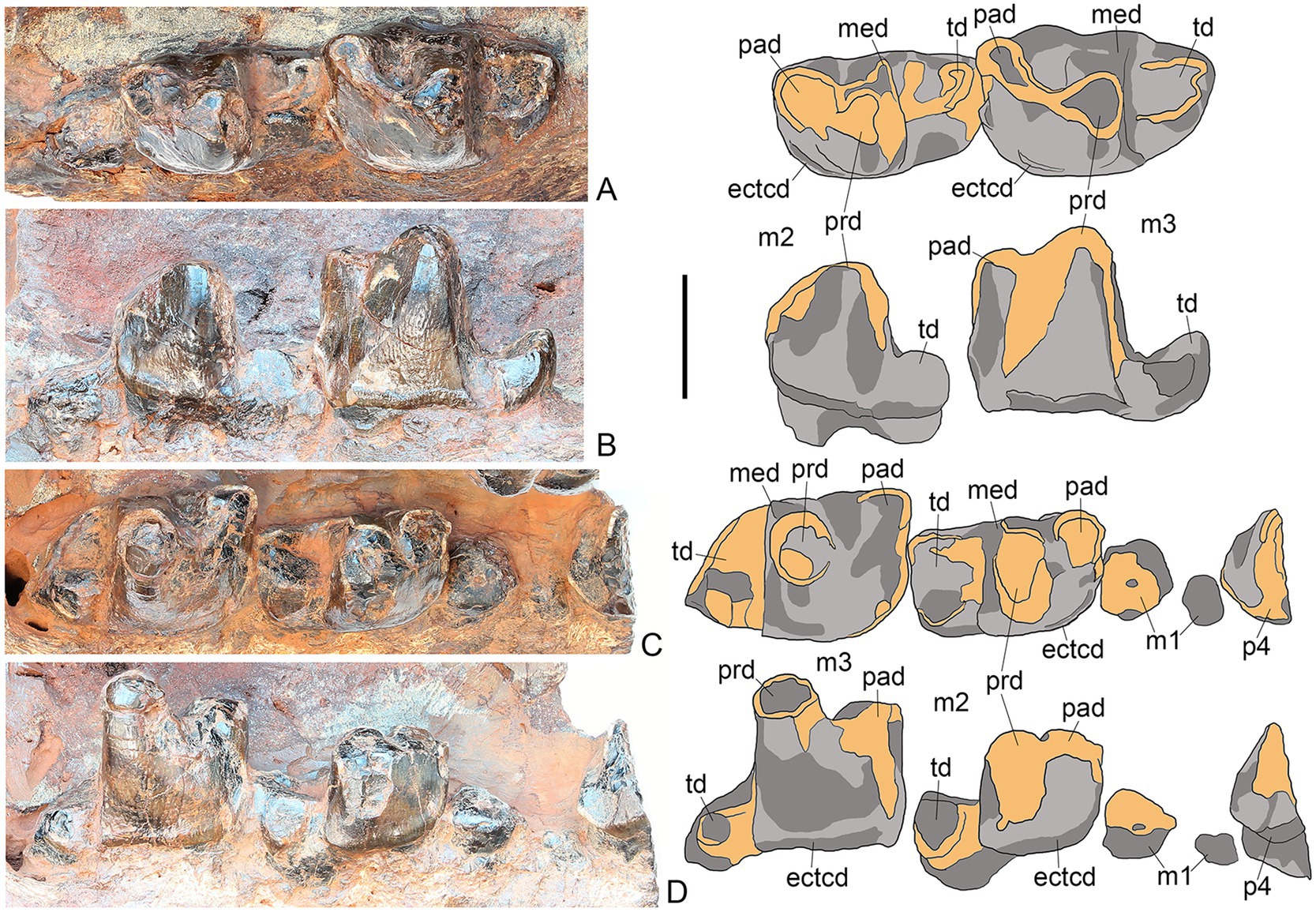
Figure 8. M. peregrinus, gen. et sp. nov., SYSU-M-5, holotype, lower dentition (photographs and explanatory drawings). Youganwo Formation, upper Eocene, Maoming locality, Guangdong Province, China. A, B, left m2-3, in occlusal (A) and labial (B) views. (C,D), right p4, m1-3, in occlusal (C) and labial (D) views. ectcd, ectocingulid; med, metaconid, pad, paraconid; prd, protoconid; td, talonid. Scale bar equals 1 cm.
