 Alexis M. Mychajliw1,2*
Alexis M. Mychajliw1,2* Audrey Y. Hsi1,3
Audrey Y. Hsi1,3 Drew An-Pham1,4Olivia L. Olson1Nanny Carder5John G. Crock5Francis “Jess” W. Robinson6
Drew An-Pham1,4Olivia L. Olson1Nanny Carder5John G. Crock5Francis “Jess” W. Robinson6- 1Department of Biology, Middlebury College, Middlebury, VT, United States
- 2Environmental Studies Program, Middlebury College, Middlebury, VT, United States
- 3Department of Anthropology, Stanford University, Stanford, CA, United States
- 4Department of Geography, Middlebury College, Middlebury, VT, United States
- 5Consulting Archaeology Program, Department of Anthropology, University of Vermont, Burlington, VT, United States
- 6Vermont Division for Historic Preservation, Montpelier, VT, United States
Shifting baselines can skew species harvest guidelines and lead to potentially inaccurate assessments of population status and range. The North American Fur Trade (~1600–1900 CE) profoundly impacted the continent’s socio-ecological systems, but its legacies are often not incorporated in management discussions. We apply a conservation paleobiology lens to address shifting baselines of nine species of fur-bearing mammals in Vermont, including seven mesocarnivores and two semi-aquatic rodents. Using a database maintained by the Vermont Division for Historic Preservation, we identified 25 existing radiocarbon dates of fur-bearer associated features from 16 archaeological localities spanning the Early-Late Holocene. We also generated 7 new radiocarbon dates on beaver and muskrat bones from the Ewing (VT-CH-005), Bohannon (VT-GI-026), and Chimney Point (VT-AD-329) localities. Our new radiocarbon dates cluster within the Late Holocene, immediately prior to and throughout the European contact period, and overlap with The Beaver Wars. We recover a ~8,000 year record of beaver harvest, affirming the millennial scale importance of beavers, a species that is often the focus of human-wildlife conflict research. Comparison of zooarchaeological occurrences with digitized natural history specimens and community science observations reveals geographic range continuity for most species except for the American marten, which was historically extirpated, and confirms the native status of the red fox. While taphonomic constraints make our dataset a conservative assessment, our case studies demonstrate how wildlife managers can employ zooarchaeological data to better understand long-term properties of coupled socio-ecological systems and highlight the cultural importance of these species to Indigenous trade networks prior to the Fur Trade in Vermont.
1. Introduction
Conservation success is often evaluated by comparing the status of the present with ecological or population targets derived from a particular past condition (e.g., Akçakaya et al., 2018). A focus on isolated time points can lead to inaccurate perceptions of what is “normal” (e.g., a shifting baseline; Pauly, 1995; Silliman et al., 2018; Cammen et al., 2019), but this pitfall can be averted by evaluating data of varying temporal depth and from multiple sources to avoid cultural and disciplinary biases (Rodrigues et al., 2019). Historical photographs, natural history specimens, and written archives have been helpful for reconstructing changes unfolding over the past decades to centuries (McClenachan et al., 2015; Turvey et al., 2015; Collins et al., 2020). However, these historical records reflect landscapes and dynamics that may have already been significantly altered by humans across millennial timescales within the context of broader climatic and environmental changes (Ellis et al., 2021). Conservation biologists and wildlife managers are, therefore, increasingly exploring datasets gleaned from both archaeological and paleontological contexts to understand the longer term ecological and evolutionary trajectories of species of concern (Dietl and Flessa, 2011; Barnosky et al., 2017).
In North America, the harvest of fur-bearing mammals (mammal species typically trapped for fur) is managed today as part of livelihood, recreational, and commercial activities, with targets often derived from data generated by annual trapper effort reports (White et al., 2021). Fur-bearers underwent a massive recent bottleneck as millions of animals were killed for pelts and shipped to European markets during The North American Fur Trade (~1600–1900 CE), resulting in ecological upheaval and cultural dislocation for Indigenous communities (Nassaney, 2015). Historical trading records from the Hudson’s Bay Company, a key fur trading business established in Canada, have revealed baselines shifts in fur-bearer populations, wherein a baseline of 1970 CE shows an increase in select Canadian fur-bearing mammal populations by 4%, whereas examining a baseline of 1850 CE instead yields an overall population decline of 15% (Collins et al., 2020). Most prior research on the impacts of the Fur Trade has centered on such European trade ledgers (see also McManus, 1972; Carlos and Lewis, 1993). However, detailed historical ledgers are not available for all locations of management interest, and commercially harvested animals were prepared as tanned pelts and other derivatives, and thus did not enter natural history collections. Historical archaeology studies have often examined fur trade outposts or Trading Company processing stations with a focus on social and economic interaction spheres (Turgeon, 1998; Veltre and McCartney, 2002), rather than whole ecosystem perspective. The North American Fur Trade itself was built on millennia of harvest and complex trade networks sustained by Indigenous people, who had important relationships with fur-bearing species and shaped their ecology prior to and throughout European contact (Thomas, 1994). Thus, contemporaneous zooarchaeological assemblages can be important windows into ecological change.
We apply a conservation paleobiology lens to identify dimensions of fur-bearer management that could benefit from engagement with archaeological datasets within the larger environmental context of the state of Vermont, United States. Recent surveys of the field of conservation paleobiology highlight the divide between academic research in paleontology and application in conservation and management (Dillon et al., 2022), with only a small percent of peer reviewed conservation paleobiology publications yielding real world impacts (Groff et al., 2022). More broadly, differences in dataset type, format, and compatibility are known barriers in crossing the research-implementation gap in conservation biology (Buxton et al., 2021). We combine radiocarbon dating, technical reports, and a state database resulting from decades of academic and regulatory archaeological activities (Robinson and Ostrum, 2016; Robinson et al., 2020) to determine a conservative estimate of the spatiotemporal span of fur-bearing species. The zooarchaeological record in Vermont is fragmentary, largely because of the regions’ acidic soils, freeze thaw cycles on shallow deposits and a bias of excavation activity toward lowlands (Lacy, 1994; Robinson, 2011). Though taphonomic biases can complicate direct mapping of past and present datasets (Barnosky et al., 2017), inclusion of archaeological datasets in conservation decision-making reflects an understanding of the true timescale of human-faunal-environmental interactions that has structured present day ecosystems (Reeder-Myers et al., 2022).
We use this database of radiocarbon dates, in conjunction with historical and present-day occurrence data, to provide a new synthesis of the Holocene history of fur-bearers and emphasize the role of these species within the dynamic human history of Vermont. Then, we address three outstanding questions surrounding fur-bearer populations in Vermont: (1) did beavers continuously exist throughout the colonial period? (2) what was the past geographic range of the American marten prior to its state-specific extirpation? and, (3) can the zooarchaeological record resolve uncertain classifications of canid species?
2. Methods
2.1. Focal taxa
We focus species currently designated by Vermont Department of Fish & Wildlife (VTFW) as fur-bearers that were also historically harvested as part of the fur trade. These taxa include mesocarnivores (Order Carnivora) across three families, including Mephitidae (striped skunk, Mephitis mephitis), Canidae (red fox, Vulpes vulpes), and Mustelidae (American mink, Neogale vison; river otter, Lontra canadensis; American marten, Martes americana; fisher, Pekania pennanti), and two semi-aquatic rodents, American beaver (Castor canadensis) and muskrat (Ondatra zibethicus). All species are listed under state ranking code S5 (common), except for marten, which is S1 (state endangered) and has no open trapping season. Beaver and muskrat are the most abundantly harvested fur-bearing mammal species in Vermont, together comprising more than half of all individuals trapped from 2020 to 2021 (VTFW, 2021).
2.2. Vermont
Vermont currently recognizes four Western Abenaki tribes; the federally recognized Stockbridge-Munsee Mohican also claim ancestral ties to several counties in southwestern Vermont. The area has a > 12,500 year record of people (families, sensu Newsom, 2022) using the landscape following the retreat of the Laurentide glacier (Robinson et al., 2017). Vermont’s archaeological and paleoecological records have been relatively underleveraged in regional and continent-wide research (e.g., Munoz et al., 2010; Chaput et al., 2015; Kelly et al., 2022). Trade in furs was a major driver in the relationship between Indigenous people and Europeans; the French made alliances and developed trade partners with people living near the St. Lawrence River and Lake Champlain as English rivals settled the Hudson and Connecticut Rivers and allied themselves more closely with Iroquoian populations (Thomas, 1979; Calloway, 1994; Figure 1). The homeland of the Western Abenaki and what is now Vermont was strategically valuable during French and Iroquois conflicts over trading rights, known generally as The Beaver Wars (1641–1701 CE; Williamson, 1949; Nassaney, 2015).
