 Cas F. Carroll
Cas F. Carroll Jacquelyn L. Gill
Jacquelyn L. Gill Caitlin McDonough MacKenzie
Caitlin McDonough MacKenzie- 1Ecology, Evolution, and Conservation Biology, University of Nevada, Reno, Reno, NV, United States
- 2Climate Change Institute, University of Maine, Orono, ME, United States
- 3Colby College, Waterville, ME, United States
Identifying refugia— specifically places where species can persist during periods of regionally unsuitable climate— is increasingly important for conservation practitioners and land managers charged with protecting biodiversity in a rapidly warming world. Currently, many researchers assist in this process by building models to predict areas of refugia using climate data projected into the future under different climate scenarios; however, the coarse spatial scale of future climate data can be orders of magnitude larger than the scale of refugia on the landscape. Conservation paleobiology is an emerging field that can contribute to the identification of climate refugia by looking at the macrofossil records contained in sediments to better understand the response of species to past climate change within a small area, and allows us to ground-truth hypotheses about specific areas functioning as climate refugia. Here, we present a conservation paleobiology case study to update vulnerability assessments for subalpine plant species in Acadia National Park and locate potential future refugia on the landscape. We analyzed plant macrofossils in a sediment core from Sargent Mountain Pond in Acadia National Park (Maine, United States) at a fine spatiotemporal resolution to test the hypothesis that the area served as a past climate refugium for the subalpine species it currently hosts. We found that, when compared to a pollen record from a forest hollow core collected on Mount Desert Island, the macrofossils reflect a more stable presence of subalpine taxa throughout the Holocene Climatic Optimum (8,000–5,000 BP) than was observed at lower elevations. Our results indicate the importance of a complementary approach that combines plant macrofossils and pollen in addition to modeling to identify refugia and better understand the vulnerability of species and communities to climate change.
Introduction
Identifying climate refugia with paleobiological data can refine climate change vulnerability assessments (Dietl and Flessa, 2011; Morelli et al., 2016). Climate refugia are local sites that support taxa unlikely to survive in the regional climate: these sites may be decoupled or buffered from regional trends in climate (Rull, 2009; Dobrowski, 2011). In the 20th century, paleobiologists developed this concept to describe the survival of temperate taxa during the last glacial maximum. The southern Appalachians were refugia supporting taxa that could not survive at the glacial margin; local persistence through the Pleistocene signaled these were warm refugia (Barrington and Paris, 2007). Today, paleobiologists are more interested in refugia for cold-adapted species in a warming world (Keppel et al., 2015). Dramatic climate change occurred throughout the late Pleistocene and early Holocene: the Bølling-Allerød warm period’s abrupt onset of warming (14,700 BP), the abrupt climatic reversal to colder conditions at the onset of the Younger Dryas (12,800 BP; Peteet et al., 1990, 1993), another abrupt warming period immediately following the Younger Dryas (11,700 BP), and, later, the Holocene Climatic Optimum (8,000–5,000 year BP). Local persistence of cold-adapted taxa throughout these past periods of climate change signal potential refugia under present and future anthropogenic climate change. Identifying and protecting climate refugia is an important conservation strategy under anthropogenic climate change: these sites may be buffered from the most intense warming while protecting rear-edge populations of species experiencing range shifts (Willis and Birks, 2006; Morelli et al., 2016). Modeling research suggests that designing conservation strategies around cool refugia, rather than current biodiversity hotspots, leads to much higher rates of species conservation in the future (Nadeau et al., 2022). Paleobiology provides tools to identify these refugia (Hunter et al., 1988). Relict populations persisting in place throughout the Holocene against regional patterns of change are evidence of locally favorable climates for these cold-adapted species (Schauffler and Jacobson, 2002).
Pollen and plant macrofossil records point to potential climate refugia in the mountains of the northeastern United States. There is a rich paleobiological record of dynamic plant assemblages shifting across the lowlands of the northeast since glacial retreat (Davis et al., 1980; Schauffler and Jacobson, 2002; Oswald et al., 2018). In contrast, only two sets of sediment cores collected along elevation gradients reach into the alpine-treeline ecotone with pollen and macrofossil data (see Figure 1 for all cores referenced in this paper). These records from New York’s Adirondack Mountains (Jackson and Whitehead, 1991) and New Hampshire’s White Mountains (Spear, 1989; Spear et al., 1994; Miller and Spear, 1999) reflect a relatively stable vegetation history at high elevations. In the Adirondacks, lower elevation sites (approximately 1,000 m and below) saw frequent shifts (500 to 4,000-year intervals) in the dominance of tree taxa, while at the highest elevation site (1,320 m) Picea and Betula papyrifera co-dominance (beginning 8,200 years BP) shifted to B. papyrifera dominance approximately at 7500 BP, then Abies balsamea dominated from 6,200 years BP to present (see Jackson and Whitehead, 1991, Figure 7). In the White Mountains, four subalpine and alpine lakes (1,140–1,676 m) record shifts in species’ elevation range and dominance over much of the 13,000-year record (Spear, 1989). However, during the post-Younger Dryas warming period (10,300 years BP) these shifts in community composition were more obvious at lower elevations while high elevations maintained communities dominated by Abies and Picea (Spear et al., 1994, Figure 9). These records from the Adirondack and White Mountains suggest a persistent presence of subalpine taxa at the highest elevations throughout the Holocene.