For dental measurements, see Table 1.
The comparison with Orienspterodon dahkoensis from the late middle Eocene of China and Myanmar (Egi et al., 2007) is limited to the dentary and lower dentition (M2-3 are unknown). Maocyon differs from Orienspterodon by smaller size, shallower mandibular body, and more reduced metaconid on lower molars.
Kerberos langebadreae from the Bartonian of France is known from a complete skull and postcranial skeleton (Solé et al., 2015). The new taxon differs from Kerberos by deeper mandibular body, vertical coronoid process of dentary, more pronounced ectocingulum and larger protocone on upper molars, more separate paracone and metacone on M2, a well developed metacone on M3, and a larger talonid on lower molars.
Maocyon differs from Pterodon dasyuroides from the Priabonian of Western Europe (Lange-Badré, 1979) by anteroposteriorly shorter occiput, lack of occipital crest, separate paracone and metacone on M2-3, much larger M3, presence of metaconid on lower molars, and larger talonid on m2-3.
The gigantic Hemipsalodon grandis from the late Eocene – early Oligocene of North America (Mellett, 1969) is similar with Maocyon by having separate paracone and metacone and pronounced ectocingulum on upper molars, and large M3. It different by its much larger size, deeper mandibular body, reduced talonid and lack of metaconid on lower molars.
The “all compat” (majority rule plus compatible groups) consensus tree produced by tip-dating Bayesian analysis (Figure 9; see Supplementary information for the complete tree) is identical to the tree obtained by Solé et al. (2021) in the interrelationships of the taxa outside the Hyainailouroidea but differs in number of details within this group. The position of Furodon crocheti and Paratritemnodon indicus is more resolved; both taxa are referred to the Teratodontinae which has the same branch support (PP = 29%). Koholia atlasense and Tritemnodon agilis do not form a clade. Orienspterodon dahkoensis is not the basalmost member of the Hyainailourinae, but placed in a polytomy with Hyainailourinae and Apterodontinae. M. peregrinus is found as a sister taxon to Orienspterodon dahkoensis. Compared with the cladogram obtained by Solé et al. (2021), our analysis reveals a better branch support for the Hyainailouroidea (PP = 55 versus 39%), Hyainailouridae (PP = 35 versus 30%), and Hyainailourinae (PP = 51 versus 26%).