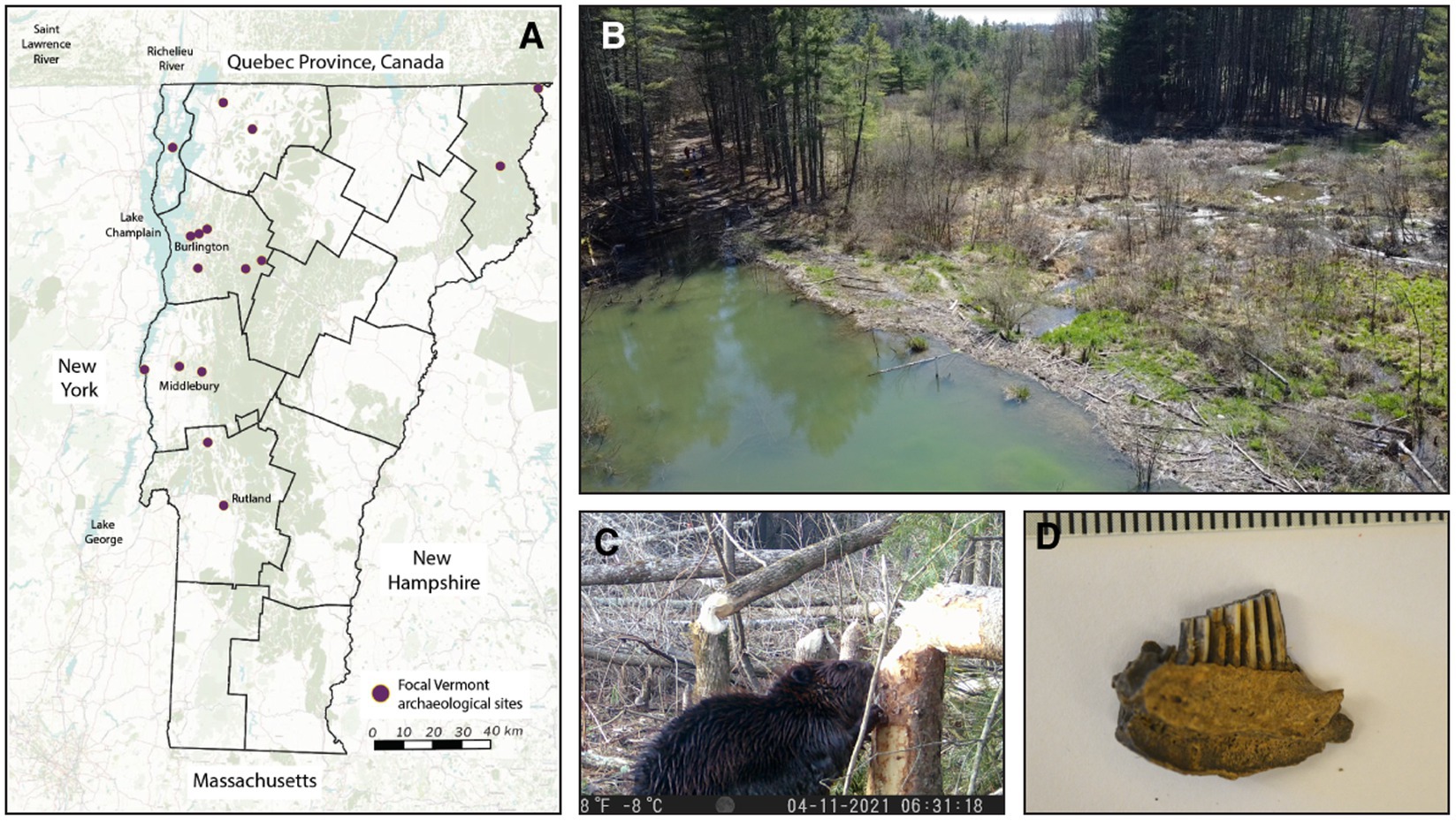
Figure 1. (A) Map of archaeological sites with confirmed fur-bearer skeletal remains used in this study, with points overlaid on an Open Street Map base map where waterways are blue, forests, farms, and agriculture are green, and black lines indicate county boundaries. (B) Drone-based imagery of a hiking trail flooded by beaver activity in Bristol, Vermont, with humans for scale in the top left (photo by Andrew Ng, with permission of The Watershed Center). (C) Camera trap footage of a beaver downing trees in areas with high human densities, such as the Middlebury College campus (photo by Andrew Ng). (D) Muskrat bones, such as the one pictured, are frequently found in archaeological sites; muskrats can also impact local wetland structure and function.
Following European contact and throughout the colonial period and post statehood (1791 CE), Vermont’s environment experienced drastic transformations due to logging and European agricultural practices that created “a world of fields and fences” (Cronon, 1983). Harvest pressure on wild mammals increased for commercial purposes and to protect livestock, resulting in the extirpation of wolves and mountain lions (Klyza and Trombulak, 1999). Compared to an early 19th century colonial baseline, Vermont is in a reforested state due to declines in agriculture and shifting demographics (Foster et al., 2002). However, present and future land use change is predicted to result in further deforestation with development and climate shifts (Pearman-Gillman, 2020).
2.3. Database of Vermont radiocarbon dates and directly associated diagnostic artifacts
Directly dated zooarchaeological assemblages or those with associated age assignments were identified using the Vermont Division for Historic Preservation (VDHP)’s Database of Vermont Radiocarbon Dates and Directly Associated Diagnostic Artifacts (Boulanger, 2007; Robinson and Ostrum, 2016; Robinson et al., 2020; Kelly et al., 2022). This database currently contains >400 radiocarbon dates generated through decades of regulatory work by archaeological consultants, federal and state agencies, and academic institutions. These radiocarbon dates were cross-referenced with VDHP’s Paleobotany & Faunal Database to identify temporal contexts for fur-bearer remains and relevant site features. While both databases are freely available through the VDHP, they have not yet been used in a conservation context. We curated a subset of the database that includes sites with faunal material identified by a zooarchaeologist and those with radiocarbon dates that can be associated with fur-bearer remains. Poor preservation of organic remains from archaeological sites in Vermont results in a large number of unidentifiable small-medium sized mammals that may include fur-bearers. Therefore, we present a conservative view of fur-bearer occurrences in Vermont and do not include sites excavated historically that lack proper identification and/or reliable radiocarbon-based temporal assignments.
2.4. Radiocarbon dating
We selected 10 bones representing semi-aquatic rodents (beaver or muskrat) from zooarchaeological materials held at VDHP and the University of Vermont Consulting Archaeology Program (UVM CAP) for radiocarbon analysis. These materials originated from sites that represent the later precontact and contact periods including Bohannon (VT-GI-026), Jonesville Bridge (VT-CH-619), Chimney Point (VT-AD-329), and Ewing (VT-CH-005) (Supplementary Table 1). Specimens were identified by Carder following standard zooarchaeological methods (Reitz and Wing, 2008) and the comparative collection held at the UVM CAP (see also Carder and Crock, 2021).
We submitted ~200 mg of whole bone to the Keck Carbon Cycle AMS Facility at UC Irvine for processing and radiocarbon analysis. Bone was decalcified in 1 N HCL, gelatinized in pH 2 at 60°C and ultrafiltered to select a high molecular weight fraction (>30 kDa). Stable isotopes reported as
where X is either 15N or 13C (‰), using standards of AIR and PeeDee Belemnite.
Stable isotope values (δ15N and δ13C) were used to assess bone preservation quality (C:N ratio). We calibrated radiocarbon dates using Oxcal v 4.4 (Bronk Ramsey, 2009) with IntCal20 (Reimer et al., 2020). One specimen (UCIAMS-259961) returned a post-bomb age and the fraction Modern was converted to calendar age using CALIBomb, Zone NH1.1
2.5. Historical and present day occurrences of focal fur-bearers
To develop a modern comparative dataset of known species locations, we queried the Global Biodiversity Informatics Facility (GBIF) for historical (i.e., catalogued museum specimens) and present day (i.e., research grade iNaturalist photographs) georeferenced occurrence records for our focal taxa within the administrative bounds of Vermont. Due to its recent extirpation, there are zero American marten records on GBIF and we relied on academic and state records of the reintroduction, with occurrence points taken from Aylward et al. (2019).