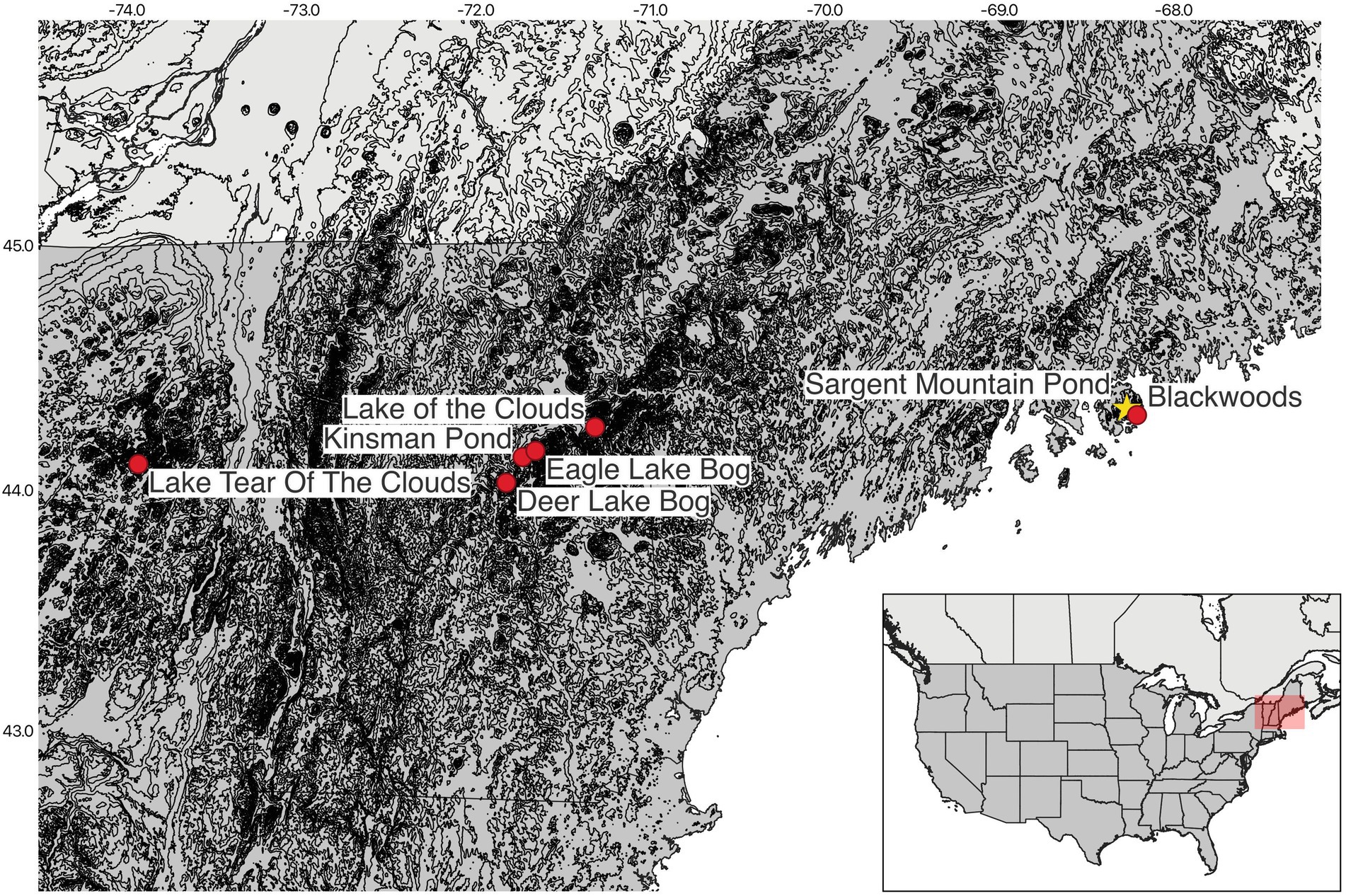
Figure 1. Location of all cores referenced in the paper. Red dots are cores referenced in the paper. The yellow star is the location of our core (as well as the location of a separate core collected by Norton et al. (2011) for limnological analyses). Cores include: Lake Tear of the Clouds (Jackson and Whitehead, 1991); Kinsman Pond, Deer Lake Bog, Eagle Lake Bog, and Lake of the Clouds (Spear, 1989; Spear et al., 1994; Miller and Spear, 1999); Blackwoods (Schauffler and Jacobson, 2002); Sargent Mountain pond (Norton et al., 2011) this study.
Assessing high elevation refugia in records from the Adirondacks and White Mountains requires macrofossils. Pollen analysis alone did not capture the contrast between dynamic lowlands and stable alpine sites (Spear, 1989; Jackson and Whitehead, 1991; Spear et al., 1994; Miller and Spear, 1999). Pollen influx at a regional scale can obscure the local vegetation signal, and, at sites near treeline, lakes will collect pollen carried upslope on the wind from forested lowlands (Webb, 1993; Birks and Birks, 2000). Thus, to identify local refugia in the paleorecord, we must look to proxies that more accurately reflect local presence. Plant macrofossils such as leaves, twigs, buds, cone scales, or needles are orders of magnitude larger and heavier than pollen, and thus tend to settle close to the parent plant, recording vegetation changes at the spatial scale of alpine refugia (Davis et al., 1980; Birks and Birks, 2000). Refugia should support a stable presence of cold-adapted taxa in the macrofossil record despite shifts in regional plant dominance (as recorded by pollen) in response to climate change during the Holocene.
Are the mountains of the northeast climate refugia? While paleobiological records from the alpine-treeline ecotone in the Adirondacks and White Mountains suggest the persistence of subalpine taxa throughout the Holocene, these studies were not designed to assess refugia or support conservation work, and represent only a few peaks in a region that stretches from upstate New York to coastal Maine (Spear, 1989; Jackson and Whitehead, 1991; Spear et al., 1994; Miller and Spear, 1999). The northeast mountains and the alpine-treeline ecotone support tundra relicts from the late Pleistocene; these alpine and subalpine plant taxa at the southern edge of their ranges are isolated by topography, ecology, and management in the northeast today, and unsurprisingly rank medium to high in climate change vulnerability assessments (Kimball and Weihrauch, 2000; Norton et al., 2011; Capers et al., 2013; Whitman et al., 2013; Ulrey et al., 2016). Identifying climate refugia with paleobiology and macrofossil analysis will help conservation managers refine such vulnerability assessments. Persistent subalpine macrofossils in local sediment records throughout the Holocene suggest the presence of a climate refugia, and indicate that these sites may buffer the vulnerability of taxa to anthropogenic climate change.
Here, we assess a climate change refugium for the alpine-treeline ecotone with paleobiological data at one of the lowest elevation sites supporting subalpine taxa and open ridges in the northeastern United States. Acadia National Park, located on the coast of Maine, is both regionally and nationally important (Publicover et al., 2021), attracting approximately 4 million visitors annually (National Park Service, 2022). Acadia’s Sargent Mountain is among the shortest peaks in the northeast to support alpine-treeline ecotone communities. Sargent Mountain Pond, a kettle hole located on the ridge, may be the oldest lake in Maine (Norton et al., 2011). The topographic complexity of the park’s summits, as well as Acadia’s proximity to the coast, are both hallmarks of potential refugia (Schauffler and Jacobson, 2002; Graae et al., 2018). The persistent presence of cold-adapted species in this macrofossil record would indicate that these peaks, and likely most taller mountains throughout Maine, served as cool climate refugia throughout the late Pleistocene and Holocene.