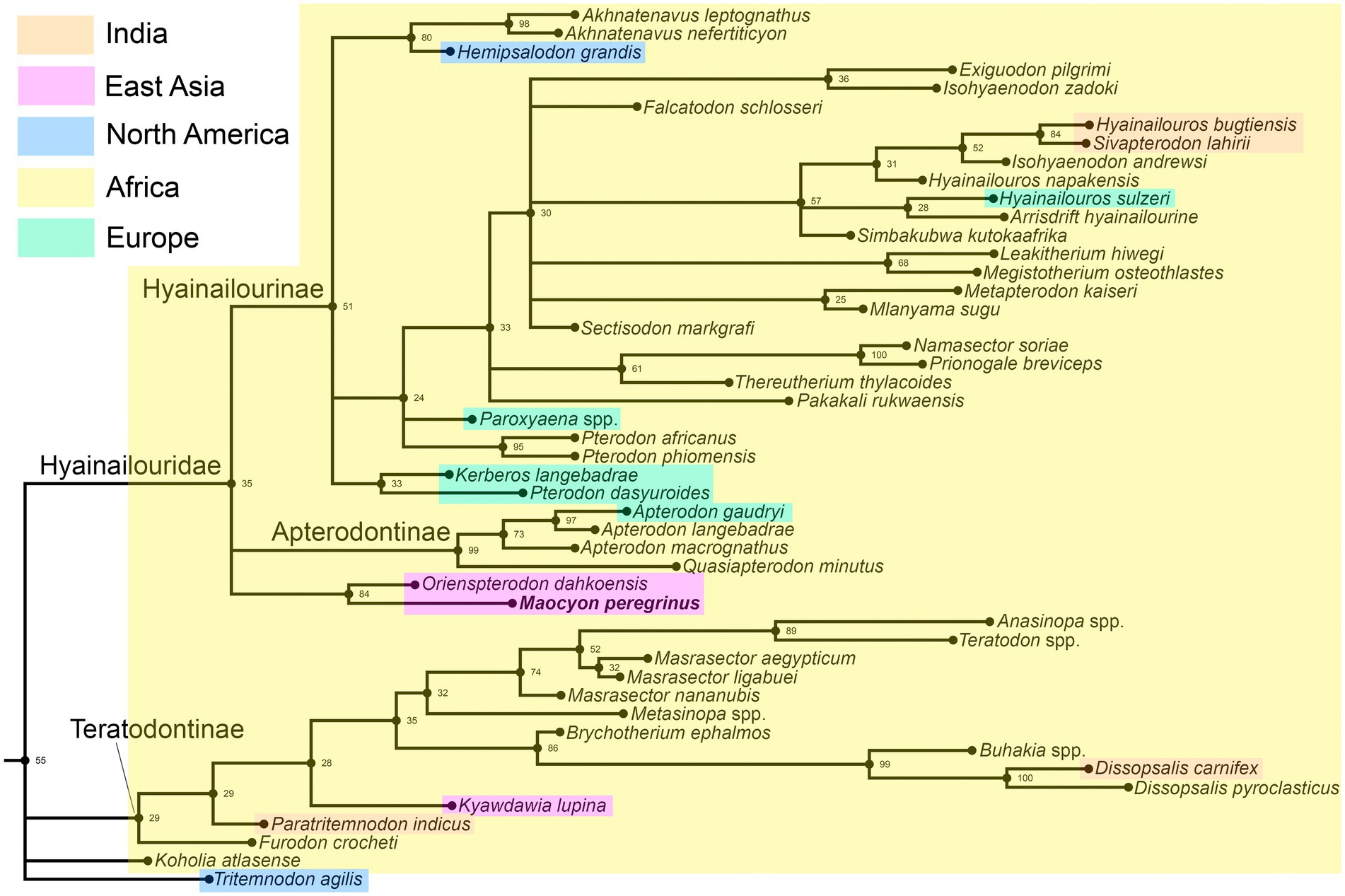
Figure 9. Segment of the “all compat” (majority rule plus compatible groups) consensus tree produced by tip-dating Bayesian analysis showing interrelationships within the Hyainailouroidea (see Supplementary material for the complete tree). Posterior probability (PP) shown at the relevant node.
The previously known members of the Hyainailouridae are large creodonts with the body mass 46–98 kg (Solé et al., 2015). The estimated body mass of M. peregrinus (39 kg) is distinctly smaller. It is less than half that of its older sister taxon Orienspterodon dahkoensis (90 kg). According to Carbone et al. (2007), carnivores larger than 20 kg generally hunt prey greater than or equal to their own body mass.