3. Results
3.1. Temporal span of fur-bearer associated radiocarbon dates
We identified 16 archaeological sites that have fur-bearing mammal remains either associated with radiocarbon-dated features or that have now been directly radiocarbon dated (Figure 1; Supplementary Table 2). Some of these sites represent occupations spanning thousands of years within the Holocene. The oldest features date as far back as the Early-Middle Archaic (~7,700 cal ybp such as Sandy Knoll/Guildhall, Mandel et al., 2022) and the Late Archaic (such as Ewing, associated lithics ~8,000 cal ybp; Petersen et al., 1985). However, most features containing fur-bearer remains date from the Early Woodland (~3,000 cal ybp) through European contact and into the present. The rarity of older remains (Figure 2) is most likely due to taphonomic biases; for example, aggregated paleobotanical data from Robinson et al. (2020) suggest that ~5,000 cal ybp, preservational quality declines dramatically, and far fewer sites overall are preserved from Early Holocene Vermont (Boulanger, 2007). This mirrors broader patterns for New England and North America in larger radiocarbon databases (Munoz et al., 2010; Chaput et al., 2015; Kelly et al., 2022). Within our curated list of sites, we found a total of 25 radiocarbon dates associated with at least one fur-bearer species. Moreover, we generated 7 new radiocarbon dates measured directly on bone (Supplementary Table 1), yielding a total of 32 radiocarbon dates representing fur-bearing mammals of Vermont.
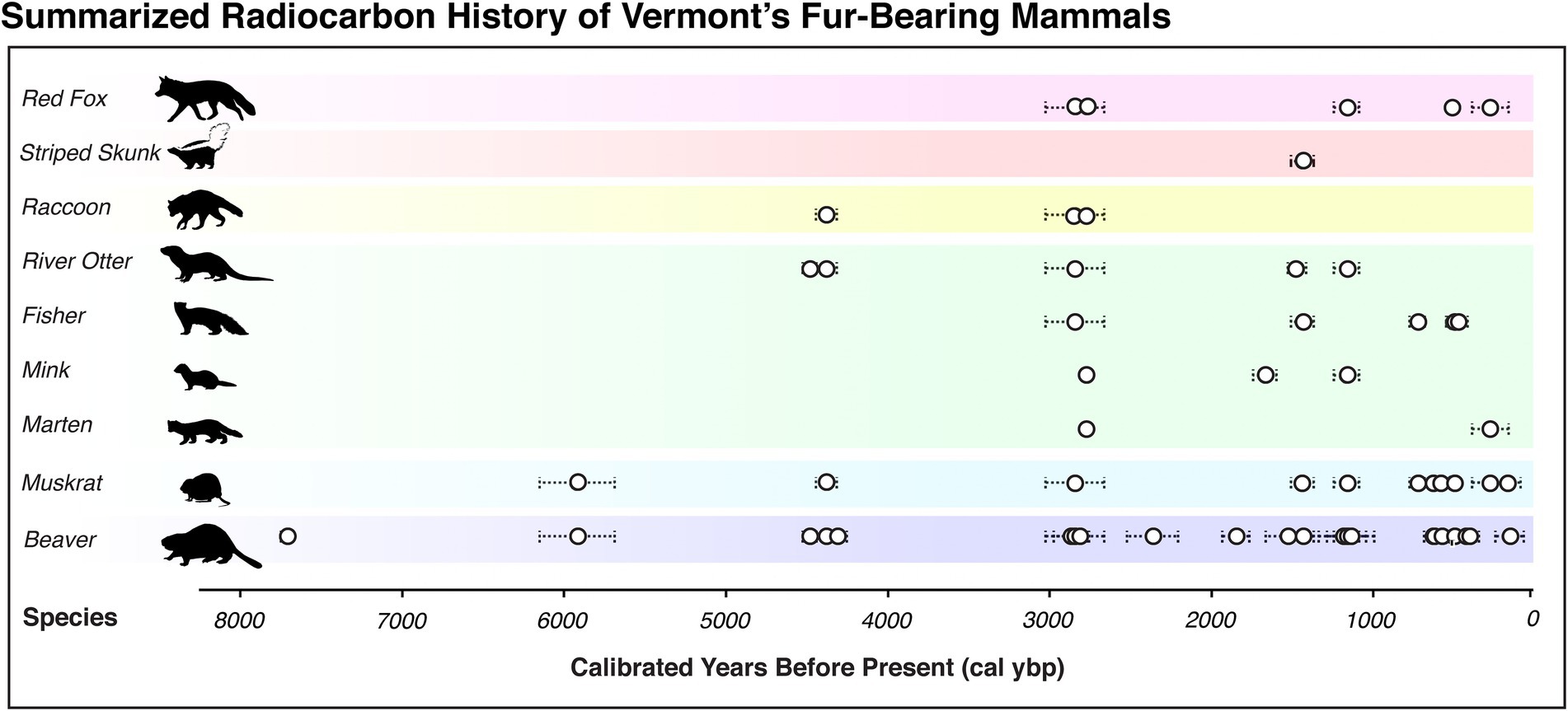
Figure 2. Radiocarbon chronology of fur-bearing mammals from Vermont archaeological sites, depicting both direct dates (for muskrats and beavers only) and radiocarbon dates from levels/loci/features that are associated with all of the fur-bearing mammal species featured. White circles show the mean calibrated date in years before present and dashed lines indicate standard deviation in calibrated years before present. Species from top to bottom include red fox (Canidae: Vulpes vulpes), striped skunk (Mephitidae: Mephitis mephitis), raccoon (Procyonidae: Procyon lotor), river otter (Mustelidae: Lontra canadensis), fisher (Mustelidae: Pekania pennanti), American mink (Mustelidae: Neovison vison), American marten (Mustelidae: Martes americana), muskrat (Cricetidae: Ondatra zibethicus), and beaver (Castoridae: Castor canadensis). See Supplementary Table 2 for the uncalibrated radiocarbon dates used to build this figure. Images courtesy of Phylopic.
3.2. New beaver and muskrat radiocarbon dates
Of the 10 samples that we submitted for radiocarbon analysis, three beaver specimens yielded zero collagen, including one specimen each from VT-CH-005 (Ewing), VT-AD-329 (Chimney Point), and VT-CH-619 (Jonesville Bridge). Our new radiocarbon dates (denoted by the UCIAMS lab identifier) cluster within the Late Holocene, with 95.4% ranges extending into the 20th century in three cases (excluding the Modern muskrat). All medians, excluding the Modern muskrat, fall within 1400–1800 CE (Figure 3), which overlapped with the period of intensive harvest for beaver pelts, including The Beaver Wars. Newly generated direct dates are consistent with existing charcoal-based radiocarbon chronologies for Ewing and Bohannon and they represent some of the most recent dates for Ewing (Supplementary Figure 1).
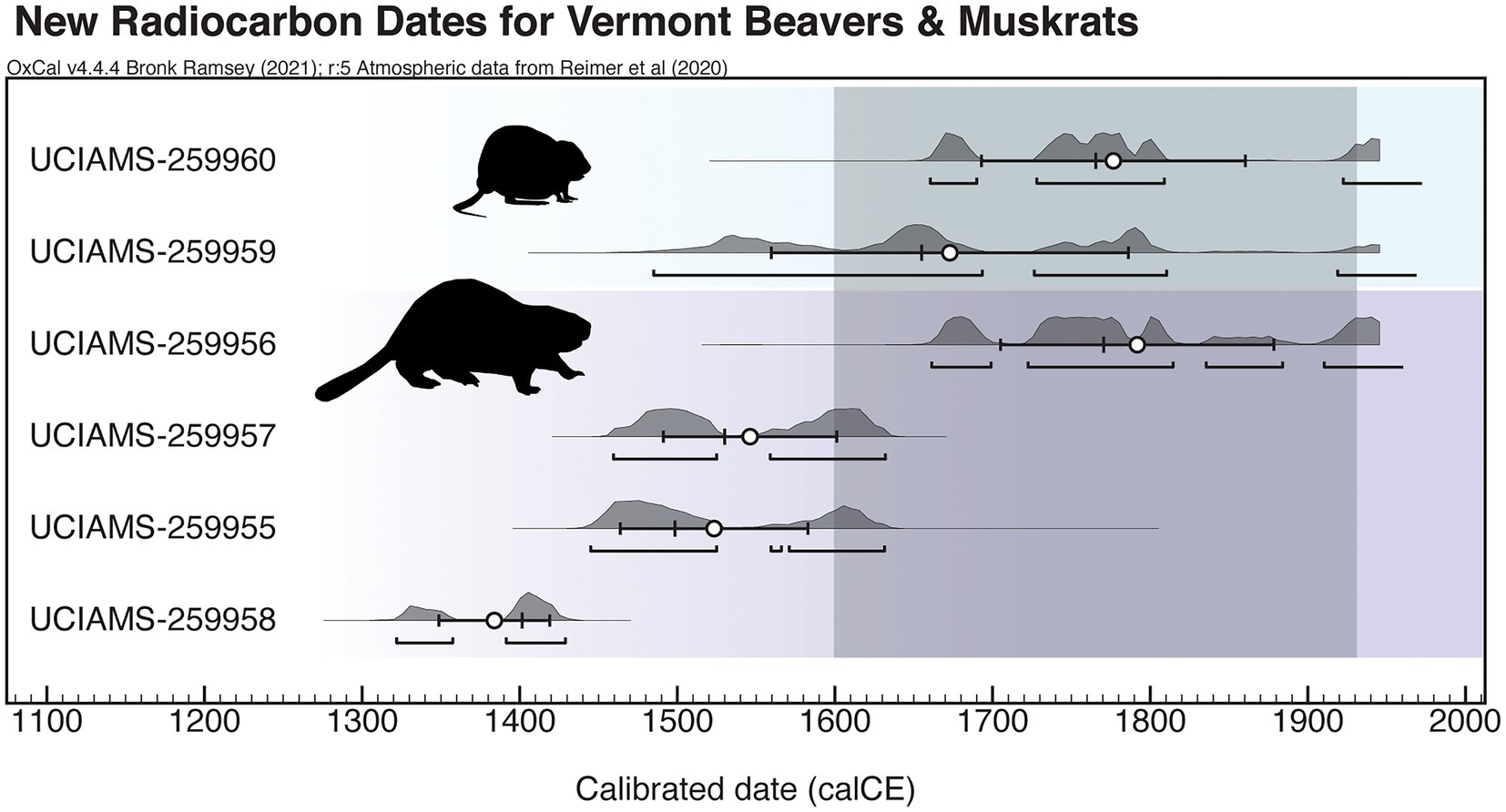
Figure 3. New radiocarbon dates spanning the North American fur trade, generated on key target species, beavers (purple) and muskrats (blue). White circles indicate mean ages, black pluses (+) indicate medians along a black line indicating sigma, and black brackets indicate the 95.4% range. The dark gray box indicates the onset and end of the period of intensive harvest and extirpation of these semi-aquatic mammals during the Fur Trade and the subsequent reintroduction of beavers (Kirk, 1923). These dates are calibrated using OxCal v. 4.4 (Bronk Ramsey, 2009) and IntCal20 (Reimer et al., 2020). Note that one muskrat from Ewing is not included as it is Modern. Images courtesy of Phylopic.