Study area
Sargent Mountain Pond (elevation 418 m; 44.334519, −68.269604) is located in Acadia National Park in Hancock County, Maine, United States (Figure 2). Sargent Mountain is the second tallest peak in Acadia and the Sargent South ridge lies to the west of the pond. The Maine Natural Areas Program classifies the system as a rocky summit heath (Gawler and Cutko, 2010), which is characterized by communities consisting of shrub or sapling Abies balsamea; shrub Betula populifolia, Thuja occidentalis, Picea rubens, Amelanchier sp., Viburnum nudum, Ilex sp.; dwarf shrubs Rhododendron candense, Rhododendron groenlandicum, Gaylussacia baccata, Kalmia angustifolia, and Vaccinium angustifolium; and Sphagnum mosses. The exposed ridge also supports Sibbaldiopsis tridentata, a common species in outcrops and alpine ridges, and Vaccinium vitis-idaea, a boreal species. The kettle hole lake Sargent Mountain Pond, on the southern ridge of Sargent Mountain was first exposed during deglaciation, approximately 16,600 years BP (Norton et al., 2011). The pond is mostly fed by precipitation and surface runoff, with a small amount of groundwater possibly entering through bedrock joints, and it drains into a fen to the southeast. Thirty-year climate normals (1991–2020) for Acadia National Park obtained from NOAA show an average range of 20.9°C in July to −4.6°C in January, with an average of 55.4 in. of precipitation yearly. Sargent Mountain pond was previously cored in March 2007 for an analysis of nutrient cycling, lake chemistry, sedimentation, and fire history (Norton et al., 2011). While nine pollen samples were analyzed at a coarse resolution spanning 16,600–9,800 BP, there has as yet been no plant macrofossil record from this site.
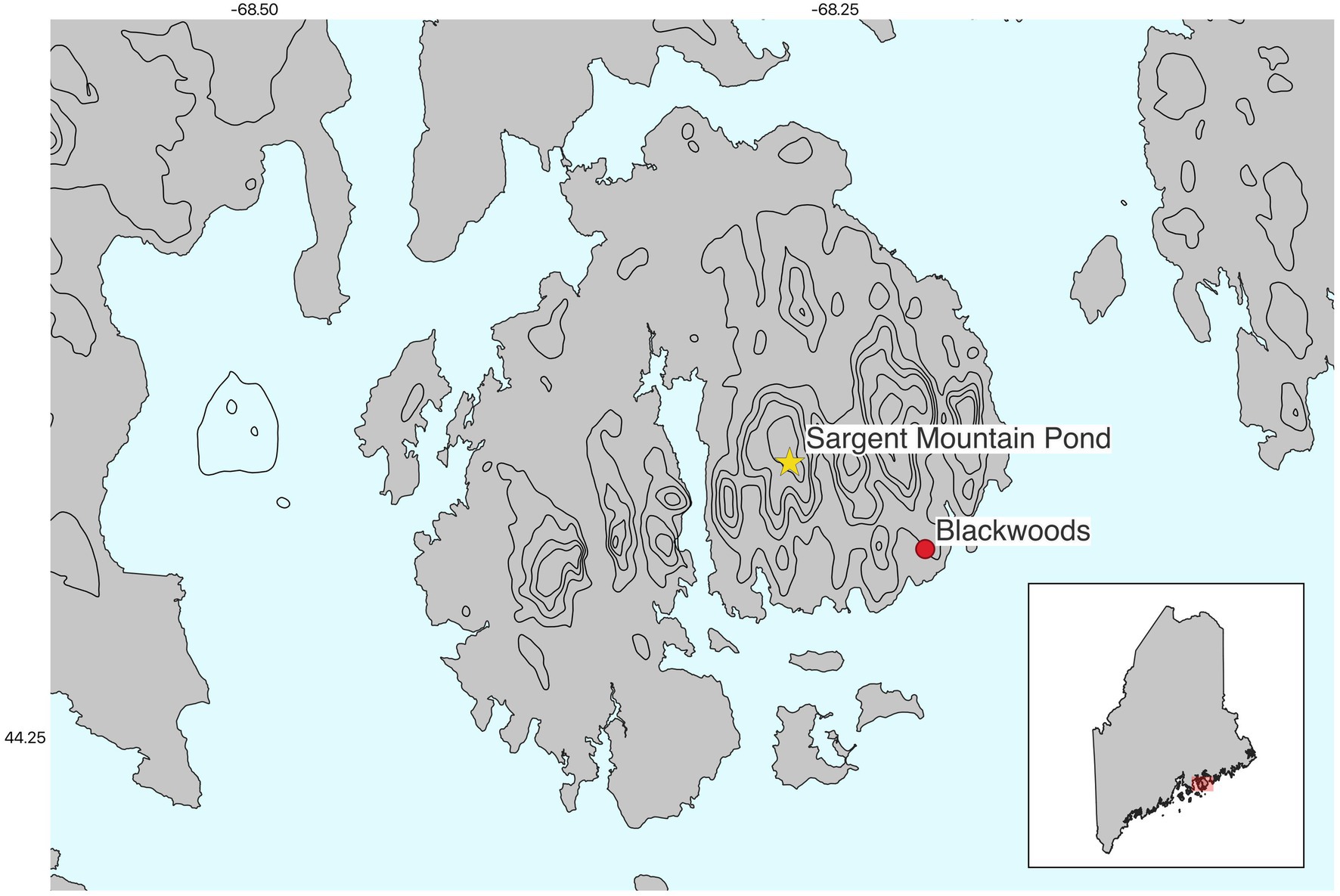
Figure 2. Location of the two cores from Mount Desert Island relative to the state of Maine. The star depicts our core, and the red dot depicts a forest hollow core collected at the Blackwoods campground (Schauffler and Jacobson, 2002).
Materials and methods
Sediment core acquisition and chronology
We collected two overlapping sediment cores in September 2017 with a modified Livingstone square-rod piston sampler, using a Bolivia adapter and polycarbonate core tube for soft surface sediments. Our core was taken from the deepest part of the pond to avoid boulders and slumping from the pond’s steep slopes. At approximately 0.75 ha, the pond is small enough that even the deepest part of the basin should contain abundant plant macrofossils. Our 409.5 cm core was built as a composite of these two overlapping cores, to avoid core gaps or disturbed sediments between drives. The core was split, imaged, and scanned, at the Continental Scientific Drilling facility at the University of Minnesota (CSD; formerly LacCore), then subsampled into 0.5 cm slices. One half of the split core remains at CSD for archiving, and the working half was transported to the University of Maine and kept in refrigeration (See Supplement 2 for the full core description).