Most taxa of the Hyainailouroidea were distributed in Africa, with few species known from Europe, East Asia, India, and North America (Figure 9). Orienspterodon and Maocyon are the only East Asian taxa referred to the Hyainailouridae (Figure 9). Orienspterodon is currently the oldest known member of that clade (late middle Eocene). The Bayesian “tip-dating” phylogenetic methods estimates the origin of the clade Orienspterodon + Maocyon as 43.68 Ma. However, estimation of the time of divergence of the Hyainailourinae is slightly older (44.35 Ma). This advocates for the origin of the Hyainailourinae in Africa and dispersal of the ancestors of the Orienspterodon-Maocyon clade from Africa, as was previously suggested (Borths and Stevens, 2019).
Orienspterodon dahkoensis is known by fragmentary specimens from three late middle Eocene localities: Rencun Member of the Heti Formation, Henan Province, central China; upper part of the Lumeiyi Formation, Yunnan Province, southern China; and upper part of the Pondaung Formation, Myanmar (Chow, 1975; Egi et al., 2007; Peigné et al., 2007). The age of the Youganwo Formation, producing M. peregrinus, is likely basal late Eocene (Averianov et al., 2019). A somewhat more derived nature of Maocyon compared with Orienspterodon, expressed by more reduced metaconid on lower molars, is consistent with its younger geological age.
The upper Eocene Youganwo Formation at Maoming locality produces abundant remains of predominantly aquatic or semiaquatic vertebrates (Table 2). Among mammals, the remains of groups that preferred mesic habitats, like amynodontid perissodactyls and anthracotheriid artiodactyls, are most common. The carnivorous mammals are extremely rare in this locality, being known previously only from a single specimen of the nimravid Maofelis cantonensis (Averianov et al., 2016). The holotype of the creodont M. peregrinus described in this paper is only the second specimen of carnivorous mammals from this fauna.
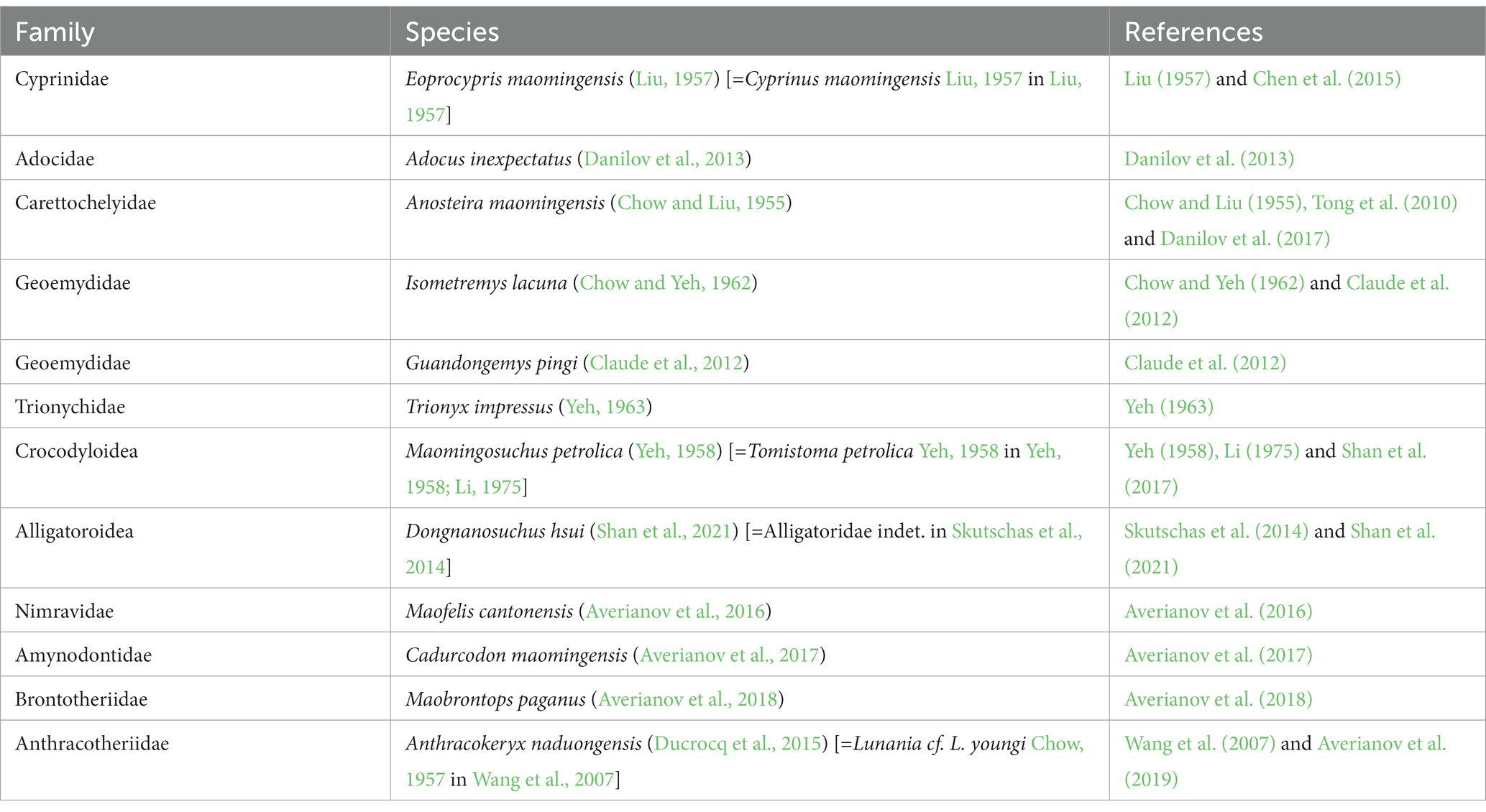
Table 2. List of the vertebrates from the upper Eocene Youganwo Formation at Maoming locality, Guangdong Province, China.
The new taxon described in this paper (Maocyon peregrinus) has been registered on ZooBank: https://zoobank.org/, 66BEBEAA-87C6-49C4-92C2-41DA70748FB4.