Radiocarbon dates from freshwater contexts could appear older than they actually are due to a freshwater reservoir effect caused by hardwater that contains calcium carbonates (Philippsen, 2013). The location of Ewing site (Shelburne Pond) contains dolomite that could cause a reservoir effect as 14C depleted carbon dissolves from bedrock into the water and is taken up into non-emergent aquatic vegetation (Lini et al., 2007). While Hart et al. (2019) found a freshwater offset of 132 ± 8 14Cyr for dogs consuming freshwater resources on the St. Lawrence River, Plint et al. (2019) did not include such an offset for paleontological beaver remains, and beavers are known to rely on terrestrial vegetation (Supplementary material); therefore, we did not apply a freshwater reservoir correction. Our muskrat and beaver radiocarbon dates are in line with expectations from contextual charcoal-based dates from the site that would be unaffected by this reservoir effect (Supplementary Figure 1). The rodent dates are among the youngest available for both Ewing and Bohannon, and a freshwater reservoir effect of ~100–200 years would place our dated specimens even more solidly within the Fur Trade time period.
3.3. Extent of historical and modern occurrence data
Over the past decade, iNaturalist occurrences of fur-bearing mammals generally fell within 200–300 observations per species (muskrat, striped skunk, river otter, fisher, American mink) with outliers of 0 observations for the American marten and >1,000 for beavers and raccoons (mean observations 474, median observations 260, range of 0–1,135; Supplementary Table 3). Digitized museum specimens for Vermont fur-bearers are comparatively rare when queried on GBIF – muskrats have the largest number available (19). The available digital records of catalogued museum specimens from GBIF cluster temporally at 1903 or 1970–1990, and these collections are held mostly by the Harvard Museum of Comparative Zoology (Supplementary Table 3). Maps comparing these occurrence points with zooarchaeological remains in general reveal continuity in the location of species in the state. The American marten is a clear exception as archaeological localities indicate their presence far outside their present-day range (Figure 4).

Figure 4. Panel of maps displaying the occurrences of focal fur-bearing mammals through time across Vermont. Black circles represent archaeological sites at which a given species is present; gray squares represent a species occurrence captured by a historical natural history collection. Present day observations (dark gray x’s) are drawn from research-grade iNaturalist observations for all species but the American marten, which reflects the present-day distribution based on Aylward et al. (2018). Maps were produced using QGIS, with state county polygons derived from the VT Open GeoData Portal.
4. Discussion
We recovered a temporal span of nearly 8,000 years of fur-bearer interactions with humans in Vermont based on dated features, including a near continuous record for beavers starting ~1,500 calibrated years before present during a time of potential extirpation (Figure 2; Supplementary Figure 2). Given the paucity of museum specimens available for our focal taxa (Figure 4; Supplementary Table 3), these zooarchaeological remains represent an untapped specimen archive that, in some cases, is contemporaneous with the early natural history collecting period and can be used to address shifting baselines as well as other questions of interest to wildlife management. Below we discuss potential biases in our dataset and conservatively integrate our results into a past socioecological context. We then consider what they mean for present day fur-bearer management and future population trends and landscape configurations.
4.1. Taphonomic and sample size considerations
Vertebrate remains are rarely preserved in Vermont due to acidic soils and other unfavorable conditions, such as freeze thaw cycles (Lacy, 1994; Robinson, 2011). These remains may not be resolved taxonomically, leaving many collections with size-based “unidentified mammal” categories. It can be difficult (if not impossible) to ascertain harvest numbers from such assemblages as increased numbers of specimens at a site may be an artifact of deposit age. Thus, the absence of a particular species from an archaeological site does not necessarily indicate its absence from that region or lack of cultural engagement (see Richmond et al., 2021 for the importance of considering such a caveat). Interpreting these records should include interdisciplinary collaboration as part of a continuing dialogue between academic researchers, consulting archaeologists, and practitioners, which can lead to revisiting historical collections or targeting new collections.
As it stands, our dataset should be interpreted with caution and at a presence/absence level within a regional context, rather than as an actual baseline of species abundance or harvest pressure. However, data at such temporal resolution and of similar sample sizes are readily used to inform conservation decision-making (see Turvey et al., 2017; West et al., 2017), for example, by making more robust species distribution models (e.g., Gibson et al., 2019) and by providing specimens that yield DNA for demographic modeling (Ramakrishnan and Hadly, 2009). Our work joins a growing body of literature demonstrating that even areas with fragmentary zooarchaeological records can benefit from engagement of archaeologists and conservation biologists, as such data consolidation and integration events can be used to establish new research questions and excavation priorities.
4.2. Millennial-scale legacies of human-fur-bearer interactions in Vermont
Here, we provide an integrated summary of fur-bearing mammal use in Vermont spanning millennia. Indigenous people in the Archaic of Vermont (~9,000–3,000 cal ybp) altered their subsistence practices to cope with the loss of marine resources and other environmental changes) following the retreat of the Champlain Sea (a post-glacial inlet that is now the freshwater Lake Champlain; Spiess and Wilson, 1987; Robinson et al., 1992; Crock and Robinson, 2012; Feranec et al., 2021). A number of sites in Vermont contain features with fur-bearers for this Archaic period: Sandy Knoll/Guildhall (VT-ES-0064) with an Early to Middle Archaic date; Weybridge Crossing (VT-AD-1744), Ewing (VT-CH-005), Headquarters (VT-FR-0318) Boucher (VT-FR-0026), and Canaan Bridge (VT-ES-002) dated to the Late Archaic-Early Woodland (Supplementary Table 2). However, the majority of fur-bearer remains instead represent the Woodland period [~3,000 cal ybp to contact (1600 CE)], which is defined by the use of ceramics, more sedentary settlement patterns, complex interaction spheres, and increased population sizes (Petersen and Power, 1985; Heckenberger et al., 1990; Haviland and Power, 1994). Our radiocarbon dates during the Woodland period are consistent with previously suggested subsistence practices (Petersen et al., 1985; Robinson, 2011; Supplementary Table 1). At Ewing, beavers and muskrats combined contributed the greatest estimated biomass and number of identified specimens among mammals following deer, and most postcranial elements exhibited cut marks, indicating entire animals were taken for processing (Carder and Crock, 2021). Thus, rodent radiocarbon dates from Ewing emphasize a trans-European contact period occupation and cultural resilience of harvest practices; as Petersen et al. (1985) note: there is “evidence for the late survival of native technologies in the late 17th century, possibly well into 18th centuries.”
While many Indigenous communities were significantly impacted by the Fur Trade and colonial activities more broadly, the presence of fur-bearers in traditional processing contexts following contact with Europeans suggest some continuity of harvest practices despite massive cultural disruptions experienced at the time. Our newly generated radiocarbon dates span this time of cultural disruption coincident with the Fur Trade (Figure 3), as dates UCIAMS 259956, 259959, and 259960 overlap with local smallpox epidemics in 1729–1733 and 1755–1758 CE (Supplementary Table 1). This period includes the introduction of diseases and firearms as well as shifts in the harvest of fur-bearers from subsistence items into commodities, though it is important to note that fur-bearing mammals may have been harvested in quantities beyond subsistence needs in some pre-contact contexts as well (Nassaney, 2015). Increased attention to archaeological sites within the past few hundred years can help us understand the interconnected socioecological system of North American fur-bearers.