Six terrestrial plant macrofossils were sent to the W.M. Keck Carbon Cycle AMS facility at UC Irvine for radiocarbon dating (Table 1). The R package bacon v 2.5.8 (Blaauw and Christen, 2011) was used to calibrate the radiocarbon dates and produce a Bayesian age-depth model for the core (see Supplement 1 for further discussion of our bacon age-depth model). Radiocarbon dates (Table 1) were calibrated using CALIB v8.2 (Stuiver and Reimer, 1993). Throughout the results, we report the median ages (Cal yr. BP) of the fossils as output by the bacon.hist function.
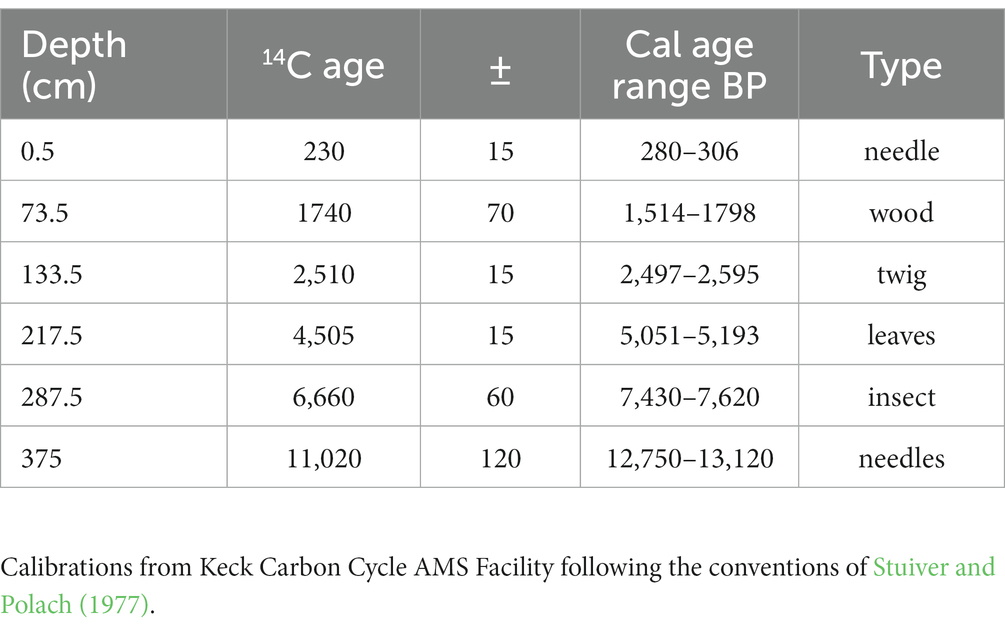
Table 1. Radiocarbon ages for Sargent Mountain Pond obtained from W. M. Keck Carbon Cycle AMS facility at UC Irvine, 2 sigma calendar year age ranges obtained from CALIB 8.2 (and rounded to the nearest 10 years), and the type of material analyzed.
Macrofossil analysis
We sieved one approximately 5 cm2 core slice every 2.5 cm for macrofossils. For this study, we focused on the bottom half of the core, which covers early- to mid-Holocene warming during the Holocene Climatic Optimum (~8,000–5,000 BP). Each sub-sample was placed in a beaker and mechanically disaggregated with DI water. The slurry was poured into a 250 μm sieve over a beaker, and DI water was used to break up any large clumps. The contents of the sieve were transferred to a petri dish, and DI water was added to cover the bottom of the dish in a thin layer of water and sediment. The petri dish was placed under a dissecting scope and systematically searched for plant fragments using 10x magnification (see Figure 3 for examples of macrofossils). As fossils were found, they were placed into 1/2 dram glass shell vials using a fine-tipped nylon paintbrush. Each plant macrofossil was identified using the reference collection at the University of Maine and CSD’s online TMI database (TMI ImageSieve, n.d.). Then the sediment was swirled and searched again under 20 × magnification. The process was repeated until all plant macrofossils were removed and identified. Vials were dried in an oven at 25°C until no water was present and then moved to a refrigerator for long-term storage or subsequent radiocarbon dating. In total, we analyzed 74 subsections of the core spanning 13,290–4,660 BP.

Figure 3. Photos of macrofossils from the Sargent Mountain Pond core, with the depth found included in parentheses: (A) Cyperaceae achene (287.5 cm), (B) Betula sp. seed (260 cm), (C) Picea sp. needle (375 cm), and (D) Betula sp. catkin scale (272.5 cm).
Results
A total of 108 terrestrial plant fossils were identified from this interval (Figure 4). These fossils are representative of plants currently found on the peak of Sargent Mountain or generally found in Acadia National Park. The oldest parts of the core (375.5–408.5 cm) consisted primarily of clay and glacial till containing no fossils. The oldest macrofossil identified for this analysis was the sporangia of a leptosporangiate fern dated at 13,090 BP. By 12,880 BP the plant macrofossils indicate that ferns, Alnus, Picea, and Abies were all present at the site. Ferns were still present in 12,750 BP alongside some unidentifiable leaf and stem tissues. 12,640 BP marks the first appearance of Juniperus in our record, most likely Juniperus communis. By 12,470 BP Betula was present at the site alongside Picea and Abies macrofossils. One Acer seed macrofossil co-occurs in the record at 12,320 BP with Abies needles. Acer was reported in the Mount Desert Island forest hollow pollen analysis and is commonly found in the park, but is not currently found on the Sargent Mountain ridge. Alnus, another taxa that is not restricted to alpine habitat in the park appears again at 11,990 BP. From 11,320 BP to 7,560 BP, the plant macrofossils observed were restricted to Abies, Betula, Juniperus, and Picea. At 7,560 BP we recorded a Cyperaceae achene. Cyperaceae does not appear again in the rest of our record. From 7,560 to 4,880 BP, the only macrofossils we recorded were from Abies, Betula, Juniperus, and Picea. The last fern sporangia in our record also occurred at the latter date.