AA designed the research, performed phylogenetic analysis, wrote the paper, and prepared illustrations and Tables 1, 2. EO cleaned the specimen from matrix. ID prepared photographs, read and approved manuscript. J-HJ conceived and acquired funding, read and approved manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (nos. 41820104002 and 42111530024). The work of AA was supported by the Zoological Institute, Russian Academy of Sciences (project 122031100282-2).
We thank Floréal Solé and two reviewers for reading the text and comments.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1076819/full#supplementary-material
Averianov, A. O., Danilov, I. G., Chen, W., and Jin, J. (2018). A new brontothere from the Eocene of South China. Acta Palaeontol. Pol. 63, 189–196. doi: 10.4202/app.00431.2017
Averianov, A. O., Danilov, I. G., Jin, J., and Wang, Y. (2017). A new amynodontid from the Eocene of South China and phylogeny of Amynodontidae (Perissodactyla: Rhinocerotoidea). J. Syst. Palaeont. 15, 927–945. doi: 10.1080/14772019.2016.1256914
Averianov, A. O., Obraztsova, E. M., Danilov, I. G., and Jin, J. (2019). Anthracotheriid artiodactyl Anthracokeryx and an upper Eocene age for the Youganwo formation of southern China. Hist. Biol. 31, 1–8. doi: 10.1080/08912963.2017.1421639
Averianov, A. O., Obraztsova, E. M., Danilov, I. G., Skutschas, P. P., and Jin, J. (2016). First nimravid skull from Asia. Sci. Rep. 6:25812. doi: 10.1038/srep25812
Borths, M. R., Holroyd, P. A., and Seiffert, E. R. (2016). Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ. 4:e2639. doi: 10.7717/peerj.2639
Borths, M. R., and Seiffert, E. R. (2017). Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS One 12:e0173527. doi: 10.1371/journal.pone.0173527
Borths, M. R., and Stevens, N. J. (2017). The first hyaenodont from the late Oligocene Nsungwe formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition. PLoS One 12:e0185301. doi: 10.1371/journal.pone.0185301
Borths, M. R., and Stevens, N. J. (2019). Simbakubwa kutokaafrika, gen. Et sp. nov. (Hyainailourinae, Hyaenodonta, ‘Creodonta,’ Mammalia), a gigantic carnivore from the earliest Miocene of Kenya. J. Vert. Paleont. 39:e1570222. doi: 10.1080/02724634.2019.1570222
Carbone, C., Teacher, A., and Rowcliffe, J. M. (2007). The costs of carnivory. PLoS Biol. 5:e22. doi: 10.1371/journal.pbio.0050022
Chen, G. J., Chang, M.-M., and Liu, H. Z. (2015). Revision of Cyprinus maomingensis Liu, 1957 and the first discovery of Procypris-like cyprinid (Teleostei, Pisces) from the late Eocene of South China. Sci. China Earth Sci. 58, 1123–1132. doi: 10.1007/s11430-015-5085-7
Chow, M.-C. (1957). On some Eocene and Oligocene mammals from Kwangsi and Yunnan. Vertebr. PalAsiatica 1, 201–214.