4.3. New insights from the zooarchaeological record for Vermont fur-bearing mammals
Ecologists and land managers in Vermont and New England more broadly are recognizing that the region is experiencing reforestation following massive deforestation from the colonial period (Foster et al., 1998, 2002). Addressing shifted baselines has been critical in modeling the region’s responses to climate and land use change, such as through the New England Landscape Futures project (Thompson et al., 2020),2 and will be vital when considering the future geographic distribution of mammal species and their interactions with each other in the context of increased construction and habitat loss (Pearman-Gillman, 2020). Using our curated dataset, we discuss how zooarchaeological data can be relevant to broad themes of interest to wildlife management such as past extirpations and species introductions as well as future translocations.
4.3.1. Historical continuity of beaver and muskrat populations
Beaver foraging and damming behavior influences plant communities and hydrological processes (Larsen et al., 2021), with cascading effects on ecological successional stages that affect overall biodiversity (Gibson and Olden, 2014; Figure 1). Though muskrats are far smaller and do not construct dams, their consumption of aquatic vegetation and lodge building behavior results in similar ecological impacts (Kua et al., 2020). Our new radiocarbon dates, in addition to those we have compiled from the VDHP database, affirm the millennial-scale significance of beavers and muskrats within coupled socioecological systems in Vermont. Both beavers and muskrats are found widely across the state’s archaeological sites (Figure 4) and represent a significant contribution to deposits where they are present, such as at the Ewing site and Jonesville Bridge (Thomas et al., 1995; Carder and Crock, 2021). Both species are foci of human-wildlife conflict across the northeastern United States, especially given stakeholder perceptions that populations are increasing and spreading into novel habitats (Siemer et al., 2013). However, examination of historical Fur Trade records suggest that recent perceptions of overpopulation are a result of shifting baselines, as use of an 1850, rather than, 1970, baseline results in a reversal of trends from apparent increase to actual decline (Collins et al., 2020). Some harvest estimates within the last few decades similarly suggest muskrats are declining across their range in Canada (Sadowski and Bowman, 2021) and the northeastern United States (Roberts and Crimmins, 2010), potentially linked to disease.
A shifted baseline could similarly be present in Vermont, where in the 1920s, it was suggested that the population was eliminated for at least half a century or more (Kirk, 1923). Historic documents report that six beavers were translocated from the Adirondack Mountains of New York and released into Bennington County in southern Vermont in 1921 (Kirk, 1923). Yet, it is unclear whether the entire extant beaver population is derived from the Adirondacks, or if pockets of native Vermont beavers persisted throughout the state in low densities. Indeed, the radiocarbon date UCIAMS-259956 exhibits a 95.4% range that extends through this period of potential extirpation (Figure 3). Because these focal taxa are ecological engineers, this shifted baseline has consequences not only for management targets of population size but also alters what is considered “normal” for a landscape in structure and function, setting the stage for stakeholder dialogues (i.e., device installation such as the Beaver Deceiver™; Goldfarb, 2018).
4.3.2. Extralimital past occurrences of the American marten
American martens are present in two sites that span the Woodland period (~3,000 years ago through contact): Boucher (VT-FR-026) and Ewing (VT-CH-005) (Supplementary Table 2). These sites fall outside the current geographic range for the species (Figure 4), which has been created by a conservation reintroduction following a hypothesized historic extirpation event. Martens received state level protection in 1972. A reintroduction effort took place in southern Vermont from 1989 to 1991, seeded by 11 martens from New York and 104 martens from Maine; this reintroduction was considered unsuccessful (Distefano et al., 1990; Royar, 1992). However, camera trap surveys in 2015–2017 located a breeding population within the Green Mountain National Forest (O’Brien et al., 2018), and it is possible a natural recolonization has occurred. Genetic diversity studies suggest the possible persistence of undetected populations throughout the 20th century (Aylward et al., 2018, 2019).
The presence of martens in the Late Holocene zooarchaeological record could therefore indicate places where martens may have cryptically persisted, or could provide discussion points for future reintroduction decisions, though it is important to consider alternate explanations for the presence of this species at archaeological sites, such as ceremonial purposes (Heckenberger et al., 1990). Recent historical records [as reviewed by Krohn (2012)] suggest that, unlike today, martens were not necessarily limited to highlands and mountains. Holocene faunal records at the presence/absence level have been successfully used to inform habitat suitability and potential distributions for conservation decision-making (e.g., Lentini et al., 2018; Gibson et al., 2019; Barlow et al., 2021). As historical records indicate a severe range contraction in the 1930s that isolated martens in northern Maine and the Adirondacks (Hagmeier, 1956; Godin, 1977), regional syntheses could facilitate larger-scale conservation dialogues for the species.
4.3.3. The red fox was present in late Holocene Vermont
Zooarchaeological records can assist in the detection of non-native species, and, conversely, affirm the native status of species categorized as having been introduced (Mychajliw and Harrison, 2014; Hofman et al., 2015; West et al., 2017). Three canid species of varying “native” status currently inhabit Vermont: coyote (Canis latrans), red fox (Vulpes vulpes), and gray fox (Urocyon cineoargentatus), and the changing dynamics between them have consequences for human-wildlife conflict and zoonotic disease (e.g., Needle et al., 2020). However, only the red fox is present in our zooarchaeological dataset and is known from three Late Holocene sites (Bohannon, Boucher, Ewing; Figure 4).
The absence of coyotes agrees with previous documentation that identifies the coyote as a new arrival which expanded its range across North America following the extirpation of wolves (Hody and Kays, 2018); the first coyote was documented in Vermont in 1948. Red foxes, on the other hand, are currently denoted as non-native by the Vermont Department of Fish & Wildlife (VTFW, n.d.). This diagnosis may stem from historical references such as sporting magazines that recorded the importation of European red foxes for hunting in some parts of eastern North America (Statham et al., 2012; Frey, 2013). While some European haplotypes have been recovered, most matrilineal ancestry of eastern red foxes is North American, including in Vermont, agreeing with our findings of red fox presence in the Late Holocene (Kasprowicz et al., 2016; Kuo et al., 2019). Conversely, far less is known about the gray fox, which may now be shifting its range northward (Reding et al., 2021). While gray foxes have been harvested over the past decade within the state (VTFW, 2021), the absence of gray fox in the zooarchaeological record could be due to the taphonomic biases in our dataset, and there is debate regarding the species range throughout the Holocene in Canada (McAlpine et al., 2016). Gray foxes only appear in the natural history record for Vermont by 1910, represented by a single skin from southern Vermont that is now stored at the Museum of Comparative Zoology (MCZ 64310). Further genetic research on extant populations of these taxa could help elucidate the human imprint on their distributions.
4.4. Looking to the past when considering the future of fur-bearers
Despite the taphonomic limitations inherent to Vermont, we have synthesized a dataset that makes decades of archaeological expertise from academic and governmental sources available for conservation and management decision-making. Conservation and restoration efforts are increasingly recognizing the importance of centering the ecological practices and relationships of Indigenous communities (e.g., Hessami et al., 2021; Lamb et al., 2022; Reeder-Myers et al., 2022). Consideration of the long history of socioecological systems is critical to parameterizing management approaches and identifying shifted baselines (see Abrams and Nowacki, 2020, Leonard et al., 2020, and Roos, 2020). Evolving molecular techniques such as zooarchaeology by mass spectrometry (ZooMS; Richter et al., 2022), ancient environmental DNA from sediments (Lentz et al., 2021), and conservation archaeogenomics (Hofman et al., 2015) all provide notable paths forward toward generating a more fully resolved and holistic picture of past and future fur-bearing mammals.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material. Both Vermont radiocarbon databases can be freely downloaded at the VAHC Collections website, https://accd.vermont.gov/historic-preservation/archaeology-center/collections.
Author contributions
AM: conceptualization, methodology, formal analysis, investigation, resources, data curation, writing–original draft, visualization, supervision, and funding acquisition. AH: investigation, writing–original draft, and funding acquisition. DA-P: data curation, resources, and visualization. OO: investigation and resources. NC: conceptualization, data curation, investigation, resources, and writing–review and editing. JC: conceptualization, data curation, investigation, resources, and writing–review and editing. FR: conceptualization, data curation, investigation, resources, and writing–review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by funds from the Senior Research Project Supplement and Department of Biology at Middlebury College (AH).