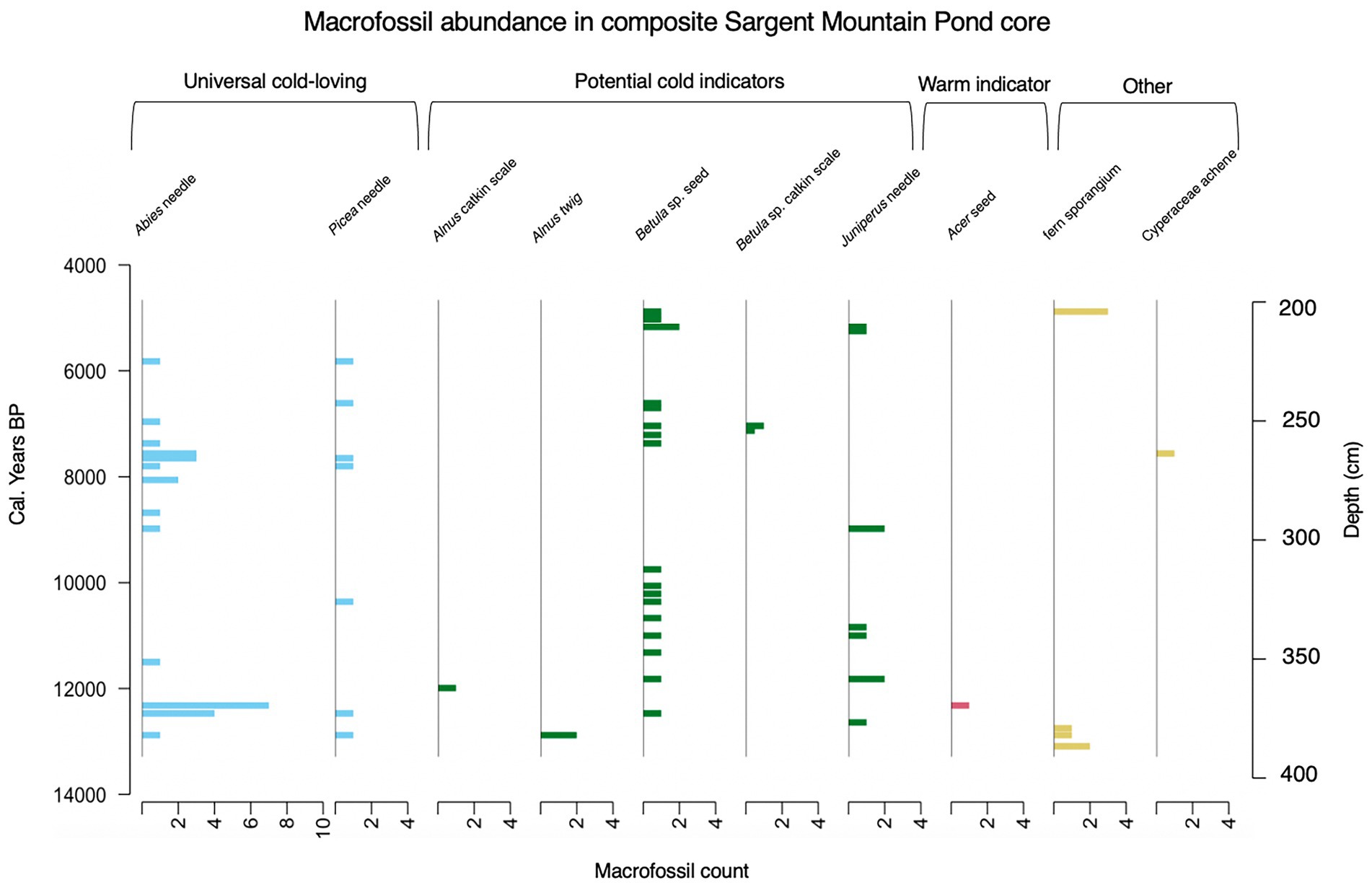
Figure 4. Plant macrofossil abundance from 13,290 BP to 4,660 BP (390–200 cm). Blue shaded graphs show counts of macrofossils of universal cold-loving species, green shaded graphs show counts of macrofossils that are potential cold indicators, pink shaded graphs show counts of macrofossils from warm indicator species, and yellow shaded graphs show counts of other, non -indicator species. Unidentifiable fragments of leaves and twigs were also collected in this core but, as they had no diagnostic characteristics, are not included in the stratigraphy. We also found evidence of Daphnia and Chara as well as other aquatic organisms, but the aquatic community in the lake is beyond the scope of this paper. We used the R package rioja (Juggins, 2020) to make this diagram.
Discussion
The Sargent Mountain Pond macrofossil record provides a proof-of-concept case study for conservation paleobiology research testing hypothesized refugia in partnership with conservation practitioners. Our results reveal that Sargent Mountain was a refugium for cold adapted taxa throughout the early Holocene and the Holocene Climatic Optimum. The persistence of macrofossils from these taxa through past rapid warming events suggests that this site—and taller mountains in Maine—may be buffered from current climate change (Dobrowski, 2011; Morelli et al., 2016). Climate change vulnerability assessments for Maine (Whitman et al., 2013; MCC STS, 2020; Publicover et al., 2021) are already considering paleobiological data: for example, Climate Change and Biodiversity in Maine: Vulnerability of Habitats and Priority Species cites Miller and Spear (1999) in its assessment of alpine habitat vulnerability. But, until now there have been no Maine-based records for vegetation at or above treeline through the early Holocene. Importantly, we co-produced our Sargent Mountain Pond work with Acadia National Park as part of a Smith Conservation Research Fellowship (Aslan et al., 2014) and an undergraduate capstone. Our research questions were designed to support park management priorities and broader efforts to understand climate change refugia (Smetzer and Morelli, 2019). Acadia National Park’s mountains support the lowest treeline in Maine. Across the state and region, researchers and managers are actively collaborating to assess and manage alpine-treeline ecotones scattered across ridges under the conservation management of federal and state agencies and NGOs (Nelson et al., 2022). Paleobiology work, especially research grounded in macrofossil analysis (Birks and Birks, 2000), will continue to support these efforts and help us refine climate change vulnerability assessments.