Chow, M.-C., and Liu, C.-L. (1955). A new anosterine turtle from Maoming, Kwangtung. Acta Palaeont. Sin. 3, 275–282.
Chow, M.-C., and Yeh, H.-K. (1962). A new emydid from Eocene of Maoming, Kwangtung. Vertebr. PalAsiatica 6, 225–229.
Claude, J., Zhang, J.-Y., Li, J.-J., Mo, J.-Y., Kuang, X.-W., and Tong, H. (2012). Geoemydid turtles from the late Eocene Maoming Basin, Southern China. Bull. Soc. Géol. France 183, 641–651. doi: 10.2113/gssgfbull.183.6.641
Danilov, I. G., Obraztsova, E. M., Chen, W., and Jin, J. (2017). The cranial morphology of Anosteira maomingensis (Testudines, pan-Carettochelys) and the evolution of pan-carettochelyid turtles. J. Vert. Paleont. 37:e1335735. doi: 10.1080/02724634.2017.1335735
Danilov, I. G., Syromyatnikova, E. V., Skutschas, P. P., Kodrul, T. T., and Jin, J. (2013). The first ‘true’ Adocus (Testudines, Adocidae) from the Paleogene of Asia. J. Vert. Paleont. 33, 1071–1080. doi: 10.1080/02724634.2013.768254
Ducrocq, S., Benammi, M., Chavasseau, O., Chaimanee, Y., Suraprasit, K., Pha, D. P., et al. (2015). New anthracotheres (Cetartiodactyla, Mammalia) from the Paleogene of northeastern Vietnam: biochronological implications. J. Vert. Paleontol. 35:e929139. doi: 10.1080/02724634.2014.929139
Egi, N., Tsubamoto, T., and Takai, F. (2007). Systematic status of Asian "Pterodon" and early evolution of hyaenaelurine hyaenodontid creodonts. J. Paleontol. 81, 770–778. doi: 10.1666/pleo0022-3360(2007)081[0770:SSOAPA]2.0.CO;2
Gheerbrant, E. (1995). Les mammifères paléocenes du Bassin d’Ouarzazate (Maroc). III. Adapisoriculidae et autres mammifères (Carnivora, ?Creodonta, Condylarthra, ?Ungulata et incertae sedis). Palaeontogr. Abt. A 237, 39–132. doi: 10.1127/pala/237/1995/39
Gill, T. (1872). Arrangement of the families of mammals. With analytical tables. Smiths. Misc. Coll. 11, 1–98.
Ginsburg, L. (1980). Hyainailouros sulzeri, mammifère créodonte du Miocène d’Europe. Ann. de Paléont. (Vert.) 66, 19–73.