Acknowledgments
This work was inspired by discussions with Kim Royar, Bill Kilpatrick, Alyssa Bennet, and others at the VT Department of Fish & Wildlife and VT Mammal Scientific Advisory Group. Archaeological data compilation was facilitated by Middlebury ENVS 401 students Shane Lusk and Steven Montilla Morantes. We thank John Southon and staff at the UCIAMS lab for processing specimens. This manuscript is a product of the Conservation Paleobiology Network working group “Leveraging lessons of the North American fur trade: integrated cultural and biological legacies.” We thank Courtney Hofman and two reviewers for feedback on this manuscript. Andrew Ng provided images for use in Figure 1. Middlebury College sits on land which has served as a site of meeting and exchange among indigenous peoples since time immemorial. The Western Abenaki are the traditional caretakers of these Vermont lands and waters, which they call Ndakinna, or “homeland.” We remember their connection to this region and the hardships they continue to endure. We give thanks for the opportunity to share in the bounty of this place and to protect it.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1065567/full#supplementary-material
Footnotes
1. ^http://calib.org accessed 2022-07-31.
References
Abrams, M. D., and Nowacki, G. J. (2020). Native American imprint in palaeoecology. Nat. Sustainability 3, 896–897. doi: 10.1038/s41893-020-0578-6
Akçakaya, H. R., Bennett, E. L., Brooks, T. M., Grace, M. K., Heath, A., Hedges, S., et al. (2018). Quantifying species recovery and conservation success to develop an IUCN green list of species. Conserv. Biol. 32, 1128–1138. doi: 10.1111/cobi.13112
Aylward, C. M., Murdoch, J. D., Donovan, T. M., Kilpatrick, C. W., Bernier, C., and Katz, J. (2018). Estimating distribution and connectivity of recolonizing American marten in the northeastern United States using expert elicitation techniques. Anim. Conserv. 21, 483–495. doi: 10.1111/acv.12417
Aylward, C. M., Murdoch, J. D., and Kilpatrick, C. W. (2019). Genetic legacies of translocation and relictual populations of American marten at the southeastern margin of their distribution. Conserv. Genet. 20, 275–286. doi: 10.1007/s10592-018-1130-3
Barlow, M. M., Johnson, C. N., McDowell, M. C., Fielding, M. W., Amin, R. J., and Brewster, R. (2021). Species distribution models for conservation: identifying translocation sites for eastern quolls under climate change. Global Ecol. Conserv. 29:e01735. doi: 10.1016/j.gecco.2021.e01735
Barnosky, A. D., Hadly, E. A., Gonzalez, P., Head, J., Polly, P. D., Lawing, A. M., et al. (2017). Merging paleobiology with conservation biology to guide the future of terrestrial ecosystems. Science 355:eaah4787. doi: 10.1126/science.aah4787
Boulanger, M. T. (2007). Database of published radiocarbon dates for Vermont with a brief discussion of the method. J. Vt. Archaeol. 8, 1–25.
Bronk Ramsey, C. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360. doi: 10.1017/S0033822200033865
Buxton, R. T., Nyboer, E. A., Pigeon, K. E., Raby, G. D., Rytwinski, T., Gallagher, A. J., et al. (2021). Avoiding wasted research resources in conservation science. Conserv. Sci. Pract. 3:e329. doi: 10.1111/csp2.329
Calloway, C. G. (1994). The Western Abenakis of Vermont, 1600–1800: war, migration, and the survival of an Indian people, vol. 197. Norman, Oklahoma: University of Oklahoma Press.
Cammen, K. M., Rasher, D. B., and Steneck, R. S. (2019). Predator recovery, shifting baselines, and the adaptive management challenges they create. Ecosphere 10:e02579. doi: 10.1002/ecs2.2579
Carder, N., and Crock, J. G. (2021). A deer camp forever: Archaeofauna from the Ewing site. Archaeol. East. N. Am. 49, 103–132.
Carlos, A. M., and Lewis, F. D. (1993). Indians, the beaver, and the bay: the economics of depletion in the lands of the Hudson's bay company, 1700–1763. J. Econ. Hist. 53, 465–494. doi: 10.1017/S0022050700013450
Chaput, M. A., Kriesche, B., Betts, M., Martindale, A., Kulik, R., Schmidt, V., et al. (2015). Spatiotemporal distribution of Holocene populations in North America. Proc. Natl. Acad. Sci. U. S. A. 112, 12127–12132. doi: 10.1073/pnas.1505657112
Collins, A. C., Böhm, M., and Collen, B. (2020). Choice of baseline affects historical population trends in hunted mammals of North America. Biol. Conserv. 242:108421. doi: 10.1016/j.biocon.2020.108421
Crock, J. G., and Robinson, F. W. IV (2012). Maritime mountaineers Paleoindian settlement patterns on the West coast of New England. Late Pleistocene Archaeol. Ecol. Far Northeast 48. doi: 10.1080/20555563.2017.1380997
Cronon, W. (1983). Changes in the land: Indians, colonists, and the ecology of New England. New York: Hill & Wang.
Dietl, G. P., and Flessa, K. W. (2011). Conservation paleobiology: putting the dead to work. Trends Ecol. Evol. 26, 30–37. doi: 10.1016/j.tree.2010.09.010
Dillon, E. M., Pier, J. Q., Smith, J. A., Raja, N. B., Dimitrijevic, D., Austin, E. L., et al. (2022). What is conservation paleobiology? Tracking 20 years of research and development. Front. Ecol. Evol. 10:1031483. doi: 10.3389/fevo.2022.1031483
Distefano, J. J., Royar, K. J., Pence, D. M., and Denoncour, J. E., (1990). Marten recovery plan for Vermont. Vol. 19. (Waterbury, Vermont, USA: Vermont Fish & Wildlife Department and USFS-GMNF. Springfield, Vermont).
Ellis, E. C., Gauthier, N., Klein Goldewijk, K., Bliege Bird, R., Boivin, N., Díaz, S., et al. (2021). People have shaped most of terrestrial nature for at least 12,000 years. Proc. Natl. Acad. Sci. U. S. A. 118:e2023483118. doi: 10.1073/pnas.2023483118
Feranec, R. S., Cournoyer, M. E., and Kozlowski, A. L. (2021). 14C dates and stable isotope ecology of marine vertebrates in the late Pleistocene-early Holocene Champlain Sea. Radiocarbon 63, 1259–1272. doi: 10.1017/RDC.2021.40
Foster, D. R., Motzkin, G., Bernardos, D., and Cardoza, J. (2002). Wildlife dynamics in the changing New England landscape. J. Biogeogr. 29, 1337–1357. doi: 10.1046/j.1365-2699.2002.00759.x
Foster, D. R., Motzkin, G., and Slater, B. (1998). Land-use history as long-term broad-scale disturbance: regional forest dynamics in Central New England. Ecosystems 1, 96–119. doi: 10.1007/s100219900008
Frey, J. K. (2013). Re-evaluation of the evidence for the importation of red foxes from Europe to colonial America: origins of the southeastern red fox (Vulpes vulpes fulva). Biol. Conserv. 158, 74–79. doi: 10.1016/j.biocon.2012.07.027
Gibson, L. M., Mychajliw, A. M., Leon, Y., Rupp, E., and Hadly, E. A. (2019). Using the past to contextualize anthropogenic impacts on the present and future distribution of an endemic Caribbean mammal. Conserv. Biol. 33, 500–510. doi: 10.1111/cobi.13290
Gibson, P. P., and Olden, J. D. (2014). Ecology, management, and conservation implications of north American beaver (Castor canadensis) in dryland streams. Aquat. Conserv. Mar. Freshwat. Ecosyst. 24, 391–409. doi: 10.1002/aqc.2432
Godin, A. J.. (1977). Wild mammals of New England. Johns Hopkins University Press, Baltimore, MD. 303.
Goldfarb, B. (2018). Eager: the surprising, secret life of beavers and why they matter. Chelsea, Vermont: Chelsea Green Publishing. 304.
Groff, D. V., McDonough, M. K. C., Pier, J. Q., Shaffer, A. B., and Dietl, G. P. (2022). Quantifying the impact of conservation paleobiology research. Geol. Soc. Am. Abstr. Programs 54. doi: 10.1130/abs/2022AM-383267
Hagmeier, E. M. (1956). Distribution of marten and fisher in North America. Can. Field Nat. 70, 149–168.
Hart, J. P., Feranec, R. S., Abel, T. J., and Vavrasek, J. L. (2019). Freshwater reservoir offsets on radiocarbon-dated dog bone from the headwaters of the St. Lawrence River, USA. PeerJ 7:e7174. doi: 10.7717/peerj.7174
Haviland, W. A., and Power, M. W. (1994). The original Vermonters: native inhabitants, past and present. Lebanon, New Hampshire: UPNE.
Heckenberger, M. J., Petersen, J. B., Basa, L. A., Cowie, E. R., Spiess, A. E., and Stuckenrath, R. E. (1990). Early woodland period mortuary ceremonialism in the far northeast: a view from the Boucher cemetery. Archaeol. East. N. Am. 18, 109–144.