Macrofossils from Sargent Mountain Pond reveal that this site was a refugium for cold-adapted plant taxa throughout two Holocene warming periods, and suggest that the low, open ridges of Acadia National Park serve as climate refugia for subalpine plant communities today. Abies, Juniperus, and Picea macrofossils appear throughout the sediment record from 12,880 to 5,170 BP. Juniperus communis is common on Acadia’s cliffs, balds, and ledges today; it is an indicator of open habitats like the alpine-treeline ecotone (Mittelhauser et al., 2010). Juniperus communis is a widespread, circumboreal species that extends from Greenland throughout Canada and Alaska, down as far as Georgia where it grows on isolated mountaintops (Tirmenstein, 1999). Abies and Picea are two of the characteristic canopy trees in the Fir-Heart-leaved Birch subalpine forest community (Gawler and Cutko, 2010). Abies currently extends from Virginia to Hudson Bay, and Picea rubens, the dominant spruce in Acadia, ranges from North Carolina to Hudson Bay, although three types of spruce—black, red, and white—are currently found in the park. Betula seeds also appear in our record, but we were unable to identify these macrofossils to species and not all birches are indicators of cool climates and subalpine plant communities. Four species of Betula are present in the park today. Betula alleghaniensis extends from Georgia to Hudson Bay; Betula cordifolia (which some indicate as a variety of B. papyrifera) extends from North Carolina to Hudson Bay, but is absent in New Jersey, Delaware, and Maryland; Betula papyrifera is present from North Carolina to Hudson Bay, extending across the plains and up into Alaska; finally, Betula populifolia is found in North Carolina to Hudson Bay. Cold adapted taxa are never completely replaced by warm adapted taxa in our macrofossil record. The persistent presence of cold-adapted macrofossils from taxa still found on Sargent Mountain ridge today illustrate local resistance to regional climatic changes.
The persistent presence of cold-adapted macrofossils on Sargent Mountain contrasts with evidence of a dynamic forest history in the early Holocene at low elevations on Mount Desert Island. Pollen analysis representing stand-scale dynamics from a low elevation (30 m) wet depression at Blackwoods, a site a few kilometers to the southeast of Sargent Mountain Pond, features taxa peaking and declining throughout the late Pleistocene and Holocene (Schauffler and Jacobson, 2002). We do not expect the Blackwoods and Sargent Mountain pond source areas to overlap, as the pollen from the forest hollow below a closed canopy should come from 50 m to a maximum of a few hundred meters. At Blackwoods, Pinus pollen jumps to peaks around 9,000 BP and 7,000 BP; Pinus does not appear in our macrofossil record. Taxa associated with warmer microclimates including Acer and Quercus appear throughout the Blackwoods pollen record; in contrast we found just a single Acer seed at 12,320 BP, and it co-occurs with Abies needles. Pollen in lake sediments is a regional record (Jackson and Whitehead, 1991; Webb, 1993); the pollen in forest hollows like Blackwoods is more local, recording stand-level taxa within a few hundred meters (Schauffler and Jacobson, 2002). Macrofossils reflect the local flora at a finer spatial scale; while pollen can easily be transferred from below treeline and deposited in sediments above treeline, the source area for macrofossils is much more limited (Birks and Birks, 2000). Together, pollen and macrofossil analyses on Mount Desert Island describe a dynamic history of cold-adapted and warm-adapted taxa shifting in abundance around a low elevation forest hollow (Schauffler and Jacobson, 2002, Figure 4C), while higher elevation sites like Sargent Mountain Pond supported stable cold-adapted plant communities throughout the first half of the Holocene.
Sargent Mountain Pond is an extraordinarily low elevation refugium for subalpine plant taxa. Our macrofossil record at Sargent Mountain Pond is consistent with macrofossil records from the highest elevation sites in the Adirondack (1,320 m) and White Mountains (1,676 m) of the northeast. In New York’s Adirondack Mountains, forest composition was never steady for longer than 2,000 years—except at 1,320 m where Abies macrofossils anchored the 8,000 year record with Picea and Betula papyrifera consistently also appearing (Jackson and Whitehead, 1991). Cold-adapted taxa persisted at this high elevation site throughout the Holocene. In New Hampshire’s White Mountains, pollen and macrofossil evidence supports a relatively stable location for treeline from 10,000 BP to present (Spear et al., 1994). Abies and Picea dominate the subalpine forest below treeline over the last 9,000 years (Spear, 1989), and sediments at the highest elevation site include macrofossils of open-habitat herbs and mosses, suggesting that it has never supported a closed canopy forest (Miller and Spear, 1999). We found similar persistent records of cold-adapted and open-habitat taxa at Sargent Mountain Pond throughout the first half of the Holocene. If this small peak (at only 418 m) was able to support subalpine taxa through the Holocene, taller mountains with more surface area above treeline across Maine may also be refugia. Sargent Mountain is not just considerably smaller than the other mountains supporting alpine habitats in Maine, it is also located on the coast. The coastal influence on the climate of Maine’s islands seems to have sustained cool refugia for spruce throughout the Holocene, and may have also supported subalpine refugia on island peaks (Schauffler and Jacobson, 2002). However, throughout the Anthropocene the Gulf of Maine has warmed, with accelerated warming recorded in the last decade; 2015–2020 was the warmest 5-year period in the instrumental record and average temperatures are expected to increase 1.1–2.4°C by 2050 (Pershing et al., 2021; Seidov et al., 2021). The influence of the Gulf of Maine and the effect of warming water temperatures on Mount Desert Island’s mountain microclimates are not fully understood. But, recognizing Sargent Mountain Pond as a refugium for subalpine and alpine-treeline ecotone taxa throughout the Holocene should shift how conservation practitioners assess and manage the climate change vulnerability of these plant communities (Hunter et al., 1988; Dietl and Flessa, 2011; Morelli et al., 2016).
Conclusion
Understanding and identifying climate refugia is a top priority for conservationists and managers currently working to protect alpine and subalpine systems in the northeastern United States. Refugia provide suitable habitat for in situ conservation under anthropogenic climate change (Keppel and Wardell-Johnson, 2012; Morelli et al., 2016). Macrofossil evidence shows the ridge around Sargent Mountain Pond consistently supported cold adapted plant taxa throughout warm climates during the first half of the Holocene, at the same time pollen data reflects dynamic vegetation changes at lower elevations at the scale of Mount Desert Island. Our macrofossil analysis indicates that plant communities at treeline in Acadia National Park may be less vulnerable than expected to climate change due to this refugium. The Sargent Mountain macrofossil record could thus provide critically timed data to support managers at Acadia National Park as they grapple with conserving and prioritizing management actions for important cultural and ecological landscapes in the face of ongoing climate change. Across the United States, National Parks are among the landscapes most exposed to climate change with high latitudes, high altitudes, and coastal locations facing dramatic impacts of climate change (Gonzalez et al., 2018). Acadia National Park and its low-elevation subalpine refugia indicate that some plant taxa may be more resilient to climate change than previously thought, though as warming extends beyond the range of the Holocene, many uncertainties remain. Our results support a growing body of conservation paleobiology literature illustrating how paleorecords can broaden our understanding of how at-risk organisms respond to rapid climatic changes, in support of their ongoing management in a warming world (Dietl and Flessa, 2011).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CC and CM wrote the paper and conducted analyses. CM and JG organized field work and collected the core. CC conducted the lab work and identification of fossils. JG edited and gave significant feedback at multiple steps of the writing process. All authors contributed to the article and approved the submitted version.