Holroyd, P. A. (1999). New Pterodontinae (Creodonta: Hyaenodontidae) from the late Eocene-early Oligocene Jebel Qatrani formation, Fayum Province. Egypt. PaleoBios. 19, 1–18.
Lange-Badré, B. (1979). Les Créodontes (Mammalia) d’Europe occidentale de l’Éocéne supérieur à l’Oligocéne supérieur. Mém. Mus. Nat. D'hist. Nat. Sér. C. 42, 1–249.
Lewis, M. E., and Morlo, M. (2010). “Creodonta” in Cenozoic mammals of Africa. eds. L. Werdelin and W. J. Sanders (Berkeley, Los Angeles, London: University of California Press), 543–560.
Li, J. L. (1975). A new material on Tomistoma petrolica of Maoming, Kwantung. Vertebr. PalAsiatica 13, 190–191.
Linnaeus, C. (1758). Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis Regnum Animale, vol. 1. Editio decima, reformata ed (Stockholm: Laurentius Salvius)
Liu, H.-T. (1957). A new fossil cyprinid fish from Maoming, Kwangtung. Vertebr. PalAsiatica 1, 151–153.
Mellett, J. S. (1969). A skull of Hemipsalodon (Mammalia, Deltatheridia) from the Clarno formation of Oregon. Am. Mus. Novitates. 2387, 1–19.
Meng, J., Zhai, R., and Wyss, A. R. (1998). The late Paleocene Bayan Ulan fauna of Inner Mongolia, China. Bull. Carnegie Mus. Nat. Hist. 34, 148–185.
Morales, J., and Pickford, M. (2017). New hyaenodonts (ferae, Mammalia) from the early Miocene of Napak (Uganda), koru (Kenya) and Grillental (Namibia). Foss. Imprint. 73, 332–359. doi: 10.2478/if-2017-0019
Morlo, M., Miller, E. R., and El-Barkooky, A. N. (2007). Creodonta and Carnivora from Wadi Moghra, Egypt. J. Vert. Paleont. 27, 145–159. doi: 10.1671/0272-4634(2007)27[145:CACFWM]2.0.CO;2
Peigné, S., Morlo, M., Chaimanee, Y., Ducrocq, S., Tun, S. T., and Jaeger, J.-J. (2007). New discoveries of hyaenodontids (Creodonta, Mammalia) from the Pondaung formation, middle Eocene, Myanmar – paleobiogeographic implications. Geodiversitas. 29, 441–458.
Polly, D. P. (1996). The skeleton of Gazinocyon vulpeculus gen. Et comb. nov. and the cladistic relationships of Hyaenodontidae (Eutheria, Mammalia). J. Vert. Paleont. 16, 303–319. doi: 10.1080/02724634.1996.10011318
Rana, R. S., Kumar, K., Zack, S. P., Solé, F., Rose, K. D., Missiaen, P., et al. (2015). Craniodental and postcranial morphology of Indohyaenodon raoi from the early eocene of India, and its implications for ecology, phylogeny, and biogeography of hyaenodontid mammals. J. Vert. Paleont. 35:e965308. doi: 10.1080/02724634.2015.965308
Rasmussen, D. T., and Gutiérrez, M. (2009). A mammalian fauna from the late Oligocene of northwestern Kenya. Palaeontogr. Abt. A 288, 1–52. doi: 10.1127/pala/288/2009/1
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhn, A. S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Savage, R. J. G. (1965). Fossil mammals of Africa: 19. The Miocene Carnivora of East Africa. Bull. Br. Mus. (Nat. Hist.) Geol. 10, 242–316.
Savage, R. J. G. (1973). Megistotherium, gigantic hyaenodont from Miocene of Gebel Zelten, Libya. Bull. Br. Mus. (Nat. Hist.) Geol. 22, 484–511.