Hessami, M. A., Bowles, E., Popp, J. N., and Ford, A. T. (2021). Indigenizing the north American model of wildlife conservation. FACETS 6, 1285–1306. doi: 10.1139/facets-2020-0088
Hody, J. W., and Kays, R. (2018). Mapping the expansion of coyotes (Canis latrans) across north and Central America. ZooKeys 759, 81–97. doi: 10.3897/zookeys.759.15149
Hofman, C. A., Rick, T. C., Fleischer, R. C., and Maldonado, J. E. (2015). Conservation archaeogenomics: ancient DNA and biodiversity in the Anthropocene. Trends Ecol. Evol. 30, 540–549. doi: 10.1016/j.tree.2015.06.008
Kasprowicz, A. E., Statham, M. J., and Sacks, B. N. (2016). Fate of the other redcoat: remnants of colonial British foxes in the eastern United States. J. Mammal. 97, 298–309. doi: 10.1093/jmammal/gyv179
Kelly, R. L., Mackie, M. E., Robinson, E., Meyer, J., Berry, M., Boulanger, M., et al. (2022). A new radiocarbon database for the lower 48 states. Am. Antiq. 87, 581–590. doi: 10.1017/aaq.2021.157
Kirk, G. L. (1923). Beaver introduced in Vermont. J. Mammal. 4, 59–60. doi: 10.1093/jmammal/4.1.59-a
Klyza, C. M., and Trombulak, S. C. (1999). The story of Vermont: a natural and cultural history. Lebanon, New Hampshire: UPNE.
Krohn, W. B. (2012). “Distribution changes of American martens and fishers in eastern North America, 1699–2001” in Biology and conservation of martens, sables, and fishers: a new synthesis. eds. K. B. Aubry, W. J. Zielinski, M. G. Raphael, G. Proulx, and S. W. Buskirk (Ithaca, NY: Cornell University Press), 58–73.
Kua, Z. X., Stella, J. C., and Farrell, J. M. (2020). Local disturbance by muskrat, an ecosystem engineer, enhances plant diversity in regionally-altered wetlands. Ecosphere 11:e03256. doi: 10.1002/ecs2.3256
Kuo, Y. H., Vanderzwan, S. L., Kasprowicz, A. E., and Sacks, B. N. (2019). Using ancestry-informative SNPs to quantify introgression of European alleles into north American red foxes. J. Hered. 110, 782–792. doi: 10.1093/jhered/esz053
Lacy, D. M. (1994). Prehistoric land-use in the Green Mountains: a view from the National Forest. J. Vermont Archaeol. 1, 92–102.
Lamb, C. T., Willson, R., Richter, C., Owens-Beek, N., Napoleon, J., Muir, B., et al. (2022). Indigenous-led conservation: pathways to recovery for the nearly extirpated Klinse-Za mountain caribou. Ecol. Appl. 32:e2581. doi: 10.1002/eap.2581
Larsen, A., Larsen, J. R., and Lane, S. N. (2021). Dam builders and their works: beaver influences on the structure and function of river corridor hydrology, geomorphology, biogeochemistry and ecosystems. Earth Sci. Rev. 218:103623. doi: 10.1016/j.earscirev.2021.103623
Lentini, P. E., Stirnemann, I. A., Stojanovic, D., Worthy, T. H., and Stein, J. A. (2018). Using fossil records to inform reintroduction of the kakapo as a refugee species. Biol. Conserv. 217, 157–165. doi: 10.1016/j.biocon.2017.10.027
Lentz, D. L., Hamilton, T. L., Dunning, N. P., Tepe, E. J., Scarborough, V. L., Meyers, S. A., et al. (2021). Environmental DNA reveals arboreal cityscapes at the ancient Maya Center of Tikal. Sci. Rep. 11, 1–10. doi: 10.1038/s41598-021-91620-6
Leonard, K., Aldern, J. D., Christianson, A., Ranco, D., Thornbrugh, C., Loring, P. A., et al. (2020). Indigenous conservation practices are not a monolith: western cultural biases and a lack of engagement with indigenous experts undermine studies of land stewardship. EcoEvoRxiv. doi: 10.32942/osf.io/jmvqy. [E-pub ahead of print].
Lini, A., Levine, S., Ostrofsky, M. L., and Dahlen, D., (2007). Trophic history of Shelburne pond, Shelburne, Vermont. Vermont Agency of Natural Resources.
Mandel, G. A., Crock, J. G., and Garcia, J. K. (2022). Archaeological supplemental phase II evaluation and phase III data recovery at the Sandy knoll site (VT-ES-0064) for the U.S. route 2 bridge replacement project, Lancaster, Coos County, New Hampshire and guildhall, Essex County, Vermont. Report prepared for the Vermont Agency of Transportation. University of Vermont Consulting Archaeology Program Report Number 1416.
McAlpine, D. F., Cormier, J., and MacLean, I. G. (2016). Second report of the gray fox, Urocyon cinereoargenteus, in New Brunswick. Ottawa Field-Naturalists’ Club 130, 164–166. doi: 10.22621/cfn.v130i2.1841
McClenachan, L., Cooper, A. B., McKenzie, M. G., and Drew, J. A. (2015). The importance of surprising results and best practices in historical ecology. Bioscience 65, 932–939. doi: 10.1093/biosci/biv100
McManus, J. C. (1972). An economic analysis of Indian behavior in the north American fur trade. J. Econ. Hist. 32, 36–53. doi: 10.1017/S0022050700075380
Munoz, S. E., Gajewski, K., and Peros, M. C. (2010). Synchronous environmental and cultural change in the prehistory of the northeastern United States. Proc. Natl. Acad. Sci. U. S. A. 107, 22008–22013. doi: 10.1073/pnas.1005764107
Mychajliw, A. M., and Harrison, R. G. (2014). Genetics reveal the origin and timing of a cryptic insular introduction of muskrats in North America. PLoS One 9:e111856. doi: 10.1371/journal.pone.0111856
Nassaney, M. S. (2015). The archaeology of the north American fur trade. Gainesville, Florida: University Press of Florida.
Needle, D. B., Marr, J. L., Park, C. J., Andam, C. P., Wise, A. G., Maes, R. K., et al. (2020). Concurrent infection of skunk adenovirus-1, listeria monocytogenes, and a regionally specific clade of canine distemper virus in one gray fox (Urocyon cinereoargenteus) and concurrent listeriosis and canine distemper in a second gray fox. Pathogens 9:591. doi: 10.3390/pathogens9070591
Newsom, B. D. (2022). “Variation amid homogeneity: an examination of early ceramic period Technologies in the Penobscot River Valley of Maine” in The far northeast: 3000 BP to contact. eds. H. Kenneth and M. G. Hrynick (Ottawa: Mercury Series, Canadian Museum of History, University of Ottawa Press)
O’Brien, P., Bernier, C., and Hapeman, P. (2018). A new record of an American marten (Martes americana) population in southern Vermont. Small Carniv. Conserv. 56, 68–75.
Pauly, D. (1995). Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 10:430. doi: 10.1016/S0169-5347(00)89171-5
Pearman-Gillman, S., (2020). Predicting wildlife distributions and resilience under alternative futures. Graduate College Dissertations and Theses. 1237.
Petersen, J. B., and Power, M. W. (1985). Three middle woodland ceramic assemblages from the Winooski site. Occasional Papers Northeast. Archaeol. 9, 109–159.
Petersen, J. B., Wolford, J. A., Hamilton, N. D., LaBar, L., and Heckenberger, M. (1985). Archaeological investigations in the Shelburne pond locality, Chittenden County, Vermont. Ann. Carnegie Mus. 54, 23–75. doi: 10.5962/p.330770
Philippsen, B. (2013). The freshwater reservoir effect in radiocarbon dating. Heritage Sci. 1, 1–19. doi: 10.1186/2050-7445-1-24
Plint, T., Longstaffe, F. J., and Zazula, G. (2019). Giant beaver palaeoecology inferred from stable isotopes. Sci. Rep. 9, 1–12. doi: 10.1038/s41598-019-43710-9
Ramakrishnan, U. M. A., and Hadly, E. A. (2009). Using phylochronology to reveal cryptic population histories: review and synthesis of 29 ancient DNA studies. Mol. Ecol. 18, 1310–1330. doi: 10.1111/j.1365-294X.2009.04092.x
Reding, D. M., Castaneda-Rico, S., Shirazi, S., Hofman, C. A., Cancellare, I. A., Lance, S. L., et al. (2021). Mitochondrial genomes of the United States distribution of gray fox (Urocyon cinereoargenteus) reveal a major phylogeographic break at the Great Plains suture zone. Front. Ecol. Evol. 9:666800. doi: 10.3389/fevo.2021.666800
Reeder-Myers, L., Braje, T. J., Hofman, C. A., Elliott Smith, E. A., Garland, C. J., Grone, M., et al. (2022). Indigenous oyster fisheries persisted for millennia and should inform future management. Nat. Commun. 13, 1–13. doi: 10.1038/s41467-022-29818-z
Reimer, P. J., Austin, W. E., Bard, E., Bayliss, A., Blackwell, P. G., Ramsey, C. B., et al. (2020). The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757. doi: 10.1017/RDC.2020.41
Reitz, E. J., and Wing, E. S. (2008). Zooarchaeology Cambridge Manuals in Archaeology, Cambridge University Press.