Funding
Funding for this project was provided by a Smith Fellowship to CM. The Sierra Club and Friends of Acadia provided support and in the field.
Acknowledgments
Lab work for this project was conducted on unceded territory of the Penobscot Nation and fieldwork was conducted on Wabanaki Confederacy land. Much of the writing and revisions were conducted on Wašiw and Numu land. Many thanks to Katie Glover, Sarah Nelson, and Kate Ruskin for inspiration and support in the completion of CC undergraduate capstone at the University of Maine. Initial core description, imaging, and scanning analysis were provided by the Continental Science Drilling Facility at the University of Minnesota. Mikayla Mason, Mario Williams, and Mac Mackenzie provided helpful support with the macrofossil analysis, and the BEAST Lab provided a supportive and encouraging space for learning and research. Kirsten Dutcher, Nathan Byer, Steve Hromada, and Anjana Parandahaman at the University of Nevada, Reno gave valuable feedback on this manuscript. Chrissy Klenke assisted CC with the map figures. The authors are grateful to Abe Miller-Rushing and the rangers and staff of Acadia National Park, as well as Karen James, Bill Buaas, and volunteers from the Sierra Club and Friends of Acadia for field assistance. We are also grateful to two reviewers, whose comments greatly improved this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1008594/full#supplementary-material
References
Aslan, C. E., Pinsky, M. L., Ryan, M. E., Souther, S., and Terrell, K. A. (2014). Cultivating creativity in conservation science. Conserv. Biol. 28, 345–353. doi: 10.1111/cobi.12173
Barrington, D. S., and Paris, C. A. (2007). Refugia and migration in the quaternary history of the New England flora. Rhodora 109, 369–386. doi: 10.3119/0035-4902(2007)109[369:RAMITQ]2.0.CO;2
Birks, H. H., and Birks, H. J. B. (2000). Future uses of pollen analysis must include plant macrofossils. J. Biogeogr. 27, 31–35. doi: 10.1046/j.1365-2699.2000.00375.x
Blaauw, M., and Christen, J. A. (2011). Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474. doi: 10.1214/ba/1339616472
Capers, R. S., Kimball, K. D., McFarland, K. P., Jones, M. T., Lloyd, A. H., Munroe, J. S., et al. (2013). Establishing alpine research priorities in northeastern North America. Northeast. Nat. 20, 559–577. doi: 10.1656/045.020.0406
Davis, M. B., Spear, R. W., and Shane, L. C. (1980). Holocene climate of New England. Quat. Res. 14, 240–250. doi: 10.1016/0033-5894(80)90051-4
Dietl, G. P., and Flessa, K. W. (2011). Conservation paleobiology: putting the dead to work. Trends Ecol. Evol. 26, 30–37. doi: 10.1016/j.tree.2010.09.010
Dobrowski, S. Z. (2011). A climatic basis for microrefugia: the influence of terrain on climate. Glob. Chang. Biol. 17, 1022–1035. doi: 10.1111/j.1365-2486.2010.02263.x
Gawler, S. C., and Cutko, A. (2010). Natural Landscapes of Maine: A Classification of Vegetated Natural Communities and Ecosystems. Augusta, ME: Maine Department of Agriculture, Conservation, and Forestry—Bureau of Geology, Natural Areas, and Coastal Resources: Maine Natural Areas Program.
Gonzalez, P., Wang, F., Notaro, M., Vimont, D. J., and Williams, J. W. (2018). Disproportionate magnitude of climate change in United States National Parks. Environ. Res. Lett. 13:104001. doi: 10.1088/1748-9326/aade09
Graae, B. J., Vandvik, V., Armbruster, W. S., Eiserhardt, W. L., Svenning, J. C., Hylander, K., et al. (2018). Stay or go–how topographic complexity influences alpine plant population and community responses to climate change. Perspect. Plant Ecol. Evol. Syst. 30, 41–50. doi: 10.1016/j.ppees.2017.09.008
Hunter, M. L. Jr., Jacobson, G. L. Jr., and Webb, T. III (1988). Paleoecology and the coarse-filter approach to maintaining biological diversity. Conserv. Biol. 2, 375–385. doi: 10.1111/j.1523-1739.1988.tb00202.x
Jackson, S. T., and Whitehead, D. R. (1991). Holocene vegetation patterns in the Adirondack Mountains. Ecology 72, 641–653. doi: 10.2307/2937204
Juggins, S. (2020). Rioja: analysis of quaternary science data, R package version (0.9-26). Available at: https://cran.r-project.org/package=rioja (Accessed December 26, 2022).
Keppel, G., Mokany, K., Wardell-Johnson, G. W., Phillips, B. L., Welbergen, J. A., and Reside, A. E. (2015). The capacity of refugia for conservation planning under climate change. Front. Ecol. Environ. 13, 106–112. doi: 10.1890/140055
Keppel, G., and Wardell-Johnson, G. W. (2012). Refugia: keys to climate change management. Glob. Chang. Biol. 18, 2389–2391. doi: 10.1111/j.1365-2486.2012.02729.x
Kimball, K. D., and Weihrauch, D. M. (2000). “Alpine vegetation communities and the alpine-treeline ecotone boundary in New England as biomonitors for climate change.” in Wilderness as a Place for Scientific Inquiry, USDA Forest Service Proceedings, 93–101.
MCC STS (2020). Scientific assessment of climate change and its effects in Maine. A report by the Scientific and Technical Subcommittee (STS) of the Maine Climate Council (MCC). Augusta, Maine, 370.
Miller, N., and Spear, R. (1999). Late-quaternary history of the alpine flora of the New Hampshire White Mountains. Géog. Phys. Quatern. 53, 137–157. doi: 10.7202/004854ar
Mittelhauser, G. H., Gregory, L. L., Rooney, S. C., and Weber, J. E. (2010). The Plants of Acadia National Park. University of Maine Press. Orono, ME.