Shan, H.-Y., Wu, X.-C., Cheng, Y.-N., and Sato, T. (2017). Maomingosuchus petrolica, a restudy of ‘Tomistoma’ petrolica Yeh, 1958. Palaeoworld 26, 672–690. doi: 10.1016/j.palwor.2017.03.006
Shan, H.-Y., Wu, X.-C., Sato, T., Cheng, Y.-N., and Rufolo, S. (2021). A new alligatoroid (Eusuchia, Crocodylia) from the Eocene of China and its implications for the relationships of Orientalosuchina. J. Paleontol. 95, 1321–1339. doi: 10.1017/jpa.2021.69
Skutschas, P. P., Danilov, I. G., Kodrul, T. T., and Jin, J. (2014). The first discovery of an alligatorid (Crocodylia, Alligatoroidea, Alligatoridae) in the Eocene of China. J. Vert. Paleont. 32, 471–476. doi: 10.1080/02724634.2013.809725
Solé, F., Amson, E., Borths, M. R., Vidalenc, D., Morlo, M., and Bastl, K. (2015). A new large hyainailourine from the Bartonian of Europe and its bearings on the evolution and ecology of massive hyaenodonts (Mammalia). PLoS One 10:e0135698. doi: 10.1371/journal.pone.0135698
Solé, F., Gheerbrant, E., Amaghzaz, M., and Bouya, B. (2009). Further evidence of the African antiquity of hyaenodontid (“Creodonta”, Mammalia) evolution. Zool. J. Linnean Soc. 156, 827–846. doi: 10.1111/j.1096-3642.2008.00501.x
Solé, F., Lhuillier, J., Adacid, M., Bensalahd, M., Mahboubi, M., and Tabuce, R. (2014). The hyaenodontids from the Gour Lazib area (?Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae. J. Syst. Palaeont. 12, 303–322. doi: 10.1080/14772019.2013.795196
Solé, F., Morlo, M., Schaal, T., and Lehmann, T. (2021). New hyaenodonts (Mammalia) from the late Ypresian locality of Prémontré (France) support a radiation of the hyaenodonts in Europe already at the end of the early Eocene. Geobios 66-67, 119–141. doi: 10.1016/j.geobios.2021.02.004
Tong, H., Zhang, J.-Y., and Li, J.-J. (2010). Anosteira maomingensis (Testudines: Carettochelyidae) from the late Eocene of Maoming, Guangdong, southern China: new material and re-description. Neues Jahrb. Geol. Palaontol. Abh. 256, 279–290. doi: 10.1127/0077-7749/2010/0053
Van Valen, L. M. (1967). New Paleocene insectivores and insectivore classification. Bull. Am. Mus. Nat. Hist. 135, 217–284.
von Stromer, E. (1926). Reste Land-und Süsswasser-Bewohnender Wirbeltiere aus den Diamantfeldern Die Diamantenwuste Südwest-Afrikas. Vol. 2 (Berlin: E. Kaiser), 107–153.
Wang, Y.-Y., Zhang, Z.-H., and Jin, J. (2007). Discovery of Eocene fossil mammal from Maoming Basin, Guangdong. Acta Sci. Nat. Univ. Sunyatseni. 46, 131–133.
Keywords: Mammalia, Hyaenodonta, Hyainailouroidea, Eocene, Asia
Citation: Averianov A, Obraztsova E, Danilov I and Jin J-H (2023) A new hypercarnivorous hyaenodont from the Eocene of South China. Front. Ecol. Evol. 11:1076819. doi: 10.3389/fevo.2023.1076819
Edited by:
Grégoire Métais, Centre National de la Recherche Scientifique, FranceReviewed by:
Yuan-Qing Wang, Institute of Vertebrate Paleontology and Paleoanthropology (CAS), ChinaCopyright © 2023 Averianov, Obraztsova, Danilov and Jin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander Averianov, ✉ ZHpoYXJha3VkdWtAbWFpbC5ydQ==; Jian-Hua Jin, ✉ bHNzampoQG1haWwuc3lzdS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.