Richmond, J. Q., Swift, C. C., Wake, T. A., Brehme, C. S., Preston, K. L., Kus, B. E., et al. (2021). Impacts of a non-indigenous ecosystem engineer, the American beaver (Castor canadensis), in a biodiversity hotspot. Front. Conserv. Sci. 2:752400. doi: 10.3389/fcosc
Richter, K. K., Codlin, M. C., Seabrook, M., and Warinner, C. (2022). A primer for ZooMS applications in archaeology. Proc. Natl. Acad. Sci. U. S. A. 119:e2109323119. doi: 10.1073/pnas.2109323119
Roberts, N. M., and Crimmins, S. M. (2010). Do trends in muskrat harvest indicate widespread population declines? Northeast. Nat. 17, 229–238. doi: 10.1656/045.017.0206
Robinson, F. W. IV (2011). “Powerful history: the archaeology of native people in the Champlain lowlands” in Native American archaeological sites along the VELCRO northwest reliability project corridor in Chittenden, Addison and Rutland counties (Rutland: Vermont Electric Power Company)
Robinson, F. W. IV, and Ostrum, B. (2016). Database of Paleobotanical and Paleofaunal remains recovered from radiocarbon-dated contexts in Vermont Precontact archaeological sites Vermont Division of Historic Preservation Available at: https://accd.vermont.gov/historic-preservation/archaeology-center/collections.
Robinson, F. W. IV, Ostrum, B., Morell, T., Boulanger, M., Dillon, R. S., and Basque, Y. B. B. (2020). Database of Paleobotanical and Paleofaunal remains recovered from radiocarbon-dated contexts in Vermont Precontact archaeological sites Vermont Division of Historic Preservation Available at: https://outside.vermont.gov/agency/ACCD/ACCD_Web_Docs/HP/Archaeology/AHC-Database-VT-C14-Dates-2020.xlsx
Robinson, F. J. IV, Springston, G. E., and Crock, J. G. (2017). Early and middle Paleoindian settlement patterns and the late-Pleistocene environment along the Champlain Sea. PaleoAmerica 3, 323–336. doi: 10.1080/20555563.2017.1380997
Robinson, B. S., Petersen, J. B., and Robinson, A. K., (1992). Early Holocene occupation in northern New England. Occasional Publications in Maine Archaeology, Number Nine. Augusta, Maine.
Rodrigues, A. S., Monsarrat, S., Charpentier, A., Brooks, T. M., Hoffmann, M., Reeves, R., et al. (2019). Unshifting the baseline: a framework for documenting historical population changes and assessing long-term anthropogenic impacts. Philos. Trans. R. Soc. B 374:20190220. doi: 10.1098/rstb.2019.0220
Roos, C. I. (2020). Scale in the study of indigenous burning. Nat. Sustainability 3, 898–899. doi: 10.1038/s41893-020-0579-5
Royar, K. J. (1992). Monitoring reintroduced marten populations in Vermont. Vermont Fish and Wildlife Department. Waterbury, Vermont, USA.
Sadowski, C., and Bowman, J. (2021). Historical surveys reveal a long-term decline in muskrat populations. Ecol. Evol. 11, 7557–7568. doi: 10.1002/ece3.7588
Siemer, W. F., Jonker, S. A., Decker, D. J., and Organ, J. F. (2013). Toward an understanding of beaver management as human and beaver densities increase. Hum.-Wildl. Interact. 7, 114–131.
Silliman, B. R., Hughes, B. B., Gaskins, L. C., He, Q., Tinker, M. T., Read, A., et al. (2018). Are the ghosts of nature’s past haunting ecology today? Curr. Biol. 28, R532–R537. doi: 10.1016/j.cub.2018.04.002
Spiess, A. E., and Wilson, D. B. (1987). Maine Archaeological Society; and Maine Historic Preservation Commission, “Michaud, a Paleoindian Site in the New England-Maritimes Region”. His. Preserv. Comm. Doc. 7.
Statham, M. J., Sacks, B. N., Aubry, K. B., Perrine, J. D., and Wisely, S. M. (2012). The origin of recently established red fox populations in the United States: translocations or natural range expansions? J. Mammal. 93, 52–65. doi: 10.1644/11-MAMM-A-033.1
Thomas, P. A. (1979). In the maelstrom of change. The Indian trade and cultural process in the middle Connecticut River valley: 1635–1665 University of Massachusetts Amherst. Amherst, Massachusetts: ProQuest Dissertations Publishing.
Thomas, P. A. (1994). Vermont archaeology comes of age: a current perspective on Vermont's prehistoric past. J. Vermont Archaeol. 1, 39–91.
Thomas, P. A., Sloma, B., Carder, N., and Florentin, R. (1995). Archaeological site identification, evaluation, and mitigation of VT-CH-619, Richmond TH07, Richmond, Vermont. Report 135. University of Vermont Consulting Archaeology Program, Burlington.
Thompson, J. R., Plisinski, J. S., Lambert, K. F., Duveneck, M. J., Morreale, L., McBride, M., et al. (2020). Spatial simulation of codesigned land cover change scenarios in New England: alternative futures and their consequences for conservation priorities. Earth's. Future 8:e2019EF001348. doi: 10.1029/2019EF001348
Turgeon, L. (1998). French fishers, fur traders, and Amerindians during the sixteenth century: history and archaeology. William Mary Q. 55, 585–610. doi: 10.2307/2674446
Turvey, S. T., Barnes, I., Marr, M., and Brace, S. (2017). Imperial trophy or island relict? A new extinction paradigm for pere David’s deer: a Chinese conservation icon. R. Soc. Open Sci. 4:171096. doi: 10.1098/rsos.171096
Turvey, S. T., Crees, J. J., and Di Fonzo, M. M. (2015). Historical data as a baseline for conservation: reconstructing long-term faunal extinction dynamics in late Imperial–modern China. Proc. R. Soc. B Biol. Sci. 282:20151299. doi: 10.1098/rspb.2015.1299
Veltre, D. W., and McCartney, A. P. (2002). Russian exploitation of Aleuts and fur seals: the archaeology of eighteenth-and early-nineteenth-century settlements in the Pribilof Islands, Alaska. Hist. Archaeol. 36, 8–17. doi: 10.1007/BF03374356
VTFW (2021). Furbearer Conservation and Management in Vermont 2021. Available at: https://vtfishandwildlife.com/sites/fishandwildlife/files/documents/Learn%20More/Library/REPORTS%20AND%20DOCUMENTS/FURBEARER%20AND%20TRAPPING/furbearer-conservation-management-report-2021.pdf
VTFW (n.d.) Red fox: Vermont wildlife fact sheet. Available at: https://vtfishandwildlife.com/sites/fishandwildlife/files/documents/Learn%20More/Library/FACTSHEETS/FURBEARER%20AND%20TRAPPING/FURBEARER%20FACTSHEETS/RED%20FOX.pdf
West, C., Hofman, C. A., Ebbert, S., Martin, J., Shirazi, S., Dunning, S., et al. (2017). Integrating archaeology and ancient DNA analysis to address invasive species colonization in the Gulf of Alaska. Conserv. Biol. 31, 1163–1172. doi: 10.1111/cobi.12865
White, H. B., Batcheller, G. R., Boggess, E. K., Brown, C. L., Butfiloski, J. W., Decker, T. A., et al. (2021). Best management practices for trapping furbearers in the United States. Wildl. Monogr. 207, 3–59. doi: 10.1002/wmon.1057
Keywords: zooarchaeology, historical ecology, mammal, fur trade, conservation paleobiology, radiocarbon dating (14C)
Citation: Mychajliw AM, Hsi AY, An-Pham D, Olson OL, Carder N, Crock JG and Robinson F&W and (2023) Zooarchaeological assemblages contextualize the historical ecology and harvest of fur-bearing mammals in Vermont. Front. Ecol. Evol. 11:1065567. doi: 10.3389/fevo.2023.1065567
Edited by:
G. Lynn Wingard, United States Geological Survey (USGS), United StatesReviewed by:
José Manuel López, CONICET Argentine Institute of Arid Zone Research (IADIZA), ArgentinaRebecca Terry, Oregon State University, United States
Copyright © 2023 Mychajliw, Hsi, An-Pham, Olson, Carder, Crock and Robinson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexis M. Mychajliw, YW15Y2hhamxpd0BtaWRkbGVidXJ5LmVkdQ==