Morelli, T. L., Daly, C., Dobrowski, S. Z., Dulen, D. M., Ebersole, J. L., Jackson, S. T., et al. (2016). Managing climate change refugia for climate adaptation. PLoS One 11:e0159909. doi: 10.1371/journal.pone.0159909
Nadeau, C. P., Giacomazzo, A., and Urban, M. C. (2022). Cool microrefugia accumulate and conserve biodiversity under climate change. Glob. Chang. Biol. 28, 3222–3235. doi: 10.1111/gcb.16143
National Park Service (2022). Acadia. Available at: https://www.nps.gov/acad/index.htm. [Accessed June 9, 2022].
Nelson, S., MacKenzie, C. M., Morelli, T. L., Wason, J., Wentzell, B., Hovel, R., et al. (2022). Introduction: climate change in the mountains of Maine and the northeast. Northeast. Nat. 28, ii–ix. doi: 10.1656/045.028.s1111
Norton, S. A., Perry, R. H., Saros, J. E., Jacobson, G. L., Fernandez, I. J., Kopáček, J., et al. (2011). The controls on phosphorus availability in a boreal lake ecosystem since deglaciation. J. Paleolimnol. 46, 107–122. doi: 10.1007/s10933-011-9526-9
Oswald, W. W., Foster, D. R., Shuman, B. N., Doughty, E. D., Faison, E. K., Hall, B. R., et al. (2018). Subregional variability in the response of New England vegetation to postglacial climate change. J. Biogeogr. 45, 2375–2388. doi: 10.1111/jbi.13407
Pershing, A. J., Alexander, M. A., Brady, D. C., Brickman, D., Curchitser, E. N., Diamond, A. W., et al. (2021). Climate impacts on the Gulf of Maine ecosystem: a review of observed and expected changes in 2050 from rising temperatures. Elem Sci Anth 9:00076. doi: 10.1525/elementa.2020.00076
Peteet, D. M., Daniels, R. A., Heusser, L. E., Vogel, J. S., Southon, J. R., and Nelson, D. E. (1993). Late-glacial pollen, macrofossils and fish remains in northeastern USA—the younger Dryas oscillation: a contribution to the ‘North Atlantic seaboard programme’of IGCP-253,‘termination of the Pleistocene’. Quat. Sci. Rev. 12, 597–612. doi: 10.1016/0277-3791(93)90002-4
Peteet, D. M., Vogel, J. S., Nelson, D. E., Southon, J. R., Nickmann, R. J., and Heusser, L. E. (1990). Younger Dryas climatic reversal in northeastern USA? AMS ages for an old problem. Quat. Res. 33, 219–230. doi: 10.1016/0033-5894(90)90020-L
Publicover, D. A., Kimball, K. D., and Poppenwimer, C. J. (2021). Northeastern high-elevation areas: ecological values and conservation priorities. Northeast. Nat. 28, 129–155. doi: 10.1656/045.028.s1108
Schauffler, M., and Jacobson, G. L. Jr. (2002). Persistence of coastal spruce refugia during the Holocene in northern New England, USA, detected by stand-scale pollen stratigraphies. J. Ecol. 90, 235–250. doi: 10.1046/j.1365-2745.2001.00656.x
Seidov, D., Mishonov, A., and Parsons, R. (2021). Recent warming and decadal variability of gulf of Maine and slope water. Limnol. Oceanogr. 66, 3472–3488. doi: 10.1002/lno.11892
Smetzer, J., and Morelli, T. L. (2019). Incorporating Climate Change Refugia Into Climate Adaptation in the Acadia National Park Region. Article, Smith College, Northampton, MA. Available at: https://scholarworks.smith.edu/second_century_refugia/44
Spear, R. W. (1989). Late-quaternary history of high-elevation vegetation in the White Mountains of New Hampshire. Ecol. Monogr. 59, 125–151. doi: 10.2307/2937283
Spear, R. W., Davis, M. B., and Shane, L. C. (1994). Late quaternary history of low-and mid-elevation vegetation in the White Mountains of New Hampshire. Ecol. Monogr. 64, 85–109. doi: 10.2307/2937056
Stuiver, M., and Reimer, P. J. (1993). CALIB rev. 8. Radiocarbon 35, 215–230. doi: 10.1017/S0033822200013904
Tirmenstein, D. (1999). Juniperus communis. In: Fire effects information system, [online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (producer). Available: https://www.fs.usda.gov/database/feis/plants/shrub/juncom/all.html [Accessed December 4, 2022].
TMI ImageSieve (n.d). https://tmi.laccore.umn.edu/is
Ulrey, C., Quintana-Ascencio, P. F., Kauffman, G., Smith, A. B., and Menges, E. S. (2016). Life at the top: long-term demography, microclimatic refugia, and responses to climate change for a high-elevation southern Appalachian endemic plant. Biol. Conserv. 200, 80–92. doi: 10.1016/j.biocon.2016.05.028
Webb, T. (1993). Constructing the past from late-quaternary pollen data: temporal resolution and a zoom lens space-time perspective. Short Courses Paleontol. 6, 79–101. doi: 10.1017/S2475263000001069
Whitman, A., Cutko, A., deMaynadier, P., Walker, S., Vickery, B., Stockwell, S., et al. (2013). Climate change and biodiversity in Maine: vulnerability of habitats and priority species. Manomet Center for Conservation Sciences (in collaboration with Maine beginning with habitat climate change working group) report SEI-2013-03. Brunswick, Maine.
Keywords: macrofossils, conservation paleobiology, subalpine, refugia, climate change vulnerability assessment
Citation: Carroll CF, Gill JL and McDonough MacKenzie C (2023) Early Holocene plant macrofossils indicate cool refugia for subalpine plant taxa in Acadia National Park, Maine. Front. Ecol. Evol. 11:1008594. doi: 10.3389/fevo.2023.1008594
Edited by:
G. Lynn Wingard, United States Geological Survey (USGS), United StatesReviewed by:
Dorothy M. Peteet, Goddard Institute for Space Studies (NASA), United StatesLysanna Anderson, United States Geological Survey (USGS), United States
Copyright © 2023 Carroll, Gill and McDonough MacKenzie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cas F. Carroll, ✉ Y2FzY2Fycm9sbEBuZXZhZGEudW5yLmVkdQ==; Caitlin McDonough MacKenzie, ✉ Y2FpdGxpbi5tY2Rvbm91Z2hAbWFpbmUuZWR1